The effect of different environmental conditions on the encystation of Acanthamoeba castellanii belonging to the T4 genotype Yousuf Aqeel, Ruqaiyyah Siddiqui, Hira Iftikhar, Naveed Ahmed Khan ⇑ Department of Biological and Biomedical Sciences, Aga Khan University, Karachi, Pakistan highlights Osmolarity is an important trigger to induce Acanthamoeba castellanii encystation. Cyst prepared from agar or liquid medium are similar in morphological and histological properties. Temperature over 37 °C or pH 9 adversely effected encystation. Light-dark cycles, 5% CO 2 and microaerophilic conditions had no effect on encystation of A. castellanii. graphical abstract article info Article history: Received 10 January 2013 Received in revised form 7 May 2013 Accepted 28 May 2013 Available online 11 June 2013 Keywords: Acanthamoeba Encystation Non-nutrient agar Osmolarity Physiological conditions Transmission electron microscopy abstract In this study, Acanthamoeba castellanii was cultivated under different stress conditions to induce possible encystation. The morphological and histological properties were analysed by light and electron micros- copy as well as cyst-specific staining. The findings revealed that cysts prepared through liquid medium using higher osmolarity as a trigger (10% glucose with 50 mM magnesium chloride for 72 h) are similar to cysts prepared using non-nutrient agar (nutrient deprivation as a trigger in plating assays for 14 days), as determined by SDS-resistance, cyst-specific Calcofluor white staining and transmission electron microscopy. Using liquid medium assay, A. castellanii encystation was studied by exposing trophozoites to media lacking growth ingredients (phosphate buffered saline or distilled water), inappropriate temper- atures (4–45 °C), pH (3–9), artificial light–dark cycles, 5% CO 2 , and microaerophilic conditions. Optimal encystation was observed when cells were incubated in PBS with 50 mM MgCl 2 and 10% glucose at 24–30 °C at pH 7. Increasing temperature over 37 °C or pH 9 adversely affected encystation, while light-dark cycles, 5% CO 2 and microaerophilic conditions had no effect on encystation of A. castellanii. None of the aforementioned conditions had any effect on the viability of A. castellanii, as determined by Trypan blue exclusion assay. A complete knowledge of encystation in A. castellanii is crucial to our understanding of the biology of these ecologically and medically important organisms. Ó 2013 Elsevier Inc. All rights reserved. 1. Introduction Acanthamoeba castellanii is a unicellular free-living protist path- ogen. The genus Acanthamoeba consists of 17 genotypes (T1–T17) (reviewed in Khan, 2012; Marciano-Cabral and Cabral, 2003), but the T4 genotype has been most frequently associated with blinding keratitis and fatal granulomatous amoebic encephalitis (GAE) which occurs mostly in immunocompromised individuals (Martinez and Visvesvara, 1997). Acanthamoeba keratitis is a pain- ful sight-threatening ulceration of the cornea and is most likely associated with the improper use of contact lenses (Marciano-Cab- ral and Cabral, 2003). The sequence of events in Acanthamoeba ker- atitis involves the breakdown of the epithelial barrier, stromal 0014-4894/$ - see front matter Ó 2013 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.exppara.2013.05.017 ⇑ Corresponding author. Address: Department of Biological and Biomedical Sciences, Aga Khan University, Stadium Road, Karachi, Pakistan. Fax: +92 (0)21 3493 4294. E-mail address: [email protected](N.A. Khan). Experimental Parasitology 135 (2013) 30–35 Contents lists available at SciVerse ScienceDirect Experimental Parasitology journal homepage: www.elsevier.com/locate/yexpr
Transcript
Experimental Parasitology 135 (2013) 30–35
Contents lists available at SciVerse ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
The effect of different environmental conditions on the encystation ofAcanthamoeba castellanii belonging to the T4 genotype
0014-4894/$ - see front matter � 2013 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.exppara.2013.05.017
⇑ Corresponding author. Address: Department of Biological and BiomedicalSciences, Aga Khan University, Stadium Road, Karachi, Pakistan. Fax: +92 (0)213493 4294.
