The role of nitric oxide on visual-evoked potentials in MPTP-induced Parkinsonism in mice Sinem Aras a , Gamze Tanriover b , Mutay Aslan c , Piraye Yargicoglu d , Aysel Agar a,⇑ a Akdeniz University, Medical School, Department of Physiology, Antalya, Turkey b Akdeniz University, Medical School, Department of Histology and Embryology, Antalya, Turkey c Akdeniz University, Medical School, Department of Biochemistry, Antalya, Turkey d Akdeniz University, Medical School, Department of Biophysics, Antalya, Turkey article info Article history: Received 16 December 2013 Received in revised form 10 April 2014 Accepted 21 April 2014 Available online 29 April 2014 Keywords: Parkinson MPTP iNOS nNOS VEPs abstract The present study aimed to elucidate visual evoked potentials (VEP) changes in MPTP induced Parkin- son’s disease (PD) and investigate the possible benefical effects of neuronal (n) and inducible (i) nitric oxide synthase (NOS) inhibitors on altered VEPs, lipid peroxidation and apoptosis. 3 months old C57BL/6 mice were randomly divided into 6 groups which included control (C), 7-nitra indazole treated (7-NI), S-methylisothiourea (SMT) treated, 1,2,3,6-tetrahydropyridine (MPTP) treated, 7-NI + MPTP trea- ted and SMT + MPTP treated. Motor activity of mice was evaluated via the pole test. At the end of the experimental period VEPs were recorded, brain and retina tissues were removed for biochemical analysis. Dopaminergic neuron death at substantia nigra (SN) was determined by immunohistochemical analysis of tyrosine hydroxylase (TH). Immunohistochemical staining was also performed to determine iNOS and nNOS in all tissue sections. Mice with experimental PD exhibited decreased motor activity. Dopaminergic cell death at pars compacta of SN (SNpc) was significantly increased in MPTP treated group compared to control. Diminished Parkinsonism symptoms were observed in 7-NI + MPTP and SMT + MPTP groups. Treatment with 7-NI and SMT decreased dopaminergic cell death in MPTP treated mice. Caspase-3 activity, nitrite/nitrate and 4-hydroxynonenal (4-HNE) levels were significantly increased in SN of MPTP treated mice compared to control. Treatment with 7-NI and SMT significantly decreased elevated cas- pase-3 activity, nitrite/nitrate and 4-HNE levels in SN of MPTP treated mice. No significant difference in above parameters were observed in the retina. VEP latencies were significantly prolonged in MPTP group compared to control group. 7-NI and SMT treatment caused a significant decrease in VEP latencies in MPTP treated mice compared to none treated MPTP group. This data shows that 7-NI and SMT improves prolonged VEP latencies. The protective effects of 7-NI and SMT on VEP alterations can be related to decreased dopaminergic cell death and reduced lipid peroxidation. Ó 2014 Elsevier Ltd. All rights reserved. 1. Introduction Parkinson’s disease (PD), first described by Dr. James Parkinson in 1817, is a progressive neurodegenerative disorder of the nervous system (Kidd, 2000). The motor symptoms of PD result from the death of dopaminergic cells in the pars compacta of the substantia nigra (SNpc) Kidd, 2000; Baraboi, 1989; Kishimoto et al., 1996. The subsequent decrease in dopamine (DA) in this area results in stri- atal dopamine deficiency. The cardinal symptoms of PD are muscle rigidity, tremor at rest, and akinesia. The symptoms first appear following 80% loss of dopaminergic cells (Kidd, 2000). Normally DA secreted in SN plays a role in the stimulation and coordination of motor functions through its effect on nucleus caudatus and nucleus putamene (Kidd, 2000). As a result of the loss of dopami- nergic neurons in SN and subsequent dopamine deficiency, the affected motor functions produce motor symptoms (Kidd, 2000). The cause of this cell death is unknown, but it is within the scope of our knowledge that free radicals and lipid peroxidation increase in PD (Baraboi, 1989). Among various factors that increase lipid peroxidation, nitric oxide (NO) is of critical importance (Kishimoto et al., 1996; Krukoff, 1999). Nitric oxide is a free radical that produces various nitrogenous substances such as nitrite and nitrate in the presence of oxygen (Gutteridge, 1994; O’Donnell et al., 1997). Formation of peroxynitrite (ONOO-) in vivo has been ascribed to the reaction of http://dx.doi.org/10.1016/j.neuint.2014.04.014 0197-0186/Ó 2014 Elsevier Ltd. All rights reserved. ⇑ Corresponding author. Address: Department of Physiology, Faculty of Medicine Akdeniz University, Arapsuyu, 07070 Antalya, Turkey. Tel.: +90 242 249 6958; fax: +90 242 227 4483. E-mail address: [email protected](A. Agar). Neurochemistry International 72 (2014) 48–57 Contents lists available at ScienceDirect Neurochemistry International journal homepage: www.elsevier.com/locate/nci
Transcript
Neurochemistry International 72 (2014) 48–57
Contents lists available at ScienceDirect
Neurochemistry International
journal homepage: www.elsevier .com/locate /nc i
The role of nitric oxide on visual-evoked potentials in MPTP-inducedParkinsonism in mice
http://dx.doi.org/10.1016/j.neuint.2014.04.0140197-0186/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author. Address: Department of Physiology, Faculty of MedicineAkdeniz University, Arapsuyu, 07070 Antalya, Turkey. Tel.: +90 242 249 6958; fax:+90 242 227 4483.
