ARCHIVES OFBIOCHEMISTRY AND BIOPHYSICS Vol. 234, No. 1, October, pp. 45-54,1934 The Effect of Hypolipidemic Drugs on Plant Lipid Metabolism JUAN SANCHEZ’ AND PAUL K. STUMPF2 Department of Biochemistry and Biophysics, University of California, Davis, Califbrnin 95616 Received February 22, 1934, and in revised form June 18, 1934 The effect of hypolipidemic drugs, WY14643 and DH990, on plant lipid metabolism has been studied. The total incorporation of [14C]acetate into lipids was inhibited by addition of both drugs to aged potato (Solarium tuberosum) tuber discs, spinach (Spinucia oleracea) leaves, and spinach chloroplasts, while the incorporation in Chbrella vulgaris cells was affected only by DH990. Moreover, DH990 inhibited the incorporation of 14C-labeled fatty acids into phosphatidylcholine and phosphatidyleth- anolamine of potato discs, and decreased the incorporation into phosphatidylglycerol of ChZoreZZu cells. DH990 inhibited the formation of polyunsaturated fatty acids in potato discs, Chlorella cells, and spinach leaves, whereas WY14643 had no effect on the formation of these fatty acids. Stearoyl-ACP desaturase from safflower (Carthumus tinctorius) seeds was very sensitive to both drugs, especially DH990, which completely blocked the activity at 2 mM levels. When safflower lysophospholipid acyltransferases were solubilized by detergent treatment, only DH990 inhibited the incorporation of [14C]oleoyl-CoA into lysophosphatidylcholine or lysophosphatidylethanolamine. Both drugs inhibited fatty acid synthesis from [‘4C]malonyl-CoA in the microsomal fraction from safflower seeds, but only DH990 inhibited FAS activity in the soluble fraction; both drugs inhibited severely the formation of stearic acid. Both acetyl-CoA carboxylase and acetyl-CoA synthetase were sensitive to both drugs. Q last Academic PM, I~~. Hypolipidemic drugs have been exten- sively used to decrease the level of blood lipid in patients affected by diseases such as atherosclerosis, but the mode of action of these drugs remains to be elucidated. Recently (1) it was reported that some hypolipidemic drugs blocked the formation of PC3 in rat liver microsomes by inhib- i Present address: ARC0 Plant Cell Research In- stitute, 6569 Trinity Court, Dublin, Calif. 94563. * To whom correspondence should be addressed. aAbbreviations used: PC, phosphatidylcholine; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; PLs, polar lipids; SL, sulfolipids; MGDG, monoga- lactosyldiacylglycerol;DGDG,digalactosyldiacylglyc- erol; FFA, free fatty acids; NL, neutral lipids (in- cluding diacylglycerol and triacylglycerol); WY, WY14643; DH, DH990; BSA, bovine serum albumin; ACP, acyl carrier protein; PCS, phase combining iting both cholinephosphotransferase (EC 2.7.8.2) and lysolecithin acyltransferase (EC 2.3.1.23). We previously found (2) that two of these drugs, WY14643 ([4-chloro-2,3-xili- dine)-2-pyrimidinylthiolacetic acid) and DH990 (2 - [3,5 - di - (t - butyl - 4 - hydroxy- phenyl)thio]hexanoic acid), affected sev- eral enzymatic activities associated with the formation of linoleic acid in safflower microsomes, namely acylthiokinase, acyl- transferases, and either the A12-oleate de- saturase itself or the electron carrier sys- tem that furnishes electrons from NADH to the enzyme. In order to obtain a more system; Hepes, 4-(2-hydroxyethyl)-l-piperazineeth- anesulfonic acid, BSA, bovine serum albumin; DTT, dithiothreitol. 45 9693-9861/34 $3.96 Copyright 8 1984 by Academic Press. Inc. All rights of reproduction in any form reserved.
Transcript
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 234, No. 1, October, pp. 45-54,1934
The Effect of Hypolipidemic Drugs on Plant Lipid Metabolism
JUAN SANCHEZ’ AND PAUL K. STUMPF2
Department of Biochemistry and Biophysics, University of California, Davis, Califbrnin 95616
Received February 22, 1934, and in revised form June 18, 1934
The effect of hypolipidemic drugs, WY14643 and DH990, on plant lipid metabolism has been studied. The total incorporation of [14C]acetate into lipids was inhibited by addition of both drugs to aged potato (Solarium tuberosum) tuber discs, spinach (Spinucia oleracea) leaves, and spinach chloroplasts, while the incorporation in Chbrella vulgaris cells was affected only by DH990. Moreover, DH990 inhibited the incorporation of 14C-labeled fatty acids into phosphatidylcholine and phosphatidyleth- anolamine of potato discs, and decreased the incorporation into phosphatidylglycerol of ChZoreZZu cells. DH990 inhibited the formation of polyunsaturated fatty acids in potato discs, Chlorella cells, and spinach leaves, whereas WY14643 had no effect on the formation of these fatty acids. Stearoyl-ACP desaturase from safflower (Carthumus tinctorius) seeds was very sensitive to both drugs, especially DH990, which completely blocked the activity at 2 mM levels. When safflower lysophospholipid acyltransferases were solubilized by detergent treatment, only DH990 inhibited the incorporation of [14C]oleoyl-CoA into lysophosphatidylcholine or lysophosphatidylethanolamine. Both drugs inhibited fatty acid synthesis from [‘4C]malonyl-CoA in the microsomal fraction from safflower seeds, but only DH990 inhibited FAS activity in the soluble fraction; both drugs inhibited severely the formation of stearic acid. Both acetyl-CoA carboxylase and acetyl-CoA synthetase were sensitive to both drugs. Q last Academic PM, I~~.
