Selective Cleavage of the Heregulin Receptor ErbB-4 by ProteinKinase C Activation*
(Received for publication, February 14, 1996, and in revised form, May 16, 1996)
Manuela Vecchi‡§, Josep Baulida‡, and Graham Carpenter‡¶i
From the Departments of ‡Biochemistry and ¶Medicine, Vanderbilt University School of Medicine,Nashville, Tennessee 37232-0146
The 180-kDa transmembrane tyrosine kinase ErbB-4is a receptor for the growth factor heregulin. 125I-Heregulin binding to NIH 3T3 cells overexpressing theErbB-4 receptor is rapidly decreased by 12-O-tetra-decanoylphorbol-13-acetate (TPA) pretreatment. Immu-nologic analysis demonstrates that TPA treatment ofcells induces the proteolytic cleavage of ErbB-4, produc-ing an 80-kDa cytoplasmic domain fragment, which con-tains a low level of phosphotyrosine, and a 120-kDaectodomain fragment, which is released into the extra-cellular medium. Cleavage of ErbB-4 was also enhancedby other protein kinase C activators, i.e. platelet-derivedgrowth factor, ionomycin, and synthetic diacylglycerol,while protein kinase C inhibition or down-regulationsuppressed the TPA stimulation of ErbB-4 degradation.TPA did not induce the degradation of related receptors(ErbB-1, ErbB-2, and ErbB-3) in the EGF receptorfamily.The phorbol ester-induced cleavage of ErbB-4 occurs
within or close to the ectodomain, as the 80-kDa cyto-plasmic domain fragment is recognized by antibody tothe ErbB-4 carboxyl terminus and is membrane-associ-ated. Coprecipitation experiments show that, while the80-kDa ErbB-4 fragment is associated with the SH2-con-taining molecules PLC-g1 and Shc, TPA did not inducethe phosphorylation of these substrates in intact cells.In addition, kinase assays in vitro indicate that the 80-kDa fragment is not an active tyrosine kinase. Theseresults show that protein kinase C negatively regulatesheregulin signaling through the ErbB-4 receptor by theactivation of a selective proteolytic mechanism.
Four transmembrane tyrosine kinases constitute the humanepidermal growth factor (EGF)1 receptor family: the EGF re-ceptor or ErbB-1, ErbB-2, ErbB-3, and ErbB-4 (1). The extra-cellular domains of all ErbB molecules have two cysteine-richdomains; the cytoplasmic tyrosine kinase domains are highlyconserved in primary sequence, while the cytoplasmic carboxyl
termini are dissimilar in sequence but not size. Understandingthe metabolic regulation of EGF receptor family members andtheir signal transduction pathways is of particular interest,as these receptors are implicated in a variety of aggressivecarcinomas (2–6).The EGF family of growth factors (7) specifically bind to the
EGF receptor, while the heregulin family of growth factors (8,9) associate with the ErbB-3 and ErbB-4 receptors. Heregulinbinds to ErbB-4 and ErbB-3, respectively, with high and mod-erate affinity (10), whereas a heterodimer of ErbB-2 andErbB-3 is reported to constitute a second high affinity receptor(11, 12). The association of heregulin with ErbB-3 or ErbB-4 isable to induce phosporylation of ErbB-2 by a mechanism thatinvolves transphosphorylation and perhaps receptor het-erodimerization (13, 14). The EGF receptor also heterodimer-izes with ErbB-2 (15, 16) and perhaps ErbB-3 (10, 17, 18).These observations may underlie the clinical importanceof coexpression of ErbB family members in many breastcarcinomas (19, 20).Ligand-induced desensitization and down-regulation mech-
anisms are important aspects of the regulation of transmem-brane receptors (21), and within the ErbB receptor familysignificant similarities and differences have emerged in atten-uation mechanisms. Ligand binding to the EGF receptor rap-idly induces receptor-mediated endocytosis through clathrin-coated pits (22). The internalized complexes are subsequentlydegraded in lysosomes and, consequently, cell surface EGFreceptors are down-regulated. In contrast to the EGF receptor,all of the other ErbB family members, including ErbB-4, arenot rapidly internalized in the presence of ligand and donot exhibit ligand-enhanced metabolic turnover and down-regulation (23, 24).Protein kinase C activity significantly influences desensiti-
zation of receptors in the ErbB family. EGF receptor activity isattenuated by protein kinase C through phosphorylation ofcytoplasmic domain serine/threonine residues which decreasesligand binding affinity and tyrosine kinase activity (25–27).Evidence for a similar phosphorylation-dependent attenuationof ErbB-2 tyrosine kinase activity by protein kinase C has beenpresented (28). In addition, heterologous attenuation of theEGF receptor by the platelet-derived growth factor (PDGF)receptor is thought to be mediated through the activation ofprotein kinase C (29–31). There are no reports of protein ki-nase C regulation of the ErbB-3 or ErbB-4 receptors. The datapresented below show that protein kinase C activators down-regulate the ErbB-4 receptor by activating a selective proteo-lytic mechanism.
EXPERIMENTAL PROCEDURES
Materials—Heregulin b1 and125I-heregulin b1 were generously sup-
plied by Mark Sliwkowski (Genentech Inc., San Francisco, CA). Mousemonoclonal antibodies to the carboxyl-terminal region of the humanEGF receptor were from Zymed, while rabbit polyclonal antibodies tothe carboxyl-terminal region of ErbB-2 and ErbB-3 and monoclonal
* This work was supported by National Institutes of Health GrantCA24071 and a Fellowship (to J. B.) from the Spanish Ministerio deEducacion y Ciencia. The costs of publication of this article were de-frayed in part by the payment of page charges. This article musttherefore be hereby marked “advertisement” in accordance with 18U.S.C. Section 1734 solely to indicate this fact.§ Supported by the Dipartimento di Scienze Biochimiche, Universita’
di Firenze.i To whom correspondence should be addressed: Dept. of Biochemis-
try, Rm. 647, Light Hall, Vanderbilt University School of Medicine,Nashville, TN 37232-0146. Tel.: 615-322--6678; Fax: 615-322-2931.
