The B. abortus general stress response 1 The Brucella abortus general stress response system regulates chronic mammalian infection, and is controlled by phosphorylation and proteolysis Hye-Sook Kim 1,2 , Clayton C. Caswell 3 , Robert Foreman 2,4 , R. Martin Roop II 3 , and Sean Crosson 1,2,4 1 Department of Biochemistry and Molecular Biology, The University of Chicago, Chicago, IL 60637, USA. 2 Howard Taylor Ricketts Laboratory, Argonne National Laboratory, Argonne, IL, 60439, USA. 3 Department of Microbiology and Immunology, East Carolina University School of Medicine, Greenville, NC 27834, USA. 4 The Committee on Microbiology, The University of Chicago, Chicago, IL 60637, USA. Running title: The B. abortus general stress response To whom correspondence should be addressed: Sean Crosson, Department of Biochemistry and Molecular Biology, The University of Chicago, 929 E. 57th St. – GCIS W138, Chicago, IL 60637, USA. Tel. (773) 834-1926; Fax (773) 702-0439. E-mail: [email protected]. Keywords: Brucella, general stress response, PhyR, signaling, proteolysis, sigma Background: Virulence of pathogenic bacteria is often determined by their ability to adapt to stress. Result: The Brucella abortus general stress response (GSR) system is required for chronic mammalian infection, and is regulated by phosphorylation and proteolysis. Conclusion: The B. abortus GSR signaling pathway has multiple layers of post-translational control, and is a determinant of chronic infection. Significance: This study provides new, molecular- level insight into chronic Brucella infection. SUMMARY Brucella spp. are adept at establishing a chronic infection in mammals. We demonstrate that core components of the α- proteobacterial general stress response (GSR) system, PhyR and σ E1 , are required for B. abortus stress survival in vitro and maintenance of chronic murine infection in vivo. Δ phyR and Δ rpoE1 null mutants exhibit decreased survival under acute oxidative and acid stress, but are not defective in infection of primary murine macrophages or in initial colonization of BALB/c mouse spleens. However, Δ phyR and Δ rpoE1 mutants are attenuated in spleens beginning one month post-infection. Thus, the B. abortus GSR system is dispensable for colonization, but is required to maintain chronic infection. A genome-scale analysis of the B. abortus GSR regulon identified stress response genes previously linked to virulence, and genes that affect immunomodulatory components of the cell envelope. These data support a model in which the GSR system affects both stress survival and the interface between B. abortus and the host immune system. We further demonstrate that PhyR proteolysis is a unique feature of GSR control in B. abortus. Proteolysis of PhyR provides a mechanism to avoid spurious PhyR protein interactions that inappropriately activate GSR- dependent transcription. We conclude that the B. abortus GSR system regulates acute stress adaptation and long-term survival within a mammalian host, and that PhyR proteolysis is a novel regulatory feature in B. abortus that ensures proper control of GSR transcription. INTRODUCTION Bacteria of the genus Brucella are the causative agents of brucellosis, which is among the most common zoonotic infections globally (1). Within the environment of a mammalian host, http://www.jbc.org/cgi/doi/10.1074/jbc.M113.459305 The latest version is at JBC Papers in Press. Published on April 1, 2013 as Manuscript M113.459305 Copyright 2013 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on March 1, 2019 http://www.jbc.org/ Downloaded from
Transcript
The B. abortus general stress response
1
The Brucella abortus general stress response system regulates chronic mammalian infection, and is controlled by phosphorylation and proteolysis
Hye-Sook Kim1,2 , Clayton C. Caswell3, Robert Foreman2,4, R. Martin Roop II3,
and Sean Crosson1,2,4
1 Department of Biochemistry and Molecular Biology, The University of Chicago, Chicago, IL 60637, USA.
2 Howard Taylor Ricketts Laboratory, Argonne National Laboratory, Argonne, IL, 60439, USA.
3 Department of Microbiology and Immunology, East Carolina University School of Medicine,
Greenville, NC 27834, USA.
4 The Committee on Microbiology, The University of Chicago, Chicago, IL 60637, USA.
Running title: The B. abortus general stress response
To whom correspondence should be addressed: Sean Crosson, Department of Biochemistry and Molecular Biology, The University of Chicago, 929 E. 57th St. – GCIS W138, Chicago, IL 60637, USA.
Tel. (773) 834-1926; Fax (773) 702-0439. E-mail: [email protected]. Keywords: Brucella, general stress response, PhyR, signaling, proteolysis, sigma Background: Virulence of pathogenic bacteria is often determined by their ability to adapt to stress. Result: The Brucella abortus general stress response (GSR) system is required for chronic mammalian infection, and is regulated by phosphorylation and proteolysis. Conclusion: The B. abortus GSR signaling pathway has multiple layers of post-translational control, and is a determinant of chronic infection. Significance: This study provides new, molecular-level insight into chronic Brucella infection. SUMMARY
Brucella spp. are adept at establishing a chronic infection in mammals. We demonstrate that core components of the α-proteobacterial general stress response (GSR) system, PhyR and σE1, are required for B. abortus stress survival in vitro and maintenance of chronic murine infection in vivo. ΔphyR and ΔrpoE1 null mutants exhibit decreased survival under acute oxidative and acid stress, but are not defective in infection of primary murine macrophages or in initial colonization of BALB/c mouse spleens. However, ΔphyR and ΔrpoE1 mutants are attenuated in spleens
beginning one month post-infection. Thus, the B. abortus GSR system is dispensable for colonization, but is required to maintain chronic infection. A genome-scale analysis of the B. abortus GSR regulon identified stress response genes previously linked to virulence, and genes that affect immunomodulatory components of the cell envelope. These data support a model in which the GSR system affects both stress survival and the interface between B. abortus and the host immune system. We further demonstrate that PhyR proteolysis is a unique feature of GSR control in B. abortus. Proteolysis of PhyR provides a mechanism to avoid spurious PhyR protein interactions that inappropriately activate GSR-dependent transcription. We conclude that the B. abortus GSR system regulates acute stress adaptation and long-term survival within a mammalian host, and that PhyR proteolysis is a novel regulatory feature in B. abortus that ensures proper control of GSR transcription. INTRODUCTION
Bacteria of the genus Brucella are the causative agents of brucellosis, which is among the most common zoonotic infections globally (1). Within the environment of a mammalian host,
http://www.jbc.org/cgi/doi/10.1074/jbc.M113.459305The latest version is at JBC Papers in Press. Published on April 1, 2013 as Manuscript M113.459305
Copyright 2013 by The American Society for Biochemistry and Molecular Biology, Inc.
brucelleae largely inhabit professional and non-professional phagocytic cells (2), where they occupy a Brucella-containing vacuole (3-5). Brucelleae replicate within this vacuole and, if left untreated, eventually spread and cause a chronic infection of multiple tissues. Even after antibiotic treatment the rate of relapse is high (6,7), and in a minority of patients chronic brucellosis persists for years despite multiple courses of treatment (8).
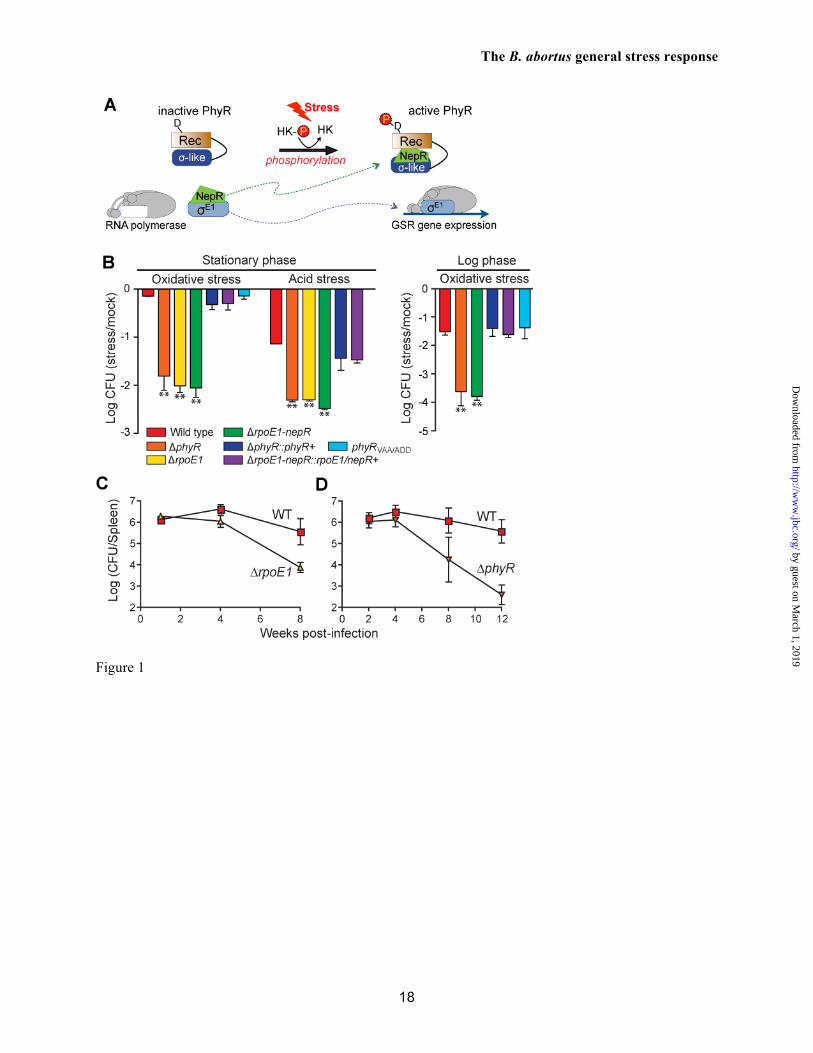
The capacity of Brucella spp. to survive the harsh environmental stressors encountered en route to, and within, their replicative niche, including reactive oxygen species, acidic pH, and nutrient deprivation is a key virulence determinant (reviewed in refs. (2,9)). In this study, we present a genetic and molecular analysis of the Brucella abortus general stress response (GSR) system, which integrates features of two-component signal transduction (TCS) and extracytoplasmic function (ECF) sigma (σ) transcriptional regulation (10,11). The central regulator in this pathway is the anti-anti-σ factor PhyR, which is composed of an ECF σ-like (SL) domain positioned amino-terminally to a two-component receiver domain (Figure 1A). Studies in related α-proteobacteria have shown that stress-induced phosphorylation of the PhyR receiver domain promotes binding of its SL domain to the anti-σ factor, NepR (10,12-15). PhyR binding titrates the NepR anti-σ factor away from a bona fide ECF σ factor (16), thereby initiating transcription of genes that mediate bacterial cell survival during stress (Figure 1A).
