Journal of Theoretical Biology 233 (2005) 271–286 Analysis of the electroencephalographic activity associated with thalamic tumors S.C. O’Connor a,b,, P.A. Robinson a,b a School of Physics, University of Sydney, Broadway, Sydney, NSW 2006, Australia b Brain Dynamics Center, Westmead Hospital, Westmead, NSW 2145, Australia Received 14 June 2004; accepted 7 October 2004 Available online 30 November 2004 Abstract A physiologically based model of corticothalamic dynamics is used to investigate the electroencephalographic (EEG) activity associated with tumors of the thala mus. Tumor activity is mode led by introducing localized two-dimensi onal spatial non- uniformities into the model parameters, and calculating the resulting activity via the coupling of spatial eigenmodes. The model is able to reproduce various qualitative features typical of waking eyes-closed EEGs in the presence of a thalamic tumor, such as the app earance of abn ormal pea ks at the ta (E3Hz) and spindl e (E12Hz) frequenci es, the attenuation of normal eyes-c losed background rhythms, and the onset of epileptic activity, as well as the relatively normal EEGs often observed. The results indicate that the abnormal activity at theta and spindle frequencies arises when a small portion of the brain is forced into an over-inhibited state due to the tumor, in which there is an increase in the firing of (inhibitory) thalamic reticular neurons. The effect is heightened when there is a concurrent decrease in the firing of (excitatory) thalamic relay neurons, which are in any case inhibited by the reticular ones. This is likely due to a decrease in the responsiveness of the peritumoral region to cholinergic inputs from the brainstem, and a corresponding depolarization of thalamic reticular neurons, and hyperpolarization of thalamic relay neurons, similar to the mechanism active during slow-wave sleep. The results indicate that disruption of normal thalamic activity is essential to generate these spectral peaks. Furthermore, the present work indicates that high-voltage and epileptiform EEGs are caused by a tumor-induced local over-excitation of the thalamus, which propagates to the cortex. Experimental findings relating to local over- inhibition and over-excitation are discussed. It is also confirmed that increasing the size of the tumor leads to greater abnormalities in the observable EEG. The usefulness of EEG for localizing the tumor is investigated. r 2004 Elsevier Ltd. All rights reserved. Keywords: Thalamic tumor; EEG; Pathological theta or delta; Continuum model 1. Introd uctio n The disruption to normal brain function induced by a tumor of the thalamus can be detected by electroence- phal ograms (EEGs), whic h measure elect rical activ ity via electrodes on the head. A typical feature of waking EEG in the presence of thalamic tumors, is unusually large activity near 3 Hz (Gibbs and Gibbs, 1964; Hirose et al., 1975; Janati and Hester, 1986; Newmark et al., 1983). This frequency is variously called ‘delta’ or ‘theta’ in thalamic literature; in the present work we refer to it as th e theta fr eque ncy. Ot he r EEG corr el ates of thalamic tumor include the focal attenuation of normal backgr ound rhythms (Jan ati and Hes ter , 198 6; New- mark et al., 1983), epileptiform discharges ( Cheek and Taveras, 1966; Jana ti and Hester, 1986 ; Hirose et al., 1975; O’Brien et al., 1997), non-reactive alpha (Janati and Hes ter, 1986), an d diff use (Hirose et al., 1975; Newmark et al., 1983) or focal (Scarff and Rahm, 1941) backgr ound slowin g. There have also been repor ts of12–14 Hz ‘spindl es’ during wakin g EEG (Hirose et al., ARTICLE IN PRESS www.elsevier.com/locate/yjtbi 002 2-51 93/$- see front matte r r 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.jtbi.2004.10.009 Corresponding author. School of Physics, CUDOS, University ofSydney, Sydney NSW 2006, Australia. Te l.: +612 93515635 ; fax: +61 2 9351 7726. E-mail address : [email protected] (S.C. O’Connor).
