Anita. Behav., 1982,30, 35-42 THE CHEMOSENSORY RESPONSES OF YOUNG CICHLID FISH TO PARENTS AND PREDATORS By COLIN BARNETT* Museum of Vertebrate Zoology, University of California, Berkeley, CA 94720 Abstract. The chemosensory responses of the fry of the Midas cichlid, Cichlasoma citrinellum (Pisces: Cichlidae), were tested in a Y-maze during their first 11 days of free-swimming. Fry preferred all con- specific females to blank water but did not distinguish between their own mother, another mother or a non-parental, adult female. Fry preferred females to males. Similar results were obtained for males. Fry preferred their siblings and (unrelated) juveniles to blank water. However, while they failed to show a preference between siblings and their mother, they preferred their mother to juveniles. Fry were able to distinguish the closely related red devil cichlid (C. labiatum) from blank water, but failed to do this for any of three heterospecifics (C. managuense, C, nigrofasciatum and the allopatric Sarotherodon mossambi- cus). The chemosensory responses demonstrated in the Y-maze are hypothesized to aid the fry in staying near the centre of the guarded territory and away from predators, particularly under murky or nocturnal conditions. Major studies of the development of parent- offspring communication have relied on birds and mammals (e.g. Beer 1979; Leon 1979), two groups which extend parental care beyond the egg stage. Parallel studies on a third major vertebrate group, the fishes, are possible only for those few fish families which also show complex care of post-egg offspring. One such family, Cichlidae, has provided almost by default the model for studies on the development of parent-offspring communication in fish. Cichlid fishes demonstrate efficient, persistent parental care towards their brood, which may consist of 5 to 10 000 fry, depending on the species (Fryer & Iles 1972; Brichard 1975). A primary function of this extended parental care is the defence of the fry from predators. If the parents of the biparental Midas cichlid, Cichlasoma citrinellum, are experimentally re- moved from a brood in the wild, their 1500 to 3000 fry will be consumed in minutes (McKaye & Hallacher 1973; Barlow 1976). Effective defence requires that fry stay near their guarding parents. Midas parents possess conspicuous black and white vertical stripes (Barlow 1976) which are presumed to function as warning coloration. However, these markings may also function as visual cues to attract the fry. In fact, vision is one of the primary senses by which cichlid parents (Noble & Curtis 1939; Myrberg 1975) and fry (yon Kuenzer 1968; Russock & Schein 1974; Hay 1978) recognize each other. Yet cichlid habitats can be turbid and murky *Present address: Departmentof Ethology, University of Bielefeld, P.O.Box 8640,4800Bielefeld 1, W. Germany. (Baylis 1974; Barlow 1976; Fernald & Hirata 1977) with visibility at times measured in centi- metres. Under these conditions, or at night, selection should favour fry who are able to stay with the brood using non-visual cues provided by chemoreception (Barnett 1977a), audition, or the lateral line system. The latter two channels have not been examined with respect to parent- offspring communication in cichlids. However, numerous studies have documented the ability of cichlid parents to recognize their fry (Kiihme 1963; Myrberg 1975; McKaye & Barlow 1976) or each other (Caprona 1974) using chemical cues alone. Barnett (1977b) provided evidence of the reciprocal ability of the fry to respond to the chemical cues from one of their parents. Barnett (1977b) showed that the fry of the Midas cichlid chemically distinguish between their mother and plain water and between another mother and plain water. However, they make no distinction between their mother and a Midas cichlid mother tending another brood, nor bet- ween their father and blank water. The purpose of the present investigation is to repeat and extend this earlier study by assessing the fry's chemical responses not only to their parents, but also to siblings, conspecific and heterospecific predators, and to other Midas cichlid parents. The guiding hypotheses are that the responses of the fry to each class of stimuli increase its chance of survival in the wild. Alternative hypotheses are that responses based on chemoreception are irrelevant to the fry's survival and thus are not acted upon by natural selection. If such is the case the fry might show the same chemoreeeptive 35
Transcript
Anita. Behav., 1982, 30, 35-42
THE CHEMOSENSORY RESPONSES OF YOUNG CICHLID FISH TO PARENTS AND PREDATORS
By COLIN BARNETT* Museum of Vertebrate Zoology, University of California, Berkeley, CA 94720
Abstract. The chemosensory responses of the fry of the Midas cichlid, Cichlasoma citrinellum (Pisces: Cichlidae), were tested in a Y-maze during their first 11 days of free-swimming. Fry preferred all con- specific females to blank water but did not distinguish between their own mother, another mother or a non-parental, adult female. Fry preferred females to males. Similar results were obtained for males. Fry preferred their siblings and (unrelated) juveniles to blank water. However, while they failed to show a preference between siblings and their mother, they preferred their mother to juveniles. Fry were able to distinguish the closely related red devil cichlid (C. labiatum) from blank water, but failed to do this for any of three heterospecifics (C. managuense, C, nigrofasciatum and the allopatric Sarotherodon mossambi- cus). The chemosensory responses demonstrated in the Y-maze are hypothesized to aid the fry in staying near the centre of the guarded territory and away from predators, particularly under murky or nocturnal conditions.