antibody to the extracellular domain of ErbB-4 were purchased fromNeo Markers (Fremont, CA). Antiserum to the carboxyl-terminal se-quence 1108–1264 of ErbB-4 (24) was a generous gift of MatthiasKraus, Istituto Europeo di Oncologia, Milan (19). Polyclonal IgG to thecarboxyl terminus (residues 1291–1308) of ErbB-4 were also purchasedfrom Santa Cruz Biotechnology, Inc. Polyclonal antibodies to Shc werepurchased from Transduction Laboratories. Antisera to phospholipaseC-g1 was described previously (32). Protein A-Sepharose and enhancedchemiluminescence (ECL) reagents were from Sigma and Immobilon-Pmembranes were from MCI. 125I-Labeled protein A was a product ofICN, and horseradish peroxidase conjugated protein A was fromZymed. 125I-Labeled goat anti-mouse IgG was purchased by ICN.[g-32P]ATP (3000 Ci/mmol) and Trans35S-label (1138 Ci/mmol) werefrom DuPont NEN. TPA, ionomycin, sn-1,2-dioctanoylglycerol, andPDGF were purchased from Sigma, while bisindolymaleimide (GF109203X) was from Calbiochem. All other chemicals were reagent gradefrom Sigma.Cell Culture—The cell line T47-14 produced by transfecting NIH 3T3
cells with the human ErbB-4 receptor, has been described elsewhere(24). The cell line Cl-17 overexpressing the human EGF receptor hasbeen previously characterized (33). T44-13 and T44-17 cell lines over-expressing, respectively, ErbB-2, or ErbB-2 and ErbB-3 were kind giftsof Dr. Paolo Di Fiore, Istituto Europeo di Oncologia, Milan. All cellswere routinely grown at 37 °C in 5% CO2 in Dulbecco’s modified Eagle’smedium (DMEM) containing 20 mM Hepes, pH 7.4, 10% calf serum, and50 mM gentamicin. Experimental cultures were generally grown in60-mm culture dishes.Immunoprecipitation and Western Blotting—T47-14 cells (70% con-
fluent) in 60-mm culture dishes were starved overnight in 0.5% calfserum. Subsequent incubations were performed in 1.0 ml of basalmedium (DMEM, 0.1% BSA, and 20 mM Hepes, pH 7.4). After theindicated treatments, cells were washed three times with ice-cold Ca21-and Mg21-free phosphate-buffered saline (CMF-PBS) and lysed with300 ml of ice-cold TGH buffer (1% Triton X-100, 10% glycerol, 20 mM
Hepes, pH 7.2, 100 mM NaCl, 1 mM phenylmethylsulfonyl fluoride, 10mg/ml leupeptin, 10 mg/ml aprotinin, and 1 mMNa3VO4). After scraping,the lysates were incubated for 30 min at 4 °C with rocking. Insolublematerial was then removed by centrifugation (14,000 3 g, 10 min) at4 °C, and the supernatant was collected. Protein concentration wasassayed with the method of Bradford (34) using bovine serum albuminas the standard. To precipitate ErbB-4, approximately 1 mg of ErbB-4antibody was incubated (2 h at 4 °C) with 300 mg of cellular proteinfollowed by a 1-h incubation with protein A-Sepharose CL-4B. Immunecomplexes were washed three times with TGH lysis buffer, resuspendedin 1 3 Laemmli buffer (35), and boiled for 5 min. Subsequently, sampleswere electrophoresed in a 7.5% SDS-polyacrylamide gel and transferredto nitrocellulose membranes for Western blotting. Membranes wereblocked by a 1-h incubation at room temperature with TBST buffer(0.05% Tween 20, 150 mM NaCl, 50 mM Tris, pH 7.4) containing 3%BSA. Subsequently, ErbB-4 antiserum (dilution, 1:3000) or phosphoty-rosine antibody (dilution, 1:1000) was added for 2 h at room tempera-ture with shaking. Membranes were then washed three times withTBST buffer, incubated either with 125I-protein A for 1 h or horseradishperoxidase-conjugated protein A, and after washing with TBST buffervisualized by autoradiography (Kodak X-Omat AR film) or ECL,respectively.Cell Fractionation—Cells were washed three times with ice-cold
CMF-PBS, scraped into 800 ml of hypotonic buffer A (20 mM Hepes, pH7.4, 10 mg/ml leupeptin, 10 mg/ml aprotinin, 1 mM phenylmethylsulfonylfluoride, and 1 mM Na3VO4), and disrupted with a Dounce homogenizer.NaCl was then added to a final concentration of 100 mM. Nuclei andunbroken cells were removed by two low speed centrifugations (800 3 g,5 min). The supernatants were ultracentrifuged (100,000 3 g, 50 min),and Triton X-100 was added to 1% to the high speed supernatants. Thehigh speed pellets were gently rinsed twice with ice-cold CMF-PBS,resuspended in hypotonic buffer A supplemented with 1% Triton X-100and 100 mM NaCl, and solubilized for 30 min at 4 °C with rocking.Subsequently, insolubilized material was removed from the Triton-solubilized pellets by centrifugation (100,000 3 g, 50 min). Equal vol-umes of cytosol and membrane fractions, representing equivalent num-bers of cells, were precipitated with antibody to ErbB-4 and subjected toWestern blot analysis.In Vitro Kinase Assay—Cell monolayers were washed twice with
ice-cold CMF-PBS and lysed in 300 ml of TGH buffer without Na3VO4.Duplicate samples (300 mg of protein) were immunoprecipitated (2 h,4 °C) with polyclonal antibody to ErbB-4 before protein A-Sepharosewas added to each tube and incubated (1 h, 4 °C). The immunoprecipi-tates were then washed twice with the TGH buffer without Na3VO4 and
twice with kinase buffer (20 mM Hepes, pH 7.4, 3 mM MnCl2, 15 mM
MgCl2, 200 mM NaCl, 10 mg/ml aprotinin, and 100 mM Na3VO4). Im-mune complexes were resuspended in 50 ml of kinase buffer containingATP (5 mM), and to half the samples [g-32P]ATP (20 mCi per sample) wasadded. All tubes were then incubated at 37 °C for 5 min. The reactionwas stopped by adding 500 ml of a buffer containing 5 mM sodiumphosphate, pH 7.0, 1% Triton X-100, 1% sodium deoxycholate, 0.1%SDS, 150 mM NaCl, 1 mM Na3VO4, 10 mg/ml aprotinin, 1 mM ATP, and5 mM EDTA. The immune complexes were washed twice with the samebuffer, and 1 3 Laemmli buffer was added. Protein in each sample werethen separated on a 7.5% polyacrylamide gel and the [g-32P]ATP-la-beled samples were analyzed by autoradiography. Nonradioactive sam-ples were analyzed for ErbB-4 protein by Western blotting with ErbB-4antiserum, and the results were quantitated using a PhosphorImager.