Hypolipidemic drugs have been exten- sively used to decrease the level of blood lipid in patients affected by diseases such as atherosclerosis, but the mode of action of these drugs remains to be elucidated. Recently (1) it was reported that some hypolipidemic drugs blocked the formation of PC3 in rat liver microsomes by inhib-
i Present address: ARC0 Plant Cell Research In- stitute, 6569 Trinity Court, Dublin, Calif. 94563.
* To whom correspondence should be addressed. a Abbreviations used: PC, phosphatidylcholine; PE,
iting both cholinephosphotransferase (EC 2.7.8.2) and lysolecithin acyltransferase (EC 2.3.1.23).
We previously found (2) that two of these drugs, WY14643 ([4-chloro-2,3-xili- dine)-2-pyrimidinylthiolacetic acid) and DH990 (2 - [3,5 - di - (t - butyl - 4 - hydroxy- phenyl)thio]hexanoic acid), affected sev- eral enzymatic activities associated with the formation of linoleic acid in safflower microsomes, namely acylthiokinase, acyl- transferases, and either the A12-oleate de- saturase itself or the electron carrier sys- tem that furnishes electrons from NADH to the enzyme. In order to obtain a more
45 9693-9861/34 $3.96 Copyright 8 1984 by Academic Press. Inc. All rights of reproduction in any form reserved.
46 SANCHEZ AND STUMPF
complete picture of the effect of these two drugs on lipid metabolism in plants, we have tested their effects, under both in vivo and in vitro conditions. The results of these experiments are reported in this paper.
MATERIALS AND METHODS
Maten’als. Potato (Solunum tuberoeum) tubers and spinach (Spinocia oleracea) leaves were purchased from the local market. Cultures of Chlorella vulgaris cells were obtained from Carolina Biological Supply Company. Developing safflower seeds (Carthamus tinctorius var. Portugal) were harvested from the field and stored at -20°C until used. [2-‘%]Acetic acid (60 mCi/mmol), [2-%]malonyl-CoA (60 mCi/ mmol), [1-‘%]stearic acid (50 mCi/mmol), and NaH’%Oa (30 mCi/mmol) were from Amersham Searle. [1-“C]Stearoyl-ACP was prepared according to Rock and Garwin (3). DH990 was obtained from Dr. J. Heeg, Merrell Dow Pharmaceuticals Inc., Indianapolis, Indiana, and WY14643 was a kind gift from Dr. W. J. Baumann, The Hormel Institute, Au&n, Minnesota. Silica sol (Percoll) and all the other reagents and enzymes were from Sigma, St. Louis, Missouri.
Preparation of tissues for in vivo exw’mats. Potato tubers were cut into discs (1 cm diameter X 1 mm) and aged in lo-’ M CaS04 for 16 h in the dark and with constant shaking. Chloreua cells were grown in a heterotrophic medium described by Harris and James (4) at room temperature, under light (66 pEs s-i m-z) at a photoperiod of 16 h light and 8 h dark. The culture was allowed to grow for 48 h after inoculation, and then the cell suspension was cen- trifuged at 1OOOg for 5 min. The pellet was resus- pended in 40 ml 0.1 M potassium phosphate, pH 7.2, and centrifuged at the same conditions. The washed pellet was resuspended in a small volume of the same phosphate buffer and immediately used in incubations.
Immature leaves were selected from spinach bunches and, after rinsing them thoroughly with distilled water, were cut into small slices, and aliquots were employed for incubations with the appropriate reaction mixtures.
Preparation of cell-free systems. Chloroplasts, pre- pared from immature spinach leaves as described by Nakatani and Barber (5), were resuspended in a small volume of 25 mM Hepes, pH 7.9, 0.33 M sorbitol, 2 mM EDTA, 0.15% BSA, and this suspension was purified through a continuous gradient of Percoll that had been prepared beforehand by centrifuging at 10,OOOg for 90 min in a mixture consisting of 25 ml Percoll and 25 ml 50 mM Hepes, pH 7.9, 0.66 M sorbitol, 4 mM EDTA, 0.3% BSA. After layering the suspension on the gradient, it was centrifuged at 10,000 g for 15 min. Intact chloroplasts, which equil-
ibrated near the bottom of the gradient, were care- fully recovered with a Pasteur pipet and immediately used in incubations.
Soluble and microsomal fractions were prepared from 5 g of developing safflower seeds after removing seed coats. Cotyledons were ground in 0.1 M potassium phosphate, pH 7.2, buffer and, after filtering through Miracloth, the brei was centrifuged at 18,OOOg for 20 min. The supernatant between the floating layer and pellet was then centrifuged at 105,000g for 1 h. The microsomal pellet was resuspended in the same buffer and, like the soluble fraction, immediately used in incubations.