Yousuf Aqeel, Ruqaiyyah Siddiqui, Hira Iftikhar, Naveed Ahmed Khan ⇑Department of Biological and Biomedical Sciences, Aga Khan University, Karachi, Pakistan
h i g h l i g h t s
� Osmolarity is an important trigger toinduce Acanthamoeba castellaniiencystation.� Cyst prepared from agar or liquid
medium are similar in morphologicaland histological properties.� Temperature over 37 �C or pH 9
adversely effected encystation.� Light-dark cycles, 5% CO2 and
microaerophilic conditions had noeffect on encystation of A. castellanii.
g r a p h i c a l a b s t r a c t
a r t i c l e i n f o
Article history:Received 10 January 2013Received in revised form 7 May 2013Accepted 28 May 2013Available online 11 June 2013
Keywords:AcanthamoebaEncystationNon-nutrient agarOsmolarityPhysiological conditionsTransmission electron microscopy
a b s t r a c t
In this study, Acanthamoeba castellanii was cultivated under different stress conditions to induce possibleencystation. The morphological and histological properties were analysed by light and electron micros-copy as well as cyst-specific staining. The findings revealed that cysts prepared through liquid mediumusing higher osmolarity as a trigger (10% glucose with 50 mM magnesium chloride for 72 h) are similarto cysts prepared using non-nutrient agar (nutrient deprivation as a trigger in plating assays for 14 days),as determined by SDS-resistance, cyst-specific Calcofluor white staining and transmission electronmicroscopy. Using liquid medium assay, A. castellanii encystation was studied by exposing trophozoitesto media lacking growth ingredients (phosphate buffered saline or distilled water), inappropriate temper-atures (4–45 �C), pH (3–9), artificial light–dark cycles, 5% CO2, and microaerophilic conditions. Optimalencystation was observed when cells were incubated in PBS with 50 mM MgCl2 and 10% glucose at24–30 �C at pH 7. Increasing temperature over 37 �C or pH 9 adversely affected encystation, whilelight-dark cycles, 5% CO2 and microaerophilic conditions had no effect on encystation of A. castellanii.None of the aforementioned conditions had any effect on the viability of A. castellanii, as determinedby Trypan blue exclusion assay. A complete knowledge of encystation in A. castellanii is crucial to ourunderstanding of the biology of these ecologically and medically important organisms.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction (reviewed in Khan, 2012; Marciano-Cabral and Cabral, 2003), but
Acanthamoeba castellanii is a unicellular free-living protist path-ogen. The genus Acanthamoeba consists of 17 genotypes (T1–T17)
the T4 genotype has been most frequently associated with blindingkeratitis and fatal granulomatous amoebic encephalitis (GAE)which occurs mostly in immunocompromised individuals(Martinez and Visvesvara, 1997). Acanthamoeba keratitis is a pain-ful sight-threatening ulceration of the cornea and is most likelyassociated with the improper use of contact lenses (Marciano-Cab-ral and Cabral, 2003). The sequence of events in Acanthamoeba ker-atitis involves the breakdown of the epithelial barrier, stromal
Y. Aqeel et al. / Experimental Parasitology 135 (2013) 30–35 31
invasion by Acanthamoeba, keratocyte depletion, induction of anintense inflammatory response and finally stromal necrosis (Gar-ner, 1993; Vemuganti et al., 2004). Treatment is problematic andconsists predominantly of hourly topical application of a mixtureof drugs including polyhexamethylene biguanide or chlorhexidinedigluconate together with propamidine isethionate or hexamidine.Moreover chloramphenicol or neomycin is also given to preventmixed bacterial infection (Perez-Santonja et al., 2003). The treat-ment lasts for several months and even then, recurrence can occur(Ficker et al., 1990). This is due to the ability of Acanthamoeba torapidly adapt to extracellular changes and differentiate into a resis-tant cyst form under harsh conditions (Byers, 1979; Hirukawaet al., 1998; Khunkitti et al., 1998; Turner et al., 2000; Weisman,1976). The Cysts are double-walled, metabolically inactive struc-tures that pose a major challenge in the successful treatment ofinfection. In the present study, we investigated encystation in A.castellanii belonging to the T4 genotype, in vitro by exposing tro-phozoites to various environmental and physiological adverseconditions.
2. Materials and methods
2.1. A. castellanii cultures
All chemicals were purchased from Sigma Labs (Poole, Dorset,England), unless otherwise stated. A. castellanii belonging to theT4 genotype (ATCC 50492) was originally isolated from a keratitispatient and was grown in 75 cm2 tissue culture flasks in the pres-ence of 10 mL PYG medium [proteose peptone 0.75% (w/v), yeastextract 0.75% (w/v) and glucose 1.5% (w/v)] without shaking at37 �C as described previously (Aqeel et al., 2012). Media were re-freshed 15–20 h prior to experiments. A. castellanii adherent toflasks represented the trophozoite form and were used in all sub-sequent assays.