RESULTS
TPA Down-modulation of 125I-Heregulin Binding to ErbB-4—To determine whether protein kinase C activation couldmodulate heregulin binding to ErbB-4, 125I-heregulin bindingassays were performed on T47-14 cells, which express approx-imately 1 3 106 ErbB-4 heregulin receptors per cell (24) fol-lowing TPA pretreatment. Cells were incubated with or with-out TPA at 37 °C for 30 min or 1 h and then incubated at 4 °Cwith a near saturating concentration of 125I-heregulin to meas-ure cell surface receptors. The results, shown in Fig. 1, dem-onstrate that TPA preincubations of 30 and 60 min decreased125I-heregulin binding by 82% and 91%, respectively. Thesedata show that the ligand binding capacity of ErbB-4 receptorscan be dramatically down-regulated by the protein kinase Cactivator TPA.Influence of TPA on ErbB-4 Protein—There are several pos-
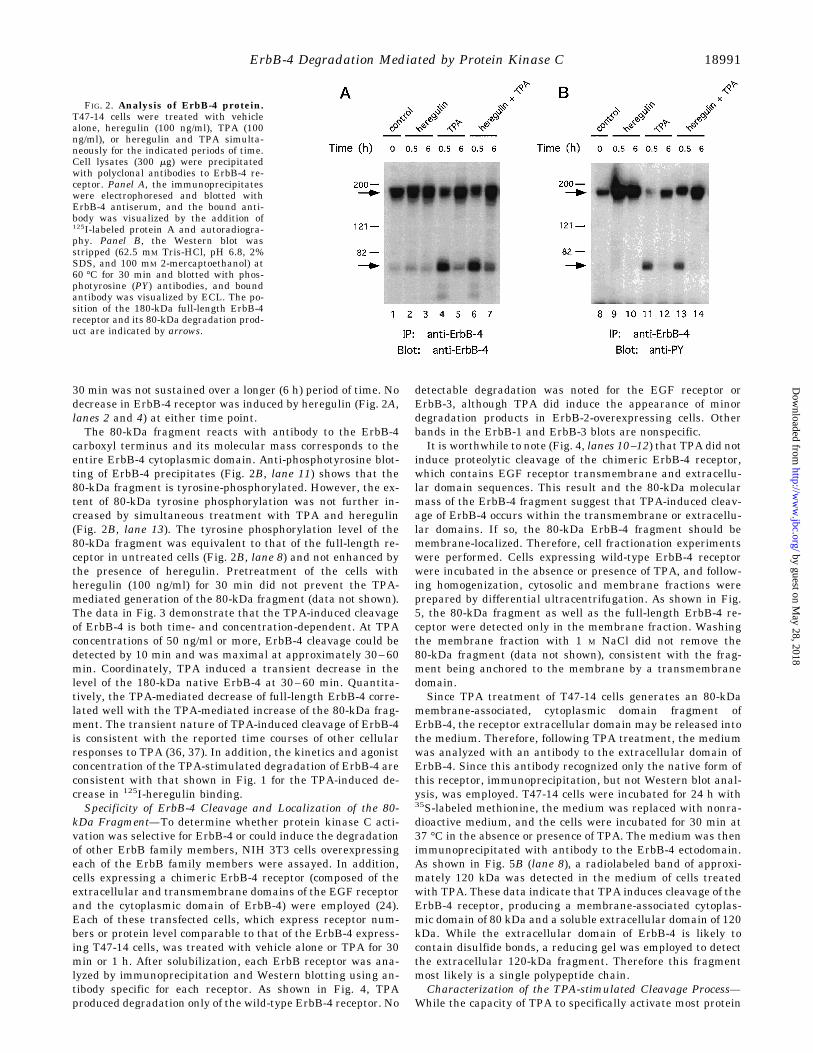
sible mechanisms by which TPAmight modulate 125I-heregulinbinding to the ErbB-4 receptor. To determine whether TPAalters the cellular level or structure of ErbB-4 protein, cellswere incubated with TPA in the presence or absence of heregu-lin for 30 min or 6 h, and detergent lysates were assayed forErbB-4 protein by immunoprecipitation and Western blotting.As shown in Fig. 2A (lane 4), TPA rapidly stimulated proteo-lytic cleavage of the ErbB-4 receptor as indicated by the loss ofnative receptor (180 kDa) and the appearance of an 80-kDafragment. Activation of this proteolytic event while evident at
FIG. 1. Influence of TPA on heregulin binding to intact cells.T47-14 cells in six-well dishes were washed twice with DMEM, and 1.0ml of binding medium (DMEM plus 0.1% BSA) was added. TPA (100ng/ml) was added to the indicated cells for 30 min or 1 h, and thecultures were incubated at 37 °C. Cell monolayers were then rapidlywashed with ice-cold DMEM and incubated for 90 min at 4 °C inbinding medium containing 125I-heregulin (100 ng/ml). After incuba-tion, the cells were washed twice with ice-cold DMEM and lysed in 1 N
NaOH. Quantitation of the amount of 125I-heregulin specifically boundwas performed with a g spectrometer (Beckman). Nonspecific 125I-heregulin binding was determined in parallel cultures containing anexcess of unlabeled heregulin.
