J Comp PhysiolA (1992) 170:565-574 Jom'l~l of Sensor-/, and Physlology Springer-Verlag 1992 The refractive development of the eye of the American kestrel (FMco sparverius): a new avian model M.E. Andison 1, J.G. Sivak 1. and D.M. Bird 2 z School of Optometry, Universityof Waterloo, Waterloo, Ontario, Canada N2L 3G1 2 Macdonald Raptor Research Centre, Macdonald Campus of McGill University, 21, 111 Lakeshore Road, Ste. Anne de Bellevue, Quebec, Canada H9X ICO Accepted February 26, 1992 Summary. Most measures of avian visual performance are carried out on commonly available domestic species such as the chicken, and most of the data on avian induced refractive error deals with chickens. Raptors are predatory birds in which good visual resolving ability is particularly important. Behavioral studies indicate that the eyes of raptors have two to three times the resolving ability of the human eye. The domestic chicken is preco- cial at hatching whereas most raptors are semi-altricial. This study was an effort to determine if the effect of early visual deprivation on the refractive development of the chicken eye can be reproduced in the American kestrel, a species which is not domesticated and in which the need for acute vision is particularly important. Visual deprivation was achieved by unilaterally apply- ing translucent plastic goggles over the eyes of kestrels two days after hatching. Refractive error was measured using a retinoscope and trial lenses. Ocular growth was monitored by A-scan ultrasonography, and frozen oc- ular sections of sacrificed birds. The effect of the experi- mental manipulation on the contralateral control eye and body weight was evaluated each day over a 42-day period. The goggles did not significantly affect the nor- mal changes in body weight or the normal pattern of ocular growth and refractive development in the un- treated eyes. An analysis of the refractive state changes as a result of form deprivation was made each week for 6 weeks after hatching on both the treated and untreated eyes in a separate group of experimental birds. Visual form deprivation caused a significant myopic shift in refractive error and a significant increase in the vitreous chamber depth in the treated eyes at 3 and 6 weeks of age. However, the amount of myopia produced is much less than that induced in chicks, and in certain cases hyper- opia is produced. The kestrels recover from myopia and hyperopia within 10 days of goggle removal, after 3 to 4 weeks of deprivation. This study is the first indication that chickens may not be a representative bird model for studying form- * To whom offprint requests should be sent deprivation myopia. First, myopia is not always produced in kestrels in response to form deprivation. Second, kestrels are severely myopic at hatching and therefore, the direction of emmetropization is opposite to that found in hatchling chicks. Key words: Eye - Refractive error - Development - Kestrel - Myopia Introduction Form-deprivation myopia is a developmental anomaly resulting from abnormal visual experience. The discovery in monkeys that form-deprivation by lid-suturing causes axial elongation and a myopic shift in refractive error was made by Hubel et al. (1975, cited by Curtin 1985) during the course of other experimental work. Since this time, lid-suture myopia has been induced in monkeys (Wiesel and Raviola 1977), tree shrews (Sherman et al. 1977), cats (Wilson and Sherman 1977) and chicks (Wall- man et al. 1978). In addition to lid-suturing and corneal opacification (Wiesel and Raviola 1979), goggles of various types have been used to degrade the retinal image. These include opaque (Sivak et al. 1989), translucent (Wallman et al. 1978; Pickett-Seltner et al. 1987) or partial occluders (Wallman et al. 1978). Concave and convex spectacle lenses (Schaeffel et al. 1988), concave and convex soft contact lenses (Sivak et al. 1990) and concave and convex hard contact'lenses fit to a modified goggle (Irving et al. 1991) have been used to produce defocus of the retinal image. Restriction of the visual environment (Young 1963), continuous light (Axsmith and Morin 1975) and low-intensity blue light (Bercovitz et al. 1972) have produced refractive errors in a variety of animal models. Raising chickens in complete darkness has been shown to produce hyperopia (Hodos 1990). A number of different models and mechanisms of myopia development have been suggested. Even within
Transcript
J Comp Physiol A (1992) 170:565-574 Jom'l~l o f
Sensor-/,
and
Physlology
�9 Springer-Verlag 1992
The refractive development of the eye of the American kestrel (FMco sparverius): a new avian model
M.E. Andison 1, J.G. Sivak 1. and D.M. Bird 2
z School of Optometry, University of Waterloo, Waterloo, Ontario, Canada N2L 3G 1 2 Macdonald Raptor Research Centre, Macdonald Campus of McGill University, 21, 111 Lakeshore Road, Ste. Anne de Bellevue,
Quebec, Canada H9X ICO
Accepted February 26, 1992
Summary. Most measures of avian visual performance are carried out on commonly available domestic species such as the chicken, and most of the data on avian induced refractive error deals with chickens. Raptors are predatory birds in which good visual resolving ability is particularly important. Behavioral studies indicate that the eyes of raptors have two to three times the resolving ability of the human eye. The domestic chicken is preco- cial at hatching whereas most raptors are semi-altricial. This study was an effort to determine if the effect of early visual deprivation on the refractive development of the chicken eye can be reproduced in the American kestrel, a species which is not domesticated and in which the need for acute vision is particularly important.
Visual deprivation was achieved by unilaterally apply- ing translucent plastic goggles over the eyes of kestrels two days after hatching. Refractive error was measured using a retinoscope and trial lenses. Ocular growth was monitored by A-scan ultrasonography, and frozen oc- ular sections of sacrificed birds. The effect of the experi- mental manipulation on the contralateral control eye and body weight was evaluated each day over a 42-day period. The goggles did not significantly affect the nor- mal changes in body weight or the normal pattern of ocular growth and refractive development in the un- treated eyes. An analysis of the refractive state changes as a result of form deprivation was made each week for 6 weeks after hatching on both the treated and untreated eyes in a separate group of experimental birds. Visual form deprivation caused a significant myopic shift in refractive error and a significant increase in the vitreous chamber depth in the treated eyes at 3 and 6 weeks of age. However, the amount of myopia produced is much less than that induced in chicks, and in certain cases hyper- opia is produced. The kestrels recover from myopia and hyperopia within 10 days of goggle removal, after 3 to 4 weeks of deprivation.
This study is the first indication that chickens may not be a representative bird model for studying form-
* To whom offprint requests should be sent
deprivation myopia. First, myopia is not always produced in kestrels in response to form deprivation. Second, kestrels are severely myopic at hatching and therefore, the direction of emmetropization is opposite to that found in hatchling chicks.
Form-deprivation myopia is a developmental anomaly resulting from abnormal visual experience. The discovery in monkeys that form-deprivation by lid-suturing causes axial elongation and a myopic shift in refractive error was made by Hubel et al. (1975, cited by Curtin 1985) during the course of other experimental work. Since this time, lid-suture myopia has been induced in monkeys (Wiesel and Raviola 1977), tree shrews (Sherman et al. 1977), cats (Wilson and Sherman 1977) and chicks (Wall- man et al. 1978).
