copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Blackwell Publishing Ltd
Review
The contribution of invertebrate taxa to moorland bird diets and the potential implications of land-use
management
GRAEME M BUCHANAN
1
MURRAY C GRANT
1
ROY A SANDERSON
2
amp JAMES W PEARCE-HIGGINS
1
1
Royal Society for the Protection of Birds Dunedin House 25 Ravelston Terrace Edinburgh EH4 3TP UK
2
Institute for Research on Environment and Sustainability Devonshire Building University of Newcastle
Newcastle upon Tyne NE1 7RU UK
We reviewed the literature to determine the importance of invertebrates for moorland-breeding birds and considered our findings with respect to the conservation of such speciesin the UK The diets of many moorland birds consist predominantly of invertebrateswith a wide range of taxa recorded in the diets of moorland birds during the breedingseason Relatively few taxa (Arachnida Coleoptera Diptera Hemiptera HymenopteraLepidoptera and Oligochaeta) were widely taken with Diptera and Coleoptera being themost important Among these latter two insect orders Carabidae Curculionidae Elateridaeand Tipulidae were the most important families Comparisons of the diets of bird familiestreated separately according to whether data were derived from adults or chicks showedthat waders take more interstitial invertebrates than passerines which themselves takemore foliage invertebrates Although we lack detailed experimental data with which todemonstrate the direct effect of particular moorland management prescriptions on some ofthese key invertebrate taxa for breeding birds available data suggest that managementregimes that create a mosaic of habitats are likely to be most beneficial In particularheterogeneity in vegetation structure and species composition and the presence of wetflushes associated with the synchronized spring emergence of adults of certain insect speciesare likely to increase invertebrate food resources for birds
Moorland habitats in the UK are of internationalconservation importance but despite this have beensubject to major land-use and management changesin recent decades Most notable among these arelarge-scale afforestation declines in grouse moormanagement and increasing numbers of sheep andin the Scottish Highlands Red Deer
Cervus elaphus
(Mackey
et al
1998 Fuller amp Gough 1999 Robertson
et al
2001) Climate change and changes in govern-ment policies that affect upland farming practices mayin time lead to further major changes in moorlandhabitats (Bardgett
et al
1995 Berry
et al
2001 TaskForce for the Hills 2001) The UK uplands hold a
unique breeding bird assemblage including eightspecies listed in Annex 1 of the EC Birds Directiveand 28 others of conservation concern in the UK(Thompson
et al
1995 Gregory
et al
2002) a highproportion of which occur on moorlands Althoughthe broad-scale habitat associations of many of thesebirds are known (Haworth amp Thompson 1990 Brownamp Stillman 1993 Pearce-Higgins amp Grant in press)a detailed understanding of their ecological require-ments is often lacking making it difficult to assess andpredict the impact of habitat and other environmentalchanges upon moorland birds Given that the UKpopulations of several moorland bird species appearto be declining (Gibbons
et al
1993 Sim
et al
2005)it is important to address this knowledge gap anddetermine the role of such changes in causing declines
Corresponding authorEmail graemebuchananrspborguk
2
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Fundamental to understanding the ecologicalrequirements of a species is knowledge of its diet andof the factors that affect food availability (Newton1998) A high proportion of bird species that typic-ally breed in moorland habitats in the UK dependupon invertebrates as a major food source forsome or all of their breeding cycle (Ratcliffe 1990)Although dietary studies have been conducted on arange of these bird species during the breedingseason this information is disparate and of varyingquality Furthermore no overall assessment has yetbeen made of the relative contribution of differentinvertebrate species or taxa to moorland birddiets Determining the main invertebrate prey ofmoorland birds is an essential prerequisite to assessingthe extent to which the effects of habitat (and otherenvironmental) changes may affect bird populationsthrough effects on food supply and the extent towhich such changes can be linked to populationdeclines
Here we review data on the diets of a range of birdspecies that are widespread breeders on UK moor-lands and for which invertebrates form a substantialcomponent of the diet for at least part of their lifehistory Using data from a range of habitats and local-ities we describe the prevalence of invertebrates intheir diets and consider differences between avianfamilies On the basis of this information we con-sider how different land-uses and managements mayaffect some of the important invertebrate prey ofmoorland birds
METHODS
Study species and data collation
Data were collated for 14 bird species that are con-sidered lsquotypicalrsquo UK moorland species (Red Grouse
Lagopus lagopus scoticus
Black Grouse
Tetrao tetrix
European Golden Plover
Pluvialis apricaria
NorthernLapwing
Vanellus vanellus
Dunlin
Calidris alpina
Common Snipe
Gallinago gallinago
Eurasian Curlew
Numenius arquata
Common Redshank
Tringa totanus
Sky Lark
Alauda arvensis
Meadow Pipit
Anthuspratensis
Northern Wheatear
Oenanthe oenanthe
Whinchat
Saxicola rubetra
Stonechat
Saxicolatorquata
Ring Ouzel
Turdus torquatus
) and in whicheither the adults or chicks (or both) rely heavilyupon invertebrate prey (Table 1 Cramp amp Simmons1980 1983 Cramp 1988 Thompson
et al
1995)These range from species for which virtually all theUK breeding population occurs on moorland (eg
Red Grouse and Ring Ouzel) to others for whichmoorland represents one of several major breedinghabitats in the UK (eg Northern Lapwing and SkyLark) We do not consider species that breed primarilyon montane as opposed to moorland habitats andalso exclude a small number of moorland specieswith relatively localized distributions in the UK (egCommon Greenshank
Tringa nebularia
and Whimbrel
Numenius phaeopus
which are restricted largely tothe far northwest of Scotland and the Northern Islesrespectively) We consider only those age classes inwhich invertebrates comprise a substantial part ofthe diet so that for both Red Grouse and Black Grouseit is the earlier chick phases only that are considered(because older chicks and adults are largely herbivorousndash eg Cramp amp Simmons 1980 Kastdalen amp Wegge1985)
Breeding season data were obtained from bothpublished and unpublished (eg university thesesand unpublished reports) sources using a combina-tion of literature searches (on the Web of Sciencesearch facility) contacting ecologists involved inornithological research in the British uplandsobtaining translated material from studies pertainingto the former USSR (accessed via Zoolit ndash httpmytele2eebirds) and the authorsrsquo existing know-ledge of relevant studies Prior to analysis the datawere distinguished by geographical locations (treat-ing data separately by study area) where this infor-mation was provided and whether they concernedadult birds or chicks hereafter termed bird speciesage category (BSAC) Thus the 14 species consideredgive a potential 26 BSACs (adult Red and BlackGrouse being excluded as largely herbivorous) althoughdata were unavailable for some (see below) Datafrom different time periods within a study area (egyears or months within a year) were pooled tominimize pseudoreplication and because such datawere rarely available
Study areas were categorized according to whetheror not they were within Britain and Ireland (sub-sequently termed lsquoBampIrsquo or lsquonon-BampIrsquo) with BampI studyareas being further divided according to whetherthey comprised moorland or non-moorland habitatsUp to ten of the 30 study areas in the moorlandcategory included habitats other than unenclosedmoorland (mainly enclosed upland grasslands andwoodlands) but were classed as such because manyof the dietary data appeared to derive from birdsforaging on typical moorland vegetation types Onlytwo of the BampI study areas (both non-moorland)were from Ireland
Moorland bird diets
3
copy 2006 T
he Authors
Journal compilation copy
2006 British O
rnithologistsrsquo Union
Table 1
The number of dietary studies for each of the 14 study species according to broad location The number of study areas providing compositional or frequency ofoccurrence data on diet and hence contributing to the analyses in Figures 4 and 5 is also shown
Studies from outside Britain amp Ireland
Britain amp Ireland non-moorland
studies British moorlands
No of studies on adults and chicks
(respectively) used in analysis of invertebrates
occurring as
SourcesAdults Chicks Not Stated Adults Chicks Adults Chicks Taken lsquoImportantrsquo
(1984) Kondratrsquoyev and Kondratrsquoyeva (1988) Nechaev (1991) OrsquoConnell
et al
(1996) 6 ndash Zinovrsquoyev (1980) Danilov (1984) Green
et al
(1990) Nechaev (1991)7 ndash Kumari (1958) Cramp and Simmons (1983) Robson (1998) Stephen (2001) Grant unpubl data 8 ndash Peresadrsquoko and Koshelev (1980) Green (1986) Beintema
et al
(1991)Nechaev (1991) 9 ndash Nekrasov (1978) Malrsquochevskiy and Pukinskiy (1983) Jenny (1990) Nechaev (1991) Donald
et al
(2001) 10 ndash Hagvar and Ostbye (1976) Walton (1979)Prokofrsquoyeva (1980) Coulson and Whittaker (1978) Hagvar and Ostbye (1976) Danilov
et al
(1984) Burges (1994) Burges and Weildinger (2000) Evans unpubl data 11 ndashAverin and Ganya (1970 as cited in Cramp 1988) Brooke (1981) Kostin (1983) Gubin and Kovshar (1985) Dorzhiev and Khertuev (1992) 12 ndash Artemrsquoyev and Popov (1978)Prokofrsquoyeva (1980) Garnett
et al
(1988) von Labhardt (1988) 13 ndash Greig-Smith and Quicke (1983) Kostin (1983) Siepel (1990) Nechaev (1991) Cummins and OrsquoHalloran(2002) 14 ndash Tyler and Green (1994) Burfield (2002)
4
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Treatment of dietary data
Comparison across studies was difficult becausedietary data were reported to varying taxonomiclevels with some studies identifying the main inver-tebrate prey to the level of family genus or occasion-ally species whereas others only identified prey tothe level of class or order Consequently analyses wereperformed at two taxonomic levels to maximize thevalue of the available data First comparisons wereundertaken at a broader taxonomic level ie orderfor insects class for arachnids and annelids (althoughall mentions of worms in diet studies referred toearthworms (Lumbricidae) we use the broaderOligocheate for consistency) and higher level
s
forother invertebrates Secondly for the subset ofstudies that identified the main prey items to at leastthe level of family for insects and order for arachnidscomparisons were made at these finer taxonomiclevels although taxa distinguished in fewer thanfive studies were not used (eg Opilione or Araneaefamilies) Percentage composition data were summedacross the relevant finer taxonomic levels to producetotals for the broader level analysis (eg figures forColeopteran families would be combined to producea total for all Coleoptera) Where frequency of occur-rence data (ie the number or proportion of samplesin which the taxa were recorded) were presented onesample may have contained several families from aparticular order In this case the maximum percent-age occurrence figure for a finer level taxon (eg amongColeoptera families) was used to represent the per-centage occurrence of the broader taxonomic level(eg all Coleoptera) Insect taxa were distinguishedby life-cycle stage (adult or larval) where sufficientinformation was provided but were otherwise cate-gorized as lsquounagedrsquo
The methods used to collect dietary data and thetreatment of these data varied between studiesDifferent sampling methods used in these studiesare each subject to biases (Moreby amp Stoate 2000)preventing direct comparisons between studiesTherefore results from the dietary studies weresummarized and for each BSAC in each study areadata on invertebrate taxa were classed according to(1) their presence or absence and (2) whether or notthey either comprised at least 5 of the diet oroccurred in at least 5 of samples (for those studiesquantifying composition or frequency of occurrence)Where results were presented in different forms com-positional data were used in preference to frequencyof occurrence and biomass composition used in
preference to numerical composition The 5 level wasselected to distinguish the lsquoimportantrsquo taxa becausethis represented the most marked discontinuity inthe decline in the contribution of different inver-tebrate taxa to the bird diets (Fig 1) and concurs witha previous review of bird diets (Wilson
et al
1999)To compare the diets of different BSACs across all
studies data were summarized according to whethereach invertebrate taxon was recorded in any study ofthat species and whether its average percentagecomposition or percentage occurrence (across studyareas providing data for that species) was at least5 This was performed at both taxonomic levelsconsidered here
Determining differences between diets of bird families and ages
Statistical comparisons of the diets of different ageclasses of different moorland bird families wereundertaken (there being insufficient data for com-parisons between BSACs) Analyses were performedat the broader taxonomic level only owing to thesmall sample size available at the finer level lsquoUnagedrsquoinvertebrates were omitted as were data from studyareas providing no information on the samplingmethods and any taxa taken by or classed as lsquoimport-antrsquo in fewer than 10 of BSACs in the respectiveanalyses (Fig 2)
Analyses were conducted using (1) the presenceor absence of each invertebrate taxon and (2) whetherthe taxon was classed as lsquoimportantrsquo (based upon the5 criterion) as dependent variables Data were
Figure 1 The decline in the proportion of broader invertebratetaxa (see text) in relation to the increasing contribution made tobird diets (ie all taxa comprise between 0 and 5 of the dietbut none contributes more than 90) Data are derived from 14bird species (with data from adults and chicks treated separatelygiving 24 different categories) using studies that present dietarydata either as percentage composition or as percentageoccurrence in diet samples The proportions for each taxon arecalculated from the average of the mean values for the differentbird species and age categories
Moorland bird diets
5
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
categorized according to whether they were grousechicks wader adults wader chicks passerine adultsor passerine chicks Variation due to different sam-pling methods and to the localities and habitats fromwhich data were derived was taken into accountbefore assessing differences between bird groups Inaddition the treatment of the data (eg percentageoccurrence or numerical composition) was takeninto account in the analyses of the lsquoimportancersquo oftaxa Thus logistic
ANOVA
was undertaken usingProc GENMOD in SAS 802 (SAS Institute 2001)specifying a logit link and binomial error structure(see Endnote) first fitting sampling method location(ie BampI moorland BampI non-moorland non-BampI)and for analyses of lsquoimportancersquo data treatment
RESULTS
Coverage of bird species age categories variation with location and sampling methods
Data on the 14 moorland bird species considered inthis review were collated from 62 different sourcesWhen treated separately by study area and bird
species age category there were 115 diet studies
minus
31from British moorland habitats 12 from otherhabitats in BampI and 72 from outside BampI (Table 1)The extent to which data were available for adultbirds and chicks varied between these three categor-ies with 70 of the dietary studies from outsideBampI concerning adult birds compared with 50 and13 of those from non-moorland and moorlandhabitats respectively in BampI (Table 1 = 984
P =
0007) There were also differences in themethods used to collect dietary data with 74 ofBampI studies using faecal analysis compared with 9elsewhere where 81 used either crop or stomachsamples ( = 425
P lt
0001) A higher proportionof the dietary studies from outside BampI reported dataas presence or absence only (ie 33 compared with15 and 0 for moorland and non-moorland habitatsin BampI respectively) although this difference was notstatistically significant ( = 51
P =
008)
Trends in the prevalence and importance of invertebrate taxa
At the broader taxonomic level (order and above)adult and unaged Coleoptera (beetles) Diptera (true
Figure 2 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different broader invertebratetaxa were recorded as occurring in the diet (entire bar) and being of importance in the diet (open and thin hatched grey sections of barsonly) Data are presented for studies from all locations (solidopen bars) and from British moorland studies only (hatched bars)Importance in the diet is defined as comprising ge 5 of diet composition or as occurring in ge 5 of dietary samples (where the studypresents frequency of occurrence but not compositional data)
χ22
χ12
χ22
6
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
flies) Hemiptera (bugs) Hymenoptera Lepidopteralarvae Arachnida (arachnids) and Oligochaeta (worms)were the most ubiquitous prey all being taken by morethan 25 of the different BSACs (Fig 2) There was asimilar pattern in the occurrence of taxa when compar-isons were limited to lsquoimportantrsquo taxa only (ie thosecomprising
ge
5 of the diet or occurring in
ge
5 of thesamples on average for a BSAC) Arachnids adult andunaged beetles true flies bugs Hymenoptera andLepidoptera larvae were most prevalent as lsquoimportantrsquotaxa in the diets (Fig 2) Differences between thefrequency of occurrence of taxa and the frequency withwhich they occurred as lsquoimportantrsquo taxa were mostmarked for arachnids and Lepidoptera larvae Bothtaxa were taken by a wide range of BSACs but oftenformed a relatively small part of the diet Thesecomparisons indicated few major differences betweenthe data from all study areas combined and those fromBritish moorland only However among the mostfrequently occurring prey there was a markedly greaterrepresentation of both Hymenoptera (adults andlarvae) and adult true flies in the moorland diets
Fewer data were available for the comparisons atthe finer taxonomic level with 66 dietary studies
from 14 bird species (nine adult 11 chick) providingsufficient data at this level (Fig 3)
These comparisons suggested that Araneae (spiders)are more prevalent than Opiliones (harvestmen)and that although a wide range of beetle and true flyfamilies are taken only a few occur frequently Carabidae(carabids) Chrysomelidae (chrysomelids) Curcu-lionidae (curculionids) Elateridae (elaterids) andStaphylinidae (staphylinids) were the most frequentlytaken adult beetles being recorded in more than 35of the BSACs (Fig 3) Beetle larvae were recordedrelatively rarely True fly families tended to be lessprevalent in bird diets with only Chironomidae(chironomid) Bibionidae (bibionid) and Tipulidae(tipulid) adults recorded in more than 20 of thediets and each recorded in less than 30 Howeverlarval tipulids and Muscidae (muscids) were recordedin 52 and 24 of the diets respectively Of theHymenoptera Formicidae (formicids) (unaged) werethe only group that were relatively prevalent in the dietsWhen classifying diet in terms of the lsquoimportantrsquo taxathe decrease in occurrence was most marked for Araneae(spiders) chrysomelids staphylinids bibionids muscidsand formicids Comparisons of the prevalence of