2.2. Encystation assays
Encystation assays were performed as described previously(Dudley et al., 2009) but with slight modifications. Briefly,2 � 106 A. castellanii were inoculated in phosphate buffered saline(PBS; containing 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4,1.8 mM KH2PO4) in the presence of 50 mM MgCl2 and 10% glucose(i.e., encystation trigger is higher osmolarity) in 24 well-tissue cul-ture plates at 37 �C. After 72 h, A. castellanii viability was quantifiedusing a haemocytometer, by Trypan blue exclusion assay. Next, so-dium dodecyl sulfate (SDS, 0.5% final concentration) was addedand placed on a shaker for 10 min to solubilize all the trophozoitesand cysts were enumerated using a haemocytometer (cysts areresistant to 0.5% SDS). The percentage encystation was determinedas follows; No. of A. castellanii post-SDS treatment/total number ofA. castellanii � 100 = % encystation. Data are represented as themean ± standard error of three independent experiments.
2.3. Environmental conditions
To study the effects of different stress conditions on the induc-tion of encystation in A. castellanii, assays were performed in thepresence of PBS and distilled water (nutrient deprivation), variabletemperatures (4–45 �C), different pH (3–9), artificial light and darkcycles, 5% CO2 and microaerophilic conditions.
To identify the optimum liquid medium and the role of nutrientdeprivation on the encystation of A. castellanii, 2 � 106 amoebaewere inoculated in the presence or absence of 10% glucose and50 mM MgCl2 in PBS or distilled water at 30 �C for 72 h. After this
incubation trophozoites were lysed with 0.5% SDS treatment andcysts were enumerated by haemocytometer counting.
To determine the optimum temperature for A. castellanii differ-entiation, encystation assays were performed at variable tempera-tures (4–45 �C) in the absence and presence of 50 mM MgCl2 and10% glucose. After this incubation trophozoites were lysed with0.5% SDS treatment and cysts counts were determined by haemo-cytometer counting.
To study the effects of pH on A. castellanii, encystation assayswere performed using different pH buffers (3.0, 5.0, 7.0 and 9.0).The pH 3 and 5 were obtained using citrate-phosphate buffer[0.2 M sodium phosphate (Na2HPO4) and 0.1 M citrate buffer andpH adjusted to 3.0 or 5.0]; pH 7 was obtained using PBS bufferand pH adjusted to 7.0; and pH 9 was obtained using glycine–so-dium hydroxide buffer (0.2 M glycine and 0.2 M sodium hydroxidebuffer and pH adjusted to 9.0). Plates were then incubated at 30 �Cin the absence and presence of 50 mM MgCl2 and 10% glucose.After 72 h, 0.5% SDS was added to lyse trophozoites and cystscounts determined using a haemocytometer.
The effect of complete light, complete dark and 12 h light/darkcycle on the encystation of A. castellanii was studied by inoculat-ing 2 � 106 amoebae in 24 well tissue culture plates in the pres-ence and absence of 10% glucose and 50 mM MgCl2 at 37 �Cincubator under complete dark or under complete light (visiblelight). For light/dark cycles, plates were incubated at 37 �C incu-bator set with automatic Panasonic timer (TB2118) and Philipsbulb (visible light). After 72 h, cells were treated with 0.5% SDSto lyse trophozoites and cysts were enumerated using ahaemocytometer.
To study the effect of 5% CO2 on encystation, 2 � 106 cells of A.castellanii were inoculated in PBS. Amoebae were then dispensed in24 well tissue culture plates in the absence or presence of 10% glu-cose and 50 mM MgCl2. Plates were then incubated at 37 �C in ashel lab water jacketed tissue culture incubator which maintains5% CO2 level. After this incubation trophozoites were lysed with0.5% SDS treatment and cysts were enumerated by haemocytome-ter counting.
The role of microaerophilic conditions on A. castellanii encysta-tion were determined by inoculating 2 � 106 amoebae in the pres-ence and absence of 10% glucose and 50 mM MgCl2 in 35 mmtissue culture dishes. Amoebae were then incubated in oxoidanaerobic jars loaded with campy gen bags (absorbs all the freeoxygen) at 37 �C. After 72 h, 0.5% SDS was added and amoeba cystswere enumerated using a haemocytometer.