The solubilization of the acyltransferase from safflower microsomes was carried out essentially as described by Moreau and Stumpf (6).
Ribulose-1,5-biphosphate carboxylase and acetyl- CoA carboxylase were assayed in a crude extract prepared from spinach leaf. About 1 g of deribbed spinach leaf was homogenized in a mortar and pestle with 3 ml 50 mM Tris-HCl, pH 8.0, 1 mM EDTA, 10 mM 2-mercaptoethanol. The brei was filtered through Miracloth, and 1 ml of the filtrate was passed through a small column of Sephadex G-25 equili- brated with the above buffer. The void volume was collected and immediately assayed.
Incubations Incorporation of [‘Cjacetate into lipids of aged potato discs and spinach leaves was examined by incubating 0.5 g tissue in 1.0 (potato) or 1.5 ml (spinach) 0.1 M potassium phosphate, pH 7.2, con- taining 2 &i [i4C]acetate (33 and 22 pM, respectively) and the indicated additions. In both cases, incubations were at room temperature for 3 h with constant shaking and, in the case of spinach leaves, under light (66 YES 5-l me*).
Chlorella cells, equivalent to 5 mg protein and 100 gg chlorophyll, were incubated with 2 &i [“Cjacetate (33 PM), in the presence or absence of hypolipidemic drug (2.5 mM), in a total volume of 1 ml. Incubations were carried out at the same conditions described above for spinach leaves for 2 h.
Spinach chloroplasts (20 pg chlorophyll) were in- cubated in 0.5 ml 25 mM Hepes, pH 7.9, 0.33 M sorbitol, 1 mM KrHPO,, 10 mM NaHCOs, 1 mM MgClz, containing 2 NCi [“Clacetate (67 pM) and, where indicated, WY14643 or DH990 (0.5 mM), CoA (0.5 mM), and ATP (2 mM). Incubations were carried out under the conditions described for spinach leaves for 1 h.
Soluble and microsomal fractions from safflower seeds were assayed for fatty acid synthesis in a reaction mixture containing 0.12 &i [“C]malonyl- CoA (2 PM), 0.5 mM NADH, 0.5 mM NADPH, 1 pM acetyl-CoA, 0.2 mg ACP, enzyme preparation equiv- alent to 2 mg protein, as well as the indicated concentration of drug, all in 1 ml 0.1 M potassium phosphate, pH 7.2, buffer. Incubations were carried out at room temperature, with constant shaking, for 1 (soluble fraction) or 4 h (microsomal fraction).
HYPOLIPIDEMIC DRUGS AND PLANT LIPID METABOLISM 47
TO test the effect of hypolipidemic drugs on the stearoyl-ACP desaturase, the soluble fraction from safflower seeds (1.5 mg protein) was incubated in a reaction mixture containing 10 mM NADPH, 2 mM DTT, 50 pg spinach ferredoxin, 0.1 IU ferre- doxin:NADP reductase, and approximately 0.1 MM [‘*Cjstearoyl-ACP (18,000 dpm) in 1 ml 0.1 M
phosphate, pH 7.2, buffer at room temperature for 30 min.
The effect of drugs on the solubilized acyltrans- ferase from safflower microsomes was tested by incubating the enzyme extract (120 pg protein) in a reaction mixture containing 1 pM [‘“Cjoleoyl-CoA (65,000 dpm) and, where stated, 50 pM lyso-PC or lyso-PE, 2 mM WY14643 or DH990, in 0.5 ml 0.1 M Tris, pH 7.9, buffer for 30 min at room temperature.
Acyl-CoA carboxylase was assayed according to Nikolau et al (7); ribulose BP carboxylase as de- scribed by Wishnick and Lane (8); glucose-6-P de- hydrogenase according to LShr and Waller (9); and acetyl-CoA synthetase as Frenkel and Kitchens (10).
Analyses. In the experiments carried out in viva with potato discs and spinach leaves, the substrate external to the tissue was removed at the end of incubation by rinsing the tissue three times with 2 ml 1% acetic acid. Lipids from potato slices were extracted according to Hara and Radin (ll), and those from spinach leaves as in Bligh and Dyer (12), using a Polytron homogenizer to grind the tissue. After partition, aliquots taken from the organic layer were submitted to transmethylation with 2.5% H2S01 in dry methanol-benzene (95:5), heating at 75°C for 2 h in screw-capped tubes. Methyl esters were extracted with petroleum ether, and the radio- activity was determined in aliquots (total incorpo- ration). 14C-Labeled fatty acid analyses were by radio-GLC (total fatty acids); polyunsaturated fatty acids were further identified by TLC on silica gel impregnated with 5% AgNOa and with petroleum ether-diethyl ether (91) as the mobile phase.