Major studies of the development of parent- offspring communication have relied on birds and mammals (e.g. Beer 1979; Leon 1979), two groups which extend parental care beyond the egg stage. Parallel studies on a third major vertebrate group, the fishes, are possible only for those few fish families which also show complex care of post-egg offspring. One such family, Cichlidae, has provided almost by default the model for studies on the development of parent-offspring communication in fish.
Cichlid fishes demonstrate efficient, persistent parental care towards their brood, which may consist of 5 to 10 000 fry, depending on the species (Fryer & Iles 1972; Brichard 1975). A primary function of this extended parental care is the defence of the fry from predators. If the parents of the biparental Midas cichlid, Cichlasoma citrinellum, are experimentally re- moved from a brood in the wild, their 1500 to 3000 fry will be consumed in minutes (McKaye & Hallacher 1973; Barlow 1976).
Effective defence requires that fry stay near their guarding parents. Midas parents possess conspicuous black and white vertical stripes (Barlow 1976) which are presumed to function as warning coloration. However, these markings may also function as visual cues to attract the fry. In fact, vision is one of the primary senses by which cichlid parents (Noble & Curtis 1939; Myrberg 1975) and fry (yon Kuenzer 1968; Russock & Schein 1974; Hay 1978) recognize each other. Yet cichlid habitats can be turbid and murky
*Present address: Department of Ethology, University of Bielefeld, P.O. Box 8640, 4800 Bielefeld 1, W. Germany.
(Baylis 1974; Barlow 1976; Fernald & Hirata 1977) with visibility at times measured in centi- metres. Under these conditions, or at night, selection should favour fry who are able to stay with the brood using non-visual cues provided by chemoreception (Barnett 1977a), audition, or the lateral line system. The latter two channels have not been examined with respect to parent- offspring communication in cichlids. However, numerous studies have documented the ability of cichlid parents to recognize their fry (Kiihme 1963; Myrberg 1975; McKaye & Barlow 1976) or each other (Caprona 1974) using chemical cues alone. Barnett (1977b) provided evidence of the reciprocal ability of the fry to respond to the chemical cues from one of their parents.
Barnett (1977b) showed that the fry of the Midas cichlid chemically distinguish between their mother and plain water and between another mother and plain water. However, they make no distinction between their mother and a Midas cichlid mother tending another brood, nor bet- ween their father and blank water. The purpose of the present investigation is to repeat and extend this earlier study by assessing the fry's chemical responses not only to their parents, but also to siblings, conspecific and heterospecific predators, and to other Midas cichlid parents. The guiding hypotheses are that the responses of the fry to each class of stimuli increase its chance of survival in the wild. Alternative hypotheses are that responses based on chemoreception are irrelevant to the fry's survival and thus are not acted upon by natural selection. If such is the case the fry might show the same chemoreeeptive
35
36 A N I M A L B E H A V I O U R , 30 , 1
response to any fish, regardless of whether it is predator or protector.
Metheds Rearing
Pairs (mean standard length of males: 160 mm; females: 140 ram) of the normal morph (Barlow 1973) of the Midas cichlid, Cichlasoma citrinellum (Fe from Lake Masaya, Nicaragua) were bred in 240-1itre aquaria (26 :L 1 C; salinity negligible; pH 7; photoperiod: 0700 hours to 2200 hours light; feeding twice daily: adults, Kordon trout chow; fry, live Artemia nauplii). The water in the aquaria was recirculated through a gravel bed. Evaporative water losses were replenished with aerated, aged, charcoal- filtered water obtained from the municipal water supply in Berkeley, CA. Some breeding tanks were on a flow-through system and did not have gravel filtration.
Each pair laid about 1500 to 2000 eggs on the inside of a terra cotta flower pot. In about 3 days the eggs hatched into non-swimming wrigglers (eleutheroembryos; Balon 1975) which the parents placed on the gravel substrate. The wrigglers adhered in a tight mass to the substrate by means of mucosal threads protruding from their heads. In about 5 days the wrigglers became free-swimming fry (larvae; Balon 1975) (stand- ard length: 5 ram).