2.4. Calcofluor white staining
A. castellanii trophozoites were inoculated on 3% non-nutrientagar plates and incubated at 37 �C for 14 days (encystation triggerwas nutrient deprivation). Cysts were collected by scraping off theagar surface using a rubber policeman and then resuspended inPBS. Simultaneously, A. castellanii cysts were also prepared by inoc-ulating trophozoites at 37 �C for 72 h with 50 mM MgCl2 and 10%glucose in PBS (encystation trigger was higher osmolarity). Cystswere centrifuged at 900g for 20 min. Following this, cysts wereresuspended in 2.5% Calcofluor white stain and incubated at30 �C for 120 min. Next, cysts were collected by centrifugation at900g and resuspended in PBS. This process was repeated twice toremove excess Calcofluor white stain, and finally cysts resus-pended in PBS. Finally, 30 lL suspension was applied to glass slidesand wet mounts of each slide were examined under Olympus, BX-41 fluorescent microscope (excitation 405 nm and emission bandpass 420–480 nm). Our previous studies have shown that Calcoflu-or white does not stain trophozoite stage of amoebae (Dudley et al.,2007), hence A. castellanii trophozoites were used as negative con-trols. Similarly, previous studies have shown that Calcofluor white
32 Y. Aqeel et al. / Experimental Parasitology 135 (2013) 30–35
stains cyst stage of Balamuthia mandrillaris (Siddiqui et al., 2009),hence B. mandrillaris cysts were used as positive controls in presentstudies.
In some experiments, excystation was determined. Briefly, cystswere collected from both methods (non-nutrient agar plate meth-od and glucose liquid medium method) and enumerated. Finally,2 � 105 cysts from both methods were inoculated in the growthmedium, i.e., PYG and plates incubated at 30 �C for 72 h to deter-mine excystation and growth of amoebae. Amoebae counts weredetermined using a haemocytometer after 3 days. The results arerepresentative of at least three independent experiments and pre-sented as the mean ± standard error.
2.5. Transmission electron microscopy
Acanthamoeba castellanii cysts were fixed in 3% gluteraldehyde(in 0.1 M veranol acetate buffer, pH 7.2) and incubated for120 min at room temperature. Next, cysts were collected by centri-fugation at 900g and resuspended in 0.1 M veranol acetate buffer.This process was repeated twice to remove primary fixative. Sec-ondary fixation was performed in 2% osmium tetroxide made in0.1 M veranol acetate buffer and incubated for 60 min at roomtemperature. Cysts were washed again as described above and cellswere embedded in 3% Nobel agar which was then sliced into 1–2 mm cubes. Next, dehydration of A. castellanii was performedusing graded series of ethanol including 60 min staining in a satu-rated solution of uranyl acetate at 70% ethanol dehydration step in1:1 ratio. Subsequently cysts were infiltrated with LR white resinand polymerized at 67 �C for 24 h. Thick sections of 0.5 lm weresliced and mounted on a glass slide and stained with toluidineand observed under light microscope. Thin sections of 80–100 nm were cut using a NOVA ultramicrotome. The sections werecollected on formvar coated grids and stained with lead citrate for20 min. After rinsing with filtered dH2O, secondary staining withuranyl acetate was performed for 5 min. The stained sections werethen viewed through a JEOL, JEM Transmission electron micro-scope at 60 kV and images were developed using Kodak X-ray films(González-Robles et al., 2008).
3. Results
3.1. Cysts prepared through nutrient deprivation or higher osmolarityare similar
