CBE 2018 - A02 1 The ecological, social and cultural origins of primate intellect The evolution of intellect Hypotheses on the evolution of intellgence in human and non-human primates The “Technological” Hypothesis Oakley (Hominins only...) The Foraging Hypothesis Milton, Clutton-Brock The “Social Brain” Hypotheses (Dunbar et al) The “Machiavellian Intelligence” Hypothesis Byrne & Whiten, Cheney & Seyfarth The “Cultural Intelligence” Hypotheses van Schaik, Tomasello 1 2
Transcript
CBE 2018 - A02
1
The ecological, social and cultural origins of primate intellect
The evolution of intellectHypotheses on the evolution of intellgencein human and non-human primates
The “Technological” Hypothesis Oakley (Hominins only...)
The Foraging HypothesisMilton, Clutton-Brock
The “Social Brain” Hypotheses (Dunbar et al)
The “Machiavellian Intelligence” HypothesisByrne & Whiten, Cheney & Seyfarth
The “Cultural Intelligence” Hypothesesvan Schaik, Tomasello
1
2
CBE 2018 - A02
2
The “Ecological” Hypotheses
The “Technological” HypothesisKenneth Oakley (1959): “Man, the toolmaker”
Jerison (1973):predators x prey
Ungulates’ and Carnivores’ brain sizes covaried through Paleogene / Neogene
3
4
CBE 2018 - A02
3
2001: A Space Odyssey (Stanley Kubrick, 1968)
Tools and hominization
Forest X savannaBipedalismFreeing of handsOpposition of thumbsEncefalization...
5
6
CBE 2018 - A02
4
“A mere 12,000 years separate the first bow and arrow from the International Space Station” (Ambrose 2001)
Tool use in nonhuman primates
7
8
CBE 2018 - A02
5
Tool use in non-primate vertebratesBottlenose dolphins (Krützen et al 2005)
Tool use in non-primate vertebrates
Egiptian vulture cracking ostrich eggOtter cracking oyster with stone
Folivores x frugivoresCorrelation between home ranges and neocortex size (...?)
The Foraging HypothesisKatherine Milton (1993)
15
16
CBE 2018 - A02
9
R. Wrangham (2010): cooking
Raw x cooked food:Digestive tract ↓ x brains ↑More free timePair bonding, sexual division of labor
Encefalizationandsocial group size(Dunbar 1992)
Neocortical Ratio XHome range residue
disconting the effect of bodily mass
Neocortical Ratio XAverage group size
17
18
CBE 2018 - A02
10
The “Social Brain” Hypotheses
Primatology:Long-term field studies, research with captive groups:
Social complexity
AlliancesWarReconciliationStatus seekingDeception
19
20
CBE 2018 - A02
11
Frans de Waal (1982, 1989)
Primate sociality
de Waal:
primates are “prisioners” of societyconflict and reconciliation
21
22
CBE 2018 - A02
12
The “Machiavellian Intelligence” Hypothesis
Byrne & Whiten 1989 Whiten & Byrne 1997
Selection pressures from complex social life:All primates (except prosimians) are socialexception of the exception: social lemuroids from Madagascar (Jolly 1966)
Harlow: social deprivation (Rhesus) → Bowlby: Attachment Theory (children)
Social Isolation inRhesus monkeys
Harlow, H. F. (1970)
25
26
CBE 2018 - A02
14
Selection pressures from complex social life:All primates (except prosimians) are socialexception of the exception: social lemuroids from Madagascar (Jolly 1966)
Effects of social deprivation (Harlow)
Social complexity:Hierarchies, alliances, reconciliation, grooming, food sharing...