Figure 3 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different finer level invertebratetaxa were recorded as occurring in the diet (filled plus open sections of bars) and being of importance in the diet (filled section only)Data are derived from studies from all locations Importance in the diet is defined as for Figure 2 For each taxon (eg Carabidae) columnsfrom left to right represent proportion of adults larvae and unaged except for Arachnids (Araneae and Opiliones) for which all are classedas adults
Moorland bird diets
7
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
invertebrates from studies on British moorlands onlywere not made at this taxonomic level because fewstudies provided such data (16 compared with 30 inFig 2) and few BSACs were represented (eightcompared with 15 in Fig 2)
Differences between bird families and age categories
Significant differences in the prevalence of inverte-brates were detected between the different samplingmethods for arachnids worms and Hymenopteralarvae and between locations for adult beetlesHymenoptera adults and Lepidoptera larvae ( gt117
P lt
001 in all cases) Among invertebrate taxathat were classed as lsquoimportantrsquo significant differencesoccurred between sampling methods for wormslocations for adult beetles and the method of datatreatment for larval true flies Gastropoda bugs andHymenoptera adults ( gt 78
P lt
005 in all cases)Owing to the high degree of association betweenthese different effects (particularly sampling methodand location ndash see above) these differences have tobe interpreted with caution
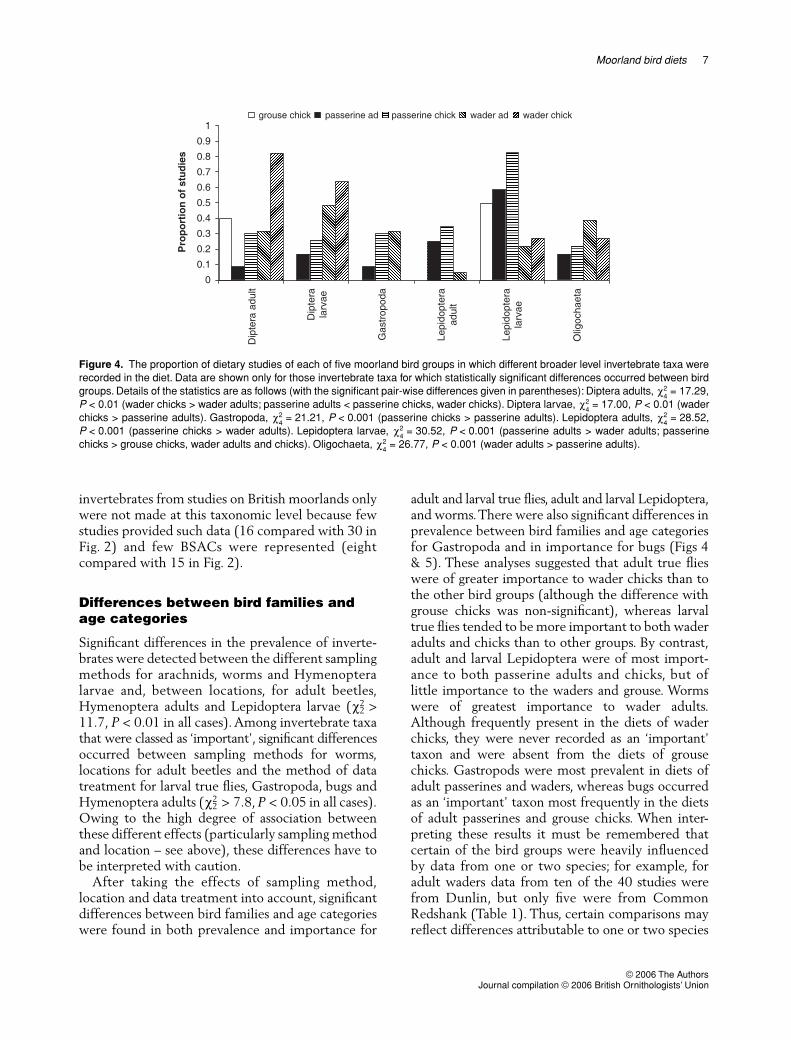
After taking the effects of sampling methodlocation and data treatment into account significantdifferences between bird families and age categorieswere found in both prevalence and importance for
adult and larval true flies adult and larval Lepidopteraand worms There were also significant differences inprevalence between bird families and age categoriesfor Gastropoda and in importance for bugs (Figs 4amp 5) These analyses suggested that adult true flieswere of greater importance to wader chicks than tothe other bird groups (although the difference withgrouse chicks was non-significant) whereas larvaltrue flies tended to be more important to both waderadults and chicks than to other groups By contrastadult and larval Lepidoptera were of most import-ance to both passerine adults and chicks but oflittle importance to the waders and grouse Wormswere of greatest importance to wader adultsAlthough frequently present in the diets of waderchicks they were never recorded as an lsquoimportantrsquotaxon and were absent from the diets of grousechicks Gastropods were most prevalent in diets ofadult passerines and waders whereas bugs occurredas an lsquoimportantrsquo taxon most frequently in the dietsof adult passerines and grouse chicks When inter-preting these results it must be remembered thatcertain of the bird groups were heavily influencedby data from one or two species for example foradult waders data from ten of the 40 studies werefrom Dunlin but only five were from CommonRedshank (Table 1) Thus certain comparisons mayreflect differences attributable to one or two species
χ22
χ22
Figure 4 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded in the diet Data are shown only for those invertebrate taxa for which statistically significant differences occurred between birdgroups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1729P lt 001 (wader chicks gt wader adults passerine adults lt passerine chicks wader chicks) Diptera larvae = 1700 P lt 001 (waderchicks gt passerine adults) Gastropoda = 2121 P lt 0001 (passerine chicks gt passerine adults) Lepidoptera adults = 2852P lt 0001 (passerine chicks gt wader adults) Lepidoptera larvae = 3052 P lt 0001 (passerine adults gt wader adults passerinechicks gt grouse chicks wader adults and chicks) Oligochaeta = 2677 P lt 0001 (wader adults gt passerine adults)
χ42
χ42
χ42 χ
42
χ42
χ42
8
G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
as opposed to being more widely representative ofthe relevant bird groups
DISCUSSION
The contribution of invertebrates to moorland bird diets
A wide range of invertebrates are consumed bymoorland birds but only a small number of thebroader level taxonomic groups are particularlyprevalent (ie arachnids beetles true flies bugsHymenoptera Lepidoptera and worms) With theexception of Hymenoptera and earthworms Coulson(1988) found that these taxa were among the mostabundant on moorland in northern England suggest-ing that birds are generally exploiting the most avail-able prey on moorlands The absence of Collembolaand enchytraeid worms from diets despite also beingnumerous is almost certainly due to their small sizeand hence lack of profitability as prey This diverserange of prey will partly reflect variation in diet betweenBSACs resulting from their different foragingstrategies (discussed below) However spatial andtemporal differences in prey availability within andbetween studies will also influence diet compositionas indicated by within-species dietary differencesassociated with different study locations and the fewstudies that have examined diet in relation to seasonaland between-habitat variation in invertebrateabundance (Picozzi amp Hepburn 1984 Starling-
Westerberg 2001 Whittingham et al 2001 Pearce-Higgins amp Yalden 2004)
Twelve studies presenting detailed compositionaldietary data for eight BSACs on British moorlands(Walton 1979 Picozzi amp Hepburn 1984 Garnettet al 1988 Cayford et al 1989 Whittingham 1996Starling-Westerberg 2001 Stephen 2001 Burfield2002 Pearce-Higgins amp Yalden 2003 2004) generallysupport the results of our wider analysis These studiesshowed that beetles formed the most importantitem for five of the BSACs considered comprisingover 15 of the diet of four cases True flies weremost important in four cases comprising over 15of the total diet in five The prevalence of adult trueflies in moorland bird diets especially among wadersand Red Grouse reflects in part the high abundanceof tipulids on some moorland areas following theirhighly synchronized emergences (Coulson 1962)Less important in this subset of more detailedmoorland studies were earthworms which were takenmainly by adult waders foraging on nearby improvedgrasslands but were also the most important item forRing Ouzel chicks As in our wider analysis arachnidsbugs Hymenoptera and Lepidoptera were alsoimportant and frequently occurring prey items inthese studies albeit less so than true flies and beetlesThus Hymenoptera were the most important preyin one case and accounted for more than 15 of thediet of three cases Arachnids bugs and Lepidopterawere never dominant possibly because of the relativelylow biomass and small prey size of some of these
Figure 5 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded as being important in the diet Importance in the diet is defined as for Figure 2 Data are shown only for those invertebrate taxafor which statistically significant differences occurred between bird groups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1389 P lt 001 (wader chicks gt wader adults passerine adults and chicks)Diptera larvae = 2130 P lt 0001 (wader adults gt passerine chicks) Hemiptera = 1178 P lt 005 (grouse chicks gt wader adultspasserine adults gt passerine chicks wader adults) Lepidoptera adults = 1714 P lt 001 (passerine chicks gt passerine adults)Lepidoptera larvae = 1254 P lt 005 (wader adults lt passerine adults and chicks) Oligochaeta = 3185 P lt 0001 (wader adultsgt passerine chicks)
χ42
χ42 χ
42
χ42
χ42 χ
42
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
2
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Fundamental to understanding the ecologicalrequirements of a species is knowledge of its diet andof the factors that affect food availability (Newton1998) A high proportion of bird species that typic-ally breed in moorland habitats in the UK dependupon invertebrates as a major food source forsome or all of their breeding cycle (Ratcliffe 1990)Although dietary studies have been conducted on arange of these bird species during the breedingseason this information is disparate and of varyingquality Furthermore no overall assessment has yetbeen made of the relative contribution of differentinvertebrate species or taxa to moorland birddiets Determining the main invertebrate prey ofmoorland birds is an essential prerequisite to assessingthe extent to which the effects of habitat (and otherenvironmental) changes may affect bird populationsthrough effects on food supply and the extent towhich such changes can be linked to populationdeclines
Here we review data on the diets of a range of birdspecies that are widespread breeders on UK moor-lands and for which invertebrates form a substantialcomponent of the diet for at least part of their lifehistory Using data from a range of habitats and local-ities we describe the prevalence of invertebrates intheir diets and consider differences between avianfamilies On the basis of this information we con-sider how different land-uses and managements mayaffect some of the important invertebrate prey ofmoorland birds
METHODS
Study species and data collation
Data were collated for 14 bird species that are con-sidered lsquotypicalrsquo UK moorland species (Red Grouse
Lagopus lagopus scoticus
Black Grouse
Tetrao tetrix
European Golden Plover
Pluvialis apricaria
NorthernLapwing
Vanellus vanellus
Dunlin
Calidris alpina
Common Snipe
Gallinago gallinago
Eurasian Curlew
Numenius arquata
Common Redshank
Tringa totanus
Sky Lark
Alauda arvensis
Meadow Pipit
Anthuspratensis
Northern Wheatear
Oenanthe oenanthe
Whinchat
Saxicola rubetra
Stonechat
Saxicolatorquata
Ring Ouzel
Turdus torquatus
) and in whicheither the adults or chicks (or both) rely heavilyupon invertebrate prey (Table 1 Cramp amp Simmons1980 1983 Cramp 1988 Thompson
et al
1995)These range from species for which virtually all theUK breeding population occurs on moorland (eg
Red Grouse and Ring Ouzel) to others for whichmoorland represents one of several major breedinghabitats in the UK (eg Northern Lapwing and SkyLark) We do not consider species that breed primarilyon montane as opposed to moorland habitats andalso exclude a small number of moorland specieswith relatively localized distributions in the UK (egCommon Greenshank
Tringa nebularia
and Whimbrel
Numenius phaeopus
which are restricted largely tothe far northwest of Scotland and the Northern Islesrespectively) We consider only those age classes inwhich invertebrates comprise a substantial part ofthe diet so that for both Red Grouse and Black Grouseit is the earlier chick phases only that are considered(because older chicks and adults are largely herbivorousndash eg Cramp amp Simmons 1980 Kastdalen amp Wegge1985)
Breeding season data were obtained from bothpublished and unpublished (eg university thesesand unpublished reports) sources using a combina-tion of literature searches (on the Web of Sciencesearch facility) contacting ecologists involved inornithological research in the British uplandsobtaining translated material from studies pertainingto the former USSR (accessed via Zoolit ndash httpmytele2eebirds) and the authorsrsquo existing know-ledge of relevant studies Prior to analysis the datawere distinguished by geographical locations (treat-ing data separately by study area) where this infor-mation was provided and whether they concernedadult birds or chicks hereafter termed bird speciesage category (BSAC) Thus the 14 species consideredgive a potential 26 BSACs (adult Red and BlackGrouse being excluded as largely herbivorous) althoughdata were unavailable for some (see below) Datafrom different time periods within a study area (egyears or months within a year) were pooled tominimize pseudoreplication and because such datawere rarely available
Study areas were categorized according to whetheror not they were within Britain and Ireland (sub-sequently termed lsquoBampIrsquo or lsquonon-BampIrsquo) with BampI studyareas being further divided according to whetherthey comprised moorland or non-moorland habitatsUp to ten of the 30 study areas in the moorlandcategory included habitats other than unenclosedmoorland (mainly enclosed upland grasslands andwoodlands) but were classed as such because manyof the dietary data appeared to derive from birdsforaging on typical moorland vegetation types Onlytwo of the BampI study areas (both non-moorland)were from Ireland
Moorland bird diets
3
copy 2006 T
he Authors
Journal compilation copy
2006 British O
rnithologistsrsquo Union
Table 1
The number of dietary studies for each of the 14 study species according to broad location The number of study areas providing compositional or frequency ofoccurrence data on diet and hence contributing to the analyses in Figures 4 and 5 is also shown
Studies from outside Britain amp Ireland
Britain amp Ireland non-moorland
studies British moorlands
No of studies on adults and chicks
(respectively) used in analysis of invertebrates
occurring as
SourcesAdults Chicks Not Stated Adults Chicks Adults Chicks Taken lsquoImportantrsquo
(1984) Kondratrsquoyev and Kondratrsquoyeva (1988) Nechaev (1991) OrsquoConnell
et al
(1996) 6 ndash Zinovrsquoyev (1980) Danilov (1984) Green
et al
(1990) Nechaev (1991)7 ndash Kumari (1958) Cramp and Simmons (1983) Robson (1998) Stephen (2001) Grant unpubl data 8 ndash Peresadrsquoko and Koshelev (1980) Green (1986) Beintema
et al
(1991)Nechaev (1991) 9 ndash Nekrasov (1978) Malrsquochevskiy and Pukinskiy (1983) Jenny (1990) Nechaev (1991) Donald
et al
(2001) 10 ndash Hagvar and Ostbye (1976) Walton (1979)Prokofrsquoyeva (1980) Coulson and Whittaker (1978) Hagvar and Ostbye (1976) Danilov
et al
(1984) Burges (1994) Burges and Weildinger (2000) Evans unpubl data 11 ndashAverin and Ganya (1970 as cited in Cramp 1988) Brooke (1981) Kostin (1983) Gubin and Kovshar (1985) Dorzhiev and Khertuev (1992) 12 ndash Artemrsquoyev and Popov (1978)Prokofrsquoyeva (1980) Garnett
et al
(1988) von Labhardt (1988) 13 ndash Greig-Smith and Quicke (1983) Kostin (1983) Siepel (1990) Nechaev (1991) Cummins and OrsquoHalloran(2002) 14 ndash Tyler and Green (1994) Burfield (2002)
4
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Treatment of dietary data
Comparison across studies was difficult becausedietary data were reported to varying taxonomiclevels with some studies identifying the main inver-tebrate prey to the level of family genus or occasion-ally species whereas others only identified prey tothe level of class or order Consequently analyses wereperformed at two taxonomic levels to maximize thevalue of the available data First comparisons wereundertaken at a broader taxonomic level ie orderfor insects class for arachnids and annelids (althoughall mentions of worms in diet studies referred toearthworms (Lumbricidae) we use the broaderOligocheate for consistency) and higher level
s
forother invertebrates Secondly for the subset ofstudies that identified the main prey items to at leastthe level of family for insects and order for arachnidscomparisons were made at these finer taxonomiclevels although taxa distinguished in fewer thanfive studies were not used (eg Opilione or Araneaefamilies) Percentage composition data were summedacross the relevant finer taxonomic levels to producetotals for the broader level analysis (eg figures forColeopteran families would be combined to producea total for all Coleoptera) Where frequency of occur-rence data (ie the number or proportion of samplesin which the taxa were recorded) were presented onesample may have contained several families from aparticular order In this case the maximum percent-age occurrence figure for a finer level taxon (eg amongColeoptera families) was used to represent the per-centage occurrence of the broader taxonomic level(eg all Coleoptera) Insect taxa were distinguishedby life-cycle stage (adult or larval) where sufficientinformation was provided but were otherwise cate-gorized as lsquounagedrsquo
The methods used to collect dietary data and thetreatment of these data varied between studiesDifferent sampling methods used in these studiesare each subject to biases (Moreby amp Stoate 2000)preventing direct comparisons between studiesTherefore results from the dietary studies weresummarized and for each BSAC in each study areadata on invertebrate taxa were classed according to(1) their presence or absence and (2) whether or notthey either comprised at least 5 of the diet oroccurred in at least 5 of samples (for those studiesquantifying composition or frequency of occurrence)Where results were presented in different forms com-positional data were used in preference to frequencyof occurrence and biomass composition used in
preference to numerical composition The 5 level wasselected to distinguish the lsquoimportantrsquo taxa becausethis represented the most marked discontinuity inthe decline in the contribution of different inver-tebrate taxa to the bird diets (Fig 1) and concurs witha previous review of bird diets (Wilson
et al
1999)To compare the diets of different BSACs across all
studies data were summarized according to whethereach invertebrate taxon was recorded in any study ofthat species and whether its average percentagecomposition or percentage occurrence (across studyareas providing data for that species) was at least5 This was performed at both taxonomic levelsconsidered here
Determining differences between diets of bird families and ages
Statistical comparisons of the diets of different ageclasses of different moorland bird families wereundertaken (there being insufficient data for com-parisons between BSACs) Analyses were performedat the broader taxonomic level only owing to thesmall sample size available at the finer level lsquoUnagedrsquoinvertebrates were omitted as were data from studyareas providing no information on the samplingmethods and any taxa taken by or classed as lsquoimport-antrsquo in fewer than 10 of BSACs in the respectiveanalyses (Fig 2)
Analyses were conducted using (1) the presenceor absence of each invertebrate taxon and (2) whetherthe taxon was classed as lsquoimportantrsquo (based upon the5 criterion) as dependent variables Data were
Figure 1 The decline in the proportion of broader invertebratetaxa (see text) in relation to the increasing contribution made tobird diets (ie all taxa comprise between 0 and 5 of the dietbut none contributes more than 90) Data are derived from 14bird species (with data from adults and chicks treated separatelygiving 24 different categories) using studies that present dietarydata either as percentage composition or as percentageoccurrence in diet samples The proportions for each taxon arecalculated from the average of the mean values for the differentbird species and age categories
