B R A I N R E S E A R C H 1 3 6 7 ( 2 0 1 1 ) 3 4 0 – 3 4 6
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
The Fos expression in rat brain following electrical stimulationof dura mater surrounding the superior sagittal sinus changedwith the pre-treatment of rizatriptan benzoate
Xiaolin Wang, Shengyuan Yu⁎, Zhao Dong, Lei JiangDepartment of Neurology, Chinese PLA General Hospital, Beijing 100853, PR China
Article history:Accepted 1 October 2010Available online 8 October 2010
Fos expression in the brain was systematically investigated by means ofimmunohistochemical staining after electrical stimulation of the dura mater surroundingthe superior sagittal sinus in conscious rats. Fos-like immunoreactive neurons are distributedmainly in the upper cervical spinal cord, spinal trigeminal nucleus caudal part, raphemagnusnucleus,periaqueductal gray, ventromedialhypothalamicnucleus, andmediodorsal thalamusnucleus. With the pre-treatment of intraperitoneal injection of rizatriptan benzoate, thenumberofFos-like immunoreactiveneuronsdecreased in the spinal trigeminalnucleuscaudalpart and raphe magnus nucleus, increased in the periaqueductal gray, and remainedunchanged in the ventromedial hypothalamic nucleus and mediodorsal thalamus nucleus.These results provide morphological evidence that the nuclei described above are involved inthe development and maintenance of the trigeminovascular headache.
Over the past decades, much work has been done to help usunderstand themigrainemechanism.Although it is still unclear,the activation of the trigeminovascular nociceptive pathway isconsidered tobeessential for thedevelopmentandmaintenanceof a migraine headache (Tepper et al., 2001). Many migraineanimal models were constructed using electrical stimulation oftrigeminal nerve-innervating tissues, such as trigeminal gangli-onmodels (Clayton et al., 1997; Goadsby and Knight, 1997), duralarteries models (De Vries et al., 1999), and dura mater andsuperior sagittal sinus models (Benjamin et al., 2004). Thesemodels have helped to greatly expand our knowledge of themechanisms underlying the trigeminovascular system inmigraines. However, they were all performed in anesthetized
com (S. Yu).
er B.V. All rights reserved
animals. As an important therapeutic method for unbearablepain, anesthesia has a great deal of influence on the nociceptiveinformation transmission and analysis during the central painprocess. This would unquestionably change the property orintensity of the pain perceived, disregarding the automaticnervous system and the emotional responses complicated withpain. In order to better mimic the physical state of a migrainepatient, the electrical stimulation of duramater surrounding thesuperior sagittal sinus (SSS) was conducted in conscious rats inthis study.
