75 TEMPORAL AND SEQUENTIAL ORGANISATION OF SONG BOUTS IN THE STARLING MARCEL EENS, RIANNE PINXTEN & RUDOLF FRANS VERHEYEN ABSTRACT Temporal analysis of Starling song revealed that males sing very long and complex song bouts, some extending over one minute and containing over ninety song bouts. Starlings sing with immediate variety. There are clear differences in average bout lenght and in singing rate be- tween males. Starling song bouts have a characteristic sequential organisa- tion which might be an important species-specific feature. Bouts are com- posed of four categories of song types. A bout typically begins with one or several whistles. These whistles are always followed by a large number of variable song types which, in thier tum, gradually pass into rattle song ty- pes. A bout usually ends with a series of high-frequency song types. A de- tailed sequential analysis of one bird showed that the Starling is a highly determinate singer. Repertoire size ranged from 43 to 48 song types. Starling song is more variable and versatile at the start of a bout than at the end. Although the primary function of the song seems to be attract a female, it is suggested that different song types have different functions. Department of Biology, University of Antwerp, U.LA., Universiteitsplein I, B-261O Wilrijk, Belgium INTRODUCTION In most songbird species the male sings several versions of the species-specific song (i.e. song types) as part of his territorial and/or mate-attrac- ting behaviour (Hartshorne 1973, Krebs & Kroodsma 1980, Catchpole 1982). The Starling Sturnus vulgaris, one of the world's commonest species of birds, is well-known for its complexity and mimicry in song (Feare 1984). Little is known, however, about the Starling's song reper- toire size and about the manner of presentation of its song types during a singing-performance. As the breeding season approaches, male Starlings sing four different categories of song: I. whistles or whistled songs (Hausberger & Guyomarc'h 1981, Adret-Hausberger 1982, 1983,1984); 2. very complex song bouts including hetero- specific imitations (Hausberger & Guyo- marc'h 1981, Hindmarsh 1984, 1986); Received 20 December 1987; revised 4 August 1988 3. ImItations of other birds or of non-avian sounds produced between song bouts or be- tween whistles (Hindmarsh 1984, 1986); 4. a very loud, high-pitched note (scream) given when other Starlings approach from a distan- ce (Davis 1959). Sonagraphic as well as quantitative analyses of the Starling'S song have so far been confined to the whistled songs and the imitations in and between song bouts. Hausberger &.Guyomarc'h (1981) showed that each male Starling has a re- pertoire of 7 to 12 whistles. Hindmarh (1981, 1986) found that Starlings have a repertoire of about 15 to 20 distinct imitations. The internal structure of the complex song bouts has not been investigated yet. Starling song bouts are usually described as a medley of a gre- at variety of sounds: "whistles, clicks, rattles, squeaks and screeches all are involved' (Feare 1984, Peterson et al. 1983). On first encounter, a Starling song bout presents itself as a rapid suc-
Transcript
75
TEMPORAL AND SEQUENTIAL ORGANISATION OF SONG BOUTS
IN THE STARLING
MARCEL EENS, RIANNE PINXTEN & RUDOLF FRANS VERHEYEN
ABSTRACT Temporal analysis of Starling song revealed that males singvery long and complex song bouts, some extending over one minute andcontaining over ninety song bouts. Starlings sing with immediate variety.There are clear differences in average bout lenght and in singing rate between males. Starling song bouts have a characteristic sequential organisation which might be an important species-specific feature. Bouts are composed of four categories of song types. A bout typically begins with one orseveral whistles. These whistles are always followed by a large number ofvariable song types which, in thier tum, gradually pass into rattle song types. A bout usually ends with a series of high-frequency song types. A detailed sequential analysis of one bird showed that the Starling is a highlydeterminate singer. Repertoire size ranged from 43 to 48 song types.Starling song is more variable and versatile at the start of a bout than atthe end. Although the primary function of the song seems to be attract afemale, it is suggested that different song types have different functions.
