BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. From baby birds to feathered dinosaurs: incipient wings and the evolution of flight Author(s): Ashley M. Heers , Kenneth P. Dial , and Bret W. Tobalske Source: Paleobiology, 40(3):459-476. 2014. Published By: The Paleontological Society DOI: http://dx.doi.org/10.1666/13057 URL: http://www.bioone.org/doi/full/10.1666/13057 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/ terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
Transcript
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
From baby birds to feathered dinosaurs: incipient wings andthe evolution of flightAuthor(s): Ashley M. Heers , Kenneth P. Dial , and Bret W. TobalskeSource: Paleobiology, 40(3):459-476. 2014.Published By: The Paleontological SocietyDOI: http://dx.doi.org/10.1666/13057URL: http://www.bioone.org/doi/full/10.1666/13057
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.
From baby birds to feathered dinosaurs: incipient wings and theevolution of flight
Ashley M. Heers, Kenneth P. Dial, and Bret W. Tobalske
Abstract.—Reconstructing the tree of life requires deciphering major evolutionary transformations andthe functional capacities of fossils with ‘‘transitional’’ morphologies. Some of the most iconic, well-studied fossils with transitional features are theropod dinosaurs, whose skeletons and featheredforelimbs record the origin and evolution of bird flight. However, in spite of over a century ofdiscussion, the functions of forelimb feathers during the evolution of flight remain enigmatic. Bothaerodynamic and non-aerodynamic roles have been proposed, but few of the form-functionrelationships assumed by these scenarios have been tested. Here, we use the developing wings of atypical extant ground bird (Chukar Partridge) as possible analogues/homologues of historical wingforms to provide the first empirical evaluation of aerodynamic potential in flapping theropod‘‘protowings.’’ Immature ground birds with underdeveloped, rudimentary wings generate usefulaerodynamic forces for a variety of locomotor tasks. Feather development in these birds resemblesfeather evolution in theropod dinosaurs, and reveals a predictable relationship between wingmorphology and aerodynamic performance that can be used to infer performance in extinct theropods.By spinning an ontogenetic series of spread-wing preparations on a rotating propeller apparatus acrossa range of flow conditions and measuring aerodynamic force, we explored how changes in wing size,feather structure, and angular velocity might have affected aerodynamic performance in dinosaurschoosing to flap their incipient wings. At slow angular velocities, wings produced aerodynamic forcessimilar in magnitude to those produced by immature birds during behaviors like wing-assisted inclinerunning. At fast angular velocities, wings produced forces sufficient to support body weight duringflight. These findings provide a quantitative, biologically relevant bracket for theropod performanceand suggest that protowings could have provided useful aerodynamic function early in maniraptoranhistory, with improvements in aerodynamic performance attending the evolution of larger wings, moreeffective feather morphologies, and faster angular velocities.
Ashley M. Heers,* Kenneth P. Dial, and Bret W. Tobalske. Division of Biological Sciences, University ofMontana, Missoula, Montana 59812, U.S.A. E-mail: [email protected]. *Present address: ComparativeBiomedical Sciences, Royal Veterinary College, Hatfield, Hertfordshire AL9 7TA, U.K.
Accepted: 17 January 2014Published online: 7 May 2014Supplemental material deposited at Dryad: doi: 10.5061/dryad.7pg3d
Introduction
Reconstructing the function(s) of forelimbfeathers in theropod dinosaurs is key tounderstanding the origin and evolution ofbirds and bird flight. Since the discovery ofArchaeopteryx in 1861, a number of featheredtheropods have been described, particularlyfrom China (Norell and Xu 2005; Xu 2006).These fossils demonstrate that both feathersand winglike structures were present in non-avian dinosaurs, in arrangements that initiallydiffered from those of extant flight-capableadult birds and that became more similar tothe extant condition throughout theropodevolution (Prum and Brush 2002; Norell andXu 2005; Xu and Guo 2009; Heers and Dial
2012). Available evidence indicates that feath-ers debuted in basal theropods as filamentousor downlike ‘‘protofeathers’’ (although fila-mentous homologues of feathers may havearisen earlier) (Norell and Xu 2005; Zheng etal. 2009; Clarke 2013). Downlike feathers wereoften complemented in maniraptorans bypennaceous (vaned) feathers, which first ap-peared as ‘‘fans’’ on the distal tail and as small‘‘protowings’’ with symmetrically vanedfeathers on the distal forelimb. Pennaceousfeathers became more widely distributed andmore asymmetric in many paravians, particu-larly in avialans, forming larger and morebirdlike wings (here, the pennaceous forelimbfeathers of extinct theropods are collectively
� 2014 The Paleontological Society. All rights reserved. 0094-8373/14/4003-0003/$1.00
Paleobiology, 40(3), 2014, pp. 459–476DOI: 10.1666/13057
referred to as ‘‘protowings’’ or ‘‘wings,’’ not toimply an aerodynamic function but merely toindicate degree of resemblance to the wings ofextant birds). Interpreting this protowing-to-wing progression is central to reconstructingthe evolutionary acquisition of bird flight.
However, the functions of protowings re-main enigmatic. Many skeletal features asso-ciated with powered flight in extant adultbirds (e.g., robust pectoral girdle, channelizedwrist) are conspicuously absent in non-aviantheropods and basal avialans (Ostrom 1976;Vazquez 1992; Heers and Dial 2012). Inaddition, the incipient wings of extinct thero-pods and the wings of extant flight-capableadult birds differ substantially in relative sizeand feather morphology; flapping velocityalso presumably differs because adult birds,but not non-avian theropods and basal avia-lans, have large ossified keels for the attach-ment of powerful flight muscles (Prum andBrush 2002; Norell and Xu 2005; Makovickyand Zanno 2011; Wang et al. 2011). It is oftenassumed that such differences would haveprecluded early winged theropods from pro-ducing the aerodynamic forces necessary forpowered (flapping) flight. For example, asym-metric primary feathers with thick rachisesand tightly interlocking barbules are thoughtto help stabilize primary feathers againstoncoming airflow (Norberg 1985), preventexcessive deformation (Nudds and Dyke2010), and reduce feather permeability (Mullerand Patone 1998), whereas large wings and alarge keel (with large flight muscles) arethought to be necessary for a fast and effectiveflight stroke. Extinct theropods with incipientwings lacked most or all of these hallmarks ofadvanced flight capacity: basal maniraptorans(e.g., Caudipteryx) and some basal paravians(e.g., Anchiornis) had relatively symmetric andpotentially more permeable feathers (Prumand Brush 2002; Xu and Guo 2009), and evenavialans like Archaeopteryx and Confusciusornishad relatively thin rachises compared toextant adult birds (Nudds and Dyke 2010).Non-avian theropods and basal avialans arethus often assumed to have used theirfeathered forelimbs for non-aerodynamic pur-poses (such as display) and/or non-poweredgliding (Heers and Dial 2012: Table 1).