Fos is the protein product of activated c-fos proto-oncogene.It has been reported to be a third-messenger molecule thatconnects extracellular signals to genetic events that result inchanges in cellular phenotypic expression (DragunowandFaull,1989; Rodella et al., 1998). Various types of peripheral stimuli,
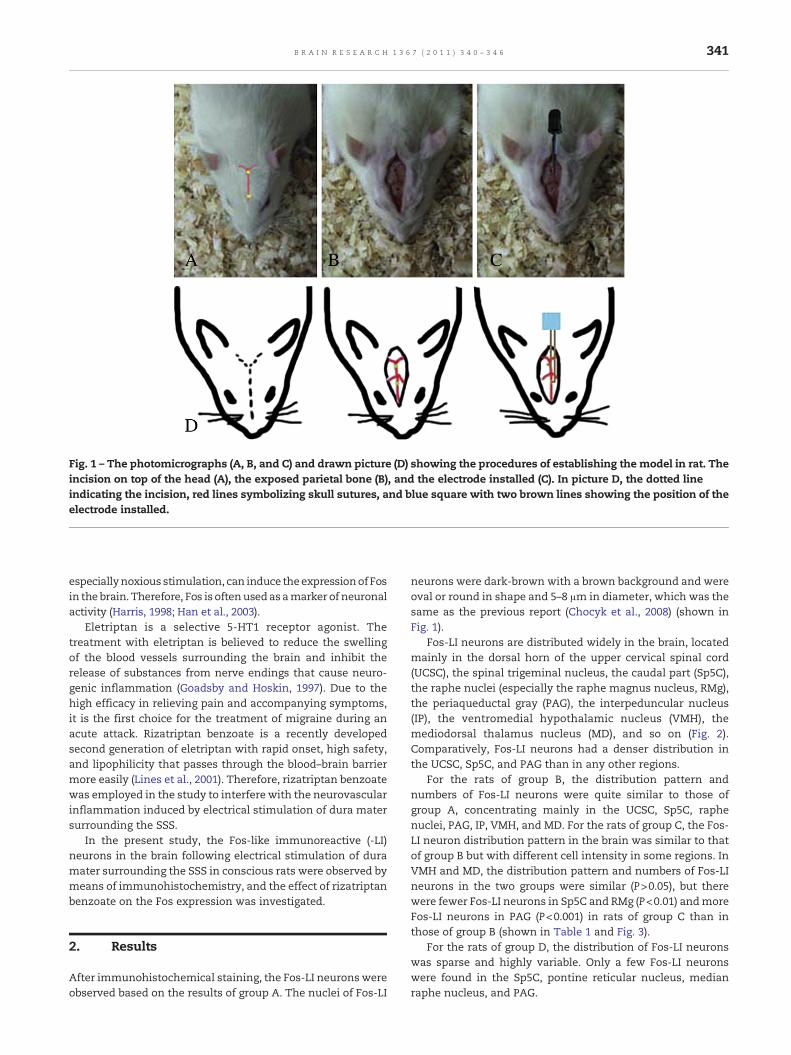
Fig. 1 – The photomicrographs (A, B, and C) and drawn picture (D) showing the procedures of establishing the model in rat. Theincision on top of the head (A), the exposed parietal bone (B), and the electrode installed (C). In picture D, the dotted lineindicating the incision, red lines symbolizing skull sutures, and blue square with two brown lines showing the position of theelectrode installed.
341B R A I N R E S E A R C H 1 3 6 7 ( 2 0 1 1 ) 3 4 0 – 3 4 6
especiallynoxious stimulation, can induce theexpressionof Fosin the brain. Therefore, Fos is oftenusedasamarker of neuronalactivity (Harris, 1998; Han et al., 2003).
Eletriptan is a selective 5-HT1 receptor agonist. Thetreatment with eletriptan is believed to reduce the swellingof the blood vessels surrounding the brain and inhibit therelease of substances from nerve endings that cause neuro-genic inflammation (Goadsby and Hoskin, 1997). Due to thehigh efficacy in relieving pain and accompanying symptoms,it is the first choice for the treatment of migraine during anacute attack. Rizatriptan benzoate is a recently developedsecond generation of eletriptan with rapid onset, high safety,and lipophilicity that passes through the blood–brain barriermore easily (Lines et al., 2001). Therefore, rizatriptan benzoatewas employed in the study to interfere with the neurovascularinflammation induced by electrical stimulation of dura matersurrounding the SSS.
In the present study, the Fos-like immunoreactive (-LI)neurons in the brain following electrical stimulation of duramater surrounding the SSS in conscious rats were observed bymeans of immunohistochemistry, and the effect of rizatriptanbenzoate on the Fos expression was investigated.
2. Results
After immunohistochemical staining, the Fos-LI neuronswereobserved based on the results of group A. The nuclei of Fos-LI
neurons were dark-brown with a brown background and wereoval or round in shape and 5–8 μm in diameter, which was thesame as the previous report (Chocyk et al., 2008) (shown inFig. 1).
Fos-LI neurons are distributed widely in the brain, locatedmainly in the dorsal horn of the upper cervical spinal cord(UCSC), the spinal trigeminal nucleus, the caudal part (Sp5C),the raphe nuclei (especially the raphe magnus nucleus, RMg),the periaqueductal gray (PAG), the interpeduncular nucleus(IP), the ventromedial hypothalamic nucleus (VMH), themediodorsal thalamus nucleus (MD), and so on (Fig. 2).Comparatively, Fos-LI neurons had a denser distribution inthe UCSC, Sp5C, and PAG than in any other regions.