Department of Biology, University of Antwerp, U.LA., UniversiteitspleinI, B-261O Wilrijk, Belgium
INTRODUCTION
In most songbird species the male sings severalversions of the species-specific song (i.e. songtypes) as part of his territorial and/or mate-attracting behaviour (Hartshorne 1973, Krebs &Kroodsma 1980, Catchpole 1982). The StarlingSturnus vulgaris, one of the world's commonestspecies of birds, is well-known for its complexityand mimicry in song (Feare 1984). Little isknown, however, about the Starling's song repertoire size and about the manner of presentation ofits song types during a singing-performance. Asthe breeding season approaches, male Starlingssing four different categories of song:I. whistles or whistled songs (Hausberger &
2. very complex song bouts including heterospecific imitations (Hausberger & Guyomarc'h 1981, Hindmarsh 1984, 1986);
Received 20 December 1987; revised 4 August 1988
3. ImItations of other birds or of non-aviansounds produced between song bouts or between whistles (Hindmarsh 1984, 1986);
4. a very loud, high-pitched note (scream) givenwhen other Starlings approach from a distance (Davis 1959).Sonagraphic as well as quantitative analyses
of the Starling'S song have so far been confinedto the whistled songs and the imitations in andbetween song bouts. Hausberger &.Guyomarc'h(1981) showed that each male Starling has a repertoire of 7 to 12 whistles. Hindmarh (1981,1986) found that Starlings have a repertoire ofabout 15 to 20 distinct imitations.
The internal structure of the complex songbouts has not been investigated yet. Starling songbouts are usually described as a medley of a great variety of sounds: "whistles, clicks, rattles,squeaks and screeches all are involved' (Feare1984, Peterson et al. 1983). On first encounter, aStarling song bout presents itself as a rapid suc-
76 ARDEA 77 (1989)
cession of an enormous number of different notes. The first subjective impression is of "variation and complexity" and almost "stereotype sequence of notes". The following analysis of theStarling's song was, therefore, carried out in order to:a. reveal and document how long and complex
song bouts are;b. describe the temporal and sequential organi
sation of the song bouts;c. determine the song repertoire size;d. find out whether interindividual vanatlOn
exists in song structure and/or in song patterning.
MATERIALS AND METHODS
This study was carried out in a colony of artificial nest boxes located on the campus of theUniversity of Antwerp in Wilrijk (51°10' N,4°25' E), Belgium. Male Starlings can be observed near their nest boxes from late January untilJune, and some also a brief time at the beginningof September ("Herbstbalz", Berthold 1964).Males were caught sleeping in the nest boxes atnight or with automatic traps when they were inspecting the nest boxes. They were ringed with anumbered metal ring, individually marked usingcolour-coded wing tags and their age was determined.
Male Starlings only actively defend their nesthole and its immediate surrounding and it is alsohere that they do most of their singing (Verheyen1969, Feare J984). Males were recorded near theirnest boxes from Mid-February to May 1984,though the majority of the recordings was madein April. All males were unmated at the time theywere recorded. Although several birds were recorded, the final analysis is based upon 3 malesfrom whom recordings of good quality were obtained. A fourth male was used to determine thesong bout length only. In the course of this studyit soon became evident that we could not get close enough to the singing males to obtain goodquality recordings by using a parabolic reflector.
Therefore all recordings were made with a Uher4400 Report Stereo IC tape recorder in connection with (via a long cable) a Sony ECM-16T ora ECM-50 PS electret microphone implanted close to or in the nest boxes. As unmated males nearly always sing on the wooden perches attachedto the bottom of the nest boxes, good recordingscan be obtained in this way. We used BASF tapestype DP 26 Ferro LH HiFi. The recording speedwas 19 cm/sec. The recordings were studied withthe aid of a Unigon spectrum analyser model4500 (Uniscan). Sonagrams on paper were madeusing a Kay 6061 B Sonagraph at the wide-bandsetting.
RESULTS
Temporal organisationSonagrams Detailed sonagraphic analyses revealed that Starling song bouts can be divided intogroups of notes forming a recognizable sequenceor pattern in time, which we call song types. Theelements or notes making up a song type are always coupled together in a fixed order. Starlingsusually repeat a given song type before theyswitch to the next (see below). For a given individual, different repetitions of the same songtype can vary slightly (see below and Fig. 4A).First of all, we wish to define song bouts andsong types on a purely temporal basis, that is ontheir duration and the intervals between them.
Song bout length The overall mean length was28.5 ± 17.7 sec (mean ± standard deviation) or237 song bouts of four males (a song bout is defined here as a succession of at least two different song types containing no pause greater that1.5 sec). The minimum bout length was 2 secand the maximum 81 sec. As Table 1 shows, wefound a highly significant difference in boutlength between the four males. Notice also that·there is a considerable amount of variation in themaximum bout length of the four males as therange of values is 46 to 81 sec.
SONG OF THE STARLING 77
Table 1. Comparison of the average song bouw length (sec) of four males. N = number of song bouts; x = average; SD =Standard deviation. (ANOVA on .vx-transformed data, F3.233 =6.69; p < 0.001).