Analysis of the electroencephalographic activity associated with
thalamic tumors
S.C. O’Connora,b,, P.A. Robinsona,b
aSchool of Physics, University of Sydney, Broadway, Sydney, NSW 2006, AustraliabBrain Dynamics Center, Westmead Hospital, Westmead, NSW 2145, Australia
Received 14 June 2004; accepted 7 October 2004
Available online 30 November 2004
Abstract
A physiologically based model of corticothalamic dynamics is used to investigate the electroencephalographic (EEG) activity
associated with tumors of the thalamus. Tumor activity is modeled by introducing localized two-dimensional spatial non-
uniformities into the model parameters, and calculating the resulting activity via the coupling of spatial eigenmodes. The model is
able to reproduce various qualitative features typical of waking eyes-closed EEGs in the presence of a thalamic tumor, such as the
appearance of abnormal peaks at theta (E3Hz) and spindle (E12Hz) frequencies, the attenuation of normal eyes-closed
background rhythms, and the onset of epileptic activity, as well as the relatively normal EEGs often observed. The results indicate
that the abnormal activity at theta and spindle frequencies arises when a small portion of the brain is forced into an over-inhibited
state due to the tumor, in which there is an increase in the firing of (inhibitory) thalamic reticular neurons. The effect is heightened
when there is a concurrent decrease in the firing of (excitatory) thalamic relay neurons, which are in any case inhibited by the
reticular ones. This is likely due to a decrease in the responsiveness of the peritumoral region to cholinergic inputs from the
brainstem, and a corresponding depolarization of thalamic reticular neurons, and hyperpolarization of thalamic relay neurons,
similar to the mechanism active during slow-wave sleep. The results indicate that disruption of normal thalamic activity is essential
to generate these spectral peaks. Furthermore, the present work indicates that high-voltage and epileptiform EEGs are caused by a
tumor-induced local over-excitation of the thalamus, which propagates to the cortex. Experimental findings relating to local over-
inhibition and over-excitation are discussed. It is also confirmed that increasing the size of the tumor leads to greater abnormalities
in the observable EEG. The usefulness of EEG for localizing the tumor is investigated.
r 2004 Elsevier Ltd. All rights reserved.
Keywords: Thalamic tumor; EEG; Pathological theta or delta; Continuum model
1. Introduction
The disruption to normal brain function induced by a
tumor of the thalamus can be detected by electroence-phalograms (EEGs), which measure electrical activity
via electrodes on the head. A typical feature of waking
EEG in the presence of thalamic tumors, is unusually
large activity near 3 Hz (Gibbs and Gibbs, 1964; Hirose
et al., 1975; Janati and Hester, 1986; Newmark et al.,
1983). This frequency is variously called ‘delta’ or ‘theta’
in thalamic literature; in the present work we refer to it
as the theta frequency. Other EEG correlates of thalamic tumor include the focal attenuation of normal
background rhythms (Janati and Hester, 1986; New-
mark et al., 1983), epileptiform discharges (Cheek and
Taveras, 1966; Janati and Hester, 1986; Hirose et al.,
1975; O’Brien et al., 1997), non-reactive alpha (Janati
and Hester, 1986), and diffuse (Hirose et al., 1975;
Newmark et al., 1983) or focal (Scarff and Rahm, 1941)
background slowing. There have also been reports of
12–14 Hz ‘spindles’ during waking EEG (Hirose et al.,
ARTICLE IN PRESS
www.elsevier.com/locate/yjtbi
0022-5193/$- see front matter r 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.jtbi.2004.10.009
Corresponding author. School of Physics, CUDOS, University of
Sydney, Sydney NSW 2006, Australia. Tel.: +612 93515635; fax:
observable signals (Nunez, 1995; O’Connor and Robin-
son, 2003). Thus, in the absence of skull volume
conduction, the power spectrum on the head is wellapproximated by the squared modulus of the signal fe;to within a constant of proportionality. The effects on
the spectrum of conduction through the cerebrospinal
fluid, skull, and scalp, have been studied in the context
of this model (O’Connor et al., 2002; Robinson et al.,
2001a); they filter out high-wavenumber activity, k
\15 m1, (O’Connor et al., 2002; Robinson et al.,
2001a) and hence high-frequency activity (via the
dispersion relation). Thus, at the frequencies of interest
in the present work,o20 Hz, the effects are minimal and
are not considered further.
In our continuum treatment of the cortex the firingrate of signals emitted by excitatory or inhibitory
neurons, which depend on individual cell body poten-
tials, are averaged to give mean values of the outgoing
pulse field faðr; tÞ; where a ¼ e; i : The mean rate of
generation of neuronal pulse density depends on the
mean local cell-body potential via a smooth sigmoidal
function that increases from 0 to its maximum value as
the potential increases from 1 to 1: We approximate
the sigmoidal function here by a linear function on the
assumption that deviations from the steady state are
small in normal, non-seizure states. This approximation
has been found to yield excellent agreement with
observed frequency spectra and other phenomena
(O’Connor et al., 2002; O’Connor and Robinson,
2003, 2004b, c; Rennie et al., 2002; Robinson et al.,
2001a, 2002, 2003; Robinson, 2003).
The local mean cell body potential of neurons of type
a in the cortex is a function of inputs from other cortical
neurons, and from excitatory subcortical neurons.Incoming activity is received in the dendritic tree and
filtered as it spreads along the dendrites to the cell body.
The quantity Lab is a dendritic low-pass filter function
which accounts for the temporal delay and smearing of
an incoming signal from a neuron of type b ¼ e; i ; s as it
travels along the dendritic tree to the cell body of a
neuron of type a ¼ e; i : It can be written (Rennie et al.,
1999)
Labðr;oÞ ¼ 1
½1 io=aabðrÞ½1 io=babðrÞ; (1)
where bab and aab are the inverse rise and decay times of the dendritic potential, respectively.
