Gene, 84 (1989) 65-72 Elsevier 65 GENE 03 197 The block to transcription elongation at the SV40 attenuation site is decreased in vitro by oligomers complementary to segments of the attenuator RNA (Recombinant DNA; RNA secondary structure; viral transcription complexes; synthetic oligodeoxyribo- nucleotides) Mark Kessler and Yosef Aloni Department of Molecular Genetics and Virology, Weizmann Institute of Science, Rehovot 76100 (Israel) Received by A. Kohn: 11 April 1989 Revised: 6 June 1989 Accepted: 12 June 1989 SUMMARY We have previously reported that a mechanism resembling attenuation in prokaryotes regulates simian virus 40 (SV40) late gene expression. We have suggested that modulation of the attenuator RNA secondary structure is an integral element regulating the elongation block at the attenuation site [Hay et al., Cell 29 (1982) 183-1931. In the present study, oligodeoxyribonucleotides (oligos), 13-19 nucleotides long, were used to probe the involvement of the attenuator RNA secondary structure in the control of elongation block at the SV40 attenuation site. These oligos are complementary to segments of the attenuator RNA suggested to play a role in the regulation of attenuation. The oligos were added to an in vitro transcription reaction containing SV40 transcription complexes, and their effect on transcription through the attenuation site was measured. As predicted, the three oligos caused specific decreases in the elongation block at the SV40 attenuation site. These results provide direct evidence for the involvement of RNA secondary structure in the attenuation mechanism in SV40. INTRODUCTION The quantity of a specific eukaryotic mRNA in the cell is thought to be primarily regulated at the level of transcription initiation, RNA processing and stability of the mature message. It is now becoming increasingly apparent that the quantity of mRNA is also determined at the level of transcription elon- gation (Aloni and Hay, 1985). A block to transcrip- tion elongation, termed attenuation, has been shown to regulate the quantity of mRNA of several animal viruses, including SV40 (Hay et al., 1982; Skolnik- Correspondenceto: Dr. Y. Aloni, Department of Genetics, Weizmann Institute of Science, Rehovot 76100 (Israel) Tel. 8-482207/8-483680; Fax (972) 8-466966. nucleotide triphosphate(s); oligo, oligodeoxyribonucleotide; PAGE, polyacrylamide-gel electrophoresis; p.i., post infection; SV40, simian virus 40; TC buffer, 40 mM Hepes-NaOH pH 8.0/5 mM KC1/0.15 M (NH&SO,; TBE, 90 mM Tris . borate Abbreviations: Ad2, adenovirus type 2; Att, attenuator(ed)(ion); pH 8.3/90 mM boric acid/2mM EDTA; TC, 40 mM bp, base pair(s); CTP, cytidine 5’-triphosphate; dA, Hepes-NaOH pH 8.0/S mM KCljO.15 M (NH&SO,; tsp, tran- deoxyadenosine; DTT, dithiothreitol; HIV-l, human immuno- scription start point(s); VTC, viral transcription complexes; deficiency virus 1; NP-40, nonidet P-40; nt, nucleotide(s); NTP, WCE, whole cell extract; wt, wild type. 0378-l 119/89/$03.50 0 1989 Elsevier Science Publishers B.V. (Biomedical Division)
Transcript
Gene, 84 (1989) 65-72
Elsevier
65
GENE 03 197
The block to transcription elongation at the SV40 attenuation site is decreased in vitro by oligomers complementary to segments of the attenuator RNA
David et al., 1982; Hay and Aloni, 1984; Aloni and Hay, 1985), polyoma (Skarnes et al., 1988), the parvovirus minute virus of mice (Ben-Asher and Aloni, 1984; Resnekov and Aloni, 1989), Ad2 (Maderious and Chen-Kiang, 1984; Seiberg et al., 1987; Kessler et al., 1989), and HIV-l and HIV-2 (Koa, et al., 1987; Toohey and Jones, 1989), of several proto-oncogenes including c-pnyc, c-myb and c-$3 (Bentley and Groudine, 1986; Eick and Bornkamm, 1986; Bender et al., 1987; Fort et al., 1987) and of the heat-shock gene in Drosophila (Rougie and Lis, 1988).