For the rats of group B, the distribution pattern andnumbers of Fos-LI neurons were quite similar to those ofgroup A, concentrating mainly in the UCSC, Sp5C, raphenuclei, PAG, IP, VMH, and MD. For the rats of group C, the Fos-LI neuron distribution pattern in the brain was similar to thatof group B but with different cell intensity in some regions. InVMH and MD, the distribution pattern and numbers of Fos-LIneurons in the two groups were similar (P>0.05), but therewere fewer Fos-LI neurons in Sp5C and RMg (P<0.01) andmoreFos-LI neurons in PAG (P<0.001) in rats of group C than inthose of group B (shown in Table 1 and Fig. 3).
For the rats of group D, the distribution of Fos-LI neuronswas sparse and highly variable. Only a few Fos-LI neuronswere found in the Sp5C, pontine reticular nucleus, medianraphe nucleus, and PAG.
Fig. 2 – Immunohistochemical photomicrograph of sections showing the Fos-LI neurons in the spinal trigeminal nucleus,caudal part. Arrowhead indicates Fos-LI neurons. Sp5C indicates spinal trigeminal nucleus, caudal part; rs, rubrospinal tract;LRt, lateral reticular nucleus. Scale bar is 50.0 μm.
Fig. 3 – Schematic picture illustrating the distribution of Fos-LI neurons in the brain in the coronal sections of one rat (rat 2 ingroup A). Sections are arranged in a rostral to caudal order. Each circle indicates five Fos-LI neurons. DV3 indicates dorsal 3rdventricle; MD, mediodorsal thalamic nucleus; VMH, ventromedial hypothalamic nucleus; PAG, periaqueductal gray; IP,interpeduncular nucleus; Aq, aqueduct; 4V, 4 ventricle; CG, central gray; PnC, pontine reticular nucleus, caudal part; Py,pyramidal tract 7n; Sp5, spinal trigeminal nucleus; Pr5VL, principal sensory trigeminal nucleus, ventrolateral part; RMg, raphemagnus nucleus; Sp5I, spinal trigeminal nucleus, interpolar part; Sp5C, spinal trigeminal nucleus, caudal part; RPa, raphepallidus nucleus; DH, dorsal horn.
342 B R A I N R E S E A R C H 1 3 6 7 ( 2 0 1 1 ) 3 4 0 – 3 4 6
343B R A I N R E S E A R C H 1 3 6 7 ( 2 0 1 1 ) 3 4 0 – 3 4 6
In the second sections of all groups, no neurons in thebrainstem showing Fos-LI were observed.
3. Discussion
Since the report by Hunt et al. (1987), many studies haveshown that various types of noxious stimulation, includingthermal, mechanical, and chemical stimuli, induce expressionof Fos in the brain and spinal cord. Fos expression induced bymigraine-like stimulus have been conducted inmonkeys, cats,guinea pigs, and rats (Goadsby and Hoskin, 1997; Hoskin et al.,1996; Gupta et al., 2006; Akerman et al., 2007). However, mostof these studies were performed on animals in a state ofanesthetization. Moreover, they paid more attention to theSp5C and the drugs' effect on it (Storer and Goadsby, 1997).However, more and more evidence shows that migraine ismuch more than a simple headache. It involves hereditaryand developmental factors, as well as estrogen levels and areferring somatic sensory, autonomic nervous, and limbicsystem. In the present study, we systematically observed theFos expression in the brain of conscious rats followingelectrical stimulation of dura mater surrounding the SSS,which imitated a migraine attack in the conscious state. Wefound that Fos-LI neurons were distributed in the UCSC, Sp5C,RMg, PAG, IP, VMH, and MD.
Fig. 4 – Graphs showing the number of Fos-LI neurons in rat braP<0.001 in the PAG (C), P>0.05 in the MD (D) and VMH (E).