In those experiments in which Chbrella cells or cell-free preparations were used, aliquots were taken from the incubation mixture and submitted to sa- ponification in 1 M KOH for 30 min at 75°C. The resulting fatty acids were methylated for 20 min as described above, the methyl esters were extracted with petroleum ether, and the radioactivity was determined in suitable aliquots. 14C-Labeled fatty acid composition was determined by radio-GLC. To determine radioactivity associated with acylthioes- ters, the aqueous layer resulting from extraction of lipids was submitted to saponification and methyl- ation, and radioactivity determined in the fatty acid methyl ester extract was taken as incorporation into acyl-CoAs.
To extract lipids from Chlurella cells, 15 v01 of chloroform-methanol (2:l) was added to the suspen- sion at the end of the incubation, and the mixture was shaken overnight at room temperature; then
the partition was carried out as in (13). Lipids from chloroplasts were extracted according to (ll), and those from other cell-free systems as in (13).
Lipids from Chlorella and spinach leaves were separated by the TLC system described by Khan and Williams (14). In other cases, lipids were chro- matographed on silica gel G plates, using petroleum ether-diethyl ether-acetic acid (4060~2) as the first solvent, followed by chloroform-methanol-water (65254) as the second solvent developed to the fatty acid band (2). In all cases, radioactive bands were located with a Packard radiochromatogram scanner (Model 7201) and scraped, and the silica gel was transferred to counting vials containing 1 ml meth- anol-water (1:l); then radioactivity was counted with 10 ml PCS (Amersham)-xylene (2:l).
Protein was determined according to Lowry et al (15), in the presence of 0.1% sodium deoxycholate, with BSA as standards, and chlorophyll was accord- ing to Bruinsma (16).
RESULTS
Eflect of Hypolipidemic Drugs on the Incorporation of [14CJAcetute in Viva
Aged potato slices incorporated [‘“Cl- acetate in both saturated (from Cl6 to C,) and unsaturated (18:l and l&2) fatty acids (17). It was found that both hypolipidemic drugs inhibited the total incorporation of [14C]acetate into lipids, with DH990 a more effective inhibitor than WY14643 (Table I). PC, which was the most heavily labeled lipid in the control, and PE were found to be very susceptible to the action of the drugs, in agreement with the results ob- tained previously by using [‘%]oleate with the same tissue (2).
The nature of the fatty acids formed from [14C]acetate was also found to be affected by addition of hypolipidemic drugs (Table II). The main effect was observed in the formation of very long chain fatty acids (C&-C&, which ac- counted for almost 40% of the total radio- active fatty acids in the control, and was inhibited severely by WY14643 or com- pletely by DH990. The formation of 18:2 was also markedly inhibited by DH990.
Intact C vz&a& cells were then ex- amined in terms of incorporation of [14C]acetate into polyunsaturated fatty ac- ids and thence into chloroplastic lipids. No significant effect was observed by
48 ShCHEZ AND STUMPF
TABLE I
EFFECT OF HYPOLIPIDEMIC DRUGS ON THE INCORPORATION OF [“‘CIACETATE IN LIPIDS BY AGED POTATO SLICES
Distribution of radioactivity (nmol [‘?]acetate) Total incorporation Control
Treatment (nmol [i4C]acetate + SD) (%) PC PE Other PLS FFA NL
Control 2.13 k 0.13 100 0.85 0.30 0.30 0.07 0.61 (40.0)" (14.1) (14.1) (3.3) (28.6)
’ Figures in parentheses express percentages of the total radioactivity incorporated into each fraction.
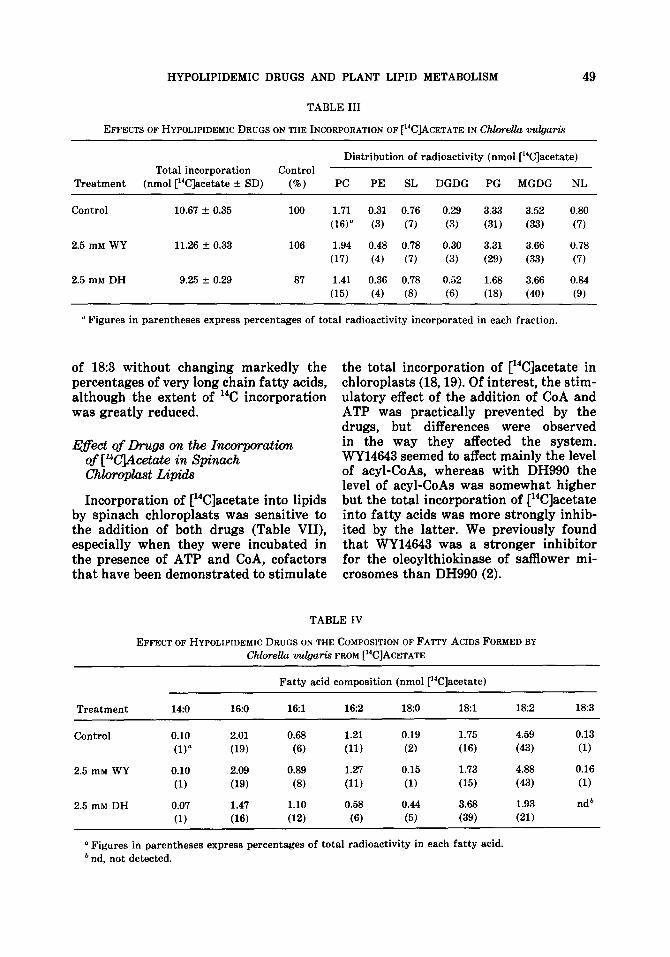
WY14643 in [14C]acetate incorporation into lipids (Table III). DH990 appeared to effect a small decrease of the total incorporation and an inhibition of radiolabel incorpo- rated into PG, the only lipid that was found to be affected by the drug. No effect was observed by addition of WY14643 on the fatty acids formed from [14C]acetate (Table IV). However, DH990 significantly inhibited the formation of polyunsatu- rated fatty acids (162, 18:2, and l&3), with the concomitant accumulation of radioactivity in monounsaturated fatty acids.