Outgoing pulses from each neuron propagate along
its axonal tree at a velocity v(r)E1 0 m s1. This
propagation can be described by damped wave equa-
tions for the fields fa (Rennie et al., 1999). After Fourier
transforming in time, one finds,
Daðr;oÞfaðr;oÞ ¼X
b
J abðr;oÞfbðr;oÞ; (2)
where
Daðr;oÞ ¼ ½1 io=gaðrÞ2 r2ar 2; (3)
J abðr;oÞ ¼ Labðr;oÞG abðrÞeiotabðrÞ; (4)
gaðrÞ ¼ vðrÞ=ra is a measure of the damping, ra is the
mean range of axons a, the gain G ab represents the
scaled response strength in neurons a due to a unit signal
incident from neurons of type b, and tab represents a
pure delay—as in signal transmission between the cortex
and the thalamus, for example—and appears as an
exponent due to the temporal Fourier transform. Note
that (2) makes explicit the approximately one-to-one
mapping between locations in the cortex and thalamus,
particularly the specific nuclei of the thalamus.
Using (2) and the connectivities shown in Fig. 1, thewave equation for excitatory cortical neurons follows as
Deðr;oÞfeðr;oÞ ¼ J eeðr;oÞfeðr;oÞ þ J ei ðr;oÞfi ðr;oÞ
þ J esðr;oÞfsðr;oÞ: ð5Þ
The analogous equations for cortical inhibitory, specific
relay, and reticular neurons can also be deduced from
(2), and the quantities fi ; fs; and fr can be eliminated to
give the transfer function of a stimulus fn to fe
(Robinson et al., 2003). This is of the form
Aðr;oÞfeðr;oÞ ¼ B ðr;oÞfnðr;oÞ; (6)
ARTICLE IN PRESS
φe, φ
i
φs
φn
φr
φe
reticularnucleus
cortex
relaynuclei
ee,ei
es
rs re
srse
sn
Fig. 1. Schematic of corticothalamic connections showing the cortex,
reticular nucleus, and relay nuclei. The cortex is extensively connected
to itself, and also projects to and receives projections from the
thalamus. There are two loops through the thalamus: a direct loop
passing only through the relay nuclei, and an indirect loop which also
passes through the reticular nucleus. There is also an intrathalamic
loop. Locations at which gains G ab act are indicated on the diagram.
The uniform case previously studied is a special case.
Activity in this model is relatively insensitive to precise
boundary conditions (O’Connor and Robinson, 2004a;
Robinson et al., 2001b; Robinson, 2003), so we
investigate a simple cortical geometry in the first
instance. Taking the Fourier transform in space, and
applying periodic boundary conditions on a rectangular
cortex of size l x l y, the expression (6) becomes
XK
Aðk K;oÞfeðK;oÞ ¼X
KB ðk K;oÞfnðK;oÞ;
(9)
where k and K range over the values kmj ; Kmj ¼
ð2pm=l x; 2p j =l yÞ; and m and j are integers. Only a finite
number of modes can be included in the calculation, and
most of the activity can actually be captured using a
relatively small number of modes (Robinson et al.,
2003); thus we choose M max such that jmj; j j jpM max:Eq. (9) can be written as a matrix equation AUe ¼ BUn;or Ue ¼ A1BUn ¼ MUn; where the size of each matrix
depends on the number of modes M max retained after
truncation [for the 2D cortex studied here, A and B are(2M max+1)2 (2M max+1)2 matrices and Ue is a
(2M max+1)2 1 column matrix] (Robinson et al.,
2003). We have shown previously (Robinson et al.,
2003) that the power spectrum at a given r is given by
P ðr;oÞ ¼ jfnðoÞ2jXm;n
exp½iðkm knÞ rðMMyÞmn (10)
for spatially white noise, where m and n label matrix
elements. By averaging (10) over position, the mean
power can be written
P ðoÞ ¼ jfnðoÞ2jTrðMMyÞ; (11)
where fn is independent of o if the noise is also
temporally white.