Our understanding of the functional rela-tionships between wing and feather morphol-ogy and aerodynamic performance is verylimited, however. Several studies, for example,highlight how many of the assumptionsunderlying traditional views (e.g., Feducciaand Tordoff 1979; Speakman and Thomson1994, 1995) of feather function are not wellunderstood and are incongruent with agrowing body of empirical evidence from liveanimals. For instance, it has recently beensuggested that symmetric feathers would havebeen useful during drag-based aerial behav-iors (Garner et al. 1999) or gliding (Dyke et al.2013), and that paravians like Anchiornis andArchaeopteryx had layered wing feathers thatmight have compensated for the ‘‘weak’’rachises of individual feathers (Longrich etal. 2012). Recent and ongoing work withextant animals shows that juvenile birds withsmall muscles and dinosaur-like protowingsand skeletons (Heers and Dial 2012: Fig. 2)negotiate complex environments through flap-ping behaviors such as wing-assisted inclinerunning (WAIR), ‘‘steaming’’ across the sur-face of water, and slowing aerial descents(Dial 2003; Dial et al. 2006; Jackson et al. 2009;Dial and Jackson 2011; Dial and Carrier 2012).Though not yet capable of level flight, thesejuveniles generate small amounts of aerody-namic force that improve locomotor perfor-mance and increase throughout ontogeny,culminating in full flight capacity (Dial 2003;Tobalske and Dial 2007; Heers et al. 2011).Recent analyses of both extant and extincttheropods thus call into question many as-sumptions about feather evolution. Thoughwe often assume that protowings were notused by extinct theropods for flapping loco-motion, developing birds demonstrate thatincipient flight structures are aerodynamicallyfunctional, for a variety of flapping locomotorbehaviors.
Flapping locomotor behaviors like WAIRand steaming are widespread across extantbirds (Tinamiformes, Anserogalliformes, Apo-diformes, Columbiformes, Procellariformes,Strigiformes, Passeriformes [http://www.youtube.com/user/UMflightlab; Dial 2011])and appear to be crucially important tojuveniles with incipient wings. These behav-
460 ASHLEY M. HEERS ET AL.
iors involve the cooperative (simultaneous)use of wings and legs, and thereby act as adevelopmental bridge between leg-based ter-restrial (or aquatic) locomotion and wing-based aerial locomotion. For example, imma-ture, flight-incapable ground birds rely onWAIR to ascend steep slopes and reachrefugia. Aerodynamic performance improvesthroughout ontogeny and allows developingbirds to flap-run up progressively steeperslopes and eventually fly (Dial 2003; Tobalskeand Dial 2007; Heers et al. 2011). By usingtheir forelimbs and hind limbs cooperatively,juvenile ground birds thus supplement theirincipient wings with their legs until the wingscan fully support body weight during flight.Similarly, flight-incapable ducklings engagetheir legs and developing wings cooperativelyto paddle rapidly across water and escapefrom terrestrial predators (steaming) (Livezeyand Humphrey 1983; Aigeldinger and Fish1995; Dial and Carrier 2012). Immature,‘‘branching’’ owls sometimes flap their devel-oping wings to ‘‘walk’’ up tree trunks andreach the safety of elevated branches (Marks etal. 1999; Marks 1982, 1986), and peachicks(Pavo cristatus) and young partridges (Alectorischukar) improve jumping performance byflapping their wings and reducing decelera-tion with small amounts of aerodynamic force(Heers and Dial 2013). Even flight-capablebirds, and birds that become highly leg-dependent (e.g., brush turkeys [Alectura lath-ami]) or flightless (some steamer ducks [Ta-chyeres]) as adults, engage their wings and legscooperatively to locomote (Livezey and Hum-phrey 1986; Dial and Jackson 2011). Wing-legcooperation is therefore common and wide-spread among birds, and facilitates the tran-sition from obligately bipedal juvenile toflight-capable adult in species with a diversearray of wing-leg morphologies and life-historystrategies. Protowings capable of producingeven small amounts of force enhance legperformance (incline-running, jumping, swim-ming), with incremental improvements inaerodynamic performance allowing juvenilesto flap-run up steeper slopes, jump higher, andeventually fly.
Given that extant juvenile birds beginproducing aerodynamic forces with veryT
AB
LE
1.B
od
ym
asse
s,ef
fect
ive
fore
lim
ble
ng
ths,
feat
her
mo
rph
olo
gie
san
dch
uk
arm
od
els
use
dto
eval
uat
eth
eae
rod
yn
amic
po
ten
tial
of
ther
op
od
pro
tow
ing
s.S
tage
iis
ph
ylo
gen
etic
ally
mo
stb
asal
and
stag
eii
iis
mo
std
eriv
ed,
wit
hso
me
ov
erla
pb
etw
een
stag
es(f
or
exam
ple
,so
me
dei
no
ny
cho
sau
rsar
est
age
iian
dso
me
are
stag
eii
i).
Th
ero
po
dm
asse
san
dfo
reli
mb
len
gth
sb
ased
on
pu
bli
shed
esti
mat
esan
dm
easu
rem
ents
(Bu
rger
san
dC
hia
pp
e19
99;S
eeb
ach
er20
01;H
olt
z20
07;T
her
rien
and
Hen
der
son
2007
;Tu
rner
etal
.20
07;
Eri
ckso
net
al.
2009
;X
uet
al.
2009
)(S
up
ple
men
tary
Tab
le1)
.
Sta
ge
(Fig
.1)
Th
ero
po
dex
amp
les
Bo
dy
mas
s(k
g)
Eff
ecti
ve
fore
lim
ble
ng
th(c
m)
Mo
rph
olo
gy
of
rem
ige
feat
her
s(i
fp
rese
nt)
On
tog
enet
icst
age
use
dfo
rm
od
elin
g
Sta
gei
Bas
alm
anir
apto
ran
sC
audi
pter
yx,
Sim
ilic
audi
pter
yxS
TM
22-6
614
–22
~S
ym
met
ric,
rela
tiv
ely
sho
rtfe
ath
ers,
mo
rep
ron
ou
nce
do
nd
ista
lfo
reli
mb
8,10
dp
hS
inos
auro
pter
yx,*
Sim
ilic
audi
pter
yxS
TM
4-1
0.55
–3.5
10
Sta
geii
Bas
alp
arav
ian
sN
oex
amp
les,
yet
0.7–
122
~S
ym
met
ric
tow
eak
lyas
ym
met
ric
feat
her
s10
,20
dp
hA
nch
iorn
is0.
1–0.
310
–14
Sta
geii
iB
asal
par
avia
ns,
bas
alav
iala
ns
Mic
rora
ptor
,Je
holo
rnis
0.7–
122
Asy
mm
etri
c,re
lati
vel
ylo
ng
feat
her
s20
,49
dp
hA
rcha
eopt
eryx
(e.g
.,E
ich
stat
t,M
un
ich
)0.
1–0.
310
–14
*H
ere,
we
foll
ow
(Mak
ov
ick
yan
dZ
ann
o20
11)
and
con
sid
erco
mp
sog
nat
hid
sto
be
bas
alm
anir
apto
ran
s.N
ote
that
Sin
osau
ropt
eryx
do
esn
ot
hav
ep
enn
aceo
us
fore
lim
bfe
ath
ers
(at
leas
tn
ot
pre
serv
ed),
and
isli
sted
on
lyto
pro
vid
ean
exam
ple
of
bo
dy
size
.