Experiments carried out with Chlorella were difficult to interpret, since the ar- gument could be made that the drugs did not readily enter the cell.
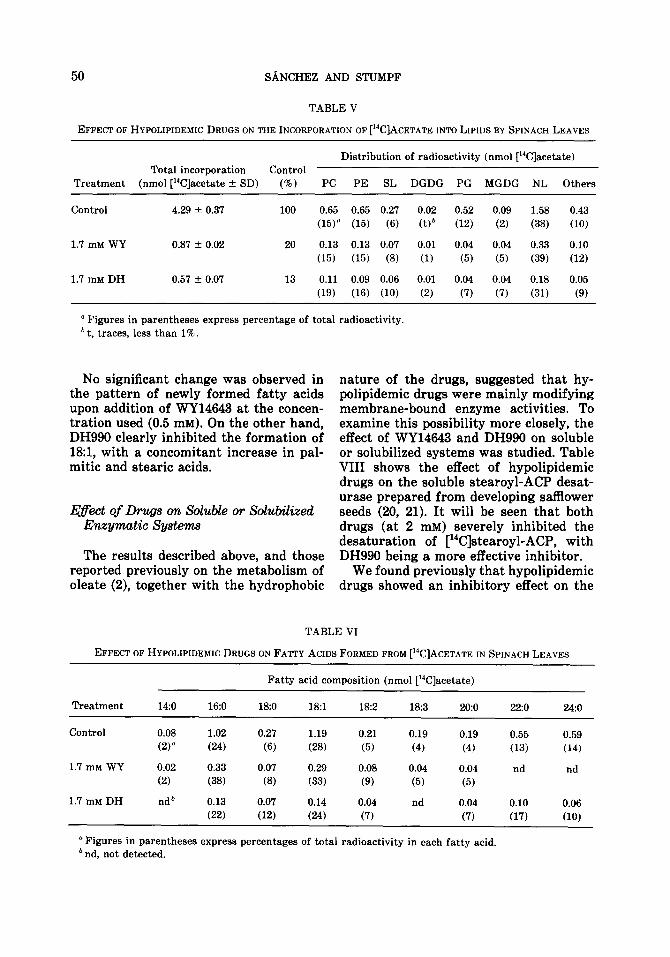
Another type of photosynthetic tissue, namely spinach leaves, was also employed. The results indicated that both drugs se- verely inhibited total incorporation of [14C]acetate (Table V) and, in agreement with the results obtained with ChloreUa (Table III), the acylation of PG as well as other phospholipids was found to be espe- cially susceptible to the addition of drugs.
Analyses of radioactive fatty acids formed in spinach leaves (Table VI) showed that each drug exerted a different effect; WY14643 inhibited the formation of very long chain fatty acids, which ac- counted for more than 30% in the con- trol, without altering the percentages of polyunsaturated fatty acids significantly, whereas DH990 inhibited the formation
TABLE II
EFFECT OF HYPOLIPIDEMIC DRUGS ON THE INCORPORATION OF [%]ACETATE INTO FATTY ACIDS BY AGED POTATO SLICES
Fatty acid composition (nmol [?]acetate)
Treatment 14:o 16:0 l&O 131 18:2 20:o 220 24:o
Control
2.5 mM WY
2.5 mM DH
0.01 (t)”
ndb
0.03 (4)
0.19 (9)
0.24 (17) 0.11 (14)
0.33 (15) 0.12
(9) 0.15 (19)
0.38 (18) 0.38 (2’0 0.41 (52)
0.42 (20) 0.61 (44) 0.09 (11)
0.17 (8)
0.04 (3)
t:,
0.34 W-3 nd
nd
0.30 (14) nd
nd
a Figures in parentheses express percentages of total radioactivity; t, traces, less than 1%. * nd, not detected.
49 HYPOLIPIDEMIC DRUGS AND PLANT LIPID METABOLISM
TABLE III
EFFEIXS OF HYPOLIPIDEMIC DRUGS ON THE INCORPORATION OF [%]ACETATE IN Chlorella vulgaris
Distribution of radioactivity (nmol [%]acetate) Total incorporation Control
Treatment (nmoi [i4C]acetate f SD) (o/o) PC PE SL DGDG PG MGDG NL
a Figures in parentheses express percentages of total radioactivity incorporated in each fraction.
of l&3 without changing markedly the percentages of very long chain fatty acids, although the extent of 14C incorporation was greatly reduced.