TheFosexpression in theUCSCandSp5C thatwas inducedbythe intervention is regarded as a standard for establishing aheadache model (Goadsby and Hoskin, 1997; Hoskin et al., 1999;Akerman et al., 2007). Our result here is quite similar to thatreported by others but with a more convergent distribution,mainly in the ventromedial or superficial layers of the Sp5C. Weassume that the fixed and limited stimulation causes thislocalization, and the comparatively dispersed distribution inthe model of chemical stimulus gives an indirect support(Mitsikostas et al., 1998, 1999). The RMg, PAG, VMH, and MDhave all been implicated in nociception, although in differentroles and mechanisms (Menétrey et al., 1982; Blomqvist andCraig, 1991; Li et al., 1993;Mukherjee et al., 2002; Pertovaara et al.,1993). The RMg and PAG are the main components of adescending pain inhibitory system (DPIS), and the VMH is oneof its origins. The MD in nociception and its modulation is lesscommonly mentioned but well documented. One noteworthyaspect here is the activation of the PAG. In the clinic, patientswho suffer infarction or infection that destroy the PAG wouldmanifest a migraine-like symptom (Haas et al., 1993). Nonmi-graine patients develop migraine-like headaches after theimplantation of a stimulating electrode in the PAG (Raskinet al., 1987). As our data prove the active state of neurons in thePAG, we prefer to interpret this contradiction in terms ofinhibition of “off neurons” and facilitation of “on neurons”(Vanegas andSchaible, 2004;Martins et al., 2010). Theother is the
in of groups B and C. P<0.01 in the Sp5C (A) and the RMg (B),
344 B R A I N R E S E A R C H 1 3 6 7 ( 2 0 1 1 ) 3 4 0 – 3 4 6
VMH. The VMH is most commonly associated with food intake.However, it has long been involved in pain modulation. It hasbeen reported to take part in acupuncture-, sucrose-fed-, as wellas many peptide- or protein-mediated analgesia (Yang et al.,2007; Mukherjee et al., 2002; Hori et al., 2000). However, Malick etal. (2001) suggest that activation of the VMH in the trigemino-vascular headache is, at least, partial responsible for pain-induced appetite loss. This, in theory, explains why migraineattacks are always associated with appetite loss. As we have notinvestigated the digestive system of rats, direct evidencesupporting this association is lacking. However, if imagingresearch, such as fMRI, was combined, this would constitute aprospective direction for further research.
Eletriptan has been widely used to abort migraine head-ache in the early phase of attacks. However, we know little ofits effect on the CNS, except the vasoconstriction and reducedtransmitter release. In the present study, with a pre-treatmentof intraperitoneal injection of rizatriptan benzoate, thenumber of Fos-LI neurons decreased in the Sp5C and RMg,increased in the PAG, and were unchanged in the VMH andMD. The changed Fos expression after the rizatriptan benzoatemay be considered to be an integration of descent nociceptiveinput and elevated antinociceptive modulation. Withoutfurther electrophysiological and ethological study, theseresults could only advance our understanding of where therizatriptan benzoate may act to reduce the pain rather thanhow it may work.
4. Conclusion
After the electrical stimulation of the SSS in conscious rats, Fosexpression in the brain as well as the effect of rizatriptanbenzoate on it were investigated. Since the purpose of the workis to provide a morphological rather than a functional evidencefor the involvement of nuclei in trigeminovascular headache,whether andhow theywork is still needed to be verified further.Recently, we just begin to identify the chemical property ofthese excited neurons in different areas. It will help us identifythe role that the nucleus may play in migraine and find outpossible targets for drug treatment.
5. Experimental procedures
5.1. Animals
Forty Sprague–Dawley male rats weighing 200–250 g each wereused. The experimental procedures were approved by theCommittee of Animal Use for Research and Education of theLaboratory Animal Center of the Chinese PLA General Hospital(Beijing, PRChina) andwere consistentwith theethical guidelinesrecommended by the International Association for the Study ofPain for experimental pain in conscious animals (Zimmermann,1983).
5.2. Groups
The rats were randomly divided into four experimental groups(n=10 for each group): stimulation group (A), physiological
saline group (B), rizatriptan benzoate group (C), and controlgroup (D).