2.1. Reduced parameter space
In this section, we give a brief overview of stability in
the model as it pertains to the present work. In previous
work using the uniform model (i.e. with spatially
uniform parameters) to analyse normal arousal
states and epileptic seizures, we found that stability
boundaries in parameter space occur approximately
where (Robinson et al., 2002)
0 ¼ ð1 io=geÞ2 x yð1 G srsÞ
1 G srsL2 eiot0 ; (12)
x ¼ G ee=ð1 G ei Þ; (13)
y ¼ G ese þ G esre
ð1 G srsÞð1 G ei Þ; (14)
is satisfied, where for brevity we have written G srs ¼
G srG rs; G ese ¼ G esG se; and G esre ¼ G esG srG re; and t0 ¼
2tes ¼ 2tse ¼ 2tre is the corticothalamic loop propaga-
tion delay. The quantities x and y relate to cortical and
corticothalamic activity, respectively, and
z ¼ G srsab=ða þ bÞ2; (15)
parametrizes intrathalamic activity; thus, the system can
be approximately parametrized in a reduced three-
dimensional (xyz) space. The stability zone in xyz spacedefined by Eq. (12) is shown in Fig. 2, for the eyes-closed
parameters used throughout this work. The alpha
stability boundary is indicated on the upper right of
the figure. Proximity to this boundary manifests itself in
the spectrum as increased activity at the alpha
frequency, since cortical activity approaches instability
and hence the maximum firing rates typical of seizures.
Indeed, if the boundary is crossed, the brain goes into a
limit cycle near 10 Hz, which is plausibly related to
seizure activity, via an instability of the alpha peak
(Robinson et al., 2002). Also shown is the slow-wave
boundary (the front unshaded surface), through which
the brain passes into slow-wave (o1 Hz) instability. This
ARTICLE IN PRESS
EO EC
S2
S4
z
1.0
y1.0
-1.0
x
1.0
theta
spindle
alpha
delta
Fig. 2. Stability zone in xyz space (Robinson et al., 2002) for the
parameters in Table 1. The surface is shaded according to instability
type: the transparent front right face corresponds to a zero-frequency
instability; the top light-shaded right face corresponds to alpha-
frequency (E10 Hz) instability; the top central surface corresponds to
a spindle-frequency (E13 Hz) instability; and the top light-shaded left
face corresponds to a theta-frequency (E3 Hz) instability. Approx-
imate locations are shown of eyes-open (EO), eyes-closed (EC), and
normal sleep stages two (S2) and four (S4), with each state located at
where P (X , Y ) takes a value P 0+P 1 at the tumor
centroid or ‘focus’ (X 1, Y 1), and the Gaussian has a
characteristic width of s: In the present work, we take
the background, or nominal value P 0 of each parameter
to be that for a waking, eyes-closed state; the parameters
have been extensively studied using the spatially uniform
model, and are consistent with anatomical and physio-
logical data (Robinson et al., 2004). Position in the two
dimensions is labeled with X and Y to avoid confusion
with the state parameters x, y, and z.In the following, we take as the linear dimension of
the brain l x=l y=0.8 m, the approximate circumference
of the brain, which was determined by scaling the head
circumference to account for cortical convolutions
(Nunez, 1981). We take the front of the head to
correspond to Y ¼ 0 m; and the back to Y ¼ 0:4 m: If
we consider a thalamic tumor that affects an area of
cortex with width s; the fraction of the cortex affected is
of order pð2s=l xÞ2; where we assume most of the affected