INCIPIENT WINGS AND THE EVOLUTION OF AVIAN FLIGHT 461
rudimentary flight apparatuses, protowings ofextinct theropods similarly might have beenaerodynamically functional. In many (most?)bird species, ontogenetic improvements in
locomotor capacity are attended by changesin feather and skeletal morphology thatresemble feather and skeletal evolution intheropod dinosaurs (Dial et al. 2006; Heers et
FIGURE 1. Incipient wings in the theropod-avian lineage. Ontogenetic trajectories in wing size and feather structure bearmany similarities to evolutionary trajectories observed among theropod dinosaurs. Younger birds and more basalmaniraptorans tend to have small wings with relatively symmetric feathers, whereas older birds and more derivedmaniraptorans tend to have larger wings with more asymmetric primaries. The feathers of younger birds also are more‘‘open’’ and more transmissive than those of older counterparts (Heers et al. 2011; Dial et al. 2012), as hypothesized forearly feathered theropods (Prum and Brush 2002). Left column: skeleton of 8-day-old chukar with wings of 8-day, 10-day, 20-day, and adult chukar attached and scaled to 8-day forelimb length; scaled wing lengths of extinct theropodsindicated by arrows (Hu et al. 2009; Wang et al. 2011); ‘‘wing size’’ defined as length of distal primary divided byforelimb length (Wang et al. 2011). Right column, stages of feather ontogeny and evolution (Table 1), with chukarprimary feathers shown in boxes and fossilized feathers on right: (stage i) 8–10-day chukar and basal maniraptorans with~symmetric feathers; (stage ii) 10–20-day chukar and basal paravians with ~symmetric to weakly asymmetric primaryfeathers; (stage iii) 20–49-day chukar, basal paravians, and basal avialans with asymmetric primary feathers. Stage i isontogenetically youngest and phylogenetically most basal, and stage iii is ontogenetically oldest and phylogeneticallymost derived, although overlap between stages occurs. Sm, Similicaudipteryx; Ca, Caudipteryx; An, Anchiornis; Bp, BPM 13–13; Mi, Microraptor; Ar, Archaeopteryx. Images of theropods from Zhou and Wang (2000), Norell et al. (2002), Xu et al.(2003, 2010), Hu et al. (2009), and Carney et al. (2012).
462 ASHLEY M. HEERS ET AL.
al. 2011; Heers and Dial 2012). Although thefossil record is complex, and extinct speciesundoubtedly utilized a variety of locomotorstrategies with both gain and loss of aerody-namic function (e.g., reduced plumage inEosinopteryx [Godefroit et al. 2013]), protow-ing-to-wing developmental transitions mirrorthe protowing-to-wing evolutionary transitionin many ways (Fig. 1). For example, bothyounger birds (6 days post-hatching, dph) andmore basal coelurosaurs (e.g., Beipiaosaurus,Similicaudipteryx STM4-1) often have distallybranched forelimb feathers, whereas olderbirds and more derived coelurosaurs tend tohave more unfurled and relatively symmetri-cally (8, 10 dph; e.g., Caudipteryx) or asym-metrically (49 dph; e.g., Archaeopteryx,Microraptor) vaned feathers (Ji et al. 1998; Xuet al. 1999; Xu et al. 2003; Dial et al. 2006; Xu etal. 2010). In addition, juvenile birds have moreflexible, more transmissive (permeable) feath-er vanes than older birds (Heers et al. 2011;Dial et al. 2012). Developmental and fossilevidence suggests that the earliest forms ofpennaceous feathers in theropod dinosaurslacked barbicels and possibly barbules (Prum
1999; Prum and Brush 2002). This wouldindicate that, like immature birds, the penna-ceous feathers of more basal coelurosaurswere less coherent and more transmissivethan the pennaceous feathers of more derivedcoelurosaurs. Finally, in immature birds and inextinct theropod dinosaurs, the first penna-ceous forelimb feathers to develop or evolveseem to be most pronounced on the distalforelimb (Garner et al. 1999; Dial et al. 2006;Xu and Guo 2009; Xu et al. 2010; Heers andDial 2012). More proximally positioned pen-naceous feathers appear in older birds andmore derived dinosaurs. Thus, there are manysimilarities between ontogenetic and evolu-tionary trajectories in feather shape, structuralintegrity, and positioning along the forelimb.Immature birds engage in flapping locomotorbehaviors at all of these stages of featherdevelopment, suggesting that extinct thero-pods might have used their feathered fore-limbs similarly.
Immature birds are certainly not identical toextinct theropods—no extant bird, for exam-ple, has a long tail, and juvenile bird feathersare probably more asymmetric than the
FIGURE 2. Phylogenetic bracket for estimating forelimb angular velocities in extinct theropod dinosaurs. Potentialangular flapping velocities achieved by dinosaurs with protowings can be phylogenetically bracketed by assuming thattheropod dinosaurs could have swung their forelimbs at least as fast as extant birds and lizards swing their limbs whilerunning (Xmin), and at most as fast as extant birds flap their wings while flying (Xmax). Theropod images adapted fromChristiansen and Bonde (2004) and Dial et al. (2008).
INCIPIENT WINGS AND THE EVOLUTION OF AVIAN FLIGHT 463
feathers of some extinct maniraptorans. How-ever, immature birds are far more similar toearly winged theropods than adult birds are,and they are the only living vertebrates thatcan be observed transitioning from an obli-gately bipedal to flight-capable state. Devel-oping birds with dinosaur-like anatomiesbridge this transition by using their legs andincipient wings cooperatively (Dial 2003; Dialet al. 2006; Jackson et al. 2009; Dial andJackson 2011; Dial and Carrier 2012; Heers andDial 2012), and wing-leg cooperation mayhave similarly facilitated the evolutionaryacquisition of flight. This is not to say thatontogeny strictly recapitulates phylogeny in alinear sense, as once proposed (Haeckel 1866).However, similarities between ontogeneticand evolutionary trajectories are fairly com-mon (Mayr 1963; Gould 1977; Heers and Dial2012) and even expected, because evolution isa process that modifies ontogeny—either byadding new characters or adjusting existingcharacters at various developmental stages(Gilbert and Epel 2009). Developing juvenilesreveal how transitional, morphing anatomiesfunction, and in this respect, the developmen-tal acquisition of flight capacity in extant birdscan help elucidate the evolutionary acquisitionof flight capacity among extinct theropods—by demonstrating the ecological utility ofdinosaur-like, incipient flight structures.
Here, we use the developing wings of atypical extant ground bird (Chukar Partridge[Alectoris chukar]) as possible analogues/ho-mologues of historical wing forms to providethe first empirical evaluation of aerodynamicpotential in flapping protowings of extincttheropods. To investigate how evolutionarytrends in feather morphology, wing size, andflapping velocity might have influenced liftand drag production in dinosaurs choosing toflap their incipient wings, we spun anontogenetic series of spread-wing prepara-tions on a rotating propeller apparatus acrossa range of flow conditions, and measuredaerodynamic force production. To the best ofour knowledge, the modeling described hererepresents the first analysis to empiricallyexplore flapping aerodynamics of the protow-ing-to-wing transition among extinct thero-pods. This analysis is the continuation of a
study documenting the ontogeny of aerody-namic force production in extant chukars(Heers et al. 2011): here, we build uponprevious work by using a developmentalseries of chukar protowings to model andexplicitly quantify aerodynamic potential offossil protowings. Although wing morpholo-gy is highly diverse among both living adultbirds and extinct theropods, extant specieswith very differently shaped wings experiencedevelopmental trajectories in feather structurethat are similar to each other and to evolu-tionary trajectories among theropod dinosaurs(Heers et al. 2011; Dial et al. 2012). Exploringfunctional morphology in chukar partridgesthus provides a useful starting point forelucidating relationships between form, func-tion, and behavior during protowing-to-wingtransitions.
Materials and Methods
To estimate lift and drag production forprotowings and wings of extinct theropods,we used a 4 3 3 3 2 factorial design with tworeplicates per treatment. Aerodynamic forceswere measured for four wing morphologies (8,10, 20, 49 dph) resembling basal to derivedmaniraptoran wings. We used younger chukarwings to model more basal maniraptorans,and older chukar wings to model morederived maniraptorans (stages i-iii in Fig. 1;Table 1). Wings were removed from deceasedanimals at the glenoid, dried in mid-down-stroke posture, and then spun like a helicopterblade on a propeller force plate apparatus,which imitates the mid-downstroke phase ofbirds flapping at low advance ratios (highflapping velocity and low translational veloc-ity, as during WAIR or takeoff) (Usherwood2009). Each wing was spun at a range ofReynolds numbers (Re) (flow conditions,proportional to wing size and velocity), tomimic extinct theropods with three differentwing sizes (forelimb lengths of 10, 14, 22 cm)flapping at slow (Xmin) and fast (Xmax) angularvelocities. The wings used in this study are thesame wings described by Heers et al. (2011),removed from deceased animals that wereraised at the Field Research Station at FortMissoula in Missoula, Montana, followingprotocols approved by the University of
464 ASHLEY M. HEERS ET AL.
Montana Institutional Animal Care and UseCommittee; for full details on animal hus-bandry, wing preparation, and the construc-tion of the propeller force plate apparatus, seeHeers et al. (2011).