In addition to lid-suturing and corneal opacification (Wiesel and Raviola 1979), goggles of various types have been used to degrade the retinal image. These include opaque (Sivak et al. 1989), translucent (Wallman et al. 1978; Pickett-Seltner et al. 1987) or partial occluders (Wallman et al. 1978). Concave and convex spectacle lenses (Schaeffel et al. 1988), concave and convex soft contact lenses (Sivak et al. 1990) and concave and convex hard contact'lenses fit to a modified goggle (Irving et al. 1991) have been used to produce defocus of the retinal image. Restriction of the visual environment (Young 1963), continuous light (Axsmith and Morin 1975) and low-intensity blue light (Bercovitz et al. 1972) have produced refractive errors in a variety of animal models. Raising chickens in complete darkness has been shown to produce hyperopia (Hodos 1990).
A number of different models and mechanisms of myopia development have been suggested. Even within
566 M.E. Andison et al. : Eye development of American kestrel
the same genus, inves t iga tors bel ieve tha t one m o d e l m a y n o t be sufficient. R a v i o l a and Wiesel (1990) hypothes ize tha t there m a y be two different p r i m a t e models , one for rhesus m o n k e y s (Macaca mulatta) a n d a no the r for s t ump- t a i l ed m o n k e y s ( Macaca arctoides ).
I t is no t poss ib le to a p p l y d i rec t ly the results f r om av ian mode l s o f m y o p i a to m a m m a l s / h u m a n s , due a va r ie ty o f a n a t o m i c a l a n d phys io log ica l differences. These inc lude such fea tures as eye shape and vascu la r iza - t ion, the presence o f a n ic t i t a t ing m e m b r a n e , annu la r p a d and scleral ossicles in b i rds (Wal l s 1942), differences in the a c c o m m o d a t i v e m e c h a n i s m (Wel ty 1982) and dif- ferences in response to p h a r m a c o l o g i c a l agents (Wal ls 1942). Howeve r , the ch icken (Gallus 9allus domesticus) has become a wide ly used and accep ted av ian mode l for s tudy ing m y o p i a . Ch ickens are inexpensive and easi ly ma in t a ined . They have a r a p i d ra te o f g rowth and de- v e l o p m e n t and have p r o p o r t i o n a t e l y large eyes ( H o d o s 1990). Howeve r , is the chicken a represen ta t ive b i rd mode l?
F a l c o n i f o r m e s such as eagles, fa lcons and hawks are r epu ted to have excep t iona l ly keen visual acuity. Thei r visual acu i ty has been empi r i ca l ly shown to exceed tha t o f h u m a n s (Shlaer 1972; F o x et al. 1976). A n a t o m i c a l evidence to s u p p o r t this c la im is p r o v i d e d by the fal- c o n i f o r m re t ina which has a h igher dens i ty o f cones than the h u m a n re t ina (Wal l s 1942). Us ing an oph tha l - m o s c o p i c m e t h o d , Sh lae r (1972) m e a s u r e d the re t inal image qua l i ty o f the A f r i c a n se rpent eagle (Dryotriorchis spectabilis) a n d d e t e r m i n e d its visual r e so lu t ion to be- tween 2.0 a n d 2.4 t imes h u m a n reso lu t ion .
A classic two-cho ice d i s c r imina t i on t a sk has been used to b e h a v i o r a l l y test the visual acu i ty o f an A m e r i c a n kestre l (Falco sparverius). The resul ts showed tha t the kestre l has a t h r e sho ld va lue o f 160 cycles /degree, a value which is 2.7 t imes the h u m a n g ra t ing acu i ty value o f 60 cycles /degree ( F o x et al. 1976).
A m a j o r difference be tween kest re ls and chick is tha t kest re ls a re semi-a l t r ic ia l while chicks are precocia l . A t ha tch ing , semi-a l t r ic ia l b i rds a re d o w n - c o v e r e d and are cha rac t e r i zed by p a r t i a l l y c losed eyes and a dependency u p o n the i r p a r e n t s for f o o d for some t ime af ter ha tching. A t ha tch ing , p recoc ia l b i rds a re down-cove red , have open eyes a n d are i n d e p e n d e n t o f thei r pa ren t s for food (Wel ty 1982). The degree o f visual acu i ty requ i red by each o f these species a t ha t ch ing and as they deve lop m a y be an a d a p t a t i o n to the s t ra tegies they use to o b t a i n food. A n o t h e r p o i n t o f difference be tween chickens and kes- t rels is t ha t ch ickens are h igh ly d o m e s t i c a t e d and kestrels a re wild. I t is conce ivab le tha t these fac tors m a y be ref lected in differences seen in the refract ive d e v e l o p m e n t o f each o f these b i rds a t ha t ch ing a n d as they mature .
The p u r p o s e o f the p resen t s tudy was to examine and eva lua te whe the r the ch icken is a represen ta t ive av ian m o d e l for s tudy ing the genesis o f f o r m - d e p r i v a t i o n m y o - pia. A n o t h e r av ian species, the A m e r i c a n kestrel , which has di f ferent behav io r a l a n d visual a d a p t a t i o n s , was used to de t e rmine w h a t effects f o r m - d e p r i v a t i o n wou ld have on its re f rac t ive deve lopmen t .
Methods
A total of 80 American kestrels were used in the study. Approxi- mately 300 kestrels are bred per year at the Macdonald Raptor Research Centre, of McGill University in Montreal, Quebec, Cana- da, primarily for pesticide research (Bird 1982). None of these birds are normally released.
Care of the kestrels. All of the birds used were hatched under artificial conditions in forced-air RolI-X incubators (Marsh Farms) maintained at 37.5 ~ and 45% humidity. At pip, eggs were placed within marked wire mesh corrals to facilitate identification. They were then transferred to a modified incubator in which the turning grid was replaced with quarter-inch mesh hardware cloth covered with krinoline cloth. The temperture was reduced to 37.0 ~ and the humidity was increased to 75 %. Within 24 h after hatching, the chicks were dry. They were weighed to the nearest 1 x 10-2 g on a Mettler top-loading balance prior to their first feeding. The chicks were subsequently placed in a wooden still-air brooder (60 cm • 55 cm x 18 cm) maintained between 34 ~ and 36 ~ at 50 to 60% humidity. They remained in this brooder for 5-7 days in cardboard corrals on a substrate of wood shavings in groups of 4-6. The chicks were then moved through a series of still-air brooders (55 cm • 55 cm x 30 cm). The temperature was reduced daily by 0.55 ~ per day and the humidity was lowered to between 35 and 40%, until the chicks were able to survive at ambient room tem- perature (27 ~ ~ at approximately two weeks of age. From this time until they were sacrificed, chicks were housed on a sub- strate of wood chips in cardboard boxes (30 cm • 30 cm x 35 cm) with removable wire mesh tops.
Control birds. A group of 10 ungoggled kestrels was used as controls to study normal refractive development and eye growth. These birds were weighed each day before their first feeding. For two days after hatching, it was not possible to measure the refractive error because the chicks' eyelids were naturally constricted. At hatch, pulling the lower lid down revealed an opening only 1.5 mm in diameter. By two days of age, it was possible to see a retinoscopic reflex and the refractive error could be assessed. Refractive measures were made for both eyes to the nearest 0.5 diopter in the horizontal meridian each day using a standard clinical streak-retinoscope and trial lenses. The retinoscope was powered via a transformer to prevent fluctuations in the beam intensity during prolonged periods of use. A working distance of 0.5 m was used. It was assumed that the kestrels were fixating the environment beyond the experimenter. Ultrasonography was performed on both eyes each day using an Oculometer 4000 A-scan ultrasound (Radionics Medical) to assess axial length. Control birds were sacrificed by decapitation at 6 weeks of age.