area lies within 2s of the tumor center. For s ¼ 0:03 m;the tumor affects less than 2% of the cortex, and for
s ¼ 0:05 m; the tumor affects around 5% of the cortex.
Thalamic tumors are particularly well suited to being
modeled in this way, since the tumor itself merely
perturbs cortical activity through distorted thalamocor-
tical projections, as seen in Fig. 3(a). Figs. 3(c) and (d)
represent schematic top views of the cortex, for thalamic
and cortical tumors, respectively. Panel (c) shows that
corticocortical fibers remain intact, albeit damaged, in
the presence of a thalamic tumor; the tumor itself does
not project to the cortex and we need therefore model
only the surrounding, damaged tissue. Cortical tumors,on the other hand, represent a region of complete
electrical silence in the cortex itself, as shown by the
black region in Fig. 3(d). The long-range corticocortical
fibers are disrupted, making the modeling of cortical
tumors significantly more complicated than modeling
subcortical ones. We therefore restrict our analysis to
thalamic tumors in the present work. The mappings
shown in Fig. 3 are deduced from experimental findings:
in tumors which invade the cortex, a zone of absolute
electrical silence in the region invaded by the tumor has
been observed (Hess, 1975; Hirsch et al., 1966). This
region is surrounded by an annulus in which normal
ARTICLE IN PRESS
side view top view
cortex
thalamus
damaged
tissue
damaged
tissue
tumor
projection of
damaged tissue
cortex
cortex
thalamus
tumor
cortex
tumor
damaged tissue
(a) (c)
(b) (d)
Fig. 3. Schematic diagram of the corticothalamic and corticocortical connections in the region near a tumor. Cross-sections through the 2D tumor
are shown from a side view (first column) and a top view (second column). Damaged connections are indicated by broken lines, or by a shaded
region; undamaged connections are indicated by unbroken lines, or by a white region. Panel (a) represents a thalamic tumor, in which the tumor itself
does not project to the cortex; intracortical connections remain intact. Panel (b) represents a cortical tumor; intracortical connections are damaged or
absent, and the tumor is more difficult to model. Panel (c) represents a top view of the cortex in the case of a thalamic tumor; a circle of cortical tissue
is damaged. Panel (d) represents a top view of the cortex in the case of a cortical tumor; corticocortical connections are absent at the tumor (black
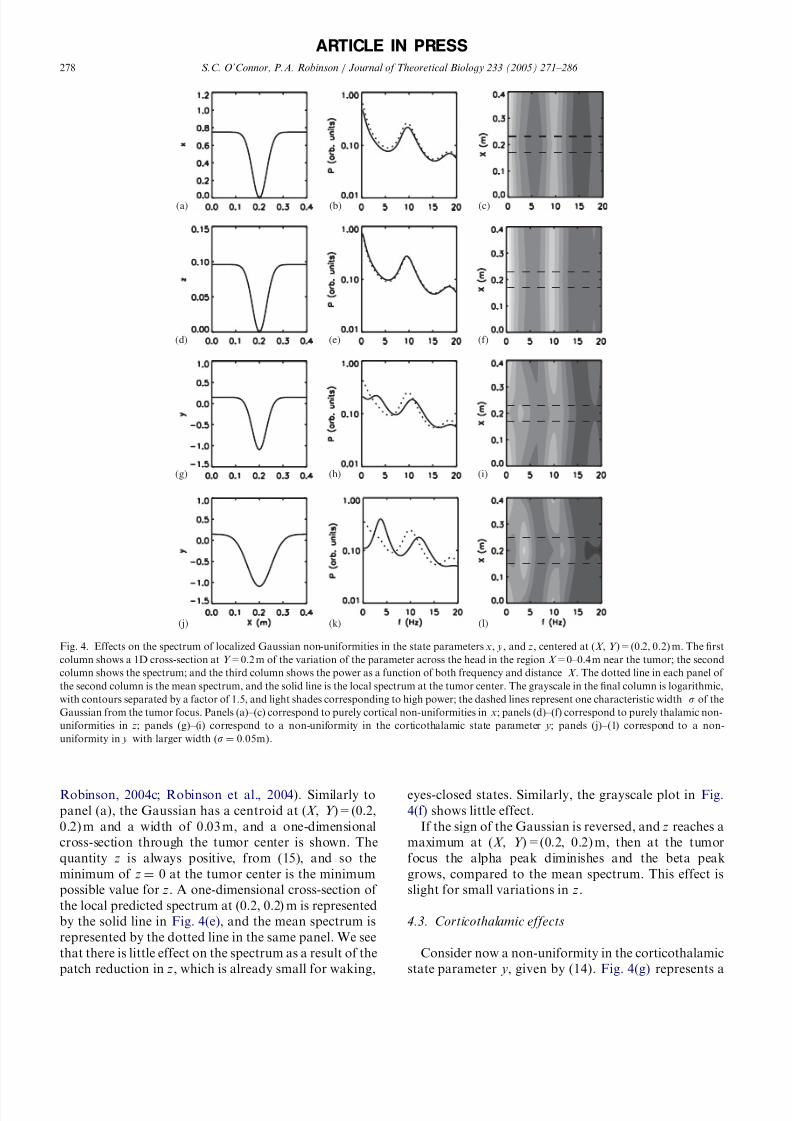
Robinson, 2004c; Robinson et al., 2004). Similarly to
panel (a), the Gaussian has a centroid at (X , Y )=(0.2,
0.2) m and a width of 0.03 m, and a one-dimensional
cross-section through the tumor center is shown. The
quantity z is always positive, from (15), and so the
minimum of z ¼ 0 at the tumor center is the minimum
possible value for z. A one-dimensional cross-section of
the local predicted spectrum at (0.2, 0.2) m is represented
by the solid line in Fig. 4(e), and the mean spectrum is
represented by the dotted line in the same panel. We see
that there is little effect on the spectrum as a result of the
patch reduction in z, which is already small for waking,
eyes-closed states. Similarly, the grayscale plot in Fig.
4(f) shows little effect.
If the sign of the Gaussian is reversed, and z reaches a
maximum at (X , Y )=(0.2, 0.2) m, then at the tumor
focus the alpha peak diminishes and the beta peak
grows, compared to the mean spectrum. This effect is
slight for small variations in z.