ErbB-4 Degradation Mediated by Protein Kinase C18990
30 min was not sustained over a longer (6 h) period of time. Nodecrease in ErbB-4 receptor was induced by heregulin (Fig. 2A,lanes 2 and 4) at either time point.The 80-kDa fragment reacts with antibody to the ErbB-4
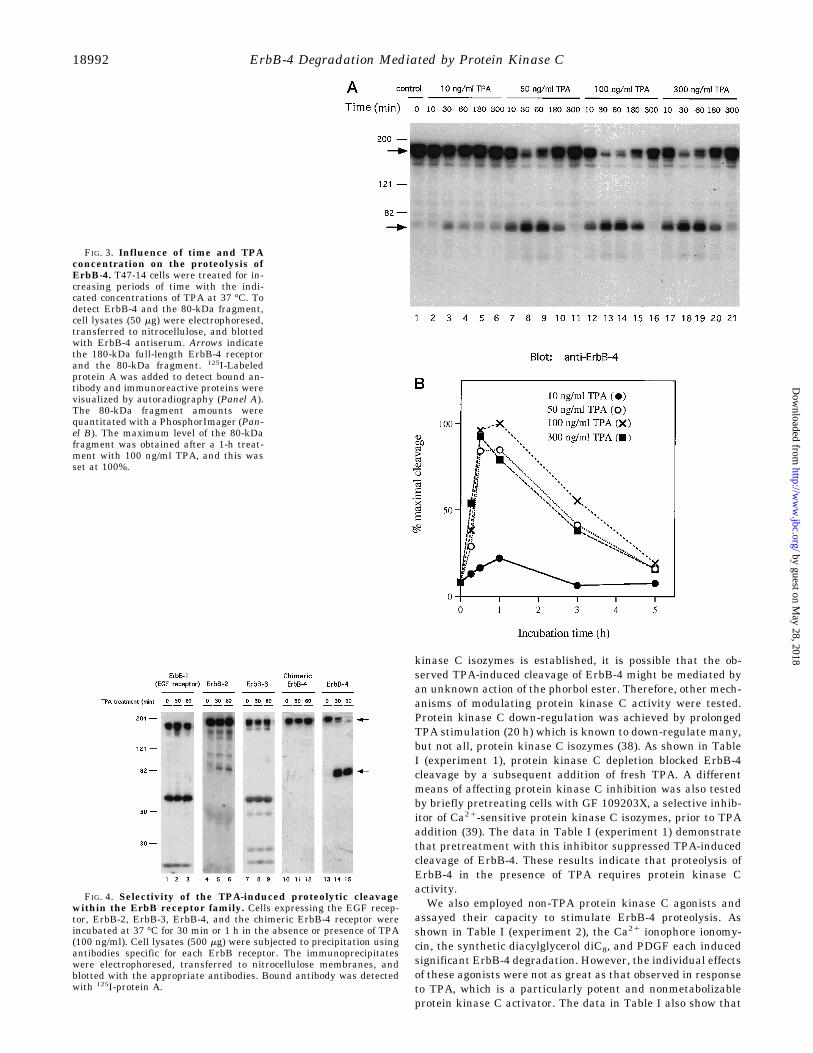
carboxyl terminus and its molecular mass corresponds to theentire ErbB-4 cytoplasmic domain. Anti-phosphotyrosine blot-ting of ErbB-4 precipitates (Fig. 2B, lane 11) shows that the80-kDa fragment is tyrosine-phosphorylated. However, the ex-tent of 80-kDa tyrosine phosphorylation was not further in-creased by simultaneous treatment with TPA and heregulin(Fig. 2B, lane 13). The tyrosine phosphorylation level of the80-kDa fragment was equivalent to that of the full-length re-ceptor in untreated cells (Fig. 2B, lane 8) and not enhanced bythe presence of heregulin. Pretreatment of the cells withheregulin (100 ng/ml) for 30 min did not prevent the TPA-mediated generation of the 80-kDa fragment (data not shown).The data in Fig. 3 demonstrate that the TPA-induced cleavageof ErbB-4 is both time- and concentration-dependent. At TPAconcentrations of 50 ng/ml or more, ErbB-4 cleavage could bedetected by 10 min and was maximal at approximately 30–60min. Coordinately, TPA induced a transient decrease in thelevel of the 180-kDa native ErbB-4 at 30–60 min. Quantita-tively, the TPA-mediated decrease of full-length ErbB-4 corre-lated well with the TPA-mediated increase of the 80-kDa frag-ment. The transient nature of TPA-induced cleavage of ErbB-4is consistent with the reported time courses of other cellularresponses to TPA (36, 37). In addition, the kinetics and agonistconcentration of the TPA-stimulated degradation of ErbB-4 areconsistent with that shown in Fig. 1 for the TPA-induced de-crease in 125I-heregulin binding.Specificity of ErbB-4 Cleavage and Localization of the 80-
kDa Fragment—To determine whether protein kinase C acti-vation was selective for ErbB-4 or could induce the degradationof other ErbB family members, NIH 3T3 cells overexpressingeach of the ErbB family members were assayed. In addition,cells expressing a chimeric ErbB-4 receptor (composed of theextracellular and transmembrane domains of the EGF receptorand the cytoplasmic domain of ErbB-4) were employed (24).Each of these transfected cells, which express receptor num-bers or protein level comparable to that of the ErbB-4 express-ing T47-14 cells, was treated with vehicle alone or TPA for 30min or 1 h. After solubilization, each ErbB receptor was ana-lyzed by immunoprecipitation and Western blotting using an-tibody specific for each receptor. As shown in Fig. 4, TPAproduced degradation only of the wild-type ErbB-4 receptor. No
detectable degradation was noted for the EGF receptor orErbB-3, although TPA did induce the appearance of minordegradation products in ErbB-2-overexpressing cells. Otherbands in the ErbB-1 and ErbB-3 blots are nonspecific.It is worthwhile to note (Fig. 4, lanes 10–12) that TPA did not
induce proteolytic cleavage of the chimeric ErbB-4 receptor,which contains EGF receptor transmembrane and extracellu-lar domain sequences. This result and the 80-kDa molecularmass of the ErbB-4 fragment suggest that TPA-induced cleav-age of ErbB-4 occurs within the transmembrane or extracellu-lar domains. If so, the 80-kDa ErbB-4 fragment should bemembrane-localized. Therefore, cell fractionation experimentswere performed. Cells expressing wild-type ErbB-4 receptorwere incubated in the absence or presence of TPA, and follow-ing homogenization, cytosolic and membrane fractions wereprepared by differential ultracentrifugation. As shown in Fig.5, the 80-kDa fragment as well as the full-length ErbB-4 re-ceptor were detected only in the membrane fraction. Washingthe membrane fraction with 1 M NaCl did not remove the80-kDa fragment (data not shown), consistent with the frag-ment being anchored to the membrane by a transmembranedomain.Since TPA treatment of T47-14 cells generates an 80-kDa
membrane-associated, cytoplasmic domain fragment ofErbB-4, the receptor extracellular domain may be released intothe medium. Therefore, following TPA treatment, the mediumwas analyzed with an antibody to the extracellular domain ofErbB-4. Since this antibody recognized only the native form ofthis receptor, immunoprecipitation, but not Western blot anal-ysis, was employed. T47-14 cells were incubated for 24 h with35S-labeled methionine, the medium was replaced with nonra-dioactive medium, and the cells were incubated for 30 min at37 °C in the absence or presence of TPA. The medium was thenimmunoprecipitated with antibody to the ErbB-4 ectodomain.As shown in Fig. 5B (lane 8), a radiolabeled band of approxi-mately 120 kDa was detected in the medium of cells treatedwith TPA. These data indicate that TPA induces cleavage of theErbB-4 receptor, producing a membrane-associated cytoplas-mic domain of 80 kDa and a soluble extracellular domain of 120kDa. While the extracellular domain of ErbB-4 is likely tocontain disulfide bonds, a reducing gel was employed to detectthe extracellular 120-kDa fragment. Therefore this fragmentmost likely is a single polypeptide chain.Characterization of the TPA-stimulated Cleavage Process—
While the capacity of TPA to specifically activate most protein
FIG. 2. Analysis of ErbB-4 protein.T47-14 cells were treated with vehiclealone, heregulin (100 ng/ml), TPA (100ng/ml), or heregulin and TPA simulta-neously for the indicated periods of time.Cell lysates (300 mg) were precipitatedwith polyclonal antibodies to ErbB-4 re-ceptor. Panel A, the immunoprecipitateswere electrophoresed and blotted withErbB-4 antiserum, and the bound anti-body was visualized by the addition of125I-labeled protein A and autoradiogra-phy. Panel B, the Western blot wasstripped (62.5 mM Tris-HCl, pH 6.8, 2%SDS, and 100 mM 2-mercaptoethanol) at60 °C for 30 min and blotted with phos-photyrosine (PY) antibodies, and boundantibody was visualized by ECL. The po-sition of the 180-kDa full-length ErbB-4receptor and its 80-kDa degradation prod-uct are indicated by arrows.