A freeze-sectioning technique was used to measure intraocular dimensions. The heads of 8 kestrels were placed on the stage of a freezing microtome which was connected to a cylinder of pres- surized carbon dioxide. The head was trimmed horizontally to the level of the eyes with the microtome blade. A single lens reflex camera was mounted on a tripod and arranged over the sectioning apparatus. Photographs were taken of the head as serial sections were removed. A millimetre ruler placed at the level of the head was included in each frame taken as a control for magnification. Contact sheets were developed and measured to establish the frame showing the largest eye diameter, and hence equatorial eye dimensions. Slides of the selected frames were made and tracings were drawn of the projected corneas in order to measure their radii of curvature. The formula used to approximate corneal radius of curvature is, r= y2/2S § S/2, where r=radius of curvature, 2y=any chord and S = sagitta of that chord. This formula assumes that the cornea is spherical within the limits of the chord (Fincham and Freeman 1974). The radius of curvature was converted to diopters using the formula, F = (n 1 _ n)/r where n is the index of refraction of air (1.0), r is the radius of curvature of the cornea and n 1 is the index of refraction of the cornea. A corneal value of 1.33 is used instead of
M.E. Andison et al.: Eye development of American kestrel 567
1.37 to compensate for the negative effect of the posterior surface of the cornea (Fincham and Freeman 1974).
Treated birds - Group L Once an initial refractive error could be determined, a group of 10 kestrels had goggles unilaterally applied to the down surrounding the right eye. The contralateral eye served as a control. The goggles, 0.40 mm thick, measured 17.0 mm in diameter and had a radius of curvature of 5.5 mm. Goggle clarity was degraded with 400-grade sandpaper to prevent sharp images from reaching the retina (Pickett 1986). This group of birds was treated in an identical manner to the control group, except for the application of the goggle.
Each day, birds were weighed prior to their first feeding. Retino- scopy and ultrasonography were performed each day on the control (untreated) eye. The external surface of the goggles was cleaned with water and tissues as needed.
At 6 weeks of age, on the day of sacrifice, goggles were removed, refractive measures were made and ultrasonography was performed on both treated and control eyes. Eight kestrels were used for freeze-sectioning to examine possible differences in intraocular di- mensions. Slides of the chosen frames were made and tracings were drawn of the projected corneas in order to measure their radii of curvature.
Treated birds - Group II. A group of 60 kestrels had goggles applied to the down surrounding one eye; the contralateral eye served as a control. Initial refractive errors were measured and ultrasound measures were made on the second day after hatching. A goggle was applied to the left eye of 10 birds and to the right eye of the remaining 50 birds in order to control for any possible bias of laterality. Fifty-five birds were sacrificed by decapitation in small groups at 7, 10, 14, 21 or 28 days of age. Birds from each of these age groups were assessed for refractive error and eye-size differences between treated and control eyes following goggle removal and before being sacrificed. Measurements from frozen sections were used to determine developmental changes for the treated and un- treated eyes over time. Slides of the appropriate frozen sections were
made and tracings were drawn of the projected corneas in order to measure their radii of curvature. Due to the fact that a limited number of such tracings were possible, the corneal curvature data for treated birds is small and sample sizes vary from only 1 to 7 for the various age groups.