The core genetic components of this regulatory system including phyR, nepR, and the ECF sigma factor rpoE1 are conserved within the Brucellaceae (11,17). It has been previously reported that a B. melitensis 16M ΔrpoE1 mutant is attenuated in BALB/c mice at 4 weeks post infection (17) and exhibits derepressed flagellar biogenesis (17,18). However, this rpoE1 mutant did not have a survival defect in a hydrogen peroxide disk diffusion assay (17). Herein, we report that the B. abortus 2308 phyR-nepR-rpoE1 system is required for survival under acute oxidative and acid stress in vitro and for maintenance of B. abortus within murine spleens during the chronic phase of infection (i.e. > 1 month post-infection). Additionally, we present a detailed molecular analysis of purified B. abortus GSR proteins that defines unusual features of
PhyR-NepR-σE1 protein-protein interactions, including high-affinity binding between unphosphorylated PhyR and NepR and a PhyR~P/NepR complex with an extremely slow dissociation half-time. The cellular concentration of PhyR is regulated post-translationally in B. abortus: PhyR is degraded in the absence of stress, likely by the essential (19) ClpXP protease, and is stabilized when the cell is exposed to either oxidative or acid stress. A B. abortus mutant strain encoding a stable version of PhyR (PhyRVAA/ADD) exhibits derepressed transcription of GSR target genes. Thus, PhyR proteolysis helps to ensure proper control of GSR-dependent transcription in B. abortus. In contrast, we did not observe PhyR protein turnover in the related α-proteobacterium, Caulobacter crescentus; this provides evidence for diverse modes of GSR regulation among α-proteobacteria. Diversity in GSR regulation likely reflects the broad range of ecological niches inhabited by α-proteobacterial species, from dilute freshwater, to plant surfaces, to mammalian phagocytic cells. EXPERIMENTAL PROCEDURES Bacterial strains and culture conditions. All studies in live B. abortus were performed at Biosafety Level 3 (BSL3) as per CDC select agent regulations at the University of Chicago H.T Ricketts Laboratory or East Carolina University School of Medicine. A detailed description of bacterial strains and culture conditions is provided in Supplemental Materials and Methods. Construction of B. abortus mutants and genetic complementation. All primers used in this study are listed in Table S4. A double recombination strategy described previously (20,21) was used to create unmarked B. abortus 2308 mutant strains in which phyR, rpoE1, and nepR-rpoE1 were deleted, or in which the wild-type phyR allele was replaced with phyRVAA/ADD, phyR(D191A), and phyR(D191E) mutant alleles. The phyR and nepR-rpoE1 deletion mutations were complemented by plasmid integration of a copy of phyR or nepR-rpoE1 plus native promoter back into B. abortus chromosome 1. A detailed description of strain and plasmid constructions is provided in Supplemental Materials and Methods.
Oxidative and acid stress assays. Brucella cells grown on Schaedler blood agar (SBA) for 48 hrs were inoculated at either 103 or 106 CFU/ml into Brucella broth and grown at 37°C for 24 hr. Final cell density was adjusted to 108 CFU/ml before oxidative and acid stress assays. For PhyR stability experiments, B. abortus strains grown on SBA supplemented with appropriate antibiotics for 48 h were harvested into Gerhardt's minimal medium (GMM) (22) and then adjusted to a cell density of 108 CFU/ml. Cells were subjected to oxidative and acid stresses by the addition of 5 mM H2O2 (final concentration) or addition of acid to adjust the medium to pH 3.9, respectively. Cell survival was measured at 1 or 2 hrs after the H2O2 or low pH exposures in a shaking incubator at 37 °C by enumerating CFU/ml. For analysis of protein turnover under stress conditions, 500 µg ml-1 of chloramphenicol was added to inhibit protein synthesis 10 min after oxidative or acid stress induction; protein samples were subsequently harvested at 0, 30, 60, and 120 min for immunoblot analysis. Microarray analysis. After 10 min exposure to 5 mM H2O2, B. abortus wild type and ΔrpoE1 cells were harvested, RNA was isolated and hybridized to URMC_Dunman_Brucella_abortus_v1.0 arrays (Affymetrix), and data were analyzed as described in Supplemental Materials and Methods. Datasets are available at the European Bioinformatics Institute server (ArrayExpress accession number E-MEXP-3722). Animal infection to assess B. abortus virulence. Female BALB/c mice (4-5 weeks old) were purchased from Harlan Laboratories, Inc. Parallel animal studies on ΔphyR and ΔrpoE1 strains were conducted at The University of Chicago and East Carolina University, respectively under protocols approved by institutional animal care and use committee (IACUC) and institutional biosafety committee (IBC). Animals used in this study were housed under specific-pathogen-free conditions in a BSL3 animal facility at the University of Chicago H.T. Ricketts Laboratory or at East Carolina University. Six-week-old, female BALB/c mice were infected with 5 x 104 CFU of wild-type B. abortus 2308, ΔphyR or ΔrpoE1 strains via the intraperitoneal route. Mouse spleens were removed aseptically, homogenized in
1 ml PBS, and serial dilutions of homogenized spleens were plated on SBA to enumerate bacterial CFU at 1, 2, 4, 8, and 12 weeks post-infection. Analytical size exclusion chromatography (SEC). SEC was performed on a 10/300 Superdex-75 column (GE healthcare) equilibrated with 20mM Tris-HCl (pH7.5), 200mM NaCl2 at a flow rate of 0.5 ml/min. Absorbance at 280 nm was used to detect proteins eluted from the column. Each fraction was subjected to SDS-PAGE and visualized by either Commassie Brilliant Blue or silver stain. Analytical ultracentrifugation (AUC). Sedimentation velocity analysis of purified B. abortus proteins was performed at 20°C in a Beckman Optima XL-A analytical ultracentrifuge equipped with a Ti60 rotor (Beckman Coulter) using absorption optics at a wavelength of 280 nm. Experiments were carried out with protein concentrations ranging from 10 to 55 µM. A 1.2-mm thick two-channel Epon centerpiece was used at a rotor speed of 40,000 rpm in a continuous-mode time intervals of 360-480s. The continuous size distribution data were analyzed using the program Sedfit (23). Surface plasmon resonance (SPR). Association and dissociation rates of PhyR and σE1 binding to surface immobilized NepR were measured using a BIAcore 3000 (GE Healthcare). Experiments were conducted at 15°C in HBS-EP buffer composed of 10mM HEPES (pH7.4), 150mM NaCl, 0.005% Surfactant P20, and 3 mM EDTA. For binding measurements of phospho-PhyR to NepR, 10 mM Acetyl-phosphate and 5 mM MgCl2 were added to the HBS-EP buffer. Briefly, His6-tagged NepR was immobilized to the surface of a nitrilotriacetic (NTA) biosensor chip charged with Ni2+. The His6-tag on SPR analytes (PhyR and σE1) were removed with TEV protease, and the tag-free digested analytes were purified by an additional round of Ni2+ affinity chromatography. Association was monitored by injecting increasing concentrations of the analyte at a flow rate of 20 µl/min for 300 seconds. Dissociation was measured for 300s for PhyR and σE1 and 1200s for phospho-PhyR. Between each injection, the surface was regenerated by washing with 200 µl of 1M EDTA and 1M imidazole. Data from a
reference flow cell not loaded with Ni2+, and a blank injection with buffer alone were used to determine baseline. Kinetic analyses were performed by fitting the data to a kinetic model for 1:1 binding with drifting baseline (Rmax local) using Biaeval software (GE Healthcare). Binding assays were carried out on, and were consistent across three independent protein preparations. Rmax for all experiments was between 100-500 RU. Protein immunoblot analysis. Rabbit PhyR polyclonal antiserum (Josman, LLC) was raised against His6-PhyR, which was purified by serial affinity and size exclusion chromatographies. Rabbits were immunized with purified PhyR on days 0, 21, 35, 49, and 63. Total B. abortus protein from liquid cell culture was concentrated by precipitation with 10% trichloroacetic acid, separated by 12 % SDS/PAGE, and then transferred to a PVDF membrane (Millipore). The membrane was probed with the primary polyclonal anti-PhyR antiserum (1:1,000) followed by HRP-conjugated goat anti-rabbit secondary antibodies (1:10,000) (Thermo Scientific). Blots were developed using SuperSignal West Femto Substrate (Thermo Scientific). The membrane was stripped using Restore Western Blot Stripping Buffer (Thermo Scientific) to probe with rabbit polyclonal ClpX antiserum (19) as described above for anti-PhyR. The signal intensity was measured using ChemiDoc MP system (Bio-Rad). RNA isolation and quantitative real-time PCR. Total RNA was isolated using modified hot-phenol extraction, including DNase I digestion. Total RNA was reverse transcribed using random primers and Superscript II reverse transcriptase (Invitrogen). qRT-PCR was performed with an ABI StepOnePlus™ Systems (ABI Prism) and the Power SYBR Green PCR Master Mix (Applied Biosystems) according to the manufacturer's instructions. Detailed descriptions of total RNA isolation and qRT-PCR are provided in Supplemental Materials and Methods. Experiments were conducted in biological triplicate with technical triplicates for each gene.
