THE FORMAL DYNAMICS OF CONTROLLED POPULATIONSAND THE ECHO, THE BOOM AND THE BUST
Ronald LeeDepartment of Economics and Population Studies Center, University of Michigan,1225 S. University Ave., Ann Arbor, Michigan 48104
Abstract-This paper analyzes the pattern of fluctuations of births in an agestructured population whose growth is subject to environmental or economic constraint. It synthesizes the traditional demographic analysis ofage-structured renewal with constant vital rates and the economic analysiswhich treats population change endogenously. When cohort fertility depends on relative cohort size, or when period fertility depends on laborforce size, fluctuations of forty or more years replace the traditional "echo"or generation-length cycle. Twentieth-century U. S. fertility change agreeswell with the theory, as the "Easterlin Hypothesis" suggests; but the periodmodel fits better than the cohort model.
1. INTRODUCTION the reversal of the echo effect, are re-All populations are subject to environ- garded from this point of view.
mental constraint in one form or another, Numerous models express an assumedand because their potential growth is so long-run tendency of population to equilrapid, they are typically found near equi- ibrate with productive capacity. Manylibrium, where the constraints operate of these derive from the populationeffectively through checks on population theory of classical economics and aregrowth and size. So Malthus argued; and exemplified by the work of Leibensteinthis paper explores the implications of his (1963), Solow (1956) and Lee (1970,view for the analysis of population dy- 1973, 1974b). Others are extensions tonamics. human populations of models devel-
However, despite an extensive litera- oped for animal ecology, as in the workture on the economics of fertility and the of Pearl (1924), Birdsell (1957), Sauvymacroeconomic effects of population (1969) and Wrigley (1969). There is regrowth, there is little direct evidence that lated work by animal population ecolpopulation is an endogenous component ogists with which this author can claimin an equilibrating system. The approach little familiarity. Most of these modelsof this paper is indirect: we assume that are static or, if dynamic, ignore the agepopulation is near equilibrium and de- structure of the population and the varirive the consequences for the dynamics ous lags which occur in the adjustmentof population renewal. Comparison of process. But oscillations are characterisactual population dynamics to the be- tic of controlled systems, since sensitivehavior implied by the model may then regulation, operating with a response lag,shed light on the questions of whether a can lead to overshooting of equilibriumcontrol system exists, and if so, how it and what we will call "control cycles."operates. In particular, the U. S. baby Thus, while abstraction from lags andboom and baby bust, characterized by age structure may be appropriate for the
563
564 DEMOGRAPHY, volume 11, number 4, November 1974
long run, for many applications we willwant to know more about the convergence path of population, about the stability of equilibrium, and about the cyclictendencies of the system.
There is also an extensive body of literature analyzing the dynamic behaviorof age-structured populations as theyproceed from an initially disturbed stateto steady exponential growth, under theassumption of constant vital rates. Thiswork derives from the models and analysis of Lotka, the pioneer mathematicaldemographer, and includes contributionsby Bernardelli (1941), Keyfitz (1965),Coale (1972) and LeBras (1969). Thiswork establishes a tendency for humanpopulations to move in cycles of onegeneration, or twenty-five to thirtythree years; we will call these "generational cycles" or "echos." However, allthe models either assume that vital ratesare constant or that they vary independently of the size and structure of thepopulation; they thus abstract from theexistence of environmental constraints.
There has not yet been a general analysis of population dynamics which incorporates both the operation of constraintsand the age structure of the populationand thus relates the control cycle andgenerational cycle (although Keyfitz,1972, touches on closely related matters).However, there is a long tradition ofinteresting speculations regarding the dynamic behavior arising from the interactions of the two. Malthus (1970) confidently asserted that population andwelfare perpetually oscillate about theirequilibria, due to the lag of labor supplybehind demand and the consequent overreaction of net reproduction and capitalformation to prevailing conditions oflabor. Alfred Marshall expressed similarviews (Liu, 1971), and Yule (1906)argued for a periodicity of fifty to onehundred years.
More recently, it has been suggestedthat the time series of fertility in theUnited States might be due to a strong
negative reaction of fertility to cohortsize or potential labor force size; alongthese lines we have the contributions ofGrauman (1960), Easterlin (1962, 1968),Coale (1963), Ryder (1971), Lee (1970)and Keyfitz (1972). Easterlin, withwhose name this hypothesis is associated,has made the most careful empiricalstudies and, together with Condran(1974), has recently extended the analysis to several other industrial countries.
These studies suggest that an economically endogenous population with agestructure might exhibit long control cycles in place of the classic echo effect. Ifso, the empirical analysis of populationfluctuations could provide evidence concerning the presence and nature of autoregulatory mechanisms governing population growth.
Regular fluctuations in human populations have long been observed. Someoscillations, with periods up to fifteenyears, are obviously imposed on demographic series by the periodicity of climate, epidemic, harvest, and the intrinsicdynamics of capitalist economies; others,like the twenty-year Kuznets cycle, mayreflect economic-demographic interactions (Easterlin, 1968). But these cyclesare superimposed on longer fluctuationsof less certain origin. The most widelyrecognized of these is a thirty-year cyclewhich is noticeable in the graphs of baptisms from many preindustrial parishes(see, e.g., Goubert, 1965). This could bean empirical manifestation of the "generational" cycle, but we cannot reallytell without analyzing it in conjunctionwith stochastic models of the sort to bedeveloped in this paper.
Some observers also see evidence oflonger cycles in human populations, oflength closer to two generations, or fortyto sixty years. Thus the fifty-year Kondratieff cycle runs through the demographic variables of nineteenth-centuryEurope and could possibly originate ina demographic control cycle. Similarly,there is a forty-year "cycle" in the fer-
Dynamics of Controlled Populations
tility of the United States in the twentieth century; it declines to a troughin the mid-thirties, rises to the "babyboom" peak in the late fifties, and declines in the "baby bust" thereafter. Thishas sometimes been interpreted as a control cycle.
This paper develops a formal modelof renewal in an age-structured population subject to environmental constraint.We analyze the dynamics of the model ina way compatible with empirical estimation and testing. We conclude with anempirical application to U. S. fertility,1917 to 1972.
2. CONTROLLED POPULATION WITHOUT
AGE STRUCTURE
By a "controlled" population, we meanone which is endogenous to a systemwhich tends toward equilibrium. This requires that population growth respondpositively to variation in material welfare, while material welfare should inturn be depressed by population growth.For some level of material welfare, population will remain stationary and thesystem will be in equilibrium. The equilibrium levels of the system, and thespeed with which it converges, depend onsocial and economic institutions, technology, and resources.
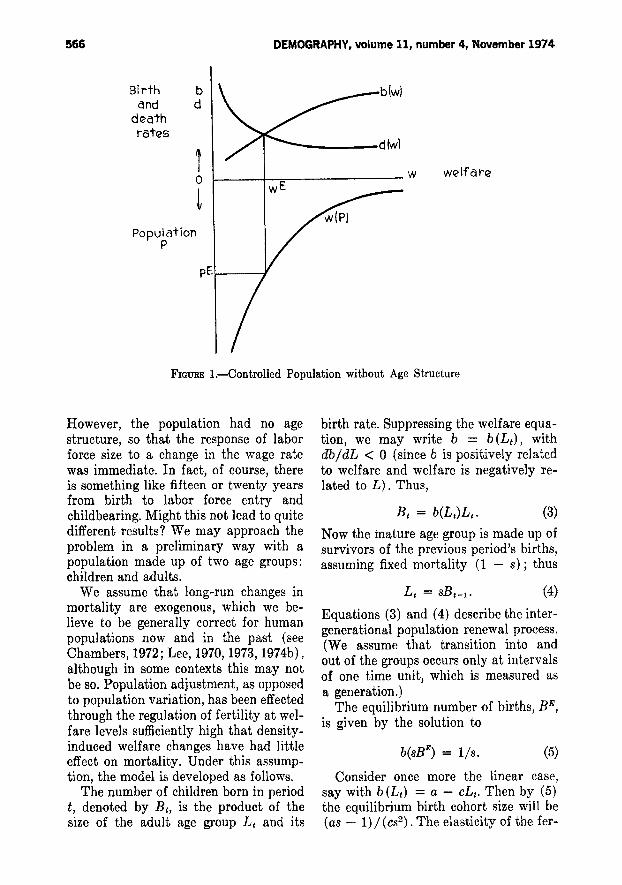
The basic system for a populationwithout age structure is illustrated inFigure 1. Aggregate fertility, measuredby the crude birth rate, b, is positivelyrelated to material welfare, w (whichcould be real wages, per capita income,average space, or any other appropriateconcept), with the form and level of therelation determined institutionally. (Fora discussion of relevant empirical work,see Simon, 1974.) This is represented bythe curve b(w). Mortality, measured bythe crude death rate, d, is negativelyrelated to w, as shown by the curve d(w) .At some level of welfare, b (w) andd (w) are equal, and the population istherefore stationary. This defines the
565
equilibrium welfare, WE, as noted on thediagram.