Effect of Llrugs cm the Inmrporatim of [V&etate in Spinach chloroplast Lipid-!3
Incorporation of [‘4C]acetate into lipids by spinach chloroplasts was sensitive to the addition of both drugs (Table VII), especially when they were incubated in the presence of ATP and CoA, cofactors that have been demonstrated to stimulate
the total incorporation of [i4C]acetate in chloroplasts (l&19). Of interest, the stim- ulatory effect of the addition of CoA and ATP was practically prevented by the drugs, but differences were observed in the way they affected the system. WY14643 seemed to affect mainly the level of acyl-CoAs, whereas with DH990 the level of acyl-CoAs was somewhat higher but the total incorporation of [14C]acetate into fatty acids was more strongly inhib- ited by the latter. We previously found that WY14643 was a stronger inhibitor for the oleoylthiokinase of safflower mi- crosomes than DH990 (2).
TABLE IV
EFFECT OF HYPOLIPIDEMIC DRUGS ON THE COMPOSITION OF FATTY ACIDS FORMED BY ChloreUa vulgaris FROM [‘%]ACETATE
a Figures in parentheses express percentage of total radioactivity. b t, traces, less than 1%.
No significant change was observed in the pattern of newly formed fatty acids upon addition of WY14643 at the concen- tration used (0.5 mM). On the other hand, DH990 clearly inhibited the formation of l&l, with a concomitant increase in pal- mitic and stearic acids.
Efect of Drugs cm Soluble OT Solubilized Enzymatic Systems
The results described above, and those reported previously on the metabolism of oleate (2), together with the hydrophobic
nature of the drugs, suggested that hy- polipidemic drugs were mainly modifying membrane-bound enzyme activities. To examine this possibility more closely, the effect of WY14643 and DH990 on soluble or solubilized systems was studied. Table VIII shows the effect of hypolipidemic drugs on the soluble stearoyl-ACP desat- urase prepared from developing safflower seeds (20, 21). It will be seen that both drugs (at 2 mM) severely inhibited the desaturation of [‘4C]stearoyl-ACP, with DH990 being a more effective inhibitor.
We found previously that hypolipidemic drugs showed an inhibitory effect on the
TABLE VI
EFFECT OF HYPOLIPIDEMIC DRUGS ON FATTY ACIDS FORMED FROM [‘“C~ACETATE IN SPINACH LEAVES
Fatty acid composition (nmol [“Clacetate)
Treatment
Control
1.7 rnM WY
1.7 mM DH
14:o
0.08 GY
0.02 (2)
ndb
l&O 18:o
1.02 0.27 (24) (6)
0.33 0.07 (38) (8)
0.13 0.07 (22) w
18:l
1.19 (28)
0.29 (33)
0.14 (24)
18:2
0.21 (5)
0.08 (9)
0.04 (7)
18:3
0.19 (4)
0.04 (5)
nd
20:o
0.19 (4)
0.04 (5)
0.04 (7)
22:o
0.55 (13)
nd
0.10 (17)
24:0
0.59 (14)
nd
0.06 (10)
a Figures in parentheses express percentages of total radioactivity in each fatty acid. b nd, not detected.
HYPOLIPIDEMIC DRUGS AND PLANT LIPID METABOLISM 51
TABLE VII
EFFECT OF HYPOLIPIDEMIC DRUGS ON THE INCORPORATION OF [%]ACETATE
INTO LIPIDS BY SPINACH CHLOROPLASTS
Cofactors* Drug
Fatty acid composition (%) Total incorporation Control Incorporation into (nmol [“Clacetate) @) acyl-CoAs” (W) 120 14:o 160 18:o 131
’ Incorporation into acyl-CoAs expresses the radioactivity found in fatty acid methyl esters, prepared by saponification and methylation of the aqueous layer remaining after extraction and partition (see Materials and Methods).
b Where indicated, 0.5 mM CoA and 2.0 mM ATP were present. ‘t, traces, less than 1%
incorporation of [14C]oleoyl-CoA into PLs in safflower microsomes (2). It was there- fore of interest to determine if the drugs exerted the same effects on a solubilized acyl-CoA transferase system (6). As shown in Table IX, only DH990 inhibited the incorporation of [14C]oleoyl-CoA into both PC and PE, although the extent of the effect was not as marked as that found when microsomal preparations were em- ployed (2). WY14643, on the other hand, did not inhibit the incorporation into PC, but actually stimulated it, in agreement with the corresponding experiment carried out with safflower microsomes (2). On the other hand, incorporation into PE, in the presence of lyso-PE, was not inhibited by WY14643, in marked contrast with the
TABLE VIII
EFFECT OF DRUGS ON THE STEAROYL-ACP DESATURASE FROMSAFFLOWERSEEDS
Note. Figures are means of three experiments f SD.
analogous experiment carried out with the microsomal fraction (2).