ErbB-4 Degradation Mediated by Protein Kinase C 18991
kinase C isozymes is established, it is possible that the ob-served TPA-induced cleavage of ErbB-4 might be mediated byan unknown action of the phorbol ester. Therefore, other mech-anisms of modulating protein kinase C activity were tested.Protein kinase C down-regulation was achieved by prolongedTPA stimulation (20 h) which is known to down-regulate many,but not all, protein kinase C isozymes (38). As shown in TableI (experiment 1), protein kinase C depletion blocked ErbB-4cleavage by a subsequent addition of fresh TPA. A differentmeans of affecting protein kinase C inhibition was also testedby briefly pretreating cells with GF 109203X, a selective inhib-itor of Ca21-sensitive protein kinase C isozymes, prior to TPAaddition (39). The data in Table I (experiment 1) demonstratethat pretreatment with this inhibitor suppressed TPA-inducedcleavage of ErbB-4. These results indicate that proteolysis ofErbB-4 in the presence of TPA requires protein kinase Cactivity.We also employed non-TPA protein kinase C agonists and
assayed their capacity to stimulate ErbB-4 proteolysis. Asshown in Table I (experiment 2), the Ca21 ionophore ionomy-cin, the synthetic diacylglycerol diC8, and PDGF each inducedsignificant ErbB-4 degradation. However, the individual effectsof these agonists were not as great as that observed in responseto TPA, which is a particularly potent and nonmetabolizableprotein kinase C activator. The data in Table I also show that
FIG. 4. Selectivity of the TPA-induced proteolytic cleavagewithin the ErbB receptor family. Cells expressing the EGF recep-tor, ErbB-2, ErbB-3, ErbB-4, and the chimeric ErbB-4 receptor wereincubated at 37 °C for 30 min or 1 h in the absence or presence of TPA(100 ng/ml). Cell lysates (500 mg) were subjected to precipitation usingantibodies specific for each ErbB receptor. The immunoprecipitateswere electrophoresed, transferred to nitrocellulose membranes, andblotted with the appropriate antibodies. Bound antibody was detectedwith 125I-protein A.
FIG. 3. Influence of time and TPAconcentration on the proteolysis ofErbB-4. T47-14 cells were treated for in-creasing periods of time with the indi-cated concentrations of TPA at 37 °C. Todetect ErbB-4 and the 80-kDa fragment,cell lysates (50 mg) were electrophoresed,transferred to nitrocellulose, and blottedwith ErbB-4 antiserum. Arrows indicatethe 180-kDa full-length ErbB-4 receptorand the 80-kDa fragment. 125I-Labeledprotein A was added to detect bound an-tibody and immunoreactive proteins werevisualized by autoradiography (Panel A).The 80-kDa fragment amounts werequantitated with a PhosphorImager (Pan-el B). The maximum level of the 80-kDafragment was obtained after a 1-h treat-ment with 100 ng/ml TPA, and this wasset at 100%.