Reynolds number (Re) is used to define theflow conditions experienced by an organismor object:
Re ¼ qlv
l; ð1Þ
where q is air density, l is mean wing chordlength (average width of the wing, measuredperpendicular to the leading edge), v is wing-tip velocity (tangential velocity of the wingtip, calculated as the product of wing length[m] and angular velocity [rad/s; rate ofangular displacement]), and l is dynamicviscosity (resistance to flow). Wings withidentical structural properties flapping atidentical Re will experience the same flowconditions at a given angle of attack. Thus,even though extinct theropods were oftenmuch larger than juvenile birds, and mighthave flapped their wings much more slowly(because they might have lacked large flightmuscles and/or neural circuitry), coefficientsof lift and drag calculated for an extantjuvenile protowing spinning at an extincttheropod Re can be used to estimate lift anddrag production for the extinct theropod(under the assumption that juvenile protow-ings are structurally similar to extinct thero-pod protowings; see Introduction). To evaluatean extinct theropod’s potential capacity forgenerating aerodynamic forces, we therefore(1) estimated a range of Re that theropodprotowings/wings might have experiencedwhile flapping, (2) used a force plate andpropeller apparatus to spin similar-lookingchukar protowings/wings at those Re andmeasure coefficients of lift and drag (CL, CD),and (3) used published body mass estimates toestimate lift and drag as a percentage of bodyweight for extinct theropods with pennaceousforelimb feathers. We did not attempt tomodel animals with only downlike feathers(e.g., Sinosauropteryx [Chen et al. 1998]),immature animals with feathers that mightstill have been growing (i.e., Similicaudipteryx
STM4-1 [Xu et al. 2010]), or animals withsecondary reductions in plumage (e.g., Eosi-nopteryx [Godefroit et al. 2013]), because theaim of this analysis was to explore theevolutionary acquisition (not loss) of flight.
Part 1: Estimating Re to Scale for Differences inWing Size and Flapping Velocity
Because Re is proportional to the product ofmean wing chord length and wing tip velocity(eq. 1), we estimated Re for flapping forelimbsof extinct theropods by estimating their meanwing chord lengths (l) and angular velocities(X; proportional to tip velocity).
Mean Wing Chord Length (l).—We usedtheropod-to-chukar scaling factors to approx-imate mean wing chord lengths for protowingsand wings of extinct theropods. First, wecalculated summed ‘‘effective’’ lengths of thehumerus, radius (ulna if data on radiusunavailable), second metacarpal, and phalan-ges II-1 through II-2 in mid-downstrokeposture for each bird wing (8, 10, 20, 49 dph)and for various basal maniraptorans, basalparavians, and basal avialans (SupplementaryTable 1, Supplementary Fig. 1; here, we refer tothe three theropod digits as I, II, and III).Effective forelimb lengths in extinct coeluro-saurs ranged from 4 to 33 cm, with mostlengths falling close to 10, 14, or 22 cm. Wethus chose lengths of 10, 14, and 22 cm foranalysis, to bracket the range of lengthsobserved in fossils. Although we cannot knowthe exact posture in which extinct theropodswould have held their forelimbs (and hencethe exact effective lengths and orientations),the aim of this project was not to estimateaerodynamic performance of specific fossilizedtheropods, but rather to determine howevolutionary changes in wing size and featherstructure generally might have influencedaerodynamic performance. Wing length andposture clearly affect aerodynamic perfor-mance, but by using chukar wings to modela range of limb lengths, feather morphologies,and body masses, and by measuring aerody-namic performance at multiple angles of attack(see Part 2 below), we hoped to account forsome of the variation in posture and anatomyand to realistically bracket the aerodynamic
INCIPIENT WINGS AND THE EVOLUTION OF AVIAN FLIGHT 465
performance that theropods might haveachieved at a given phylogenetic stage.
Next, we computed scaling factors as theratio of theropod-to-chukar effective bonelengths. We chose bone lengths to calculatescaling factors rather than wing or chordlengths because of the difficulty in measuringwing and chord lengths for fossils, whosewings are often folded or positioned at oddangles. Using theropod-to-chukar bone scalingfactors and mean chord lengths of the fourchukar age classes, we then estimated meanchord lengths (l) for theropod protowingswith each type of feather morphology (8, 10,20, 49 dph) and bone length (10, 14, 22 cm),using the following equation:
Theropod effective bone length
Chukar effective bone length
¼ Theropod chord length ðlÞChukar chord length
: ð2Þ
Given that wing size and shape are highlyvariable in both extant and extinct taxa, theanalysis presented here best applies to thero-pods with relatively short, low-aspect-ratiowings. However, using chukar wings to modelthe wings of extinct theropods with similarfeather morphologies is a logical startingpoint, because developing chukar wings aresimilar in shape and proportion to theincipient wings of many extinct theropods(Supplementary Box 1).
Flapping Velocities.—Potential flapping ve-locities achieved by extinct theropods withpennaceous forelimb feathers can be phyloge-netically bracketed (Witmer 1995) by assum-ing that theropods could have swung theirforelimbs at least as fast as extant birds andlizards swing their limbs while running(Xmin), and at most as fast as extant birds flaptheir wings while flying (Xmax) (Fig. 2).
1. Minimum angular velocities (Xmin). Frompreviously published data on maximalrunning speeds in birds and lizards (Sup-plementary Table 2, Supplementary Fig. 2),we estimated the relationship betweenlimb length and running angular velocityfor extant lizards and birds as
log10ðrunning angular velocity; XminÞ¼ �0:60
�log10ðeffective leg lengthÞ
�þ 0:60;
ð3Þ
where
Running angular velocity ðXmin; rad=sÞ
¼ Leg tip velocity
Effective leg length; ð4Þ
and
Leg tip velocityðm=sÞ
¼ Stride length
Swing duration� running velocity: ð5Þ
Minimum angular velocities for theropodprotowings were estimated using equation (3)by substituting theropod wing length forbird/lizard effective leg length. These valueswere then used to calculate minimum pro-towing tip velocities (vmin):
vmin ¼ Minimum wing tip velocityðm=sÞ¼ ðwing lengthÞðXminÞ; ð6Þ
where theropod wing lengths were estimatedfor theropod forelimbs with each type offeather morphology (8, 10, 20, 49 dph) andeach bone length (10, 14, 22 cm), using theequation:
Theropod effective bone length
Chukar effective bone length
¼ Theropod wing length
Chukar wing length: ð7Þ
2. Maximum angular velocities (Xmax). Frompreviously published data on flying angu-lar velocities in birds during takeoff (Sup-plementary Table 3, Supplementary Fig. 3),the relationship between wing length andwing angular velocity was estimated as
log10ðflying angular velocity; XmaxÞ¼ �0:54
�log10ðwing lengthÞ
�þ 1:45: ð8Þ
Maximum protowing angular velocitieswere estimated using equation (8) by substi-tuting theropod wing length (eq. 7) for birdwing length. These values were then used to