Moorland bird diets
5
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
categorized according to whether they were grousechicks wader adults wader chicks passerine adultsor passerine chicks Variation due to different sam-pling methods and to the localities and habitats fromwhich data were derived was taken into accountbefore assessing differences between bird groups Inaddition the treatment of the data (eg percentageoccurrence or numerical composition) was takeninto account in the analyses of the lsquoimportancersquo oftaxa Thus logistic
ANOVA
was undertaken usingProc GENMOD in SAS 802 (SAS Institute 2001)specifying a logit link and binomial error structure(see Endnote) first fitting sampling method location(ie BampI moorland BampI non-moorland non-BampI)and for analyses of lsquoimportancersquo data treatment
RESULTS
Coverage of bird species age categories variation with location and sampling methods
Data on the 14 moorland bird species considered inthis review were collated from 62 different sourcesWhen treated separately by study area and bird
species age category there were 115 diet studies
minus
31from British moorland habitats 12 from otherhabitats in BampI and 72 from outside BampI (Table 1)The extent to which data were available for adultbirds and chicks varied between these three categor-ies with 70 of the dietary studies from outsideBampI concerning adult birds compared with 50 and13 of those from non-moorland and moorlandhabitats respectively in BampI (Table 1 = 984
P =
0007) There were also differences in themethods used to collect dietary data with 74 ofBampI studies using faecal analysis compared with 9elsewhere where 81 used either crop or stomachsamples ( = 425
P lt
0001) A higher proportionof the dietary studies from outside BampI reported dataas presence or absence only (ie 33 compared with15 and 0 for moorland and non-moorland habitatsin BampI respectively) although this difference was notstatistically significant ( = 51
P =
008)
Trends in the prevalence and importance of invertebrate taxa
At the broader taxonomic level (order and above)adult and unaged Coleoptera (beetles) Diptera (true
Figure 2 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different broader invertebratetaxa were recorded as occurring in the diet (entire bar) and being of importance in the diet (open and thin hatched grey sections of barsonly) Data are presented for studies from all locations (solidopen bars) and from British moorland studies only (hatched bars)Importance in the diet is defined as comprising ge 5 of diet composition or as occurring in ge 5 of dietary samples (where the studypresents frequency of occurrence but not compositional data)
χ22
χ12
χ22
6
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
flies) Hemiptera (bugs) Hymenoptera Lepidopteralarvae Arachnida (arachnids) and Oligochaeta (worms)were the most ubiquitous prey all being taken by morethan 25 of the different BSACs (Fig 2) There was asimilar pattern in the occurrence of taxa when compar-isons were limited to lsquoimportantrsquo taxa only (ie thosecomprising
ge
5 of the diet or occurring in
ge
5 of thesamples on average for a BSAC) Arachnids adult andunaged beetles true flies bugs Hymenoptera andLepidoptera larvae were most prevalent as lsquoimportantrsquotaxa in the diets (Fig 2) Differences between thefrequency of occurrence of taxa and the frequency withwhich they occurred as lsquoimportantrsquo taxa were mostmarked for arachnids and Lepidoptera larvae Bothtaxa were taken by a wide range of BSACs but oftenformed a relatively small part of the diet Thesecomparisons indicated few major differences betweenthe data from all study areas combined and those fromBritish moorland only However among the mostfrequently occurring prey there was a markedly greaterrepresentation of both Hymenoptera (adults andlarvae) and adult true flies in the moorland diets
Fewer data were available for the comparisons atthe finer taxonomic level with 66 dietary studies
from 14 bird species (nine adult 11 chick) providingsufficient data at this level (Fig 3)
These comparisons suggested that Araneae (spiders)are more prevalent than Opiliones (harvestmen)and that although a wide range of beetle and true flyfamilies are taken only a few occur frequently Carabidae(carabids) Chrysomelidae (chrysomelids) Curcu-lionidae (curculionids) Elateridae (elaterids) andStaphylinidae (staphylinids) were the most frequentlytaken adult beetles being recorded in more than 35of the BSACs (Fig 3) Beetle larvae were recordedrelatively rarely True fly families tended to be lessprevalent in bird diets with only Chironomidae(chironomid) Bibionidae (bibionid) and Tipulidae(tipulid) adults recorded in more than 20 of thediets and each recorded in less than 30 Howeverlarval tipulids and Muscidae (muscids) were recordedin 52 and 24 of the diets respectively Of theHymenoptera Formicidae (formicids) (unaged) werethe only group that were relatively prevalent in the dietsWhen classifying diet in terms of the lsquoimportantrsquo taxathe decrease in occurrence was most marked for Araneae(spiders) chrysomelids staphylinids bibionids muscidsand formicids Comparisons of the prevalence of
Figure 3 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different finer level invertebratetaxa were recorded as occurring in the diet (filled plus open sections of bars) and being of importance in the diet (filled section only)Data are derived from studies from all locations Importance in the diet is defined as for Figure 2 For each taxon (eg Carabidae) columnsfrom left to right represent proportion of adults larvae and unaged except for Arachnids (Araneae and Opiliones) for which all are classedas adults
Moorland bird diets
7
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
invertebrates from studies on British moorlands onlywere not made at this taxonomic level because fewstudies provided such data (16 compared with 30 inFig 2) and few BSACs were represented (eightcompared with 15 in Fig 2)
Differences between bird families and age categories
Significant differences in the prevalence of inverte-brates were detected between the different samplingmethods for arachnids worms and Hymenopteralarvae and between locations for adult beetlesHymenoptera adults and Lepidoptera larvae ( gt117
P lt
001 in all cases) Among invertebrate taxathat were classed as lsquoimportantrsquo significant differencesoccurred between sampling methods for wormslocations for adult beetles and the method of datatreatment for larval true flies Gastropoda bugs andHymenoptera adults ( gt 78
P lt
005 in all cases)Owing to the high degree of association betweenthese different effects (particularly sampling methodand location ndash see above) these differences have tobe interpreted with caution
After taking the effects of sampling methodlocation and data treatment into account significantdifferences between bird families and age categorieswere found in both prevalence and importance for
adult and larval true flies adult and larval Lepidopteraand worms There were also significant differences inprevalence between bird families and age categoriesfor Gastropoda and in importance for bugs (Figs 4amp 5) These analyses suggested that adult true flieswere of greater importance to wader chicks than tothe other bird groups (although the difference withgrouse chicks was non-significant) whereas larvaltrue flies tended to be more important to both waderadults and chicks than to other groups By contrastadult and larval Lepidoptera were of most import-ance to both passerine adults and chicks but oflittle importance to the waders and grouse Wormswere of greatest importance to wader adultsAlthough frequently present in the diets of waderchicks they were never recorded as an lsquoimportantrsquotaxon and were absent from the diets of grousechicks Gastropods were most prevalent in diets ofadult passerines and waders whereas bugs occurredas an lsquoimportantrsquo taxon most frequently in the dietsof adult passerines and grouse chicks When inter-preting these results it must be remembered thatcertain of the bird groups were heavily influencedby data from one or two species for example foradult waders data from ten of the 40 studies werefrom Dunlin but only five were from CommonRedshank (Table 1) Thus certain comparisons mayreflect differences attributable to one or two species
χ22
χ22
Figure 4 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded in the diet Data are shown only for those invertebrate taxa for which statistically significant differences occurred between birdgroups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1729P lt 001 (wader chicks gt wader adults passerine adults lt passerine chicks wader chicks) Diptera larvae = 1700 P lt 001 (waderchicks gt passerine adults) Gastropoda = 2121 P lt 0001 (passerine chicks gt passerine adults) Lepidoptera adults = 2852P lt 0001 (passerine chicks gt wader adults) Lepidoptera larvae = 3052 P lt 0001 (passerine adults gt wader adults passerinechicks gt grouse chicks wader adults and chicks) Oligochaeta = 2677 P lt 0001 (wader adults gt passerine adults)
χ42
χ42
χ42 χ
42
χ42
χ42
8
G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
as opposed to being more widely representative ofthe relevant bird groups
DISCUSSION
The contribution of invertebrates to moorland bird diets
A wide range of invertebrates are consumed bymoorland birds but only a small number of thebroader level taxonomic groups are particularlyprevalent (ie arachnids beetles true flies bugsHymenoptera Lepidoptera and worms) With theexception of Hymenoptera and earthworms Coulson(1988) found that these taxa were among the mostabundant on moorland in northern England suggest-ing that birds are generally exploiting the most avail-able prey on moorlands The absence of Collembolaand enchytraeid worms from diets despite also beingnumerous is almost certainly due to their small sizeand hence lack of profitability as prey This diverserange of prey will partly reflect variation in diet betweenBSACs resulting from their different foragingstrategies (discussed below) However spatial andtemporal differences in prey availability within andbetween studies will also influence diet compositionas indicated by within-species dietary differencesassociated with different study locations and the fewstudies that have examined diet in relation to seasonaland between-habitat variation in invertebrateabundance (Picozzi amp Hepburn 1984 Starling-
Westerberg 2001 Whittingham et al 2001 Pearce-Higgins amp Yalden 2004)
Twelve studies presenting detailed compositionaldietary data for eight BSACs on British moorlands(Walton 1979 Picozzi amp Hepburn 1984 Garnettet al 1988 Cayford et al 1989 Whittingham 1996Starling-Westerberg 2001 Stephen 2001 Burfield2002 Pearce-Higgins amp Yalden 2003 2004) generallysupport the results of our wider analysis These studiesshowed that beetles formed the most importantitem for five of the BSACs considered comprisingover 15 of the diet of four cases True flies weremost important in four cases comprising over 15of the total diet in five The prevalence of adult trueflies in moorland bird diets especially among wadersand Red Grouse reflects in part the high abundanceof tipulids on some moorland areas following theirhighly synchronized emergences (Coulson 1962)Less important in this subset of more detailedmoorland studies were earthworms which were takenmainly by adult waders foraging on nearby improvedgrasslands but were also the most important item forRing Ouzel chicks As in our wider analysis arachnidsbugs Hymenoptera and Lepidoptera were alsoimportant and frequently occurring prey items inthese studies albeit less so than true flies and beetlesThus Hymenoptera were the most important preyin one case and accounted for more than 15 of thediet of three cases Arachnids bugs and Lepidopterawere never dominant possibly because of the relativelylow biomass and small prey size of some of these
Figure 5 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded as being important in the diet Importance in the diet is defined as for Figure 2 Data are shown only for those invertebrate taxafor which statistically significant differences occurred between bird groups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1389 P lt 001 (wader chicks gt wader adults passerine adults and chicks)Diptera larvae = 2130 P lt 0001 (wader adults gt passerine chicks) Hemiptera = 1178 P lt 005 (grouse chicks gt wader adultspasserine adults gt passerine chicks wader adults) Lepidoptera adults = 1714 P lt 001 (passerine chicks gt passerine adults)Lepidoptera larvae = 1254 P lt 005 (wader adults lt passerine adults and chicks) Oligochaeta = 3185 P lt 0001 (wader adultsgt passerine chicks)
χ42
χ42 χ
42
χ42
χ42 χ
42
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
Moorland bird diets
3
copy 2006 T
he Authors
Journal compilation copy
2006 British O
rnithologistsrsquo Union
Table 1
The number of dietary studies for each of the 14 study species according to broad location The number of study areas providing compositional or frequency ofoccurrence data on diet and hence contributing to the analyses in Figures 4 and 5 is also shown
Studies from outside Britain amp Ireland
Britain amp Ireland non-moorland
studies British moorlands
No of studies on adults and chicks
(respectively) used in analysis of invertebrates
occurring as
SourcesAdults Chicks Not Stated Adults Chicks Adults Chicks Taken lsquoImportantrsquo
(1984) Kondratrsquoyev and Kondratrsquoyeva (1988) Nechaev (1991) OrsquoConnell
et al
(1996) 6 ndash Zinovrsquoyev (1980) Danilov (1984) Green
et al
(1990) Nechaev (1991)7 ndash Kumari (1958) Cramp and Simmons (1983) Robson (1998) Stephen (2001) Grant unpubl data 8 ndash Peresadrsquoko and Koshelev (1980) Green (1986) Beintema
et al
(1991)Nechaev (1991) 9 ndash Nekrasov (1978) Malrsquochevskiy and Pukinskiy (1983) Jenny (1990) Nechaev (1991) Donald
et al
(2001) 10 ndash Hagvar and Ostbye (1976) Walton (1979)Prokofrsquoyeva (1980) Coulson and Whittaker (1978) Hagvar and Ostbye (1976) Danilov
et al
(1984) Burges (1994) Burges and Weildinger (2000) Evans unpubl data 11 ndashAverin and Ganya (1970 as cited in Cramp 1988) Brooke (1981) Kostin (1983) Gubin and Kovshar (1985) Dorzhiev and Khertuev (1992) 12 ndash Artemrsquoyev and Popov (1978)Prokofrsquoyeva (1980) Garnett
et al
(1988) von Labhardt (1988) 13 ndash Greig-Smith and Quicke (1983) Kostin (1983) Siepel (1990) Nechaev (1991) Cummins and OrsquoHalloran(2002) 14 ndash Tyler and Green (1994) Burfield (2002)
4
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Treatment of dietary data
Comparison across studies was difficult becausedietary data were reported to varying taxonomiclevels with some studies identifying the main inver-tebrate prey to the level of family genus or occasion-ally species whereas others only identified prey tothe level of class or order Consequently analyses wereperformed at two taxonomic levels to maximize thevalue of the available data First comparisons wereundertaken at a broader taxonomic level ie orderfor insects class for arachnids and annelids (althoughall mentions of worms in diet studies referred toearthworms (Lumbricidae) we use the broaderOligocheate for consistency) and higher level
s
forother invertebrates Secondly for the subset ofstudies that identified the main prey items to at leastthe level of family for insects and order for arachnidscomparisons were made at these finer taxonomiclevels although taxa distinguished in fewer thanfive studies were not used (eg Opilione or Araneaefamilies) Percentage composition data were summedacross the relevant finer taxonomic levels to producetotals for the broader level analysis (eg figures forColeopteran families would be combined to producea total for all Coleoptera) Where frequency of occur-rence data (ie the number or proportion of samplesin which the taxa were recorded) were presented onesample may have contained several families from aparticular order In this case the maximum percent-age occurrence figure for a finer level taxon (eg amongColeoptera families) was used to represent the per-centage occurrence of the broader taxonomic level(eg all Coleoptera) Insect taxa were distinguishedby life-cycle stage (adult or larval) where sufficientinformation was provided but were otherwise cate-gorized as lsquounagedrsquo
The methods used to collect dietary data and thetreatment of these data varied between studiesDifferent sampling methods used in these studiesare each subject to biases (Moreby amp Stoate 2000)preventing direct comparisons between studiesTherefore results from the dietary studies weresummarized and for each BSAC in each study areadata on invertebrate taxa were classed according to(1) their presence or absence and (2) whether or notthey either comprised at least 5 of the diet oroccurred in at least 5 of samples (for those studiesquantifying composition or frequency of occurrence)Where results were presented in different forms com-positional data were used in preference to frequencyof occurrence and biomass composition used in
preference to numerical composition The 5 level wasselected to distinguish the lsquoimportantrsquo taxa becausethis represented the most marked discontinuity inthe decline in the contribution of different inver-tebrate taxa to the bird diets (Fig 1) and concurs witha previous review of bird diets (Wilson
et al
1999)To compare the diets of different BSACs across all
studies data were summarized according to whethereach invertebrate taxon was recorded in any study ofthat species and whether its average percentagecomposition or percentage occurrence (across studyareas providing data for that species) was at least5 This was performed at both taxonomic levelsconsidered here
Determining differences between diets of bird families and ages
Statistical comparisons of the diets of different ageclasses of different moorland bird families wereundertaken (there being insufficient data for com-parisons between BSACs) Analyses were performedat the broader taxonomic level only owing to thesmall sample size available at the finer level lsquoUnagedrsquoinvertebrates were omitted as were data from studyareas providing no information on the samplingmethods and any taxa taken by or classed as lsquoimport-antrsquo in fewer than 10 of BSACs in the respectiveanalyses (Fig 2)
Analyses were conducted using (1) the presenceor absence of each invertebrate taxon and (2) whetherthe taxon was classed as lsquoimportantrsquo (based upon the5 criterion) as dependent variables Data were
Figure 1 The decline in the proportion of broader invertebratetaxa (see text) in relation to the increasing contribution made tobird diets (ie all taxa comprise between 0 and 5 of the dietbut none contributes more than 90) Data are derived from 14bird species (with data from adults and chicks treated separatelygiving 24 different categories) using studies that present dietarydata either as percentage composition or as percentageoccurrence in diet samples The proportions for each taxon arecalculated from the average of the mean values for the differentbird species and age categories
Moorland bird diets
5
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