The remaining 5 kestrels had their goggles removed at 30 days of age and served as a recovery group. Each day refractive error changes were assessed by retinoscopy. Ultrasound measures were made to determine changes in the treated or control eyes. These birds were sacrificed at 40 days of age by decapitation.
Goggle transmittance properties. The transmittance properties of the goggles were determined using a portable spectroradiometer (model LI-1800, LI-COR, Inc., Lincoln, Nebraska, USA). Wavelengths from 400 nm to 800 nm in 2 nm intervals were used. A tungsten 100 W light was used as the source illumination. Five goggles were tested and a composite plot of percent transmittance versus wavelength was produced.
R e s u l t s
B o d y w e i g h t d a t a
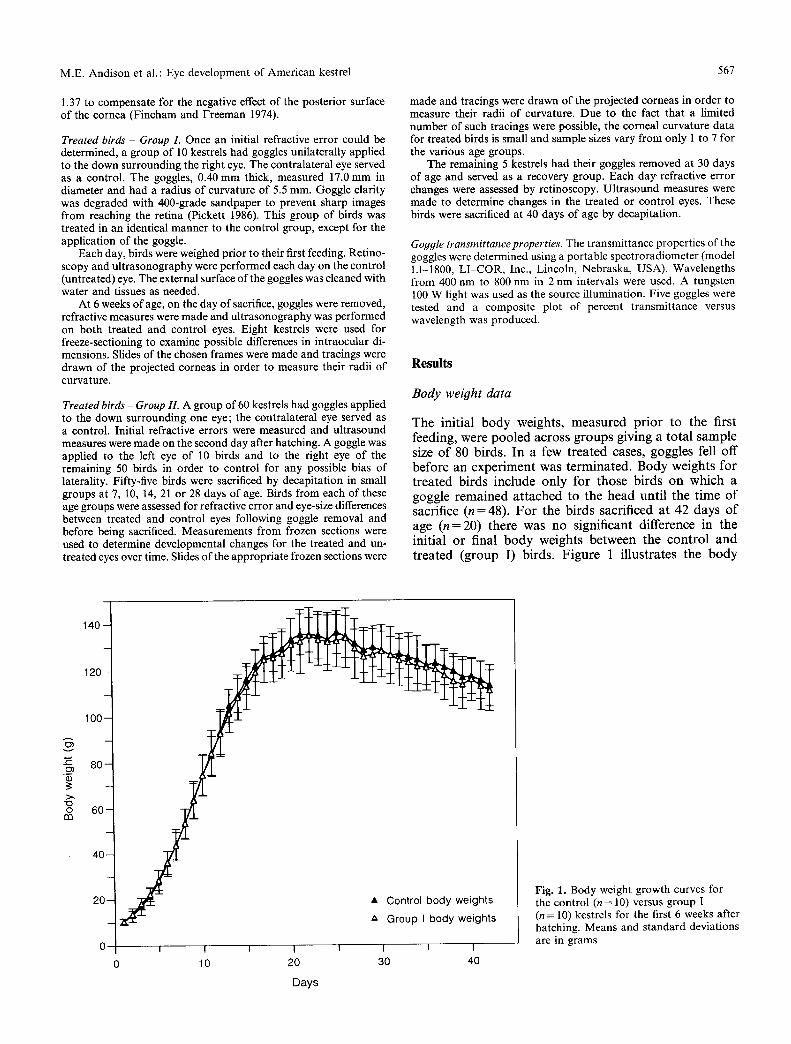
The initial b o d y weights , measu red p r io r to the first feeding, were poo led across g roups giving a to ta l sample size o f 80 birds. In a few t rea ted cases, goggles fell off before an exper iment was te rminated . Body weights for t rea ted birds include only for those birds on which a goggle r emained a t t ached to the head unt i l the t ime o f sacrifice (n = 48). F o r the birds sacrificed at 42 days o f age ( n = 20) there was no significant difference in the init ial or final b o d y weights be tween the con t ro l and t rea ted (group I) birds. F igure 1 i l lustrates the b o d y
_E C~ .m {33
"O O
m
140
I I 10
I
20
Days
�9 Control body weights
A Group I body weights
I I I
30 40
Fig. 1. Body weight growth curves for the control (n = 10) versus group I (n = 10) kestrels for the first 6 weeks after hatching. Means and standard deviations are in grams
568
10
M.E. Andison et al. : Eye development o f American kestrel
O . o
" 0
>
O
N rr
O -
-10 -
- 2 0 -
-30
0
�9 Control OD
z~ Control OS
t I t I ~ I t I
10 20 30 40
Group I OS
Days
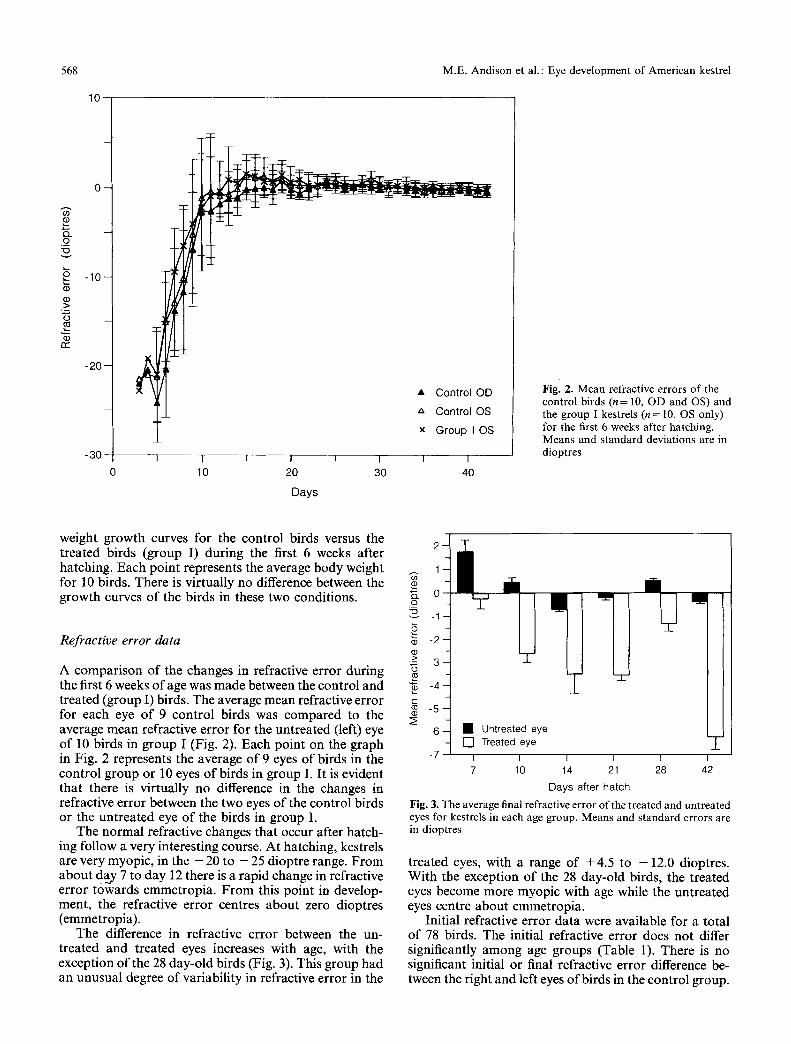
Fig. 2. Mean refractive errors o f the control birds (n = 10, OD and OS) and the group I kestrels (n = 10, OS only) for the first 6 weeks after hatching. Means and s tandard deviations are in dioptres
weight growth curves for the control birds versus the treated birds (group I) during the first 6 weeks after hatching. Each point represents the average body weight for 10 birds. There is virtually no difference between the growth curves of the birds in these two conditions.
Refractive error data
A comparison of the changes in refractive error during the first 6 weeks o f age was made between the control and treated (group I) birds. The average mean refractive error for each eye of 9 control birds was compared to the average mean refractive error for the untreated (left) eye of 10 birds in group I (Fig. 2). Each point on the graph in Fig. 2 represents the average of 9 eyes of birds in the control group or 10 eyes of birds in group I. It is evident that there is virtually no difference in the changes in refractive error between the two eyes of the control birds or the untreated eye of the birds in group I.
The normal refractive changes that occur after hatch- ing follow a very interesting course. At hatching, kestrels are very myopic, in the - 20 to - 25 dioptre range. From about day 7 to day 12 there is a rapid change in refractive error towards emmetropia. F rom this point in develop- ment, the refractive error centres about zero dioptres (emmetropia).
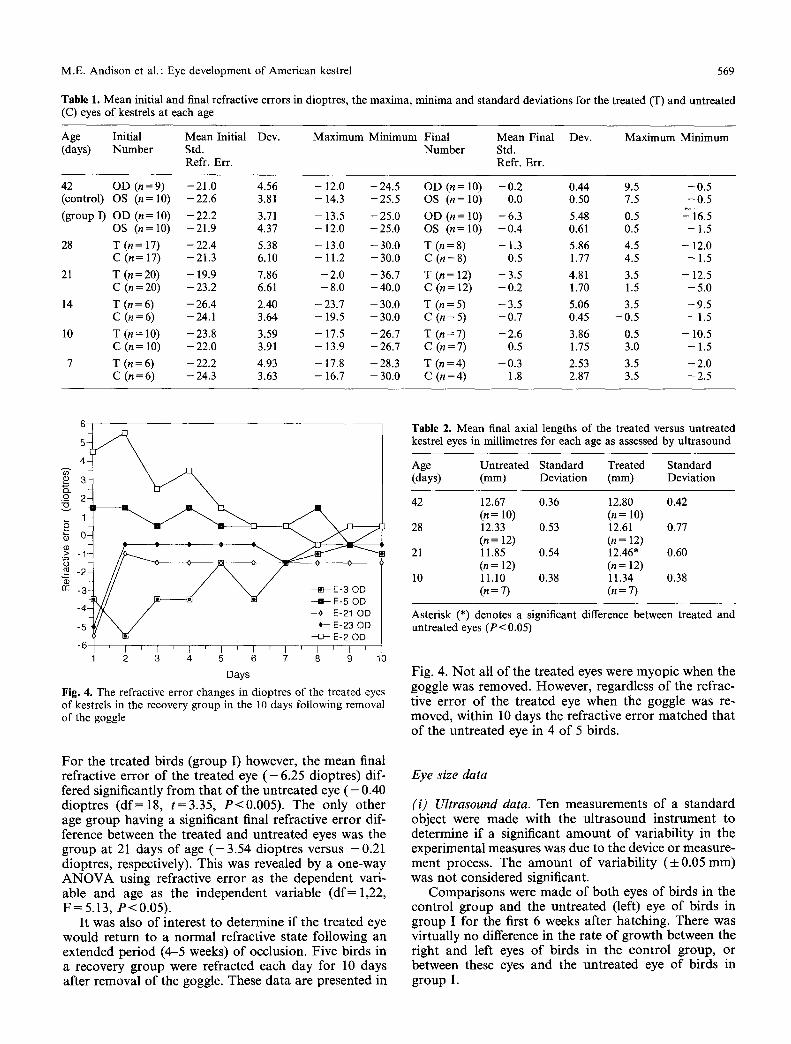
The difference in refractive error between the un- treated and treated eyes increases with age, with the exception of the 28 day-old birds (Fig. 3). This group had an unusual degree of variability in refractive error in the
2
~_ 0 O
b -2
"> -3 "6
c
-6
-7
�9 Untreated eye [ ] Treated eye "~
I I J I I I
7 10 14 21 28 42
Days after hatch
Fig. 3. The average final refractive error o f the treated and untreated eyes for kestrels in each age group. Means and s tandard errors are in dioptres
treated eyes, with a range of +4.5 to - 1 2 . 0 dioptres. With the exception of the 28 day-old birds, the treated eyes become more myopic with age while the untreated eyes centre about emmetropia.