RESULTS The phyR-nepR-rpoE1 locus determines B. abortus survival during oxidative and acid stress
To test the role of the predicted phyR-nepR-rpoE1 general stress regulatory system (GSR) in B. abortus 2308 stress survival, we first measured differences in cell viability between wild-type strain 2308 and single and double in-frame deletion strains of phyR, nepR, and rpoE1 that were grown to early stationary phase, and subjected to equivalent levels of oxidative or acid stress. Deletion of the rpoE1, nepR-rpoE1 or phyR genes resulted in a ≈30-fold (1.5 log) reduction in cell survival relative to wild type after a 1-hour exposure to 5 mM H2O2. Single-copy integration of phyR or nepR-rpoE1 with their native promoters back into B. abortus chromosome 1 (ΔphyR::phyR+ and ΔnepR-rpoE1::nepR-rpoE1+) restored the survival defect under this oxidative stress condition (Figure 1B). A similar survival defect was observed in the deletion strains relative to wild type after incubation of cells at pH 3.9 for 2 hours (Figure 1B). This low pH survival defect was also restored in the ΔphyR::phyR+ and ΔnepR-rpoE1::nepR-rpoE1+ strains. These data demonstrate that phyR-nepR-rpoE1 genetic locus is required for normal cell survival under two distinct stress conditions.
In enteric bacteria, entry into stationary phase confers protection against multiple environmental stressors via gene regulation by σS (24). We tested whether B. abortus cells grown in log phase exhibited higher sensitivity to oxidative stress than stationary phase cells. Whereas 5 mM H2O2 treatment of wild-type B. abortus harvested from stationary phase had no significant effect on viability, an equivalent treatment of cells harvested from logarithmic phase resulted in a 1.5 log decrease of viable CFU/ml. Survival of B. abortus ΔphyR harvested from logarithmic growth phase was severely compromised after peroxide treatment, decreasing 4 logs relative to a mock treated control (Figure 1B). However, the survival differential between wild-type and ΔphyR strains of B. abortus is equivalent in cells from logarithmic phase and stationary phase (i.e. phyR confers an approximate 2 log survival advantage under oxidative stress regardless of growth phase).
This result suggests that PhyR-independent pathways are involved in the stationary phase-dependent stress protection that we observe. B. abortus GSR system is required for maintenance of chronic infection in a mouse model
Genes regulating stress physiology are important virulence determinants in Brucella spp. (25) and in other bacterial pathogens. To test whether the GSR system plays a role in B. abortus colonization or survival within mammalian cells or an animal host, we infected primary murine macrophages with wild-type B. abortus 2308 and a ΔrpoE1 mutant and BALB/c mice with wild-type B. abortus 2308, ΔphyR, and ΔrpoE1. We observed no defect in replication of a B. abortus ΔrpoE1 mutant within primary murine macrophages (Figure S1) or in the initial colonization of the animal as assessed by enumerating brucelleae in the mouse spleen; bacterial counts (CFU/spleen) were statistically identical at 2 and 4 weeks post intraperitoneal (IP) infection (n = 5; p > 0.05) (Figure 1C). 8 weeks post-infection, however, the load of cells in the mouse spleen was significantly reduced in mice infected with both ΔphyR and ΔrpoE1 (p < 0.009) null strains. By 12 weeks, mice infected with B. abortus ΔphyR had 3 logs fewer splenic bacteria than mice infected with wild-type B. abortus 2308 (p < 0.0001). These results demonstrate that B. abortus GSR system regulates pathogenesis specifically at the chronic phase of infection. rpoE1 regulates transcription of stress response genes and genes affecting the biogenesis of cell envelope immune effectors
We next conducted a genome-scale analysis of the B. abortus GSR regulon. Specifically, we measured transcript levels in a ΔrpoE1 deletion and wild type parent strains subjected to equivalent oxidative stress conditions using a custom tiled Affymetrix GeneChip (see Supplemental Materials and Methods). We identified 81 genes that were expressed at levels above background and whose expression changed more than twofold (Table S1 and S2). We validated six of these σE1/GSR gene targets by qRT-PCR; all six validation experiments confirmed our GeneChip measurements (Figure S2). Our data demonstrate that σE1 directly or
indirectly activates a group of 73 genes and represses 8 genes in the presence of oxidative stress in vitro. These experiments yield a clear set of regulated genes that fall into three main categories (Table 1): (1) acute stress response genes including dps, urease, and cydAB, which are activated by σE1 and are known determinants of B. abortus virulence (26,27), (2) transcriptional regulatory proteins including the heat shock factor and virulence determinant rpoH1 (17), which is strongly activated by σE1, (3) cell envelope genes including the flagellar gene cluster, which is repressed by σE1 (18), and effectors of LPS O-chain ba14K (28) and pgm (29,30), which are activated by σE1. We note that both ΔphyR and ΔrpoE1 mutants appear smooth by both crystal violet and acriflavine staining suggesting loss of the GSR system does not result in gross changes in LPS O-chain structure (data not shown).
Genes including cydB, pgm, and the urease cluster, which are transcriptionally activated by σE1 under oxidative stress in vitro, are required for the early stage of Brucella spp. animal infection (26,27,29). This stands in contrast to the specific chronic infection defect of ΔphyR and ΔrpoE1 strains. However, we note that assignment of genes such as cydB, pgm and urease as determinants of acute Brucella infection is based on loss-of-function gene deletion data; the effects of transcriptional dysregulation of these virulence genes on animal infection is not known. It may be that the specific chronic defect we observe in ΔphyR and ΔrpoE1 strains is a result of missexpression of multiple GSR gene targets.
We further identified conserved σEcfG binding motifs (GGAACC-N15-CGTT) upstream of 11 σE1-regulated genes using the MAST algorithm (31) (Table S3). This sequence matches the known σEcfG regulatory sequence described by Mascher and colleagues (32). Our data inform a model in which the PhyR/σE1 regulatory system controls acute stress response genes, but also functions to modulate components of the B. abortus cell envelope (O-chain and flagellum). Regulated remodeling of cell envelope composition by PhyR/σE1 under select stress conditions may play an important role in long-term immune evasion or immune modulation during chronic mammalian infection.
A molecular-level characterization of the B. abortus GSR system
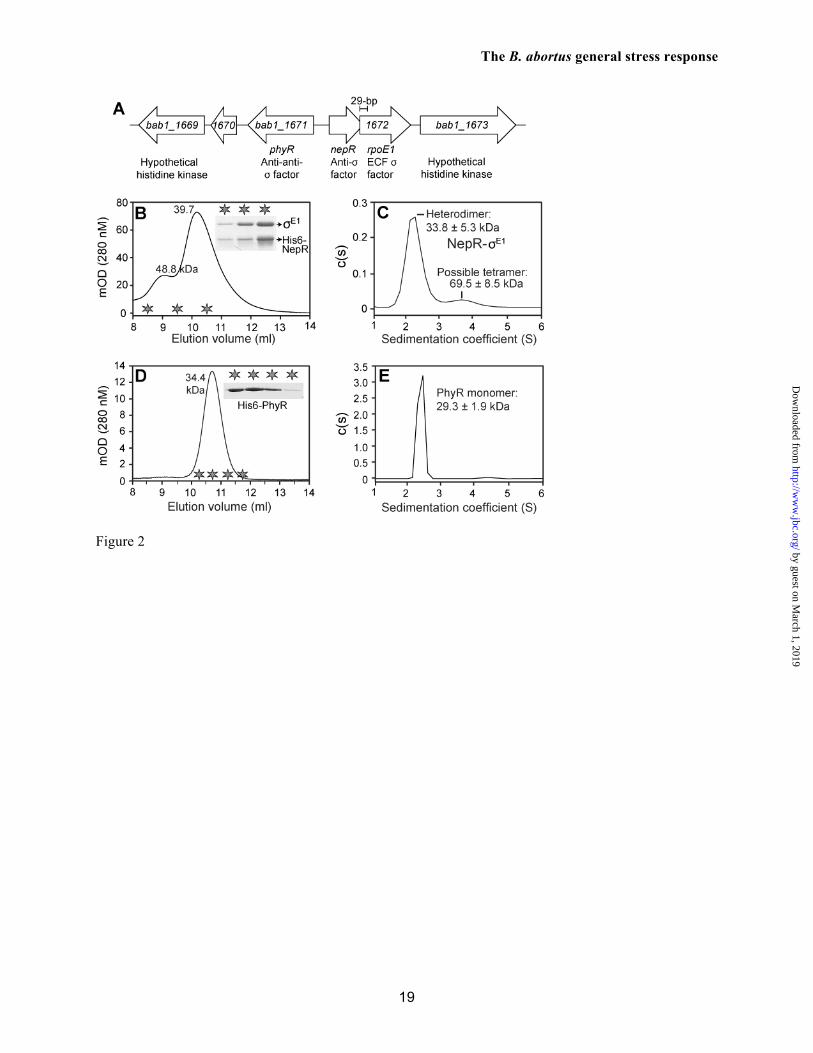
PhyR is encoded from a highly conserved gene cluster on B. abortus 2308 chromosome 1. As in other α-proteobacteria, a predicted anti-σ factor (nepR) and an ECF σ factor (rpoE1) are adjacent to phyR on the chromosome but are oriented in the opposite direction. The nepR and rpoE1 coding regions overlap by 29 base pairs (Figure 2A). The unusual overlap between nepR and rpoE1 does not appear to be result of a genome sequencing error or misannotation; we confirmed the sequence of this locus, there is no alternative rpoE1 translational start site that would produce a sense rpoE1 gene product. To test whether the overlapped genes in this putative operon yield discrete NepR and RpoE1 (σE1) proteins from a single promoter, we cloned the complete nepR-rpoE1 locus into the hexahistidine-fusion expression plasmid, pET151/D-TOPO. This construct placed transcription of nepR-rpoE1 under the control of a single T7 promoter, and was minimally predicted to produce a His6-NepR fusion protein. We induced expression from this plasmid in E. coli BL21(DE3) and purified protein from the cell lysate on Ni2+-NTA resin. Two major proteins eluted from Ni2+-NTA of matching the predicted sizes of His6-NepR and σE1.