466 ASHLEY M. HEERS ET AL.
calculate maximum protowing tip velocities(vmax):
vmax ¼ Maximum wing tip velocityðm=sÞ¼ ðwing lengthÞðXmaxÞ: ð9Þ
Part 2: Using a Force Plate and PropellerApparatus to Measure Lift and Drag
Using the mean chord lengths and mini-mum and maximum velocities describedabove, we computed Re representing 10, 14,and 22 cm effective bone lengths swinging atminimum (Xmin) and maximum (Xmax) angu-lar velocities (Supplementary Table 4), for atotal of 24 feather morphology–bone length–angular velocity combinations. To measureaerodynamic force production at minimumangular velocities (Xmin), we followed themethods of Usherwood and Ellington (Usher-wood and Ellington 2002) and spun each wingon a propeller force plate apparatus, at anglesof attack of 158, 308, 458, and 608 (Supplemen-tary Movie 1; for full details on this procedurerefer to Heers et al. [2011], and for a sketch ofthe experimental setup, see Crandell andTobalske [2011]). Vertical and horizontal aero-dynamic forces generated by the spinningwings were converted into coefficients of lift(CL) and drag (CD):
CV ¼2FV
qX2S2
; ð10Þ
CH ¼2Q
qX2S3
; ð11Þ
CL ¼ ðCVcoseþ CHsineÞ 1
cos e
� �2
; ð12Þ
and
CD ¼ ðCHcose� CVsineÞ 1
cos e
� �2
; ð13Þ
where CV is the coefficient of vertical force, CH
is the coefficient of horizontal force, FV isvertical force measured by the force plate (N),Q is torque (Nm) about the z-axis, q is airdensity at Missoula, Montana (1.07 kg/m3), Xis angular velocity of the spinning chukarwing (rad/s), S2 is the second moment of area
of the chukar wing (m4), S3 is the thirdmoment of area of the chukar wing (m5),and e is the downwash angle; induceddownwash velocity was modeled assuming aRankine-Froude momentum jet and a trian-gular distribution for local induced down-wash along the wing (see Usherwood andEllington 2002). Lift and drag coefficients werethen used to estimate lift and drag productionby theropod protowings (see Part 3 below).For maximum angular velocities (Xmax), coef-ficients of lift and drag were recorded at Rebased on in vivo kinematic measurements onchukars (Heers et al. 2011), under the assump-tion that these coefficients would be represen-tative of wings moving at flying angularvelocities regardless of scaling.
Part 3: Estimating Lift and Drag as aPercentage of Body Weight for ExtinctTheropods with Pennaceous ForelimbFeathers
We used coefficients of lift and drag toestimate lift and drag production by theropodprotowings using a blade element model:
Resultant forceðNÞ
¼ 0:5qCR
Z r¼R
r¼0
�ðXrÞ2 þ VT
2�
cdr; ð14Þ
where q is air density at Missoula (1.07 kg/m3), CR is the resultant coefficient (CR¼ (CL
2þCD
2)0.5) at a 458 angle of attack (in vivo angleof attack at mid-downstroke in chukars[Jackson et al. 2009; Heers et al. 2011]), R istheropod wing length (m), X is angularvelocity (rad/s), VT is translational velocity(m/s), and c is mean chord length (m).Translational velocities were conservativelyset at 1.5 m/s for all theropods. During WAIRthere is no clear relationship between bodysize and velocity, but both juvenile and adultchukars (Jackson et al. 2009) and peafowl(Heers and Dial 2013)—weighing between 100g and 5 kg and spanning the range ofestimated weights for theropods (Table 1)—can flap-run at least at 1.5 m/s. Birds in thissize range move more rapidly when runningon horizontal surfaces, jumping into the air, orusing their wings to take off (SupplementaryFig. 4), and so 1.5 m/s is a conservative
INCIPIENT WINGS AND THE EVOLUTION OF AVIAN FLIGHT 467
translational velocity irrespective of behavior.Angular velocities (X) and translational veloc-ities (VT) were also conservatively modeled asperpendicular to one another, and summed toobtain the net wing velocity (Vnet ¼ [(Xr)2 þVT
2]0.5). Resultant forces were multiplied bytwo (to account for both wings), then ex-pressed as a percentage of body weight bydividing by published estimates of theropodbody weights (Table 1).
Because propeller models mimic only forcesproduced at mid-downstroke, each forceestimate for extinct theropods was standard-ized by the average resultant force (as apercentage of body weight) of two adultchukar wings spinning at flying angularvelocities (Xmax). Thus a force of 100% bodyweight suggests that a theropod could havefully supported body weight during flight. Aforce of ~6–8% body weight suggests that atheropod could have engaged in WAIR,because the resultant force produced by 6–8-day-old chukars during WAIR is ~6–8% of theforce produced by adult chukars during flight(based on in vivo measurements [Tobalskeand Dial 2007] and on the force platemeasurements described here).
Following standardization, all force esti-mates for maximum (but not minimum)angular velocities were reduced by 20% toaccount for the additional flight musculaturethat was not considered in calculations ofbody mass, and that would presumably benecessary to oscillate wings at high speed (ourgoal was to estimate the maximal flightperformance possible—as if extinct theropodswere capable of oscillating their forelimbs likeextant birds—which presumably would re-quire extant-like values of muscle mass).Forelimb muscle mass varies widely acrossextant species (Hartman 1961), but is notnecessarily dictated by size. Chukars (~500g) and peafowl (~4–5 kg, approaching theupper limit of sizes considered here), forexample, are both capable of rapid bursttakeoffs and both have pectoral muscle (pec-toralis þ supracoracoideus) masses that are20–21% of their body mass (Heers and Dial2013). Galliforms are known for their rapidwingbeat frequencies and powerful burstflight, so a theropod with pectoral muscles of
20% body mass should have been capable ofhigh wingbeat frequencies. Finally, the small-est body sizes (100 g with a 14-cm forelimb,700 g with a 22-cm forelimb) were eliminatedfrom calculations at maximum angular veloc-ities (Xmax) because such small animals prob-ably could not have oscillated 14- or 22-cmforelimbs at such high speeds, given that a 49-dph chukar with an 11.2-cm forelimb weighs~250 g. For a sample calculation, see Supple-mentary Box 2.