Song type length and time intervals between songWe found no statistically significant differencesin average song type length between the threemales (ANOVA F 2.305 = 1.24; p > 0.25). Themean length of a song type was 0.64 ± 0.22 sec.(N = 308; 3 males). Song type lengths rangedfrom 0.16 to 2.40 sec. As Fig. 1A shows, morethan 95% of the lengths were between 0.20 and1.00 sec. Time intervals between song types didnot differ significantly between the three males
(ANOVA F 2,288 = 1.107; p > 0.25). The overallmean for 291 intervals was 0.14 ± 0.26 sec. Timeintervals ranged from 0 to 1.4 sec. Of these intervals, more than 95% were between 0 and 0.4 sec(Fig. 1B).
Repetition of song types As Fig. 2 shows, Starlings tend to repeat a given songtype before switching to another. On average, a song type wassung 2.38 ± 1.22 times (N = 872; 3 males) before
o .1 .2 .3 .4 >.4
200
60
o .1 .2 .3 .4 .5 .6 .7 .8 .9 1.0 1.1 >1.1
80
150>u
> Zu wZ 40 ;:)w 0 100:J W0 0::u... U.0::u..
20 50
SONG TYPE LENGTHS (SEC.)
Fig. lA. Frequency distribution of song type lengths.Data of three males are combined.
SONG TYPE INTERVALS (SEC.)
Fig. lB. Frequency distribution of intervals betweensong types. Data of three males are combined.
78 ARDEA 77 (1989)
NUMBER OF TIMES A SONG TYPE IS SUNG
Fig. 2. Frequency plot of the number of times a sungtype is sung before another is introduced. Data of threemales are combined.
another was introduced. The averages of bird 1,bird 2 and 3 respectively, were 2.25 ± 1.12(N = 569), 2.94 ± 1.20 (N = 139) and 2.38 ± 1.45(N = 164). The frequency - Fig. 2 en Fig. lA enIB - distributions of the three males differedhighly significant (X21O = 49.39; p < 0.001)which means that bird 2 repeated its song typessignificantly more than the two others. It can beclearly seen in Fig. 2 that a given song type issung very rarely more than 5 times. The observed maximuffi' was 12.
Number of song types sung per song bout Theoverall mean number of song types sung persong bout was 34.2 (SD = 24.3) for 57 songbouts belonging to the three males. A KruskalWallis analysis of variance showed that therewas no statistically significant difference in thenumber of song types sung per bout between thethree males (H = 2.43, df = 2; p > 0.20). Thenumber of song types sung per song bout dividedby the song bout length, however, did differhighly significant (H = 13.0, df = 2; p < 0.001)indicating that there was a significant differencein the number of song types sung per unit of time
Number of different song types sung per song boutThe three males sang, on average, 13.4 differentsong types per song bout (SD = 7.9; N = 57).Neither the number of different song types sungper bout (Kruskal-Wallis analysis of varianceH =0.43, df = 2; p < 0.8) nor the former dividedby the bout length (H = 0.14, df = 2; P > 0.9)differed significantly between the three males.The maximum recorded number of different songtypes sung per song bout was nearly the same forall three males: bird 1 and 3 sang a maximum of26 different song types per bout, bird 2 sang amaximum of 25. We found a strong positive linear correlation between bout length and the number of different song types sung for the three males (p < 0.001 for the three males; bird 1:r =0.937, df =8; bird 3: r =0.927, df =7) meaning that longer song bouts contain more different song types.
between the three males. When plotting songbout length against the number of song typessung per song bout, a strong positive linear correlation was found for all three males (bird 1:r =0.930, df =36, p < 0.001; bird 2: r =0.992,df = 8, p < 0.001; bird 3: r = 0.991, df = 7,p < 0.001) which means that longer song boutscontain more song types.
Sequential organisationSonagrams Detailed analysis by sonagram revealed that Starling song bouts are composed offour general categories of song types (Fig. 3,4B). These are:1. whistles or whistled songs;2. variable song types;3. rattle song types;4. terminal or high-frequency song types.
Whistles (1) Whistles are pure, tonelike soundswith or without frequency modulation. Of allStarling song types, these are the simplest havinga sonagraphic picture which permits easy measurement (Fig. 3, 4B). A Starling song bouttypically begins with one or several whistles(X21 = 65.7, N = 223; p < 0.001).
4 '.. .' \ 'I' "f III II I~ .............. It· ,.- \. UM',..
2
",'" \ f ItI ~~'I~
." ,'1'
r--_~V~20"'-------'i ,r--_V:..;;2:..;;1__-,
R4R3
V13 V14 V15r---1 r--I
~~I 1\ ,
·11I ,1.• /.
,I; '-'
\ \~r' !;II! MI ,H~ .t,fu!
T1 T2 T3 T4 T5I
~'i,~~~ .. :fIUI
\~ Vi ......~&«. ~_ ;vr~
. .j
A :1 11 "'I·;'1, ; .) .'