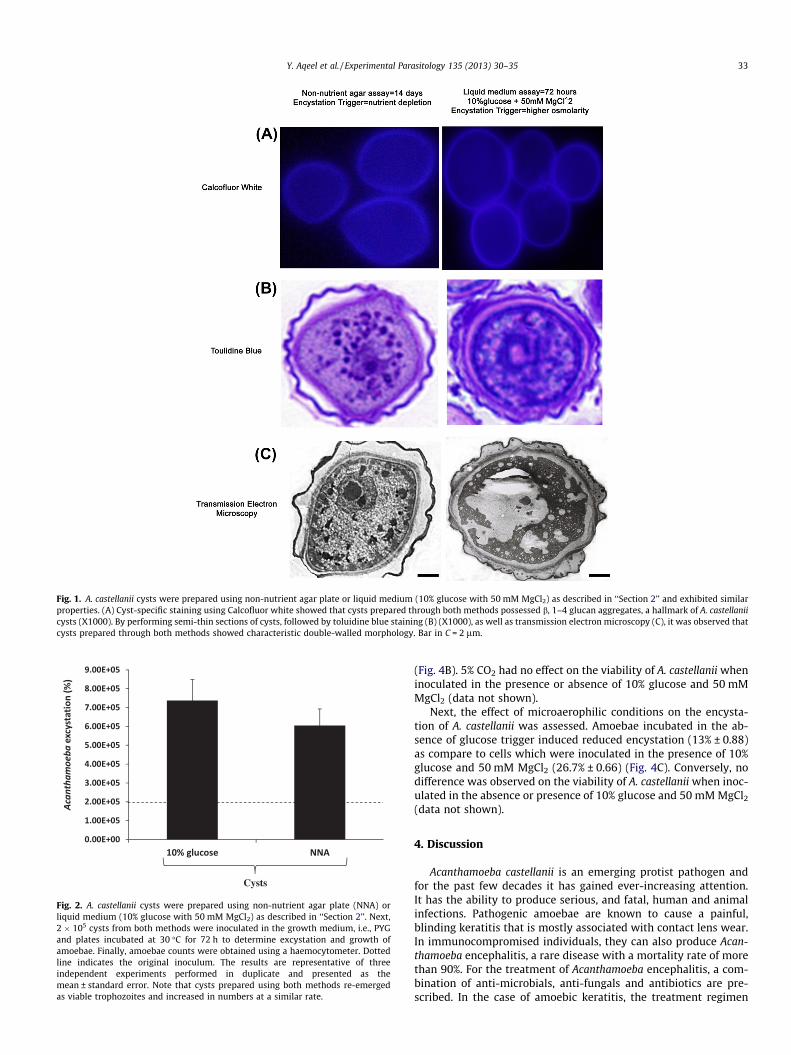
To determine if the A. castellanii cysts prepared using non-nutri-ent agar are similar to cysts prepared using 10% glucose, encysta-tion assays were performed. Cysts were stained with Calcofluorwhite staining. Calcofluor white binds specifically to b-1, 3 andb-1, 4 linked glycans present in the cyst wall of Acanthamoebabut not trophozoites (Dudley et al., 2007). The staining revealedthat there is no difference in the cysts prepared in non-nutrientagar or 10% glucose in 24 well plates (Fig 1A). When cysts werevisualized under the fluorescent microscope, distinct layers werevisible, and fluorescence was observed in cysts prepared throughboth encystation methods. For controls, Calcofluor white did notstain trophozoites of A. castellanii but exhibited fluorescence whentested against cysts of B. mandrillaris as demonstrated previously(data not shown) (Dudley et al., 2007; Siddiqui et al., 2009). To fur-ther evaluate any morphological differences between cysts pro-duced using the two encystation methods, toluidine blue stainingand transmission electron microscopy were performed. The resultsshowed comparatively similar cyst wall structure when both medi-ums were used (Fig. 1B and C). A distinct outer layer and innerlayer were present in cysts prepared through both encystationmethods. However, the percentage encystation was greater when
amoebae were inoculated on non-nutrient agar (up to 100% ency-station), compared with �40% encystation using higher osmolarity(i.e., 10% glucose). When incubated in the growth medium, i.e.,PYG, cysts prepared using both methods re-emerged as viable tro-phozoites and increased in numbers at a similar rate (Fig. 2).
3.2. Phosphate buffered saline or distilled water had no effect, whereaspH above 9 and temperatures over 37 �C adversely effected A.castellanii encystation
To determine the optimum liquid medium for inducing ency-station in A. castellanii, encystation assays were performed usingPBS and distilled water in the presence and absence of 10% glucoseas a trigger for encystation. No considerable difference in the ency-station rate was observed when cells were inoculated in PBS or dis-tilled water in the presence of 10% glucose and 50 mM MgCl2
(35.8 ± 3.5% and 30 ± 5.4%, respectively). Simultaneously no majoreffect was observed on the encystation of A. castellanii when cellswere inoculated without glucose (4.3 ± 0.38% in PBS and2.8 ± 1.18% in distilled water). However, as 10% glucose serves asa strong stimulus for encystation (i.e., nutrient depletion and high-er osmolarity), therefore more encystation was observed in thismedia as compared to the cells which are inoculated without glu-cose (Fig. 3A). PBS and distilled water had no effects on the viabilityof A. castellanii (data not shown).
To identify the optimum temperature for A. castellanii encysta-tion, assays were performed at variable temperatures ranging from4–45 �C. In the absence of the glucose trigger, maximum encysta-tion was observed at room temperature (24 �C). In the presenceof 10% glucose and 50 mM MgCl2, a comparatively similar encysta-tion pattern was observed at temperatures ranging from 4 �C,24 �C, 30 �C and 37 �C. However, an increase in temperatures above37 �C had deleterious effects on encystation and this was reducedmarginally both in the presence or absence of the glucose triggerand 50 mM MgCl2 (Fig. 3B). All the temperatures tested had no ef-fects on the viability of A. castellanii, either in the presence or ab-sence of 10% glucose and 50 mM MgCl2 (data not shown).