Welfare, w, is in turn negatively related to population size, P, by a functionreflecting resources, reproducible capital,technology and social organization. Thebottom half of the diagram shows thisrelation, w(P), in inverted form. Finally,the relation w(P), together with WE, determines the equilibrium population size,P", as shown.
Every population size has a specificwage and a specific rate of populationgrowth, b (w) - d (w), associated withit. Inspection of Figure 1 shows that,when population is above equilibrium,mortality exceeds fertility and the population declines; when population is below equilibrium, the opposite occurs.Thus, the equilibrium is stable.
We can be more explicit about thedynamics of the population when particular equations for i. d and wareknown. For example, suppose that theseequations are all linear. Then the differential equation relating populationgrowth rates (PIP) to population sizewill also be linear:
PIP = a - cP (1)
for appropriate a and c. The solution tothis equation is the well-known logisticcurve, applied to human populations byPearl (1924) and many others. It has theform
P(t) = kl(l + e-a t) , (2)
where k = ale is the equilibrium population size.
For further development of this modeland for its estimation, testing and simulation based on historical data, see Lee(1970, 1973, 1974b).
3. CONTROLLED POPULATIONS WITH
Two AGE GROUPS
The model outlined in the last partconverged smoothly and steadily towardequilibrium following a disturbance.
566 DEMOGRAPHY, volume 11, number 4, November 1974
welfare
b(wl
l--__+--;:;- w
bd
io
t
pE\----f
PopulationP
Birthand
dC!othrates
FIGURE I.-Controlled Population without Age Structure
Consider once more the linear case,say with b (L t ) = a - cl.; Then by (5)the equilibrium birth cohort size will be(as - 1)/ (cs2 ) • The elasticity of the fer-
B l = b(LI)LI. (3)
Now the mature age group is made up ofsurvivors of the previous period's births,assuming fixed mortality (1 - s); thus
birth rate. Suppressing the welfare equation, we may write b = b (L t ) , withdb/dL < 0 (since b is positively relatedto welfare and welfare is negatively related to L). Thus,
(5)
(4)
Equations (3) and (4) describe the intergenerational population renewal process.(We assume that transition into andout of the groups occurs only at intervalsof one time unit, which is measured asa generation.)
The equilibrium number of births, BE,is given by the solution to
However, the population had no agestructure, so that the response of laborforce size to a change in the wage ratewas immediate. In fact, of course, thereis something like fifteen or twenty yearsfrom birth to labor force entry andchildbearing. Might this not lead to quitedifferent results? We may approach theproblem in a preliminary way with apopulation made up of two age groups:children and adults.
We assume that long-run changes inmortality are exogenous, which we believe to be generally correct for humanpopulations now and in the past (seeChambers, 1972; Lee, 1970, 1973, 1974b),although in some contexts this may notbe so. Population adjustment, as opposedto population variation, has been effectedthrough the regulation of fertility at welfare levels sufficiently high that densityinduced welfare changes have had littleeffect on mortality. Under this assumption, the model is developed as follows.
The number of children born in periodt, denoted by B t , is the product of thesize of the adult age group L, and its
Dynamics of Controlled Populations
tility function with respect to L is(1 - as). The difference equation will be
B, = (as)B I - 1 - (cl)Bl-1 2• (6)
Now this difference equation is veryclosely related to the logistic differentialequation, since with a little manipulationit can be represented in the form
tiBI/BI = e - fBI, (7)
with e = as - 1, and f = cs,". This ishardly surprising; the question is: Willthe solution also closely resemble thelogistic curve? This can be explored mostsimply with a diagram.
Let us represent the number of personsin the adult age group, L, on the horizontal axis and the number of births onthe vertical axis. Equations (3) and (4)may then be plotted as in Figure 2A.
From a point on the L axis we movevertically to find the number of birthson the B curve. We then advance a timeperiod (one generation or about thirtyyears) by moving horizontally to thenumber of adult survivors on the Lcurve. The process may now be repeated,leading to a steady convergence towardequilibrium from above or below theequilibrium point, indicated by the intersection of the two curves. Recalling thateach horizontal move represents a generation of time, we can plot the convergence of numbers of children or adults asin Figure 2B. The path closely resemblesa logistic.
However, the situation depicted inFigure 2 is not the only one possible. Ifthe birth rate is very sensitive (i.e., elastic) to the welfare of adults, which isin turn sensitive to their numbers, thenfertility may overreact to the size of theparent generation, leading to oscillationsabout equilibrium. This case is illustrated in Figure 3. Near equilibrium, thesize of, each birth cohort is inverselyrelated to the size of the mature agegroup; if the adult age group is aboveequilibrium in one period, it will restrict
567
its fertility so sharply that, when its children become adults, their numbers willbe below equilibrium. Thus each agegroup oscillates about equilibrium, witha period equal to twice the length of ageneration. The system mayor may noteventually converge.
The dynamic behavior in Figure 3 corresponds roughly to recent U. S. demographic history: the small cohorts bornin the 1930's had a very large number ofbirths; the larger cohorts born after the1930's are producing a much smallernumber of births.
There are other possibilities not illustrated in Figures 2 and 3: the equilibriumpoint may be unstable, in which caseviolent oscillations may lead to extinction (if B, :s; 0) ; or a stable "limit cycle"may occur, with perpetual oscillation.However, neither is likely to occur in ahuman population, as we now show.
If we use equation (6) to find thederivative of B, with respect to B t- 1 andevaluate this derivative at equilibrium[B= (as - 1)/cs2], we find it equal to2 - as. From this we may conclude thatfor 1 < as :s; 2 convergence will be directas in Figure 2. When 2 < as :s; 3, convergence will be oscillating, as in Figure 3. When as > 3, the equilibrium isunstable and either extinction or a limitcycle will result.
Let us consider the interpretation ofas. Since a is the maximum generationalbirth rate, occurring when L is verysmall, and s is the constant generationalsurvival rate, as must be the maximumnet reproduction rate, which obtainswhen population is so small that it cangrow essentially without constraint. Sincenet reproduction rates for human populations hardly ever exceed 3, we may conclude that as < 3, and therefore steadyor oscillating convergencewill be the rule.
Malthus looked to the North AmericanColonies to exemplify the power ofhuman reproduction under ideal conditions and argued that population therewas at least doubling every twenty-five
568
2A:- 24
20
16
Births B 12
8
DEMOGRAPHY, volume 11, number 4, November1974
Lt-sBt_1
Bt-LtCa-cLtJ
as" 1.5
a - 1.875s -.8C ...03907
BE- 20
Adults L
ZB: 24
8i rths B
Tim~) in generations
FIGURE 2.-Convergence of a Weakly Controlled Population
years through natural increase. With ageneration length of thirty years, this implies a net reproduction rate (as) of 2.3.When continuous logistic curves are fit toU. S. data, the parameters imply an unconstrained population growth rate of.0314 per year, which corresponds to anet reproduction rate (as) of 2.6 for ageneration length of thirty years (seeDavis, 1963, p. 257).