ErbB-4 Degradation Mediated by Protein Kinase C18992
the TPA-induced cleavage of ErbB-4 is not inhibited by cyclo-heximide pretreatment and, therefore, does not require thesynthesis of new proteins.Influence of TPA on ErbB-4 Kinase Activity—While TPA
induces the cleavage of the ErbB-4 receptor, it is plausible thatthe resulting 80-kDa fragment might be an activated tyrosinekinase. In vivo and in vitro assays were performed to assess thekinase activity of the 80-kDa ErbB-4 fragment.To determine whether the TPA cleaved fragment was an
active tyrosine kinase in intact cells, cells were incubated withTPA or heregulin and the tyrosine phosphorylation of Shc andPLC-g1 were assayed. As shown in Fig. 6A, the tyrosine phos-phorylation of Shc and PLC-g1 by heregulin activation of thenative ErbB-4 receptor is evident (Fig. 6A, lanes 2 and 5), whileTPA treatment did not stimulate the tyrosine phosphorylationof either substrate (Fig. 6A, lanes 3 and 6). When unfraction-ated lysates from cells treated with TPA or heregulin wereprobed by anti-phosphotyrosine blotting, there was no evidenceof TPA-induced increases in total cell tyrosine phosphorylation(data not shown). However, the Shc and PLC-g1 immunopre-cipitates from TPA-treated cells do indicate the presence ofcoprecipitating proteins whose migration corresponds to that ofthe 80-kDa ErbB-4 fragment or the 180-kDa native ErbB-4receptor. Therefore, the blots were stripped and reprobed withantibody to ErbB-4 (Fig. 6B). The results show that both the80-kDa and native ErbB-4 molecules are coprecipitated withthese two substrates. The amount of Shc or PLC-g1 proteinpresent in each precipitate was similar regardless of cell treat-ment (data not shown). These results suggest that the 80-kDaErbB-4 fragment, although able to associate with Shc andPLC-g1, does not phosphorylate these potential substrates inintact cells.The tyrosine kinase activity of the 80-kDa ErbB-4 fragment
was also assayed in an immune complex assay in vitro. Controland TPA-treated cells were solubilized and ErbB-4 moleculesimmunoprecipitated from each lysate. The immunoprecipitateswere then resuspended in kinase buffer containing unlabeled
FIG. 5. Localization of the 80-kDa ErbB-4 fragment and release of the ErbB-4 extracellular domain into themedium. Panel A, T47-14cells were incubated in the absence or presence of TPA (100 ng/ml) for the indicated times. The cells were fractionated, as described under“Experimental Procedures,” into cytosolic (C) and membrane (M) fractions, and each fraction was probed with a polyclonal ErbB-4 antibody.Immunoprecipitates were separated on a polyacrylamide gel, transferred to nitrocellulose, and analyzed by Western blotting with anti-ErBb-4.Bound antibody was detected with 125I-labeled protein A and autoradiography. Solid arrows indicate positions of the full-length receptor and the80-kDa fragment. Panel B, subconfluent T47-14 cells grown in 60-mm dishes were incubated for 24 h in methionine-free Dulbecco’s modifiedEagle’s medium containing 1% dialyzed fetal calf serum and Trans35S-label (100 mCi/ml). After one wash, 0.5 ml of methionine-free Dulbecco’smodified Eagle’s medium containing 1% dialyzed fetal calf serum, 20 mM Hepes, pH 7.4, and 0.1% BSA was added. Cells were then treated withvehicle alone or TPA (100 ng/ml) for 30 min at 37 °C. The medium was then recovered and subjected to precipitation using an antibody to theextracellular domain of ErbB-4. The immunoprecipitates were extensively washed and loaded onto a polyacrylamide gel. The gel was dried andexposed to x-ray film. The open arrow indicates the position of the 120-kDa extracellular ErbB-4 domain.
TABLE IInfluence of protein kinase C modulators on formation of the
80-kDa ErbB-4 fragmentCells were incubated at 37 °C with the indicated modulators of pro-
tein kinase C for the indicated periods of time and subsequently as-sayed by Western blotting for the ErbB-4 80-kDa fragment, as dis-cussed under “Experimental Procedures.” 80-kDa fragment amountswere quantitated using a densitometer (Bio-Rad) and the amounts ofthe 80-kDa fragment obtained with TPA treatment considered 100%.For each treatment 10-, 30-, and 60-min time points were analyzed, andthe time at which the amount of the 80-kDa fragment was maximal isreported.
Additions 80-kDa fragment
relative %
Experiment 1Control 3TPA (100 ng/ml) 1 h 100TPA (100 ng/ml) 20 h 3TPA (100 ng/ml) 20 h 1 TPA (100 ng/ml) 1 h
3
GF 109203X (5 mM) 10 min 3GF 109203X (5 mM) 10 min 1 TPA (100ng/ml) 1 h
2
Experiment 2Control 12TPA (100 ng/ml) 1 h 100PDGF (100 ng/ml) 30 min 40sn-1, 2-Dioctanoylglycerol (200 mM) 30min
35
Cycloheximide (10 mg/ml) 15 min 1TPA (100 ng/ml) 1 h
77
Ionomycin (10 mM) 10 min 40
ErbB-4 Degradation Mediated by Protein Kinase C 18993
ATP with or without [g-32P]ATP. After incubation, the immu-noprecipitates containing unlabeled ATP were analyzed byWestern blotting with anti-ErbB-4, and those containing[g-32P]ATP were analyzed by autoradiography. As shown inFig. 7A, the native 180-kDa ErbB-4 receptor was intenselyphosphorylated in vitro, but the 80-kDa fragment was weaklyphosphorylated. The anti-ErbB-4 Western blot (Fig. 7B) indi-cates that approximately equal amounts of 180-kDa native and80-kDa fragment ErbB-4 protein were present in the assays.Normalizing for protein levels, the 80-kDa fragment is esti-mated to be 20% as active as the native ErbB-4 molecule.
DISCUSSION
Activators of protein kinase C modulate the activity and/orstructure of many transmembrane molecules encoding growthfactors and their receptors within the EGF/ErbB family. Reg-ulation of the EGF receptor and its ligands by protein kinase Coccurs at several levels: stimulation of transcription of mRNAencoding the ligands heparin-binding EGF (40, 41) and amphi-regulin (42); stimulation of the cleavage of the transmembraneforms of transforming growth factor-a (43–45) and heparin-binding EGF (46) to produce diffusable, extracellular, low mo-lecular weight ligands; and attenuation of EGF receptor func-tions by phosphorylation of serine/threonine residues within itscytoplasmic domain (47). ErbB-2 does not bind a known ligand,but rather forms heterodimeric complexes with the EGF recep-tor (15, 16), as well as ErbB-3 (11, 12) and ErbB-4 (14). ErbB-2is phosphorylated by protein kinase C and its tyrosine kinaseactivity decreased (28).Heregulin isoforms, which serve as ligands for the ErbB-3
and ErbB-4 members of the EGF receptor family (10), are alsosynthesized as transmembrane molecules and protein kinase Cactivation stimulates their cleavage leading to the release ofsoluble extracellular growth factors (48). The data in this re-port demonstrate that protein kinase C activation regulates theErbB-4 receptor by inducing a proteolytic cleavage that yieldsa membrane-localized cytoplasmic domain of 80 kDa and a
soluble ectodomain of 120 kDa. This is the first demonstrationof protein kinase C control of transmembrane receptors thatdirectly bind heregulin. Whether ErbB-3 is directly modulatedby protein kinase C activity is unknown, although our studiesdemonstrate that any such regulation would not likely includestimulation of proteolysis. Hence, the protein kinase C-stimu-lated cleavage of ErbB-4 is unique within the EGF receptorfamily of related transmembrane molecules.Protein kinase C activation does induce the cleavage of sev-
FIG. 6. Interaction of the 80-kDa fragment with Shc and PLC-g1 in vivo. T47-14 cells were incubated with vehicle alone, heregulin (100ng/ml), or TPA (100 ng/ml) for 1 h at 37 °C. After solubilization, cell lysates (500 mg) were precipitated with either anti-Shc or anti-PLC-g1. PanelA, the immunoprecipitates were analyzed by Western blotting with anti-phosphotyrosine, and bound antibody was visualized with 125I-labeledprotein A. Panel B, the anti-phosphotyrosine blots were stripped and reblotted with anti-ErbB-4. Bound antibody was detected using ECL. Solidarrows indicate the position of the full-length ErbB-4 receptor and the 80-kDa fragment. Open arrows indicate the position of Shc and PLC-g1.