Sinem Aras a, Gamze Tanriover b, Mutay Aslan c, Piraye Yargicoglu d, Aysel Agar a,⇑a Akdeniz University, Medical School, Department of Physiology, Antalya, Turkeyb Akdeniz University, Medical School, Department of Histology and Embryology, Antalya, Turkeyc Akdeniz University, Medical School, Department of Biochemistry, Antalya, Turkeyd Akdeniz University, Medical School, Department of Biophysics, Antalya, Turkey
a r t i c l e i n f o a b s t r a c t
Article history:Received 16 December 2013Received in revised form 10 April 2014Accepted 21 April 2014Available online 29 April 2014
Keywords:ParkinsonMPTPiNOSnNOSVEPs
The present study aimed to elucidate visual evoked potentials (VEP) changes in MPTP induced Parkin-son’s disease (PD) and investigate the possible benefical effects of neuronal (n) and inducible (i) nitricoxide synthase (NOS) inhibitors on altered VEPs, lipid peroxidation and apoptosis. 3 months oldC57BL/6 mice were randomly divided into 6 groups which included control (C), 7-nitra indazole treated(7-NI), S-methylisothiourea (SMT) treated, 1,2,3,6-tetrahydropyridine (MPTP) treated, 7-NI + MPTP trea-ted and SMT + MPTP treated. Motor activity of mice was evaluated via the pole test. At the end of theexperimental period VEPs were recorded, brain and retina tissues were removed for biochemical analysis.Dopaminergic neuron death at substantia nigra (SN) was determined by immunohistochemical analysisof tyrosine hydroxylase (TH). Immunohistochemical staining was also performed to determine iNOS andnNOS in all tissue sections. Mice with experimental PD exhibited decreased motor activity. Dopaminergiccell death at pars compacta of SN (SNpc) was significantly increased in MPTP treated group compared tocontrol. Diminished Parkinsonism symptoms were observed in 7-NI + MPTP and SMT + MPTP groups.Treatment with 7-NI and SMT decreased dopaminergic cell death in MPTP treated mice. Caspase-3activity, nitrite/nitrate and 4-hydroxynonenal (4-HNE) levels were significantly increased in SN of MPTPtreated mice compared to control. Treatment with 7-NI and SMT significantly decreased elevated cas-pase-3 activity, nitrite/nitrate and 4-HNE levels in SN of MPTP treated mice. No significant differencein above parameters were observed in the retina. VEP latencies were significantly prolonged in MPTPgroup compared to control group. 7-NI and SMT treatment caused a significant decrease in VEP latenciesin MPTP treated mice compared to none treated MPTP group. This data shows that 7-NI and SMTimproves prolonged VEP latencies. The protective effects of 7-NI and SMT on VEP alterations can berelated to decreased dopaminergic cell death and reduced lipid peroxidation.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Parkinson’s disease (PD), first described by Dr. James Parkinsonin 1817, is a progressive neurodegenerative disorder of the nervoussystem (Kidd, 2000). The motor symptoms of PD result from thedeath of dopaminergic cells in the pars compacta of the substantianigra (SNpc) Kidd, 2000; Baraboi, 1989; Kishimoto et al., 1996. Thesubsequent decrease in dopamine (DA) in this area results in stri-atal dopamine deficiency. The cardinal symptoms of PD are musclerigidity, tremor at rest, and akinesia. The symptoms first appear
following 80% loss of dopaminergic cells (Kidd, 2000). NormallyDA secreted in SN plays a role in the stimulation and coordinationof motor functions through its effect on nucleus caudatus andnucleus putamene (Kidd, 2000). As a result of the loss of dopami-nergic neurons in SN and subsequent dopamine deficiency, theaffected motor functions produce motor symptoms (Kidd, 2000).The cause of this cell death is unknown, but it is within the scopeof our knowledge that free radicals and lipid peroxidation increasein PD (Baraboi, 1989).
Among various factors that increase lipid peroxidation, nitricoxide (NO) is of critical importance (Kishimoto et al., 1996;Krukoff, 1999). Nitric oxide is a free radical that produces variousnitrogenous substances such as nitrite and nitrate in the presenceof oxygen (Gutteridge, 1994; O’Donnell et al., 1997). Formation ofperoxynitrite (ONOO-) in vivo has been ascribed to the reaction of
S. Aras et al. / Neurochemistry International 72 (2014) 48–57 49
the free oxygen radicals formed by auto-oxidation with the freeradical nitric oxide (Hesslinger et al., 2009; Tieu et al., 2003).Peroxynitrite is a powerful oxidant; because of its oxidizingproperties, peroxynitrite can damage a wide array of moleculesin the body’s cells (Hesslinger et al., 2009; Tieu et al., 2003). Theproduction of nitric oxide in mammals is catalyzed by threeenzyme isoforms (Sener et al., 2001). These three enzymes are neu-ronal nitric oxide synthase (nNOS-NOS I) in the neurons, induciblenitric oxide synthase (iNOS-NOS II), and endothelial induciblenitric oxide synthase (eNOS-NOS III) in the endotelium (Tieuet al., 2003). Experimental animal models of PD induced by neuro-toxic substances 1-methyl 4-phenyl 1,2,3,6-tetrahydropridine(MPTP) and 6-hydroxydopamine (6-OHDA) have revealed anincrease in the expression and activity of iNOS and nNOS enzymes(Langheinrich et al., 2000). The mechanisms of these neurotoxinshave been related to the pathogenesis of nigral cell death in PD(Langheinrich et al., 2000). In studies where inhibitors of iNOS venNOS are used, neurodegeneration has been found to be less(Langheinrich et al., 2000).