The parents, but primarily the female (see Discussion), fanned the eggs and wrigglers with pectoral fins and removed dead individuals with their mouths. When the fry were free-swimming, the parents retrieved those fry that wandered from the school. At night, the parents gathered the fry into a tight cluster underneath them in a small depression in the substrate. Fish demon- strating such care of offspring are referred to in this paper as parental,
transparent boxes gave a full view of the experi- menter and his movements. This no doubt stressed the fish. At times, those stimulus fish that were parental would even lose their parental coloration and adopt a blanched appearance. Surprisingly, the responses of the fry toward such apparently stressed fish seemed to be more consistent and reliable than when the stimulus fish were in the cave-like shelter. This effect was not investigated further in these experiments.
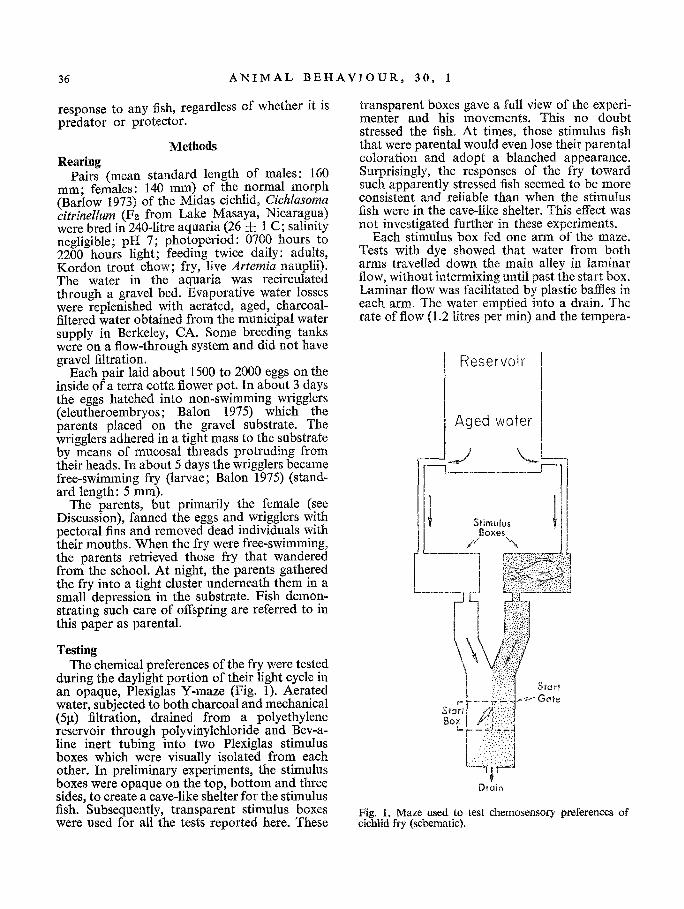
Each stimulus box fed one arm of the maze. Tests with dye showed that water from both arms travelled down the main alley in laminar flow, without intermixing until past the start box. Laminar flow was facilitated by plastic baffles in each arm. The water emptied into a drain. The rate of flow (1.2 litres per min) and the tempera-
Reservoir
Aged water
Boxes
Testing The chemical preferences of the fry were tested
during the daylight portion of their light cycle in an opaque, Plexiglas Y-maze (Fig. 1). Aerated water, subjected to both charcoal and mechanical (5Ix) filtration, drained from a polyethylene reservoir through polyvinylchloride and Bev-a- line inert tubing into two Plexiglas stimulus boxes which were visually isolated from each other. In preliminary experiments, the stimulus boxes were opaque on the top, bottom and three sides, to create a cave-like shelter for the stimulus fish. Subsequently, transparent stimulus boxes were used for all the tests reported here. These
Sf B~
Drain
Fig. 1. Maze used to test chemosensory preferences of cichlid fry (schematic).
BARNETT: CHEMOSENSORY RESPONSES OF CICHLID FISH 37
ture o f the water (26 C) enter ing each a r m of the maze were a b o u t equal .
Protocol Eighteen different tests, encompass ing a
var ie ty o f na tura l ly occurr ing stimuli , were presented to the fry (Table I). Idea l ly each test should have been presented to equivalent ly aged fry, bu t p rob lems o f scheduling and o f b reed ing synchrony prevented this. However , p i lo t experi- ments on fry ranging in age f rom 1 to 11 days f ree-swimming indica ted that , except for tests 9 and 10 (Table I), the age o f the fry at test ing had no effect on the results.