It was of interest to determine if hy- polipidemic drugs affected the fatty acid synthesis from [14C]malonyl-CoA in both the soluble and microsomal fractions from safflower seeds. As shown in Table X, the soluble fraction was more active in incor- porating [14C]malonyl-CoA into fatty acids than the microsomal fraction, and fatty acid patterns were also different. Both drugs inhibited more markedly the micro- somal elongation system. Interestingly, WY14643, which did not affect the total incorporation of [‘4C]malonyl-CoA in the soluble fraction, induced a remarkable change in the fatty acid pattern by inhib- iting the formation of [14C]stearate. This result would suggest that this drug may inhibit @-ketoacyl-ACP synthetase II (22). The same effect on the fatty acid pattern was observed by addition of DH990, but in this case the total incorporation, as well as the formation of l&l, were af- fected too.
Finally, Table XI shows the effect of hypolipidemic drugs on the activity of a number of soluble enzymes, including two that are critical in lipid metabolism, ace- tyl-CoA synthetase and acetyl-CoA car- boxylase. Interestingly, DH990 markedly inhibited acetyl-CoA carboxylase, and also acetyl-CoA synthetase; WY14643 inhibited
52 SANCHEZ AND STUMPF
TABLE IX
EFFECT OF HYPOLIPIDEMIC DRUGS ON THE SOLUBILIZED ACYL-CoA:Luso-PL ACYLTRANSFERASE FROM SAFFLOWER MICROSOMES
the same activities but to a lesser extent. These results fit in well with the inhibition of total incorporation of [14C]acetate in potato discs (Table I), spinach leaves (Ta- ble V), and spinach chloroplasts (Table VII). On the other hand, ribulose bisphos- phate carboxylase was also inhibited by both drugs, whereas glucose-&phosphate dehydrogenase was not affected.
DISCUSSION
The results described in this paper, together with those reported previously
(2), demonstrate that hypolipidemic drugs, WY14643 and DH990, are capable of per- turbing plant lipid metabolism at a num- ber of enzymatic sites. On the basis of the highly hydrophobic structures of these drugs and their great affinity for the microsomal membranes (23), it was thought that hypolipidemic drugs mainly affected membrane-bound enzymes. How- ever, the results shown here indicate that this interpretation must be broadened.
Results from the in viva experiments carried out with different tissues (Tables I-VI), together with those others obtained from spinach chloroplasts (Table VII),
TABLE X
EFFECT OF HYPOLIPIDEMIC DRUGS ON THE INCORPORATION OF [%]MALONYL-COA INTO FATTY ACID IN THE SOLUBLE AND MICROSOMAL FRACTIONS FROM MATURING SAFFLOWER SEEDS
Fraction”
Total incorporation (nmol Fatty acid composition (W) [“C]maIonyl-CoA/mg Control
Microsome Control 0.06 f 0.01 100 1 2 67 22 t 8 2mMwY 0.04 + 0.01 67 not determined 2mMDH 0.03 AI 0.00 54 not determined
’ The soluble fraction was incubated for 1 h, the microsomal fraction for 4 h. b nd, not detected. ’ t, traces, less than 1%.
HYPOLIPIDEMIC DRUGS AND PLANT LIPID METABOLISM 53
TABLE XI
EFFECT OF HYPOLIPIDEMIC DRUGS ON DIFFERENT ENZYMES
Relative activity (% of control)
Enzyme WY14643” DH990 ”
Acetyl-CoA synthetase* Glucose-g-phosphate
dehydrogenase” Ribulose biphosphate
carboxylase” Acetyl-CoA carboxylase”
71 67
100 100
53 41 67 26
“Drugs were tested at 1 mM concentration. * Purified enzymes purchased from Sigma. “Determined on an extract from spinach leaves
(see Materials and Methods) by Dr. B. J. Nikolau.
clearly indicated that total incorporation of [14C]acetate into fatty acids and complex lipids was inhibited by both drugs. It was also found that both drugs inhibited ace- tyl-CoA synthetase and acetyl-CoA car- boxylase (Table XI), two key enzymes for the utilization of [14C]acetate for fatty acid synthesis that have been demon- strated to be soluble (7, 24-26), and this finding in part explains the decrease in total incorporation of [14C]acetate in those experiments where this precursor was employed. However, analyses of the in- cubation products from those experiments showed that those enzymatic activities that have been reported to be associated with membranes underwent the most marked inhibition:incorporation into PC, PE, and PG (27), and formation of poly- unsaturated (28) and very long chain fatty acids (29), as well as acyl-CoAs (30).
In summary, it would appear that hy- polipidemic drugs, WY14643 and DH990, have wide inhibitory effects on the plant lipid metabolism. Both drugs were found to affect both membrane-bound and sol- uble enzymatic activities, although the first seem to be more sensitive to the inhibition. The results reported here in- dicate that these drugs may be useful as probes for the manipulation of the lipid metabolism in plants.
However, it is unusual that these drugs inhibit such a large number of enzymes and activities, implying little specificity in their mode of action. A commonality of inhibitory effects does not appear to occur, making it difficult to pinpoint the site of action of these drugs in plant lipid metabolism. Further experiments are needed in order to define more precisely the mode of action of these two interesting drugs.