4.3. Corticothalamic effects
Consider now a non-uniformity in the corticothalamic
state parameter y, given by (14). Fig. 4(g) represents a
ARTICLE IN PRESS
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
(j) (k) (l)
Fig. 4. Effects on the spectrum of localized Gaussian non-uniformities in the state parameters x, y, and z, centered at (X , Y )=(0.2, 0.2) m. The first
column shows a 1D cross-section at Y =0.2 m of the variation of the parameter across the head in the region X =0–0.4 m near the tumor; the second
column shows the spectrum; and the third column shows the power as a function of both frequency and distance X . The dotted line in each panel of
the second column is the mean spectrum, and the solid line is the local spectrum at the tumor center. The grayscale in the final column is logarithmic,
with contours separated by a factor of 1.5, and light shades corresponding to high power; the dashed lines represent one characteristic width s of the
Gaussian from the tumor focus. Panels (a)–(c) correspond to purely cortical non-uniformities in x; panels (d)–(f) correspond to purely thalamic non-
uniformities in z; panels (g)–(i) correspond to a non-uniformity in the corticothalamic state parameter y; panels (j)–(1) correspond to a non-
one-dimensional cross-section through Y =0.2 m of the
variation of y over the head where, as above, the patch is
a Gaussian localized at (X , Y )=(0.2, 0.2) m, with a
width of 0.03m, and the results are shown over
X =0–0.4 m. The background value of y 0:15 is typical
of the waking, eyes-closed state (O’Connor and Robin-
son, 2004c; Robinson et al., 2004). We reduce y to avalue of 1:1 at the patch center in Fig. 4(g), by
reducing G ese and increasing |G esre|; these parameters are
studied individually later in this subsection. Fig. 4(h)
shows the local spectrum at the tumor focus (solid line),
and the mean spectrum across the head (dotted line) for
the non-uniformity shown in panel (g). The local
spectrum differs in several important ways from the
mean spectrum: the background activity is attenuated at
low frequencies, and at alpha and beta frequencies; there
is an additional peak in the theta range near 3 Hz (often
called the delta range); and, the alpha peak is shifted to a
higher frequency. These effects are quite localized, as
seen from the grayscale plot in Fig. 4(i).
We now increase the size of the tumor, as seen in Fig.
4(j), where that the affected cortical area has a width of
0.05 m, which affects around 5% of the cortex. Fig. 4(h)
shows that there is a peak near 12 Hz, which lies in the
spindle frequency range. The amplitude of the theta
peak near 3 Hz also increases as the size of the tumor
increases. Figs. 4(k) and (l) show the large peak near
3 Hz, the attenuation of background frequencies, and
the peak in the spindle range, near 12 Hz. With reference
to the discussion in Section 2.1, we note that a localized
change in the sign of y would take that local portion of
the brain into a region of the stability zone in Fig. 2which is normally associated with sleep states; hence the
appearance of the typical sleep peaks near 3 and 12 Hz.
We explore the possibility that the 12Hz rhythm is
‘fast alpha’, rather than a spindle, in conjunction with
Fig. 6 below. This is also discussed thoroughly in
Section 6.
Consider first the effects on the spectrum of changing
the sign of the Gaussian patch, so that y takes amaximum at the patch focus. From Fig. 2, we see that
increasing y takes the brain nearer both the alpha and
slow-wave stability boundaries; indeed, in such a case we
see a corresponding increase in the power at alpha and
low frequencies. If the increase in y takes the brain
outside the stability boundaries in Fig. 2 then the brain
becomes unstable at those frequencies. This mechanism
could account for the epileptiform discharges sometimes
observed in the EEGs of subjects with a thalamic tumor
(Cheek and Taveras, 1966; Hirose et al., 1975; Janati
and Hester, 1986; Robinson et al., 2002), and is
discussed further in Section 6.