Before each test the st imulus fish was p laced in the st imulus box and a g roup o f 10 fry were r emoved f rom the rear ing aqua r ium into a hold- ing t ank o f clean water . Five minutes la ter the flow th rough the maze was star ted. One fry was
p laced in the s tar t box. Af te r 30 s, the a m o u n t o f t ime (to the neares t second) tha t the fry spent on the r ight , left or over the naidline o f the s ta r t box was recorded for the next 30 s; then the fry was d iscarded. Thus each fry was used only once. The fry was scored as showing a preference for the st imulus tha t p rov ided water to the side o f the s ta r t box on which the fry spent the mos t t ime. The number of fry preferr ing each side was c o m p a r e d using a Binomia l Test (Siegel 1956). I omi t t ed f rom the analysis fry who spent equal t ime on bo th sides. Pi lo t exper iments us ing dye revealed tha t the l aminar flow was res tored bet- ween the remova l o f one fry and the test ing o f the next one.
To increase the general i ty o f the conclusions, I used four b roods for tests 1 to 9 (Table I). A reduc t ion in the number o f avai lable breed ing pai rs forced me to use less than four b roods for
Table I. Chemosensory Tests Presented to Fry of Cichlasoma citrinellum
Fig. Test No.
No. of No. of fry broods tested tested* per broodt Age when tested:~
*Drawn randomly from a population of 29 pairs according to availability of fry from each pair. Hence pairs that spawned more often were used more frequently, although an effort was made to use as many different pairs as possible throughout the sequence of 18 tests. Four different spawning pairs were not available for tests 10 through 18.
tWith the switch to a more sensitive testing protocol (see Fig. caption 2A) after test 3, ten fry per brood became a suffi- ciently large sample size and reduced the time required foreach test. Test 14 was done when only one brood was avail- able; hence all fry had to come from that brood.
SNumbers are days free-swimming at the time of testing for each brood. w in parentheses are the ages of the stimulus fish's brood. **The approximately 3-month-old juveniles were significantly younger than the earliest known age at first reprodnction of the Midas cichlid (12 to 18 months), and were therefore considered sexually immature. They lacked the secondary sexual characteristics, including the sexually diagnostic shape of the genital papilla, which permitted sexing of the adults; hence they were not sexed. Three different juveniles were used for each test. tThe hetcrospecific fish were non-breeding adult males. (The sex of N. nematopus was not determined.)
38 A N I M A L B E H A V I O U R , 30 , 1
tests 10 to 18. Pre l iminary exper iments indica ted no consis tent differences between b roods for these tests; tha t is, no fry f rom any given b r o o d were consistent ly m o r e or less successful in mak ing discr iminat ions than fry f rom any other b rood . Therefore groups o f fry f rom more than one b r o o d were combined in the analysis. The number o f fry tested f rom each b r o o d (10) was t oo few to permi t a meaningful analysis o f the i n t r a - b r o o d var ia t ion. F o r each g roup o f fry f rom a single b rood , the pos i t ion o f the st imuli was reversed after ha l f the fry were tested by switching the hoses leading into the arms o f the maze. This ensured tha t i f fry were choos ing on the basis o f pos i t ion ra ther than st imulus, they would fail to demons t r a t e a significant preference for one s t imulus o r the other . Before switching hoses, the maze was r insed using warm water a n d p a p e r towels.
To test i f darkness would affect the chemosen- sory responses o f the fry, one exper iment (mother versus b l ank water) was pe r fo rmed dur ing the middle o f the f ry ' s da rk cycle. A dim red l ight was used to see the fry.
Results
F r y prefer red all females, pa ren ta l or not , and al l males, pa ren ta l or not , to b l ank water (Fig. 2). F r y fai led to show a preference between m o t h e r a n d a non-pa ren ta l female bu t preferred a non- pa ren ta l male to their fa ther (Fig. 3).
On the first day o f f ree-swimming, fry pre- fer red a female, whether it was their m o t h e r or a non-pa ren ta l fish, to a male, whether i t was their fa ther or a non-pa ren ta l fish (Fig. 4). F r y fai led to show a preference between the sexes on sub- sequent days free-swimming.