“Neocortical suppression of autonomic functions”
(Chance & Mead 1953)
“Tripartite relationships” in baboons (Kummer 1967)
Children understand social causality before physical causality(Laurendeau & Pinard 1962)
29
30
CBE 2018 - A02
16
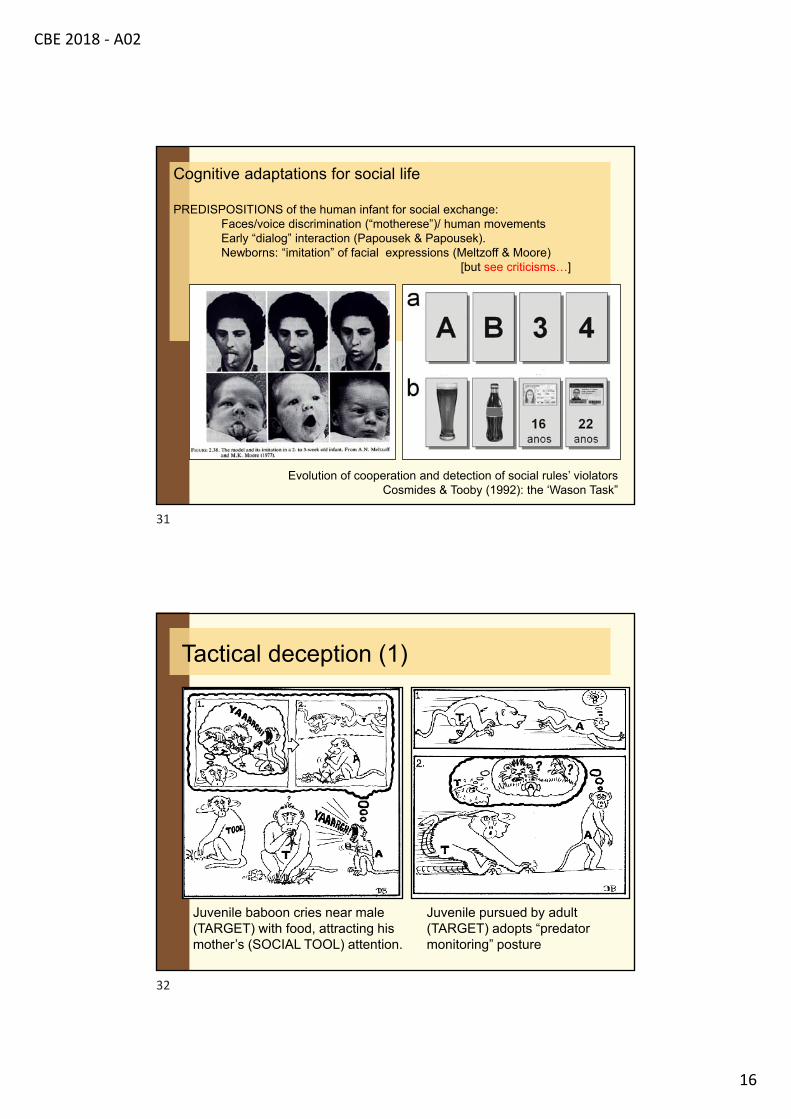
Cognitive adaptations for social life
PREDISPOSITIONS of the human infant for social exchange:Faces/voice discrimination (“motherese”)/ human movementsEarly “dialog” interaction (Papousek & Papousek).Newborns: “imitation” of facial expressions (Meltzoff & Moore)
[but see criticisms…]
Evolution of cooperation and detection of social rules’ violatorsCosmides & Tooby (1992): the ‘Wason Task”
Tactical deception (1)
Juvenile baboon cries near male (TARGET) with food, attracting his mother’s (SOCIAL TOOL) attention.