ACKNOWLEDGMENTS
However, although no inhibition was observed in the formation of l&l in in viva experiments (actually, an accumula- tion of radioactivity in this fatty acid was usually observed in the presence of drugs), in isolated chloroplasts a clear inhibition was consistently noted by these drugs (Table VII); moreover, when safflower stearoyl-ACP desaturase activity was challenged with these drugs, the inhibitory effect of both drugs was clearly demon-
Dr. J. SLnchez thanks the Fundacion Juan March (Spain) for his travel grant. We also thank Dr. N. W. Lem for the preparation of [‘4C]stearoyl-ACP, Dr. B. J. Nikolau for the determination of enzyme activities, and Dr. V. P. Agrawal for useful discussion. We are indebted to Professor W. Baumann and Dr. J. Heeg for providing us with the hypolipidemic drugs, and to Ms. Billie Gabriel for the preparation of the manuscript.
REFERENCES
1. PARTHASARATHY, S., KRITCHEVSKY, D., AND BAU- MANN, W. J. (1982) Proc Natl. Acad Sci USA 79, 68904893.
strated (Table VIII). Furthermore, the formation of stearate, which has been demonstrated to be specifically formed from palmitoyl-ACP in the soluble frac- tion from safflower seeds (22, 31), was markedly affected by these hypolipidemic drugs (Table X).
The effects of DH990 on the solubilized acyl-CoA:lyso-PL acyltransferase (Table IX) are in the line of the results we previously found with the same activities in safflower microsomes (2) but, once again, the inhibitory effect was smaller with the solubilized preparation.
54 SANCHEZ AND STUMPF
2. SANCHEZ, J., AND STUMPF, P. K. (1984) Arch. B&hem. Biophys. 228,185-196.
3. ROCK, C. O., AND GARWIN, J. L. (1979) J. BioL Chem. 254, 7123-7128.
4. HARRIS, R. V., AND JAMES, A. T. (1965) B&him Biophys. Acta 106,456-464.
5. NAKATANI, H. Y., AND BARBER, J. (1977) Biochim Biophys. Acta 461, 510-512.
6. MOREAU, R. A., AND STUMPF, P. K. (1982) Plant Physid 69, 1293-1297.
7. NIKOLAU, B. J., HAWKE, J. C., AND SLACK, C. R. (1981) Arch. Biochem. Biophga 211, 605-612.
8. WISHNICK, M., AND LANE, M. D. (1971) in Methods in Enzymology (San Pietro, A., ed.), Vol. 23, pp. 570-577, Academic Press, New York.
9. LBHR, G. W., AND WALLER, H. D. (1965) in Methods of Enzymatic Analysis (Bergmeger, H. V., ed.), pp. 744-751, Academic Press, New York.
10. FRENKEL, E. P., AND KITCHENS, R. L. (1981) in Methods in Enzymology (Lowenstein, J. W., ed.), Vol. 71, pp. 317-324, Academic Press, New York.
11. HARA, A., AND RADIN, N. S. (1978) And Biochem SO, 420-426.
12. BLIGH, E. G., AND DYER, W. J. (1959) Caned J. Biochem Physiol 37,911-917.
13. FOLCH, J., LEES, M., AND SLOANE-STANLEY, G. H. (1957) J. Biol Chem 226, 497-509.
14. KHAN, M., AND WILLIAMS, J. P. (1977) .I. Chw wzatogr. 140,179-185.
15. LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL, R. J. (1951) J. BioL Chem 193. 265-275.
16. BRUINSMA, J. (1961) Biochim Biophys. Acta 52, 576-578.
17. WILLEMOT, C., AND STUMPF, P. K. (1967) Canad J. Bat. 45,579-584.
18. SANCHEZ, J., AND MANCHA, M. (1981) Plunta 153, 519-523.
19. ROUGHAN, P. G., AND SLACK, C. R. (1981) FEBS L&t. 135, 182-186.
20. JAWORSKI, J. G., AND STIJMPF, P. K. (1974) Arch, Biochem Biophys. 162,158-165.
21. MCKEON, T. A., AND STUMPF, P. K. (1982) .I. Biol Chem 257, 12141-12147.
22. SHIMAKATA, T., AND STUMPF, P. K. (1982) Proc Nat1 Acud Sci USA 79, 5808-5812.
23. PARTHASARATHY, S., ABDEL-RAHMAN, M. M. A., AND BAUMANN, W. J. (1981) Biochim Biophys. Acta 665, 345-351.
24. HUANG, K. P., AND STUMPF, P. K. (1970) Arch. Biochem Biophys. 140,158-173.
25. KUHN, D. N., KNAUF, M. J., AND STUMPF, P. K. (1981) Arch Biochem Biophys. 209, 441-450.
26. ROUGHAN, P. G., AND SLACK, C. R. (1977) Biochm. J. 162,457-459.
27. MOORE, T. S. JR. (1982) Annzc, Rev. Plant PhysioL 33,235-259.
28. ROUGHAN, P. G., AND SLACK, C. R. (1982) Annu. Rev. Plant PhysioL 33, 97-132.
29. HARWOOD, J. L. (1979) Prog. Lipid Res. 18, 55- 86.
30. JOYARD, J., AND STUMPF, P. K. (1981) Plant PhysioL 67, 250-256.
31. JAWORSKI, J. G., GOLDSCHMIDT, E. E., AND STIJMPF, P. K. (1974) Arch. Biochem Biuphys. 163,769- 776.