The neurotransmitters DA and NO are amply present in thevisual pathway and central nervous system (Sener et al., 2001;Langheinrich et al., 2000). PD also affects the visual system. Nitricoxide, found in many areas of the body, is also produced in thevisual system. The enzymes nNOS and iNOS are also localized inthe retina. The NO in the retina contributes to the regulation ofbasal retinal blood flow. Dopamine is affected by NO secretion.Nitric oxide directly reduces DA through oxidation (Djamgozet al., 1995; Neal et al., 1999). Parallel to these findings, the inhibi-tion of NOS increases the basal DA level in the retina, independentof the cGMP pathway (Bugnon et al., 1994).
Considering that the measurement of visual evoked potentials(VEP) is a reliable method of functional evaluation of the visualsystem (Bodis-Wollner and Yahr, 1978; Celesia, 1984), the changein VEP latencies noted in our former experimental Parkinsonmodels is the indispensable proof of the effect of PD on the visualsystem. Moreover, our former investigations have elucidated theeffect of lipid peroxidation on VEP latencies (Kucukatay et al.,2006).
Studies have proved that NO, iNOS and nNOS together playimportant roles in neuronal death in SN. In view of these data, thisstudy was planned to assess, with VEP recordings, the roles of iNOSand nNOS in the mechanism of changes in the visual system ofexperimental Parkinson’s disease models where specific enzymeinhibitors were used.
2. Materials and methods
Animal maintenance and treatment were carried out in accor-dance with the the Institutional Animal Care and Use Committeeat Akdeniz University Medical School. Male C57BL/6 mice(3 months old, 25–30 g) were obtained from Akdeniz UniversityAnimal Care Unit. Animals were housed at an ambient temperatureof 22 ± 1 �C, a 12 h light/dark cycle and were fed standart micechow and tap water ad libitum up to day of the experiments.
2.1. Drug administration
Animals were assigned to six groups: (1) control group (n = 30;intraperitoneally peanut oil injected group), (2) MPTP group(n = 30; intraperitoneally MPTP (Sigma-M 0896) injected group),(3) MPTP + 7-NI (Sigma-N778) group (n = 30; intraperitoneallyspecific nNOS inhibitor (50 mg/kg) plus after 1 h MPTP receivedgroup), (4) MPTP + SMT group (n = 30; intraperitoneally specificiNOS inhibitor plus after 15 min MPTP received group), (5) 7-NIgroup (n = 30; intraperitoneally specific nNOS inhibitor treated
group) and (6) SMT group (n = 30; intraperitoneally specific iNOSinhibitor treated group). SMT dissolved in normal saline wasadministered 12 h intervals for 2 days consecutively (Iravaniet al., 2002). 7-NI dissolved in peanut oil (Di Matteo et al., 2009)was administered 12 h intervals for 2 days consecutively. MPTPat 20 mg/kg dissolved in 3 mg/ml saline was injected 12 h intervalsfor 2 days (Yokoyama et al., 2011).
3. Measurement of motor activity
3.1. Pole test
The pole test has been utilized to measure motor coordinationand balance in mouse models of PD (Ogawa, 1997). We performedthe pole test on the 7th day after the last MPTP injection. In thistest, animals were placed on top of a rough-surfaced iron pole(50 cm in length and 0.8 cm in diameter) and allowed to climbdown to the base of the pole. Sticking plaster was wrapped aroundthe iron pole to increase traction. Mice were placed at the top ofthe pole facing downward and latency to climb down the polewas measured. Trials were eliminated if the mouse jumped or sliddown the pole rather than climbed down. On testing day, animalswere placed head-up on the top of the pole. The time it took for theanimal to turn its head downwards (movement initiation) andclimb down the entire length of the pole was taken. Average ofthe best three measurements were taken as results.
3.2. VEPs recordings
On the 7th day of MPTP and drug treatment, VEPs wererecorded in a darkened room. The mice were anesthetized with acombination of ketamine (80 mg/kg, ip) and xylazine (16 mg/kg,ip) diluted in saline. For longer recording session, supplementalanesthesia (20% of initial dose) was given when required(Peachey and Ball, 2003). Recordings were collected with stainlesssteel sub-dermal electrodes (Nihon Kohden NE-223S, Tokyo Japan)shortened to 7 mm in length, resharpened and inserted along thelongitudinal axis of the mouse. Electrode placements were Fpz(midline, just distal to the interorbital line; negative electrode)and Oz (midline, nuchal crest; positive electrode), with a groundelectrode placed on the tail of the mouse (Strain and Tedford,1993). After 5 min of dark adaptation, a photic stimulator (BiopacSystem; Nova-Strobe AB, Santa Barbara, California, USA) at thelowest intensity setting was used to provide the flash stimulus ata distance of 10 cm, which allowed the lighting of the entire pupillafrom the temporal visual field. The repetition rate of the flashstimulus was 0.1 J. Throughout the experiments, the eye not underinvestigation was occluded by black carbon paper and cotton,allowing VEP recordings for both right and left eyes to be obtained.Meanwhile, the body temperature was maintained at 37.5–38 �Cby a heating pad. The averaging of 100 responses was accom-plished with the averager in the Biopac MP100 data acquisitionequipment. Analysis time was 300 ms. The frequency bandwidthof the amplifier was 1–100 Hz. The gain was selected as 20–50 lV/div. The microprocessor was programmed to reject anysweeps contaminated with larger artifacts, and at least twoaverages were obtained to ensure response reproducibility. Peaklatencies of the components were measured from the stimulusartifact to the peak in millisecond. Amplitudes were measured asthe voltage between successive peaks.