Results
Aerodynamic performance improves dur-ing protowing-to-wing transitions, by increas-ing aerodynamic force production and lift-to-drag ratios (rs¼ 1 for peak CL and peak CL/CD
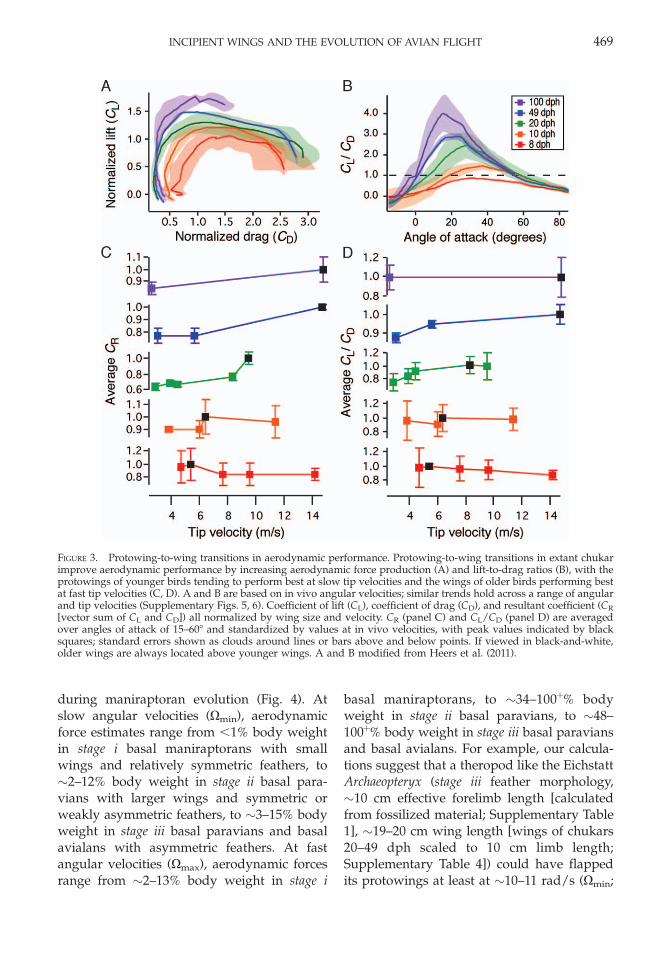
vs. age [Heers et al. 2011]) (Fig. 3A,B). At allcombinations of bone size and angular veloc-ity (all Re categories), older wings with stiffer,more structurally coherent (less transmissive),and more asymmetric primary feathers pro-duce larger aerodynamic force coefficientsthan younger protowings with more flexible,more transmissive, and weakly asymmetric orsymmetric feathers (i.e., older wings [repre-senting the wings of more derived coeluro-saurs] have greater aerodynamic performancethan younger wings [representing the wingsof more basal coelurosaurs]; stage iii . stage ii. stage i) (Supplementary Figs. 5, 6). At allcombinations of bone size and feather mor-phology, fast angular velocities (Xmax) yieldhigher aerodynamic forces than slow angularvelocities (Xmin) (because force � velocity2).However, when aerodynamic forces (lift, drag)are normalized for wing size and velocity (andexpressed as CL, CD), there appears to be aslight tendency for younger (i.e., more basal)protowings to perform best at slower tipvelocities, and older (i.e., more derived) wingsto perform best at faster tip velocities (tipvelocity ¼ product of angular velocity andwing length; rs ¼ 1 for age vs. tip velocity atwhich peak CR and peak CL/CD occur) (Fig.3C,D).
Chukar wing models and published esti-mates of forelimb lengths and body masses fortheropod dinosaurs (Table 1, SupplementaryTable 1) suggest that weight-specific aerody-namic force production would have increased
468 ASHLEY M. HEERS ET AL.
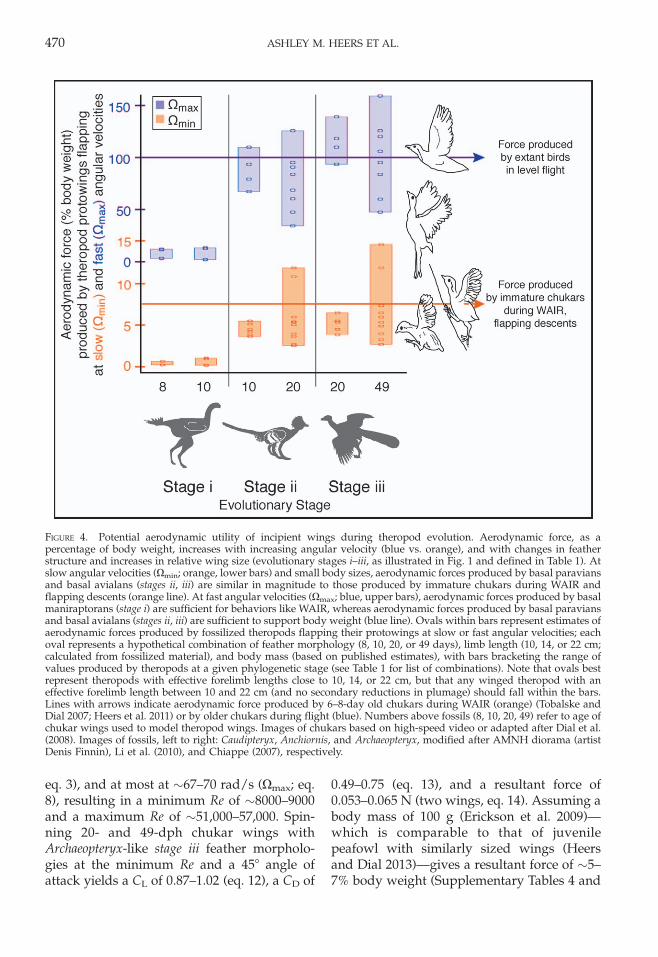
during maniraptoran evolution (Fig. 4). Atslow angular velocities (Xmin), aerodynamicforce estimates range from ,1% body weightin stage i basal maniraptorans with smallwings and relatively symmetric feathers, to~2–12% body weight in stage ii basal para-vians with larger wings and symmetric orweakly asymmetric feathers, to ~3–15% bodyweight in stage iii basal paravians and basalavialans with asymmetric feathers. At fastangular velocities (Xmax), aerodynamic forcesrange from ~2–13% body weight in stage i
basal maniraptorans, to ~34–100þ% bodyweight in stage ii basal paravians, to ~48–100þ% body weight in stage iii basal paraviansand basal avialans. For example, our calcula-tions suggest that a theropod like the EichstattArchaeopteryx (stage iii feather morphology,~10 cm effective forelimb length [calculatedfrom fossilized material; Supplementary Table1], ~19–20 cm wing length [wings of chukars20–49 dph scaled to 10 cm limb length;Supplementary Table 4]) could have flappedits protowings at least at ~10–11 rad/s (Xmin;
FIGURE 3. Protowing-to-wing transitions in aerodynamic performance. Protowing-to-wing transitions in extant chukarimprove aerodynamic performance by increasing aerodynamic force production (A) and lift-to-drag ratios (B), with theprotowings of younger birds tending to perform best at slow tip velocities and the wings of older birds performing bestat fast tip velocities (C, D). A and B are based on in vivo angular velocities; similar trends hold across a range of angularand tip velocities (Supplementary Figs. 5, 6). Coefficient of lift (CL), coefficient of drag (CD), and resultant coefficient (CR
[vector sum of CL and CD]) all normalized by wing size and velocity. CR (panel C) and CL/CD (panel D) are averagedover angles of attack of 15–608 and standardized by values at in vivo velocities, with peak values indicated by blacksquares; standard errors shown as clouds around lines or bars above and below points. If viewed in black-and-white,older wings are always located above younger wings. A and B modified from Heers et al. (2011).