From (14) the sign of y depends only on the relative
magnitudes of G ese and G esre, since G srs and G ei are
always negative. We now examine the effects of G ese and
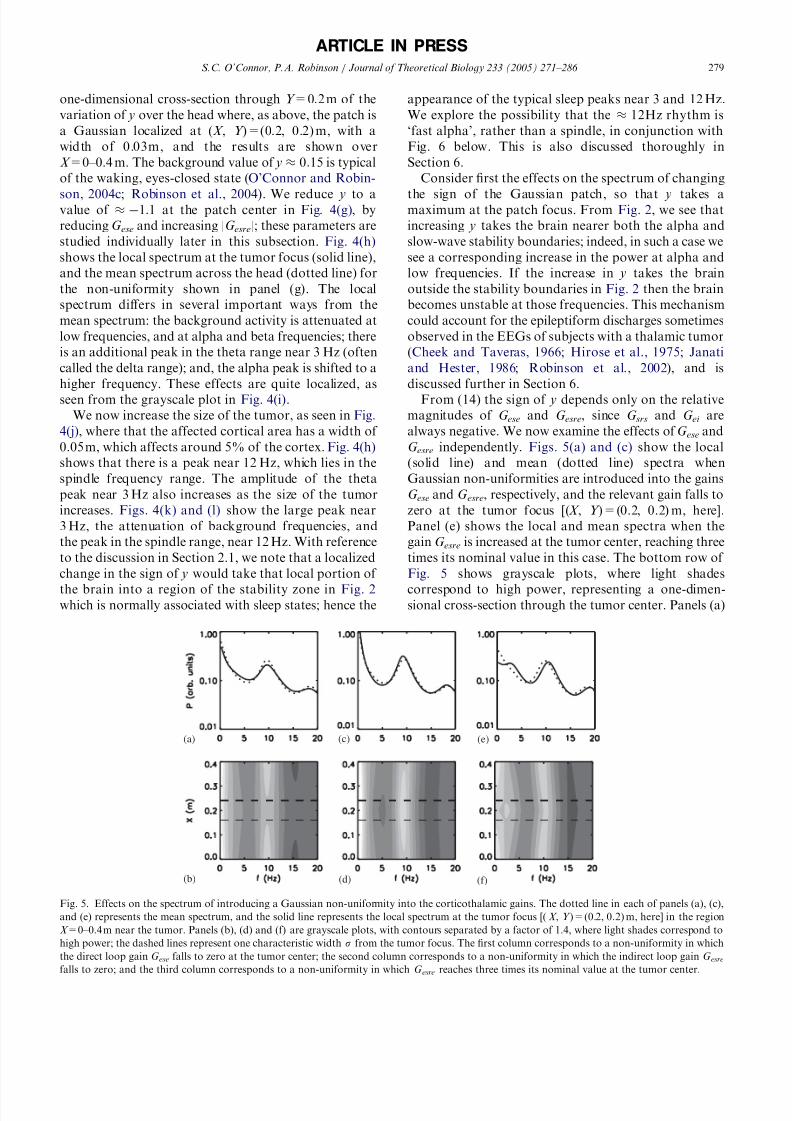
G esre independently. Figs. 5(a) and (c) show the local
(solid line) and mean (dotted line) spectra when
Gaussian non-uniformities are introduced into the gains
G ese and G esre, respectively, and the relevant gain falls to
zero at the tumor focus [(X , Y )=(0.2, 0.2) m, here].
Panel (e) shows the local and mean spectra when the
gain G esre is increased at the tumor center, reaching three
times its nominal value in this case. The bottom row of
Fig. 5 shows grayscale plots, where light shades
correspond to high power, representing a one-dimen-sional cross-section through the tumor center. Panels (a)
ARTICLE IN PRESS
(a)
(b)
(c) (e)
(d) (f)
Fig. 5. Effects on the spectrum of introducing a Gaussian non-uniformity into the corticothalamic gains. The dotted line in each of panels (a), (c),
and (e) represents the mean spectrum, and the solid line represents the local spectrum at the tumor focus [( X , Y )=(0.2, 0.2) m, here] in the region
X =0–0.4 m near the tumor. Panels (b), (d) and (f) are grayscale plots, with contours separated by a factor of 1.4, where light shades correspond to
high power; the dashed lines represent one characteristic width s from the tumor focus. The first column corresponds to a non-uniformity in which
the direct loop gain G ese falls to zero at the tumor center; the second column corresponds to a non-uniformity in which the indirect loop gain G esrefalls to zero; and the third column corresponds to a non-uniformity in which G esre reaches three times its nominal value at the tumor center.
and (b) show that the reduction in G ese and hence y, and
consequently increased distance from the slow-wave and
alpha stability boundaries indicated in Fig. 2, produces
a focal attenuation of background activity, similar to
that in Figs. 4(h) and (k). In panels (c) and (d), we see
the opposite effect: the reduction in |G esre| and hence
increase in y, and consequently increased proximity tothe slow-wave and alpha stability boundaries indicated
in Fig. 2, produces a focal increase of background
rhythms. Furthermore, there is a slight downward shift
of the alpha peak frequency in panel (c), or ‘background
slowing’, due to disruption of the propagation delay in
the corticothalamic pathway. In panels (e) and (f), there
is a focal theta (E3 Hz) peak, and a focal shift in the
alpha peak towards the spindle frequency. We thus see
that reducing G ese attenuates the background peaks by
flattening the spectrum, and increasing |G esre| introduces
the abnormal theta peak. The latter effect is due to the
sign change of the state parameter y and consequent
localized over-inhibited state.
The spectral effects of non-uniformities in the
parameters G ei and G srs are more complicated, since
they affect more than one of the state parameters x, y,
and z. In general, decreasing |G ei | increases both x and y,
and hence gives higher power at spectral peaks.
Decreasing |G srs| increases y but decreases z, from Eqs.