If we accept the structural homogene-
ity of two centuries of U. S. populationgrowth despite an industrial revolution, increased life expectancy, and varying net migration, then these estimates ofas suggest that the U. S. populationshould conform to the oscillating growthpath of Figure 3B rather than the smoothconvergence of Figure 2B. Indeed, onecan interpret the behavior of U. S. fertility since 1917 in these terms.
We will now extend this analysis of
Dynamics of Controlled Populations 569
3A:2B
Births B
Births B
24
20
16
12
8
4
24
as"" 2..8d - 3.5s = .8c - .141BE - 2.0
4 8 12 16 2.0 24 28Adults L
7 B 9 10
Time) in gCi/nC'lrotions
FIGURE a.-Convergence of a Strongly Controlled Population
the two-age-group case to a populationwith an arbitrarily fine age structure.
4. A GENERAL MODEL OF RENEWAL
FOR CONSTRAINED POPULATIONS
We begin by reviewing the Lotkamodel for renewal of populations subjectto constant age-specific rates. Let Bedenote the number of births at time t;
Pa the proportion of births surviving toage a; and ma the number of births persurviving member of the population ofage a in the appropriate time interval.The product maPa is called the "net maternity function," epa. The sum of epa overall reproductive ages equals the "net reproduction rate," denoted by R (for details, see Keyfitz, 1968, p. 195) .
570 DEMOGRAPHY, volume 11, number 4, November 1974
The population renewal equation, indiscrete form (and using a slightly modified definition of cf>a), is then
45
B, = L cf>.B,-.. (8)0-15
It is well known that such a populationwill converge to steady state growth, withbirths following an exponential path. Therate of growth, called the "intrinsicgrowth rate," may be found as the uniquereal solution for r in the equation
1 = Le-racf>.. (9)
We note that, when the NRR equals 1,then r =0, and the birth series and population converge to a constant level.
The population described by equation(8) is unconstrained; its growth rate isindependent of its size and hence independent of its environment.
As before, we incorporate control inthe model by permitting welfare, andhence fertility, to depend on the population size and age distribution. Since thesefeatures of the population depend entirely on the sequence of previous births(under the assumptions of closure andconstant mortality), and since as beforewe may "solve out" the welfare variable,we can express the control assumption ingeneral form as follows:
m.,t = ma(B'_i, B t - 2 , ••• ,Bt - ,,) , (10)
where w is the oldest age of survival. Fornotational convenience, we may let B,denote the vector (Bt-I, ... , B,_,,) andlet cf>.(B,) = Pam.(B,) denote the netmaternity function. Then (10) may berewritten as
cf>." = cf>,,(B,). (11)
The renewal equation for the controlledpopulation is now
The population is in equilibrium whenthe net reproduction rate R is unity andthe number of births is constant over
time. Thus if B* is the equilibrium birthcohort size and B* denotes the vector B,with all entries equal to B*, then
1 = Lcf>.(B*). (13)
We are interested in models for whichthis kind of equilibrium exists. We nextconsider whether the equilibrium is stableand the kind of fluctuations that wouldbe generated by displacements from equilibrium. To carry out this analysis, wemust first derive a linear approximationto equation (12), which will generallybe nonlinear.
Let c/>. = cf>.(B*) denote the net maternity function in equilibrium. Let 1/. =(aR,1aBt-.)B* denote the elasticity atequilibrium of the net reproduction ratewith respect to the number of birthsa years earlier. Then it is shown in thesecond part of the Appendix that thelinear approximation to (12) is given by
B, == B* + L(c/>. + 1/.)(B,-. - B*). (14)
This equation may be conveniently rewritten in terms of proportional deviations of births from their equilibriumvalue, B*. Thus let b, = iB, - B*) IB" ;then equation (14) can be rewritten as
The result expressed in (15) is surprisingly simple and suggestive, but it isvery general and says nothing of thelikely magnitudes and patterns by age ofthe elasticities 7]a. These will depend onspecific hypotheses about the way thematerial welfare and fertility depend onpopulation size and age structure. Thereare two specifications we will discuss:the "period" model and the "cohort"model.
The period model is essentially a neoclassical growth model with endogenousage-structured population, and it conforms closely to the classical view thatin the long run the supply of labor wasendogenous and infinitely elastic. In theperiod model, each age-specific fertility
Dynamics of Controlled Populations
rate depends on the real wage, and thereal wage varies inversely with the supply of labor. Both relations may incorporate a "time-shift" factor: the fertility-wage relation may shift because ofchanging wage expectations, and thewage-labor relation because of capitalaccumulation and technical progress.These generalizations are discussed insection 1 of the Appendix. It should benoted that we are ignoring all Keynesiancomplications, such as the stagnationistview that population growth may increase income per head by stimulatingdemand. We instead focus on supplyconstraints in a fully employed economy.
The cohort specification formalizesthe view, now accepted by many demographers, that "the relative size of a cohort may be an important influence onits entire life cycle" (Ryder, 1971) andthus that cohort fertility may be inversely related to cohort size. Explanations for this cohort effect on materialwelfare are seldom spelled out explicitlybut might include the effects of lowerhuman capital per head due to the impact of numbers of siblings on healthand LQ. (Wray, 1971) and of crowdingon educational quality, and the effectsof cohort size on wages and promotionin an age-segmented labor market (Keyfitz, 1972; Coale, 1963). Some very preliminary empirical work by Winsborough(1974) suggests the existence of a negative impact of cohort size on wages whenseveral other factors are controlled.
Thus, while the period model assum~s
that all age groups of labor are of urnform quality and perfectly substitutable,the cohort model emphasizes the heterogeneity of the labor force with respect tohuman capital endowments (which areheld to vary inversely with cohort size)and with respect to age proper. A moregeneral specification would incorporateboth period and cohort effects.
Period ModelConsider the size of the labor force,
571
denoted Lt. This will depend on the sizeof surviving birth cohorts (pjB t- h for j= 1, " " w) and on the age-specific participation rates (nj, j = 1, '" , w)which are assumed constant over time.Specifically,
L, = 1:ni(PiBI-i)' (16)
We assume that the demand for laborschedule is constant, so that real wagesand fertility vary inversely with thelabor supply Lt. We thus have
m.,l = m.(L I ) , dm.ldL I < O. (17)
This may be compared to the more general (10) in which age-specific fertilitywas a function of the entire vector ofprevious births; in (17) the effects ofvariations in that vector are summarizedby a single number, Lt.
We now calculate the elasticity "fJa,which equals the product of the elasticityof R; with respect to Lt , and the elasticity of L t with respect to Bt- a . Denotethe elasticity of R, with respect to L,by -/3. The elasticity of L, with respectto Bi.; can be calculated from (16) andequals naPa/:£' njPh which we denote ka.This is simply the proportional share ofage group a in the total labor force inequilibrium. It will often be convenientto assume that nj is constant from age 15to 64 and zero elsewhere, so that
/
64
k. = P. 1:Pi'15
We may now write the approximaterenewal equation for the period model:
b, == 1: (cf>. - {31c.)b l - . , (18)
where ka is the proportion of the totallabor force supplied by age group a inequilibrium, and -{3 is the elasticity ofthe net reproduction rate with respectto labor force size, at equilibrium. Equation (18) completely defines the age pattern of the elasticities "fJa, although it doesnot determine their absolute level, whichdepends on /3.
