J. Cell Sci. 4, 751-762 (1969) 751 Printed in Great Britain THE NUTRITIONAL RELATIONSHIP OF ANEMONIA SULCATA (PENNANT) AND ITS DINOFLAGELLATE SYMBIONT D. L. TAYLOR* Department of Zoology, University College of Swansea and The Laboratory, Citadel Hill, Plymouth, England SUMMARY High resolution autoradiography in the optical and electron microscopes has been used to define the nature of the nutritional relationship between Anemonia sulcata and its algal symbiont. Grain counts from these autoradiographs have been examined quantitatively, and the results compared with those obtained from an in vitro analysis of the excreted products of the sym- bionts. These investigations clearly indicate that a system of carbon translocation between the alga and the host does exist in situ, and that over 60 % of the carbon fixed in photosynthesis is transferred to the host under the conditions of these experiments. INTRODUCTION It has been known for some time that algae in culture will commonly excrete a great deal of their photosynthetic products in the form of mucilages, peptides and organic acids (Lewin, 1956, 1961; Fogg, 1962; McLaughlin, Zahl, Nowak & Marchi- sotto, 1963), and that in the case of symbiotic species these metabolites may be of some potential value to the host. Nevertheless, Yonge & Nicholls (1931) concluded from their studies on Pacific Madreporaria that the host species does not actually receive anything of nutritional value from its symbionts, and that the beneficial aspects of the association are realized through the production of additional oxygen and the removal of the host's metabolic wastes by the algae. Similarly, Goreau & Goreau (i960) were unable to demonstrate the transfer of metabolites in situ using standard autoradiographic techniques on the West Indian corals Manicina areolata and Montastrea annularis. Their conclusions are in contrast with the work of both Odum & Odum (1955) and Muscatine & Hand (1958), who have used similar methods on coral reef communities and the anemone, Anthopleura elegantissima. They are also at variance with the more recent biochemical and chromatographic data of Muscatine (1967) and von Holt & von Holt (1968 a, b). The in vitro experiments of these authors have successfully demonstrated that some symbiotic algae are capable of excreting large quantities of extracellular metabolites in a form that can be directly utilized by the host. On the basis of this evidence, they postulate the transfer of these soluble products to the host where they could be used as a potential source of nourishment. • Present address: Haskins Laboratories, 305 East 43rd Street, New York, N.Y., 10017, U.S.A.
Transcript
J. Cell Sci. 4, 751-762 (1969) 751
Printed in Great Britain
THE NUTRITIONAL RELATIONSHIP OF
ANEMONIA SULCATA (PENNANT) AND ITS
DINOFLAGELLATE SYMBIONT
D. L. TAYLOR*Department of Zoology, University College of Swansea andThe Laboratory, Citadel Hill, Plymouth, England
SUMMARY
High resolution autoradiography in the optical and electron microscopes has been used todefine the nature of the nutritional relationship between Anemonia sulcata and its algal symbiont.Grain counts from these autoradiographs have been examined quantitatively, and the resultscompared with those obtained from an in vitro analysis of the excreted products of the sym-bionts. These investigations clearly indicate that a system of carbon translocation between thealga and the host does exist in situ, and that over 60 % of the carbon fixed in photosynthesis istransferred to the host under the conditions of these experiments.
INTRODUCTION
It has been known for some time that algae in culture will commonly excrete agreat deal of their photosynthetic products in the form of mucilages, peptides andorganic acids (Lewin, 1956, 1961; Fogg, 1962; McLaughlin, Zahl, Nowak & Marchi-sotto, 1963), and that in the case of symbiotic species these metabolites may be ofsome potential value to the host. Nevertheless, Yonge & Nicholls (1931) concludedfrom their studies on Pacific Madreporaria that the host species does not actuallyreceive anything of nutritional value from its symbionts, and that the beneficial aspectsof the association are realized through the production of additional oxygen and theremoval of the host's metabolic wastes by the algae. Similarly, Goreau & Goreau(i960) were unable to demonstrate the transfer of metabolites in situ using standardautoradiographic techniques on the West Indian corals Manicina areolata andMontastrea annularis. Their conclusions are in contrast with the work of both Odum& Odum (1955) and Muscatine & Hand (1958), who have used similar methods oncoral reef communities and the anemone, Anthopleura elegantissima. They are also atvariance with the more recent biochemical and chromatographic data of Muscatine(1967) and von Holt & von Holt (1968 a, b). The in vitro experiments of these authorshave successfully demonstrated that some symbiotic algae are capable of excretinglarge quantities of extracellular metabolites in a form that can be directly utilized bythe host. On the basis of this evidence, they postulate the transfer of these solubleproducts to the host where they could be used as a potential source of nourishment.