To study the effects of variable pH on A. castellanii differentia-tion, encystation assays were performed at pHs ranging from 3 to9. The results revealed that in the absence of 10% glucose and50 mM MgCl2, maximum encystation occurred at pH 3. Howeverin the presence of glucose and 50 mM MgCl2, pH 7 was found tobe the optimum for encystation (approx. 33 ± 5.31%). A relativelysimilar pattern in encystation was observed from pH 3 to pH 7.However higher pH adversely effected encystation in the presenceor absence of 10% glucose and 50 mM MgCl2 (Fig. 3C). No effect onthe viability of A. castellanii was observed when cells are inoculatedfor encystation at different pH with or without the presence of 10%glucose and 50 mM MgCl2 (data not shown).
3.3. Light/dark cycles, 5% CO2 and microaerophilic conditions had noeffect on A. castellanii encystation
The effect of light and dark conditions (complete light, completedark, 12 h light/dark) on the encystation of A. castellanii was as-sessed in the presence and absence of 10% glucose and 50 mMMgCl2. In the presence of 10% glucose and 50 mM MgCl2,46.5% ± 0.48, 44.1% ± 3.66 and 42.8% ± 4.6 (light, dark, 12 h light/dark cycles, respectively) encystation was observed in A. castellanii(Fig. 4A). The survival of A. castellanii was not affected by variouslight and dark conditions (data not shown).
To determine the effect of 5% CO2 on the encystation A. castella-nii, amoebae were incubated in a tissue culture CO2 incubator. Inthe absence of glucose trigger, CO2 did not serve as a stimulus toinduce encystation. Whereas amoebae inoculated with 10%glucoseand 50 mM MgCl2 induced substantial encystation (31.7% ± 2.97)
Fig. 1. A. castellanii cysts were prepared using non-nutrient agar plate or liquid medium (10% glucose with 50 mM MgCl2) as described in ‘‘Section 2’’ and exhibited similarproperties. (A) Cyst-specific staining using Calcofluor white showed that cysts prepared through both methods possessed b, 1–4 glucan aggregates, a hallmark of A. castellaniicysts (X1000). By performing semi-thin sections of cysts, followed by toluidine blue staining (B) (X1000), as well as transmission electron microscopy (C), it was observed thatcysts prepared through both methods showed characteristic double-walled morphology. Bar in C = 2 lm.
Fig. 2. A. castellanii cysts were prepared using non-nutrient agar plate (NNA) orliquid medium (10% glucose with 50 mM MgCl2) as described in ‘‘Section 2’’. Next,2 � 105 cysts from both methods were inoculated in the growth medium, i.e., PYGand plates incubated at 30 �C for 72 h to determine excystation and growth ofamoebae. Finally, amoebae counts were obtained using a haemocytometer. Dottedline indicates the original inoculum. The results are representative of threeindependent experiments performed in duplicate and presented as themean ± standard error. Note that cysts prepared using both methods re-emergedas viable trophozoites and increased in numbers at a similar rate.
Y. Aqeel et al. / Experimental Parasitology 135 (2013) 30–35 33
(Fig. 4B). 5% CO2 had no effect on the viability of A. castellanii wheninoculated in the presence or absence of 10% glucose and 50 mMMgCl2 (data not shown).
Next, the effect of microaerophilic conditions on the encysta-tion of A. castellanii was assessed. Amoebae incubated in the ab-sence of glucose trigger induced reduced encystation (13% ± 0.88)as compare to cells which were inoculated in the presence of 10%glucose and 50 mM MgCl2 (26.7% ± 0.66) (Fig. 4C). Conversely, nodifference was observed on the viability of A. castellanii when inoc-ulated in the absence or presence of 10% glucose and 50 mM MgCl2
(data not shown).
4. Discussion
Acanthamoeba castellanii is an emerging protist pathogen andfor the past few decades it has gained ever-increasing attention.It has the ability to produce serious, and fatal, human and animalinfections. Pathogenic amoebae are known to cause a painful,blinding keratitis that is mostly associated with contact lens wear.In immunocompromised individuals, they can also produce Acan-thamoeba encephalitis, a rare disease with a mortality rate of morethan 90%. For the treatment of Acanthamoeba encephalitis, a com-bination of anti-microbials, anti-fungals and antibiotics are pre-scribed. In the case of amoebic keratitis, the treatment regimen
(A)
(B)
(C)
Fig. 3. To determine the effects of PBS and distilled water, variable pH andtemperatures on A. castellanii encystation, assays were performed by inoculating2 � 106 amebae trophozoites under different environmental conditions for 72 h.After this incubation, 0.5% SDS was added to solubilize trophozoites and cysts werecounted using a haemocytometer as described in ‘‘Section 2’’. Note that the use ofPBS or distilled water had no effect on A. castellanii encystation (A), whiletemperatures above 37 �C (B) and pH above 7.0 (C) adversely affected A. castellaniiencystation. The results are presented as the mean ± standard error of at least threeindependent experiments performed in duplicate.