Initial refractive error data were available for a total of 78 birds. The initial refractive error does not differ significantly among age groups (Table 1). There is no significant initial or final refractive error difference be- tween the right and left eyes of birds in the control group.
M.E. Andison et al. : Eye development of American kestrel 569
Table 1. Mean initial and final refractive errors in dioptres, the maxima, minima and standard deviations for the treated (T) and untreated (C) eyes of kestrels at each age
Age Initial Mean Initial Dev. Maximum Minimum Final Mean Final Dev. Maximum Minimum (days) Number Std. Number Std.
Refr. Err. Refr. Err.
42 OD (n=9) -21.0 4.56 - 12 .0 -24.5 OD (n = 10) -0 .2 0.44 9.5 -0 .5 (control) OS (n=10) -22.6 3.81 -14.3 -25.5 OS (n=10) 0.0 0.50 7.5 -0 .5
(group I) OD (n= 10) -22.2 3.71 - 13 .5 -25.0 OD (n= 10) -6 .3 5.48 0.5 -"i6.5 OS (n=10) -21.9 4.37 -12.0 -25.0 OS (n=10) - 0 . 4 0.61 0.5 -1 .5
28 T (n = 17) - 22.4 5.38 - 13.0 - 30.0 T (n = 8) - 1.3 5.86 4.5 - 12.0 C (n= 17) -21.3 6.10 -11.2 -30.0 C (n=8) 0.5 1.77 4.5 -1 .5
21 T (n=20) -19.9 7.86 -2 .0 -36.7 T (n=12) -3 .5 4.81 3.5 -12.5 C (n=20) -23.2 6.61 -8 .0 -40.0 C (n= 12) -0 .2 1.70 1.5 -5 .0
14 T (n=6) -26.4 2.40 -23.7 -30.0 T (n=5) -3 .5 5.06 3.5 -9 .5 C (n=6) -24.1 3.64 - 19 .5 -30.0 C (n=5) -0 .7 0.45 -0 .5 -1 .5
10 T (n= 10) -23.8 3.59 -17.5 -26.7 T (n=7) -2 .6 3.86 0.5 -10.5 C (n = 10) - 22.0 3.91 - 13.9 - 26.7 C (n = 7) 0.5 1.75 3.0 - 1.5
7 T (n=6) -22.2 4.93 - 17 .8 -28.3 T (n=4) -0 .3 2.53 3.5 -2 .0 C (n=6) -24.3 3.63 -16.7 -30.0 C (n=4) 1.8 2.87 3.5 -2 .5
g~
2 3 4 5 6 7 8 9 10
Days
Fig. 4. The refractive error changes in dioptres of the treated eyes of kestrels in the recovery group in the 10 days following removal of the goggle
F o r the t r ea t ed b i rds (g roup I) however , the m e a n final ref rac t ive e r ro r o f the t r ea t ed eye ( - 6.25 d iop t res ) dif- fered s ignif icant ly f rom tha t o f the u n t r e a t e d eye ( - 0 . 4 0 d iop t r e s ( d f = 1 8 , t = 3 . 3 5 , P < 0 , 0 0 5 ) . The on ly o the r age g r o u p hav ing a s ignif icant f inal ref rac t ive e r ro r dif- ference be tween the t r ea ted a n d u n t r e a t e d eyes was the g r o u p a t 21 days o f age ( - 3 . 5 4 d iop t r e s versus - 0 . 2 1 d iop t res , respect ively) . This was revea led by a one -way A N O V A using ref rac t ive e r ro r as the d e p e n d e n t var i - ab le and age as the i n d e p e n d e n t va r i ab le ( d f = 1,22, F = 5.13, P < 0.05).
I t was also o f in teres t to de t e rmine i f the t r ea ted eye w o u l d r e tu rn to a n o r m a l ref rac t ive s ta te fo l lowing an ex tended pe r iod (4 -5 weeks) o f occlus ion. F ive b i rds in a r ecovery g r o u p were re f rac ted each d a y for 10 days af te r r e m o v a l o f the goggle. These d a t a a re p resen ted in
Table 2. Mean final axial lengths of the treated versus untreated kestrel eyes in millimetres for each age as assessed by ultrasound
Age Untreated Standard Treated Standard (days) (mm) Deviation (mm) Deviation
42 12.67 0.36 12.80 0.42 (n = 10) (n = 10)
28 12.33 0.53 12.61 0.77 (n = 12) (n = 12)
21 11.85 0.54 12.46" 0.60 (n = 12) (n = 12)
10 11.10 0.38 11.34 0.38 (n = 7) (n = 7)
Asterisk (*) denotes a significant difference between treated and untreated eyes (P< 0.05)
Fig . 4. N o t all o f the t r ea ted eyes were m y o p i c when the goggle was removed . Howeve r , regard less o f the ref rac- t ive e r ro r o f the t r ea t ed eye when the goggle was re- moved , wi th in 10 days the ref rac t ive e r ro r m a t c h e d tha t o f the u n t r e a t e d eye in 4 o f 5 b i rds .
Eye size data
(i) Ultrasound data. Ten m e a s u r e m e n t s o f a s t a n d a r d objec t were m a d e wi th the u l t r a s o u n d i n s t rum en t to de te rmine i f a s ignif icant a m o u n t o f va r i ab i l i t y in the expe r imen ta l measures was due to the device o r measu re - m e n t process . The a m o u n t o f va r i ab i l i t y (4-0.05 m m ) was n o t cons ide red significant .
C o m p a r i s o n s were m a d e o f b o t h eyes o f b i rds in the c on t ro l g r o u p and the u n t r e a t e d (left) eye o f b i rds in g r o u p I for the first 6 weeks af te r ha tch ing . There was v i r tua l ly no difference in the ra te o f g r o w t h be tween the r ight and left eyes o f b i rds in the con t ro l g roup , or be tween these eyes a n d the u n t r e a t e d eye o f b i rds in g roup I.