• Present address: Haskins Laboratories, 305 East 43rd Street, New York, N.Y., 10017,U.S.A.
752 D. L. Taylor
These conflicting points of view suggest that the problem of carbon translocationbetween symbiotic algae and their coelenterate hosts has not been adequately resolved,particularly with respect to the in situ demonstration of the actual transfer. A furtherinvestigation of the question was therefore extremely desirable. Anemonia sulcata(Pennant) was selected as the experimental host because previous investigationsprovided sufficient data on the physiology of the association (Trendelenburg, 1909;Putter, 1911; Smith, 1939) and the structural relationships of the host and its sym-biont (Taylor, 1968) to form a basis for the present study which can be used effectivelyin the interpretation of the final results. This work is concerned primarily with thein situ demonstration of an existing system of carbon translocation and includesreference to some in vitro experiments on the quantity of the alga's extracellularproducts.
MATERIALS AND METHODS
Specimens of Anemonia sulcata were collected from St Bride's Haven, Pembroke-shire, and maintained in circulating sea water until needed. The autoradiography ofspecimens for optical and electron microscopy was carried out using Na2
14CO3 (Radio-chemical Centre, Amersham) at a final concentration of 30 /tc/ml.
The experiment involved twenty specimens divided equally into two groups(A and B), each maintained at a constant temperature of 16 °C. Group A was kept incontinuous darkness, and group B under illumination for 18 h each day. Two speci-mens from each group were fixed for examination after 12, 24, 96, 168, and 240 hcontinuous exposure to the label. All sea water and dilutions of label were filteredthrough no. HAWG 04700 Millipore filters. These were tested for bacterial contami-nation and found to be sterile.
At the appropriate time intervals, the specimens were removed and fixed in 4%glutaraldehyde in cacodylate buffer (pH 7-0) for 12-20 h and post-fixed in 1 % OsO4
for 4 h. Following dehydration in alcohol, these were embedded in a standard mixtureof Epon 812 (Manton, 1964) and later sectioned on a Huxley ultramicrotome, usingglass knives, and left unstained. The reagents used for these procedures were laterexamined for the presence of radioactivity to detect the possible loss of 14C in solublecompounds. No losses were detected above the normal background af the samples. Asa further check on this important source of error, some specimens from each samplewere freeze-dried and directly infiltrated with Epon 812. Although tissue preserva-tion of these specimens leaves much to be desifed, they do serve to confirm theretention of the label in the preparations used for the interpretation of this work.
Thick plastic sections (1000-1500 A) of the glutaraldehyde-osmium fixed materialwere mounted on clean glass slides, coated with 50-60 A of evaporated carbon andcovered with a monolayer of Ilford L—4 emulsion. Unlabelled material was alsocoated in the same way as a control for chemography. Exposure was in total darknessat 4 °C and varied from 1 to 10 weeks. Development at weekly intervals consisted ofgold latensification (Rogers, 1967) followed by processing in Ilford Microphendeveloper. The developed autoradiographs were then air dried and the sections
Nutrition ofalgal-actinian symbiosis 753
stained through the emulsion with a 1 % aqueous solution of toluidine blue in caco-dylate buffer (pH 6-o) (Aoki & Gutierrez, 1967), washed in distilled water and airdried again. The preparations were examined with a Zeiss photomicroscope usingphase-contrast illumination and photographed on Ilford Micro Neg Pan film. Theresulting negatives were enlarged photographically (x5"s) and grain counts of theareas under consideration were made using the methods employed by Ostrowski &Sawicki (1961). The technique of visual grain counting in the optical microscope wasalso used to a limited extent. Corrections for background interference and differences
Table 1. Autoradiographic analysis of percentage excretionThe mean percentage excretion is derived directly from individual percentages
observed in the unit surface area of the algal and animal cells in the section weremade (Rogers, 1967), and the results expressed as mean grain counts/10/i2 algae andmean grain counts/io/*2 host tissue. The percentage of algal photosynthate excretedto the host was also calculated from these grain counts (Table 1, Fig. 1 B).