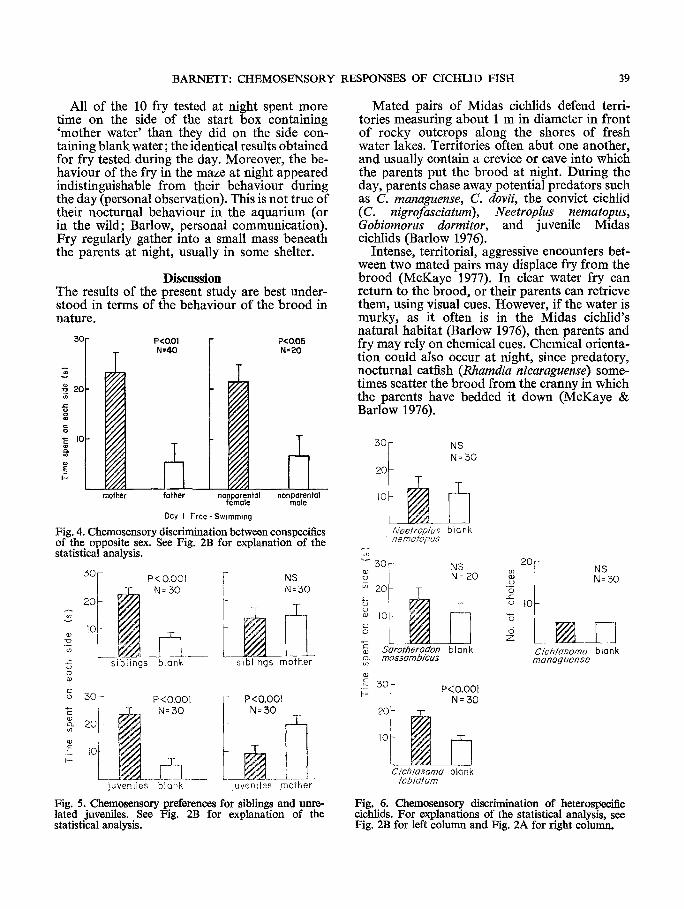
F r y prefer red ei ther thei r siblings o r (un- related) juveni les to b l ank water (Fig. 5). However , while they fai led to show a preference between their sibs and their mother , they pre- fe r red thei r mo the r to juveniles, who are m a j o r p reda to r s on the fry in the wild (Barlow, per- sonal communica t ion) .
W h e n offered a choice between b l ank water and ei ther o f two heterospecif ic p reda to r s (C. managuense and Neetroplus nematopus), fry showed no preference (Fig. 6). A th i rd hetero- specific, the he rb ivorous Afr ican cichlid, Sarotherodon mossambicus, also fai led to elicit a preference, while a four th species, the sympat r ic red devil cichlid (C. labiatum) was prefer red to b l ank water.
mother blank aon erental blank poreniol hP]nk ~m~!e ema e
50 - ~(not I~other)
~ P<O.OOI P < 0.001 P< O.Ol �9 N=40 2O c ~ N = 40 N = 40
B ~e
�9 father blonk nonparental blank parental blen~' ,'hale me[e
(not father)
Fig. 2. Chemosensory preferences of the fry for con- specifics.
A. After the fry was in the start box for 30 s, the start gate was raised. As soon as the fry entered an arm of the maze, defined as swimming 2 cm past the choice point, it was removed with a small net and discarded. The fry was scored either as showing a preference for the stimulus providing water to the arm of the maze it entered or, if it failed to enter an arm in 30 s, as showing 'no choice.' The number of choices made for each side was compared using a binomial test (Siegel 1956), with the 'no choice' scores omitted from the analysis. This method was not used for the remainder of the tests because it failed to evaluate fry who scored 'no choice,' despite the fact that these fry often spent significantly more time on one side of the main alley of the maze than on the other. Thus this method was viewed as less sensitive than the method described in the text.
B. Bars depict the mean time ( § sn~) spent on each side of the start box. The data could not be statistically evaluated in this form using a t-test since they did not have a normal distribution. Therefore each fry was as- signed a preference for the stimulus providing water to the side of the start box on which the fry spent the most time. The number of fry preferring each stimulus was compared using a sign test, with fry spending equal time on both sides omitted from the analysis.
NS N=40
~ 2o
g
.~_ I -
mother nonporenloJ femaIe
A
fo[her
P< 0.05 N=40
nonporenIo l mate
B
Fig. 3. Chemosensory discrkmination between parental and non-parental conspecifics. See Fig. 2B for explanation of the statistical analysis.
BARNETT: CHEMOSENSORY RESPONSES OF CICHLID FISH 39
All of the 10 fry tested at night spent more time on the side of the start box containing 'mother water' than they did on the side con- taining blank water; the identical results obtained for fry tested during the day. Moreover, the be- haviour of the fry in the maze at night appeared indistinguishable from their behaviour during the day (personal observation). This is not true of their nocturnal behaviour in the aquarium (or in the wild; Barlow, personal communication). Fry regularly gather into a small mass beneath the parents at night, usually in some shelter.
Discussion The results of the present study are best under- stood in terms of the behaviour of the brood in nature.