In SV40, Att. RNA transcripts resulting from a specific block to ~~s~ription elongation, have been observed in association with late gene expression (Hay et al., 1982). The 3’ ends of the Att transcripts were mapped to a dA-rich region in the noncoding DNA strand situated 93 nt downstream from the major late fsp at nt 325 (Hay et al., 1982; Skomik- David et al., 1982; Hay and Aloni, 1984; Aloni and Hay, 1985). Located immediately 5’ to the dA-rich sequences, there are two regions of dyad symmetry which enable the RNA transcript of this region to fold into two hairpin structures. It has been suggested that these hairpin structures, designated 1 + 2 and 3 + 4 in Fig. 1, act synergistically as the Att signal. In support of this suggestion is the observation that there is a significant reduction in the synthesis of Att transcripts when GTP is replaced by ITP in in vitro tr~sc~ption, as a result of the destabi~zation of RNA secondary structure (Hay and Aloni, 1984) and in the presence of proflavine which intercalates into RNA secondary structure (Hay et al., 1982). Moreover, recent studies using site-directed muta- genesis to create templates which either strengthened or weakened the proposed hairpin structures have shown that the extent of the block to elongation is directly dependent on the stability of the hairpin structure of the RNA (Resnekov et al., 1989).
To test this model further, we have employed an approach that was su~cess~lly exploited by Yanofsky and his colleagues in their studies on the trp attenuator (Winkler et al., 1982; Fisher and Yanofsky, 1983). We synthesized three oligos, com- plementary to strands 1, 2 and 3 (see Fig. 1), and added them to an in vitro tr~sc~ption mixture containing SV40 transcriptional complexes, to inter- fere with the folding of the RNA into the predicted secondary structures. Indeed, the addition of the
oligos specifically decreased the elongation block at the Att site. These results provide additional support for the involvement of RNA secondary structures in the regulation at the SV40 Att. site.
MATERIALS AND METHODS
(a) Preparation of viral transcription complexes
VTC were prepared from BSC-1 cells infected with SV40 (strain 776) as previously described (Laub and Aloni, 1976). Briefly, 58-h post~fection cells were harvested and the nuclei isolated by treatment with 0.5 % NP-40. They were suspended in TC buffer and treated with 0.3 y0 Sarkosyl. Cellular chromatin was pelleted by centrifugation at 30000 x g for 30 min at 2” C, The supe~atant containing the VTC was collected and dialyzed for 12 h against TC buffer containing 5 mM /I-mercaptoethanol. Aliquots (300 ~1) were then frozen in liquid nitrogen in TC buffer containing 30% glycerol and 5 mM DTT. Frozen VTC were maintained at -70 * C for up to 1 year without a noticeable loss in transc~ption activity.
(b) Preparation of oligos
Oligo l(5 ~-GCAGCACCATGGC~-3 ’ ), oligo 2 (5’-CGGAGGCCTGGCGTGACAG-3’), and oligo 3 (5 ‘-CAGTCCATGACCT-3’), complemen- tary to strands 1,2 and 3, respectively, of the hairpin structures shown in Fig. 1, as well as the noncom- plement~ oligo (S’XXACGGTGGTATATCC- AGTG-3’) were synthesized using standard syn- thetic methods and then gel purified. (Narang et al., 1980). The sequence of each of the oligos was con- firmed by sequencing prior to their use.
(c) Binding of oligos and in vitro elongation of nascent RNA associated with viral transcription complexes
Ohgo 1 and oligo 2 were allowed to hybridize to nascent RNA of the VTC by incubating them for 10 min at 37” C prior to adding NTP. Oligo 3 was not preincubated with the VTC, but was added with the transcription mixture. Unless otherwise indicated,
67
600 pmol of oligo were added to transcription
reactions. The nascent RNA transcripts were elong-
ated by adding ATP, GTP and CTP to 0.5 mM, and
[a-32P]UTP to 1 ,uM (3000 Ci/mmol, Amersham,
U.K.). The RNA was purified by phenol-chloroform
extraction followed by passage through 5 ml Se-
phadex G-25 columns and ethanol precipitation.