FIG. 7. Autophosphorylation activity of the 80-kDa fragmentin vitro. T47-14 cells were treated at 37 °C with vehicle alone or TPA(100 ng/ml) for 1 h. Lysates (300 mg) were precipitated in duplicate fromeach sample, using a polyclonal antibody to the ErbB-4. The immuno-precipitates were washed, and the kinase assay was performed asdescribed under “Experimental Procedures,” at 37 °C for 5 min in theabsence or presence of [g-32P]ATP. Panel A, 32P-labeled samples wereseparated by gel electrophoresis and detected by autoradiography.Panel B, nonradioactive samples were separated by gel electrophoresisand Western blotted with anti-ErbB-4. Solid arrows mark the positionof the full-length ErbB-4 and the 80-kDa fragment.
ErbB-4 Degradation Mediated by Protein Kinase C18994
eral other receptor tyrosine kinases, including the colony-stim-ulating factor-1 receptor (150 kDa), the c-kit receptor (150kDa), and the receptor tyrosine kinase Axl (140 kDa). In eachcase, as with ErbB-4, proteolytic cleavage occurs at or near theextracellular face of the plasma membrane generating solubleectodomains of 100 kDa for the colony-stimulating factor-1receptor (49), 95 kDa for the c-kit receptor (50, 51), and 80 kDafor the Axl receptor (52). However, the protein kinase C acti-vated protease(s) for any growth factor precursor or transmem-brane receptor has not been identified. Cleavage of thesegrowth factor receptor tyrosine kinases produces cytoplasmicfragments that each contains a complete tyrosine kinase do-main. There is no evidence, however, that these membrane-localized or cytoplasmic tyrosine kinases are activated. Ourexperiments demonstrate that the tyrosine kinase fragment ofErbB-4 does not phosphorylate substrates in intact cells follow-ing TPA treatment and is also inactive in kinase assays invitro. The lack of activity in these tyrosine kinase moleculesmay be due to the loss of dimerization capacity which is gen-erally facilitated by the ligand-binding extracellular domains.The ErbB-4-derived 80-kDa cytoplasmic domain fragment doescontain phosphotyrosine and is, therefore, able to associatewith SH2 domain-containing molecules (Shc and PLC-g1) thatare ErbB-4 tyrosine phosphorylation substrates. The questionmay arise as to why this fragment is tyrosine-phosphorylatedin cells, if its kinase domain is not active. There are twoplausible rationales for this circumstance. The basal level ofautophosphorylation of ErbB-4 is high, and the tyrosine phos-phate present on the 80-kDa fragment may have been presenton the native receptor prior to proteolysis. Alternatively, thenative ErbB-4 receptor or another tyrosine kinase may utilizethe 80-kDa fragment as a substrate.We detect the ErbB-4 ectodomain as a 120-kDa fragment
present in the medium of cells exposed to TPA. In manypolypeptide hormone systems, soluble receptor ectodomainsare able to bind ligand and may, in some cases, serve asnegative regulators. We do not have evidence as to whether theErbB-4 120-kDa ectodomain fragment detected in these studiesis able to bind heregulin. However, Dong et al. (53) have ex-pressed the ectodomain of ErbB-4 as a soluble fusion proteinand shown that it is able to block the activity of neural differ-entiation factor, a heregulin isoform.
Acknowledgments—We thank Sandra Ermini for technical assist-ance and Sue Carpenter for assistance with manuscript preparation.Drs. Matthias Kraus and Paolo Di Fiore are acknowledged for antibodyand cell lines, and Dr. Mark Sliwkowski for heregulin.
REFERENCES
1. Earp, H. S., Dawson, T. L., Li, X., and Yu, H. (1995) Breast Cancer Res. Treat.35, 115–132
2. Gullick, W. J. (1990) Int. J. Cancer Suppl. 5, 55–613. Slamon, D. J., Clark, G. M., Wong, S. G., Levin, W. J., Ullrich, A., and
McGuire, W. L. (1987) Science 235, 177–1824. Slamon, D. J., Godolphin, W., Jones, L. A., Holt, J. A., Wong, S. G., Keith, D.
E., Levin, W. J., Stuart, S. G., Udove, J., Ullrich, A., and Press, M. F. (1989)Science 244, 707–712
5. Kraus, M. H., Issing, M., Popescu, N. C., and Aaronson, S. A. (1989) Proc. Natl.Acad. Sci. U. S. A. 86, 9193–9197
6. Lemoine, N. R., Barnes, D. M., Hollywood, D. P., Hughes, C. M., Smith, P.,Dublin, E., Prigent, S. A., Gullick, W. J., and Hurst, H. C. (1992) Br. J.Cancer 66, 1116–1121
7. Soler, C., and Carpenter, G. (1995) in Guidebook to Cytokines and theirReceptors (Nicola, N. A., ed) pp 194–197, Oxford University Press, Oxford
8. Mudge, A. W. (1993) Curr. Biol. 3, 361–3649. Marchionni, M. A., Goodearl, A. D. J., Chen, M. S., Bermingham-McDonogh,
O., Kirk, C., Hendricks, M., Danehy, F., Misumi, D., Sudhalter, J., Koba-yashi, K., Wroblewski, D., Lynch, C., Baldassare, M., Hiles, I., Davis, J. B.,Hsuan, J. J., Totty, N. F., Otsu, M., McBurney, R. N., Waterfield, M. D.,Stroobant, P., and Gwynne, D. (1993) Nature 362, 312–318
10. Tzahar, E., Levkowitz, G., Karunagaran, D., Yi, L., Peles, E., Lavi, S., Chang,D., Liu, N., Yayon, A., Wen, D., and Yarden, Y. (1994) J. Biol. Chem. 269,25226–25233
11. Sliwkowski, M. X., Schaefer, G., Akita, W. A., Lofgren, J. A., Fitzpatrick, V. D.,Nuijens, A., Fendly, B. M., Cerione, R. A., Vandlen, R. L., and Carraway, K.L., III (1994) J. Biol. Chem. 269, 14661–14665
12. Carraway, K. L., III, Sliwkowski, M. X., Akita, R., Platko, J. V., Guy, P. M.,Nuijens, A., Diamonti, A. J., Vandlen, R. L., Cantley, L. C., and Cerione, R.A. (1994) J. Biol. Chem. 269, 14303–14306
13. Plowman, G. D., Green, J. M., Culouscou, J.-M., Carlton, G. W., Rothwell, V.M., and Buckley, S. (1993) Nature 366, 473–475
14. Karunagaran, D., Tzahar, E., Beerli, R. R., Chen, X., Graus-Porta, D., Ratzkin,B. J., Seger, R., Hynes N. E., and Yarden, Y. (1996) EMBO J. 15, 254–264
15. Wada, T., Qian, X., and Green, M. (1990) Cell 61, 1339–134716. Goldman, R., Levy, R. B., Peles, E., and Yarden, Y. (1990) Biochemistry 29,
11024–1102817. Karunagaran, D., Tzahar, C., Liu, N., Wen, D., and Yarden, Y. (1995) J. Biol.