~ ; .~..,
V17 R1 R2
, ( ,ll '11"'1''1'\1 -\i '11111 ~ "111111 I'J,"\ ' '. p.!! 'f! I' t~. I""I . 1\, , f II. " ~ , I I 'I " ."
i\\i\\~ 111111'i lji~.!.'\HI~I.I: '1\1':I'J~: . 'll. ll~I' f I} \Iflli~ I!: ~d~ 1M I I " ~J I! I .I \,,, ",·'t I I .., .. I I II I II ij " I II':I~~ " 'I I If I 11: '1
r---,-,R5.:....---" r-I------.
V22 V23 V12
8 .. .. I • , ...6 t \ ... 1 _" \ ;~' l "
'!. \, , r4
"Ik:-'!if .,,, · .IL 8\ '" I ,11M I;.' ,,"{ n2 • I i:I'~~ I'" ~ I II
". ,till 1"" I".." i •. -11111 1 I I\ ".,.,.
• .
12108642
>uzw::>aw0:::LL
. : ....
o 1 2 3 4
TIME (SEC>
Fig. 3. Illustration of a complete Starling song bout, that is a bout containing all four categories of song types.This bout of bird 1 lasted 61 seconds and contained 23 different song types (W = whistle, V = variable song type,R = rattle song type, T = terminal of high-frequency song type). Apart from the introducing whistle, every songtype was repeated before the next was sung. Due to lack of space, every song type is presented only once. Formore details see text.
Bird 1 had a repertoire of 10 whistle typeswhile bird 2 and 3 sang 9. Many of these whistles are also sung between song bouts, withoutfanning part of a song bout. Some whistles,however, are never sung as part of a bout. Whistles sung as part of a bout are mostly not repeated(X2I = 160, N = 172; p < 0.001). Some whistles
could be recognized in the repertoires of all threemales while others were unique. Whistles aresung very loudly and can be heard over long distances.
Variable song types (2) In a song bout, the introductory whistles are always followed by a large
80
--N:I:~......>UZw:::Jow0::LL
8642
12108642
o
w
1
ARDEA 77 (1989)
v
2
R
3
A
T
~,'4 Ii p","
B
4
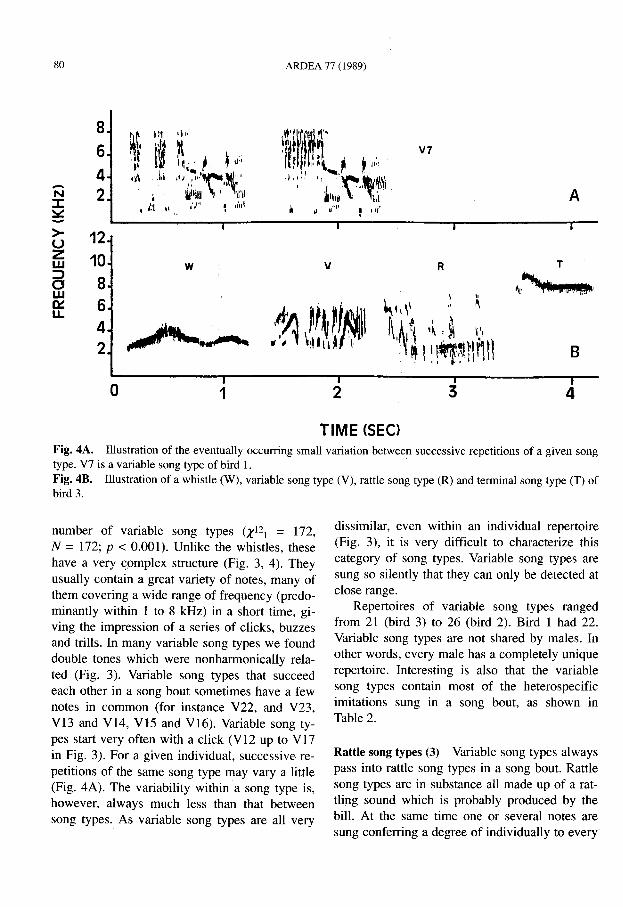
TIME (SEC)Fig. 4A. . Illust~ation of the eventually occurring small variation between successive repetitions of a given songtype. V7 IS a vanable song type of bird 1.F,ig. 4B. Illustration of a whistle (W), variable song type (V), rattle song type (R) and terminal song type (T) ofbird 3.