572 DEMOGRAPHY, volume 11, number 4, November 1974
Cohort Model
In the cohort model, each age-specificrate ma.t is a function of the relevantcohort size Bi.; and is independent of allother cohort sizes. Thus
ma. t = ma(Bt- a) , (19)
and once again the vector of past birthsis reduced to a single number. In thiscase,aRtlaBt- a equals arPa.tlaBt-a. If ,aadenotes the elasticity of net maternityat age a with respect to cohort size, thenn« = -,aarPa' Then by (15) the approximate renewal relation for the cohortmodel is
bt == :E (1 - cxa)cf>abt- a. (20)
Suppose all the age-specific elasticitiesequal the same value, say -ex, so that aone percent increase in the size of a cohort would reduce its completed fertility,and its fertility at each age, by ,a percent. Then the renewal equation wouldtake on the particularly simple form
b, == (1 - cx) :E cf>abt- a. (21)
This last equation again determines theage pattern of the 7]a, while leaving theirabsolute level, which depends on 'lX,
indeterminate.
Other ApproachesThere are, of course, many other pos
sible models. The work of Easterlin,which has been widely discussed and hasbeen formalized by Keyfitz (1972), emphasizes the size of cohorts relative toone another, rather than relative to anequilibrium size. Such models imply anequilibrium growth rate but are compatible with any corresponding growth pathor population size. For this reason theyare inconsistent with the approach of thispaper, which emphasizes environmentalconstraints. Such models may accuratelyportray the dynamics of population renewal over a period of several generations, and their conceptual unsuitabilityfor the very long run may be of little
practical importance. Similarly, the kindof model presented above may be appropriate for some hypothetical long runbut may not capture the dynamic behavior over the shorter period for whichthe interplay of age structure and constraint is really of interest. We, ofcourse, hope that this is not so.
5. CONSTRAINED POPULATIONS SUBJECT
TO STOCHASTIC DISTURBANCE
We must now acknowledge the obviousfact that the net maternity rate is notcompletely determined endogenously byB, but also reflects exogenous factorssuch as climate, business cycles, war, andpure demographic randomness. We mayformalize this indeterminancy by writingcf>a.t = cf>a(B t) +ea.t•The net reproductionrate, Ru which equals the sum of theage-specific net maternity rates, is thengiven by R, = R(B,) + :E ea • l • It willbe convenient to denote this last term,which is the sum of all the individualperturbations in the age-specific netmaternity rates, by e..
The renewal equation is now
n, = L [cf>a(B t) + ea.t]Bt- a. (22)
Recalling the definitions of bt and ei,this can be rewritten as,
bt == L: (cf>a + 7]a)bt- a
+ L ea.tbt - a + et· (23)
The second summation is composed ofterms which are second order in deviations from equilibrium and which maytherefore be ignored for small variations.This leaves us with the following simpleapproximation for the renewal processin a stochastically disturbed constrainedpopulation:
b, == L: (4)0 + 'T/a)b t - a + e., (24)
where e, is the random variation in thenet reproduction rate let = R, - R(Bt)].This result is perfectly general and soapplies to the period and cohort specifications of (18) and (21). (This stochastic
Dynamics of Controlled Populations
renewal equation, with or without controlpresent, may be used to generate population forecasts and their confidence intervals [see Lee, 1974a].)
The two-age-group model of section 3provides a simple illustration of all theanalyses so far. The net reproductionrate R, is given by B,/B'_I = as cs2B t-!. Its elasticity with respect toB'_I' at equilibrium, is '171 = 1 - as.In equilibrium we will also have R =CPI = 1. So by (24) the approximaterenewal equation is
b, == (2 - as)bt - I + e.. (25)
This is a first-order Markov processwhich will either tend to converge steadily toward equilibrium (as in Figure 2)if 1 < as ~ Z or tend to oscillate whileconverging to equilibrium if Z < as ~ 3(as in Figure 3). If as >3, the oscillations will be explosive. These are thesame conclusions we reached in section 3,except that in a linear system no limitcycle can occur.
6. POPULATION FLUCTUATIONS
In the preceding part we derived thelinear stochastic difference equation (24)approximately describing the renewalprocess for a constrained population. Thenext step is to find the dynamic implications for population renewal by analyzing the coefficients in the equation.
Demographers have studied fluctuations principally in deterministic populations with constant vital rates (seeBernardelli, 1941; Keyfitz, 1965; Coale,1972; LeBras, 1969; Pollard, 1973), although there have been important exceptions (LeBras, 1971; Keyfitz, 1972;Coale, 1972). The method has been toderive an explicit expression for futurebirths as a function of time,given aninitial age distribution. The expressioncontains an exponential trend, withdamped oscillations superimposed; andthe periodicity of the oscillations provides insights into the pattern of fluctuation of births about trend. However,
573
this approach has serious shortcomings:it is conditional on a particular initialage structure; it is unrealistic in assuming constant vital rates; in actual populations, fluctuations are observed in theabsence of the severe initial distortionrequired by this model; and actual population waves are much more persistentthan those of the model, which dampvery rapidly. An adequate treatment ofthe problem must be based on a stochastic model.
There are several ways to analyze thedynamics of stochastic linear differenceequations; for a discussion of the relations among them, see Howrey (1968).We will use two tools: the theoreticalautocorrelation function and the theoretical spectrum. The autocorrelation isthe correlation of the process with itselfat each lag; for example, the autocorrelation of the b-series at lag m, denotedr,.., is the covariance of b, and bi:«(which for a stationary process is independent of t) divided by the varianceof b. The plot of r; against m is calledthe correlogram. Periodic oscillations inthe correlogram indicate a tendency forthe series to oscillate with the sameperiod. The autocorrelation, as a functionof the lag, can be written as the weightedsum of the same damped oscillatory components as occur in the deterministicexpression of the classic demographicanalysis. However, the weights are independent of any initial age distribution, and therefore the analysis is moregeneral.
The use of the autocorrelation functioncan be illustrated with the two-age groupmodel, as given in (25). The process willbe stationary when 1 < as < 3. Theautocorrelation function is given by:r-:> (2 - as)m. When 1 > (2 - as) > 0,rm will decline exponentially to zero, andno oscillations will be generated. Butwhen 2 - as < 0, rm will oscillate aboutzero with a period of two generations.This indicates that the population, whensubject to continual random shocks, will
574 DEMOGRAPHY, volume 11, number 4, November 1974
continually exhibit waves of roughly twogenerations length.
To what extent may these results begeneralized to models with many agegroups? The two-age-group model, byconstruction, can exhibit only the control cycle and excludes the generationalcycle. However, it will still be of interestto compare its behavior to that of morecomplete models to determine whetherthe preconditions of the control cycle andits period are similar.
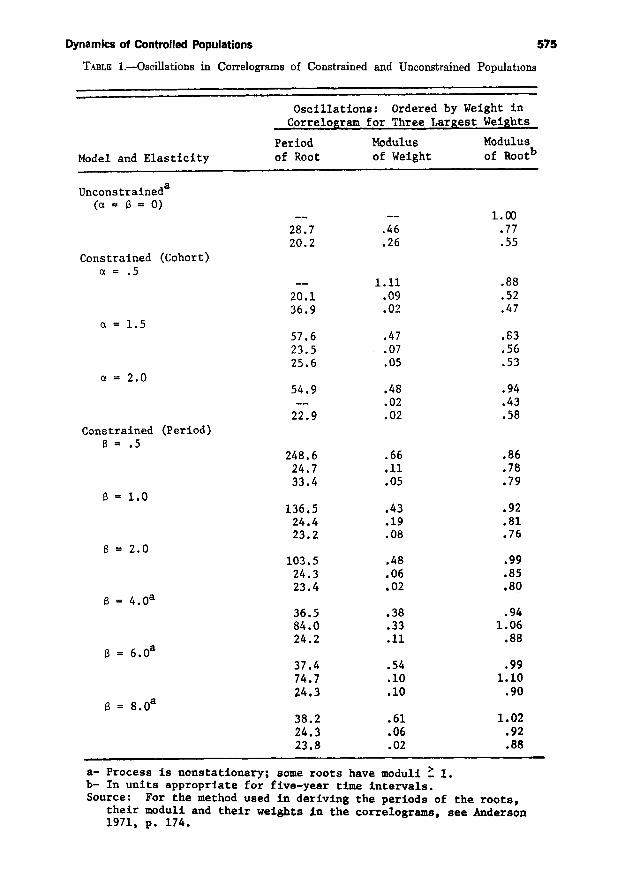
Table 1 shows the periods of the rootsof the characteristic polynomials, theirmoduli, and their weights in the correlogram for each model (for the methodused to derive these, see Anderson, 1971).The roots are ranked according to theirweights, and only the three most important are shown. We note that theuncontrolled model has a root with unitmodulus and is hence nonstationary.Otherwise nonstationarity due to explosive oscillations arises from high controlelasticities (e.g., for values of 13 abovetwo).