570 M.E. Andison et al. : Eye development of American kestrel
Table 3. Average dimensions and standard deviations in millimetres of the treated and untreated eyes of birds at each age from frozen sections
UTdenotes untreated eye, Tdenotes treated eye, M denotes myopic eye. n refers to the number of eyes. * P < 0 . 0 5 , ** P < 0 . 0 1 , *** P < 0 . 0 0 5 , **** P < 0 . 0 0 0 5
Examinat ion of the ultrasound data revealed that for each age, the treated eye was longer axially than the untreated eye. A one-way A N O V A using axial length as the dependent variable and age as the independent vari- able revealed that this difference was only significant for birds at 21 days of age (df = 1,22, F=6 .88 , P<0 .05 ) (Table 2).
between the untreated eyes of the treated birds (group I) and both eyes of the control group on any of the dimen- sions. The treated eyes tended to be larger equatorially (d f= 14, t = - 3.35, P<0 .005) and had deeper vitreous chambers (d f=14 , t = - 2 . 7 6 , P<0 .05 ) than the un- treated eyes for the birds in group I. Since only 6 of the 8 treated eyes were myopic, averages and standard devia- tions of each dimension using only the myopic eyes were calculated and compared with the dimensions obtained for the untreated eyes of birds in group I (n = 8). The values obtained using the myopic eyes tended to give the largest values for the equatorial dimensions of the globe (d f= 12, t = - 5.02, P < 0.0005) and the vitreous chamber depth (d f= 12, t = - 2 . 96 , P < 0.01). For this reason, only frozen sections of birds that had manifested a myopic refractive error in the treated eye were used for each age.
The treated birds (group I) were the only group having a significant difference in the average equatorial diameter between the untreated and treated eyes (df = 12, t = - 5 . 02 , P<0.0005). For all of the other age groups, the treated eye tended to be larger equatorially, but the difference was not significant. The treated birds at 21 days of age were the only group showing a significant difference in the average axial diameter between the untreated and treated eyes (df = 12, t = - 2.38, P < 0.05). Again, the treated eyes for all ages tended to be larger than the untreated eyes with respect to axial diameter. There was no significant difference between the treated and untreated eyes of birds at any of the ages with respect to the average anterior chamber depth, and average axial or equatorial lens dimensions. There was a significant difference for birds at 42 days (df=12 , t = - 2 . 9 6 , P<0 .01 ) and 21 days (df=12 , t = - 5 . 2 3 , P<0.0005) with respect to the average vitreous chamber depth. The treated eyes for the birds at 14 days had a deeper average vitreous chamber depth than the untreated eyes, but the difference was not significant. There was no difference in the average vitreous chamber depth between treated and untreated eyes of birds at 10 and 28 days of age (Table 3).
The frozen sections versus ultrasound measurements were well fitted by a regression line (Y=4.01 +0.59X, df = 17, F = 26.86, P < 0.0001). These two measurements were positively correlated (R = 0.78). These results sup- port the expectation that the ultrasound measures would be smaller due to liquid expansion on freezing. Also, the ultrasound measures were based on an instrument calibrated for average sound transmission through the human eye. I f average sound transmission in the kestrel eye is higher, due to a relatively larger lens, such mea- sures may have underestimated axial length.
Corneal curvature data
(ii) Frozen sections data. Frozen sections were used f rom a total o f 32 birds. F r o m the frozen sections, com- parisons were made of the anterior and vitreous chamber depths, the axial and equatorial lens dimensions and the total axial length and equatorial diameter of the globe for the treated and untreated eyes of birds at each age at sacrifice (Table 3). There was no significant difference
The measurement accuracy of the radii of curvature was estimated to be + 0.05 mm. The average corneal radii of curvature in the untreated eyes for the birds at 42, 28, 21, and 14 days of age were not significantly different. The range of curvatures (3.3 m m to 3.5 mm) represented a range of corneal powers f rom 94.3 to 100 dioptres. The radii o f curvature of corneas for birds at 10 and 7 days
M.E. Andison et al. : Eye development of American kestrel 571
and at hatch were progressively shorter with decreasing age. The range of curvatures (3.1 mm to 2.1 mm) repre- sented a range of corneal powers from 103.1 to 157.1 dioptres in the untreated eyes. The radii of curvature were not significantly different in the untreated and treated eyes for birds aged 42, 28, 21, 14 and 10 days.
The small radius of curvature (2.1 mm) and associated corneal power (157.1 dioptres) of the hatchlings indicates that the cornea may be involved in the extreme myopia measured in the young kestrels. The radii of curvature are progressively longer with increasing age up to 14 days, which corresponds to the progressively less myopic refractive errors measured in the untreated eyes up to this age.
Goggle transmittance
The transmittance properties of 5 goggles were tested to ensure that the kestrels were all receiving the same qual- ity and quantity of light. There is no significant difference between the 5 measures. The curves are slightly non- monotonic, having a small decrease between 560 nm and 630 nm. The longer (red) wavelengths are transmitted more by almost 15% than those in the blue range. This is to be expected because the goggles are slightly bluish in colour. Light transmittance of the goggles varied from about 58 to 64% from 400 to 700 nm.
Discussion
This study is an attempt to determine if the chicken is a representative avian model for studying form-depriva- tion myopia. It uses an accepted technique with a dif- ferent species, the American kestrel. The untreated eye of a monocularly occluded kestrel appears not to be affected by the contralateral treated eye, a finding which is consistent with the results of form-deprivation experi- ments using chickens. In both species, the untreated eye does not differ from the eyes of control birds with respect to refractive error, intraocular dimensions, external di- mensions, weight or corneal curvature.
Using chickens, previous investigators have reported substantial myopic shifts in refractive error and large increases in the external dimensions and weight of the treated eye following visual field restriction (Wallman et al 1978) and form deprivation (Pickett 1986). The sever- ity of the myopia produced in the kestrels was, on av- erage, relatively minor even after prolonged periods of form deprivation. Thus, the kestrel data revealed that the dimensions of the treated eyes were only marginally larger than those of the untreated eyes.
From hatching to about 12 days of age, the eyes of all young kestrels approach emmetropia from an average refractive error of approximately - 22 dioptres. The av- erage refractive state of the untreated eyes of kestrels aged 14 days to 6 weeks of age is emmetropia.
The dioptric power of the cornea was inversely proportional to age. The corneas having the shortest radii of curvature were from eyes which manifested the
largest myopic refractive errors. Thus, the hatchling kestrels had the shortest corneal radii of curvature. The radii of curvature of the corneas from progressively older kestrels increased up to 14 days, whereupon they did not differ significantly to 6 weeks of age. Thus, the corneal results support the refractive error measurements which demonstrate extreme myopia in the very young chicks and emmetropia from about day 12 to 6 weeks of age in the untreated eyes. However, the small sample sizes must be kept in mind in interpreting this data. Troilo (1990) reported that dark-rearing produced both hyperopia and ocular enlargement in chickens, but attributed the hyperopia to an increase is corneal radius of curvature.