3.3. Tissue collection
After motor performance test, at the end of the treatment per-iod, mice were anesthetized with a combination of ketamine
50 S. Aras et al. / Neurochemistry International 72 (2014) 48–57
(80 mg/kg, ip) and xylazine (16 mg/kg, ip) diluted in saline. Micewere perfused transcardially with heparinized saline and theirbrain and retina tissues were removed immediately and stored at�80 �C for later biochemical analysis (Smeyne and Smeyne,2002). For immunohistochemical studies, brain tissues containingSN were fixed in 10% formaldehyde for 24 h and washed withtap water for approximately 6 h.
3.4. Immunohistochemistry
For tyrosine hydroxylase (TH), iNOS and nNOS immunohisto-chemistry, paraffin sections were deparaffinized in xylene andrehydrated in a graded series of ethanol. Sections were immersedin 3% hydrogen peroxide in methanol for 20 min to block endog-enous peroxidase activity. Slides were then incubated withuniversal blocking reagent (TA-125-UB, LabVision Ultra V Block-THERMO) for 7 min at room temperature. Afterwards, excessserum was drained and sections were incubated with primaryantibodies rabbit polyclonal anti-TH (1/400; Abcam, #ab-112),rabbit monoclonal iNOS (1/500; Cell signaling; #4236), rabbitmonoclonal nNOS 1/200; Cell signaling; #4231) in a humidifiedchamber overnight at +4�. For negative controls, the primaryantibodies were replaced by PBS. After several washes in PBS,sections were incubated with biotinylated goat anti-rabbit IgGantibody (1/400; Vector, #BA-1000) for 30 min followed by LSABstreptavidin-peroxidase complex (Vector ABC kit, VECTOR#PK400) incubation for 45 min and were rinsed with PBS.Antibody complexes were visualized by incubation with diam-inobenzidine (DAB) chromogen (TA-125-HD, DAB chromogenSubstrate System-THERMO). Sections were counterstained withMayer’s hematoxylin (Dako), dehydrated, mounted and examinedby an Axioplan microscope (Zeiss, Oberkochen, Germany). Theimages were taken using a 5MP Canon A95 camera integratedto the microscope.
We used semi-quantitative evaluations to determine theamount of immunohistochemical staining. Three randomlyselected slides, each of ten different fields of SN were evaluatedat 40� magnification. The distinct labeled cells with observedimmunostaining were counted in every lm2.
3.5. Nitrite/nitrate assay
Nitrite and nitrate, oxidized forms of NO, were determined bythe use of the nitrite/nitrate colorimetric assay kit (CAYMAN-780001). Nitrate cannot be measured directly. In this method,the nitrate in the SN and retina samples were converted into nitriteby nitrate reductase, and the total nitrite levels were determined asthe total nitrate/nitrite (NOx).
3.6. Caspase 3 activity assay
Activity of caspase 3 was measured by the colorimetric assaywith (_Invitrogen-KHZ220) following manufacturer’s protocol.Whole SN and retina tissues were homogenized in 200 lL lysisbuffer supplied with the kit and incubated on ice for 10 min. Aftercentrifugation for 15 min at 10,000g, supernatants were trans-ferred to clean tubes and assayed for caspase activity based onspectrophotometric detection of the chromophore p-nitroaniline(pNA) after cleavage from the labeled substrates. Samples of SNand retina tissues were mixed with 50 lL reaction buffer and50 lL 10 mM DTT, subsequently DEVDpNA was added and sampleswere incubated at 37 �C for 2 h in the dark. The absorbance wasthen measured at 405 nm in a microplate reader.
3.7. 4-HNE assay
Lipid peroxide levels were measured in SN and retina tissuesusing a 4-hydroxynonenal (4-HNE) assay kit (Cell Bio Labs,STA-334) following manufacturer’s protocol. The utilized protocolis an enzyme immunoassay developed for detection and quantita-ion of HNE-His protein adducts. Whole SN and retina tissues werehomogenized in 200 lL PBS buffer and centrifuged for 15 min at10,000g, supernatants were transferred to clean tubes and assayedfor quantitation of HNE-His protein adduct. The quantitiy ofHNE-His adduct in tissue samples were determined by comparingthe absorbance measured at 450 nm with that of known HNE-BSAstandards via a standard curve.
3.8. Protein measurements
Protein concentrations were measured at 595 nm by a modifiedBradford assay using Coomassie Plus reagent with bovine serumalbumin as a standard (Pierce Chemical Company, Rockford, IL,USA).
3.9. Statistical analysis
Analysis of variance (ANOVA) was performed on all parametersof VEPs for the factors of side (right and left) and groups. Differ-ences of other data were also analyzed by ANOVA followed up withTukey’s Post Hoc Test. Significance levels were set at P < 0.05.
4. Results
4.1. Animal health and survival
All animals appeared generally healthy during experimentalperiod. There were no difference in body weight gain and survivalswere similar for all experimental groups.
4.2. Motor activity
Motor activity are shown in Fig. 1A and B. Bradykinesia deter-mined turn time and total time. Turn time and total timeprolonged MPTP group compared with control group. Turn timedecrease in MPTP + 7-NI and MPTP + SMT group compared withMPTP group.