Our major interest is in the dominantperiodicities and the way in which theyare affected by control. For the uncontrolled model (ex = 13 = 0) there is onlya generational cycle, with period roughlyequal to the mean age of the net maternity function. This corroborates theresults of previous deterministic analyses. For the cohort-controlled model witha control elasticity (a) of .5, the generational cycle virtually disappears, and thecorrelogram is dominated by an exponential decline to zero, indicated by thereal root. This is entirely consistent withthe two-age-group model with 1 < as <2. For the cohort-controlled model withcontrol elasticity greater than one, theprocess exhibits a control cycle of twogenerations length, and the generationalcycle is of little importance. Thus in thiscase also, the two-age-group model provides an accurate representation of thedynamics.
The behavior of the period control
model (which we prefer) is much morecomplicated. For control elasticities (13)less than one, there is a very long controlcycle of well over a hundred years; andsuperimposed on it is a modified generational cycle of about twenty-five years.As the elasticity increases, the controlcycle period becomes steadily shorterand loses importance relative to themodified generational cycle. At 13 = 4,which is unstable, the process is dominated by fluctuations of 36.5 and 84.5years. Above this elasticity, the controlcycle and generational cycle merge at aperiod of about thirty-eight years, whichdominates all else. We note that withthe period model the elasticity must bevery high to produce a control cycle neartwo generations in length; and when ashort control cycle does emerge, its period is substantially less than two generations. The two-age-group model nolonger provides a satisfactory simplification.
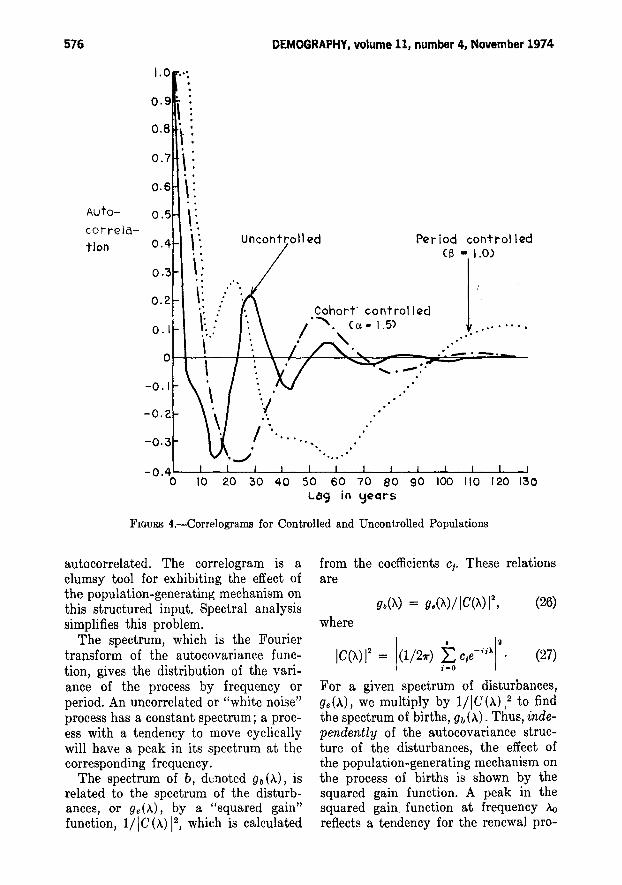
We now consider Figure 4, which showsseveral theoretical correlograms. For theuncontrolled model, the pseudo-correlogram shows sharply damped oscillationsof about twenty-eight years length, asexpected. The correlogram for ex = 1.5shows a damped 57.6-year cycle andconforms closely to the correlogram of afirst-order Markov process with ri =- .37 and time measured in twenty-eightyear units. The correlogram for the period model with 13 = 1 shows a 24.4-yeargenerational cycle superimposed on a136-year control cycle.
As we have mentioned, the correlogram interests us because its oscillationsreflect a tendency for the process to oscillate with the same periodicity. However, the correlogram depends not onlyon the coefficients OJ but also On the autocorrelation structure of the disturbanceterm, e, which we have implicitly assumed to be serially uncorrelated. Infact, however, disturbances reflect economic, meteorological and epidemeological conditions, each of which is itself
Dynamics of Controlled Populations
TABLE i.-Oscillations in Correlograms of Constrained and Unconstrained Populauons
a- Process is nonstationary; some roots have moduli ~ 1.b- In units appropriate for five-year time intervals.Source: For the method used in deriving the periods of the roots,
their moduli and their weights in the correlograms, see Anderson1971, p. 174.
575
576 DEMOGRAPHY, volume 11, number 4, November 1974
o 10 20 30 40 50 60 70 80 90 100 110 120 130Lag in \jeers
FIGURE 4.-Correlograms for Controlled and Uncontrolled Populations
autocorrelated. The oorrelogram is aclumsy tool for exhibiting the effect ofthe population-generating mechanism onthis structured input. Spectral analysissimplifies this problem.
The spectrum, which is the Fouriertransform of the autocovariance function, gives the distribution of the variance of the process by frequency orperiod. An uncorrelated or "white noise"process has a constant spectrum; a process with a tendency to move cyclicallywill have a peak in its spectrum at thecorresponding frequency.
The spectrum of b, denoted gb (A), isrelated to the spectrum of the disturbances, or ge(A), by a "squared gain"function, 1/IC(A) 12 , which is calculated
from the coefficients Ci' These relationsare
where
IC(>-)12 = 1(1/2'11) ~Cje-ijXr, (27)
For a given spectrum of disturbances,ge (A), we multiply by 1/ IC (A) 1
2 to findthe spectrum of births, gb (A). Thus, independently of the autocovariance structure of the disturbances, the effect ofthe population-generating mechanism onthe process of births is shown by thesquared gain function. A peak in thesquared gain function at frequency Aoreflects a tendency for the renewal pro-
Dynamics of Controlled Populations
cess to create cycles of frequency x, andperiod l/Ao, out of random disturbance.When the disturbance term, e, is seriallyuncorrelated, then its spectrum is a horizontal line, and the squared gain func-
577
tion represents exactly the shape of thespectrum of births.
Figure 5 plots squared gain againstfrequency for the cohort-controlled modelwith various elasticities, a, including zero
10090807060
50
40
30
987
.~54.5 years
I~i \..I \
II,I
2 3 4 5 6
Cycles per centurlj
FIGURE 5.-Squared Gain for the Cohort Model
o
3
2
109876
5
4
1.0f--4-#---~1-----~~~~=~"!:30_-----.9
F.8.7.6
.5
s9u~red 20Gain
578 DEMOGRAPHY, volume 11, number 4, November 1974
(no control). We see that the uncontrolled population amplifies variation atperiods of about twenty-eight years, andalso at very long periods (low frequencies). (For a much more detailed discussion of the squared gain of an uncontrolled population in a nonstochasticcontext, see Coale, 1972.) This amplification at long periods reflects the absence of control; when fertility remainsabove equilibrium for a long time, thebirth series will grow exponentially without restraint. The effect of cohort control with elasticities (a) less than oneis to flatten out the spectrum and reducethe generational cycle, or echo. Note inparticular that low frequency (long period) fluctuations are drastically reducedin amplitude, which is the expected effectof control. When the elasticity is aboveone, there is an amplification of varianceat periods of two generations; the echodisappears and is replaced by a controlcycle.