categorized according to whether they were grousechicks wader adults wader chicks passerine adultsor passerine chicks Variation due to different sam-pling methods and to the localities and habitats fromwhich data were derived was taken into accountbefore assessing differences between bird groups Inaddition the treatment of the data (eg percentageoccurrence or numerical composition) was takeninto account in the analyses of the lsquoimportancersquo oftaxa Thus logistic
ANOVA
was undertaken usingProc GENMOD in SAS 802 (SAS Institute 2001)specifying a logit link and binomial error structure(see Endnote) first fitting sampling method location(ie BampI moorland BampI non-moorland non-BampI)and for analyses of lsquoimportancersquo data treatment
RESULTS
Coverage of bird species age categories variation with location and sampling methods
Data on the 14 moorland bird species considered inthis review were collated from 62 different sourcesWhen treated separately by study area and bird
species age category there were 115 diet studies
minus
31from British moorland habitats 12 from otherhabitats in BampI and 72 from outside BampI (Table 1)The extent to which data were available for adultbirds and chicks varied between these three categor-ies with 70 of the dietary studies from outsideBampI concerning adult birds compared with 50 and13 of those from non-moorland and moorlandhabitats respectively in BampI (Table 1 = 984
P =
0007) There were also differences in themethods used to collect dietary data with 74 ofBampI studies using faecal analysis compared with 9elsewhere where 81 used either crop or stomachsamples ( = 425
P lt
0001) A higher proportionof the dietary studies from outside BampI reported dataas presence or absence only (ie 33 compared with15 and 0 for moorland and non-moorland habitatsin BampI respectively) although this difference was notstatistically significant ( = 51
P =
008)
Trends in the prevalence and importance of invertebrate taxa
At the broader taxonomic level (order and above)adult and unaged Coleoptera (beetles) Diptera (true
Figure 2 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different broader invertebratetaxa were recorded as occurring in the diet (entire bar) and being of importance in the diet (open and thin hatched grey sections of barsonly) Data are presented for studies from all locations (solidopen bars) and from British moorland studies only (hatched bars)Importance in the diet is defined as comprising ge 5 of diet composition or as occurring in ge 5 of dietary samples (where the studypresents frequency of occurrence but not compositional data)
χ22
χ12
χ22
6
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
flies) Hemiptera (bugs) Hymenoptera Lepidopteralarvae Arachnida (arachnids) and Oligochaeta (worms)were the most ubiquitous prey all being taken by morethan 25 of the different BSACs (Fig 2) There was asimilar pattern in the occurrence of taxa when compar-isons were limited to lsquoimportantrsquo taxa only (ie thosecomprising
ge
5 of the diet or occurring in
ge
5 of thesamples on average for a BSAC) Arachnids adult andunaged beetles true flies bugs Hymenoptera andLepidoptera larvae were most prevalent as lsquoimportantrsquotaxa in the diets (Fig 2) Differences between thefrequency of occurrence of taxa and the frequency withwhich they occurred as lsquoimportantrsquo taxa were mostmarked for arachnids and Lepidoptera larvae Bothtaxa were taken by a wide range of BSACs but oftenformed a relatively small part of the diet Thesecomparisons indicated few major differences betweenthe data from all study areas combined and those fromBritish moorland only However among the mostfrequently occurring prey there was a markedly greaterrepresentation of both Hymenoptera (adults andlarvae) and adult true flies in the moorland diets
Fewer data were available for the comparisons atthe finer taxonomic level with 66 dietary studies
from 14 bird species (nine adult 11 chick) providingsufficient data at this level (Fig 3)
These comparisons suggested that Araneae (spiders)are more prevalent than Opiliones (harvestmen)and that although a wide range of beetle and true flyfamilies are taken only a few occur frequently Carabidae(carabids) Chrysomelidae (chrysomelids) Curcu-lionidae (curculionids) Elateridae (elaterids) andStaphylinidae (staphylinids) were the most frequentlytaken adult beetles being recorded in more than 35of the BSACs (Fig 3) Beetle larvae were recordedrelatively rarely True fly families tended to be lessprevalent in bird diets with only Chironomidae(chironomid) Bibionidae (bibionid) and Tipulidae(tipulid) adults recorded in more than 20 of thediets and each recorded in less than 30 Howeverlarval tipulids and Muscidae (muscids) were recordedin 52 and 24 of the diets respectively Of theHymenoptera Formicidae (formicids) (unaged) werethe only group that were relatively prevalent in the dietsWhen classifying diet in terms of the lsquoimportantrsquo taxathe decrease in occurrence was most marked for Araneae(spiders) chrysomelids staphylinids bibionids muscidsand formicids Comparisons of the prevalence of
Figure 3 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different finer level invertebratetaxa were recorded as occurring in the diet (filled plus open sections of bars) and being of importance in the diet (filled section only)Data are derived from studies from all locations Importance in the diet is defined as for Figure 2 For each taxon (eg Carabidae) columnsfrom left to right represent proportion of adults larvae and unaged except for Arachnids (Araneae and Opiliones) for which all are classedas adults
Moorland bird diets
7
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
invertebrates from studies on British moorlands onlywere not made at this taxonomic level because fewstudies provided such data (16 compared with 30 inFig 2) and few BSACs were represented (eightcompared with 15 in Fig 2)
Differences between bird families and age categories
Significant differences in the prevalence of inverte-brates were detected between the different samplingmethods for arachnids worms and Hymenopteralarvae and between locations for adult beetlesHymenoptera adults and Lepidoptera larvae ( gt117
P lt
001 in all cases) Among invertebrate taxathat were classed as lsquoimportantrsquo significant differencesoccurred between sampling methods for wormslocations for adult beetles and the method of datatreatment for larval true flies Gastropoda bugs andHymenoptera adults ( gt 78
P lt
005 in all cases)Owing to the high degree of association betweenthese different effects (particularly sampling methodand location ndash see above) these differences have tobe interpreted with caution
After taking the effects of sampling methodlocation and data treatment into account significantdifferences between bird families and age categorieswere found in both prevalence and importance for
adult and larval true flies adult and larval Lepidopteraand worms There were also significant differences inprevalence between bird families and age categoriesfor Gastropoda and in importance for bugs (Figs 4amp 5) These analyses suggested that adult true flieswere of greater importance to wader chicks than tothe other bird groups (although the difference withgrouse chicks was non-significant) whereas larvaltrue flies tended to be more important to both waderadults and chicks than to other groups By contrastadult and larval Lepidoptera were of most import-ance to both passerine adults and chicks but oflittle importance to the waders and grouse Wormswere of greatest importance to wader adultsAlthough frequently present in the diets of waderchicks they were never recorded as an lsquoimportantrsquotaxon and were absent from the diets of grousechicks Gastropods were most prevalent in diets ofadult passerines and waders whereas bugs occurredas an lsquoimportantrsquo taxon most frequently in the dietsof adult passerines and grouse chicks When inter-preting these results it must be remembered thatcertain of the bird groups were heavily influencedby data from one or two species for example foradult waders data from ten of the 40 studies werefrom Dunlin but only five were from CommonRedshank (Table 1) Thus certain comparisons mayreflect differences attributable to one or two species
χ22
χ22
Figure 4 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded in the diet Data are shown only for those invertebrate taxa for which statistically significant differences occurred between birdgroups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1729P lt 001 (wader chicks gt wader adults passerine adults lt passerine chicks wader chicks) Diptera larvae = 1700 P lt 001 (waderchicks gt passerine adults) Gastropoda = 2121 P lt 0001 (passerine chicks gt passerine adults) Lepidoptera adults = 2852P lt 0001 (passerine chicks gt wader adults) Lepidoptera larvae = 3052 P lt 0001 (passerine adults gt wader adults passerinechicks gt grouse chicks wader adults and chicks) Oligochaeta = 2677 P lt 0001 (wader adults gt passerine adults)
χ42
χ42
χ42 χ
42
χ42
χ42
8
G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
as opposed to being more widely representative ofthe relevant bird groups
DISCUSSION
The contribution of invertebrates to moorland bird diets
A wide range of invertebrates are consumed bymoorland birds but only a small number of thebroader level taxonomic groups are particularlyprevalent (ie arachnids beetles true flies bugsHymenoptera Lepidoptera and worms) With theexception of Hymenoptera and earthworms Coulson(1988) found that these taxa were among the mostabundant on moorland in northern England suggest-ing that birds are generally exploiting the most avail-able prey on moorlands The absence of Collembolaand enchytraeid worms from diets despite also beingnumerous is almost certainly due to their small sizeand hence lack of profitability as prey This diverserange of prey will partly reflect variation in diet betweenBSACs resulting from their different foragingstrategies (discussed below) However spatial andtemporal differences in prey availability within andbetween studies will also influence diet compositionas indicated by within-species dietary differencesassociated with different study locations and the fewstudies that have examined diet in relation to seasonaland between-habitat variation in invertebrateabundance (Picozzi amp Hepburn 1984 Starling-
Westerberg 2001 Whittingham et al 2001 Pearce-Higgins amp Yalden 2004)
Twelve studies presenting detailed compositionaldietary data for eight BSACs on British moorlands(Walton 1979 Picozzi amp Hepburn 1984 Garnettet al 1988 Cayford et al 1989 Whittingham 1996Starling-Westerberg 2001 Stephen 2001 Burfield2002 Pearce-Higgins amp Yalden 2003 2004) generallysupport the results of our wider analysis These studiesshowed that beetles formed the most importantitem for five of the BSACs considered comprisingover 15 of the diet of four cases True flies weremost important in four cases comprising over 15of the total diet in five The prevalence of adult trueflies in moorland bird diets especially among wadersand Red Grouse reflects in part the high abundanceof tipulids on some moorland areas following theirhighly synchronized emergences (Coulson 1962)Less important in this subset of more detailedmoorland studies were earthworms which were takenmainly by adult waders foraging on nearby improvedgrasslands but were also the most important item forRing Ouzel chicks As in our wider analysis arachnidsbugs Hymenoptera and Lepidoptera were alsoimportant and frequently occurring prey items inthese studies albeit less so than true flies and beetlesThus Hymenoptera were the most important preyin one case and accounted for more than 15 of thediet of three cases Arachnids bugs and Lepidopterawere never dominant possibly because of the relativelylow biomass and small prey size of some of these
Figure 5 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded as being important in the diet Importance in the diet is defined as for Figure 2 Data are shown only for those invertebrate taxafor which statistically significant differences occurred between bird groups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1389 P lt 001 (wader chicks gt wader adults passerine adults and chicks)Diptera larvae = 2130 P lt 0001 (wader adults gt passerine chicks) Hemiptera = 1178 P lt 005 (grouse chicks gt wader adultspasserine adults gt passerine chicks wader adults) Lepidoptera adults = 1714 P lt 001 (passerine chicks gt passerine adults)Lepidoptera larvae = 1254 P lt 005 (wader adults lt passerine adults and chicks) Oligochaeta = 3185 P lt 0001 (wader adultsgt passerine chicks)
χ42
χ42 χ
42
χ42
χ42 χ
42
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
4
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Treatment of dietary data
Comparison across studies was difficult becausedietary data were reported to varying taxonomiclevels with some studies identifying the main inver-tebrate prey to the level of family genus or occasion-ally species whereas others only identified prey tothe level of class or order Consequently analyses wereperformed at two taxonomic levels to maximize thevalue of the available data First comparisons wereundertaken at a broader taxonomic level ie orderfor insects class for arachnids and annelids (althoughall mentions of worms in diet studies referred toearthworms (Lumbricidae) we use the broaderOligocheate for consistency) and higher level
s
forother invertebrates Secondly for the subset ofstudies that identified the main prey items to at leastthe level of family for insects and order for arachnidscomparisons were made at these finer taxonomiclevels although taxa distinguished in fewer thanfive studies were not used (eg Opilione or Araneaefamilies) Percentage composition data were summedacross the relevant finer taxonomic levels to producetotals for the broader level analysis (eg figures forColeopteran families would be combined to producea total for all Coleoptera) Where frequency of occur-rence data (ie the number or proportion of samplesin which the taxa were recorded) were presented onesample may have contained several families from aparticular order In this case the maximum percent-age occurrence figure for a finer level taxon (eg amongColeoptera families) was used to represent the per-centage occurrence of the broader taxonomic level(eg all Coleoptera) Insect taxa were distinguishedby life-cycle stage (adult or larval) where sufficientinformation was provided but were otherwise cate-gorized as lsquounagedrsquo
The methods used to collect dietary data and thetreatment of these data varied between studiesDifferent sampling methods used in these studiesare each subject to biases (Moreby amp Stoate 2000)preventing direct comparisons between studiesTherefore results from the dietary studies weresummarized and for each BSAC in each study areadata on invertebrate taxa were classed according to(1) their presence or absence and (2) whether or notthey either comprised at least 5 of the diet oroccurred in at least 5 of samples (for those studiesquantifying composition or frequency of occurrence)Where results were presented in different forms com-positional data were used in preference to frequencyof occurrence and biomass composition used in
preference to numerical composition The 5 level wasselected to distinguish the lsquoimportantrsquo taxa becausethis represented the most marked discontinuity inthe decline in the contribution of different inver-tebrate taxa to the bird diets (Fig 1) and concurs witha previous review of bird diets (Wilson
et al
1999)To compare the diets of different BSACs across all
studies data were summarized according to whethereach invertebrate taxon was recorded in any study ofthat species and whether its average percentagecomposition or percentage occurrence (across studyareas providing data for that species) was at least5 This was performed at both taxonomic levelsconsidered here
Determining differences between diets of bird families and ages
Statistical comparisons of the diets of different ageclasses of different moorland bird families wereundertaken (there being insufficient data for com-parisons between BSACs) Analyses were performedat the broader taxonomic level only owing to thesmall sample size available at the finer level lsquoUnagedrsquoinvertebrates were omitted as were data from studyareas providing no information on the samplingmethods and any taxa taken by or classed as lsquoimport-antrsquo in fewer than 10 of BSACs in the respectiveanalyses (Fig 2)
Analyses were conducted using (1) the presenceor absence of each invertebrate taxon and (2) whetherthe taxon was classed as lsquoimportantrsquo (based upon the5 criterion) as dependent variables Data were
Figure 1 The decline in the proportion of broader invertebratetaxa (see text) in relation to the increasing contribution made tobird diets (ie all taxa comprise between 0 and 5 of the dietbut none contributes more than 90) Data are derived from 14bird species (with data from adults and chicks treated separatelygiving 24 different categories) using studies that present dietarydata either as percentage composition or as percentageoccurrence in diet samples The proportions for each taxon arecalculated from the average of the mean values for the differentbird species and age categories
Moorland bird diets
5
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
categorized according to whether they were grousechicks wader adults wader chicks passerine adultsor passerine chicks Variation due to different sam-pling methods and to the localities and habitats fromwhich data were derived was taken into accountbefore assessing differences between bird groups Inaddition the treatment of the data (eg percentageoccurrence or numerical composition) was takeninto account in the analyses of the lsquoimportancersquo oftaxa Thus logistic
ANOVA
was undertaken usingProc GENMOD in SAS 802 (SAS Institute 2001)specifying a logit link and binomial error structure(see Endnote) first fitting sampling method location(ie BampI moorland BampI non-moorland non-BampI)and for analyses of lsquoimportancersquo data treatment
RESULTS
Coverage of bird species age categories variation with location and sampling methods
Data on the 14 moorland bird species considered inthis review were collated from 62 different sourcesWhen treated separately by study area and bird
species age category there were 115 diet studies
minus
31from British moorland habitats 12 from otherhabitats in BampI and 72 from outside BampI (Table 1)The extent to which data were available for adultbirds and chicks varied between these three categor-ies with 70 of the dietary studies from outsideBampI concerning adult birds compared with 50 and13 of those from non-moorland and moorlandhabitats respectively in BampI (Table 1 = 984
P =
0007) There were also differences in themethods used to collect dietary data with 74 ofBampI studies using faecal analysis compared with 9elsewhere where 81 used either crop or stomachsamples ( = 425
P lt
0001) A higher proportionof the dietary studies from outside BampI reported dataas presence or absence only (ie 33 compared with15 and 0 for moorland and non-moorland habitatsin BampI respectively) although this difference was notstatistically significant ( = 51