30 P<O~I P<Q05 N=40 N=20
-8 20
g ",d
mother
L
father nonparental nonparental female male
Day I Free-Swlmrmng
Fig. 4. Chemosensory discrimination between conspecifics of the opposite sex. See Fig. 2]3 for explanation of the statistical analysis.
u3 . . . . . .
sibl ings blank s ib l ings mother o o)
~ N=3o N~ # 20[- ': L _ N . . . . . . . . . .
juveniles blank juveniles mother
Fig. 5. Chemosensory preferences for siblings and unre- lated juveniles. See Fig. 2B for explanation of the statistical analysis.
Mated pairs of Midas cichlids defend terri- tories measuring about 1 m in diameter in front of rocky outcrops along the shores of fresh water lakes. Territories often abut one another, and usually contain a crevice or cave into which the parents put the brood at night. During the day, parents chase away potential predators such as C. managuense, C. dovii, the convict cichlid ( C. nigrofaseiatum), Neetroplus nematopus, Gobiomorus dormitor, and juvenile Midas cichlids (Barlow 1976).
Intense, territorial, aggressive encounters bet- ween two mated pairs may displace fry from the brood (McKaye 1977). In clear water fry can return to the brood, or their parents can retrieve them, using visual cues. However, if the water is murky, as it often is in the Midas cichlid's natural habitat (Barlow 1976), then parents and fry may rely on chemical cues. Chemical orienta- tion could also occur at night, since predatory, nocturnal catfish (Rhamdia niearaguense) some- times scatter the brood from the cranny in which the parents have bedded it down (McKaye & Barlow 1976).
30 NS N=30
2O io T]
A/eetrop/us blank nemofopus
-~ N : 20 2O
g io / ] g .
SaroTherodon blank o_ mossarnb/cus t o
ig: P<O, O01 N=30 20 '~
C/ch/asoma blank /cb/dfum
N= 30 o t0 ~d
C/ch/axoma blank monaguense
Fig. 6. Chemosensory discrimination of heterospecifie cichlids. For explanations of the statistical analysis, see Fig. 2B for left column and Fig. 2A for right column.
40 A N I M A L B E H A V I O U R , 3 0 , 1
If a displaced fry is to survive, it needs to return to the brood. Surprisingly, it does not matter which brood, since parents will adopt foreign fry i~to their brood and defend them (McKaye & McKaye 1977) if the foreign fry are the same age or younger than their own (Noakes & Barlow 1973). Since breeding in nature is synchronous (McKaye 1977), fry from different broods are likely to be of approximately the same age. McKaye & MeKaye (1977) have argued that interbrood adoptions are advanta- geous for both the fry and the foster parents.
The general impression of field workers ex- perienced in observing New World cichlids is that Midas cichlid parents have different roles in defending the brood (Barlow, McKaye, Bayliss, personal communications). During the egg and wriggler stages, the female spends more time tending the brood, while the male remains at the periphery of the territory or beyond, often threatening encroachers. When the fry hatch, the male moves nearer the brood, but still is not with them as much as the female is, and is always more easily chased away (Barlow 1976). Although quantitative data are lacking, such role division appears to be a general pattern for other Cichlasoma species (Meral 1973; Davies & McKaye 1978; Loiselle 1979; Mortensen 1979; Wilson 1979) and for New World substrate spawning cichlids generally (Barlow 1974; Cooper & Cooper 1979; Keenleyside 1979). This asymmetry in parental defence of the brood suggests that a displaced fry would be somewhat more likely to find safety by swimming to a mother rather than to a father. However, if only one conspecific stimulus is detectable, whether it be from any mother, any father, or other fry, then the fry should approach it rather than re- main alone. The data reported here (Figs 2, 4 and 5) suggest that fry can make these responses using chemical cues alone. Unlike the subjects of many studies of sibling recognition (Porter & Wyrick 1979; Waldman & Adler 1979) the fry of the Midas cichlid do not appear to gain an advantage by recognizing kin, since protection is derived from returning to any brood.
That the chemical preference for a male over a female (Fig. 4) did not persist after day 1 free- swimming may be an artifact of confinement in an aquarium where, in the absence of predators, the school of fry eventually disperses throughout the tank, thereby preventing significant asym- metry in parental proximity or defence (Barnett 1980).
Fry fail to show a preference for hetero- specifics whether they are predators of C. citrinellum ( C. managuense and C. nigrofasciatum) or not (S. mossambicus) (Fig. 6). The exception to this statement is that fry prefer the red devil cichlid (C. Iabiatum) to blank water. This can perhaps be explained by the close systematic relationship of C. labiatum and C. citrinellum. Although they are distinct species (Barlow and Munsey 1976) their close morphological simi- larity suggests they have only recently diverged. Perhaps these sympatric species have not differ- entiated enough to be distinct from each other in whatever chemical cues the fry use to detect con- specifics. In contrast, C. managuense, N. neetroplus, and S. mossambicus not only look radically different but are apparently chemically different as well.