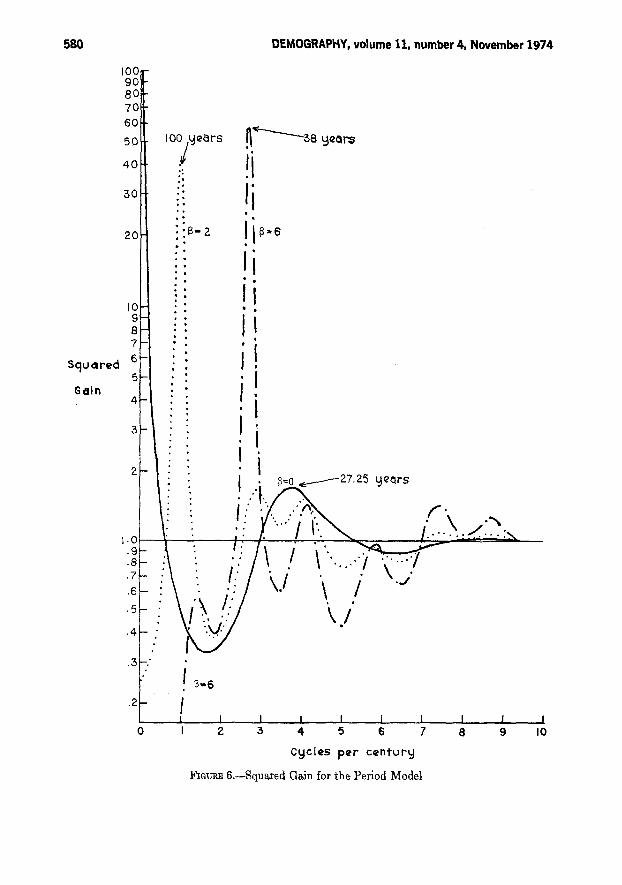
Figure 6 shows the inverse squaredgain for f3 = 2 and 6 in the period model.At f3 = 2, there is a very strong controlcycle with a period of one hundred years,which swamps the generational cycle,represented by two small peaks. Whenf3 =6, there is a very strong control cyclewith period thirty-eight years. For thismodel also, we note the attenuation ofall low frequency variations, in sharpcontrast to the uncontrolled model whichamplifies them.
Let us recapitulate the main results ofthis part. The dynamics of controlledpopulations, are, indeed, quite differentfrom those of uncontrolled populations,at least for moderately high control elasticities. Uncontrolled populations tend tofluctuate with a period of one generation.In cohort-controlled populations, evenwith low elasticities, the importance ofthis generational cycle is sharply reduced. When the control elasticity exceeds one, the generational cycle disappears altogether and is replaced by acontrol cycle twice as long. This latterresult is sufficiently strong and un-
ambiguous to permit inference from observed dynamics to the presence orabsence of strong control, provided thecohort model is an appropriate specification. We believe, however, that theperiod-control model is more reasonable.And unfortunately the dynamics of thismodel are more complicated and provideless clearcut indications. For low elasticities, the generational cycle remains,superimposed on a very long controlcycle of over a century. At very highelasticities there is a single cycle withperiod less than two generations; butbefore this appears, the system becomesunstable.
7. PRELIMINARY RESULTS FOR THE
UNITED STATES
The empirical analysis which is undertaken in this part is intended to be illustrative and suggestive, not a formaltest or demonstration. We begin with adiscussion of the correlogram of birthsfor the United States, 1900 to 1972, anda comparison of this correlogram withthe correlogram of baptisms for a preindustrial French parish, 1590-1800. Wethen estimate the fertility relations assumed by each of the two models wedeveloped and consider whether the estimated relations are consistent with theobserved dynamic behavior.
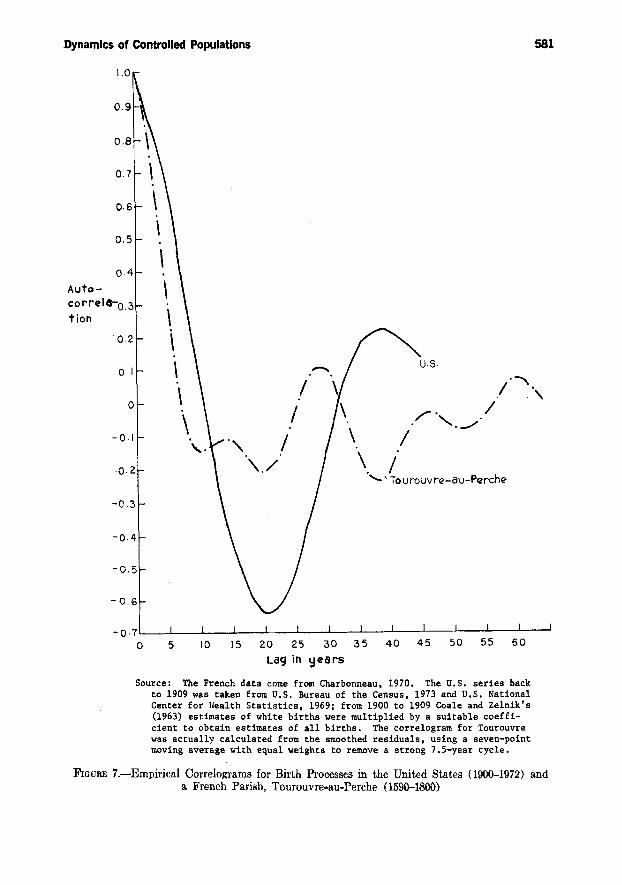
The correlograms for the United Statesand Tourouvre-au-Perche are shown inFigure 7. These are calculated from theproportional deviations of births fromexponential trend, estimated as the residuals from the regression of the logof annual births on time. Tourouvre waschosen arbitrarily from those parisheswith long published series. The correlogram for Tourouvre shows a twentynine-year cycle, which is also found,plus or minus a few years, in the otherthree parish baptism series I have analyzed. The work of the previous sectionshows that these would be characteristicof uncontrolled or weakly controlledpopulations, though the possibility thatthese cycles are forced oscillations due
.812.
(29)
Dynamics of Controlled Populations
to climate or prices cannot yet be entirely ruled out. The fifteen-year cyclewhich the eorrelogram also reveals isalmost certainly due to a correspondingcycle in grain prices.
The correlogram for the United States,on the other hand, shows a strong peakat about thirty-nine years. Relative tothe hypothesized "cycle" length the seriesis much too short for accurate estimation. But taken at face value, the estimate conforms very closely to the predicted cycle length of thirty-eight yearsgenerated by the period model with ahigh control elasticity. According to themodel (see Table 1), the oscillationsshould be explosive, but the empiricalcorrelogram cannot provide informationon this score.
We now consider the estimated fertility equations. The first model is basedon the assumption that period fertilityrates depend on the size of the (discounted) potential labor force, as inequation (17). To test this assumption,we first find the deviations of potentiallabor force size from exponential trendby calculating the residuals of the regression of the log of the population aged15 to 64 on time. (We use Potentialrather than actual labor force size, because it is the base population size inrelation to available nonlabor inputswhich determines the feasible set of aggregate income-leisure choices for thesociety.) Then the log of the total fertility rate (equal to twice the sum of allthe ma,t for period t) is regressed on theseresiduals. The results are shown belowfor U. S. data, 1917 to 1972, aggregatedinto eleven observations of quinquennialaverages (the standard errors of the estimates are shown in parentheses) :
In (P/S-64)
= 11.102 + .01l4t + el , R 2 = .988;
(.0142) (.000419) (28)
In (TFR.)= 7.942 - 7.724e. + U" R 2
=
(.0247) (1.241)
579
We thus find a highly significant estimate of 7.7 for [3, and 80 percent of thevariance in fertility is "explained." When[3 actually has this value in the periodmodel, births tend to move in an explosive thirty-eight-year cycle. Thus, theestimated elasticity is strikingly consistent with the observed dynamic behavior,which helps to confirm the model. On theother hand, if this fluctuation in fertility were forced on the system by exogenous factors, then we would necessarilyestimate a high, though spurious, elasticity [3. The consistency of the estimated [3 with the dynamic behavior cutsboth ways.