Thin sections for electron microscopy were prepared according to the methods ofSalpeter & Bachmann (1964) using Ilford L-4 emulsion. These were exposed asabove and developed in Kodak Microdol X with gold latensification. The finishedautoradiographs were examined with an AEI EM6 electron microscope operated at100 kV and a Philips EM 200 operated at 80 kV to detect the possible pathways ofcarbon translocation in situ. These preparations were not used in the relative measure-ments of radioactivity described above.
Homogenates of Anemonia and its symbiont were prepared and separated followingthe methods described by Muscatine (1967). After separation the algae were re-suspended in either (1) ASP 8-A medium (McLaughlin & Zahl, 1966), (2) fresh hosthomogenate (Muscatine, 1967) or (3) host homogenate boiled for 2 min and allowed to
754 D. L. Taylor
equilibrate for one hour. These suspensions were then incubated in stopperedcentrifuge tubes with 30 /ic Na2
14CO3 for 30 min at 16 °C under the same illumina-tion used above. Following incubation, the algae were separated from the suspendingmedium by centrifugation and washing, and the total radioactivity in each (i.e. thealgae and the suspending medium) was assayed. The results of this analysis aresummarized in Table 2.
2 20m
30 -
10 -
RESULTS
Optical microscopy
Group A. Sections examined from specimens kept in the dark confirm that label isnot present in the animal tissues as a result of the independent fixation of 14C by thehost (Fig. 2). In view of this, all of the label recorded below is considered to be thedirect result of the fixation of 14C by the photosynthetic activity of the algal symbiont.
~ 60fl r 50 -
40 -
30 -
20 -
,2 10 -
o
60
°5 50 -t-t
| 40 -
. A
-
(«•<>
( 5 2 3 ^
(28-1)
. B
1 1 1 |
(52-5)'
,
i i i i
(511)]T
,— " 1(48-8)
I I I I
20 40 60 160 180 200 220 24080 100 120 140Hours of total exposure
Fig. 1 A. Total uptake of 14C during the experimental period, derived from relativemeasurements of radioactivity in autoradiographic sections.Fig. 1 B. Percentage excreted photosynthate detected at 12, 24, 96, 168, 240 h exposureto the label (see p. 753).