Chem. 270, 9982–999018. Riese, D. J., II, van Raaij, T. M., II, Plowman, G. D., Andrews, G. C., and Stern,
D. F., (1995) Mol. Cell. Biol. 15, 5770–577619. Kraus, M. H., Fedi, P., Starks, V., Muraro, R., and Aaronson, S. A. (1993) Proc.
Natl. Acad. Sci. U. S. A. 90, 2900–290420. Plowman, G. D., Culouscou, J.-M., Whitney, G. S., Green, J. M., Carlton, G. W.,
Foy, L., Neubauer, M. G., and Shoyab, M. (1993) Proc. Natl. Acad. Sci.U. S. A. 90, 1746–1750
21. Sorkin, A., and Waters, C. M. (1993) BioEssays 15, 375–38222. Sorkin, A., and Carpenter, G. (1993) Science 261, 612–61523. Sorkin, A., Di Fiore, P. P., and Carpenter, G. (1993) Oncogene 8, 3021–302824. Baulida, J., Kraus, M. H., Alimandi, M., Di Fiore, P. P., and Carpenter, G.
(1996) J. Biol. Chem. 271, 5251–525725. Brown, K. D., Dicker, P., and Rozengurt, E. (1979) Biochem. Biophys. Res.
Commun. 86, 1037–104326. Shoyab, M., De Larco, J. E., and Todaro, G. J. (1979) Nature 279, 387–39127. Magun, B. E., Matrisian, L. M., and Bowden, T. (1980) J. Biol. Chem. 255,
6373–638128. Cao, H., Decker, S., and Stern, D. (1991) Oncogene 6, 705–71129. Olashaw, N. E., O’Keefe, E. J., and Pledger, W. (1986) Proc. Natl. Acad. Sci.
U. S. A. 83, 3834–383830. Olashaw, N. E., and Pledger, W. J. (1987) J. Cell. Biochem. 34, 143–14931. Friedman, B.-A., and Rosner, M. R. (1987) J. Cell. Biochem. 34, 1–1132. Arteaga, C. L., Johnson, M. D., Todderud, G., Coffrey, R J., Carpenter, G., and
Page, D. L. (1991) Proc. Natl. Acad. Sci. U. S. A. 88, 10435–1043933. Sorkin, A., Helin, K., Waters, C. M., Carpenter, G., and Beguinot, L. (1992)
J. Biol. Chem. 267, 8672–867834. Bradford, M. M. (1976) Anal. Biochem. 72, 248–25435. Laemmli, U. K. (1970) Nature 227, 680–68536. Ballester, R., and Rosen, O M. (1985) J. Biol. Chem. 260, 15194–1519937. Chida, K., Kato, N., and Kuroki, T. (1986) J. Biol. Chem. 261, 13013–1301838. Nishizuka, Y. (1995) FASEB J. 9, 484–49639. Toullec, D., Pianetti, P., Coste, H., Bellevergue, P., Grand-Perret, T., Ajakane,
M., Baudet, V., Boissin, P., Boursier, E., Loriolle, F., Duhamel, L., Charon,D., and Kirilovsky, J. (1991) J. Biol. Chem. 266, 15771–15781
40. Temizer, D. H., Yoshizumi, M., Perrella, M. A., Susanni, E. E., Quertermous,T., and Lee, M.-E. (1992) J. Biol. Chem. 267, 24892–24896
41. Dluz, S. M., Higashiyama, S., Damm, D., Abraham, J. A., and Klagsbrun, M.(1993) J. Biol. Chem. 268, 18330–18334
42. Shoyab, M., McDonald, V., Bradley, J. G., and Todaro, G. J. (1988) Proc. Natl.Acad. Sci. U. S. A. 85, 6528–6532
43. Pandiella, A., and Massague, J. (1991) Proc. Natl. Acad. Sci. U. S. A. 88,1726–1730
44. Pandiella, A., and Massague, J. (1991) J. Biol. Chem. 266, 5769–577345. Pandiella, A., Bosenberg, M. W., Huang, E. J., Besmer, P., and Massague, J.
(1992) J. Biol. Chem. 267, 24028–2403346. Raab, G., Higashiyama, S., Hetelekidis, S., Abraham, J. A., Damm, D., Ono,
M., and Klagsbrun, M. (1994) Biochem. Biophys. Res. Commun. 204,592–597
47. Carpenter, G., and Cohen, S. (1990) J. Biol. Chem. 265, 7709–771248. Burgess, T. L., Ross, S. L., Qian, Y., Brankow, D., and Hu, S. (1995) J. Biol.
Chem. 270, 19188–1919649. Downing, J. R., Roussel, M. F., and Sherr, C. J. (1989) Mol. Cell. Biol. 9,
2890–289650. Yee, N. S., Langen, H., and Besmer, P. (1993) J. Biol. Chem. 268, 14189–1420151. Yee, N. S., Hsiau, C.-W. M., Serve, H., Vosseller, K., and Besmer, P. (1994)
J. Biol. Chem. 269, 31991–3199852. O’Bryan, J. P., Fridell, Y.-W., Koski, R., Varnum, B., and Liu, E. T. (1995)
J. Biol. Chem. 270, 551–55753. Dong, Z., Brennan, A., Liu, N., Yarden, Y., Lefkowitz, G., Mirsky, R., and
Jessen, K. R. (1995) Cell 15, 585–596
ErbB-4 Degradation Mediated by Protein Kinase C 18995