number of variable song types (X12 \ = 172,N = 172; P < 0.001). Unlike the whistles, thesehave a very ~.omplex structure (Fig. 3, 4). Theyusually contain a great variety of notes, many ofthem covering a wide range of frequency (predominantly within 1 to 8 kHz) in a short time, giving the impression of a series of clicks, buzzesand trills. In many variable song types we founddouble tones which were nonharmonically related (Fig. 3). Variable song types that succeedeach other in a song bout sometimes have a fewnotes in common (for instance V22, and V23,VB and V14, V15 and VI6). Variable song types start very often with a click (V12 up to V17in Fig. 3). For a given individual, successive repetitions of the same song type may vary a little(Fig. 4A). The variability within a song type is,however, always much less than that betweensong types. As variable song types are all very
dissimilar, even within an individual repertoire(Fig. 3), it is very difficult to characterize thiscategory of song types. Variable song types aresung so silently that they can only be detected atclose range.
Repertoires of variable song types rangedfrom 21 (bird 3) to 26 (bird 2). Bird 1 had 22.Variable song types are not shared by males. Inother words, every male has a completely uniquerepertoire. Interesting is also that the variablesong types contain most of the heterospecificimitations sung in a song bout, as shown inTable 2.
Rattle song types (3) Variable song types alwayspass into rattle song types in a song bout. Rattlesong types are in substance all made up of a rattling sound which is probably produced by thebill. At the same time one or several notes aresung conferring a degree of individually to every
SONG OF THE STARLING 81
Table 2. Number of heterospecific imitations occurring in each of the four categories of song types. Dataof three males are combined. W = whistles; V = variable song types; R = rattle song types; H = highfrequency song types; N = total number of song typesper category.
Table 3. Number of song types per category and total repertoire size (TR) for each of the three males.W =whistles; V =variable song types; R =rattle songtypes; T = terminal or high-frequency song types.
ImitationsN
W V
I 1728 69
R
I24
H
116
WVRTR
Bird 1 Bird 2 Bird 3
10 9 922 26 21
5 6 343 48 46
four categories of song types in a bout, rattlesong types are, nevertheless, the most difficult todivide into simple units. In several rattle song types we found double tones which were non-harmonically related. Another typical feature of thiscategory of song types are sharp changes in amplitude, such as sudden onsets and terminations.
Repertoires of this category ranged from 5(bird 1) to 13 (bird 3). Bird 2 had 6. Rattle songtypes are, like variable song types only audible ata fairly close distance.
Terminal or high-frequency song types (4) Rattlesong types are always followed by a series ofhigh pitched notes in a Starling song bout. Theseare predominantly within the frequency range 7to 12 kHz (Fig. 3, 4B). Many of them are frequency-modulated and often a clear division intofrequencies is not possible, giving the impressionof a harsh, squealing or screeching cry. The highfrequency song types, which are mostly accompanied by vigorous wing-waving, are the loudestof a song bout. Some terminal song types wereshared by the three males. Bird 1 has a repertoireof 6 terminal song types, bird 2 of 7 and bird 3sang only 3.
It needs to be stressed, however, that less than20% of the bouts end with terminal song typessince the song can be interrupted at any stage during a bout. As the high-frequency song typesterminate the bouts, they are sung the least. Acomplete song bout, being a bout containing allfour categories of song types, contains never allthe variable song types that a male can sing whe-
reas it contains mostly all the rattle and the terminal song types.
It can be seen (Table 3) that the total repertoire size of the three males ranged from 43 (bird 1)to 48 (bird 2). Bird 3 sang in total 46 song types.These figures do not include the heterospecificimitations which are sung between song bouts.
So far we showed that Starling song bouts arecomposed of 4 categories of song types following each other in a fixed order. A next questionwe wished to answer was whether the sequencing of song types in a song bout was random orwhether it was deterministic.
For the study of sequential relationships ofsong types within song bouts, it is essential thatall song types be known. When plotting the cumulative number of song types against consecutive song bouts, an asymptote was reached for thethree males. Bird 3 needed only 8 song bouts tosing his full repertoire, bird 2, 12 and bird 1, 16.Although we analysed more than 60 song boutsof bird 1, no new song types were found after thesixteenth. Of bird 2 and 3 we analysed in total respectively 16 and 40 song bouts.
The first step in looking for song type sequence patterns is to determine whether or notthe arrangement is random and independent(Isaac & Marler 1963). We tested our data by making up a transition matrix of sequence patternswithin song bouts for bird 1. Since 5 whistleswere not or only very rarely sung as part of asong bout, we used 38 of 43 song types for thisanalysis. Since we are at present only interestedin the transitions between different song types,
82
\I)
wQ.>...ozo\I)
ozowUwCIl:Q.