Nutrition of algal-actinian symbiosis 755
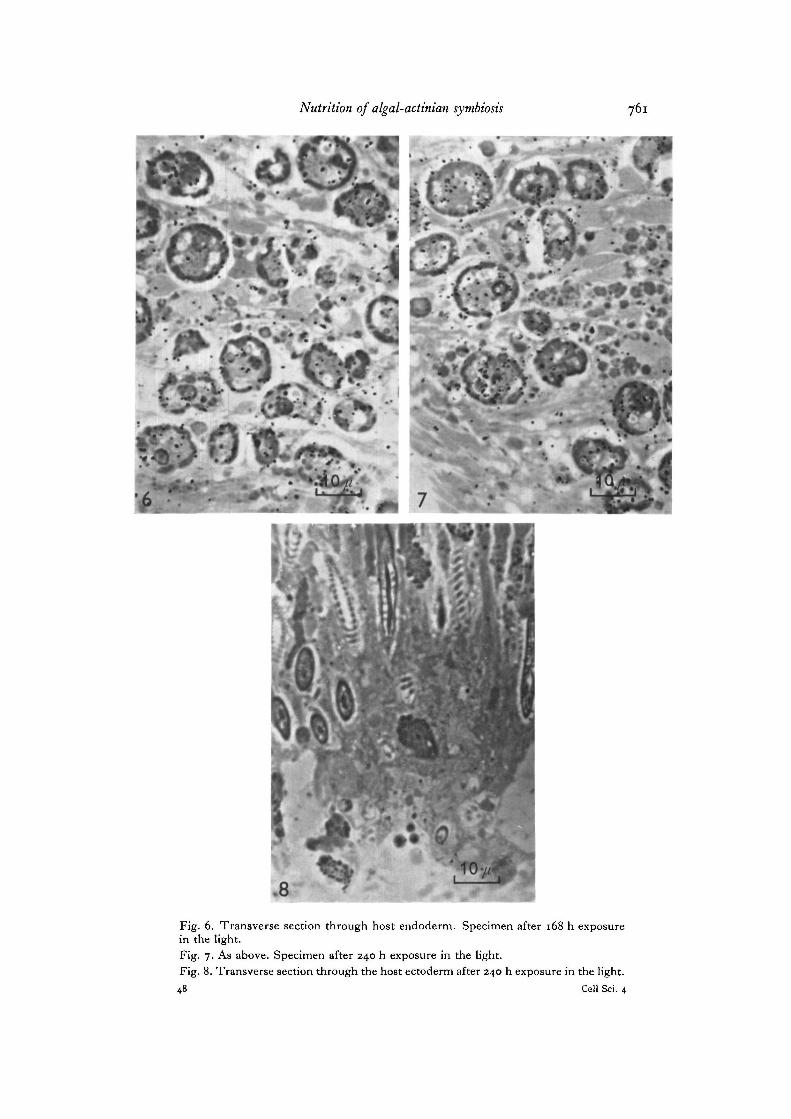
Group B. Thick plastic sections of specimens from this group exhibit a generalincrease in the total number of silver grains recorded within the algal cells andthroughout the endodermal tissues of the animal (Fig. 1 A). The uptake associatedwith this increase apparently begins before the first sample was taken at 24 h (shownby the in vitro experiments discussed below) and continues for the duration of theexperiment. Grain counts of these autoradiographs analysed by the method previouslydescribed (Table 1, Figs. 3-7) show that a fairly constant level of excretion is main-tained by the algae throughout the experiment (Fig. IB), and that these excretedproducts are apparently incorporated into the host tissues by the activities of theanimal. Further examination reveals thatvery littleof this photosynthate is subsequentlytransported to the host's mesoderm, and that no trace of it is found amongst the cellsof the ectoderm even after 240 h continuous exposure to the label (Fig. 8). In view ofthis, it is thought that the mesoderm may possibly act to slow down or exclude thetransport of the algal photosynthate to the ectodermal cells. A considerably longerperiod of initial exposure to the label appears to be needed before this problem can beadequately resolved. This, unfortunately, was beyond the capacity of these studies.However, similar results have been recorded previously by Muscatine & Hand (1958)and by Taylor (1967) in experiments using standard autoradiography and strippingfilm techniques.
Electron microscopy
High resolution autoradiographs were examined in the electron microscope todefine the nature of the carbon translocations detected above, and to determinewhether the mechanism involved was an active or passive process on the part of thehost. The results of these autoradiographs are represented in Figs. 9 and 10.