5.3. Surgical procedures
The rats of groupA, B, and Cwere anesthetizedwith 10% chloralhydrate (4 ml/kg, i.p.) and then placed into a stereotactic frame.The scalp covering the dorsal surface of each skull was incisedand the connective tissue and muscle were removed, leavingthe parietal bone exposed. Two cranial windows (4mm beforeand 6 mm after the bregma on the midline suture and 1 mm indiameter) for electrical stimulation (Fig. 4) were carefully drilledinto the parietal bone, and the skull was opened to expose thedura mater adjacent to the superior sagittal sinus (Kurosawa etal., 1995). Care was taken to drill with a constant application ofice-cold saline. A pair of stimulation electrodes (0.8 mm indiameter; Donghengyu Ltd., PR China) were oriented in thedrilled holes and placed onto the dural surface. Except for theirends, the electrodes were insulated. It is important to avoidlacerating the dural mater with the electrodes. Then, theelectrodes were attached to the skull, using 502 glue and dentalcement, and the sectioned skin was sutured, leaving only thewire connection outside of the skull. All of the rats receivedprophylactic treatment with antibiotic injections (penicillin,0.04million IU/100 g) for two days following surgery. Then, theywere housed individually at a constant ambient room temper-ature and a 12-h light–dark cycle and given unrestricted accessto food andwater following the surgical procedure. Testing tookplace on the fourth day in the test room,whichwasmaintainedat about 25 °C.
5.4. Experimental procedures
Rats were placed into a cage of 40 cm in diameter and 17.5 cm inheight,which is convenient for bothobservationand recordingbyavideocamera.Theelectrodesoneachrat'sheadwereconnectedto a stimulator (SEN-7103; Japan) and separator (SS-102J; Japan) bya matching wire connection. The electrical stimulus parameterswere established on the basis of our preliminary experiment(monophasic square-wave pulses, 0.25ms in pulse duration and20 Hz in stimuli frequency). Before the electrical stimulus stage ofthe experimental procedures, all rats received a transientelectrical stimulation (5min) to ensure that the electrodes hadgood contact and to determine the current intensity applied later,which was the minimum that would elicit an eligible behavioralresponse to pain, usually 2.5–3.5 mA. After a 30-min periodhabituation in the same cage tominimize stress, rats in groups Band C received i.p. injections of normal saline solution andrizatriptan benzoate [0.083mg/kg (5mg/60 kg, adult human)],30 min before the electrical stimulation. Then, an electricalstimulation was applied to the rats in groups A, B, and C for 1 h,with rats in group D connected to the stimulator, but notstimulated.
5.5. Perfusion and tissue preparation
One hour after electrical stimulation, all rats were re-anesthe-tizedwith an overdose of 10% chloral hydrate (15 ml/kg, i.p.) andperfused transcardially through the ascending aorta with
345B R A I N R E S E A R C H 1 3 6 7 ( 2 0 1 1 ) 3 4 0 – 3 4 6
500 ml of 0.1 M phosphate buffer (PB, pH 7.4) containing 4%paraformaldehyde. Their brains were immediately removed,placed into the same fresh fixative for 4 h (4 °C), and thenplacedinto 0.1 M PB (pH 7.4) containing 30% sucrose overnight at 4 °C.The brainstems were cut into serial frontal sections of 30 μmthickness on a freezing microtome (Cryostat 1720; Leitz,Wetzlar, Germany). The sections were collected at intervals inthree dishes containing 0.01 M phosphate buffered saline (PBS,pH 7.4). All sections were washed with 0.01 M PBS.
5.6. Immunohistochemistry
The first set of sections in the first dish were incubated atroom temperaturewith rat antisera against Fos (1:500 dilution;Santa Cruz Biotechnology, Santa Cruz, CA, USA) in 0.01 M PBScontaining 5% normal donkey serum, 0.3% Triton X-100, 0.05%NaN 3, and 0.25% carrageenan (PBS–NDS, pH 7.4) for 24 h. Thiswas followed by goat anti-rat IgG (1:100 dilution; Dako, FortCollins, CO, USA) in PBS–NDS for 3 h and then by peroxidaseantiperoxidase (PAP) made from rat (1:200 dilution; Dako) inPBS containing 0.3% Triton X-100 (PBS-X, pH 7.4) for 1 h.Between each step, the sections were washed completely with0.01 M PBS three times (10 min per time). Finally, the sectionswere reacted with 0.05 M Tris–HCl buffer (pH 7.6) containing0.04% diaminobenzidine tetrahydrochloride (DAB) (Doujin,Kumamoto, Japan), 0.003% H2O2, and 0.04% Ni(NH4)2(SO4)2 for30–40 min to intensify the DAB-based reaction of Fos-LIneurons.