Juvenile pursued by adult (TARGET) adopts “predator monitoring” posture
31
32
CBE 2018 - A02
17
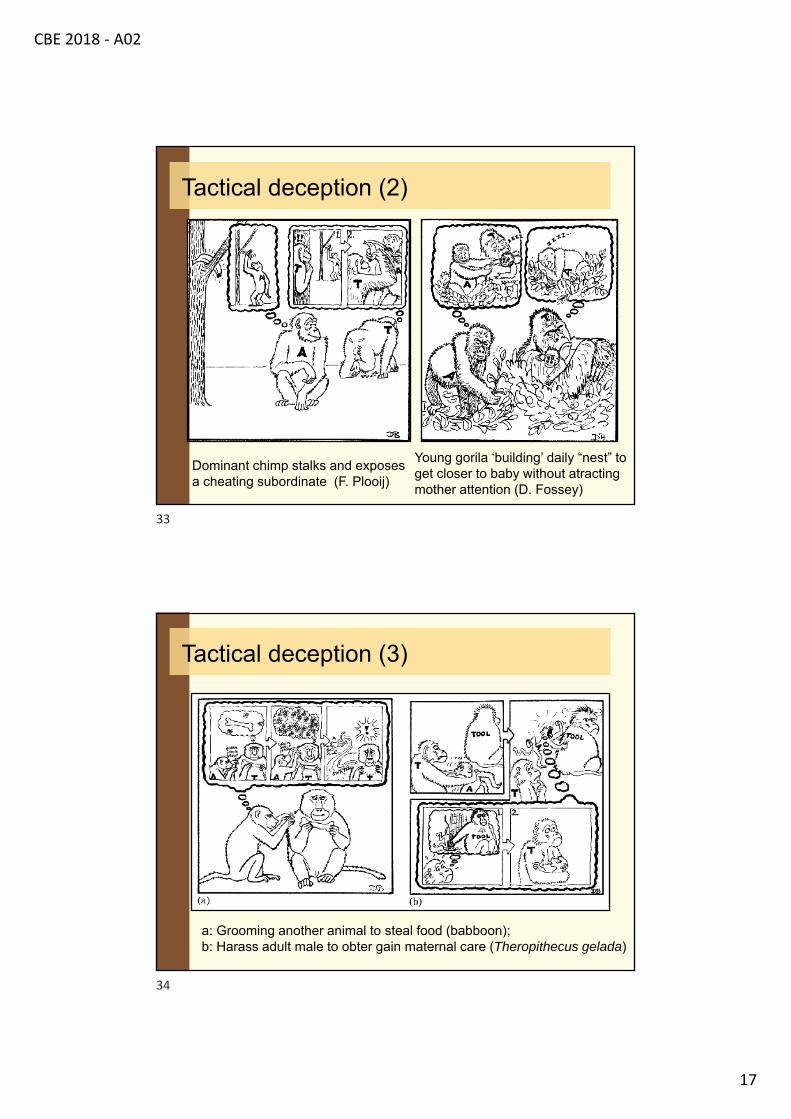
Dominant chimp stalks and exposes a cheating subordinate (F. Plooij)
Young gorila ‘building’ daily “nest” to get closer to baby without atracting mother attention (D. Fossey)
Tactical deception (2)
a: Grooming another animal to steal food (babboon);b: Harass adult male to obter gain maternal care (Theropithecus gelada)
Tactical deception (3)
33
34
CBE 2018 - A02
18
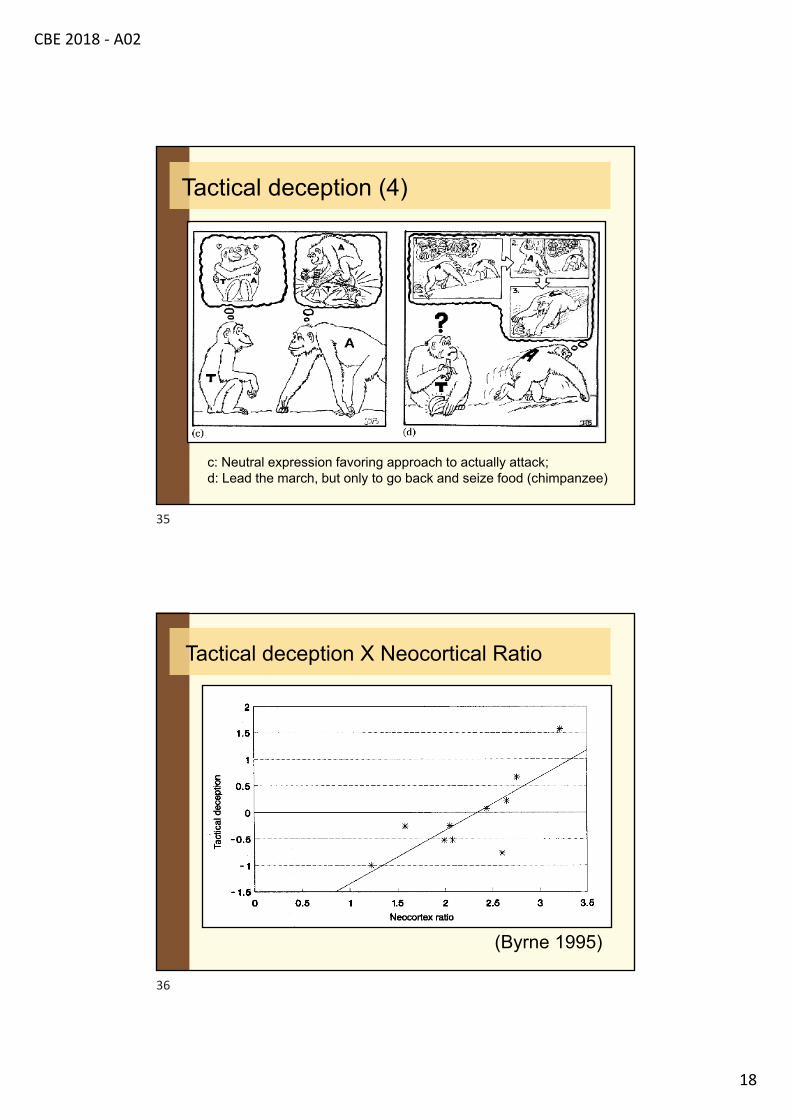
c: Neutral expression favoring approach to actually attack;d: Lead the march, but only to go back and seize food (chimpanzee)
Tactical deception (4)
Tactical deception X Neocortical Ratio
(Byrne 1995)
35
36
CBE 2018 - A02
19
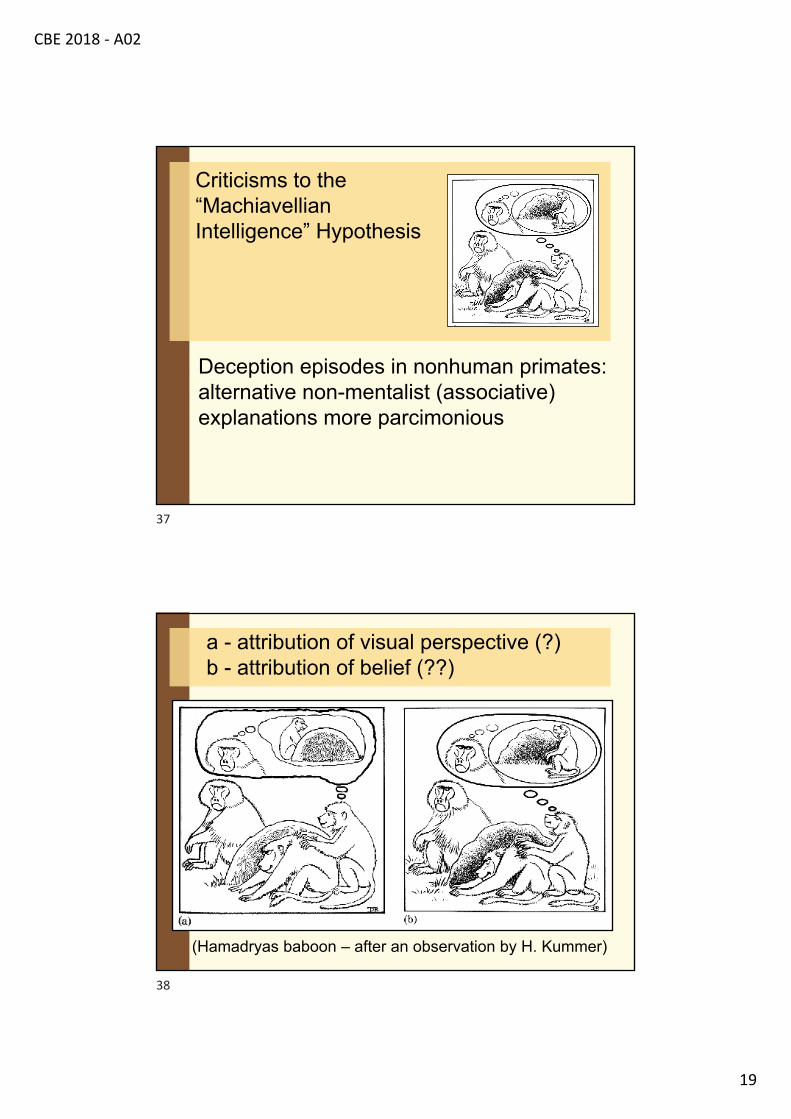
Criticisms to the “Machiavellian Intelligence” Hypothesis
Deception episodes in nonhuman primates: alternative non-mentalist (associative) explanations more parcimonious