Regardless of the period of exposure to the label, the overall appearance of theseautoradiographs at the subcellular level is the same in all cases. In some of the sectionsnumerous vesicles are seen in the cytoplasm of the host cells, suggesting the possibilitythat the host is actively taking up the excreted photosynthate through pinocytoticactivity in its endodermal cells. However, the actual stages in the formation of pino-cytotic vesicles have not been observed here, and there is no direct evidence from theautoradiographs themselves to support the view that this represents a mechanism ofuptake. Instead, it appears as if the opposite is the more correct, as shown in Fig. 10,where the vesicles noted above are almost entirely absent from the host cytoplasm.This appearance suggests that the translocation observed above is more likely totake place as the photosynthetic products of the algae are leaked into the surroundinghost tissues and passively absorbed by the endodermal cells. The random scatteringof the label seen in the host's cytoplasm tends to support this view still further.Nevertheless, the active uptake of algal photosynthate by pinocytosis in the hostshould not be entirely ruled out, since the precise localization of label in the vesicles ofhost cells surrounding the symbiont may be beyond the resolving power of the presenttechnique in most instances. At the present time, however, circumstantial evidenceseems to exclude this possibility.
756 D. L. Taylor
Excretion of the photosynthate in vitro
Algae which were separated from their host tissues by the method described above,and incubated in ASP-8A medium with Na2
14CO3) were found to fix high levels oflabelled carbon, but excreted very little of their photosynthate when compared withsimilar suspensions incubated in the presence of fresh host homogenate (Table 2).Under the latter conditions a maximum of up to 58% of the labelled photosynthatewas found to be present in the incubating medium after 30 min, suggesting that thereis some degree of host control over the level of excretion. When boiled host homogenatewas substituted for the fresh suspension used above, the level of excretion was found tobe noticeably lower (28 %), but not as low as that observed with algae incubated inASP-8A alone (Table 2). Animal homogenate incubated as above, but without thepresence of the algae, did not fix appreciable carbon in the course of theseexperiments.
DISCUSSION
The results obtained from previous studies on the symbiotic association of A. sulcataand its alga clearly demonstrate that the relationship results in numerous beneficialeffects for the host which may be detected as a general increase in the biologicalefficiency of the anemone (Trendelenburg, 1909; Putter, 1911; Smith, 1939;Taylor, 1967, 1968). The present investigation shows that some of these benefitsare realized through an endogenous system of carbon translocation which has beenfound to exist between the host and its symbiont and is capable of yielding a maximumof over 60% of the algal photosynthate to the host's metabolism. This conclusion isbased upon relative measurements of radioactivity obtained from high resolutionautoradiographs and compares favourably with the results obtained from an in vitroanalysis of the percentage excreted photosynthate. Observations from autoradiographsexamined in the electron microscope suggest that the transfer of this material involvesa passive uptake on the part of the host. The possibility that an active uptake oftranslocated carbon takes place by pinocytosis in the host cells remains, and will beconsidered again in the discussion which follows.
On the whole, these findings are consistent with the results obtained by Muscatine(1967) from in vitro studies on the excreted products of the symbionts from a tridacnidand from a Pacific coral, and confirm that the system of carbon translocation which heoriginally described can be clearly demonstrated in situ. Muscatine (1967) found thatthe symbionts which he studied liberated glycerol as their principal extracellularproduct, and that the rate of excretion was influenced by the presence of host cells.At the present time, there are no data available on the exact chemical nature of thephotosynthate excreted by the Ammonia symbionts; it seems very likely, however,that this also will be found to be glycerol, since the symbionts of the Tridacnidae andthose of Ammonia seem to be taxonomically identical (Taylor, unpublished obser-vations). Control over the rate of excretion by the presence of host cells has also beendemonstrated here by an in vitro study of the percentage excretion (Table 2). The fact
Nutrition of algal-actinian symbiosis 757
that the results obtained from autoradiographs are significantly close to those derivedfrom isolated symbionts incubated in the presence of host homogenate suggests thathost control over the rate of excretion by the algae was fully operative throughout theautoradiographic experiments. Previous studies on the ultrastructure of this symbiont(Taylor, 1968) have shown that the algal cell wall is a morphologically complexstructure which could serve to block the type of translocation which has been foundhere. The observation that high levels of translocation can take place only in thepresence of host cells suggests that the mechanism of host control is fully capable ofovercoming this barrier and therefore seems to represent a very powerful influenceover the functioning of the algal cell. In view of this, it may be thought surprising thatthe uptake of the algal photosynthate is apparently a passive process (see above).Nevertheless, the intracellular situation of the symbionts (Taylor, 1968) would seemto exclude the necessity for a complex mechanism for the uptake of excreted carbon bythe host. Indeed this intracellular position may explain why no definite proof ofpinocytosis exists, since this mechanism would be superfluous in a situation wherepassive uptake in the cytoplasm of the host cells would be a completely adequatemechanism.