P =
008)
Trends in the prevalence and importance of invertebrate taxa
At the broader taxonomic level (order and above)adult and unaged Coleoptera (beetles) Diptera (true
Figure 2 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different broader invertebratetaxa were recorded as occurring in the diet (entire bar) and being of importance in the diet (open and thin hatched grey sections of barsonly) Data are presented for studies from all locations (solidopen bars) and from British moorland studies only (hatched bars)Importance in the diet is defined as comprising ge 5 of diet composition or as occurring in ge 5 of dietary samples (where the studypresents frequency of occurrence but not compositional data)
χ22
χ12
χ22
6
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
flies) Hemiptera (bugs) Hymenoptera Lepidopteralarvae Arachnida (arachnids) and Oligochaeta (worms)were the most ubiquitous prey all being taken by morethan 25 of the different BSACs (Fig 2) There was asimilar pattern in the occurrence of taxa when compar-isons were limited to lsquoimportantrsquo taxa only (ie thosecomprising
ge
5 of the diet or occurring in
ge
5 of thesamples on average for a BSAC) Arachnids adult andunaged beetles true flies bugs Hymenoptera andLepidoptera larvae were most prevalent as lsquoimportantrsquotaxa in the diets (Fig 2) Differences between thefrequency of occurrence of taxa and the frequency withwhich they occurred as lsquoimportantrsquo taxa were mostmarked for arachnids and Lepidoptera larvae Bothtaxa were taken by a wide range of BSACs but oftenformed a relatively small part of the diet Thesecomparisons indicated few major differences betweenthe data from all study areas combined and those fromBritish moorland only However among the mostfrequently occurring prey there was a markedly greaterrepresentation of both Hymenoptera (adults andlarvae) and adult true flies in the moorland diets
Fewer data were available for the comparisons atthe finer taxonomic level with 66 dietary studies
from 14 bird species (nine adult 11 chick) providingsufficient data at this level (Fig 3)
These comparisons suggested that Araneae (spiders)are more prevalent than Opiliones (harvestmen)and that although a wide range of beetle and true flyfamilies are taken only a few occur frequently Carabidae(carabids) Chrysomelidae (chrysomelids) Curcu-lionidae (curculionids) Elateridae (elaterids) andStaphylinidae (staphylinids) were the most frequentlytaken adult beetles being recorded in more than 35of the BSACs (Fig 3) Beetle larvae were recordedrelatively rarely True fly families tended to be lessprevalent in bird diets with only Chironomidae(chironomid) Bibionidae (bibionid) and Tipulidae(tipulid) adults recorded in more than 20 of thediets and each recorded in less than 30 Howeverlarval tipulids and Muscidae (muscids) were recordedin 52 and 24 of the diets respectively Of theHymenoptera Formicidae (formicids) (unaged) werethe only group that were relatively prevalent in the dietsWhen classifying diet in terms of the lsquoimportantrsquo taxathe decrease in occurrence was most marked for Araneae(spiders) chrysomelids staphylinids bibionids muscidsand formicids Comparisons of the prevalence of
Figure 3 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different finer level invertebratetaxa were recorded as occurring in the diet (filled plus open sections of bars) and being of importance in the diet (filled section only)Data are derived from studies from all locations Importance in the diet is defined as for Figure 2 For each taxon (eg Carabidae) columnsfrom left to right represent proportion of adults larvae and unaged except for Arachnids (Araneae and Opiliones) for which all are classedas adults
Moorland bird diets
7
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
invertebrates from studies on British moorlands onlywere not made at this taxonomic level because fewstudies provided such data (16 compared with 30 inFig 2) and few BSACs were represented (eightcompared with 15 in Fig 2)
Differences between bird families and age categories
Significant differences in the prevalence of inverte-brates were detected between the different samplingmethods for arachnids worms and Hymenopteralarvae and between locations for adult beetlesHymenoptera adults and Lepidoptera larvae ( gt117
P lt
001 in all cases) Among invertebrate taxathat were classed as lsquoimportantrsquo significant differencesoccurred between sampling methods for wormslocations for adult beetles and the method of datatreatment for larval true flies Gastropoda bugs andHymenoptera adults ( gt 78
P lt
005 in all cases)Owing to the high degree of association betweenthese different effects (particularly sampling methodand location ndash see above) these differences have tobe interpreted with caution
After taking the effects of sampling methodlocation and data treatment into account significantdifferences between bird families and age categorieswere found in both prevalence and importance for
adult and larval true flies adult and larval Lepidopteraand worms There were also significant differences inprevalence between bird families and age categoriesfor Gastropoda and in importance for bugs (Figs 4amp 5) These analyses suggested that adult true flieswere of greater importance to wader chicks than tothe other bird groups (although the difference withgrouse chicks was non-significant) whereas larvaltrue flies tended to be more important to both waderadults and chicks than to other groups By contrastadult and larval Lepidoptera were of most import-ance to both passerine adults and chicks but oflittle importance to the waders and grouse Wormswere of greatest importance to wader adultsAlthough frequently present in the diets of waderchicks they were never recorded as an lsquoimportantrsquotaxon and were absent from the diets of grousechicks Gastropods were most prevalent in diets ofadult passerines and waders whereas bugs occurredas an lsquoimportantrsquo taxon most frequently in the dietsof adult passerines and grouse chicks When inter-preting these results it must be remembered thatcertain of the bird groups were heavily influencedby data from one or two species for example foradult waders data from ten of the 40 studies werefrom Dunlin but only five were from CommonRedshank (Table 1) Thus certain comparisons mayreflect differences attributable to one or two species
χ22
χ22
Figure 4 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded in the diet Data are shown only for those invertebrate taxa for which statistically significant differences occurred between birdgroups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1729P lt 001 (wader chicks gt wader adults passerine adults lt passerine chicks wader chicks) Diptera larvae = 1700 P lt 001 (waderchicks gt passerine adults) Gastropoda = 2121 P lt 0001 (passerine chicks gt passerine adults) Lepidoptera adults = 2852P lt 0001 (passerine chicks gt wader adults) Lepidoptera larvae = 3052 P lt 0001 (passerine adults gt wader adults passerinechicks gt grouse chicks wader adults and chicks) Oligochaeta = 2677 P lt 0001 (wader adults gt passerine adults)
χ42
χ42
χ42 χ
42
χ42
χ42
8
G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
as opposed to being more widely representative ofthe relevant bird groups
DISCUSSION
The contribution of invertebrates to moorland bird diets
A wide range of invertebrates are consumed bymoorland birds but only a small number of thebroader level taxonomic groups are particularlyprevalent (ie arachnids beetles true flies bugsHymenoptera Lepidoptera and worms) With theexception of Hymenoptera and earthworms Coulson(1988) found that these taxa were among the mostabundant on moorland in northern England suggest-ing that birds are generally exploiting the most avail-able prey on moorlands The absence of Collembolaand enchytraeid worms from diets despite also beingnumerous is almost certainly due to their small sizeand hence lack of profitability as prey This diverserange of prey will partly reflect variation in diet betweenBSACs resulting from their different foragingstrategies (discussed below) However spatial andtemporal differences in prey availability within andbetween studies will also influence diet compositionas indicated by within-species dietary differencesassociated with different study locations and the fewstudies that have examined diet in relation to seasonaland between-habitat variation in invertebrateabundance (Picozzi amp Hepburn 1984 Starling-
Westerberg 2001 Whittingham et al 2001 Pearce-Higgins amp Yalden 2004)
Twelve studies presenting detailed compositionaldietary data for eight BSACs on British moorlands(Walton 1979 Picozzi amp Hepburn 1984 Garnettet al 1988 Cayford et al 1989 Whittingham 1996Starling-Westerberg 2001 Stephen 2001 Burfield2002 Pearce-Higgins amp Yalden 2003 2004) generallysupport the results of our wider analysis These studiesshowed that beetles formed the most importantitem for five of the BSACs considered comprisingover 15 of the diet of four cases True flies weremost important in four cases comprising over 15of the total diet in five The prevalence of adult trueflies in moorland bird diets especially among wadersand Red Grouse reflects in part the high abundanceof tipulids on some moorland areas following theirhighly synchronized emergences (Coulson 1962)Less important in this subset of more detailedmoorland studies were earthworms which were takenmainly by adult waders foraging on nearby improvedgrasslands but were also the most important item forRing Ouzel chicks As in our wider analysis arachnidsbugs Hymenoptera and Lepidoptera were alsoimportant and frequently occurring prey items inthese studies albeit less so than true flies and beetlesThus Hymenoptera were the most important preyin one case and accounted for more than 15 of thediet of three cases Arachnids bugs and Lepidopterawere never dominant possibly because of the relativelylow biomass and small prey size of some of these
Figure 5 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded as being important in the diet Importance in the diet is defined as for Figure 2 Data are shown only for those invertebrate taxafor which statistically significant differences occurred between bird groups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1389 P lt 001 (wader chicks gt wader adults passerine adults and chicks)Diptera larvae = 2130 P lt 0001 (wader adults gt passerine chicks) Hemiptera = 1178 P lt 005 (grouse chicks gt wader adultspasserine adults gt passerine chicks wader adults) Lepidoptera adults = 1714 P lt 001 (passerine chicks gt passerine adults)Lepidoptera larvae = 1254 P lt 005 (wader adults lt passerine adults and chicks) Oligochaeta = 3185 P lt 0001 (wader adultsgt passerine chicks)
χ42
χ42 χ
42
χ42
χ42 χ
42
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
Moorland bird diets
5
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
categorized according to whether they were grousechicks wader adults wader chicks passerine adultsor passerine chicks Variation due to different sam-pling methods and to the localities and habitats fromwhich data were derived was taken into accountbefore assessing differences between bird groups Inaddition the treatment of the data (eg percentageoccurrence or numerical composition) was takeninto account in the analyses of the lsquoimportancersquo oftaxa Thus logistic
ANOVA
was undertaken usingProc GENMOD in SAS 802 (SAS Institute 2001)specifying a logit link and binomial error structure(see Endnote) first fitting sampling method location(ie BampI moorland BampI non-moorland non-BampI)and for analyses of lsquoimportancersquo data treatment
RESULTS
Coverage of bird species age categories variation with location and sampling methods
Data on the 14 moorland bird species considered inthis review were collated from 62 different sourcesWhen treated separately by study area and bird
species age category there were 115 diet studies
minus
31from British moorland habitats 12 from otherhabitats in BampI and 72 from outside BampI (Table 1)The extent to which data were available for adultbirds and chicks varied between these three categor-ies with 70 of the dietary studies from outsideBampI concerning adult birds compared with 50 and13 of those from non-moorland and moorlandhabitats respectively in BampI (Table 1 = 984
P =
0007) There were also differences in themethods used to collect dietary data with 74 ofBampI studies using faecal analysis compared with 9elsewhere where 81 used either crop or stomachsamples ( = 425
P lt
0001) A higher proportionof the dietary studies from outside BampI reported dataas presence or absence only (ie 33 compared with15 and 0 for moorland and non-moorland habitatsin BampI respectively) although this difference was notstatistically significant ( = 51
P =
008)
Trends in the prevalence and importance of invertebrate taxa
At the broader taxonomic level (order and above)adult and unaged Coleoptera (beetles) Diptera (true
Figure 2 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different broader invertebratetaxa were recorded as occurring in the diet (entire bar) and being of importance in the diet (open and thin hatched grey sections of barsonly) Data are presented for studies from all locations (solidopen bars) and from British moorland studies only (hatched bars)Importance in the diet is defined as comprising ge 5 of diet composition or as occurring in ge 5 of dietary samples (where the studypresents frequency of occurrence but not compositional data)
χ22
χ12
χ22
6
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
flies) Hemiptera (bugs) Hymenoptera Lepidopteralarvae Arachnida (arachnids) and Oligochaeta (worms)were the most ubiquitous prey all being taken by morethan 25 of the different BSACs (Fig 2) There was asimilar pattern in the occurrence of taxa when compar-isons were limited to lsquoimportantrsquo taxa only (ie thosecomprising
ge
5 of the diet or occurring in
ge
5 of thesamples on average for a BSAC) Arachnids adult andunaged beetles true flies bugs Hymenoptera andLepidoptera larvae were most prevalent as lsquoimportantrsquotaxa in the diets (Fig 2) Differences between thefrequency of occurrence of taxa and the frequency withwhich they occurred as lsquoimportantrsquo taxa were mostmarked for arachnids and Lepidoptera larvae Bothtaxa were taken by a wide range of BSACs but oftenformed a relatively small part of the diet Thesecomparisons indicated few major differences betweenthe data from all study areas combined and those fromBritish moorland only However among the mostfrequently occurring prey there was a markedly greaterrepresentation of both Hymenoptera (adults andlarvae) and adult true flies in the moorland diets
Fewer data were available for the comparisons atthe finer taxonomic level with 66 dietary studies
from 14 bird species (nine adult 11 chick) providingsufficient data at this level (Fig 3)
These comparisons suggested that Araneae (spiders)are more prevalent than Opiliones (harvestmen)and that although a wide range of beetle and true flyfamilies are taken only a few occur frequently Carabidae(carabids) Chrysomelidae (chrysomelids) Curcu-lionidae (curculionids) Elateridae (elaterids) andStaphylinidae (staphylinids) were the most frequentlytaken adult beetles being recorded in more than 35of the BSACs (Fig 3) Beetle larvae were recordedrelatively rarely True fly families tended to be lessprevalent in bird diets with only Chironomidae(chironomid) Bibionidae (bibionid) and Tipulidae(tipulid) adults recorded in more than 20 of thediets and each recorded in less than 30 Howeverlarval tipulids and Muscidae (muscids) were recordedin 52 and 24 of the diets respectively Of theHymenoptera Formicidae (formicids) (unaged) werethe only group that were relatively prevalent in the dietsWhen classifying diet in terms of the lsquoimportantrsquo taxathe decrease in occurrence was most marked for Araneae(spiders) chrysomelids staphylinids bibionids muscidsand formicids Comparisons of the prevalence of
Figure 3 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different finer level invertebratetaxa were recorded as occurring in the diet (filled plus open sections of bars) and being of importance in the diet (filled section only)Data are derived from studies from all locations Importance in the diet is defined as for Figure 2 For each taxon (eg Carabidae) columnsfrom left to right represent proportion of adults larvae and unaged except for Arachnids (Araneae and Opiliones) for which all are classedas adults
Moorland bird diets
7
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
invertebrates from studies on British moorlands onlywere not made at this taxonomic level because fewstudies provided such data (16 compared with 30 inFig 2) and few BSACs were represented (eightcompared with 15 in Fig 2)
Differences between bird families and age categories
Significant differences in the prevalence of inverte-brates were detected between the different samplingmethods for arachnids worms and Hymenopteralarvae and between locations for adult beetlesHymenoptera adults and Lepidoptera larvae ( gt117
P lt
001 in all cases) Among invertebrate taxathat were classed as lsquoimportantrsquo significant differencesoccurred between sampling methods for wormslocations for adult beetles and the method of datatreatment for larval true flies Gastropoda bugs andHymenoptera adults ( gt 78
P lt
005 in all cases)Owing to the high degree of association betweenthese different effects (particularly sampling methodand location ndash see above) these differences have tobe interpreted with caution
After taking the effects of sampling methodlocation and data treatment into account significantdifferences between bird families and age categorieswere found in both prevalence and importance for
adult and larval true flies adult and larval Lepidopteraand worms There were also significant differences inprevalence between bird families and age categoriesfor Gastropoda and in importance for bugs (Figs 4amp 5) These analyses suggested that adult true flieswere of greater importance to wader chicks than tothe other bird groups (although the difference withgrouse chicks was non-significant) whereas larvaltrue flies tended to be more important to both waderadults and chicks than to other groups By contrastadult and larval Lepidoptera were of most import-ance to both passerine adults and chicks but oflittle importance to the waders and grouse Wormswere of greatest importance to wader adultsAlthough frequently present in the diets of waderchicks they were never recorded as an lsquoimportantrsquotaxon and were absent from the diets of grousechicks Gastropods were most prevalent in diets ofadult passerines and waders whereas bugs occurredas an lsquoimportantrsquo taxon most frequently in the dietsof adult passerines and grouse chicks When inter-preting these results it must be remembered thatcertain of the bird groups were heavily influencedby data from one or two species for example foradult waders data from ten of the 40 studies werefrom Dunlin but only five were from CommonRedshank (Table 1) Thus certain comparisons mayreflect differences attributable to one or two species
χ22
χ22