For the second set of sections, normal rat serum was usedto replace rat anti-Fos antisera. The following stainingprocedures were the same as those used for the first set ofserial sections. All of the sections were then mounted ontogelatin-coated glass slides.
The third set of sections weremounted onto gelatin-coatedglass slides and processed for Nissl staining.
5.7. Statistical analysis
Five rats from groups B and C were randomly selected forstatistical analysis. For each rat, the first sections from the firstdish were used for the study of quantification or mapping of theimmunoreactive neuronal cell bodies, which usually comprisedabout30sectionsof 30 μmthickness, underanopticalmicroscope(VANOX; Olympus, Tokyo, Japan).
Cells were counted in squares at 40× magnification (onesquare randomlyper brain regionper section). The brain regionswere identified according to the atlas of Paxinos and Watson(2004). The data for different brain regions from five rats shownin Tables 1 and 2 were the sum of the cell numbers from allsquares in the same brain region, which represent the cellintensity in the regions. For normal distribution, the numericaldata were expressed as xP±s. For abnormal distribution orunequal variance, thenumerical datawereexpressedasmedian(.25 and .75 quartiles).
The Student Newman–Keuls test was used as a comparisonmethod to compare differences between the groups. In cases ofabnormal distribution or unequal variance, the Kruskall–Wallistest was used (nonparametric ANOVA). A statistical differencewas considered to be significant if P<0.05. The statistical analysiswas performed using SPSS13 software.
Acknowledgments
We are grateful to Mr. X. H. Wang for animal ethological help.This work was supported in part by the National NaturalScience Foundation of China (30570264).
Benjamin, L., Levy, M.J., Lasalandra, M.P., Knight, Y.E., Akerman, S.,Classey, J.D., Goadsby, P.J., 2004. Hypothalamic activation afterstimulation of the superior sagittal sinus in the cat: a Fos study.J. Neurobiol. Dis. 16, 500–505.
Blomqvist, A., Craig, A.D., 1991. Organization of spinal andtrigeminal input to the PAG. In: Depaulis, A., Bandler, R. (Eds.),The Midbrain Periaqueductal Gray Matter: Functional,Anatomical and Neurochemical Organization. London PlenumPress, New York, pp. 345–363.
Chocyk, A., Czyrak, A., Wedzony, K., 2008. Dopamine D1-likereceptors agonist SKF 38393 increases c-fos expression in theparaventricular nucleus of the hypothalamus—impact of acuteand chronic cocaine. J. Physiol. Pharmcol. 59 (3), 425–440.
Clayton, J.S., Gaskin, P.J., Beattie, D.T., 1997. Attenuation of Fos-likeimmunoreactivity in the trigeminal nucleus caudalis followingtrigeminovascular activation in the anaesthetised guinea-pig.J. Brain Res. 775, 74–80.
De Vries, P., Villalon, C.M., Saxena, P.R., 1999. Pharmacologicalaspects of experimental headache models in relation to acuteantimigraine therapy. Eur. J. Pharmacol. 375, 61–74.
Dragunow, M., Faull, R., 1989. The use of c-fos as a metabolicmarker in neuronal pathway tracing. J. Neurosci. Meth. 29,261–266.
Paxinos, G., Watson, C., 2004. The Rat Brain in StereotaxicCoordinates, 5th ed. Academic Press, New York.
Goadsby, P.J., Hoskin, K.L., 1997. The distribution oftrigeminovascular afferents in the nonhuman primatebrain Macaca nemestrina: a c-fos immunocytochemical study.J. Anat. 190, 367–375.
Goadsby, P.J., Knight, Y.E., 1997. Inhibition of trigeminal neuronesafter intravenous administration of naratriptan through anaction at 5-hydroxy-tryptamine (5-HT(1B/1D)) receptors. Br. J.Pharmacol. 122, 918–922.
Gupta, S., Akerman, S., van den Maagdenberg, A.M., Saxena, P.R.,Goadsby, P.J., van den Brink, A.M., 2006. Intravital microscopyon a closed cranial window in mice: a model to studytrigeminovascular mechanisms involved in migraine.Cephalalgia 26, 1294–1303.
Haas, D.C., Kent, P.F., Friedman, D.I., 1993. Headache caused by asingle lesion of multiple sclerosis in the periaqueductal grayarea. Headache 33, 452–455.