4.3. Visual evoked potentials
The means and standart errors of peak latencies, peak to peakamplitudes of VEP components and the results of statisticalanalysis in all experimental groups shown in Tables 1A and 1B.Differences in VEP parameters were analyzed by ANOVA. Measure-ments were made on five positive and four negative potentialswhich were seen all of the groups we did not find significant differ-ences in latencies and amplitudes between right and left eyes.Therefore the data from stimulation of both eyes were aver-aged.iNOS and nNOS inhibitor caused a significant delay in P1,N1, P2,N2, P3, N3, P4, N4, P5 components of VEPs in 7 NI andSMT group compared with control group. P1, N1, P2, N2, P3, P5latencies of VEP component prolonged in MPTP group comparedwith control group. P1, N1,N2, P3, N3 and P5 latencies of VEP com-ponent were significantly prolonged in 7-NI + MPTP group com-pared with control groups. P1 latency is decrease in 7-NI + MPTPgroup compared with MPTP group.
The means and standart errors of peak to peak amplitudes ofVEP components of all groups are given in Table 1B. ComponentP1N, N2P3 were significantly increased in SMT group compared
B
A*
0
2
4
6
8
Tim
e re
quir
ed to
turn
(T-T
urn)
(S)
T-TURN (S)
*
0
5
10
15
20
Tot
al T
urn
Tim
e (S
)
TOTAL TURN TIME (S)
Control 7-NI SMT MPTP 7-NI+MPTP SMT+MPTP
Control 7-NI SMT MPTP 7-NI+MPTP SMT+MPTP
Fig. 1. Determination of bradykinesia by pole test. (A) Time required to turn (s). (B)Total turn time. At day 7 post-intoxication, each mouse was placed head upward atthe top of a rough-surfaced pole double wrapped with gauze to prevent slipping.Data are means ± SEM. P < 0.05. � vs. Control.
S. Aras et al. / Neurochemistry International 72 (2014) 48–57 51
with control. P2N2, P4N4, N4P5 amplitude were significantly ele-vated in 7-NI + MPTP group compared with MPTP (Table 1B).
4.4. Substantia nigra and retina caspase-3 activity
Fig. 2A shown SN caspase 3 activity levels of study groups at theend of experimental period. Caspase 3 activity increased in theMPTP group compared with control group. SMT and 7-NI had noeffect in caspase 3 activity of SN. However 7-NI, SMT decreasedcaspase 3 activity in 7-NI + MPTP and SMT + MPTP compared with
The means and standart errors of peak to peak amplitudes of each groups. The mean valexpressed as means ± SEM for 10 mice each group.
# Compared with control P < 0.05.* Compared with MPTP P < 0.05.
MPTP group. Caspase 3 activity did not show any significantchange in the retina of all groups (Fig. 2B).
4.5. Nitrite/nitrate levels
Nitrite/nitrate levels of SN shown Fig. 3A. Nitrite/nitrate levelsincreased in MPTP group compared with control group. Nitrite/nitrate level was not found to be different in 7- NI and SMT groupcompared with control. Nitrite/nitrate levels of SN decreased in7-NI + MPTP and SMT + MPTP group compared with MPTP group.Nitrite/nitrate levels did not show any significant change in theretina of all groups (Fig. 3B).
4.6. 4-HNE levels
4-HNE is an indicator of lipid peroxidation. The level of 4-HNEincreased in MPTP group compared control group. Specific NOSinhibitors decreased the level of 4-HNE in SN of 7-NI + MPTP andSMT + MPTP groups compared with MPTP (Fig. 4A). 4-HNE levelsdid not show any significant change in the retina of all groups(Fig. 4B).
4.7. Expression of TH in the SN
The compact, reticular and lateral parts of mice SN were easilydistinguished by TH immunochemistry. The immunoreactivity forTH was observed in neuron bodies and processes. NO immunoreac-tivity was observed in glial cells and endothelium.
The TH immunoreactive neuron number in all groups werecounted independently by two observers blinded to the sourceand type of tissue according to these counts the dopaminergicneuron numbers in MPTP groups (%63) were significantly lowercompared to all the other groups. The quantitative number ofimmunostained cells are presented in Fig. 5C given below. Dopami-nergic neuron numbers significantly increased in 7-NI + MPTP andSMT + MPTP group compared with MPTP group. There were no sig-nificant differences in neuron number of SN in 7-NI and SMTgroups were compared control (Fig. 5C).
ue of each component was determined by averaging the data both eyes. Values are
A
B
**
*
**
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
SN C
aspa
se-3
act
ivity
(fol
d in
crea
se)
0
0,2
0,4
0,6
0,8
1
1,2
Ret
ina
Cas
pase
-3 a
ctiv
ity(f
old
incr
ease
)
Control 7-NI SMT MPTP 7-NI+MPTP SMT+MPTP
Control 7-NI SMT MPTP 7-NI+MPTP SMT+MPTP
Fig. 2. The effect of 7-NI and SMT on caspase-3 activity in the control and MPTPtreated groups in SN and retina tissues. (A) Substantia nigra Caspase-3 activity. (B)Retina Caspase-3 activity. Caspase-3 activities in all groups were measured by acaspase colorimetric activity assay kit. Data are means ± SEM. � Compared withcontrol group P < 0.05, �� compared with MPTP group P < 0.001.
** **
*
**
0
2
4
6
8
10
SN N
itra
te/N
itri
te(n
mol
/mg
prot
ein)
0
5
10
15
20
25
Ret
ina
Nit
rate
/Nit
rite
(nm
ol/m
g pr
otei
n)
Control 7-NI SMT MPTP 7-NI+MPTP SMT+MPTP
Control 7-NI SMT MPTP 7-NI+MPTP SMT+MPTP
A
B
Fig. 3. Nitrite/nitrate levels (A) substantia nigra and (B) retina. Data are mean-s ± SEM. � Compared control and 7-NI groups P < 0.05, �� compared with MPTPgroup P < 0.001.