ARDEA 77 (1989)
FOLLOWING SONG TYPES
vlv V V V V V V V V V V V V VWW WW V V V V V V V 1 1 1 1 1 1 1 1 R R R R R T T T T T T W 1 1 2 2 21 2 3 4 1 2 3 4 5 6 7 a 9 0 1 2 3 4 5 6 7 1 2 3 4 5 1 2 3 4 5 6 5 a 9 0 1 2
Fig. 5. Transition matrix of preceding and following song types from bird I. W = whistles; V = variable songtypes; R - rattle song types; T = terminal song types or high-frequency song types.
we chose a matrix without a descending diagonal.
It can be clearly seen in Table 4 that there arefewer pair combinations than we would expect ifthe transitions were random and independent: ofthe thirty-seven possible song types that couldfollow any given song type, the maximum observed was four and in most cases there was only
one. This extremely high level of dependence results in a nearly determined system. Song typescould be arranged in such an order that the majority of the entries fell in the upper sub-major diagonal. It can also be seen that the variable songtypes containing heterospecific imitations as forinstance V5. V8 and Vl2 behave in the sameway as those without heterospecific imitations.
SONG OF THE STARLING 83
Above results also allow us to conclude thatStarling song bouts are more variable and versatile at the start, whereas the latter parts of a boutare much more predictable:1. most rattle and terminal song types are prece
ded and followed by only one song type whereas most whistles and several variable songtypes are preceded and/or followed by morethan one song type (Fig. 5);
2. a Starling song bout can start in several ways;3. a complete song bout contains mostly all the
rattle and terminal song types a male cansing, but never all the whistles and variablesong types.
DISCUSSION
Although the Starling's song is said to be a medley of a great many sounds (Gallacher 1978,Peterson et ai. 1983, Feare 1984), our study revealed that Starling song types are not sung independent from each other in a song bout. Bothtemporal and sequential analysis revealed someconsistent, species-specific characteristics in thesong bouts, whereas the temporal analysis alsorevealed some individual differences.
Like most songbird species, Starlings singmore than one version of the species-specificsong. When a bird has a repertoire of differentsong types, there are two extreme ways in whichthe repertoire can be presented during a singingperformance (Hartshorne 1956, 1973). Males mayrepeat a song type many times before introducinganother, what Hartshorne called "eventual variety", or they may sing with "immediate variety",where successive song types are different fromone another. We have clearly demonstrated thatStarlings sing with immediate variety. It will beuseful at this stage to recall how long and complex Starling song bouts are. They have an average duration of 28.5 sec and the longest was timedat 81 sec. On average a bout contains 34.2 songtypes, and the maximum found was 94. Time intervals between successive song types are verysmall. This combined with the rapid switching of
song types make that, when first heard, Starlingbouts seem to consist of an almost continuousdisplay of unrelated, random notes.
We were able to show, however, that Starlingsong bouts have a characteristic sequential pattern which might be an important species-specific feature. A bout typically begins with one orseveral whistles and ends with a series of highpitched notes, as has already been described byWitchell (1896): "... I allude to the high,squealing and almost toneless notes which, in thebreeding season, nearly always conclude itssong-phrases". The largest part of the repertoire,however, consists of variable song types whichfollow closely on the whistles and gradually passinto rattle song types. By making up a transitionmatrix of the sequencing of song types withinsong bouts of one bird, we were able to showthat Starlings are highly determinate singers. According to Slater (1973) this type of sequencing,when song types follow each other in a fixed order, is the simplest. In this case song types can bearranged in such an order that the upper sub-major diagonal contains most entries. Similar matrices have been found in the Mistle ThrushTurdus viscivorus by Isaac & Marler (1963) andin the Nightingale Luscinia megarhynchos byTodt (1971).
Although we found highly significant differences in bout length, the variation in repertoiresize was rather small. This can be due to the liemited sample size and/or to the fact that all3 males were probably of similar age (older than2 years). Male Starlings are namely thought to becapable of learning new song types at any timeof life (Krebs & Kroodsma 1980, Feare 1984)what probably means that repertoire size increases with age. The fact that every male has an unique repertoire of variable song types stronglysuggest that these are the result of improvisation(Marler & Peters 1982). From our sequential ana·lysis it appeared that variable song types containing heterospecific imitations behave in a similarway as those without imitations. This stronglysubscribes the view of Hindmarsh (1986) that vocal mimicry in Starlings probably has no func-
84 ARDEA 77 (1989)
tion Qther than the role it plays in the song as awhole.