The second model is based on the assumption that cohort fertility rates aredependent on cohort size, as in equation(19). To test this hypothesis, we firstobtained the residuals from the regressionof the log of cohort size on time, as ameasure of the deviation of cohort sizefrom its stable growth path. We usedfive-year birth cohorts, 1890-1894 to1950-1954. The estimates given in equations (30) .and (31) are based on cohortsize measured at ages 0-4. We found verysimilar results measuring cohort size atages ~24, which reflects changes inmigration and mortality, as well as insize at birth.
We then regressed the "completed" cohort fertility on the residual from equation (30). For many cohorts this involved some projection of fertility, butfor all except the last, which had borneonly about half its expected number ofchildren, this cannot have led to largeerrors. We followed Ryder (1971) forcohorts through 1919; after that we followed U. S. Bureau of the Census (1972).
!i~ i!i~a6Iii i·.II· iI .· II .i ~· II .· II .-· :~. .
!/ .....·......I·:~( r: \ ."".' ......~ ... :
\. .' \....... ,,/ . '\..'/\/ " i .J
\j
cycles per century
FIGURE 5.-Squared Gain for the Period Model
Dynamics of Controlled Populations 581
1.0
504540
.,/ '. '\
r: /. '- .../'/
, /........ ' tcurouvre-eu-Perche
20 25 30 35
Lag in ~ears
1510
0.8 \0.7 \0.6 \0.5
\
\0.4
Auto- \correI4-0.3
\tion
0·2 \01 \
0 \
-0.1\
-0.2
-0.3
-0·4
-0.5
- 0.6
-0·70 5
Source: The French data come from Charbonneau, 1970, The U.S. series backto 1909 was taken from U.S. Bureau of the Census, 1973 and U.S. NationalCenter for }Lealth Statistics, 1969; from 1900 to 1909 Coale and Zelnik's(1963) estimates of white births were multiplied by a suitable coefficient to obtain estimates of all births. The correlogram for Tourouvrewas actually calculated from the smoothed residuals, using a seven-pointmoving average with equal weights to remove a strong 7.S-year cycle.
FIGURE 7.-Empirical Correlograms for Birth Processes in the United States (1900-1972) anda French Parish, Tourouvre-au-Perche (1590--1800)
582 DEMOGRAPHY, volume 11, number 4, November 1974
This model also gives a good fit,though not so close as the previous one.However, in this case the estimated elasticity, - .954, is not consistent with thedynamic behavior of the system. Anelasticity of -1 would immediatelydamp any disturbance, and the expectednumber of births would be the same, regardless of cohort size. The model withthis estimated value of a is thus incapable of generating the observed patternof fluctuation in births or fertility. Apparently, therefore, this version of themodel is inconsistent with the dynamicbehavior of population in the UnitedStates.
8. DISCUSSION
Two distinct kinds of cycles may arisein populations: the "generational" cycleor "echo" due to the intrinsic dynamicsof population renewal as an age-structured process and the "control" cycledue to the lagged operation of equilibrating mechanisms. In populations growingwithout effective constraint, such asmight occur temporarily in newly settledareas, only the generational cycle willarise. This case has previously been analyzed in a deterministic context; we haveextended the analysis to a stochasticcontext and derived the theoretical correlogram and spectrum of the birth process. The cycles are shown to arise fromthe "filtering" of random variations bythe age structure of reproduction, ratherthan from the repercussions of a singledistorting shock. '
When homeostatic control is effectivelypresent, the pattern of fluctuation depends on the form of the control mechanism and its sensitivity. We have considered two kinds of mechanisms. In theperiod model, the labor supply exertsdownward pressure on welfare and fertility in each time period, while in thecohort model the size of each cohortaffects its welfare and fertility throughout its life.
When control is weak, the period
model continues to exhibit generationalcycles, although these are superimposedon a very long control cycle of one ortwo centuries periodicity. In the cohortmodel, no control cycle emerges, and thegenerational cycles are strongly damped.We conclude that, when control is weak,it will be difficult or impossible to drawconclusions about possible control froman analysis of population fluctuations; itwill be necessary to examine their longrun behavior.
When control is strong, the cohortmodel exhibits strikingly different behavior; the generational cycle disappearsand is replaced by a control cycle oflength two generations. As the controlelasticity of the period model increases,it first becomes unstable and then develops a very strong thirty-eight-yearcycle. We conclude that at high controlelasticities either system would presentsufficiently distinctive dynamics to berecognizable.
Preliminary empirical analysis suggests that only the generational cyclewas prominent in preindustrial Europe.This would indicate relatively low control elasticities, which is consistent withthe low long-run elasticity estimated forpreindustrial England by Lee (1970,1973, 1974b). But in the nineteenth century, dominated by the Kondratieff longcycle, the generational cycle is conspicuously absent. Thus the correlogram ofthe birth series for nineteenth-centuryFrance (not shown in this paper) has asingle strong peak at fifty-five years.And in the twentieth century, the UnitedStates, relatively undisturbed by war,has a single and very strong peak in itsbirth correlogram at thirty-nine years.Might these longer fluctuations representcontrol cycles? Is it possible that thelaws of motion for demographic processes underwent a radical change withindustrialization and/or the transition tolower fertility? Are the populations ofmodern industrial nations, far fromgrowing out of control, actually subject
(A.8)
Dynamics of Controlled PopulatiOns
to a more sensitive regulation than theirpreindustrial predecessors? These arehighly speculative hypotheses, based onthe slenderest evidence, but they invitefurther investigation.
ApPENDIX
Analysis of a Growing System
We have developed and analyzed amodel of population renewal under theassumption that the equilibrium population size was constant. We now supposethat the constraint to which the population is subject is receding at a constantrate u, as the result of capital accumulation, technical change, increasing qualityof labor, or other unspecified factors.Then the population size which is consistent with any given level of welfareis increasing at rate u. To illustrate forthe model of section 2, without age structure, we would have U', = U'(e-u,p,) .
We may also suppose that the relationbetween fertility and welfare is changing with time at some constant rate v,due to rising expectations; then the levelof welfare necessary to induce a givenlevel of fertility increases at a rate v. Inthe model without age structure, wewould have b, = b(e--v'U',). The systemwill have a steady state growth rate rfor population, which will depend on therates u and v and on the functiondetermining U'.
Now let us consider the more generalage-structured model of section 4. Let rdenote the steady state growth rate ofpopulation and of births resulting fromthe interplay of changes in productivityand in expectations. When the stream ofbirths grows at rate r, the net maternityfunction will be constant. The net maternity function, then, may be writtenas follows:
¢a.,(B,)
- A. [e-r<c-IlB . . . -r<C-w)B 1- '/'0 '-I" e c-,. ,
(A.l)
583
or, to slightly extend our earlier notation,
ePa.,(B,) = ePo(e-r'B,). (A.2)
The renewal equation based on (A.2) is
B, = L ePo(e-rIBc)B,_o. (A.3)
Now consider the transformed birthsequence, B,*, and the net maternityfunction, ePa* (B,*) , defined, respectively,as follows:
and
ePo*(B,*) = e-r°ePo(B,*). (A.5)
Substituting into the renewal equation(A.3) and dividing both sides by e:"we find
B,* = L ePo*(B,*)B,- o*' (A.6)
But this is precisely the equation analyzed in section 4.
In section 6 we showed how to determine the autocorrelation function and thespectrum for birth fluctuations in a constrained system with a constant equilibrium, where fluctuations were measured as proportional deviations of birthsfrom equilibrium. The analysis wasbased on coefficients (ePa + "1a) in equation (15). Our discussion in this part ofthe Appendix has shown that the entireanalysis in section 6 carries over to agrowing system; we need only multiplyeach coefficient in (15) by e-ra and proceed as before.