INCIPIENT WINGS AND THE EVOLUTION OF AVIAN FLIGHT 469
eq. 3), and at most at ~67–70 rad/s (Xmax; eq.8), resulting in a minimum Re of ~8000–9000and a maximum Re of ~51,000–57,000. Spin-ning 20- and 49-dph chukar wings withArchaeopteryx-like stage iii feather morpholo-gies at the minimum Re and a 458 angle ofattack yields a CL of 0.87–1.02 (eq. 12), a CD of
0.49–0.75 (eq. 13), and a resultant force of0.053–0.065 N (two wings, eq. 14). Assuming abody mass of 100 g (Erickson et al. 2009)—which is comparable to that of juvenilepeafowl with similarly sized wings (Heersand Dial 2013)—gives a resultant force of ~5–7% body weight (Supplementary Tables 4 and
FIGURE 4. Potential aerodynamic utility of incipient wings during theropod evolution. Aerodynamic force, as apercentage of body weight, increases with increasing angular velocity (blue vs. orange), and with changes in featherstructure and increases in relative wing size (evolutionary stages i–iii, as illustrated in Fig. 1 and defined in Table 1). Atslow angular velocities (Xmin; orange, lower bars) and small body sizes, aerodynamic forces produced by basal paraviansand basal avialans (stages ii, iii) are similar in magnitude to those produced by immature chukars during WAIR andflapping descents (orange line). At fast angular velocities (Xmax; blue, upper bars), aerodynamic forces produced by basalmaniraptorans (stage i) are sufficient for behaviors like WAIR, whereas aerodynamic forces produced by basal paraviansand basal avialans (stages ii, iii) are sufficient to support body weight (blue line). Ovals within bars represent estimates ofaerodynamic forces produced by fossilized theropods flapping their protowings at slow or fast angular velocities; eachoval represents a hypothetical combination of feather morphology (8, 10, 20, or 49 days), limb length (10, 14, or 22 cm;calculated from fossilized material), and body mass (based on published estimates), with bars bracketing the range ofvalues produced by theropods at a given phylogenetic stage (see Table 1 for list of combinations). Note that ovals bestrepresent theropods with effective forelimb lengths close to 10, 14, or 22 cm, but that any winged theropod with aneffective forelimb length between 10 and 22 cm (and no secondary reductions in plumage) should fall within the bars.Lines with arrows indicate aerodynamic force produced by 6–8-day old chukars during WAIR (orange) (Tobalske andDial 2007; Heers et al. 2011) or by older chukars during flight (blue). Numbers above fossils (8, 10, 20, 49) refer to age ofchukar wings used to model theropod wings. Images of chukars based on high-speed video or adapted after Dial et al.(2008). Images of fossils, left to right: Caudipteryx, Anchiornis, and Archaeopteryx, modified after AMNH diorama (artistDenis Finnin), Li et al. (2010), and Chiappe (2007), respectively.
470 ASHLEY M. HEERS ET AL.
5, Supplementary Box 2). This range is verysimilar in magnitude to the aerodynamicforces produced by immature chukars duringWAIR, controlled flapping descents, andjump-flapping up to elevated perches (~6–8% body weight in birds 6–8 dph; orange linein Fig. 4) (Tobalske and Dial 2007; Jackson etal. 2009; Heers et al. 2011), suggesting thatArchaeopteryx-like theropods would have beencapable of comparable behaviors. By similarreasoning, at maximum angular velocities(~67–70 rad/s, Xmax), Archaeopteryx-like the-ropods would have produced forces sufficientto support their body weight during flight(�100% body weight).
Discussion
Our findings, in conjunction with work ondeveloping birds, suggest that flapping pro-towings could have provided useful aerody-namic function early in the history ofManiraptora, with improvements in aerody-namic performance attending the evolution oflarger wings, more effective feather morphol-ogies, and faster flapping velocities. Thoughfeathered forelimbs serve many functions inextant birds, and were probably used for avariety of purposes before being exapted forpowered flight (e.g., insulation [Norell and Xu2005], display [Xu and Guo 2009; Dimond etal. 2011], and/or balance [Fowler et al. 2011]),our results indicate that forelimb feathers alsocould have functioned aerodynamically forflapping locomotor behaviors like WAIR. Ourexperiments indicate that at slow angularvelocities (Xmin), maniraptorans with stage iiand stage iii feather morphologies could haveproduced aerodynamic forces sufficient forflap-running up slopes, jumping and flappingup to elevated surfaces, and slowing aerialdescents (Fig. 4, orange line). At fast angularvelocities (Xmax), all maniraptorans (stages i–iii) would have been capable of WAIR, flap-jumps and controlled aerial descents, andmaniraptorans with stage ii and stage iii feathermorphologies would have been able to sup-port their body weight during flight as well(Fig. 4, blue line). Our ontogenetic modelsusing Re to account for variation in size and tobracket potential flapping velocities (Fig. 2)thus indicate that protowings could have
functioned aerodynamically relatively earlyin maniraptoran history, with evolutionarychanges in feather structure, relative wingsize, and flapping velocity dramatically im-proving aerodynamic performance.
Feather Structure
Feather morphology changes considerablyduring bird ontogeny and theropod evolution.In chukars, flight feathers begin to emerge at~6 days post-hatching, in the form of pro-towings that enable chicks to produce aero-dynamic forces and flap-run up slopes or slowaerial descents. Protowings with flexible,transmissive, and relatively symmetric feath-ers therefore provide useful aerodynamicfunction early in chukar ontogeny. In speciesexamined thus far, developmental changes infeather structure that are similar to changesoccurring during theropod evolution arecorrelated with increases in aerodynamic forceproduction and higher lift-to-drag ratios (Dialet al. 2006; Tobalske and Dial 2007; Heers et al.2011; Dial et al. 2012; Heers and Dial 2012)(Fig. 3A,B), most likely due to increases inflexural stiffness and feather asymmetry, and/or decreases in wing and feather transmissiv-ity. Asymmetric primary feathers with thickrachises and tightly interlocking barbules mayimprove performance by stabilizing primaryfeathers against oncoming airflow (Norberg1985), preventing excessive deformation(Nudds and Dyke 2010), reducing featherpermeability (Muller and Patone 1998; Heerset al. 2011; Dial et al. 2012), and/or influencingthree-dimensional wing morphing. Improvedperformance cannot be explained by ontoge-netic changes in static wing shape (aspectratio, camber), wing size, or flapping velocity(Heers et al. 2011; Dial et al. 2012). Forexample, by 49 dph, chukars have grown allof their flight feathers (primaries, secondaries,coverts) and differ from adults only withrespect to feather structure and the relativelengths of the two distal-most primaries(Supplementary Fig. 7). Given that adultwings produce more aerodynamic force andmore lift per unit drag than 49-dph wings (Fig.3), even when standardized for wing size andvelocity, feather structure must play an im-portant role in aerodynamic force produc-
INCIPIENT WINGS AND THE EVOLUTION OF AVIAN FLIGHT 471
tion—though even symmetric, flexible, andtransmissive pennaceous feathers are aerody-namically useful.
In short, protowings are immediately func-tional in extant birds, and potentially inextinct theropods. Developmental changes infeather morphology and attending improve-ments in aerodynamic performance suggestthat comparable evolutionary changes infeather structure might similarly have im-proved aerodynamic performance amongextinct theropods. According to our experi-ments, at slow angular velocities (Xmin),maniraptorans with stage ii and stage iii feathermorphologies could have produced aerody-namic forces sufficient for WAIR and similarbehaviors (Fig. 4, orange line); at fast angularvelocities (Xmax), all maniraptorans (stages i–iii) would have been capable of WAIR-likebehaviors, and maniraptorans with stage ii andstage iii feather morphologies would havebeen able to engage in flight as well (Fig. 4,blue line).
Flapping Velocity
Irrespective of feather morphology, allwings produce more aerodynamic force athigher angular velocities (force � velocity2).However, when aerodynamic force is stan-dardized for wing size and flapping velocity(CL, CD), wings seem to perform better at invivo velocities, with the wings of older birdstending to perform best at higher velocitiesand the wings of younger birds tending toperform best at lower velocities (Fig. 3C,D).This pattern may suggest that the incipientwings of younger birds and more basalmaniraptorans are(were) tuned to flapping atlower tip velocities, and that the wings ofolder birds and more derived maniraptoransare(were) tuned to flapping at higher tipvelocities. Such aeroelastic ‘‘tuning’’ is consis-tent with ontogenetic and evolutionary in-creases in wing length and in keel size (Wanget al. 2011; Heers and Dial 2012)—for a givenangular velocity, increasing wing length in-creases wing tip velocity (as observed inchukars that do not change X much duringdevelopment [Jackson et al. 2009; Heers et al.2011]), whereas increasing keel size is presum-ably associated with increases in pectoral
muscle mass and the ability to achieve higherflapping velocities. Feather morphology andwing kinematics thus may be developmental-ly and evolutionarily tuned or coupled (seeShyy et al. 2010).