These co-eluted proteins were subsequently purified by size exclusion chromatography (SEC) on Superdex 75 resin and eluted in a major peak at 10.2 ml that contained His6-NepR and σE1. These same two proteins also eluted in minor peak (9.0 ml) that was presumably composed of a higher-order oligomer (Figure 2B). Based on the elution volume of the major peak, we calculated a predicted molecular weight of 39 kDa for the His6-NepR/σE1 complex. This result did not permit us to draw a firm conclusion regarding complex stoichiometry. As such, we further characterized this purified complex by sedimentation velocity analytical ultracentrifugation (AUC). His6-NepR/σE1 primarily sediments as a 2.4 S particle (Figure 2C). A molecular weight distribution associated with this 2.4 S particle was calculated from a distribution of Lamm equation solutions and yielded a predicted mass of 33.8 ± 5.3 kDa; this measurement is congruent with the calculated 33.3 kDa mass of a 1:1 His6-NepR/σE1 complex. As in
our SEC experiments, we also observe a larger (3.8 S) minor species that corresponds to a calculated molecular mass of 69.5 ± 8.5 kDa. These size exclusion and sedimentation data are consistent with a model in which His6-NepR/σE1 primarily form a 1:1 heterodimeric complex at the assessed concentration of 12 µM. Under these conditions, a small fraction of these two proteins form a higher-order oligomer, which may be a 2:2 heterotetramer. We conclude that the overlapping nepR-rpoE1 open reading frames yield discrete NepR and σE1 proteins, and that these proteins primarily form a 1:1 heterodimeric complex.
We next assessed the oligomeric state of B. abortus PhyR by SEC and AUC. PhyR elutes from a Superdex 75 column in a single peak at a volume corresponding to an apparent molecular mass of 34.4 kDa (Figure 2D). Analysis of PhyR by AUC is consistent with SEC; PhyR sediments as a single 2.3 S particle with a calculated molecular mass of 29 ± 1.9 kDa (Figure 2E). Thus, B. abortus PhyR is monomeric at the assessed concentrations of 10-50 µM. Evidence for atypical protein-protein interactions between B. abortus NepR and its regulatory partners, PhyR and σE1
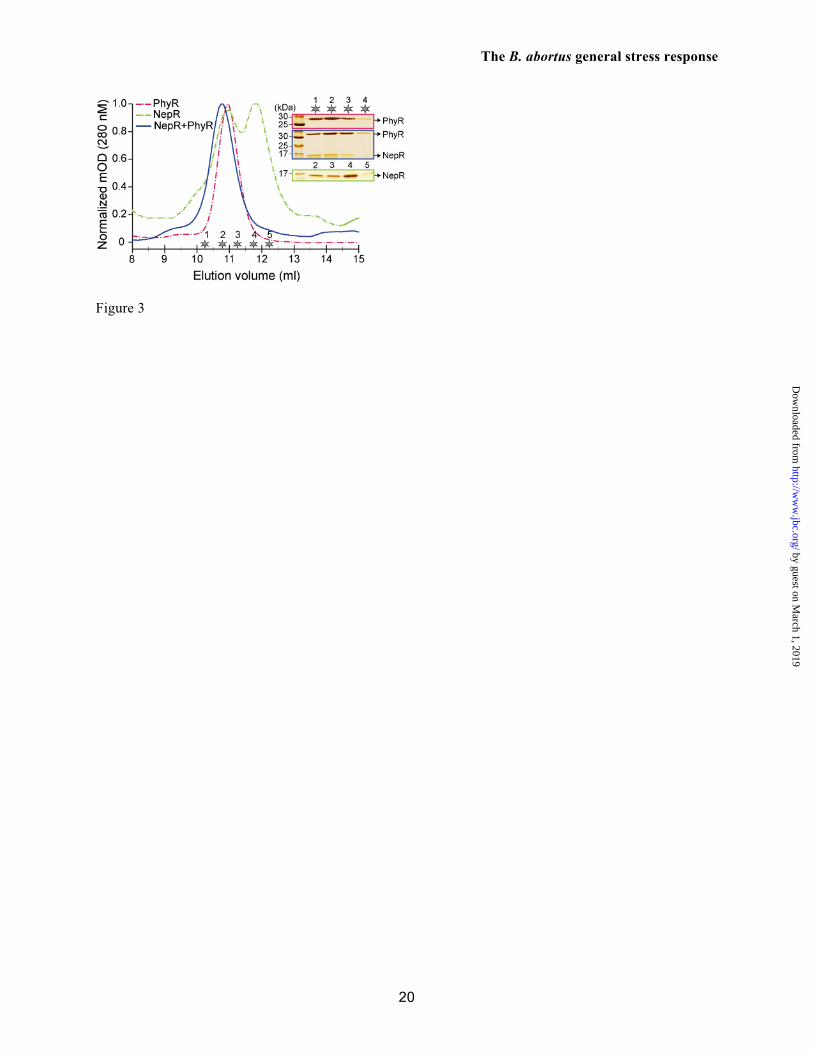
In related α-proteobacteria, aspartyl phosphorylation of the PhyR receiver domain (RD) is required for the high-affinity interaction between PhyR and the anti-σ factor, NepR. Through this mechanism, phospho-PhyR recruits NepR from its cognate ECF σ factor and promotes initiation of transcription during stress. To test this protein partner-switching model in B. abortus, we first co-expressed His6-NepR with either wild-type PhyR or with PhyR point mutants in which the aspartyl phosphorylation site (D191) was mutated to alanine (D191A) or glutamic acid (D191E). Surprisingly, all three versions of PhyR strongly co-purified with His6-NepR from a Ni2+-NTA affinity column (Figure S3). We further assayed binding between unphosphorylated PhyR and NepR, by individually purifying PhyR and NepR proteins and mixing them at a concentration of 10 µM each. Purified PhyR and NepR proteins have distinct elution profiles from a Superdex 75 size exclusion column (11.1 and 11.1/11.8 ml, respectively). The 10 µM PhyR:NepR protein mixture eluted in a single peak at a lower volume
(10.7 ml) providing evidence for formation of a higher molecular weight complex (Figure 3). This result differs from what our group (33) and others (12-16) have observed for purified PhyR and NepR orthologs from related species, which do not stably interact at similar concentrations in the absence of PhyR phosphorylation. Based on these experiments we can conclude that unphosphorylated B. abortus PhyR and NepR form a stable complex at 10 µM.
To quantify the protein-protein interactions between B. abortus NepR and its partners, PhyR and σE1, we next measured association (ka) and dissociation (kd) rate constants by surface plasmon resonance (SPR). In these experiments His6-NepR was immobilized on a nitrilotriacetic (NTA) biosensor chip and purified PhyR or σE1 were flowed across the chip at increasing concentrations. NepR binding to PhyR and σE1 is characterized by slow association and dissociation kinetics and similar equilibrium affinities (57 nM and 67 nM, respectively) (Figure 4), though ka and kd of NepR-σE1 binding are each approximately 5-fold faster than NepR-PhyR binding.
To test whether the unusual high-affinity interaction we observe between NepR and unphosphorylated PhyR is unique to the B. abortus protein pair, we measured binding of immobilized B. abortus NepR to unphosphorylated Caulobacter crescentus PhyR, which is 55% identical and 70% similar to B. abortus PhyR. We could only detect weak binding signal at a C. crescentus PhyR concentration ≈10 times higher (1 µM) than the highest concentration measured for B. abortus PhyR (Figure S4). At this concentration, binding quickly saturated at 13 response units (RU), which is two orders of magnitude below the calculated maximal binding signal (Rmax) and indicative of non-specific interaction. These data provide evidence that the high affinity interaction we observe between NepR and unphosphorylated PhyR is specific to the B. abortus PhyR-NepR pair.