Derivation of the Linear Approximation
The linear approximation is given by
B, == B* + L (fJB,/fJB'_o)(B,_o - B*),(A.7)
where the partial derivatives are to beevaluated at equilibrium. These partialderivatives may be calculated from (12) I
yielding
fJB,/fJB,-o = ePo(B,)
+ L [fJeP;(B.)/fJB,-.]B,-i-
n; = (B*/1.0) (aRt/aBt-a) IB,-B>. (A.11)
It therefore follows from (A.9) that
DEMOGRAPHY, volume 11, number 4, November 1974
Chambers, J. D. 1972. Population, Economyand Society in Preindustrial England. Oxford: Oxford University Press.
Charbonneau, Hubert. 1970. Tourouvre-auPerche Aux XVII" et XVIII" Siecles. Paris:Presses Universitaires de France.
Coale, Ansley. 1963. The Economic Effects ofFertility Control in Underdeveloped Areas.In Roy Greep (ed.), Human Fertility andPopulation Problems. Cambridge: Schenkman Publishing Co.
--. 1972. The Growth and Structure ofHuman Populations: A Mathematical Investigation. Princeton: Princeton UniversityPress.
--, and Melvin Zelnik. 1963. New Estimates of Fertility and Population in theUnited States: A Study of Annual WhiteBirths from 1855 to 1960 and of Completeness of Enumeration in the Censuses from1880to 1960. Princeton: Princeton UniversityPress.
Davis, Harold T. 1963. The Analysis of Economic Time Series (reissued ed.). San Antonio: Principia Press of Trinity University.
Easterlin, Richard. 1962. The American BabyBoom in Historical Perspective. Occasionalpaper no. 79. New York: National Bureaufor Economic Research.
--. 1968. Population, Labor Force, andLong Swings in Economic Growth. NewYork: National Bureau for Economic Research.
--, and Gretchen Condran. 1974. A Noteon the Recent Fertility Swing in Australia,Canada, England and Wales, and the UnitedStates. In Migration, Foreign Capital andEconomic Development: Essays in Honor ofBrinley Thomas. Forthcoming.
Goubert, Pierre. 1965. Recent Theories andResearch in French Population between 1500and 1700. Pp. 457-473 in D. V. Glass andD. E. C. Eversley (eds.). Population in History. Chicago: Aldine Publishing Co.
Grauman, John. 1960. Comment. In NationalBureau for Economic Research (ed.), Demographic and Economic Change in DevelopedCountries. Princeton: Princeton UniversityPress.
Howrey, P. E. 1968. A Spectrum Analysis ofthe Long-Swing Hypothesis. InternationalEconomic Review 9 :228-252.
Keyfitz, Nathan. 1965. Estimating the Trajectory of a Population. Proceedings of theFifth Berkeley Symposium on MathematicalStatistics and Probability 4 :81-113.
__. 1968. Introduction to the Mathematicsof Population. Reading: Addison-Wesley.
__. 1972. Population Waves. Pp, 1-38 inT. N. E. Greville (ed.). Population Dynamics. New York: Academic Press.
(A.12)
(A.l4)
REFERENCES
Evaluated at equilibrium, this is
584
L: ar/>;(B,)/OBt-a = aRt/aBt-a, (A.lO)
and in equilibrium R equals one. Theelasticity at equilibrium of R, with respect to Bt-a, which we denote 7Ja, therefore is given by
aBt/OBt-a IB,-B> = r/>a + 1/a,
and hence from (A.7) that
Anderson, T. W. 1971. The Statistical Analysisof Time Series. New York: John Wiley &Sons.
Birdsell, Joseph. 1957. Some Population Problems Involving Pleistocene Man. ColdSprings Harbor Symposia on QuantitativeBiology 22:47-69.
Bernardelli, Harre. 1941. Population Waves.Journal of the Burma Research Society31 :1-18.
aBt/aBt-a = r/>a + B*
..l: ar/>;(Bt)/aBt-a IB,-B>. (A.9)
Now by definition of R, we have
ACKNOWLEDGMENTS
The research on which this paper isbased was supported by the PopulationStudies Center of the University ofMichigan. Professors Saul Hymans andPhil Howrey and two referees made useful comments on an earlier draft. Theauthor gratefully acknowledges the research assistance of Martha Hill andDavid Levinson and the typing andpreparation of diagrams by CarolynCopley.
B, == B* + L: (r/>a + 1/a)(Bt-a - B*).
(A.l3)
Finally, letting bt = iB, - B*)/B*,(A.l3) implies that
Dynamics of Controlled Populations
Le Bras, Herve. 1969. Retour d'une population a l'etat stable apres une "catastrophe."Population 24:861--s96.
--. 1971. Elements pour une theorie despopulations instables. Population 26:525-572.
Lee, Ronald. 1970. Econometric Studies ofTopics in Demographic History. Unpublished Ph. D. dissertation. Cambridge: Harvard University.
--. 1973. Population in Preindustrial England: An Econometric Analysis. QuarterlyJournal of Economics 87:581--Q07.
--. 1974a. Forecasting Births in Post-Transition Populations: Stochastic Renewal withSerially Correlated Fertility. Journal of theAmerican Statistical Association 69:607-617.
--. 1974b. Models of Preindustrial Population Dynamics with Applications to England. In Charles Tilly (ed.), Early Industrialization, Shifts in Fertility and Changesin Family Structure. Forthcoming. Princeton: Princeton University Press.
Leibenstein, Harvey. 1963. Economic Backwardness and Economic Growth. New York:John Wiley & Sons.
Liu, Paul. 1971. Marshall's Economic Theoryof Population. Unpublished manuscript. EastLansing: Department of Economics, Michigan State University.
Malthus, Thomas Robert. 1970. An Essay onthe Principle of Population (1798). Editedby Anthony Flew. Baltimore: PenguinBooks Inc.
Pearl, Raymond. 1924. Studies in Human Biology. Baltimore: Williams & Wilkins.
Pollard, J. H. 1973. Mathematical Models forthe Growth of Human Populations. Cambridge: Cambridge University Press.
Ryder, Norman B. 1971. The Time Series ofFertility in the United States. In Interna-
585
tional Union for the Scientific Study ofPopulation (ed.), International PopulationConference, London, 1969. Vol. 1. London:International Union for the Scientific Studyof Population.
Sauvy, Alfred. 1969. General Theory of Population. New York: Basic Books.
Solow, Robert. 1956. A Contribution to theTheory of Economic Growth. QuarterlyJournal of Economics 79:65-94.
Simon, Julian L. 1974. The Effects of Incomeon Fertility Control. Chapel Hill: CarolinaPopulation Center.
U. S. Bureau of the Census. 1972. Projectionsof the Population of the United States, byAge and Sex: 1972 to 2020. Current Population Reports, Series P-25, No. 493. Washington, D. C.: Government Printing Office.
--. 1973. Statistical Abstract of the UnitedStates: 1973. Washington, D. C.: Government Printing Office.
U. S. National Center for Health Statistics.1969. Vital Statistics of the United States:1967. Vol. 1. Natality. Washington, D. C.:Government Printing Office.
Winsborough, H. H. 1974. Age, Period, Cohortand Education Effects on Earnings by Race.In Kenneth Land and Seymour Spilerman(eds.), Social Indicator Models. Forthcoming.New York: Russell Sage Foundation.
Wray, Joe D. 1971. Population Pressure onFamilies: Family Spacing. In National Academy of Sciences (ed.), Rapid PopulationGrowth: Consequences and Policy Implications. Baltimore: Johns Hopkins Press.
Wrigley, E. A. 1969. Population and History.New York: McGraw-Hill.
Yule, G. Udney. 1906. Changes in the Marriageand Birth Rates in England and Wales During the Past Half Century. Journal of theRoyal Statistical Society 69:18-132.