a - attribution of visual perspective (?)b - attribution of belief (??)
(Hamadryas baboon – after an observation by H. Kummer)
37
38
CBE 2018 - A02
20
Criticisms to the “Machiavellian Intelligence” Hypothesis
“Situated action”/ “Socially distributed cognition”(Strum et al 1997, Hutchins 1991, Johnson 2001)
Encefalization and social group size(Dunbar 2003)
Neocortex X social variables
• social group size• grooming clique size• social skill in male mating strategies• frequency of tactical deception• frequency of play
ALL SIGNIFICANTBUT
Neocortex X ecological variables
• fruit in diet• home range size• day journey length• foraging style (categories of extractive foraging)
NOT SIGNIFICANT(when ecological vars. corrected for relative body size)
(of course, correlation ≠ causality...)
39
40
CBE 2018 - A02
21
Criticism: Reader & Laland (2002)
Frequencies of behavioral innovation / social learning / tool use: allsignificantly correlated with absolute and relative volume ofexecutive brain (neocortex plus striate cortex) in primates, whenappropriate controls are made for phylogeny and research effort.
Innovation and social learning co-vary across species: underminesthe claim of an evolutionary trade-off between reliance on socialexperience and on individual experience.Reader & Lefebvre (2001): no correlation between the sociallearning index and social group size once brain volume has beentaken into account.
Ecological factors may have been as (or more) important thansocial factors in primate brain evolution.
Encefalization and social group size(Dunbar 2003)
Dunbar replies to R&L criticisms:
Behaviors analized by R&L all refer to foraging situations: they do not comparecognitive demands in ecological problem solving (including social learning tosolve these problems) X management of social relationships.
Lack of correlation between social learning index used by R&L and group size:
(1) Cognitive demands for purely social tasks not required for foraging (ToM?)(2) Social tasks involve components not included in the executive brain
(Limbic System?)