Our experiments provide support for atypical interactions among B. abortus GSR proteins, in which NepR forms complexes with unphosphorylated PhyR and σE1 with near-equivalent affinity (≈60 nM). Experiments that
test the functional implications of these binding interactions are described in a later section. B. abortus phospho-PhyR forms a high affinity, long-lived complex with NepR
As described in the introduction, studies in other species of α-proteobacteria have demonstrated that phosphorylation of PhyR is required for high affinity binding to the anti-σ factor, NepR. To test whether PhyR phosphorylation modulates its affinity for NepR, we added acetyl phosphate (AcP) and Mg2+ to purified B. abortus PhyR; this buffer condition is known to induce in vitro phosphorylation of the receiver domain in related species (10,12,14,15,33). Under this buffer condition, we measured binding of PhyR to surface immobilized NepR by SPR. The equilibrium affinity of PhyR~P for NepR is 4400 times higher (12 pM) than unphosphorylated PhyR (Figure 4). The observed increase in affinity is determined by a dissociation rate constant (Kd) that slows from 2.8 x 10-4 s-1 to 6.2 x 10-8 s-1. Thus the in vitro half lifetime of the PhyR~P/NepR complex is 129 days. So, as in other species, phosphorylation of B. abortus PhyR increases its affinity for NepR. However, the absolute affinity of PhyR for NepR is substantially higher both in its phosphorylated and unphosphorylated states than PhyR/NepR pairs from other α-proteobacteria (16,33). Moreover, the measured association and dissociation rate constants for these B. abortus proteins are orders of magnitude slower than what our group has previously measured on orthologous proteins from Caulobacter crescentus (33). The slow binding kinetics and high affinities reported here were consistent across three replicate experiments using independent protein preparations. Growth phase-dependent regulation of PhyR protein concentration
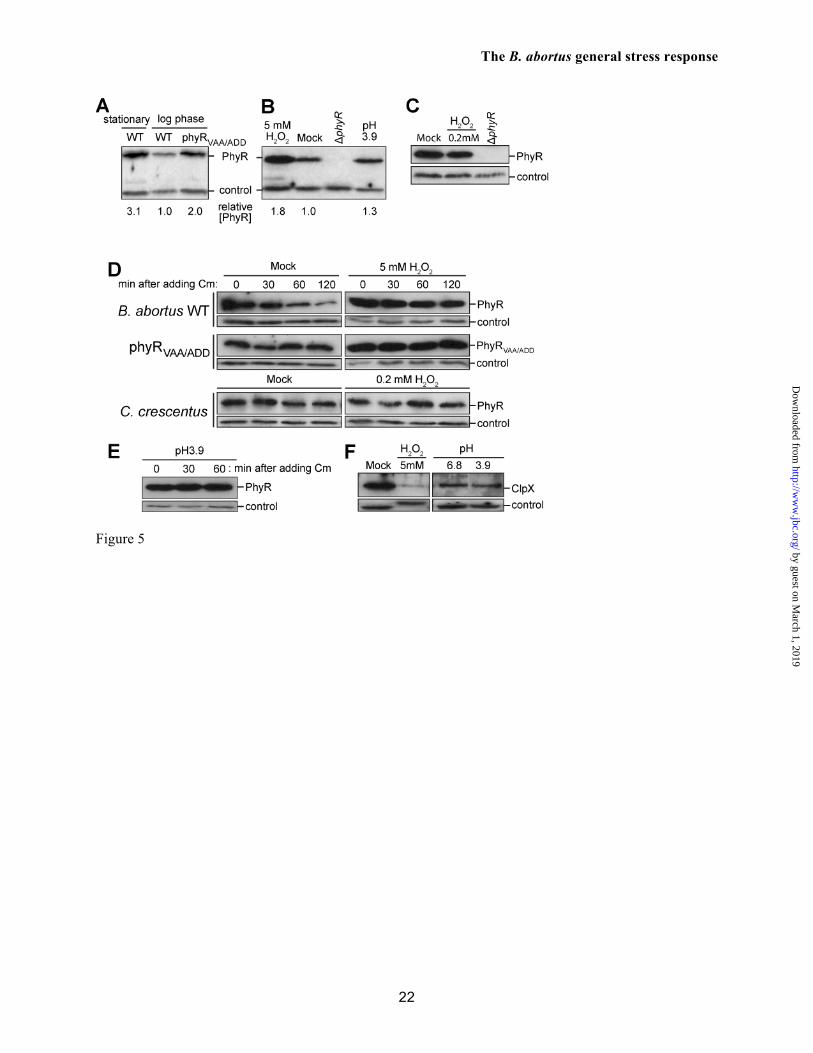
As detailed above, the anti-σ factor NepR forms a high-affinity complex with unphosphorylated PhyR. This result suggests that PhyR protein levels should be regulated in the cell to avoid constitutive NepR binding and activation of σE1-dependent transcription. We tested whether PhyR protein levels are regulated in B. abortus under different growth conditions. PhyR protein level, as assessed by Western blot, is 3-fold lower
in logarithmic growth phase relative to stationary phase (normalized to loading control bands) (Figure 5A). These data provide evidence that PhyR protein concentration is regulated as a function of growth phase.
There are multiple ways by which PhyR concentration could be regulated in the B. abortus cell including modulation of transcription, translation, or protein stability. We observed that B. abortus PhyR harbors a C-terminal Val-Ala-Ala sequence (Figure S5), which resembles a ssrA-like degron for ClpX-dependent protein degradation (34). Therefore, we tested whether the level of PhyR in the cell is affected by the presence of this putative ClpXP proteolysis tag at the C-terminus. Replacement of wild-type phyR on chromosome 1 with a mutant allele that encodes Ala-Asp-Asp at its C-terminus phyRVAA/ADD (V262A, A263D, A264D) results in a 2-fold increase in the steady-state concentration of PhyR in the logarithmic phase of growth relative to a loading control band (Figure 5A). This result is consistent with a model in which ClpXP-dependent proteolysis is involved in constitutive PhyR protein turnover in the unstressed, log phase B. abortus cell. Stress-dependent inhibition of PhyR protein turnover in B. abortus
We subjected stationary phase B. abortus cells to the same oxidative and acid stress conditions assayed in our cell survival experiments (see Figure 1B) and measured PhyR protein levels in B. abortus cells. The steady-state concentration of PhyR protein showed a 1.8-fold increase under peroxide stress relative to the loading control (Figure 5B). As a cross-species comparison, we measured C. crescentus PhyR levels under peroxide stress but observed no measureable increase in PhyR protein (Figure 5C). phyR transcription did not increase during peroxide treatment in B. abortus (Figure S6), demonstrating that increased transcription is not a primary mechanism controlling increased PhyR protein during oxidative stress. As such, we assessed the effect of stress treatment on PhyR protein stability. In these experiments, we inhibited PhyR protein synthesis with chloramphenicol, and measured PhyR decay upon treatment with peroxide or acid stress; PhyR protein decay was compared to a mock treated control by Western blot. In the control, PhyR decays with an approximate 60-
minute half lifetime. The PhyRVAA/ADD tail mutant protein is stable across the same time scale, providing additional support for a model in which PhyR is degraded by ClpXP (Figure 5D; Figure S7). In contrast, both PhyR and PhyRVAA/ADD proteins were stable after peroxide treatment (Figure 5D; Figure S7). PhyR protein was similarly stabilized in cells exposed to acid pH (Figure 5E). From this, we conclude that oxidative and acid stresses stabilize PhyR against proteolysis.
There are several possible mechanisms by which PhyR protein may be stabilized during stress. It is known that oxidative damage to a Cys4-containing zinc-binding domain (ZBD) in Bacillus subtilis ClpX results in protein degradation in vivo (35). As B. abortus ClpX also bears a Cys4 ZBD at its amino terminus, we tested whether ClpX degradation is correlated with increased PhyR protein during H2O2 stress. Treatment of B. abortus cells with 5 mM H2O2 resulted in a sharp decrease in the level of ClpX protein detected by Western blot (Figure 5F). Acid treatment (pH 3.9) had no effect on ClpX protein level (Figure 5F). From this, we conclude that the specific increase in steady-state PhyR levels that we observe in H2O2 stress (Figure 5B) is likely affected by the degradation of ClpX. However, there must be additional mechanisms beyond ClpX-dependent proteolysis that control stabilization of PhyR during stress; ClpX is not degraded after acid stress and yet PhyR stability increases under this condition (Figure 5E).
The carboxy terminus of C. crescentus PhyR does not have an apparent ssrA-like ClpX degron, and we did not observe an increase in steady-state PhyR level upon H2O2 stress treatment (Figure 5C), or PhyR degradation in C. crescentus in the absence of stress (Figure 5D). Thus, the regulated PhyR proteolysis that we observe in B. abortus is likely not a universal feature of the α-proteobacterial general stress response system.
Functional roles of PhyR proteolysis and phosphorylation in the B. abortus GSR
As unphosphorylated PhyR binds NepR with high affinity (Figure 4), we predicted that inhibition of PhyR proteolysis will increase the steady-state level of PhyR protein in the cell, and
thereby increase basal transcription of σE1-regulated genes in the absence of stress. To test this hypothesis, we compared the level of two GSR-regulated transcripts, rpoH1 and dps, from wild-type and the B. abortus phyRVAA/ADD mutant strain that expresses stable PhyR protein. The level of rpoH1 and dps mRNA was significantly higher, 3- and 8-fold respectively, in B. abortus phyRVAA/ADD relative to wild-type in the absence of stress treatment (Figure 6A). These data provide evidence that PhyR proteolysis has a functional role in the control of σE1-dependent gene transcription in B. abortus. We further tested the effect of PhyR proteolysis on cell adaptation to stress. However, we did not observe a difference in survival of wild-type versus B. abortus phyRVAA/ADD under oxidative stress (Figure 1B).