Figure 4 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded in the diet Data are shown only for those invertebrate taxa for which statistically significant differences occurred between birdgroups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1729P lt 001 (wader chicks gt wader adults passerine adults lt passerine chicks wader chicks) Diptera larvae = 1700 P lt 001 (waderchicks gt passerine adults) Gastropoda = 2121 P lt 0001 (passerine chicks gt passerine adults) Lepidoptera adults = 2852P lt 0001 (passerine chicks gt wader adults) Lepidoptera larvae = 3052 P lt 0001 (passerine adults gt wader adults passerinechicks gt grouse chicks wader adults and chicks) Oligochaeta = 2677 P lt 0001 (wader adults gt passerine adults)
χ42
χ42
χ42 χ
42
χ42
χ42
8
G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
as opposed to being more widely representative ofthe relevant bird groups
DISCUSSION
The contribution of invertebrates to moorland bird diets
A wide range of invertebrates are consumed bymoorland birds but only a small number of thebroader level taxonomic groups are particularlyprevalent (ie arachnids beetles true flies bugsHymenoptera Lepidoptera and worms) With theexception of Hymenoptera and earthworms Coulson(1988) found that these taxa were among the mostabundant on moorland in northern England suggest-ing that birds are generally exploiting the most avail-able prey on moorlands The absence of Collembolaand enchytraeid worms from diets despite also beingnumerous is almost certainly due to their small sizeand hence lack of profitability as prey This diverserange of prey will partly reflect variation in diet betweenBSACs resulting from their different foragingstrategies (discussed below) However spatial andtemporal differences in prey availability within andbetween studies will also influence diet compositionas indicated by within-species dietary differencesassociated with different study locations and the fewstudies that have examined diet in relation to seasonaland between-habitat variation in invertebrateabundance (Picozzi amp Hepburn 1984 Starling-
Westerberg 2001 Whittingham et al 2001 Pearce-Higgins amp Yalden 2004)
Twelve studies presenting detailed compositionaldietary data for eight BSACs on British moorlands(Walton 1979 Picozzi amp Hepburn 1984 Garnettet al 1988 Cayford et al 1989 Whittingham 1996Starling-Westerberg 2001 Stephen 2001 Burfield2002 Pearce-Higgins amp Yalden 2003 2004) generallysupport the results of our wider analysis These studiesshowed that beetles formed the most importantitem for five of the BSACs considered comprisingover 15 of the diet of four cases True flies weremost important in four cases comprising over 15of the total diet in five The prevalence of adult trueflies in moorland bird diets especially among wadersand Red Grouse reflects in part the high abundanceof tipulids on some moorland areas following theirhighly synchronized emergences (Coulson 1962)Less important in this subset of more detailedmoorland studies were earthworms which were takenmainly by adult waders foraging on nearby improvedgrasslands but were also the most important item forRing Ouzel chicks As in our wider analysis arachnidsbugs Hymenoptera and Lepidoptera were alsoimportant and frequently occurring prey items inthese studies albeit less so than true flies and beetlesThus Hymenoptera were the most important preyin one case and accounted for more than 15 of thediet of three cases Arachnids bugs and Lepidopterawere never dominant possibly because of the relativelylow biomass and small prey size of some of these
Figure 5 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded as being important in the diet Importance in the diet is defined as for Figure 2 Data are shown only for those invertebrate taxafor which statistically significant differences occurred between bird groups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1389 P lt 001 (wader chicks gt wader adults passerine adults and chicks)Diptera larvae = 2130 P lt 0001 (wader adults gt passerine chicks) Hemiptera = 1178 P lt 005 (grouse chicks gt wader adultspasserine adults gt passerine chicks wader adults) Lepidoptera adults = 1714 P lt 001 (passerine chicks gt passerine adults)Lepidoptera larvae = 1254 P lt 005 (wader adults lt passerine adults and chicks) Oligochaeta = 3185 P lt 0001 (wader adultsgt passerine chicks)
χ42
χ42 χ
42
χ42
χ42 χ
42
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
6
G M Buchanan
et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
flies) Hemiptera (bugs) Hymenoptera Lepidopteralarvae Arachnida (arachnids) and Oligochaeta (worms)were the most ubiquitous prey all being taken by morethan 25 of the different BSACs (Fig 2) There was asimilar pattern in the occurrence of taxa when compar-isons were limited to lsquoimportantrsquo taxa only (ie thosecomprising
ge
5 of the diet or occurring in
ge
5 of thesamples on average for a BSAC) Arachnids adult andunaged beetles true flies bugs Hymenoptera andLepidoptera larvae were most prevalent as lsquoimportantrsquotaxa in the diets (Fig 2) Differences between thefrequency of occurrence of taxa and the frequency withwhich they occurred as lsquoimportantrsquo taxa were mostmarked for arachnids and Lepidoptera larvae Bothtaxa were taken by a wide range of BSACs but oftenformed a relatively small part of the diet Thesecomparisons indicated few major differences betweenthe data from all study areas combined and those fromBritish moorland only However among the mostfrequently occurring prey there was a markedly greaterrepresentation of both Hymenoptera (adults andlarvae) and adult true flies in the moorland diets
Fewer data were available for the comparisons atthe finer taxonomic level with 66 dietary studies
from 14 bird species (nine adult 11 chick) providingsufficient data at this level (Fig 3)
These comparisons suggested that Araneae (spiders)are more prevalent than Opiliones (harvestmen)and that although a wide range of beetle and true flyfamilies are taken only a few occur frequently Carabidae(carabids) Chrysomelidae (chrysomelids) Curcu-lionidae (curculionids) Elateridae (elaterids) andStaphylinidae (staphylinids) were the most frequentlytaken adult beetles being recorded in more than 35of the BSACs (Fig 3) Beetle larvae were recordedrelatively rarely True fly families tended to be lessprevalent in bird diets with only Chironomidae(chironomid) Bibionidae (bibionid) and Tipulidae(tipulid) adults recorded in more than 20 of thediets and each recorded in less than 30 Howeverlarval tipulids and Muscidae (muscids) were recordedin 52 and 24 of the diets respectively Of theHymenoptera Formicidae (formicids) (unaged) werethe only group that were relatively prevalent in the dietsWhen classifying diet in terms of the lsquoimportantrsquo taxathe decrease in occurrence was most marked for Araneae(spiders) chrysomelids staphylinids bibionids muscidsand formicids Comparisons of the prevalence of
Figure 3 The number of bird species age categories (ie adults or chicks or both of 14 species) in which different finer level invertebratetaxa were recorded as occurring in the diet (filled plus open sections of bars) and being of importance in the diet (filled section only)Data are derived from studies from all locations Importance in the diet is defined as for Figure 2 For each taxon (eg Carabidae) columnsfrom left to right represent proportion of adults larvae and unaged except for Arachnids (Araneae and Opiliones) for which all are classedas adults
Moorland bird diets
7
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
invertebrates from studies on British moorlands onlywere not made at this taxonomic level because fewstudies provided such data (16 compared with 30 inFig 2) and few BSACs were represented (eightcompared with 15 in Fig 2)
Differences between bird families and age categories
Significant differences in the prevalence of inverte-brates were detected between the different samplingmethods for arachnids worms and Hymenopteralarvae and between locations for adult beetlesHymenoptera adults and Lepidoptera larvae ( gt117
P lt
001 in all cases) Among invertebrate taxathat were classed as lsquoimportantrsquo significant differencesoccurred between sampling methods for wormslocations for adult beetles and the method of datatreatment for larval true flies Gastropoda bugs andHymenoptera adults ( gt 78
P lt
005 in all cases)Owing to the high degree of association betweenthese different effects (particularly sampling methodand location ndash see above) these differences have tobe interpreted with caution
After taking the effects of sampling methodlocation and data treatment into account significantdifferences between bird families and age categorieswere found in both prevalence and importance for
adult and larval true flies adult and larval Lepidopteraand worms There were also significant differences inprevalence between bird families and age categoriesfor Gastropoda and in importance for bugs (Figs 4amp 5) These analyses suggested that adult true flieswere of greater importance to wader chicks than tothe other bird groups (although the difference withgrouse chicks was non-significant) whereas larvaltrue flies tended to be more important to both waderadults and chicks than to other groups By contrastadult and larval Lepidoptera were of most import-ance to both passerine adults and chicks but oflittle importance to the waders and grouse Wormswere of greatest importance to wader adultsAlthough frequently present in the diets of waderchicks they were never recorded as an lsquoimportantrsquotaxon and were absent from the diets of grousechicks Gastropods were most prevalent in diets ofadult passerines and waders whereas bugs occurredas an lsquoimportantrsquo taxon most frequently in the dietsof adult passerines and grouse chicks When inter-preting these results it must be remembered thatcertain of the bird groups were heavily influencedby data from one or two species for example foradult waders data from ten of the 40 studies werefrom Dunlin but only five were from CommonRedshank (Table 1) Thus certain comparisons mayreflect differences attributable to one or two species
χ22
χ22
Figure 4 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded in the diet Data are shown only for those invertebrate taxa for which statistically significant differences occurred between birdgroups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1729P lt 001 (wader chicks gt wader adults passerine adults lt passerine chicks wader chicks) Diptera larvae = 1700 P lt 001 (waderchicks gt passerine adults) Gastropoda = 2121 P lt 0001 (passerine chicks gt passerine adults) Lepidoptera adults = 2852P lt 0001 (passerine chicks gt wader adults) Lepidoptera larvae = 3052 P lt 0001 (passerine adults gt wader adults passerinechicks gt grouse chicks wader adults and chicks) Oligochaeta = 2677 P lt 0001 (wader adults gt passerine adults)
χ42
χ42
χ42 χ
42
χ42
χ42
8
G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
as opposed to being more widely representative ofthe relevant bird groups
DISCUSSION
The contribution of invertebrates to moorland bird diets
A wide range of invertebrates are consumed bymoorland birds but only a small number of thebroader level taxonomic groups are particularlyprevalent (ie arachnids beetles true flies bugsHymenoptera Lepidoptera and worms) With theexception of Hymenoptera and earthworms Coulson(1988) found that these taxa were among the mostabundant on moorland in northern England suggest-ing that birds are generally exploiting the most avail-able prey on moorlands The absence of Collembolaand enchytraeid worms from diets despite also beingnumerous is almost certainly due to their small sizeand hence lack of profitability as prey This diverserange of prey will partly reflect variation in diet betweenBSACs resulting from their different foragingstrategies (discussed below) However spatial andtemporal differences in prey availability within andbetween studies will also influence diet compositionas indicated by within-species dietary differencesassociated with different study locations and the fewstudies that have examined diet in relation to seasonaland between-habitat variation in invertebrateabundance (Picozzi amp Hepburn 1984 Starling-
Westerberg 2001 Whittingham et al 2001 Pearce-Higgins amp Yalden 2004)
Twelve studies presenting detailed compositionaldietary data for eight BSACs on British moorlands(Walton 1979 Picozzi amp Hepburn 1984 Garnettet al 1988 Cayford et al 1989 Whittingham 1996Starling-Westerberg 2001 Stephen 2001 Burfield2002 Pearce-Higgins amp Yalden 2003 2004) generallysupport the results of our wider analysis These studiesshowed that beetles formed the most importantitem for five of the BSACs considered comprisingover 15 of the diet of four cases True flies weremost important in four cases comprising over 15of the total diet in five The prevalence of adult trueflies in moorland bird diets especially among wadersand Red Grouse reflects in part the high abundanceof tipulids on some moorland areas following theirhighly synchronized emergences (Coulson 1962)Less important in this subset of more detailedmoorland studies were earthworms which were takenmainly by adult waders foraging on nearby improvedgrasslands but were also the most important item forRing Ouzel chicks As in our wider analysis arachnidsbugs Hymenoptera and Lepidoptera were alsoimportant and frequently occurring prey items inthese studies albeit less so than true flies and beetlesThus Hymenoptera were the most important preyin one case and accounted for more than 15 of thediet of three cases Arachnids bugs and Lepidopterawere never dominant possibly because of the relativelylow biomass and small prey size of some of these
Figure 5 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded as being important in the diet Importance in the diet is defined as for Figure 2 Data are shown only for those invertebrate taxafor which statistically significant differences occurred between bird groups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1389 P lt 001 (wader chicks gt wader adults passerine adults and chicks)Diptera larvae = 2130 P lt 0001 (wader adults gt passerine chicks) Hemiptera = 1178 P lt 005 (grouse chicks gt wader adultspasserine adults gt passerine chicks wader adults) Lepidoptera adults = 1714 P lt 001 (passerine chicks gt passerine adults)Lepidoptera larvae = 1254 P lt 005 (wader adults lt passerine adults and chicks) Oligochaeta = 3185 P lt 0001 (wader adultsgt passerine chicks)
χ42
χ42 χ
42
χ42
χ42 χ
42
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
Moorland bird diets
7
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
invertebrates from studies on British moorlands onlywere not made at this taxonomic level because fewstudies provided such data (16 compared with 30 inFig 2) and few BSACs were represented (eightcompared with 15 in Fig 2)
Differences between bird families and age categories
Significant differences in the prevalence of inverte-brates were detected between the different samplingmethods for arachnids worms and Hymenopteralarvae and between locations for adult beetlesHymenoptera adults and Lepidoptera larvae ( gt117
P lt
001 in all cases) Among invertebrate taxathat were classed as lsquoimportantrsquo significant differencesoccurred between sampling methods for wormslocations for adult beetles and the method of datatreatment for larval true flies Gastropoda bugs andHymenoptera adults ( gt 78
P lt
005 in all cases)Owing to the high degree of association betweenthese different effects (particularly sampling methodand location ndash see above) these differences have tobe interpreted with caution
After taking the effects of sampling methodlocation and data treatment into account significantdifferences between bird families and age categorieswere found in both prevalence and importance for
adult and larval true flies adult and larval Lepidopteraand worms There were also significant differences inprevalence between bird families and age categoriesfor Gastropoda and in importance for bugs (Figs 4amp 5) These analyses suggested that adult true flieswere of greater importance to wader chicks than tothe other bird groups (although the difference withgrouse chicks was non-significant) whereas larvaltrue flies tended to be more important to both waderadults and chicks than to other groups By contrastadult and larval Lepidoptera were of most import-ance to both passerine adults and chicks but oflittle importance to the waders and grouse Wormswere of greatest importance to wader adultsAlthough frequently present in the diets of waderchicks they were never recorded as an lsquoimportantrsquotaxon and were absent from the diets of grousechicks Gastropods were most prevalent in diets ofadult passerines and waders whereas bugs occurredas an lsquoimportantrsquo taxon most frequently in the dietsof adult passerines and grouse chicks When inter-preting these results it must be remembered thatcertain of the bird groups were heavily influencedby data from one or two species for example foradult waders data from ten of the 40 studies werefrom Dunlin but only five were from CommonRedshank (Table 1) Thus certain comparisons mayreflect differences attributable to one or two species
χ22
χ22
Figure 4 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded in the diet Data are shown only for those invertebrate taxa for which statistically significant differences occurred between birdgroups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1729P lt 001 (wader chicks gt wader adults passerine adults lt passerine chicks wader chicks) Diptera larvae = 1700 P lt 001 (waderchicks gt passerine adults) Gastropoda = 2121 P lt 0001 (passerine chicks gt passerine adults) Lepidoptera adults = 2852P lt 0001 (passerine chicks gt wader adults) Lepidoptera larvae = 3052 P lt 0001 (passerine adults gt wader adults passerinechicks gt grouse chicks wader adults and chicks) Oligochaeta = 2677 P lt 0001 (wader adults gt passerine adults)
χ42
χ42
χ42 χ
42
χ42
χ42
8
G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
as opposed to being more widely representative ofthe relevant bird groups
DISCUSSION
The contribution of invertebrates to moorland bird diets
A wide range of invertebrates are consumed bymoorland birds but only a small number of thebroader level taxonomic groups are particularlyprevalent (ie arachnids beetles true flies bugsHymenoptera Lepidoptera and worms) With theexception of Hymenoptera and earthworms Coulson(1988) found that these taxa were among the mostabundant on moorland in northern England suggest-ing that birds are generally exploiting the most avail-able prey on moorlands The absence of Collembolaand enchytraeid worms from diets despite also beingnumerous is almost certainly due to their small sizeand hence lack of profitability as prey This diverserange of prey will partly reflect variation in diet betweenBSACs resulting from their different foragingstrategies (discussed below) However spatial andtemporal differences in prey availability within andbetween studies will also influence diet compositionas indicated by within-species dietary differencesassociated with different study locations and the fewstudies that have examined diet in relation to seasonaland between-habitat variation in invertebrateabundance (Picozzi amp Hepburn 1984 Starling-