Prior to electrophoresis, samples in H,O were equal-
ized according to Cherenkoff counts. The H,O was
removed by vacuum centrifugation (Speed Vat Con-
centrator) and the samples resuspended in 80% for-
mamide and 1 x TBE. Following heating for 1 min
at 100°C they were quickly cooled and loaded onto
8% polyacrylamide (bis-acrylamide 1: 19) gels,
containing 7 M urea in TBE. The amount of cpm
loaded on the gels, in the various experiments, varied
between 800 and 1200. However, in each experiment
equal numbers of cpm were loaded in each lane.
RESULTS AND DISCUSSION
(a) Characterization of the in vitro system
To verify whether the RNA secondary structure
shown in Fig. 1 is involved in inducing the transcrip-
tion elongation block at the SV40 Att site situated
93 nt downstream from the major late tsp, oligos
complementary to the individual strands of the stems
of the putative hairpin structures of the SV40 Att
RNA (Fig. 1) were added to in vitro transcription
reactions, and their effect on transcription elongation
through the Att site was measured. We assumed
that if the RNA secondary structures played a role
in inducing the polymerase to stop at the Att. site,
then the addition of excess amounts of oligos com-
plementary to the individual strands of the stem
structure would interfere with the formation of the
hairpin structures and a reduction in the elongation
block at the Att site should be observed.
We chose to use VTC isolated late in SV40 in-
fection, as the transcription system. The advantage
of this in vitro transcription elongation system over
other in vitro systems, such as nuclei or minichromo-
somes, is that VTC synthesize viral RNA exclusively,
the RNA is relatively free of proteins and trans-
cription is initiated in vivo (Laub and Aloni, 1976).
Fig. 1. Schema of alternative conformations of the Att. RNA.
(A) Attenuation conformation showing the 1 + 2 and 3 + 4
hairpin structures with the 3 t 4 structure situated immediately
5’ to the U-rich sequences. (B) Readthrough conformation in
which strands 2 and 3 form the alternative 2 + 3 hairpin. The
tsp at nt 325 is indicated. The AG’s were calculated according to
Tinoco et al. (1973). Oligos 1, 2 and 3 are complementary to
strands 1, 2 and 3, respectively, of the Att. conformation.
relative usage of the multiple tsp, using systems like
HeLa whole cell extract or nuclear extract, of the late
SV40 transcript are avoided. However, since in VTC
only pausing and not true termination occurs at the
Att. site, we carried out the in vitro transcription
elongation only for a short pulse period prior to the
extraction of the RNA and quantitation of the
paused and ‘run-on’ transcripts.
(h) Transcription elongation in the presence of oligos
We have previously reported that when VTC are
extracted from SV40-infected cells, the majority of
transcriptional complexes are paused at nt 365
68
(Skolnik-David and Aloni, 1983). This has recently been confirmed using the WCE system (Resnekov et al., 1989). In initial experiments, an oligo comple- mentary to nt 330-345 (designated oligo 1; Fig. 1) was added to the in vitro elongation reactions. This oligo can, therefore, hybridize to the complementary RNA strand 1 in the nascent RNA transcripts of the VTC, prior to carrying out in vitro elongation. The hybridization reaction was carried out by pre- incubating the VTC with the oligo for 10 min at 37°C. The nascent RNA transcripts were then elongated in vitro in the presence of [ c+~~P]UTP for 2 min at 30’ C. The RNA was extracted and purified by passage through 5 ml G-25 Sephadex columns, to remove unincorporated labeled NTP. Prior to separ- ation by PAGE, the labeled samples from the differ- ent reactions were carefully equalized (with regard to their cpm).
Since the quantity of RNA present in the VTC preparation, which could potentially bind to the oligo, and the efficiency with which binding would take place were unknown, oligo 1 was initially added to the VTC preparations in increasing quantities (Fig. 2). In vitro elongation of nascent RNA asso- ciated with VTC, in the presence of 50, 100,300, and 600 pmol of oligo 1 was carried out (lanes 2-5). It is apparent from these reactions that the addition of 300 pmol or more of oligo to the elongation reactions resulted in a small but reproducible reduction in the amount of transcription complexes, which remain paused at the Att site at the time of extraction of the RNA. Based on scanning of the gel, the quantity of complexes paused at this attenuation site was re- duced by 36 % in the reaction containing 300 pmol of oligo and 41 y0 in the reaction containing 600 pmol of oligo (lanes 4 and 5, respectively). The reduction is specific because noncomplementary oligo failed to do so (Fig. 3). The reduction of Att RNA when oligo 1 is added to the transcription elongation reactions implies that RNA strand 1 (Fig. 1) is in- volved in inducing the elongation block at the Att site.