Finally, we sought to test the role of PhyR phosphorylation in the regulation of PhyR function as a stress regulator, and in the control of PhyR stability in vivo. As in other species, the SPR binding assays presented in Figure 4 clearly demonstrate that PhyR phosphorylation increases its affinity for NepR. We replaced the wild-type chromosomal copy of phyR with mutant alleles in which the conserved aspartyl phosphorylation site was mutated to an alanine (D191A) or glutamic acid (D191E). Both of these mutant strains were defective in oxidative stress survival relative to wild-type B. abortus (Figure 6). This defect is likely due to decreased levels of these mutant PhyR proteins in the B. abortus cell (Figure S8). However, these data do not permit us to discern the effect of D191 on phyR expression versus PhyR stability. Nonetheless, we can conclude that the conserved aspartyl phosphorylation site D191 is required for the appropriate regulation of stress physiology in vivo. DISCUSSION
Bacterial stress physiology is controlled at multiple levels, from gene transcription to the post-translational regulation of protein-protein interactions and protein stability. Examples of these layers of control are well described in the rpoS- and sigB-dependent general stress response systems of select Gram-negative (24) and Gram-positive species (36), respectively. Until very recently, a functional analog of the rpoS and sigB systems was unknown in the α-proteobacterial clade. It is now known that α-proteobacterial
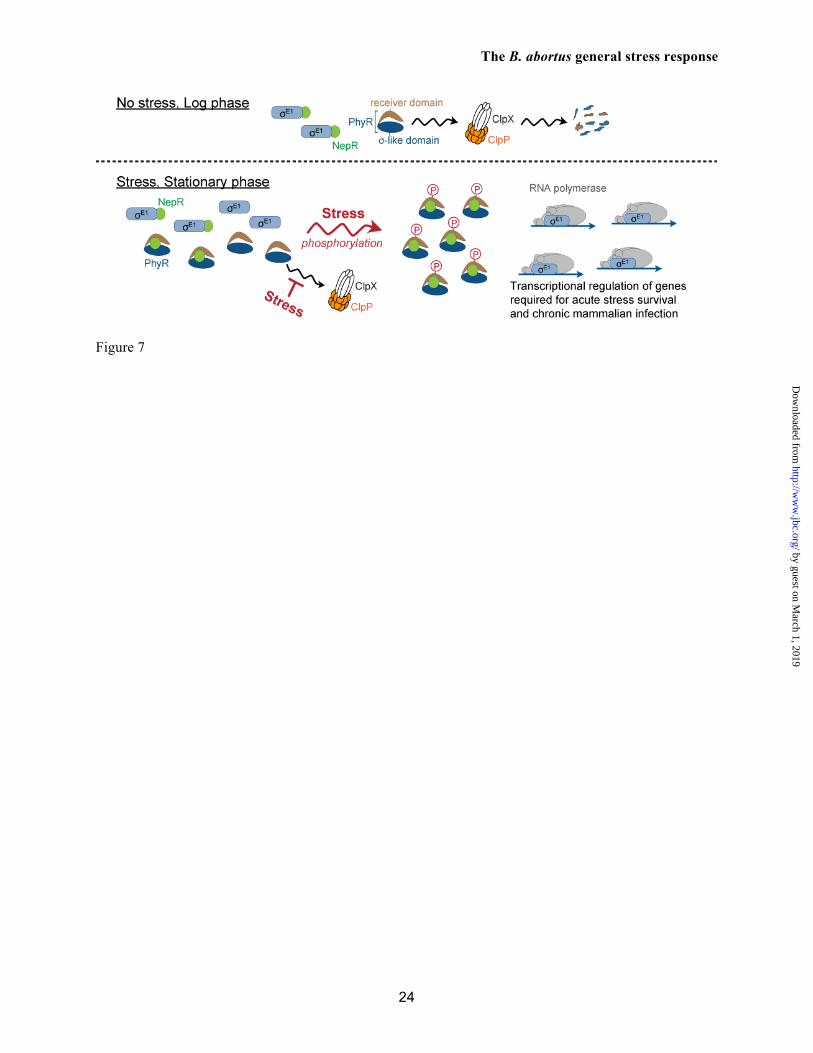
species encode a novel regulatory system in which the hybrid σ-like/receiver protein, PhyR, functions as a positive regulator of the σEcfG-dependent general stress response (GSR) (10). Specifically, phosphorylated PhyR (PhyR~P) activates an extracytoplasmic function (ECF) sigma factor of the EcfG family (32) by directly binding the anti-σEcfG protein, NepR (10). In other words, PhyR is a phosphorylation-dependent anti-anti-σ factor.
The mechanism of PhyR-dependent stress signaling has been assessed in several free-living and plant-associated α-proteobacteria (10,12-15,37). However, within this clade there are species that primarily inhabit mammalian host cells and cause disease in livestock and in humans. Among these animal pathogens, species of the genus Brucella contain a genetic locus that is predicted to encode a fully-intact GSR system including phyR, nepR, and the ecfG-family sigma factor known as rpoE1. The function of these putative GSR genes in Brucella spp. was largely uncharacterized prior to this study, though it has been reported that a B. melitensis rpoE1 mutant negatively regulates flagellar biosynthesis (18) and is attenuated at 4 weeks post-infection in a mouse model (17). In this study, we have defined a functional role for the B. abortus GSR system in response to acute acid and oxidative stress in vitro, and in control of chronic infection in mice (i.e. maintenance of B. abortus within a murine host >1 month post-infection). In addition to our functional analysis of the system, we have identified unique features of the molecular interactions among the core GSR proteins, PhyR, NepR, and σE1 and demonstrated a role for PhyR proteolysis in the control of GSR transcription in B. abortus. Regulated PhyR proteolysis is a GSR control feature in B. abortus.
Previous biochemical analyses of the interaction between PhyR and the anti-σ factor, NepR support a model in which high-affinity association between these proteins requires aspartyl phosphorylation of the PhyR receiver domain (10,12,13,15,16,33). The equilibrium affinity of PhyR~P for NepR has been measured in the tens (16) to hundreds (33) nanomolar range, using purified proteins from Sphingomonas sp. Fr1 and Caulobacter crescentus, respectively. The
extent to which PhyR phosphorylation increases its affinity for NepR has not been described prior to this study, though Herrou et al. reported no measureable binding between C. crescentus PhyR and NepR in SPR experiments in which they immobilized MBP-NepR and flowed PhyR at concentrations up to 3 µM (33).
When we investigated the interaction between B. abortus PhyR and NepR, we observed an unexpected high-affinity interaction between unphosphorylated PhyR and NepR. Specifically, PhyR co-purified with NepR by affinity chromatography and size exclusion chromatography (Figure 3 and S3). This high-affinity interaction was confirmed in SPR experiments in which we measured a PhyR/NepR equilibrium binding affinity of 57 nM (Figure 4). Surprisingly, the measured affinity between NepR and its cognate ECF sigma factor, σE1 is in the same range: 67 nM. Though PhyR phosphorylation does increase its affinity for NepR by ≈5000 fold, the similar affinity of NepR for both unphosphorylated PhyR and σE1 suggested to us that PhyR concentration must be tightly regulated in B. abortus for the GSR to function properly. Specifically, these data point to a model in which low cellular levels of PhyR are required to avoid constitutive PhyR/NepR binding and, hence, constitutive activation of σE1-dependent transcription.
To test this hypothesis, we measured in vivo levels of PhyR by Western blot under multiple growth and stress conditions. The concentration of PhyR increased in stationary phase relative to log phase, and also increased upon stress treatment (Figure 5). Mutation of a ssrA-like proteolysis degron (34) at the carboxy terminus of PhyR (Val-Ala-Ala) increased steady-state levels of PhyR protein in log phase and expression of two σE1-regulated genes , rpoH1 and dps. These data provide evidence that B. abortus utilizes a ClpXP-dependent mechanism to control cytosolic PhyR concentration and, hence, basal σE1-dependent transcription.
Both acid and oxidative stress treatments increased the stability of PhyR protein: PhyR has a half-lifetime of approximately 60 minutes in unstressed liquid culture, but is stable over a two-hour window after stress treatment (Figure 5D). Mutation of the PhyR C-terminus from VAA to
ADD results in complete stabilization of the protein across a two-hour window, again providing evidence for a ClpX-dependent mechanism of PhyR degradation.
Our data demonstrate that the PhyR protein is stabilized under stress conditions, and that phospho-PhyR and NepR form an extremely high affinity complex with a long dissociation half-life (Figure 4). This suggests a model in which Brucella abortus PhyR, once activated by phosphorylation, would lock a cell into a state in which σE1 is permanently active as a transcriptional regulator. The lifetime of this active state may be longer than the lifetime of the cell, which could provide B. abortus with a “memory” of prior stress encounters and thus preadapt the cell to future endogenous (38) or exogenous stresses encountered during host residence. However, PhyR dephosphorylation mechanisms may exist in B. abortus that switch the system off, as has been outlined in Sphingomonas sp. Fr1 (14) and C. crescentus (39). The B. abortus GSR and chronic infection
In the process of animal infection, Brucella sp. must survive a range of host-generated stressors (reviewed in ref. (25)). Given our discovery that B. abortus strains lacking an intact GSR system are deficient in survival under acute oxidative and acid stress in vitro (Figure 1B), we initially predicted that there would be some defect in the acute phase of animal infection where the B. abortus cell is exposed to oxidative burst and acidification of the early phagosome. However, our data indicate that phyR and rpoE1 are dispensable in the context of initial colonization of both murine spleens (Figure 1 C and D) and primary murine macrophages (Figure S2). These results provide evidence that other stress adaptation systems not under the control of PhyR-NepR-σE1 are sufficient to mediate bacterial cell survival in the face of stresses encountered during the first month of infection; it is only during the chronic phase of infection that we observe a difference in the rate of clearance of B. abortus ΔphyR and ΔrpoE1 strains relative to wild-type.
We envision two models by which the B. abortus GSR system mediates long-term cell survival in an animal host: 1) GSR is required for adaptation to a stress condition(s) that is unique to
the chronic phase of infection or 2) the B. abortus GSR system regulates genes that are required for subversion or evasion of host immunity during the chronic stage. In regard to the latter model, our microarray analysis confirms that rpoE1 controls expression of genes required for flagellum biosynthesis. It is known that expression of this classical pathogen-associated molecular pattern (PAMP) is required for B. melitensis host persistence (40) and interacts with the host innate immune system (41). rpoE1 also regulates genes involved in synthesis of the LPS O-chain, which has known immunomodulatory properties (42). Of course, neither of these models is mutually exclusive: regulation of chronic infection by the GSR system may involve changes in both Brucella stress physiology and modulation of its interactions with multiple arms of the host immune system, thus ensuring it remains weakly immunogenic (43) throughout the course of infection. Future study of this regulatory system
will provide insight into host side stresses and signals that are unique to the chronic phase of infection, and to which Brucella spp. must adapt in order to maintain long-term host colonization.
Acknowledgements The authors acknowledge support for this project from the NIH-NIAID Regional Center of Excellence for Biodefense and Emerging Infectious Diseases Research (RCE) Program (Region V ‘Great Lakes’ RCE; NIH award 1U54AI057153) and NIAID (award R01AI048499). We thank Aretha Fiebig for experimental suggestions and for comments on drafts of this manuscript, and Paul Dunman for the design and construction of the microarray chips. The support of the biophysics core facility at the University of Chicago, in particular Elena Solomaha, is greatly appreciated.