Given the fact that a system of carbon translocation exists between Anemonia andits symbiont, one is compelled to ask whether the host derives a large portion ofits nutritional requirements from this source. The experimental evidence of Smith(1939) and Taylor (1967) suggests that this is clearly not the case. It has been shownrepeatedly that if Anemonia is supplied with an external source of nourishment, itcan survive indefinitely without the aid of the photosynthetic products of its sym-bionts. If, on the other hand, it is deprived of an exogenous food source and made torely solely on the excreted photosynthate of its algae, depression phenomena resultwhich lead to reduced biological activity and ultimately death in the majority ofcases (Taylor, 1967, unpublished observations). It would seem from this that theexcreted products of the algae are of value only as a supplement to the host's metabolism,and that the primary source of nourishment comes from an exogenous food supply.It would be easy to overemphasize the importance of carbon translocation in thisparticular symbiotic system.
Although the present investigations and the recent studies of Muscatine (1967) andvon Holt & von Holt (1968a, b) suggest that some form of carbon translocation islikely to take place in almost all symbiotic associations of this type, the type of meta-bolic interdependence which can exist is likely to vary in degree. One extreme givessemi- or complete independence of exogenous food to host species like Zoanthus(Goreau, 1967), whilst the opposite situation is found amongst predatory hosts such asAnemonia and the hermatypic corals.
If the results of the present study are examined in their proper context along withthe information gained from previous investigations (Trendelenburg, 1909; Putter,1911; Smith, 1939; Taylor, 1967, 1968), the true nature of the association becomesapparent. It now appears certain that Anemonia receives only secondary nutritionalbenefits from the endogenous system of carbon translocation provided by its associa-tion with a symbiotic alga, and that this cannot replace the natural feeding habit of the
758 D. L. Taylor
animal as a primary source of nourishment. The supplementary nourishment whichthis system does supply could, however, be of great ecological benefit to the entirepopulation, helping to account for Anemonia being one of the commonest actinians onBritish and European coasts.
The author wishes to thank Professor E. W. Knight-Jones for advice and laboratory facilities,including equipment provided by the Science Research Council, and the Director of the Ply-mouth Laboratory for laboratory facilities there, including the use of a Philips electron micro-scope provided by the Wellcome Fund. The author was in receipt of a N.A.T.O. PostdoctoralFellowship.
REFERENCES
AOKI, A. & GUTIERREZ, L. S. (1967). A simple toluidine blue-basic fuchsin stain for spermatozoain epoxy sections. Stain Technol. 42, 307-310.
FOGG, G. E. (1962). Extracellular products. In Physiology and Biochemistry of tlie Algae (ed.R. A. Lewin), pp. 475-493. New York and London: Academic Press.
GOREAU, T. F. (1967). Quoted from von Holt, C. & von Holt, M. (1968). Transfer of photo-synthetic products from zooxanthellae to coelenterate hosts. Comp. Biochem. Pliysiol. 24,73-8i.
GOREAU, T. F. & GOREAU, N. I. (i960). Distribution of labelled carbon in reef building coralswith and without zooxanthellae. Science, N. Y. 131, 668-669.
HOLT, C. VON & HOLT, M. VON (1968a). Transfer of photosynthetic products from zooxan-thellae to coelenterate hosts. Comp. Biochem. Physiol. Z4, 73-81.
HOLT, C. VON & HOLT, M. VON (19686). The secretion of organic compounds by zooxanthellaeisolated from various types of Zoanthus. Comp. Biochem. Physiol. 24, 83-92.