To check the possibility that oligos complementary to strands 2 and 3 are also involved in inducing the elongation block at the Att site, oligos complemen- tary to nt 350-368 and 380-392 (oligo 2 and oligo 3, respectively; Fig. I), were added to the transcription reactions. Oligo 2 was also allowed to bind to the nascent RNA transcripts by preincubating it with the
Att. RNA
Fig. 2. Transcription elongation of nascent RNA associated
with VTC in the presence of various concentrations of oligo 1.
SV40 VTC, isolated late in infection (56 h post infection), were
preincubated for 10 min at 37°C with 0, 50, 100, 300 and
600 pmol of oligo 1 (lanes l-5, respectively). The nascent RNA
transcripts were then elongated in vitro for 2 min at 28°C in the
presence of [cc-~*P]UTP, and the labeled RNA was extracted and
analyzed by 8% PAGE. Equal amounts of labeled RNA were
loaded in each lane. In all the figures, the position of the Att.
RNA is marked with an arrow. Lane M shows DNA markers (in
bp) obtained by digesting pBR322 DNA with HpaII and end-
labeling with 3ZP.
VTC prior to carrying out transcription elongation. Transcription reactions, which included no oligo or equal amounts of oligo 1, oligo 2 and a noncomple- mentary oligo of about the same length, were carried
69
1234 M
110
90
Fig. 3. Transcription elongation of nascent RNA associated
with VTC in the absence and presence of oligo 1, oligo 2, and a
noncomplementary oligo. SV40 VTC, isolated late in infection
(56 h post infection), were preincubated for 10 min at 37°C with
no oligo (lane l), 600 pmol of oligo 1 or oligo 2 (lanes 2 and 3),
and noncomplementary oligo (lane 4). In vitro transcription was
then carried out for 2 min at 28 o C. The RNA was extracted and
analyzed by 8% PAGE. For other details see Fig. 2.
out and the amounts of Att transcripts produced
were compared (Fig. 3, lanes 1-4, respectively). As
can be seen in lanes 2 and 3, the addition of oligo 1
and oligo 2 to the elongation reactions had a very
similar effect on the elongation block at the Att site.
In these reactions there was a 53 and 60 % reduction,
respectively, in Att RNA, as compared to the
reaction in which no oligo or noncomplementary
oligo was added.
In the present in vitro system, the polymerase
pauses following the 1 + 2 hairpin structure at
residue 365 (Skolnik-David and Aloni, 1983; see
Fig. 1). Oligo 3 is complementary to sequences
which are 3’ to this pause site. Binding of oligo 3 to
the nascent RNA transcripts could, therefore, be
carried out only in the course of in vitro transcription
elongation. Moreover, because of the close proximity
of strand 3 to the Att site, the rate of transcription
was slowed down, by lowering the transcription
temperature, to allow more time between the syn-
thesis of the RNA complementary to oligo 3 and the
arrival of the transcription complexes at the Att site.
Fig. 4 shows the results of a comparison between
transcription kinetics of two separate reactions in
which no oligo was added (lanes l-3 and 4-6) and
a third reaction containing oligo 3 (lanes 7-9). The
two separate experiments with no oligo are included
to show the reproducibility of the RNA pattern. The
RNA of samples removed after 4, 8 and 12 min of
transcription elongation was isolated and separated
M 123 456 789M
Att. RN IA-
Fig. 4. Kinetics of transcription elongation ofnascent RNA asso-
ciated with VTC in the absence and presence of oligo 3.
Nascent RNA transcripts associated with SV40 VTC, at 56 h
post infection, were elongated in vitro in the absence (lanes l-3
and 4-6) or in the presence of 600 pmol of oligo 3 (lanes 7-9).
Samples were removed after 4 (lanes 1,4 and 7), 8 (lanes 2,5 and
8) and 12 min (lanes 3, 6 and 9) of incubation at 16°C in the
presence of [c+~*P]UTP. Labeled RNA was extracted and
analyzed by 8% PAGE. For other details see Fig. 2.