1. Pappas, G., Papadimitriou, P., Akritidis, N., Christou, L., and Tsianos, E. V. (2006) The new global map of human brucellosis. Lancet Infect Dis 6, 91-99
2. Roop, R. M., Bellaire, B. H., Valderas, M. W., and Cardelli, J. A. (2004) Adaptation of the brucellae to their intracellular niche. Mol Microbiol 52, 621-630
3. Arenas, G. N., Staskevich, A. S., Aballay, A., and Mayorga, L. S. (2000) Intracellular trafficking of Brucella abortus in J774 macrophages. Infect Immun 68, 4255-4263
4. Naroeni, A., Jouy, N., Ouahrani-Bettache, S., Liautard, J. P., and Porte, F. (2001) Brucella suis-impaired specific recognition of phagosomes by lysosomes due to phagosomal membrane modifications. Infect Immun 69, 486-493
5. Celli, J., de Chastellier, C., Franchini, D.-M., Pizarro-Cerda, J., Moreno, E., and Gorvel, J.-P. (2003) Brucella evades macrophage killing via VirB-dependent sustained interactions with the endoplasmic reticulum. J Exp Med 198, 545-556
6. Moreno, E., and Moriyon, I. (2006) The Genus Brucella. in The Prokaryotes: A Handbook on the Biology of Bacteria (Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., and Stackebrandt, E. eds.), Springer, New York. pp
7. Solera, J. (2010) Update on brucellosis: therapeutic challenges. Int J Antimicrob Agents 36, S18-20
8. Castano, M. J., and Solera, J. (2009) Chronic brucellosis and persistence of Brucella melitensis DNA. J Clin Microbiol 47, 2084-2089
9. Köhler, S., Michaux-Charachon, S., Porte, F., Ramuz, M., and Liautard, J. P. (2003) What is the nature of the replicative niche of a stealthy bug named Brucella? Trends Microbiol 11, 215-219
10. Francez-Charlot, A., Frunzke, J., Reichen, C., Ebneter, J. Z., Gourion, B., and Vorholt, J. A. (2009) Sigma factor mimicry involved in regulation of general stress response. Proc Natl Acad Sci USA 106, 3467-3472
11. Gourion, B., Francez-Charlot, A., and Vorholt, J. A. (2008) PhyR is involved in the general stress response of Methylobacterium extorquens AM1. J Bacteriol 190, 1027-1035
12. Bastiat, B., Sauviac, L., and Bruand, C. (2010) Dual control of Sinorhizobium meliloti RpoE2 sigma factor activity by two PhyR-type two-component response regulators. J Bacteriol 192, 2255-2265
13. Gourion, B., Sulser, S., Frunzke, J., Francez-Charlot, A., Stiefel, P., Pessi, G., Vorholt, J. A., and Fischer, H.-M. (2009) The PhyR-σEcfG signalling cascade is involved in stress response and symbiotic efficiency in Bradyrhizobium japonicum. Mol Microbiol 73, 291-305
14. Kaczmarczyk, A., Campagne, S., Danza, F., Metzger, L. C., Vorholt, J. A., and Francez-Charlot, A. (2011) Role of Sphingomonas sp. strain Fr1 PhyR-NepR-sigmaEcfG cascade in general stress response and identification of a negative regulator of PhyR. J Bacteriol 193, 6629-6638
15. Lourenço, R. F., Kohler, C., and Gomes, S. L. (2011) A two-component system, an anti-sigma factor and two paralogous ECF sigma factors are involved in the control of general stress response in Caulobacter crescentus. Mol Microbiol 80, 1598-1612
16. Campagne, S., Damberger, F. F., Kaczmarczyk, A., Francez-Charlot, A., Allain, F. H., and Vorholt, J. A. (2012) Structural basis for sigma factor mimicry in the general stress response of Alphaproteobacteria. Proc Natl Acad Sci U S A 109, E1405-1414
17. Delory, M., Hallez, R., Letesson, J. J., and De Bolle, X. (2006) An RpoH-like heat shock sigma factor is involved in stress response and virulence in Brucella melitensis 16M. J Bacteriol 188, 7707-7710
18. Ferooz, J., Lemaire, J., Delory, M., De Bolle, X., and Letesson, J. J. (2011) RpoE1, an extracytoplasmic function sigma factor, is a repressor of the flagellar system in Brucella melitensis. Microbiology 157, 1263-1268
19. Robertson, G. T. (2000) The Brucella abortus Lon and ClpXP proteases participate in adaptation to conditions encountered in the murine host. in Department of Microbiology and Immunology, Louisiana State University Health Sciences Center, Shreveport
20. Fiebig, A., Castro Rojas, C. M., Siegal-Gaskins, D., and Crosson, S. (2010) Interaction specificity, toxicity and regulation of a paralogous set of ParE/RelE-family toxin–antitoxin systems. Mol Microbiol 77, 236-251
21. Ried, J. L., and Collmer, A. (1987) An nptI-sacB-sacR cartridge for constructing directed, unmarked mutations in gram-negative bacteria by marker exchange-eviction mutagenesis. Gene 57, 239-246
22. Gerhardt, P. (1958) The nutrition of brucellae. Microbiol Rev 22, 81-98 23. Schuck, P. (2000) Size-distribution analysis of macromolecules by sedimentation velocity
ultracentrifugation and lamm equation modeling. Biophys J 78, 1606-1619 24. Battesti, A., Majdalani, N., and Gottesman, S. (2011) The RpoS-mediated general stress response
in Escherichia coli. Annu Rev Microbiol 65, 189-213 25. Roop, R. M., Gaines, J. M., Anderson, E. S., Caswell, C. C., and Martin, D. W. (2009) Survival
of the fittest: how Brucella strains adapt to their intracellular niche in the host. Med Microbiol Immunol 198, 221-238
26. Endley, S., McMurray, D., and Ficht, T. A. (2001) Interruption of the cydB locus in Brucella abortus attenuates intracellular survival and virulence in the mouse model of infection. J Bacteriol 183, 2454-2462
27. Sangari, F. J., Seoane, A., Rodriguez, M. C., Aguero, J., and Garcia Lobo, J. M. (2007) Characterization of the urease operon of Brucella abortus and assessment of its role in virulence of the bacterium. Infect Immun 75, 774-780
28. Vemulapalli, T. H., Vemulapalli, R., Schurig, G. G., Boyle, S. M., and Sriranganathan, N. (2006) Role in virulence of a Brucella abortus protein exhibiting lectin-like activity. Infect Immun 74, 183-191
29. Ugalde, J. E., Czibener, C., Feldman, M. F., and Ugalde, R. A. (2000) Identification and characterization of the Brucella abortus phosphoglucomutase gene: role of lipopolysaccharide in virulence and intracellular multiplication. Infect Immun 68, 5716-5723
30. Ugalde, J. E., Comerci, D. J., Leguizamon, M. S., and Ugalde, R. A. (2003) Evaluation of Brucella abortus phosphoglucomutase (pgm) mutant as a new live rough-phenotype vaccine. Infect Immun 71, 6264-6269
31. Bailey, T. L., and Gribskov, M. (1998) Combining evidence using p-values: application to sequence homology searches. Bioinformatics 14, 48-54
32. Staroń, A., Sofia, H. J., Dietrich, S., Ulrich, L. E., Liesegang, H., and Mascher, T. (2009) The third pillar of bacterial signal transduction: classification of the extracytoplasmic function (ECF) σ factor protein family. Mol Microbiol 74, 557-581
33. Herrou, J., Rotskoff, G., Luo, Y., Roux, B., and Crosson, S. (2012) Structural basis of a protein partner switch that regulates the general stress response of alpha-proteobacteria. Proc Natl Acad Sci U S A 109, E1415-1423
34. Flynn, J. M., Neher, S. B., Kim, Y.-I., Sauer, R. T., and Baker, T. A. (2003) Proteomic discovery of cellular substrates of the ClpXP protease reveals five classes of ClpX-recognition signals. Mol Cell 11, 671-683
35. Zhang, Y., and Zuber, P. (2007) Requirement of the zinc-binding domain of ClpX for Spx proteolysis in Bacillus subtilis and effects of disulfide stress on ClpXP activity. J Bacteriol 189, 7669-7680
36. Hecker, M., Pane-Farre, J., and Volker, U. (2007) SigB-dependent general stress response in Bacillus subtilis and related gram-positive bacteria. Annu Rev Microbiol 61, 215-236
37. Herrou, J., Foreman, R., Fiebig, A., and Crosson, S. (2010) A structural model of anti-anti-σ inhibition by a two-component receiver domain: the PhyR stress response regulator. Mol Microbiol 78, 290-304
38. Steele, K. H., Baumgartner, J. E., Valderas, M. W., and Roop, R. M., 2nd. (2010) Comparative study of the roles of AhpC and KatE as respiratory antioxidants in Brucella abortus 2308. J Bacteriol 192, 4912-4922
39. Foreman, R., Fiebig, A., and Crosson, S. (2012) The LovK-LovR two-component system is a regulator of the general stress pathway in Caulobacter crescentus. J Bacteriol 194, 3038-3049
40. Fretin, D., Fauconnier, A., Kohler, S., Halling, S., Leonard, S., Nijskens, C., Ferooz, J., Lestrate, P., Delrue, R. M., Danese, I., Vandenhaute, J., Tibor, A., DeBolle, X., and Letesson, J. J. (2005) The sheathed flagellum of Brucella melitensis is involved in persistence in a murine model of infection. Cell Microbiol 7, 687-698