*
0
1
2
3
4
5
SN 4
-HN
E(n
g/ug
tis
sue
prot
ein)
0
8
16
24
32
40
Ret
ina
4-H
NE
(ng/
ug t
issu
e pr
otei
n)
Control 7-NI SMT MPTP 7-NI+MPTP SMT+MPTP
Control 7-NI SMT MPTP 7-NI+MPTP SMT+MPTP
A
B
Fig. 4. 4-Hydroxynonenal levels. (A) Substantia nigra and (B) retina. Data aremeans ± SEM. � Compared with control group P < 0.05.
52 S. Aras et al. / Neurochemistry International 72 (2014) 48–57
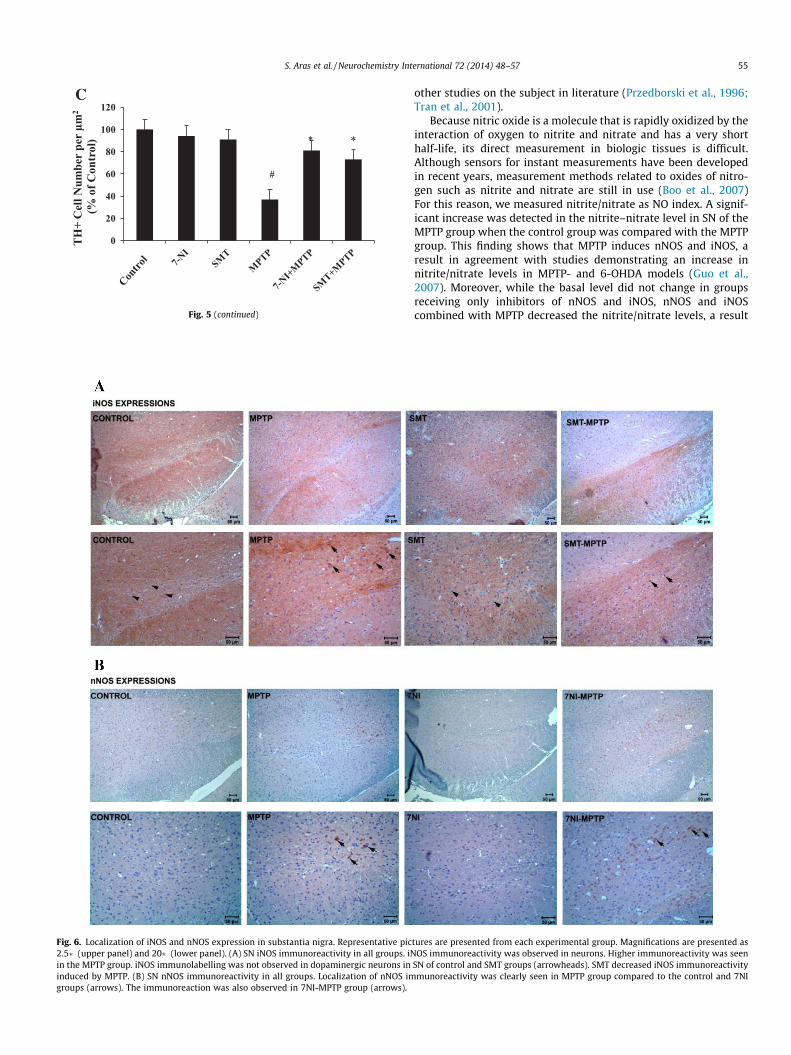
4.8. NOS immunoreactivity
iNOS and nNOS immunoreactivities were slightly observed inneuron cyctoplasm in control and 7NI, SMT treatment groups.Moreover, both of the antibodies immunoreaction resulted in mod-erate staining intensities in 7NI-MPTP and SMT-MPTP groups.However, iNOS and nNOS showed strong immunoreactivities in
MPTP group. The reactions were clearly increased in MPTP groups(Fig. 6A and B).
5. Discussion
Parkinson’s disease also affects the visual system; this effect iselucidated by a reliable parameter, the elongation of VEP latencies(O’Mahony et al., 1993). In our study, the use of specific inhibitorsof iNOS and nNOS resulted in improvement. This finding was sup-ported by biochemical parameters. NO plays a role in the process-ing of vision from the lowest level of retinal tranduction to thecontrol of neuronal excitability in the visual cortex (Lima et al.,2014). Thus, there is no doubt that parkinson induced lipid perox-idation and increase in NO synthesis can cause changes in brainand retinal functions. Consequently, it could be expected that iNOSand nNOS inhibitors improve visual system changes. In this regard,our study was undertaken to investigate the effect of iNOS andnNOS on visual system by means of visual evoked potentials whichconsist of several components arising from retina, optic pathway,subcortical and cortex. We have focused on the effect of lipid per-oxidation on altered VEP latencies in this study. Other parametersreported within the manuscript validate the MPTP-induced Parkin-sonism in our experimental settings.
Since induction of experimental PD in man is not possible, ani-mal models of PD have been used in the search for clues to theunderlying cause of the disorder and in the discovery of noveltreatments. The neurotoxic chemicals widely used in animal mod-els of PD are 6-OHDA, MPTP, paraquat, rotenon and manneb(Baltazar et al., 2014; Qi et al., 2014). These neurotoxins causemitochondrial dysfunction by inhibiting the Complex-I or Com-plex-III in the mitochondria. 6-OHDA is the hydroxyl analog ofDA. It destroys almost all dopaminergic neurons and also adrener-gic and serotonergic neurons. Since 6-OHDA cannot pass throughthe blood–brain barrier, it is injected into the Medial Forebrain
Fig. 5. (A) Localization of tyrosine hydroxylase (TH) protein in SN. TH immunoreactivity in the control group was almost the same as in 7NI and SMT groups. Theimmunostaining observed in the MPTP group was lower than other Parkinson experimental groups. Also, the immunoreactivity of 7NI-MPTP and SMT-MPTP groups werealmost the same as in control, 7NI and SMT groups. No immunoreactivity was observed on when TH antibody was replaced with normal rabbit IgG (negative control).Magnification is presented 2.5�. (B) Photomicrographs of TH immunohistochemistry in the dopaminergic neurons. The neuronal morphological alterations, the decrease ofTH immunoreactivity and loss dopaminegic fibers can be seen in the MPTP group. TH increased in 7-NI and SMT groups. Representative pictures are presented in highmagnification (10�). (C) TH+ cell number per lm2.