Finally, there remains the question why theStarlings sings such long and complex songbouts. Cuthill & Hindmarsh (1985) demonstratedthat the function of song in the Starling is primarily concemed with mate attraction. If maleStarlings do sing in order to attract a mate, itmight be that larger repertoires and/or longersong bouts are more effective in doing so as hasbeen found for Sedge Warblers Acrocephatusschoenobaenus and Great Reed Warblers A.arundinaceus (Catchpole 1980, 1986). The Starling would be a particularly appropriate speciesto test the sexual selection hypothesis sincemales have a high variance in reproductive success due to the frequently occurring polygyny(Merkel 1980, Pinxten et at. 1986, 1988).
When there are such great differences instructure and in loudness between (categories of)song types as in the Starling, one may wonderwhether these all carry the same message. Thewhistles clearly stand out from the other 3 categories because they are also sung separately between song bouts. Furthermore, song matchingbetween neighbours occurs only with the whistled song (own obs.), suggesting that these couldbe important for social relationships betweenmales. Hausberger & Guyomarc'h (1981) andAdret-Hausberger (1984) showed that the relativefrequency of whistle types varies with the season, the time of day and the type of activity, whatcould mean that different whistles have differentfunctions.
Cuthill & Hindmarsh (1985) showed experimentally that unmated males sing longer songbouts than mated. In view of the results of oursequential analysis, this means that unmatedmales sing more often the highly stereotype ending of a bout. This includes the high-frequencysong types which are the loudest of a bout (Feare1984) and which are mostly accompanied by vigorous wing-waving (Witchell 1896, Verheyen1968, 1969, Feare 1984, own obs.). Since the mainfunction of wing-waving seems to be mate attraction, the difference in bout length between
mated and unmated males might indicate that thehigh-frequency song types play an important rolein mate attraction. Song activity decreases markedly at pairing in the Starling (Cuthill &Hindmarsh 1985, own obs.). Pinxten et at. (1987)showed that from 4 days prior to laying until thelaying of the second or third egg, males followtheir females wherever they go. During this period of mateguarding, the male stays close to hismate and often sings (Feare 1984, own obs.). Simulating the female might be an important function of song at this moment. During the breedingseason of 1984, we witnessed more than 30 copulations, all of which were preceded by song, indicating that it is important in inviting and stimulating the female to copulations. As the highfrequency song types are very rarely sung at thismoment, we suggest that the variable and rattlesong types, which can only be heard at close range, are hereby most important. That most copulations in the Starling are preceded by "low-intensity" song, has already been described by Hartby(1969). A more experimental approach is, however, necessary to test this hypothesis.
ACKNOWLEDGEMENTS
We wish to thank Marcel Lambrechts, Ranga BabuMyneni and Luc Bijnens for their suggestions andcomments on earlier versions of this paper. This studyis financed by a fellowship of the Instituut totAanmoediging van het Wetenschappelijk Onderzoekin Nijverheid en Landbouw to M. Eens while R.Pinxten is supported by a Research Assistantship ofthe Belgian National Fund for Scientific Research. K.Verbeeck drew the figures. We are also grateful to thetwo referees whose comments were useful to improvethis paper.
REFERENCES
Adret-Hausberger, M. 1982. Social influences on thewhistled songs of Starlings. Behav. Ecol.Sociobiol. 11: 241-246.
Adret-Hausberger, M. 1983. Variations dialectales dessifflements de l'etoumeau sansonnet sedentaire enBretagne. Z. Tierpsychol. 62: 55-71.
SONG OF THE STARLING 85
Adret-Hausberger, M. 1984. Seasonal quantitative variations of the whistled songs of Starlings. Ibis126: 372-378.
Berthold, P. 1964. Uber den Fortpflanzungszyklus stidwest-deutscher Stare und tiber bedingendeFaktoren der Brutreife beim Star. Vogelwarte 22:236-275.
Catchpole, C.K. 1980. Sexual selection and the evolution of complex songs among European warblersof the genus Acrocephalus. Behaviour 74: 149166.
Catchpole, C.K. 1982. The evolution of bird sounds inrelation to mating and spacing behaviour. In: D.E.Kroodsma & E.H. Miller (eds.). Acoustic communication in birds, Vol. 1. Academic Press, NewYork.
Catchpole, C.K. 1986. Song repertoires and reproductive success in the Great Reed Warbler Acrocephalus arundinaceus. Behav. Sociobio!. 19:439-445.
Cuthill, I. & A.M. Hindmarsh 1985. Increase in Starling song activity with removal of mate. Anim.Behav. 33: 326-328.
Davis, D.E. 1959. Territorial rank in Starlings. Anim.Behav. 7: 214-221.
Feare, C. 1984. The Starling. Oxford Univ. Press,Oxford.
Gallacher, H. 1978. De Spreeuw. Spectrum, Utrecht.Hartby, E. 1969. The calls of the Starling (Sturnus
vulgaris). Dansk. Om. Foren. Tidsskr. 62:205-230.