Westerberg 2001 Whittingham et al 2001 Pearce-Higgins amp Yalden 2004)
Twelve studies presenting detailed compositionaldietary data for eight BSACs on British moorlands(Walton 1979 Picozzi amp Hepburn 1984 Garnettet al 1988 Cayford et al 1989 Whittingham 1996Starling-Westerberg 2001 Stephen 2001 Burfield2002 Pearce-Higgins amp Yalden 2003 2004) generallysupport the results of our wider analysis These studiesshowed that beetles formed the most importantitem for five of the BSACs considered comprisingover 15 of the diet of four cases True flies weremost important in four cases comprising over 15of the total diet in five The prevalence of adult trueflies in moorland bird diets especially among wadersand Red Grouse reflects in part the high abundanceof tipulids on some moorland areas following theirhighly synchronized emergences (Coulson 1962)Less important in this subset of more detailedmoorland studies were earthworms which were takenmainly by adult waders foraging on nearby improvedgrasslands but were also the most important item forRing Ouzel chicks As in our wider analysis arachnidsbugs Hymenoptera and Lepidoptera were alsoimportant and frequently occurring prey items inthese studies albeit less so than true flies and beetlesThus Hymenoptera were the most important preyin one case and accounted for more than 15 of thediet of three cases Arachnids bugs and Lepidopterawere never dominant possibly because of the relativelylow biomass and small prey size of some of these
Figure 5 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded as being important in the diet Importance in the diet is defined as for Figure 2 Data are shown only for those invertebrate taxafor which statistically significant differences occurred between bird groups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1389 P lt 001 (wader chicks gt wader adults passerine adults and chicks)Diptera larvae = 2130 P lt 0001 (wader adults gt passerine chicks) Hemiptera = 1178 P lt 005 (grouse chicks gt wader adultspasserine adults gt passerine chicks wader adults) Lepidoptera adults = 1714 P lt 001 (passerine chicks gt passerine adults)Lepidoptera larvae = 1254 P lt 005 (wader adults lt passerine adults and chicks) Oligochaeta = 3185 P lt 0001 (wader adultsgt passerine chicks)
χ42
χ42 χ
42
χ42
χ42 χ
42
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
8
G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
as opposed to being more widely representative ofthe relevant bird groups
DISCUSSION
The contribution of invertebrates to moorland bird diets
A wide range of invertebrates are consumed bymoorland birds but only a small number of thebroader level taxonomic groups are particularlyprevalent (ie arachnids beetles true flies bugsHymenoptera Lepidoptera and worms) With theexception of Hymenoptera and earthworms Coulson(1988) found that these taxa were among the mostabundant on moorland in northern England suggest-ing that birds are generally exploiting the most avail-able prey on moorlands The absence of Collembolaand enchytraeid worms from diets despite also beingnumerous is almost certainly due to their small sizeand hence lack of profitability as prey This diverserange of prey will partly reflect variation in diet betweenBSACs resulting from their different foragingstrategies (discussed below) However spatial andtemporal differences in prey availability within andbetween studies will also influence diet compositionas indicated by within-species dietary differencesassociated with different study locations and the fewstudies that have examined diet in relation to seasonaland between-habitat variation in invertebrateabundance (Picozzi amp Hepburn 1984 Starling-
Westerberg 2001 Whittingham et al 2001 Pearce-Higgins amp Yalden 2004)
Twelve studies presenting detailed compositionaldietary data for eight BSACs on British moorlands(Walton 1979 Picozzi amp Hepburn 1984 Garnettet al 1988 Cayford et al 1989 Whittingham 1996Starling-Westerberg 2001 Stephen 2001 Burfield2002 Pearce-Higgins amp Yalden 2003 2004) generallysupport the results of our wider analysis These studiesshowed that beetles formed the most importantitem for five of the BSACs considered comprisingover 15 of the diet of four cases True flies weremost important in four cases comprising over 15of the total diet in five The prevalence of adult trueflies in moorland bird diets especially among wadersand Red Grouse reflects in part the high abundanceof tipulids on some moorland areas following theirhighly synchronized emergences (Coulson 1962)Less important in this subset of more detailedmoorland studies were earthworms which were takenmainly by adult waders foraging on nearby improvedgrasslands but were also the most important item forRing Ouzel chicks As in our wider analysis arachnidsbugs Hymenoptera and Lepidoptera were alsoimportant and frequently occurring prey items inthese studies albeit less so than true flies and beetlesThus Hymenoptera were the most important preyin one case and accounted for more than 15 of thediet of three cases Arachnids bugs and Lepidopterawere never dominant possibly because of the relativelylow biomass and small prey size of some of these
Figure 5 The proportion of dietary studies of each of five moorland bird groups in which different broader level invertebrate taxa wererecorded as being important in the diet Importance in the diet is defined as for Figure 2 Data are shown only for those invertebrate taxafor which statistically significant differences occurred between bird groups Details of the statistics are as follows (with the significant pair-wise differences given in parentheses) Diptera adults = 1389 P lt 001 (wader chicks gt wader adults passerine adults and chicks)Diptera larvae = 2130 P lt 0001 (wader adults gt passerine chicks) Hemiptera = 1178 P lt 005 (grouse chicks gt wader adultspasserine adults gt passerine chicks wader adults) Lepidoptera adults = 1714 P lt 001 (passerine chicks gt passerine adults)Lepidoptera larvae = 1254 P lt 005 (wader adults lt passerine adults and chicks) Oligochaeta = 3185 P lt 0001 (wader adultsgt passerine chicks)
χ42
χ42 χ
42
χ42
χ42 χ
42
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
Moorland bird diets 9
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
taxa (Coulson 1988) Arachnid prevalence in thesestudies supports the trend in our wider analyses thatalthough widespread in the bird diets they oftenform a relatively small part of the total prey intake
Prey size is likely to be important in determiningthe extent to which birds take different invertebratesbut few studies have identified the main prey to thelevel of species or even genus and considerable sizevariation exists within insect families and otherinvertebrate orders Of the insect families identifiedas important in the current review several include alarge number of relatively large species (eg carabidsstaphylinids and adult and larval tipulids) butothers tend to comprise relatively small species suchas chironomids (eg Chinery 1986) The evidencesuggests that moorland birds often prey upon rela-tively large and presumably more profitable inver-tebrates especially in the case of adult waders foragingfor soil invertebrates on nearby grasslands Existingdata prevent any detailed assessment of the prey sizesthat are most often taken or selected although inEuropean Golden Plover the mean prey size takenby chicks foraging on blanket bog habitats increasedwith chick age from 312 mg (1- to 8-day-old chicks)to 834 mg (over 32 days) with this increase beingat least partly due to the greater exploitation oftipulid larvae by older chicks (Pearce-Higgins ampYalden 2004) Beintema et al (1991) found thatwader chicks (including Northern Lapwing andCommon Redshank) foraging on agricultural grass-lands tended to select invertebrate taxa in the largestsize class (4ndash20 mm) rather than taxa in smaller sizeclasses (0ndash4 and 2ndash6 mm) relative to measures oftheir abundance However in some instances rela-tively small prey can be important (eg Red Grousechicks and Molophilus ater ndash Savory 1977)
The comparison of diets between bird families andage classes highlights some broad differences relatedlargely to differing foraging strategies surface andsubsurface feeders vs foliage gleaners and aerialsalliers Waders comprising mainly the former group-ing tended to take higher proportions of interstitialinvertebrates such as true fly larvae and worms thandid most passerines which took more foliar inver-tebrates such as Lepidoptera and bugs Ring Ouzelswere an exception among the passerines in takingearthworms This is presumably associated with theirstrong preferences for foraging on short grass (Burfield2002)
Dietary differences between wader adults andchicks reflect the fact that during the prelaying andincubation periods at least adult Eurasian Curlew
and European Golden Plover forage off moorland onimproved pastures and other grasslands where theyexploit soil invertebrates while chicks generally remainon moorland where they exploit surface-active inver-tebrates (Whittingham et al 2000 2001 Robsonet al 2002 Pearce-Higgins amp Yalden 2003 2004)Differences in the diets of passerine adults and chicksare less readily explained given that adult passerinesprovision their chicks in the nest and would beexpected to have similar diets However variation innutritional value between invertebrate taxa could beimportant in this respect (eg Gosler 1993 Ramsayamp Houston 2003) or there may be seasonal differ-ences in the times at which adult and chick dietshave been studied
Invertebrate habitat associations
As indicated above a relatively small range ofinvertebrates tend to be important components ofmoorland bird diets although a wide range of inver-tebrates are taken reflecting the pattern found inlowland farmland birds (Wilson et al 1999) Thus itis likely that the abundance and distribution of somemoorland bird species are influenced by factorsaffecting habitat suitability for these key invertebratetaxa The diversity and abundance of many inver-tebrate taxa are heavily dependent on soil type andmoisture content For example beetles (which oftenhave interstitial larvae) display strong associationswith both characteristics (Butterfield amp Coulson1983 Dennis et al 1997 Gardner et al 1997) withspecifically the abundance of carabids being greateston sites with a mix of both wet and dry habitats(Butterfield amp Coulson 1983) Among the true fliestipulid larvae are most abundant on peaty soilsespecially where annual rainfall is high (Butterfield ampCoulson 1975 Coulson 1988) probably due to theirvulnerability to desiccation (Coulson 1962) Thereare no strong general associations with physical char-acteristics of soil among two of the three remainingtaxa (spiders and Lepidoptera) that formed a substan-tial proportion of bird diets (Downie et al 1995) butgiven that soil type can determine the distribution ofthe vegetation it can indirectly affect bug distributionand probably that of other less well-studied taxa aswell (Sanderson et al 1995 Hartley et al 2003)Despite being prevalent in the diets of some moorlandbirds worms are virtually absent from peat soilsHowever they are abundant on mineral soilsespecially pastures near moorland edges wherethe adults of some species forage regularly (eg
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
10 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Whittingham et al 2000 Burfield 2002 Pearce-Higgins amp Yalden 2003)
Moorland vegetation composition is also an impor-tant determinant of the invertebrate community (egButterfield amp Coulson 1983 Coulson amp Butterfield1986 Gardner 1991 Gardner et al 1997) On sedge-dominated blanket bogs true flies especially tipulidstend to predominate although this may also reflectthe wet peaty substrate Conversely drier heathcommunities dominated by dwarf shrubs such asHeather Calluna vulgaris are characterized by a highLepidoptera biomass (Coulson 1988) and cansupport more beetles than graminoid-dominatedhabitats (Gardner et al 1997) The differing habitatassociations of invertebrates mean that sites with amosaic of habitat types are liable to support a greaterdiversity and abundance of invertebrates thanhomogeneous sites (Coulson amp Butterfield 1985Usher amp Thompson 1993) potentially resulting in agreater diversity and abundance of moorland birds
The effects of moorland management
Vegetation and management are important determi-nants of bird abundance (eg Tharme et al 2001Calladine et al 2002 Pearce-Higgins amp Grant in press)Changes in vegetation or management affectingvegetation may affect bird populations throughaltering invertebrate abundance Large-scale changesin moorland habitats will alter invertebrate commun-ity composition (Coulson 1988 Usher amp Thompson1993) while smaller-scale changes in vegetationcomposition and structure can have major impactson both species richness and abundance of themajor invertebrate groups (Southwood et al 1979Gimingham 1985) Trends in moorland managementsince the middle of the last century will have changedthe soil conditions vegetation structure and compo-sition on moorlands and hence affected invertebrateabundance and community composition Notabletrends are(1) Increases in sheep and Red Deer numbersaffecting moorland vegetation across many uplandareas particularly in causing the expansion of grass-land into heather moorland (Mackey et al 1998Fuller amp Gough 1999)(2) Declines in grouse moor management (egRobertson et al 2001) may have reduced the extentand frequency of muirburn so reducing spatial varia-tion in vegetation composition and height (althoughspecific evidence for declines in muirburn extent islacking ndash Hester amp Sydes 1992)
(3) The widespread construction of drainage ditches(grips) between the 1960s and 1990s changed soilconditions and vegetation composition (Coulsonet al 1990)
Where relationships between moorland man-agement vegetation and invertebrates have beeninvestigated by experimental manipulation focus hasbeen on the impacts of relatively short-term changesin vegetation structure mediated through differentialgrazing regimes (eg McFerran et al 1994a 1994bDennis et al 1997 1998 2001 2005) or muirburnmanagement (Gardner 1991 Haysom amp Coulson1998) Few experimental studies have investigatedthe impacts of different management regimes overlonger periods (decades) replicated across severaldifferent sites In particular relatively few experimentalstudies have focused on the effects of moorland man-agement on the true flies especially tipulids whichare a key component in the diet of many bird speciesHowever their general ecology has been well studiedallowing some management impacts to be inferred(Coulson amp Butterfield 1985 Coulson et al 1990)
The effects of mixed grazing regimes (sheep andcattle) on invertebrates appear to vary Sheep andcattle grazing reduce beetle abundance throughtrampling and dunging by cattle while increasedgrazing can reduce overall arthropod abundance onNardus-dominated grasslands by damaging thetussocks where invertebrates shelter (Dennis et al1998) Increased dunging may however also benefitinvertebrates by providing food for larvae (egMcCracken amp Foster 1993) Overall invertebratebiomass may increase at lower grazing intensitieswith the biomass of surface-dwelling invertebrates(as sampled by suction sampling) doubling after a66 decrease in stocking (Dennis et al 2005) How-ever reduced grazing did not benefit all invertebratesin that study and larval tipulid biomass decreasedunder the reduced stocking levels There is alsoevidence of an interaction between cattle grazing andHeather Beetle Lochmaea suturalis on dry heaths inthe Netherlands (Bokdam 2001) Patches of defoli-ated heather are vulnerable to invasion by grasses(especially Molinia on wet heaths and Deschampsiaon dry heaths) although cattle grazing of the grassleads to more rapid heather recovery
Among the invertebrates that are key componentsof moorland bird diets changes in vegetation structurecan produce rapid responses in the abundance ofspiders which favour tall complex swards (Coulsonamp Butterfield 1986 McFerran et al 1994b Denniset al 1998 2004 Bell et al 2001) and some bugs
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
Moorland bird diets 11
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
(Gimingham 1985) which increased after a reductionin grazing pressure (Dennis et al 2005) Similarlybeetles especially staphylinids increased with adecrease in grazing pressure (Dennis et al 2005)Hymenoptera and Lepidoptera larvae both increasein abundance with increased vegetation height orstructural complexity (Haysom amp Coulson 1998) Theshort open vegetation patches produced by muirburnbenefit mobile predatory and scavenging species andpioneer species that colonize the new growth ofvegetation (Gardner amp Usher 1989 Gardner 1991Bell et al 2001) Moorland drainage schemes wereformerly used to improve heather growth and henceprovide better habitats for grouse and sheepalthough the wet areas affected by such drainage mayprovide important food for grouse chicks (Coulsonamp Butterfield 1985) The impacts of drainage appearto be relatively minor in areas with heavy rainfallbut on drier areas it may remove small pockets ofwet peat reducing the overall invertebrate diversityand perhaps more importantly in the context of birddiets depressing the spring emergence of insectsespecially tipulids (Coulson amp Butterfield 1985Coulson et al 1990)
Overall these studies suggest that managementregimes that create habitats characterized by a mosaicof vegetation communities and structures are likelyto support the greatest invertebrate diversity andabundance (Coulson amp Butterfield 1985 Usheramp Thompson 1993) potentially benefiting themoorland birds Thus the generally positive effects ofincreasing vegetation structure on overall arthropodabundance are likely to benefit foliage gleaners andaerial salliers such as Whinchat and Stonechat thatassociate with taller denser vegetation (eg Allen1995 Pearce-Higgins amp Grant in press) Converselyshort open vegetation can be associated with a greaterabundance of important subsurface invertebrate preyand can also increase the availability of surfaceinvertebrates so benefiting such birds as EuropeanGolden Plover and Ring Ouzel which tend to requireshort open vegetation (Burfield 2002 Pearce-Higginsamp Yalden 2003)
In conclusion we have summarized the importanceof different invertebrate taxa in the diets of moor-land birds highlighting the most important taxa andbased upon current knowledge summarized howmoorland management may affect their abundanceWe have also identified gaps in current knowledgeFor example diets remain poorly described for themajority of moorland bird species Furthermore tomaximize the potential use of such data we recom-
mend that future studies quantify diet in terms ofnumerical or preferably biomass composition to asfine a taxonomic level as possible (at least family)assess prey-size selection and target bird speciesfor which dietary data are scarce To understand theeffects of management on invertebrate prey experi-mental studies are required particularly on theeffects of burning or cutting moorland vegetationand on grip blocking Fundamentally investigationsof relationships between the diversity and abundanceof invertebrates and the abundance of moorlandbirds are required to determine the extent to whichhabitat change affects these bird populations viaeffects on their invertebrate prey