41. Terwagne, M., Ferooz, J., Rolan, H. G., Sun, Y. H., Atluri, V., Xavier, M. N., Franchi, L., Nunez, G., Legrand, T., Flavell, R. A., De Bolle, X., Letesson, J. J., and Tsolis, R. M. (2012) Innate immune recognition of flagellin limits systemic persistence of Brucella. Cell Microbiol doi: 10.1111/cmi.12088
42. Gorvel, J. P., and Moreno, E. (2002) Brucella intracellular life: from invasion to intracellular replication. Vet Microbiol 90, 281-297
43. Barquero-Calvo, E., Chaves-Olarte, E., Weiss, D. S., Guzman-Verri, C., Chacon-Diaz, C., Rucavado, A., Moriyon, I., and Moreno, E. (2007) Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PLoS One 2, e631
44. Sun, Y. H., de Jong, M. F., den Hartigh, A. B., Roux, C. M., Rolan, H. G., and Tsolis, R. M. (2012) The small protein CydX is required for function of cytochrome bd oxidase in Brucella abortus. Front Cell Infect Microbiol 2, 47
FIGURE LEGENDS Figure 1. B. abortus phyR and rpoE1 are required for in vitro stress survival and chronic infection of a mammalian host. (A) Canonical model of PhyR-dependent regulation of transcription during stress (based on ref. 10). The hybrid response regulator, PhyR, is composed of a N-terminal σ-like domain (orange) and C-terminal receiver (Rec) domain (blue). Stress-dependent phosphorylation of PhyR increases its affinity for the anti-σ factor, NepR (green). PhyR~P/NepR binding releases an EcfG-family σ factor (σE1: orange) to bind RNA polymerase and direct transcription. (B) Survival of wild type B. abortus 2308, ΔrpoE1, ΔnepR-rpoE1, ΔphyR, and phyRVAA/ADD strains upon oxidative and acid stress treatment in stationary and logarithmic growth phases. ΔphyR and ΔnepR-rpoE1 null phenotypes are complemented by single copy integration of these genes plus their native promoter into the chromosome (phyR+ and nepR-rpoE1+). Cell viability was assessed by measuring colony forming units (CFU) after 1 hr of oxidative (5 mM H2O2) or 2 hrs acid (pH 3.9) treatment and comparing to mock treated controls. Error bars represent standard deviation of at least 3 independent replicates. Statistical significance of the survival difference between wild-type B. abortus and ΔphyR was assessed by one-way ANOVA followed by Dunnett’s post comparison test to wild-type (** p < 0.001 between wild-type and ΔphyR). (C and D) Kinetics of BALB/c mouse spleen colonization by wild-type B. abortus 2308, and its derivatives ΔrpoE1 and ΔphyR. Mice were infected intraperitoneally with ≈5 x 104 CFU and spleens were collected at the indicated time points. Error bars indicate standard deviation. Statistical significance of the spleen colonization difference between wild-type B. abortus and mutants was assessed by one-way ANOVA followed by Bonferroni’s multiple comparison test (p > 0.05 at week 2 and week 4; ** p < 0.001 at week 8 and week 12). Data are representative of two independent experiments with five mice per time point. Figure 2. Biophysical analysis of purified B. abortus His6-NepR-σE1complex and His6-PhyR. (A) Organization of the B. abortus gene cluster encoding PhyR, NepR, σE1, and two predicted histidine kinases. (B) Size-exclusion chromatography (SEC) elution profile of NepR-σE1 complex; (C) c(S) distribution calculated from sedimentation velocity measurements of the NepR-σE1 complex. (D) SEC elution profile of PhyR; (E) c(S) distribution calculated from sedimentation velocity measurements of PhyR. The estimated molecular mass based on SEC elution standards or c(M) calculations in Sedfit are indicated above the peaks; RMSD of the fit sedimentation data is marked on panels C and E. Stars denote the SEC fractions analyzed by 14% SDS-PAGE. Figure 3. Atypical interactions between B. abortus NepR and unphosphorylated PhyR. PhyR elutes as a single peak by size-exclusion chromatography (SEC) (pink line; 10.9 ml); NepR elutes as multiple conformational species (dashed green line; 10.9 and 11.8 ml); an equimolar mixture of individually-purified NepR and PhyR (10 uM each) elutes as a single peak with a volume shift that is consistent with complex formation (10.7 ml). Stars indicate the fractions analyzed by 14% SDS-PAGE (inset). Figure 4. B. abortus NepR forms a high-affinity complex with unphosphorylated PhyR; phosphorylation of PhyR yields an extremely high-affinity, long-lived PhyR~P/NepR complex. Sensorgrams and fit residuals from surface plasmon resonance (SPR) experiments measuring association and dissociation of surface-immobilized NepR to analyte proteins PhyR, phospho-PhyR (PhyR~P) and σE1. Immobilized ligand (Rligand) ≈ 400 RU for PhyR and PhyR~P binding experiments; Rligand ≈ 100 for σE1 binding experiment. Analytes were flowed at marked concentrations. Black lines indicate global fit of association and dissociation curves using a 1:1 binding model (Rmax local) with drifting baseline. Table presents equilibrium affinities (KD) calculated from association (Ka) and dissociation (Kd) rate constants. Figure 5. Regulated proteolysis of PhyR protein and mutant variants. (A) Immunoblot analysis of B. abortus wild-type PhyR and the C-terminal mutant variant PhyRVAA/ADD in stationary growth phase and logarithmic phase. PhyR concentration [PhyR] relative to WT log phase cells is normalized to loading
control band (B) Blot measuring B. abortus PhyR in oxidative stress (5 mM H2O2) and acid stress (pH 3.9) versus a mock treated control; a ΔphyR control is included. PhyR concentration [PhyR] relative to mock treated cells is normalized to loading control band. (C) Immunoblot of C. crescentus wild-type PhyR in the presence of an activating peroxide stress (0.2 mM H2O2) versus a mock treated control. (D; top) To probe PhyR stability in vivo, B. abortus cells from early stationary phase were treated 10 minutes post-stress with chloramphenicol (Cm) to halt protein synthesis. PhyR decay was assessed by immunoblot using protein samples collected across a 120-minute window (after Cm treatment). PhyR and PhyRVAA/ADD protein decay as a function of 5 mM H2O2 treatment was compared to a mock treated control. (D; bottom) As above, C. crescentus PhyR stability was assessed across a 120-minute window (after Cm treatment). Protein decay was measured as a function of peroxide stress treatment. (E) B. abortus PhyR stability was assessed by immunoblot as a function of acid treatment (pH 3.9) across a 60-minute window (after Cm treatment). (F) B. abortus ClpX protein as a function of 5 mM H2O2 treatment and acid pH treatment (pH 3.9) was assessed by immunoblot. In all immunoblots, a non-specific loading control band is shown below the PhyR band. Figure 6. Assessing the functional role of PhyR degradation and phosphorylation. (A) Relative rpoH1 and dps transcript levels in phyRVAA/ADD versus wild-type (WT) strains during unstressed logarithmic growth. Transcript levels were measured by qRT-PCR, and normalized to rplK mRNA. (B) Cell viability of B. abortus 2308 and mutant derivatives upon treatment with 5 mM H2O2. Assessed strains include the ΔphyR in-frame deletion mutant and strains in which the wild-type phyR allele was replaced with aspartyl phosphorylation site point mutants, D191A and D191E. Survival was measured relative to a mock treated control. Figure 7. Proposed molecular model of the PhyR-NepR-σE1 GSR regulatory system of B. abortus. PhyR protein concentration is maintained at lower levels in the absence of stress by an ATP-dependent protease, ClpXP; PhyR proteolysis helps to ensure NepR (orange) remains bound to σE1 (blue). Under stress and stationary growth conditions, the PhyR is phosphorylated, stabilized, and binds NepR. This releases σE1 to activate transcription of genes required for stress survival and chronic animal infection.
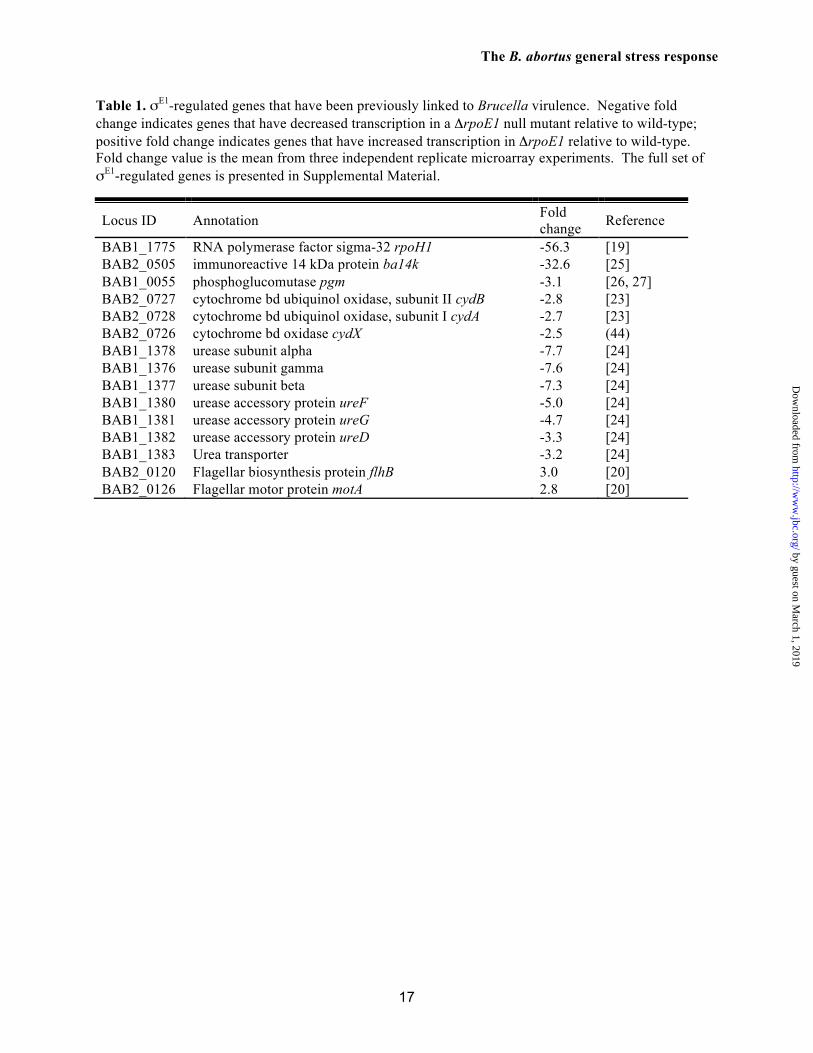
Table 1. σE1-regulated genes that have been previously linked to Brucella virulence. Negative fold change indicates genes that have decreased transcription in a ΔrpoE1 null mutant relative to wild-type; positive fold change indicates genes that have increased transcription in ΔrpoE1 relative to wild-type. Fold change value is the mean from three independent replicate microarray experiments. The full set of σE1-regulated genes is presented in Supplemental Material.