The fact that such a high degree of variability was found in the refractive errors of the treated eyes was surprising. The goggle was applied before emmetropia had been reached and the refractive error was still one of severe myopia. The refractive errors of the treated eyes from which goggles were removed before emmetropia should have been reached (7 and 10 day-olds) were closer to emmetropia than when they were occluded. This was not anticipated since the goggle was expected to cause a myopic shift in refractive error. The treated eyes which could have emmetropized (14, 21, 28 and 42 days-olds) manifested progressively larger myopic shifts in refrac- tive error /f the initial refractive error was emmetropia, with the exception of the group at 28 days of age. How- ever, the initial refractive error was, on average, - 2 2 dioptres, which means that these eyes were manifesting progressively smaller myopic shifts in refractive error. Since only the initial and final refractive errors of the birds sacrificed at each of the 5 youngest ages were measured, it is assumed that their refractive errors follow the same developmental changes as those of the 20 birds refracted each day for 6 weeks.
It is possible that short periods of occlusion do not interfere with the normal emmetropization process. The average refractive error in the treated eyes of birds at 10 days of age (-2.61 + 3.86 dioptres, S.D.) matches the average refractive error of the untreated eyes of the control and treated groups' refractive errors for this age. Therefore, the eyes might be emmetropizing to a certain extent despite the goggle. In fact, the extreme myopia in the early period may mean that the occluders make little difference in vision. By two weeks of age, the effect of blurred retinal images may be affecting the refractive development of the eye. When compared to average refractive errors of the eyes of the control group and the untreated eyes of group I, the relative amount of myopia from 14 days onwards is increasing (with the exception of the 28 day-old birds.) Among all of the age groups, there is the most variability in the refractive errors in the treated eyes of the 28 day-old birds. This group also had the second largest range of refractive errors of any of the treated eyes. There may be too much variability in the refractive errors up to the first 10 days after hatching to draw any real conclusions about what is happening during this period.
If the distribution of the refractive errors of the un- treated eyes is considered, there were 3-4 birds showing hyperopia in the untreated eye at 7 days of age. At 10 and
572 M.E. Andison et al.: Eye development of American kestrel
28 days of age, there was one bird in each group showing hyperopia (3.0 and 4.5 dioptres), respectively. When the refractive errors of the birds in the control group and group I are examined at 7, 10 and 28 days of age, the variability in these data are sufficient to account for the hyperopic refractive errors measured at the younger ages. For this reason, it is assumed that the slightly hyperopic average refractive errors in the untreated eyes of the groups at 7 and 10 days are within the expected range of refractive errors. The use of larger sample sizes would decrease the variability measured in these groups. Sam- piing the birds at 4, 6, 8 and 10 days of age may give a better indication of what is happening to the treated eye as the other eye is emmetropizing. It may be more valu- able to concentrate on the period between 10 and 42 days after hatching.
Emmetropization and recovery
Experiments in which the development of refractive er- rors result from manipulations of visual experience sup- port the claim that visual regulation of eye growth oc- curs. In addition, near work in humans (Curtin 1985) and experimental restriction of the visual environment in monkeys (Young 1963) have both been positively cor- related with myopia.
Wallman and Adams (1987) have suggested that emmetropization is a vision-dependent phenomenon. In chickens, if the visual field is restricted during the period of rapid eye growth, high levels of myopia are produced. A return to a normal refractive state follows the restora- tion of clear vision during this time. Up to 4 weeks of age, the refractive state of the kestrel eye is "plastic" enough to recover from induced refractive errors. It seems that when the kestrel retina is presented with a blurred image, the eye does not "know" in which direction to find em- metropia because the refractive errors are both hyperopic and myopic after 3 or 4 weeks of occlusion. It is interest- ing to note that the eyes were capable of reaching em- metropia from either direction within 10 days.
In young kestrels, the earliest refractive errors mea- sured are extremely myopic. For the first two days after hatching their eyes are partially closed. Therefore, no clear images are reaching the retina during this time. Over the next few days the young kestrels keep their eyes open for more extended periods of time. This factor might be involved in the process of emmetropization. The eyes become more emmetropic with more prolonged periods of clear form vision. Troilo (1990) reported that in chickens, emmetropization is achieved by changes in the vitreous chamber depth. Myopic and hyperopic eyes show decreased and increased growth rates, respectively, in the vitreous chamber depth when recovering from induced refractive errors.
Models of emmetropization are based on research using chickens. The present study has indicated a number of differences between chickens and kestrels in their re- fractive development and response to occlusion. Just prior to hatching, chickens are extremely hyperopic. A pilot study carried out prior to the present study exam-
ined the normal refractive errors of chickens just before hatching and at hatching. At 21 days of incubation, shortly before hatching, the average refractive error was + 15.5 + 4.70 dioptres (S.D.). At hatching, the refractive error averaged +11.3:t:3.08 dioptres (S.D.). In a separate study, Pickett (1986) found the refractive errors of chickens to average + 2.7 + 0.2 dioptres (S.E.) within one day after hatching. Within the first 36 hours after hatching, their refractive errors decreased rapidly and within 7 days they were within + 1.0 dioptres of em- metropia (Pickett 1986). The kestrels have an average myopic refractive error at hatching of -22.2 4- 5.5 diop- tres (S.D.). They emmetropize within 12 days after hatch- ing. This rate of emmetropization occurs at a slower rate than that of the chickens.
After an extended period (42 days) of occlusion, the average amount of myopia produced in the kestrel eyes ( - 6.25 4- 5.48 dioptres, S.D.) was substantially less than that measured in the chickens. The trend towards an increasingly more myopic refractive state with longer periods of occlusion agrees with studies of chickens (Selt- ner et al. 1988). The magnitude of eye growth in the treated versus the untreated eyes of the kestrels was also significantly smaller than that found in chickens (0.5 mm versus 1.5 mm at 14 days), but again the direction of the change was the same. A longer period of occlusion was accompanied by an increase in the axial difference be- tween the treated and untreated eyes. The form-depriva- tion experiments of Seltner et al. (1988) using chickens showed that within 14 days, there was a substantial myopic shift in refractive error, averaging - 10.7 + 3.12 dioptres (S.D.). These changes in refractive error were accompanied by a 19% increase in the axial length and a 21% increase in the wet weight of the treated eyes when compared to the contralateral controls (Seltner et al. 1988).
A separate experiment examined the development of experimentally induced myopia in the Japanese quail, Coturnix coturnix japonica (Andison, unpublished re- suits). The quail became significantly myopic in the treated eye within 7 days after hatching when they were monocularly occluded. The quail are precocial hatch- lings, like the chickens. These data agree with the chicken results. The findings with the quail are also valuable because they provide a control for the methodology used with the kestrels. The goggles used were identical to those applied to the kestrels; the refractive technique was also identical. The principal difference between the quail and kestrels is that the quail are precocial and the kestrels are semi-altricial. It was not possible to study quail em- metropization in more detail, but the results of this initial study do indicate that they are hyperopic when they hatch, like the chickens.
Is the chicken a representative bird model?
The results found with the quail corroborated previous findings with chickens. Both of these species are preco- cial. When the same technique was used with the semi- altricial kestrel, not only were the expected refractive
M.E. Andison et al. : Eye development of American kestrel 573
error changes and ocular enlargement less spectacular, but the normal pat tern of emmetropizat ion also differed f rom that of chickens. This may be an indication that precocial and semi-altricial birds, as classes of birds, differ with respect to their processes of emmetropizat ion. Inter-genus differences in the response to form depriva- tion have been suggested (Raviola and Wiesel 1990). These authors reported that even within the same genus of monkeys, two different mechanisms of axial elonga- tion may be in effect.