S. Aras et al. / Neurochemistry International 72 (2014) 48–57 53
Bundle (MFB) unilaterally or bilaterally by using the stereotaxicmethod (Betarbet et al., 2002).
The MPTP-treated primate model of PD, which closely mimicsthe clinical features of PD is undoubtedly the most clinically-relevant of all available models. This model which also mimicsthe first stage of PD is the most widely-used model for the studyof neurochemical and neuropathologic changes in this disorder(Wichmann and DeLong, 2003). There are also MPTP-treatedmouse and rat models for the study of PD (Bazzu et al., 0000. MPTPselectively shows toxic effect on dopaminergic neurons. Thischemical is administered to rats by using the stereotaxic method.In mice it destroys the dopaminergic neurons in the SN (Betarbetet al., 2002). Lesion formation is dependent on the dose of MPTPadministered. Rats are resistant to the degenerative effect of MPTPon dopaminergic neurons (Betarbet et al., 2002). MPTP can beadministered to mice by gavage feeding, subcutaneously,
intravenously, intraperitoneally or intramuscularly (Bazzu et al.,0000). The degree of dopaminergic damage depends on the doseand application protocol of the chemical. The C57BL/6 strain ofmice is more susceptible to systemic MPTP than other strains ofmice, where the chemical selectively damages the dopaminergicneurons (Castagnoli et al., 2001). In view of these data in literature,in our animal model we preferred to administer MPTP intraperito-neally at a dose of 4 � 20 mg/kg with 12-h intervals for 2 days(Date et al., 1990). The purpose of this application was to decreasethe risk of death in the animals. In experiments with higher dosesthere was a higher rate of death.
The damage induced in animal models, particularly the damagein their dopaminergic and adrenergic neurons, is assessed withmotor performance tests. In our study, peg test was used to assessthe motor activity in Parkinson models. With peg test we deter-mined the degree of bradykinesia, one of the cardinal symptoms
Fig. 5 (continued)
54 S. Aras et al. / Neurochemistry International 72 (2014) 48–57
of PD. Time of delay in executing the command to move shows thedegree of bradykinesia (Ogawa, 1997). In our model, prolongedreaction time was noted both in starting to move as well as inreturn. This result showed that our animal model was successfullybuilt. Upon administration of iNOS and nNOS with MPTP, the levelof bradykinesia fell down to that of the control, a result whichunderlines the role of iNOS and nNOS in motor movement impair-ement in PD.
To obtain additional proof of the success of our animal model,the number of TH immunoreactive cells was assessed with immu-nohistochemical methods. We found a decrease of 63% in the cellsof SN in the MPTP group. This result also indicates the success ofour model. We also noted the protective effect of the inhibitorsof iNOS and nNOS, a finding supported by biochemical parameters.
Although the pathogenesis of PD has not been fully clarifieduntil present time, studies have elucidated the role of lipid perox-idation, a process implicated in the pathogenesis of various disor-ders (Pall et al., 1986). Nitric oxide is quite reactive and unstablegas taking part with other agents in lipid peroxidation (Jenner,1998). Studies have shown the role of NO in forming free radicalsand neurodegeneration (Jenner, 1998). Nitric oxide has been foundto be localized to the central nervous system (hippocampus, stria-tum) as well as to the visual system (Vielma et al., 2012). Formerstudies have shown an increase in the expression and activity ofnNOS and iNOS in PD (Levecque et al., 2003). In relevance to thesefindings, we assessed the role of NO in visual system changes in PD,using the inhibitors of nNOS and iNOS. The doses and routes ofadministration of the inhibitors were determined after considering
Fig. 5 (continued)
Fig. 6. Localization of iNOS and nNOS expression in substantia nigra. Representative pic2.5� (upper panel) and 20� (lower panel). (A) SN iNOS immunoreactivity in all groups. iNin the MPTP group. iNOS immunolabelling was not observed in dopaminergic neurons ininduced by MPTP. (B) SN nNOS immunoreactivity in all groups. Localization of nNOS imgroups (arrows). The immunoreaction was also observed in 7NI-MPTP group (arrows).
S. Aras et al. / Neurochemistry International 72 (2014) 48–57 55
other studies on the subject in literature (Przedborski et al., 1996;Tran et al., 2001).