Han, F., Zhang, Y.F., Li, Y.Q., 2003. Fos expression in tyrosinehydroxylase-containing neurons in rat brainstem after visceralnoxious stimulation: an immunohistochemical study. World J.Gastroenterol. 9, 1045–1050.
Harris, J.A., 1998. Using c-fos as a neural marker of pain. Brain Res.Bull. 45, 1–8.
Hoskin, K.L., Bulmer, D.C., Goadsby, P.J., 1999. Fos expression inthe trigeminocervical complex of the cat after stimulation ofthe superior sagittal sinus is reduced by L-NAME. Neurosci.Lett. 266, 173–176.
Hoskin, K.L., Kaube, H., Goadsby, P.J., 1996. Sumatriptan can inhibittrigeminal afferents by an exclusively neural mechanism.Brain 119, 1419–1428.
346 B R A I N R E S E A R C H 1 3 6 7 ( 2 0 1 1 ) 3 4 0 – 3 4 6
Hori, T., Oka, T., Hosoi, M., Abe, M., Oka, K., 2000. Hypothalamicmechanisms of pain modulatory actions of cytokines andprostaglandin E2. Ann. NY Acad. Sci. 917, 106–120.
Hunt, S.P., Pini, A., Evan, G., 1987. Induction of c-fos-like protein inspinal cord neurons following sensory stimulation. Nature 328,632–634.
Mukherjee, K., Mathur, R., Nayar, U., 2002. Hyperalgesic responsein rats fed sucrose from weaning to adulthood: role of VMH.Pharmacol. Biochem. Behav. 73, 601–610.
Kurosawa, M., Messlinger, K., Pawlak, M., Schmidt, R.F., 1995.Increase of meningeal blood flow after electrical stimulation ofrat dura mater encephali: mediation by calcitonin gene-relatedpeptide. Br. J. Pharmacol. 114, 1397–1402.
Li, Y.Q., Takada, M., Shinonaga, Y., Mizuno, N., 1993. Directprojections from the midbrain periaqueductal gray and dorsalraphe nucleus to the trigeminal sensory complex in the rat.Neuroscience 54, 431–443.
Martins, M.A., De Castro, Bastos L., Melo, N.E., Tonussi, C.R., 2010.Dependency of nociception facilitation or inhibition afterperiaqueductal gray matter stimulation on the context. Behav.Brain Res. 214, 260–267.
Malick, A., Jakubowski, M., Elmquist, J.K., Saper, C.B., Burstein, R.,2001. A neurohistochemical blueprint for pain-induced loss ofappetite. Proc. Natl Acad. Sci. USA 98, 9930–9935.
Menétrey, D., Chaouch, A., Binder, D., Besson, J.M., 1982. The originof the spinomesencephalic tract of the rat: an anatomical studyusing the retrograde transport of horseradish peroxidase.J. Comp. Neurol. 206, 193–207.
Mitsikostas, D.D., Sanchez del Rio, M., Waeber, C., Moskowitz,M.A., Cutrer, F.M., 1998. The NMDA receptor antagonistMK-801 reduces capsaicin-induced c-fos expressionwithin rat trigeminal nucleus caudalis. Pain 76,239–248.
Mitsikostas, D.D., Sanchez del Rio, M., Waeber, C., Huang, Z.,Cutrer, F.M., Moskowitz, M.A., 1999. Non-NMDA glutamatereceptors modulate capsaicin induced c-fos expressionwithin trigeminal nucleus caudalis. Br. J. Pharmacol. 127,623–630.
Pertovaara, A., Bravo, R., Herdegen, T., 1993. Induction andsuppression of immediate-early genes in the rat brain by aselective alpha-2-adrenoceptor agonist and antagonistfollowing noxious peripheral stimulation. Neuroscience 54,117–126.
Raskin, N.H., Hosobuchi, Y., Lamb, S., 1987. Headache may arisefrom perturbation of brain. J. Headache 27 (8), 416–420.
Rodella, L., Rezzani, R., Gioia, M., Tredici, G., Bianchi, R., 1998.Expression of fos immunoreactivity in the rat supraspinalregions following noxious visceral stimulation. Brain Res. Bull.47, 357–366.