The initial refractive states and speed with which emmetropia is reached are likely a reflection of the needs of each species at hatching. The hatchling kestrels prob- ably have no requirement for clear form vision because the adults feed the young and care for them in the nest until fledging age (between 3 and 4 weeks after hatching). However, within the first couple of weeks, the chicks are strong enough to tear their own food, so the adults no longer do this for them. It is within the first two weeks after hatching that the young kestrels are the most vul- nerable and rely the most heavily on the adults, and it is during this time they are emmetropizing f rom a high degree of myopia. I t is possible that the myopia of nest- bound chicks may be of some functional value in feeding. However, it seems that at hatching, the kestrels are less completely developed than the quail or chickens. No t only is their visual development at a more immature stage, they also lack the necessary muscle tone and, hence, the mobility to seek their own food (Welty 1982). Chickens and quail are self-sufficient shortly after they hatch. The quail were hatched in large, commercial in- cubators and then moved to pens which provided only food, water and warmth. Hatchery-raised chickens are raised in much the same way. These two species have a more immediate need for clear vision than the kestrels.
A more thorough study of the refractive development of a number of altricial or semi-altricial and precocial species is necessary to make any generalizations. How- ever, it is reasonable to speculate that the normal refrac- tive development observed in such species as kestrels and chickens reflects the time at which each of these species requires clear vision. Gibson and Walk (1960) have shown, in a number of young mammals , that depth perception is correlated with the time at which they are capable of locomotion.
The present study represents the first indication that chickens may not be a representative bird model for studying form-deprivat ion myopia. N o t only do kestrels and chickens differ with respect to the results obtained using an identical form-deprivat ion technique and meth- od of assessment, their normal emmetropizat ion patterns are diametrically opposed. These differences may be due to the extent to which each species is developed at the time of hatching. Possibly the accommodat ive appara tus of the very young kestrel is not fully functional. Schaeffel et al. (1986) reported that, at hatching, the chicken is capable of an adult level o f accommodat ion (17 diop- tres). They relate this ability to its precocity. Rouse and Ryan (1990) have indicated that the accommodat ive re- sponse of the human infant is not equivalent to adult capabilities for the first 3 or 4 months after birth. Weale
(1982) suggests that the myopia measured in premature infants is related to the incomplete development of the eye at the time of birth (in Goss 1985). Nissenkorn et al. (1983) have positively correlated prematur i ty and myo- pia in pre-term human infants. Is it conceivable that hatchling kestrels are just "premature" birds at an in- complete level o f ocular development?
G i b s o n E J, Walk RD (1960) The "visual cliff". Sci Amer 202: 64-71 Goss DA (1985) Refractive status and premature birth. Optometric
Monthly 76:109-111 Hodos W (1990) Avian models of experimental myopia: environ-
mental factors in the regulation of eye growth. In: Myopia and the control of eye growth. Ciba Foundation Symposium 155. John Wiley & Sons Ltd., pp 149-156
Hubel DH, Wiesel TN, Le Vay S (1975) Functional architecture of area 17 in normal and monocularly deprived macaque monkeys. Cold Spring Harbor Symp Quant Biol 40:581-589
Irving EL, Callender MG, Sivak JG (1991) Inducing myopia, hyperopia and astigmatism in chicks. Optom Vision Sci 68 : 364-368
Nissenkorn I, Yassur Y, Mashkowski D, Sherf I, Ben-Sira I (1983) Myopia in premature babies with and without retinopathy of prematurity. Br J Ophthalmol 67:170-173
Pickett RL (1986) Experimentally induced myopia in chicks: characterization of the physical and biochemical changes occur- ring in the first two weeks post-hatching. M Sc thesis, University of Waterloo, Ontario
Pickett-Seltner RL, Weerheim JA, Sivak JG, Pasternak JJ (1987) Experimentally induced myopia does not affect post-hatching development of the chick lens. Vision Res 27:179-1782
Raviola E, Wiesel TN (1990) Neural control of eye growth and experimental myopia in primates. In: Myopia and the control of eye growth. Ciba Foundation Symposium 155. John Wiley & Sons Ltd., pp 22-39
Rouse MW, Ryan JM (1990) The optometric examination and management of children. In: Rosenbloom AA, Morgan MW (ed) principals and practice of pediatric optometry. JB Lippin- cott Company, Philadelphia, pp 155-191
Schaeffel F, Glasser A, Howland HC (1988) Accommodation, re- fractive error and eye growth in chickens. Vision Res 28 : 639-657
Schaeffel F, Howland HC, Farkas L (1986) Natural accommoda- tion in the growing chicken. Vision Res 26:197%1993
Seltner RL, Sivak JG, Pasternak JJ (1988) Experimentally induced myopia in chicks: morphometric and biochemical analysis during the first 14 days after hatching. Vision Res 28:323-328
Sherman SM, Norton TT, Casagrande VA (1977) Myopia in the lid-sutured tree shrew (Tupaia 9lis). Brain Res 124:154-157
Shlaer R (1972) An eagle's eye: quality of the retinal image. Science 176: 920-922
Sivak JG, Barrie DL, CaUender MG, Doughty M J, Seltner RL, West JA (1990) Optical causes of experimental myopia. In:
574 M.E. Andison et al. : Eye development of American kestrel
Myopia and the control of eye growth. Ciba Foundation Sym- posium 155. John Wiley & Sons Ltd., pp 160-172
Sivak JG, Barrie DL, Weerheim JA (1989) Bilateral experimental myopia in chicks. Optom Vision Sci 66:854-858
Troilo D (1990) Experimental studies of emmetropization in the chick. In: Myopia and the control of eye growth. Ciba Founda- tion Symposium 155. John Wiley & Sons Ltd., pp 89-102
Wallman J, Adams JI (1987) Developmental aspects of experi- mental myopia in chicks: susceptibility, recovery and relation to emmetropization. Vision Res 27:1139-1163
Wallman J, Turkel J, Trachtman J (1978) Extreme myopia produced by modest change in early visual experience. Science 201 : 124~1251
Walls GL (1942) The vertebrate eye and its adaptive radiation. Hafner Publishing Company, New York London
Weale RA (1982) A biography of the eye development, growth and age. HK Lewis and Co
Welty JC (1982) The life of birds. Third Edition. Saunders College Publishing, NY
Wiesel TN, Raviola E (1977) Myopia and eye enlargement after neonatal lid fusion in monkeys. Nature 266:66
Wiesel TN, Raviola E (1979) Increase in axial length of the macaque monkey eye after corneal opacification. Invest Ophthalmol Vis Sci 18:1232-1236
Wilson JR, Sherman SM (1977) Differential effects of early monoc- ular deprivation on the binocular and monocular segments of cat striate cortex. J Neurophysiol 40 : 891
Young FA (1963) The effect of restricted visual space on the refrac- tive error of the young monkey eye. Invest Ophthalmol 2:571-577