(A)
(B)
(C)
Fig. 4. Light and dark cycles, 5% CO2 and microaerophilic environment had no effecton the encystation of A. castellanii. (A) Encystation of A. castellanii in the presence ofcomplete light, complete dark and 12 h light/dark cycles were studied as describedin ‘‘Materials and Methods’’. Similarly, 5% CO2 (B) and microaerophilic conditions(C) had no effect on A. castellanii encystation. The results are presented as themean ± standard error of at least three independent experiments performed induplicate.
34 Y. Aqeel et al. / Experimental Parasitology 135 (2013) 30–35
Y. Aqeel et al. / Experimental Parasitology 135 (2013) 30–35 35
includes the topical application of polyhexamethylene biguanideor chlorhexidine together with pentamidine or hexamidine, de-spite this recurrence of infection is commonly observed (Khan,2012; Marciano-Cabral and Cabral, 2003). It is the ability of Acan-thamoeba to transform into a dormant, resistant cyst form whichleads to resistance to treatment. Once encystation occurs, theorganism is resistant to harsh environmental conditions and mostantimicrobial drugs. Consequently, it is pivotal to understand theprocess of encystation.
A common method used to induce encystation in Acanthamoebais to inoculate the organism onto non-nutrient agar plates thuseliminating the food source for amoeba and providing a stimulusfor encystation. The amoebae can take up to 2 weeks to form cysts,which is time consuming. Moreover, it is difficult to study the ef-fects of compounds or environmental conditions quantitativelyand robustly using this method. Previous studies have suggestedosmolarity as a trigger to induce encystation in Acanthamoebausing liquid medium (Cordingley et al., 1996; Dudley et al.,2005), however it has not been determined whether this methodproduce cysts of similar properties. For the first time, here wedetermined whether cysts produced using both of these methodsare similar in morphology and histology.
Recently, the total carbohydrate composition of A. castellaniicyst walls using gas chromatography/mass spectrometry showedthat approximately 22.2% of the cyst walls of A. castellanii are com-posed of 1–4 linked glycan sugars (Dudley et al., 2009). Further-more, the fluorescent brightener Calcofluor White (4,4⁄Œ-bis[4-anilino-6-bis(2-hydroxyethyl)amino-s-trizin-2-ylamino]-2-2⁄Œstilbenedisulfonic-acid), a stilbene dye is shown to bind to b-glucan aggregates in A. castellanii cysts and emits fluorescencewhen observed under fluorescent microscope, but not trophozoites(Dudley et al., 2007). Therefore, Calcofluor white staining is a veryuseful marker to differentiate A. castellanii cysts from trophozoites.Using Calcofluor white staining, the present findings showedclearly that cysts prepared through both methods (agar platemethod or liquid medium method) are similar. Also, the resistanceof cysts (prepared through both methods) to 0.5% SDS impliedresemblance in their properties. Another key feature of A. castella-nii cysts is its characteristic double-wall. By performing semi-thinsections of cysts, followed by toluidine blue staining as well astransmission electron microscopy, the results showed characteris-tic double-walled morphology of cysts prepared through bothmethods.
Having established that 10% glucose along with MgCl2 producedcysts comparable to the non-nutrient agar assay, we determinedthe effect of various environmental conditions on encystation.The results revealed that CO2, microaerophilic environment andlight/dark cycles had no effect on encystation or viability of A. cas-tellanii. However, further studies are needed when Acanthamoebaspp. are cultured in such conditions for a number of generationsto verify our findings. The acidic pH 3.0 did not serve as a strongstimulus for encystation of A. castellanii. These findings are similarto other protists, Giardia lamblia and Entamoeba histolytica, whichexhibit high excystation at acidic pH (Boucher and Gillin, 1990;Makioka et al., 2006), suggesting possible conserve mechanisms.A neutral pH 7.0 served as an optimum medium in which approx-imately 30–40% encystation of A. castellanii was observed. It wasinteresting that A. castellanii in the presence of 10% glucose at4 �C exhibited increased encystation compared with higher tem-peratures. A likely explanation is that these conditions provide dif-ferent triggers (i.e., lack of nutrients, higher osmolarity, lowtemperature) that act in synergy to stimulate encystation. Overall,
the findings demonstrated that varying environmental conditionsalone without 10% glucose and 50 mM MgCl2 did not serve as astrong stimulus to induce encystation and higher osmolarity isan important trigger for A. castellanii encystation. The possiblemechanisms of osmolarity-induced encystation have been sug-gested to induce conformational changes in the receptor(s) ex-pressed on the surface of A. castellanii (Cordingley et al., 1996),and being investigated further. Future studies in the understandingof the molecular mechanisms associated with Acanthamoeba ency-station will provide insights into the biology of these importantorganisms, which will in turn be of value in the rationale develop-ment of therapeutic interventions.