Dunbar (2003): frontal lobe: even better fit to social group sizeFrontal lobe + Amygdala: social emotions, moral judgement etcSize of basolateral complex of the amygdala correlates w group size
Does not discard a role for ecological cognition in the start of theencefalization process: ex: color vision
Encefalization and social group size(Dunbar 2003)
41
42
CBE 2018 - A02
22
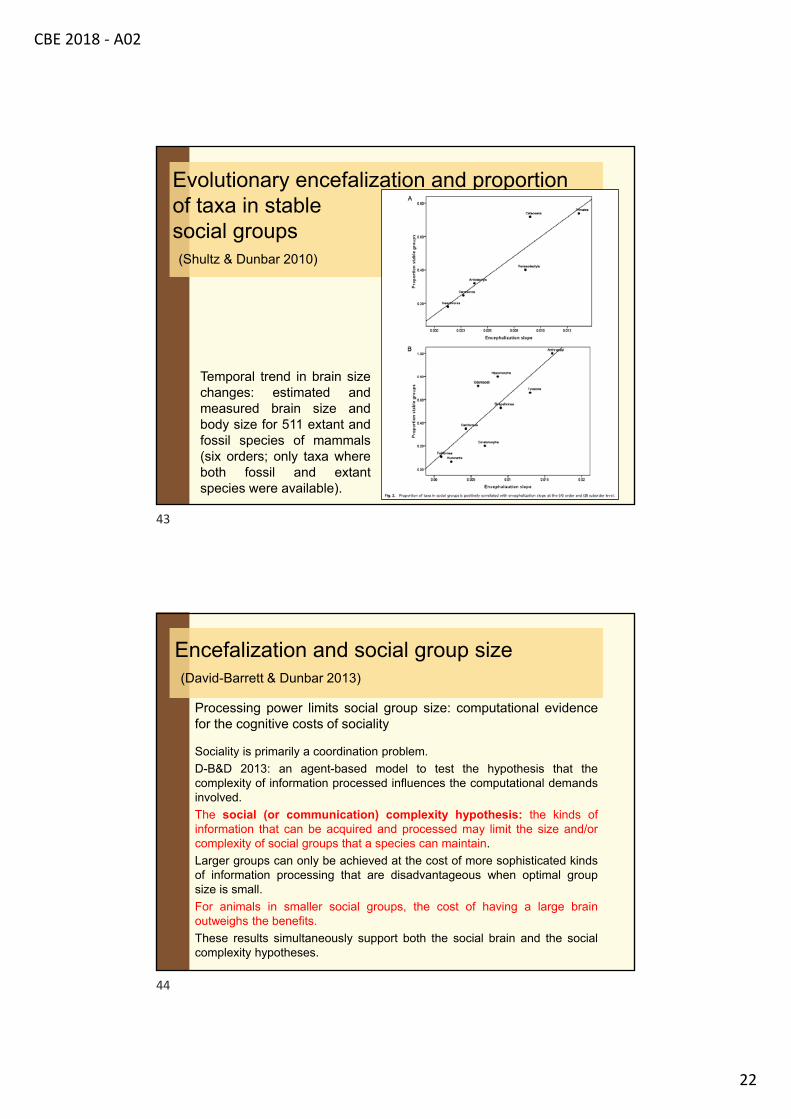
Evolutionary encefalization and proportionof taxa in stablesocial groups(Shultz & Dunbar 2010)
Temporal trend in brain sizechanges: estimated andmeasured brain size andbody size for 511 extant andfossil species of mammals(six orders; only taxa whereboth fossil and extantspecies were available).
Encefalization and social group size(David-Barrett & Dunbar 2013)
Processing power limits social group size: computational evidencefor the cognitive costs of sociality
Sociality is primarily a coordination problem.D-B&D 2013: an agent-based model to test the hypothesis that thecomplexity of information processed influences the computational demandsinvolved.The social (or communication) complexity hypothesis: the kinds ofinformation that can be acquired and processed may limit the size and/orcomplexity of social groups that a species can maintain.Larger groups can only be achieved at the cost of more sophisticated kindsof information processing that are disadvantageous when optimal groupsize is small.For animals in smaller social groups, the cost of having a large brainoutweighs the benefits.These results simultaneously support both the social brain and the socialcomplexity hypotheses.
43
44
CBE 2018 - A02
23
The “Communication complexity” hypothesis(Dunbar)
Origins of languageMusic: common origin or “cheesecake” (Pinker)?