ENDNOTE
Analyses were performed by first fitting samplingmethod (ie stomach or crop faecal neck collarfeeding observation) location (ie BampI moorlandBampI non-moorland non-BampI) and for analyses oflsquoimportancersquo data treatment (ie biomass composi-tion numerical composition percentage occurrence)A step-down procedure was used deleting the factorwith the lowest significance level at each stage untilonly the significant (P lt 005) factors remained Effectsof bird families and age were assessed at this stage bytesting whether the addition of this factor produceda further significant change in model devianceStatistical significance was determined by treatingthe change in residual deviance associated with eachterm as χ2 with the appropriate degrees of freedom(Crawley 1993 SAS Institute 2001) Changing theorder in which the levels of the factor (bird familyand age) were entered allowed the standard errorsaround each estimated mean to be compared andhence which categories differed significantly fromthe others (Crawley 1993 SAS Institute 2001)
This work was part of research funded by DEFRA EnglishNature and the Countryside Council for Wales underDEFRA project no BD1228 Thanks to Lynn GiddingsDr Jevgeni Shergalin and Dr David Carroll for helping sourceand provide journals and theses and Darren Evans for pro-viding unpublished data We are grateful to Jeremy Wilsonand Mark Whittingham and two anonymous referees forcomments on the manuscript
REFERENCES
Allen DS 1995 Factors influencing Habitat Associations ofUpland Passerines particularly the Whinchat (Saxicola rubetra)the Importance of Scale DPhil thesis University of York
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
12 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Andreev AV 1988 Ecological energetics of Palearctic Tetrao-nidae in relation to chemical composition and digestibility oftheir winter diets Can J Zool 66 1382ndash1388
Andrusenko NN 1980 K pitaniyu nekotorykh maibolee mass-ovykh vidov kulikov na ozere Tengiz Novoe v izucheniibiologii i rasprostranenii kulikov Moscow Nauka
Anon 1983 Osnovnye osobennosti pishchevykh svyazeikulikov v basseine rShchuchrsquoei Moscow Nauka
Artemrsquoyev YT amp Popov VA 1978 Ptitsy Volzhsko-KamskogoKraya Moscow Nauka
Baines D 1990 The roles of predation food and agriculturalpractice in determining the breeding success of the lapwingVanellus vanellus on upland grasslands J Anim Ecol 59915ndash929
Baker MC 1977 Shorebird food habits in the easternCanadian arctic Condor 79 56ndash62
Bardgett RD Marsden JH Howard DC amp Hossell JE1995 The extent and condition of heather in moorland andthe potential impact of climate change In Thompson DBAHester AJ amp Usher MB (eds) Heaths and MoorlandCultural Landscapes 43ndash50 Edinburgh HMSO
Beintema AJ Thissen JB Tensen D amp Visser GH1991 Feeding ecology of Charadriiform chicks in agriculturalgrassland Ardea 79 31ndash44
Bell JR Wheater CP amp Cullen WR 2001 The implicationsof grassland and heathland management for the conservation ofspider communities a review J Zool Lond 255 377ndash387
Berry PM Vanhinsberg D Viles HA Harrison PAPearson RG Fuller RJ Butt N amp Miller F 2001Impacts on terrestrial environments In Harrison PA BerryPM amp Dawson TP (eds) Climate Change and Nature Con-servation in Britain and Ireland Modelling Natural ResourceResponses to Climate Change the MONARCH Project 43ndash149 UKCIP Technical Report Oxford UKCIP
Bokdam J 2001 Effects of browsing and grazing on cyclicsuccession in nutrient-limited ecosystems J Veg Sci 12875ndash886
Borchtchevski VG 2000 Diet of the Black Grouse Tetrao tetrixin Vodlozerski National Park North-western Russia GameWildlife Sci 17 29ndash42
Brooke M 1981 How an adult wheatear Oenanthe oenanthe usesits territory when feeding nestlings J Anim Ecol 50 683ndash696
Brown AF amp Stillman RA 1993 Birdndashhabitat associations inthe Eastern Highlands of Scotland J Appl Ecol 20 31ndash42
Burfield IJ 2002 The Breeding Ecology and Conservationof the Ring Ouzel Turdus torquatus in Britain PhD thesisUniversity of Cambridge
Burges S 1994 Segregation of diet in water pipit Anthusspinoletta and Meadow Pipit Anthus pratensis nestlings in anarea damaged by air pollution Folia Zool 43 43ndash48
Burges S amp Weildinger K 2000 Estimation of calcium intakeby Meadow Pipit nestlings in an acidified area J Ornithol 31426ndash429
Butterfield J amp Coulson JC 1983 The Carabid communitieson peat and upland grasslands in northern England HolarctEcol 6 163ndash174
Byrkjedal I 1980 Summer food of the Golden Plover Pluvialisapricaria at Hardangervidda southern Norway Holarct Ecol3 40ndash58
Calladine J Baines D amp Warren P 2002 Effects of reducedgrazing on population density and breeding success of blackgrouse in northern England J Appl Ecol 39 772ndash780
Cayford JT Tyler G amp MacIntosh-Williams L 1989 TheEcology and Management of Black Grouse in Conifer Forestsin Wales Sandy RSPB
Chinery M 1986 A Field Guide to the Insects of Britain andNorthern Europe 2nd edn London Collins
Coulson JC 1962 The biology of Tipula subnodicornisZetterstedt with comparative observations on Tipula paludosaMeigen J Anim Ecol 31 1ndash21
Coulson JC 1988 The structure and importance of inverte-brate communities on peatlands and moorlands and effectsof environmental and management changes In Usher MBamp Thompson DBA (eds) Ecological Change in the Uplands365ndash380 Oxford Blackwell Scientific Publications
Coulson JC amp Butterfield JEL 1985 The invertebratecommunities of peat and upland grasslands in the north ofEngland and some conservation implications Biol Conserv34 197ndash225
Coulson JC amp Butterfield J 1986 The spider communitieson peat and upland grasslands in northern England HolarctEcol 9 229ndash239
Coulson JC Butterfield JEL amp Henderson E 1990 Theeffect of open drainage ditches on the plant and invertebratecommunities of moorland and on the decomposition of peatJ Appl Ecol 27 549ndash561
Coulson JC amp Whittaker JB 1978 Ecology of moorlandanimals In Heal OW amp Perkins DF (eds) Production Eco-logy of British Moors and Montane Grasslands 52ndash93 BerlinSpringerndashVerlag
Cramp S (ed) 1988 The Birds of the Western Palearctic Vol 5Oxford Oxford University Press
Cramp S amp Simmons KEL 1980 The Birds of the WesternPalearctic Vol 2 Oxford Oxford University Press
Cramp S amp Simmons KEL 1983 The Birds of the WesternPalearctic Vol 3 Oxford Oxford University Press
Crawley MJ 1993 GLIM for Ecologists Oxford BlackwellScientific Publications
Cummins S amp OrsquoHalloran J 2002 An assessment of the dietof nestling Stonechats Saxicola torquata using compositionalanalysis Bird Study 49 139ndash145
Danilov NN Ryzhanovskiy VN amp Ryabitsev VK 1984Ptitsy Yamala Moscow Nauka
Dennis P Elston D Evans DM Evans SA Gordon IJGrant M Kunaver A Marquiss M Mayes RMcCracken DI Pakeman R Pearce-Higgins JRedpath SM Skartveit J Stephen L Benton T ampBryant D 2005 Effects of Grazing Management on UplandBird Populations Disentangling Habitat Structure andArthropod Food Supply at Appropriate Spatial Scales (GRUB)Unpublished report to The Scottish Executive Environmentand Rural Affairs Department Edinburgh
Dennis P Young MR amp Bentley C 2001 The effects ofvaried grazing management on epigeal spiders harvestmenand pseudoscorpions of Nardus stricta grassland in uplandScotland Agric Ecosyst Environ 86 39ndash57
Dennis P Young MR amp Gordon IJ 1998 Distribution andabundance of small insects and arachnids in relation tostructural heterogeneity of grazed indigenous grasslandsEcol Entomol 23 253ndash264
Dennis P Young MR Howard CL amp Gordon IJ 1997
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
Moorland bird diets 13
copy 2006 The AuthorsJournal compilation copy 2006 British Ornithologistsrsquo Union
The response of epigeal beetles (Col Carabidae Staphylinidae)to varied grazing regimes on upland Nardus stricta grasslandsJ Appl Ecol 34 433ndash443
Donald PF Muirhead LB Buckingham DL Evans ADKirby WB amp Gruar DJ 2001 Body condition growth ratesand diet of Sky Lark Alauda arvensis nestlings on lowlandfarmland Ibis 143 658ndash669
Dorzhiev TZ amp Khertuev VN 1992 Ekologiya kamenok vZabaikalie Ulan-Ude Buryatian Scientific Centre of SiberianBranch of Russia Ac of Sc 148
Downie IS Butterfield JEL amp Coulson JC 1995 Habitatpreferences of sub-montane spiders in northern EnglandEcography 18 51ndash61
Filimonova NP 1972 Materialy o pitanii tetereva i seroikuropatki v Kustanaiskoi oblasti Novosibirsk MoscowNauka
Fuller RJ amp Gough SJ 1999 Changes in sheep numbers inBritain implications for bird populations Biol Conserv 9173ndash89
Galbraith H 1989 The diet of Lapwing Vanellus vanelluschicks on Scottish farmland Ibis 131 80ndash84
Gardner SM 1991 Ground Beetle Coleoptera Carabidaecommunities on upland heath and their association withheathland flora J Biogeogr 18 281ndash289
Gardner SM Hartley SE Davies A amp Palmer SCF1997 Carabid communities on heather moorlands in north-east Scotland the consequences of grazing pressure forcommunity diversity Biol Conserv 81 275ndash286
Gardner SM amp Usher MB 1989 Insect abundance onburned and cut upland Calluna heath Entomologist 108145ndash200
Garnett MC MacFadzean S amp Townsend M 1988 TheForaging Ecology of the Whinchat Saxicola rubetra in WalesUnpublished RSPB report Sandy RSPB
Gibbons DW Reid JB amp Chapman RA 1993 The NewAtlas of Breeding Birds in Britain and Ireland 1988ndash1991London T amp AD Poyser
Gosler A 1993 The Great Tit London HamlynGreen RE 1986 Breeding Waders of the Somerset Moors
Factors Affecting Their Distribution and Breeding SuccessUnpublished report to NCC Peterborough NCC
Green RE Hirons GJM amp Cresswell BH 1990 Foraginghabitats of female common snipe Gallinago gallinago duringthe incubation period J Appl Ecol 27 325ndash335
Gregory RD Wilkinson NI Noble DG Robinson JABrown AF Hughes J Proctor D Gibbons DW ampGalbraith CA 2002 The population status of birds in theUnited Kingdom Channel Islands and Isle of Man an analysisof conservation concern 2002ndash07 Br Birds 95 410ndash448
Greig-Smith PW amp Quicke DLJ 1983 The diet of nestingStonechats Bird Study 30 47ndash50
Gubin BM amp Kovshar AF 1985 K biologii obyknovennoikamenki v vysokogorrsquoie Tien-Shanya Ornitologiya 20 53ndash59
Hagvar S amp Ostbye E 1976 Food habits of the Meadow PipitAnthus pratensis in alpine habitats at Hardangervidda southNorway Norw J Zool 24 53ndash67
Hartley SE Garner SM amp Mitchell RJ 2003 Indirecteffects of grazing and nutrient addition on the hemipterancommunity of heather moorlands J Appl Ecol 40 793ndash803
Haworth PF amp Thompson DBA 1990 Factors associatedwith the breeding distribution of upland birds in the SouthPennines England J Appl Ecol 27 562ndash577
Haysom KA amp Coulson JC 1998 The Lepidoptera faunaassociated with Calluna vulgaris effects of plant architectureon abundance and diversity Ecol Entomol 23 377ndash385
Hester AJ amp Sydes C 1992 Changes in burning of Scottishheather moorland since the 1940s from aerial photographsBiol Conserv 60 25ndash30
Holmes RT 1966 Feeding ecology of the red backed sandpiperCalidris alpina in arctic Alaska Ecology 47 32ndash45
Holmes RT 1970 Differences in population density territorialityand food supply of dunlin on arctic and subarctic tundra InWatson A (ed) Animal Populations in Relation to Their FoodResources 303ndash319 Symposium of the British EcologicalSociety Oxford Blackwell
Jenny M 1990 Nahrungsokologie der Feldlerche Alaudaarvensis in einer intensiv genutzten Agrarlandschaft desschweizerischen Mittellandes Der Ornithologische Beobachter87 31ndash53
Kastdalen L amp Wegge P 1985 Animal food in Capercaillie andBlack Grouse chicks in south east Norway ndash a preliminaryreport In Lovel TWI amp Hudson PJ (eds) Proceedings ofthe Third International Grouse Symposium 499ndash513 Read-ing UK World Pheasant Association
Kondratrsquoyev AY amp Kondratrsquoyeva LF 1988 Ispolrsquozovanierezulrsquotatov gelmintologicheskikh vskrytiy dlya analiza pish-chevykh svyazei kulikov Moscow Nauka
Kostin YV 1983 Ptitsy Kryma Moscow NaukaKumari A 1958 Rabakurvitsaliste toitumisest ndash Rmt-s E
Kumari toim Ornitoloogiline kogumik 1 Eesti NSV TeadusteAkadeemia Toimetus-Kirjastusnotildeukogu Tartu Lk 195ndash215
von Labhardt A 1988 Zur nahrung des BraunkehlchensSaxicola rubetra wahrend der Jungenaufzucht Beih VeroffNaturschutz Landschaftspflege Bad-Wurtt 51 179ndash185
Linsley MD 1999 The Effects of Intensive Agriculture on theBreeding Ecology of the Lapwing Vanellus vanellus PhDthesis Liverpool John Moores University
Mackey EC Shewry MC amp Tudor GJ 1998 Land CoverChange Scotland from the 1940s to the 1980s EdinburghHMSO
Malrsquochevskiy AS amp Pukinskiy YB 1983 Ptitsy Leningrad-skoi oblasti i sopredelrsquonykh territoriy 2 Leningrad LeningradUniversity Press
McCracken DI amp Foster GN 1993 Surface-active inverte-brate communities and the availability of potential food for theChough Pyrrhocorax pyrrhocorax L on pastures in north-west Islay Pedobiologia 37 141ndash158
McFerran DM Montgomery WI amp McAdam JH 1994aEffects of grazing intensity on heathland vegetation andground beetle assemblages of the uplands of County Antrimnortheast Ireland Biol Environ 94B 41ndash52
McFerran DM Montgomery WI amp McAdam JH 1994bThe impact of grazing on communities of ground-dwellingspiders Araneae in upland vegetation types Biol Environ94B 119ndash126
Moreby SJ amp Stoate C 2000 A quantitative comparison ofneck collar and faecal analysis to determine passerinenestling diet Bird Study 47 320ndash331
Nechaev AV 1991 Ptitsy ostrova Sakhalin Vladivostok Vladi-vostok Institute of Biology and Pedology
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006
14 G M Buchanan et al
copy 2006 The Authors Journal compilation copy 2006 British Ornithologistsrsquo Union
Newton I 1998 Population Limitation in Birds London AcademicPress
Niewold FJJ 1990 The decline of Black Grouse in the Neth-erlands In Lumeij JT amp Hoogeveen YR (eds) The Futureof Wild Galliformes in the Netherlands 71ndash81 AmersfoortNetherlands Organisatiecommissie Nederlandse WildeHoenders
OrsquoConnell M Thomas CJ Twiss SD Downie ISEvans PR amp Whitfield DP 1996 Functional Ecology ofPeatland Animals in the Flow Country of Northern ScotlandI Habitat Requirements of Breeding Waders CharadriiEdinburgh Research and Advisory services DirectorateReport SNH
Park KJ Robertson PA Campbell ST Foster RRussell ZM Newborn D amp Hudson PJ 2001 The roleof invertebrates in the diet growth and survival of RedGrouse Lagopus lagopus scoticus chicks J Zool Lond254 137ndash145
Pearce-Higgins JW 1999 The Ecology of Golden PloversPluvialis apricaria in the Peak District PhD thesis Universityof Manchester
Pearce-Higgins JW amp Grant MC 2006 Relationshipsbetween bird abundance and the composition and structureof moorland vegetation Bird Study 53 112ndash125
Pearce-Higgins JW amp Yalden DW 2003 Variation in the useof pasture by breeding European Golden Plovers Pluvialisapricaria in relation to prey availability Ibis 145 365ndash381
Pearce-Higgins JW amp Yalden DW 2004 Habitat selectiondiet arthropod availability and growth of a moorland waderthe ecology of European Golden Plover Pluvialis apricariachicks Ibis 146 335ndash346
Peresadrsquoko LV amp Koshelev AI 1980 K pitaniyu kulikov nayuge Zapadnoi Sibiri Novoe v izuchenii biologii i raspros-tranenii kulikov Moscow Nauka
Picozzi N amp Hepburn LV 1984 A study of Black Grouse innorth east Scotland Proc Int Grouse Symp 3 462ndash480
Ramsay SL amp Houston DC 2003 Amino acid compositionof some woodland arthropods and its implications for breed-ing tits and other passerines Ibis 145 227ndash232
Ratcliffe DA 1976 Observations on the breeding of GoldenPlover in Great Britain Bird Study 23 63ndash116
Ratcliffe D 1990 Bird Life of Mountains and Uplands Cam-bridge Cambridge University Press
Robertson PA Park KJ amp Barton AF 2001 Loss ofheather Calluna vulgaris moorland in the Scottish uplandsthe role of Red Grouse Lagopus lagopus scoticus manage-ment Wildlife Biol 7 11ndash16
Robson G 1998 The Breeding Ecology of Curlew Numeniusarquata on North Pennine Moorland PhD thesis Universityof Sunderland
Robson G Percival SM amp Brown AF 2002 The use ofmarginal farmland by Curlew Numenius arquata breeding onupland moors Aspects Appl Biol 67 75ndash84
Sanderson RA Rushton SP Cherrill AJ amp Byrne JP1995 Soil vegetation and space ndash an analysis of their effects
on the invertebrate communities of a moorland in northeastEngland J Appl Ecol 32 506ndash518
SAS Institute 2001 SASSTAT Userrsquos Guide Version 802Cary NC SAS Institute
Savory CJ 1977 The food of Red Grouse chicks Lagopuslagopus scoticus Ibis 119 1ndash9
Siepel H 1990 The influence of management on food size inthe menu of insectivorous animals Proc Exp Appl Entomol1 69ndash74
Sim IMW Gregory RD Hancock MH amp Brown AF2005 Recent changes in the abundance of British uplandbreeding birds Bird Study 52 261ndash275
Southwood TRE Brown VK amp Reader PM 1979 Therelationships of plant and insect diversities in successionBiol J Linn Soc 12 327ndash348
Starling-Westerberg A 2001 The habitat use and diet of BlackGrouse Tetrao tetrix in the Pennine hills of northern EnglandBird Study 48 76ndash89
Stephen L 2001 An Investigation into the Dietary Require-ments of Curlew Numenius arquata Chicks BSc thesisUniversity of Edinburgh
Task Force for the Hills 2001 Report of Findings to the Ministerfor Agriculture Fisheries and Food London HMSO
Tharme AP Green RE Baines D Bainbridge IP ampOrsquoBrien M 2001 The effect of management for Red Grouseshooting on the population density of breeding birds onheather dominated moorland J Appl Ecol 38 439ndash457
Thompson DBA MacDonald AJ Marsden JH ampGalbraith CA 1995 Upland heather moorland in GreatBritain a review of international importance vegetationchange and some objectives for nature conservation BiolConserv 71 163ndash178
Tyler SJ amp Green M 1994 The status and breeding ecologyof Ring Ouzels Turdus torquatus in Wales with reference tosoil acidity Welsh Bird Report 7 78ndash89
Usher MB amp Thompson DBA 1993 Variation in the uplandheathlands of Great Britain ndash conservation importance BiolConserv 66 69ndash81
Walton KC 1979 Diet of Meadow Pipits Anthus pratensis onmountain grassland in Snowdonia Ibis 121 325ndash334
Whittingham MJ 1996 Habitat Requirements of BreedingGolden Plover Pluvialis apricaria PhD thesis University ofSunderland
Whittingham MJ Percival SM amp Brown AF 2000 Timebudgets and foraging of breeding golden plover Pluvialisapricaria J Appl Ecol 37 632ndash646
Whittingham MJ Percival SM amp Brown AF 2001 Habitatselection by Golden Plover Pluvialis apricaria chicks BasicApp Ecol 2 177ndash191
Wilson JD Morris AJ Arroyo BE Clark SC ampBradbury RB 1999 A review of the abundance anddiversity of invertebrate and plant food of granivorous birdsin northern Europe in relation to agricultural change AgrEcosyst Environ 75 13ndash30
Zinovrsquoyev VI 1980 Ptitsy lesnoi zony Evropeiskoi chasti SSSRrzhankoobraznye Kalinin Kalinin State University Press
Received 8 June 2005 revision accepted 28 March 2006