70
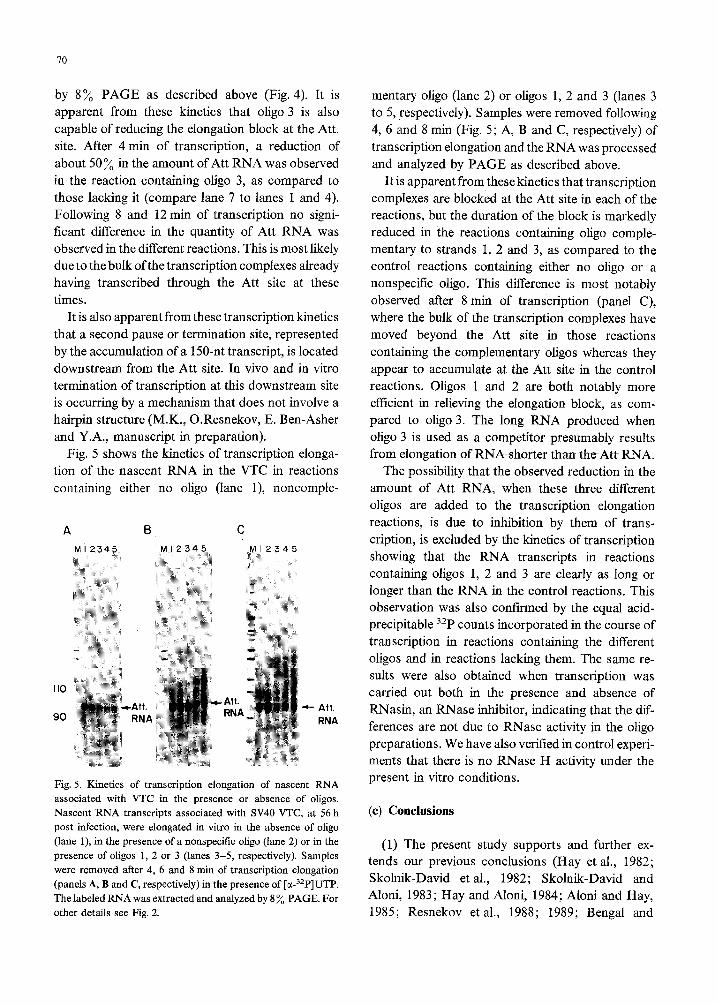
by 8% PAGE as described above (Fig. 4). It is
apparent from these kinetics that oligo 3 is also
capable of reducing the elongation block at the Att.
site. After 4 min of transcription, a reduction of
about 50% in the amount of Att RNA was observed
in the reaction containing oligo 3, as compared to
those lacking it (compare lane 7 to lanes 1 and 4).
Following 8 and 12 min of transcription no signi-
ficant difference in the quantity of Att RNA was
observed in the different reactions. This is most likely
due to the bulk of the transcription complexes already
having transcribed through the Att site at these
times.
It is also apparent from these transcription kinetics
that a second pause or termination site, represented
by the accumulation of a 150-nt transcript, is located
downstream from the Att site. In vivo and in vitro
termination of transcription at this downstream site
is occurring by a mechanism that does not involve a
hairpin structure (M.K., O.Resnekov, E. Ben-Asher
and Y.A., manuscript in preparation).
Fig. 5 shows the kinetics of transcription elonga-
tion of the nascent RNA in the VTC in reactions
containing either no oligo (lane l), noncomple-
A B C
MI 2345
i
. Att. RNA
4- Att.
RNA
Fig. 5. Kinetics of transcription elongation of nascent RNA
associated with VTC in the presence or absence of oligos.
Nascent RNA transcripts associated with SV40 VTC, at 56 h
post infection, were elongated in vitro in the absence of oligo
(lane l), in the presence of a nonspecific oligo (lane 2) or in the
presence of oligos 1, 2 or 3 (lanes 3-5, respectively). Samples
were removed after 4, 6 and 8 min of transcription elongation
(panels A, B and C, respectively) in the presence of [LY-~~P]UTP.
The labeled RNA was extracted and analyzed by 8 % PAGE. For
other details see Fig. 2.
mentary oligo (lane 2) or oligos 1, 2 and 3 (lanes 3
to 5, respectively). Samples were removed following
4, 6 and 8 min (Fig. 5; A, B and C, respectively) of
transcription elongation and the RNA was processed
and analyzed by PAGE as described above.