Hartshorne, C. 1956. The monotony-threshold in singing birds. Auk 83: 176-192.
Hartshorne, C. 1973. Born to sing. Indiana Univ.Press, Bloomington.
Hausberger, M. & J. C. Guyomarc'h 1981. Contribution it l'etude des voca1isations territoriales sifflees de l'etourneau sansonnet en Bretegne. Bio!.Behav. 6: 79-98.
Hindmarsh, A.M. 1984. Vocal mimicry in Starlings.Behaviour 90: 304-324.
Hindmarsh, A.M. 1986. The functional significance ofvocal mimicry in song. Behaviour 99: 87-100.
Isaac, D. & P. Marler 1963. Ordening of sequences ofsinging behaviour of Mistle Thrushes in relationto timing Anim. Behav. 11: 179-188.
Marler, P. & S. Peters 1982. Subsong and plastic song:their role in the vocal learning process. In: D.E.Kroodsma & E.H. Miller (eds.). Acoustic communication in birds, Vol. 2. Academic Press, NewYork.
Merkel, EW. 1980. Sozialverhalten von individuellmarkierten Staren Sturnus vulgaris in einer kleinen Nistkastenkolonie. III. Die Rolle der Polygynie. Luscinia 44: 133-158.
Peterson, R.T., G. Mountfort & PA.D. Hollom 1983.A field guide to the birds of Britain and Europe.Houghton Mifflin Company, Boston.
Pinxten, R., L. van Elsacker & R.E Verheyen 1986.Observations on polygyny in the Starling(Sturnus vulgaris). In: IEC Toulouse Volume.Behavior and the dynamics of animal populations. Privat of Toulouse, France.
Pinxten, R., L. van Elsacker & R.E Verheyen 1987.Duration and temporal pattern of mate guardingin the Starling. Ardea 75: 263-269.
Pinxten, R., M. Eens & R.E Verheyen 1988. An extreme case of polygyny in the European Starling.Bird Study.
Slater, P.J.B. 1973. Describing sequences of behaviour.In: P.P.G. Bateson & P.H. Klopfer (eds.). PlenumPress, New York & London.
Todt, D. 1971. Aquivalente und konvalente gesang1iche Reaktionen einer extrem regelmaBig singenden Nachtigall (Luscinia megarhynchos). Z. verg.Psycho!. 71: 262-285.
Verheyen, R.E 1968. De baits en copulatie van deSpreeuw, Sturnus v. vulgaris L. Gerfaut 58: 239259.
Verheyen, R.E 1968. Bijdrage tot de ethologie en deoecologie van de Spreeuw, Sturnus v. vulgaris L.,gedurende de voortp1antingsperiode. Ph. D. Thesis. R.U. Gent.
Witchell, C.A. 1986. The evolution of bird song.Adam & Charles Black, London.
SAMENVATTING
De verschillende elementen van de spreeuwenzang, de tijdsduur ervan en de tijdsduur van deintervallen werden geanalyseerd. Uit deze analyse bleek dat mannetjes zeer lange en complexezangbeurten zingen. Een zangbeurt bestaat uiteen snelle opeenvolging van een groot aantalverschillende zangtypes. Mannetjesspreeuwenzingen met 'onmiddellijke variatie'. Er bestaanduidelijke individuele verschillen tussen de mannetjes in hun gemiddelde zangbeurtlengte en inhun 'singing rate'.
Spreeuwenzangbeurten hebben een karakteristieke sequentiele organisatie. Ze zijn samengesteld uit 4 categorieen van zangtypes. Een zang-
86 ARDEA 77 (1989)
beurt begint nagenoeg altijd met een of meerfluittonen. Deze fluittonen worden gevolgd dooreen groot aantal variabele zangtypes die, op hunbeurt, geleidelijk overgaan in ratel zangtypes.Een zangbeurt eindigt gewoonlijk met een serievan hoge-frequentie zangtypes. Een gedetailleerde sequentiele analyse van een mannetje toondeaan dat de spreeuw een zeer gedetermineerdezanger is. Dat wil zeggen dat de verschillende
zangtypes elkaar niet willekeurig opvolgen, maareen soort-specifiek patroon volgen. Repertoirgrootte varieerde van 43 tot 48 zangtypes. Dezang van de Spreeuw is meer gevarieerd bij deaanvang van een zangbeurt dan aan het einde.
Alhoewel de belangrijkste functie van despreeuwenzang wellicht het aanlokken van wijfjes is, wordt er gesuggereerd dat verschillendezangtypes verschillende functies vervullen.