The general lack of response to heterospecifics cannot be interpreted as a mechanism designed for avoiding predators, although it may inci- dentally have that effect. One would imagine that an effective predator avoidance mechanism based on chemical cues might cause either a freezing response in the fry or an actual avoid- ance of the side of the maze that contained heterospecific water. Neither of these occurred; rather, the fry simply chose both arms of the maze a statistically equal number of times. Either the fry do not chemically perceive hetero- specifics, or they perceive them but fail to show a response, perhaps because they have had no prior experience with heterospecific chemical cues.
A number of results do not fit the adaptive scheme sketched here. Fry prefer a non-parental male to their father and show no preference between their mother and a non-parental female (Fig. 3). A priori, one would expect the fry to choose a parental fish, which would pro- tect it, over a non-parental fish, which may eat it. Certainly one would not expect it actually to prefer a non-parental fish over a parental one (Fig. 3B). This result is not likely the result of variation in size of the stimulus fish, since there was no correlation between size of stimulus fish and degree of response. Moreover all stimulus fish were handled in the same manner.
A second set of results also do not appear to be adaptive. Fry prefer over blank water both conspecific non-parental fish (Fig. 2) and con- specific juveniles (Fig. 5). Both these classes of fish will eat fry; the juveniles will do so vo- raciously. The conclusion to be drawn is that fry of the Midas cichlid chemically prefer any con-
BARNETT: CHEMOSENSORY RESPONSES OF CICHLID FISH 41
specific to b l ank water and general ly show no preference for heterospecifics over b l ank water , regardless o f whether the conspecific or he tero- specific is a p r e d a t o r on the fry or not . A pre- vious conclus ion (Barnet t 1977b) tha t fry canno t chemical ly d iscr iminate between thei r fa ther and b l ank water , is therefore reversed by this more sensitive present study.
Ur ine is bel ieved to be the source o f the chemi- cal cues involved (Barnet t , 1981) b u t the ac tua l compounds a t t ended to, and their diffusion rates, are unknown.
A comple te exp lana t ion o f the adap t ive significance o f the f ry ' s chemosensory responses to o ther fish mus t t ake into account the re la t ion- ship between chemorecep t ion and o ther sensory modes , par t i cu la r ly vision.
Acknowledgments I t h a n k Len Welsh for helping wi th the experi- ments and George Bar low, Steve Gl i ckman , and R o y Caldwel l for their comments on the manu- script. Paul Siri, H a n k Fuj ishige and D o n He lmgren gave invaluable technical suppor t . This research was pa r t o f m y Ph.D. d isser ta t ion at the Univers i ty o f Cal i fornia , Berkeley, and was suppor t ed in large measure by N S F G r a n t BNS 76-19918 to G. W. Barlow.
R E F E R E N C E S Balon, E. K. 1975. Terminology of intervals in fish
development, d. Fish. Res. Bd. Can., 32, 1663-1670. Barlow, G. W. 1973. Competition between color morphs
of the polychromatic Midas cichlid Uichlasoma citrinellum. Science, N.Y., 179, 806-807.
Barlow, G. W. 1974. Contrasts in social behavior between Central American cichlid fishes and coral-reef surgeon fishes. Am. ZooL, 14, 19-34.
Barlow, G. W. 1976. The Midas ciehlid in Nicaragua. In: Investigations of the Ichthyofauna of Nicaraguan Lakes (Ed. by T. B. Thorson), pp. 333-358. Lincoln: University of Nebraska.
Barlow, G. W. & Munsey, J. 1976. The Red Devil- Midas-Arrow Cichlid species complex in Nicaragua. In: Investigations of the Ichthyofauna of Nicaraguan Lakes (Ed. by T. B. Thorson), pp. 359-370. Lincoln: University of Nebraska.
Barnett, C. 1977a. Aspects of chemical communication with special reference to fish. BioscL Commun., 3, 331-392.
Barnett, C. 1977b. Chemical recognition of the mother by the young of the ciehlid fish Cichlasoma citrinellum, d. chem. EcoL, 3, 463--468.
Barnett, C. 1980. The development of chemical communi- cation in cichlid fish. Ph.D. thesis, University of California, Berkeley.
Barnett, C. 1981. ~The role of urine in parent offspring communication in a cichlid fish. Z. Tierpsychol., 55, 173-182.
Baylis, J. 1974. The behavior and ecology of Herotilapia multispinosa (Teleostei, Cichlidae). Z. TierspychoL, 34, 115-146.
Beer, C. G. 1979. Vocal communication between laughing gull parents and chicks. Behaviour, 70, 118-146.