(14) and (15), so its effect depends on the location in the
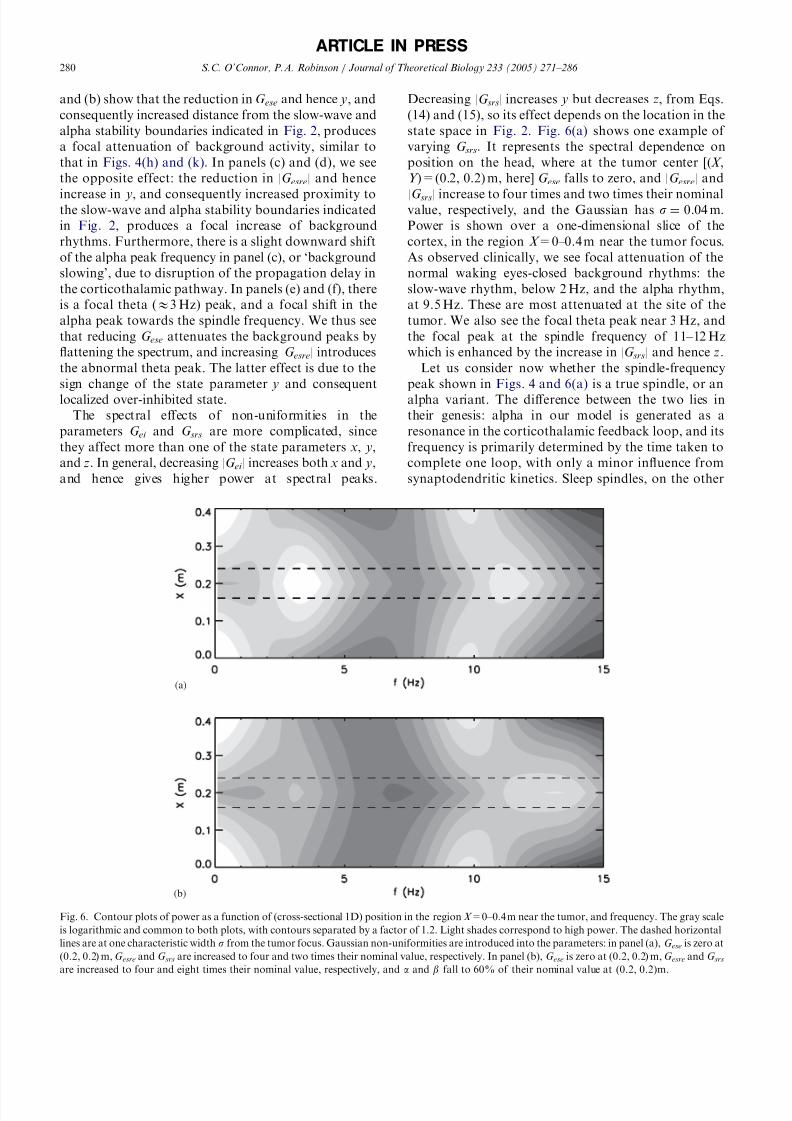
state space in Fig. 2. Fig. 6(a) shows one example of
varying G srs. It represents the spectral dependence on
position on the head, where at the tumor center [(X ,
Y )=(0.2, 0.2) m, here] G ese falls to zero, and |G esre| and
|G srs| increase to four times and two times their nominalvalue, respectively, and the Gaussian has s ¼ 0:04 m:Power is shown over a one-dimensional slice of the
cortex, in the region X =0–0.4 m near the tumor focus.
As observed clinically, we see focal attenuation of the
normal waking eyes-closed background rhythms: the
slow-wave rhythm, below 2 Hz, and the alpha rhythm,
at 9.5 Hz. These are most attenuated at the site of the
tumor. We also see the focal theta peak near 3 Hz, and
the focal peak at the spindle frequency of 11–12 Hz
which is enhanced by the increase in |G srs| and hence z.
Let us consider now whether the spindle-frequency
peak shown in Figs. 4 and 6(a) is a true spindle, or an
alpha variant. The difference between the two lies in
their genesis: alpha in our model is generated as a
resonance in the corticothalamic feedback loop, and its
frequency is primarily determined by the time taken to
complete one loop, with only a minor influence from
synaptodendritic kinetics. Sleep spindles, on the other
ARTICLE IN PRESS
(a)
(b)
Fig. 6. Contour plots of power as a function of (cross-sectional 1D) position in the region X =0–0.4 m near the tumor, and frequency. The gray scale
is logarithmic and common to both plots, with contours separated by a factor of 1.2. Light shades correspond to high power. The dashed horizontal
lines are at one characteristic width s from the tumor focus. Gaussian non-uniformities are introduced into the parameters: in panel (a), G ese is zero at
(0.2, 0.2) m, G esre and G srs are increased to four and two times their nominal value, respectively. In panel (b), G ese is zero at (0.2, 0.2) m, G esre and G srsare increased to four and eight times their nominal value, respectively, and a and b fall to 60% of their nominal value at (0.2, 0.2)m.