“Hmmmm” (Mithen)
The “Cultural Intelligence” hypothesisvan Schaik & Pradhan 2003Whiten & van Schaik 2007
Positive selection of characters associated to social learning
Evolution of social tolerance
45
46
CBE 2018 - A02
24
Social cognition and “Theory of Mind”:anecdotes x experiments
Premack & Woodruff 1978
Povinelli & Vonk 2003
Tomasello, Call & Hare 2003
Do primates have a “Theory of Mind”?Understanding the other
as a causal agency
X understanding mental states
“Theory of Mind” (ToM): attribution of mental states to outher individuals, using them to predict and explain their behavior.
Premack & Woodruff (1978). Does a chimpanzee have a “Theory of Mind”?
A “limited” ToMin chimpanzees?
Povinelli et al 1994
47
48
CBE 2018 - A02
25
Mirror self-recognition
Chimpanzees,
(Gallup & Povinelli)
bonobos,
orangutans.
Gorillas?
but also
elephants,
dolphins,
corvids...
Povinelli et al 1998
Understanding the difference between intention and accident?
49
50
CBE 2018 - A02
26
Povinelli, Nelson & Boysen,1990
X
Povinelli & Eddy, 1996
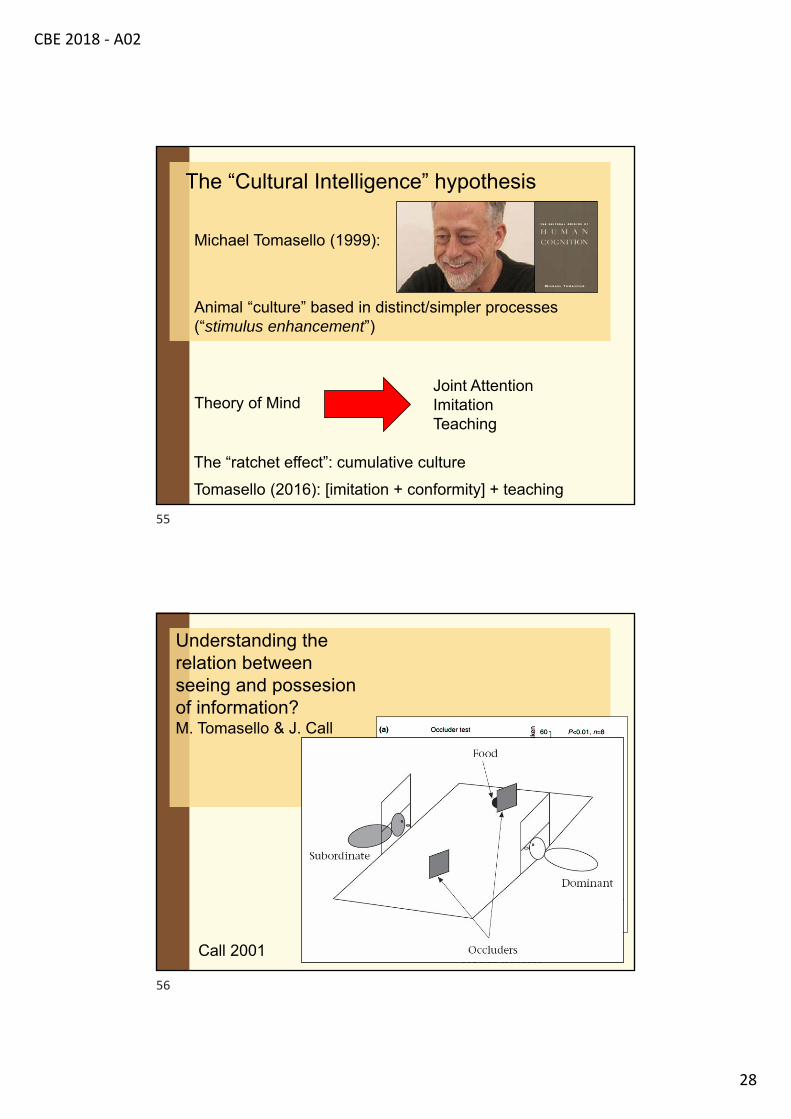
Understanding the relation between seeing and possession of information?