Brichard, P. 1975. R6flexions sur le choix de la nidificafion ou de l'incubation buccale comme mode de repro- duction chez certaines populations de poissons Cichlidae du lac Tanganyika. Revue ZooL Aft., 89, 871-888.
Caprona, M. D. C. 1974. The effect of chemical stimuli from conspecifics on the behavior of Haplochromis burtoni (Cichlidae, Pisces). Experientia, 30, 1394-- 1395.
Cooper, J. & Cooper, J. 1979. Aequidens dorsigerus (Heckel 1840). American Cichlid Assoc. Cichlid Index, 4(11), 1-2.
Davies, J. K. & McKaye, K. R. 1978. Cichlasoma dovii (Gtinther 1864). American Cichlid Assoc. Cichlid Index, 3(9), 1-2.
Fernald, R. & Hirata, N. R. 1977. Field study of Haplochromis burton# habitats and co-habitants. Env. BioL Fish., 2, 299-308.
Fryer, G. & Iles, T. D. 1972. The Cichlid Fishes of the Great Lakes o f Africa: Their Biology and Evolution. Edinburgh: Oliver and Boy&
Hay, T. 1978. Filial imprinting in the convict cichlid fish Cichlasoma nigrofaseiatum. Behaviour, 65, 138-160.
Keenleyside, M. H. A. 1979. Diversity and Adaptation in Fish Behavior. Berlin: Springer-Verlag.
Kiihrne, W. 1963. Brutpflege-und Sehwarmreaktionen bei Hemichromis bimaculatus (Pisces). Z. Tierpsychol., 20, 687-7O4.
Leon, M. 1979. Mother-young reunions. Prog. Psychobiol. Physiol. PsychoL, 8, 301-334.
Loiselle, P. V. 1979. Cichlasoma synspilum Hubbs 1935. American Cichlid Assoc. Cichlid Index, 4(3), 1-2.
McKaye, K. R. 1977. Competition for breeding sites between the cichlid fishes of Lake Jiloa, Nicaragua. Ecology, 58, 291-302.
McKaye, K. R. & Barlow, G. W. 1976. Chemical recog- nition of young by the Midas cichlid, Cichlasoma eitrinellum. Copeia, 1976, 276-282.
McKaye, K. R. & Hallacher, L. E. 1973. The midas cichlid in Nicaragua. Pae. Discovery, 25, I-8.
McKaye, K. R. & McKaye, N. M. 1977. Communal care and kidnapping of young by parental cichlids. Evolution, 31, 674-681.
Meral, G. H. 1973. The adaptive significance of terri- toriality in New World cichlidae. Ph.D. thesis, University of California, Berkeley.
Mortensen, L H. 1979. Cichlasoma salvini (Giinther 1964). American Cichlid Assoc. Cichlid Index, 4(9), 1-2.
Myrberg, A. A. 1975. The role of chemical and visual stimuli in the preferential discrimination of young by the eichlid fish Cichlasoma nigrofasciatum (Giinther). Z. TierpsychoL, 37, 274-297.
Noakes, D. L. G. & Barlow, G. W. 1973. Cross-fostering and parent offspring responses in Cichlasoma citrinellum (Pisces, Cichlidae). Z. Tierpsychol., 33, 147-152.
Noble, G. K. & Curtis, B. 1939. The social behavior of the jewel fish, Hemichrom bimaculatus Gill. Bull. Am. Mus. nat. Hist., 76, 1-46.
42 A N I M A L B E H A V I O U R , 3 0 , 1
Porter, R. H. & Wyrick, M. 1979. Sibling recognition in spiny mice (Acomys cahirinus): influence of age and isolation. Anita. Behav., 27, 761-766.
Russock, H. I. & Schein, M. W. 1974. Effect of early experience and age on initial responsiveness of Tilapia mossambica fry to a maternal model. Anita. Learn. Behav., 2, 111-114.
Siegel, S. 1956. Nonparametric Statistics for the Behavioral Sciences. New York: McGraw-Hill.
yon Kuenzer, P. 1968. Die AuslSsung der Nachfolgereak- tion bel erfahrungslosen Jungfischen von Nanna-
cara anomala (Cichlidae). Z. Tierpsychol., 25, 257-314.
Waldman, B. & Adler, K. 1979. Toad tadpoles associate preferentially with siblings. Nature, Lond., 282, 611-613.
Wilson, M. J. 1979. Cichlasoma spilurum (Giinther 1862). American Cichlid Assoc. Cichlid Index, 4(1), 1-2.
(Received 10 November 1980; revised 19 March 1981 ; MS. number: A2564)