It is apparent from these kinetics that transcription
complexes are blocked at the Att site in each of the
reactions, but the duration of the block is markedly
reduced in the reactions containing oligo comple-
mentary to strands 1, 2 and 3, as compared to the
control reactions containing either no oligo or a
nonspecific oligo. This difference is most notably
observed after 8 min of transcription (panel C),
where the bulk of the transcription complexes have
moved beyond the Att site in those reactions
containing the complementary oligos whereas they
appear to accumulate at the Att site in the control
reactions. Oligos 1 and 2 are both notably more
efficient in relieving the elongation block, as com-
pared to oligo 3. The long RNA produced when
oligo 3 is used as a competitor presumably results
from elongation of RNA shorter than the Att RNA.
The possibility that the observed reduction in the
amount of Att RNA, when these three different
oligos are added to the transcription elongation
reactions, is due to inhibition by them of trans-
cription, is excluded by the kinetics of transcription
showing that the RNA transcripts in reactions
containing oligos 1, 2 and 3 are clearly as long or
longer than the RNA in the control reactions. This
observation was also confirmed by the equal acid-
precipitable 32P counts incorporated in the course of
transcription in reactions containing the different
oligos and in reactions lacking them. The same re-
sults were also obtained when transcription was
carried out both in the presence and absence of
RNasin, an RNase inhibitor, indicating that the dif-
ferences are not due to RNase activity in the oligo
preparations. We have also verified in control experi-
ments that there is no RNase H activity under the
present in vitro conditions.
(c) Conclusions
(1) The present study supports and further ex-
tends our previous conclusions (Hay et al., 1982;
Skolnik-David et al., 1982; Skolnik-David and
Aloni, 1983; Hay and Aloni, 1984; Aloni and Hay,
1985; Resnekov et al., 1988; 1989; Bengal and
71
Aloni, 1989) that the control at the SV40 Att site is
mediated by the two mutually exclusive RNA
secondary structures, designated as attenuation and
readthrough conformations (Fig. 1, A and B, respec-
tively), contained in a region of the SV40 genome
spanning between nt 330 and 4 18 (SV40 numbering).
Moreover, the 1 + 2 and 3 + 4 hairpin structures
(Fig. 1) act synergistically as the Att. conformation.
Thus, the relatively unstable 3 + 4 hairpin structure
will only form once the 1 + 2 structure has been
formed. Disruption of the 1 + 2 secondary structure
would subsequently preclude the formation of the
3 + 4 hairpin structure.
(2) The utilization of oligos, in a manner similar
to that presented here, was used successfully to
establish the RNA secondary structures directing
termination at the trp Att (Winkler et al., 1982;
Fisher and Yanofsky, 1983). The present study sup-
ports the idea that complementary oligos are useful
probes in the study of transcription-related pheno-
mena.
(3) The prokaryotic RNA polymerase and the
eukaryotic RNA polymerase II recognize common
elements as signals for transcription Att and it is
suggested that Att with the adequate adjustments
has been conserved during evolution.
(4) Transcription Att in eukaryotes can occur at
various distances from the tsp and they can function
by various mechanisms (Aloni and Hay, 1985;
Rougie and Lis, 1988; Bently and Groudine, 1986;
M.K., O.Resnekov, E. Ben-Asher and Y.A., manu-
script in preparation).
(5) Transcription Att is a general form of gene
regulation in eukaryotes.
ACKNOWLEDGEMENTS
We thank Michael Laspia for critical reading of
this paper. This research was supported by the fol-
lowing grants: The United States Public Health
Service-CA 14995; Minerva Foundation, Munich,
F.R.G. ; the Leo Forcheimer Center for Molecular
Genetics and the Rockefeller-WIF Collaboration
Trust Fund.
REFERENCES
Aloni, Y. and Hay, N.: Attenuation may regulate gene expression
in animal viruses and cells. CRC Crit. Rev. Bioch. 18 (1985)
327-383.
Ben-Asher, E. and Aloni, Y.: Transcription of minute virus of
mice, an autonomous parvovirus may be regulated by
attenuation. J. Virol. 52 (1984) 266-276.
Bender, T.P., Thomas, C.B. and Kuehl, W.M.: Differential ex-
pression of c-myb mRNA in murine lymphomas by a block to