Esta é a SallySally põe suabola na caixa
Sally vai embora
Aonde a Sally vaiprocurar pela bola? Esta é a AnneAnne move a bolapara o seu cesto
The Sally-Anne task (Baron-Cohen,Leslie, & Frith, 1985)
ToM e Autism/AspergerBaron-Cohen 1995“Mindblindness”
Reality-biased responses: verbal tasks?“Where” question prematurely interpretedby young children as referring to thelocation of the hidden object?(Southgate et al 2007) The “Sally & Anne” [or “false belief”] test
Great apes distinguish true from false beliefs in an interactive helping taskButtelmann et al 2017 (16/18-mo children: Buttelmann et al 2009)
Helping behavior: true x false belief x ignorance conditions
“Niveis” de Teoria da Mente?
Leslie 1994:
ToBy (“Theory of Body”): agentes x não-agentes (início do 1º ano)
ToM I: pessoas como entes que percebem o ambiente e buscam certos propósitos (“agentes intencionais”) (final do 1º ano)
ToMII: mecanismo para lidar com os agentes como seres capazes de atitudes proposicionais (acreditar/ imaginar/ fingir) falsas ou verdadeiras (2º ano).
59
60
CBE 2018 - A02
31
Teoria da Mente: competências distintas?
Compreensão de intenções, desejos, pensamentos, conhecimento, crenças... Provavelmente não se desenvolvem ao mesmo tempo.
Bartsch & Wellman (1995)
Sequência de desenvolvimento:
1. 2 anos: compreensão de estados como atenção, emoções e percepçõessimples (não-representacional, sem interpretação das conexões daspessoas com as coisas em termos de “estados mentais”);
2. 3 anos: “pensamentos” e “desejos” como representações falsas ouverdadeiras, que podem variar de pessoa para pessoa;
3. 4 anos: ToM adulta, onde desejos e crenças interagem na causação docomportamento.
ToM and Levels of Intentionality(Dennett, DC)
Dunbar 2003:
Monkeys: 1st orderChimpanzees: 2nd orderAdult humans: 4th order
Ottoni, Rodriguez & Barreto 2006
3-13 yo:
• Understanding graphic representation (“thought bubbles”): 4/5 to 7 yo
• Understanding of “false belief”: 4/5 - 9 yo• 2nd / 3rd order (“nested”) representations: 9 yo.
La Frenière 1988The ontogeny of tactical deception in humans
“Cultural Intelligence” 2 information channels•Inheritable individual variation + Nat. selection•Non-genetic Social Information TransferNiche Construction: phenotypes as AGENTS of the evolutionary process (change selective pressures acting on genes)Gene-Culture Coevolution
Mammals and birds?
(... not necessarily mutually exclusive)
Evolutionary approachesto human behavior
Next:
63
64
CBE 2018 - A02
34
Adaptações cognitivas para a socialidade
PREDISPOSIÇÕES do bebê humano para a interação:Discriminação de rostos/ voz (“motherese”)/ movimento humanosInteração “dialogada” precoce (Papousek & Papousek).Recém-nascidos: “imitação” da expressão facial (Meltzoff & Moore)
Michael Tomasello: cultural learning and joint attention
67
68
CBE 2018 - A02
35
“Teoria da Mente”: Modulos Inatos x Construção de Teorias?
Autismo (Asperger)X
Sindrome de Williams
Prosopagnosia
“Estágios” no Desenvolvimentoda Teoria da MenteX
Flavell (1999): uma teoria adequada terá de integrar ambas as perspectivas...