Also irrespective of feather morphology, allwings produce aerodynamic force at slowangular velocities (Xmin). Developing birdsdemonstrate that even small amounts ofaerodynamic force (,10% body weight) im-prove locomotor performance during behav-iors like WAIR, controlled flapping descent, orjumping and flapping to elevated perches(Dial 2003; Tobalske and Dial 2007; Jacksonet al. 2009; Heers et al. 2011; Heers and Dial2013). Thus, extinct theropods that were notcapable of flapping rapidly (Fig. 4, orangebars), or of producing a fully birdlike wingstroke (with high stroke amplitudes or chan-nelized kinematics), might still have benefitedfrom flapping their feathered forelimbs andproducing some aerodynamic force. Fastangular velocities and ‘‘birdlike’’ wing strokesmay not be prerequisites for locomotor be-haviors requiring little aerodynamic force,such as controlled flapping descent and WAIRor jumping.
Relative Wing Size
Published estimates of forelimb lengths andbody masses for theropod dinosaurs (Table 1,Supplementary Table 1) indicate that relativewing size, and hence the potential to produceuseful aerodynamic forces, increased duringmaniraptoran evolution. At slow angularvelocities (Xmin), aerodynamic force estimatesrange from ~1% to ~15% body weight, withmore derived maniraptorans tending to pro-duce larger forces (stage iii . stage ii . stage i;Fig. 4, orange bars). At fast angular velocities(Xmax), aerodynamic force estimates rangefrom ~2% to 100þ% body weight, with largerforces again being produced by more derivedmaniraptorans (blue bars). Immature chukarscan flap-run up steep inclines (.658), jumpand flap up to low perches, and slow aerialdescents by producing aerodynamic forcesthat are less than 10% of their body weight(~6–8% body weight for birds 6–8 dph duringWAIR [Tobalske and Dial 2007]), indicatingthat small theropods with stage ii or stage iii
472 ASHLEY M. HEERS ET AL.
feather morphologies may have achievedsimilar behaviors, even when flapping slowly.Basal paravians and basal avialans capable offlapping more rapidly might have been able togenerate forces sufficient to support bodyweight for level flight.
Although maniraptorans with stage i feath-ers seemingly had wings that were too smallto generate much aerodynamic force relativeto adult mass at slow angular velocities (Xmin;Fig. 4, orange bars), it is plausible thatjuveniles of even large-bodied theropodsmight have benefited from flapping theirfeathered forelimbs (Dial and Jackson 2011).Owing to a paucity of data, this study couldnot consider extinct juveniles. However, manytheropod species required several years toreach adult mass (Padian et al. 2001; Ericksonet al. 2004). If wings developed early, as inmany precocial ground birds, then immatureindividuals might have been able to generateuseful aerodynamic forces prior to outgrow-ing their wings as adults. Brush turkeys (Dialand Jackson 2011) and peafowl (Heers andDial 2013), for example, have relatively largerwings and greater wing performance asjuveniles. Similarly, Velociraptor (Turner et al.2007b) and other maniraptorans that wererelatively large and flight-incapable as adultsmay have had aerodynamically functionalwings as juveniles. Although we often focuson adult stages, selection on aerodynamiccapacity might have originally occurredamong juveniles, and only later among adultsthat did not attain large body size.
Wing-Leg Cooperation
An important theme that has emerged fromwork with extant developing birds is that evensmall amounts of aerodynamic force improvelocomotor performance, particularly duringbehaviors involving the cooperative use ofwings and legs. Wing-leg cooperation enhanc-es locomotor performance throughout ontog-eny, bridging obligately bipedal and flight-capable developmental stages by allowingjuveniles to transition from (1) leg-basedterrestrial (or aquatic) locomotion, to (2) wing-and leg-based locomotion on inclined (oraquatic) substrates, to (3) wing-based aeriallocomotion (Dial et al. in press). Behaviors like
WAIR, steaming, or jumping and flapping toelevated surfaces are ubiquitous across extantbirds and crucial to immature birds withincipient wings (Dial 2011), leading to thehypothesis that wing-leg cooperation mightalso have served as an evolutionary bridgebetween leg- and wing-based locomotion.
This hypothesis is consistent with the fossilrecord. With some extant exceptions (e.g.,Apodiformes), hind limbs have always beena conspicuous part of theropod anatomy andthus probably played an important roleduring the evolutionary acquisition of flight.In some coelurosaurs, hind limb input mayhave occurred in the form of ground reactionforces during WAIR (Dial 2003) and/orlaunching and landing (Earls 2003; Heersand Dial 2013). In other coelurosaurs—the‘‘four-winged’’ paravians—hind limbs mayhave also generated aerodynamic forces (e.g.,Xu et al. 2003). Extinct species undoubtedlyutilized a variety of locomotor strategies andengaged their fore- and hind limbs for manypurposes, including non-locomotor behaviorssuch as feeding or signaling. In all cases,however, the hind limbs probably facilitatedwing-based locomotion, playing a crucial rolein coelurosaurs with protowings and poten-tially becoming less important as larger andmore robust flight apparatuses evolved (seeDial 2003; Xu and Zhang 2005; Heers and Dial2012; Zheng et al. 2013).
Conclusions
Collectively, our findings suggest that ‘‘pro-towings’’ may have provided useful aerody-namic function early in the history ofmaniraptoran theropods, particularly amongsmall or immature individuals. Improvementsin aerodynamic performance would haveoccurred as (1) changes in feather structureincreased force production and lift-to-dragratios, (2) reductions in body mass andincreases in wing size increased relative wingsize, and (3) increases in flight musculatureallowed for faster flapping velocities. Extantjuvenile birds demonstrate that incipient wingscan function aerodynamically, especially whenwings are supplemented by legs during behav-iors like jumping and flapping to elevatedsurfaces or WAIR (Tobalske and Dial 2007;
INCIPIENT WINGS AND THE EVOLUTION OF AVIAN FLIGHT 473
Heers et al. 2011; Heers and Dial 2013). Giventhat developing birds rely on small musclesand dinosaur-like protowings and skeletons(Heers and Dial 2012) to bridge the develop-mental transition from obligately bipedal juve-nile to flight-capable adult, such behaviors mayhave been similarly important to theropodsduring the evolutionary acquisition of flight. Byusing a protowing-to-wing developmentaltransition to model the protowing-to-wingevolutionary transition among theropod dino-saurs, and aerodynamic theory to account fordifferences in size and potential flappingvelocity, this study provides the first experi-mental evidence to suggest that feathereddinosaurs choosing to flap their incipient wingswould have been capable of producing usefulaerodynamic forces, similar in magnitude tothose produced by immature birds using theirwings and legs cooperatively or flapping toslow aerial descents. Protowings are aerody-namically functional, and incremental changesin form, function, and behavior that facilitatethe development of flight among extant birdsmay have similarly facilitated the evolution offlight among extinct theropods.
Acknowledgment
The authors wish to thank B. Jackson andnumerous other colleagues for thoughtfulcomments and invaluable discussions. Sup-ported by National Science Foundation grantsGRFP-2007057068 (to A.M.H.) and NSF IOS0919799 (to K.P.D. and B.W.T.).
Literature Cited
Aigeldinger, T., and F. Fish. 1995. Hydroplaning by ducklings:
overcoming limitations to swimming at the water surface.
Journal of Experimental Biology 198:1567–1574.
Burgers, P., and L. M. Chiappe. 1999. The wing of Archaeopteryx as
a primary thrust generator. Nature 399:60–62.
Carney, R. M., J. Vinther, M. D. Shawkey, L. D’Alba, and J.
Ackermann. 2012. New evidence on the colour and nature of the