Acknowledgments
The authors are thankful to Abdus Salam, Jinnah PostgraduateMedical Centre for his support related to TEM studies. This workwas supported by the Aga Khan University
References
Aqeel, Y., Siddiqui, R., Khan, N.A., 2012. Anti- Acanthamoebic properties ofresveratrol and de-methoxy curcumin. Exp. Parasitol. 132, 519–523.
Byers, T.J., 1979. Growth, reproduction, and differentiation in Acanthamoeba. Int.Rev. Cytol. 61, 283–338.
Cordingley, J.S., Wills, R.A., Villemez, C.L., 1996. Osmolarity is an independenttrigger of Acanthamoeba castellanii differentiation. J. Cell. Biochem. 61, 167–171.
Dudley, R., Matin, A., Alsam, S., Sissons, J., Maghsood, A.H., Khan, N.A., 2005.Acanthamoeba isolates belonging to T1, T2, T3, T4 but not T7 encyst in responseto increased osmolarity and cysts do not bind to human corneal epithelial cells.Acta Trop. 95, 100–108.
Dudley, R., Alsam, S., Khan, N.A., 2007. Cellulose biosynthesis pathway is a potentialtarget in the improved treatment of Acanthamoeba keratitis. Appl. Microbiol.Biotechnol. 75, 133–140.
Dudley, R., Jarroll, E.L., Khan, N.A., 2009. Carbohydrate analysis of Acanthamoebacastellanii. Exp. Parasitol. 122, 338–343.
Ficker, L., Seal, D., Warhurst, D., Wright, P., 1990. Acanthamoeba keratitis: resistanceto medical therapy. Eye 4, 835–838.
Garner, A., 1993. Pathogenesis of acanthamoebic keratitis: hypothesis based on ahistological analysis of 30 cases. Br. J. Ophthalmol. 77, 366–370.
González-Robles, A., Castañón, G., Hernández-Ramírez, V.I., Salazar-Villatoro, V.,González-Lázaro, M., Omaña-Molina, M., Talamás-Rohana, P., Martínez-Palomo,A., 2008. Acanthamoeba catellanii: identification and distribution ofcytoskeleton. Exp. Parasitol. 119, 411–417.
Hirukawa, Y., Nakato, H., Izumi, S., Tsuruhara, T., Tomino, S., 1998. Structure andexpression of a cyst specific protein of Acanthamoeba castellanii. Biochim.Biophys. Acta 1398, 47–56.
Khan, N.A., 2012. Biology and pathogenesis of Acanthamoeba. Parasit. Vectors 5, 6.Khunkitti, W., Hann, A.C., Lloyd, D., Furr, J.R., Russell, A.D., 1998. Biguanide-induced
changes in Acanthamoeba castellanii: an electron microscopic study. J. Appl.Microbiol. 84, 53–62.
Makioka, A., Kumagai, M., Kobayashi, S., Takeuchi, T., 2006. Effect of artificialgastrointestinal fluids on the excystation and metacystic development ofEntamoeba invadens. Parasitol. Res. 98, 443–446.
Marciano-Cabral, F., Cabral, G., 2003. Acanthamoeba spp. as agents of disease inhumans. Clin. Microbiol. Rev. 16, 273–307.
Perez-Santonja, J.J., Kilvington, S., Hughes, R., Tufail, A., Metheson, M., Dart, J.K.G.,2003. Persistently culture positive Acanthamoeba keratitis; in vivo resistanceand in vitro sensitivity. Ophthalmology 110, 1593–1600.
Siddiqui, R., Jarroll, E.L., Khan, N.A., 2009. Balamuthia mandrillaris: stainingproperties of cysts and trophozoites and the effect of 2,6-dichlorobenzonitrileand calcofluor white on encystment. J. Eukaryot. Microbiol. 56 (2), 136–141.
Turner, N.A., Russell, A.D., Furr, J.R., Lloyd, D., 2000. Emergence of resistance tobiocides during differentiation of Acanthamoeba castellanii. J Antimicrob.Chemother. 46, 27–34.
Vemuganti, G.K., Reddy, K., Iftekhar, G., Garg, P., Sharma, S., 2004. Keratocyte loss incorneal infection through apoptosis: a histologic study of 59 cases. BMCOphthalmol. 4, 16.