LEWIN, R. A. (1956). Extracellular polysaccharides of green algae. Can.J. Microbiol. 2, 665-672.LEWIN, R. A. (1961). Phytoflagellates and algae. In Handbuch der Pflanzenpliysiologie, vol. 4
(ed. W. Ruhland), pp. 401-417. Berlin: Springer.MANTON, I. (1964). Observations on the fine structure of the zoospore and germling of Stigeo-
clonium.J. exp. Bot. 15, 399-411.MCLAUGHLIN, J. J. A. & ZAHL, P. A. (1966). Endozoic algae. In Symbiosis, vol. 1 (ed. M. S.
Henry), pp. 257-297. New York: Academic Press.MCLAUGHLIN, J. J. A., ZAHL, P. A., NOWAK, A. & MARCHISOTTO, J. (1963). Some constituents
of zooxanthellae grown in axenic culture. Proc. 1st int. Congr. Protozool., Prague, pp. 204-205. Prague: Czech. Acad. Sci.
MUSCATINE, L. (1967). Glycerol excretion by symbiotic algae from corals and Tridacna and itscontrol by the host. Science, N.Y. 156, 516-519.
MUSCATINE, L. & HAND, C. (1958). Direct evidence for the transfer of materials from sym-biotic algae to the tissues of a coelenterate. Proc. natn. Acad. Sci. U.S.A. 44, 1259-1263.
ODUM, H. T. & ODUM, E. P. (1955). Trophic structure and productivity of a windward coralreef community on Enjwetok Atoll. Ecol. Monogr. 25, 291—320.
OSTROWSKI, K. & SAWACKI, W. (1961). Photomicrographic method for counting photographicgrains in autoradiograms. Expl Cell Res. 24, 625-628.
POTTER, A. (1911). Der Stoffwechsel der Aktinien. Z. allg. Physiol. 12, 297-322.ROGERS, A. W. (1967). Techniques of Autoradiography. London, Amsterdam and New York:
Elsevier.SALPETER, M. M. & BACHMANN, L. (1964). Autoradiography with the electron microscope. I.
A procedure for improving resolution, sensitivity and contrast. .7. Cell Biol. 16, 383-391.SMITH, H. G. (1939). The significance of the relationship between actinians and zooxanthellae.
J. exp. Biol. 16, 334-345-TAYLOR, D. L. (1967). Symbiotic Relationships Between Some Marine Plants and Animals.
Ph.D. Thesis, The University of Wales.TAYLOR, D. L. (1968). In situ studies on the cytochemistry and ultrastructure of a symbiotic
marine dinoflagellate. J. mar. biol. Ass. U.K. 48, 349-366.
Nutrition of algal-actinian symbiosis 759
TRENDELENBURG, W. (1909). Versuche iiber den Gaswechsel bei Symbiose zwischen Alga undTier. Arch. Anat. Physiol. 42, 42-70.
YONGE, C. M. & NICHOLLS, A. G. (1931). Studies on the physiology of corals. V. The effects ofstarvation, in light and in total darkness, on the relationship between corals and zooxan-thellae. Scient. Rep. Gt Barrier Reef Exped. 1, 177-211.
(Received 5 July 1968)
D. L. Taylor
Fig. 2. Transverse section through host endoderm. Dark control after 240 h exposureto the label, (h, host tissue; z, zooxanthellae).Fig. 3. As above. Specimen after 12 h exposure in the light.Fig. 4. As above. Specimen after 24 h exposure in the light.Fig. 5. As above. Specimen after 96 h exposure in the light.
Nutrition of algal-actinian symbiosis 761
#1
Fig. 6. Transverse section through host endoderm. Specimen after 168 h exposurein the light.Fig. 7. As above. Specimen after 240 h exposure in the light.Fig. 8. Transverse section through the host ectoderm after 240 h exposure in the light.48 Cell Sci. 4
762 D. L. Taylor
10
Fig. 9. Electron-microscopic autoradiograph of a zooxanthella (a) in the host ectoderm(h) after 24 h exposure to the label in the light (see text p. 755).Fig. 10. As above, after Z40 h exposure to the label in the light.