Because nitric oxide is a molecule that is rapidly oxidized by theinteraction of oxygen to nitrite and nitrate and has a very shorthalf-life, its direct measurement in biologic tissues is difficult.Although sensors for instant measurements have been developedin recent years, measurement methods related to oxides of nitro-gen such as nitrite and nitrate are still in use (Boo et al., 2007)For this reason, we measured nitrite/nitrate as NO index. A signif-icant increase was detected in the nitrite–nitrate level in SN of theMPTP group when the control group was compared with the MPTPgroup. This finding shows that MPTP induces nNOS and iNOS, aresult in agreement with studies demonstrating an increase innitrite/nitrate levels in MPTP- and 6-OHDA models (Guo et al.,2007). Moreover, while the basal level did not change in groupsreceiving only inhibitors of nNOS and iNOS, nNOS and iNOScombined with MPTP decreased the nitrite/nitrate levels, a result
tures are presented from each experimental group. Magnifications are presented asOS immunoreactivity was observed in neurons. Higher immunoreactivity was seen
SN of control and SMT groups (arrowheads). SMT decreased iNOS immunoreactivitymunoreactivity was clearly seen in MPTP group compared to the control and 7NI
56 S. Aras et al. / Neurochemistry International 72 (2014) 48–57
supporting former studies on the subject (Levecque et al., 2003).This result indicates the importance of nNOS and iNOS in neuronalapoptosis. No change was detected in the levels of nitrite/nitrate inthe retina. Therefore, our result indicates that the administration ofMPTP does not cause an increased expression of the neurotrans-mitter NO or of nNOS and iNOS in the retina as much as it doesin SN, or it indicates that the neuromelanin-containing dopaminer-gic neurons in the retina are less than those found in SN. Anotherinterpretation of this result is that MPTP in the retina cannot beconverted to MPP+. The active form of MPTP is the free radicalMPP+; conversion to MPP+ is mediated by the enzyme MAO-B inglial cells (Kalgutkar and Castagnoli, 1992). This mechanism couldnot be efficient in the retina, or other mechanisms could be moreeffective.
In this study, the levels of lipid peroxidation were determinedwith 4-HNE measurements in order to evaluate the relationshipbetween changes in VEP and oxidative damage. By dissociationof lipid peroxides, aldehydes such as 2-alkenes, 2,4 alkenes and4-hydroxyalkenes form. The hexzonal Omega-6, formed by theoxidation of fatty acids and 4-HNE are the primary aldehydes.4-HNE bears cytotoxic, mutagenic and genotoxic properties(Soulere et al., 2007). Oxygen radicals make up the secondarytoxic messenger of lipid peroxidation. The role of lipid peroxida-tion in certain pathologies is well known. In PD, lipid peroxidationand the level of 4-HNE in SN are increased (Andersen, 2004). Theincrease in 4-HNE stimulates the caspase 3-8-9 activation whichtriggers the apoptotic process (Ji et al., 2001). Moreover, 4-HNEcauses aggregation of a-synuclein (Hattoria et al., 2009) and frag-mentation of DNA (Zhou and Zhu, 2009). Our study showed that4-HNE increased quantitatively when MPTP was administered.The effect of MPTP is mediated by nNOS and iNOS (Levecqueet al., 2003). This increase in 4-HNE is mediated by NO and freeradicals formed by nNOS and iNOS (Levecque et al., 2003). Inagreement with this fact, in groups that received iNOS and nNOSinhibitors we found a significant decrease in 4-HNE in SN, but nosignificant decrease in the retina. These results indicate that,beside the roles of iNOS and nNOS, the COX-2 pathway and otherpathways might play a role in lipid peroxidation. In our formerstudy we showed that MPTP caused an increase in the COX-2activity and lipid peroxidation (Ozsoy et al., 2011). On the otherhand, there was no difference between the groups in regard tothe levels of MPTP in the retina. This result indicates that the ret-ina is not affected by MPTP as much as SN. In literature, no dataexists on the effect of MPTP on 4-HNE and nitrite–nitrate levelsin the retina.
When the caspase-3 activity as indicator of apoptosis was stud-ied, in SN and retina the following were observed: in MPTP-groupsof PD the caspase-3 activity in the retina showed no change, butwas increased in the SN, and in groups receiving nNOS and iNOSinhibitors, the caspase-3 activity was less than that found inMPTP-groups. These findings are in agreement with the results ofthe study by Brzozowski et al. (2011). It has been reported thatnNOS and iNOS have roles in apoptosis, and inhibition of iNOSleads to a decrease in caspase-3 activity (Du et al., 2001). Theseresults support our study. The inhibition of nNOS and iNOS showsa protective effect (Brzozowski et al., 2011).
The immunohistochemical results and biochemical parametersfound in this study affirm that in experimental PD the inhibitionof iNOS and nNOS exerts a corrective effect. The examination ofTH-stained dopaminergic neurons in SN showed a decrease inthe number of TH-stained neurons when MPTP was administered.This result is in agreement with literature (Kastner et al., 1994).When MPTP was given in combination with SMT and 7-NI, theconsecutive specific inhibitors of iNOS and nNOS, the formerlydecreased neuron count showed an increase. This result indicatesthe crucial roles played by iNOS and nNOS in apoptosis and
neurodegeneration. Nevertheless, the inhibitors of iNOS and nNOSnever caused neuron counts equal to those in the controls. Thisresult points out the role of other mechanisms in apoptosis. Withconversion of MPTP into MPP+ apoptosis is induced. As MPTPturns into MPP+, mitochondria damage 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and increase NOproduction, thus by violating redox balance cause the death ofdopaminergic cells. Excess production of NO causes the loss ofmitochondrial membrane potential, secretion of cytochrome-cand binding with APAF-1 which together increase the caspase-3activity (Eberhardt and Schulz, 2003).
nNOS, iNOS and eNOS can also be expressed under normal cir-cumstances. The conversion of MPTP into MPP+ causes an eleva-tion of intracellular Ca+2 which stimulates nNOS activity andalso causes increased NO production. Moreover, factors secretedby microglia and astrocytes play a role in apoptosis by increasingthe expression of iNOS and leading to the production of MPP+.Our findings show that the inhibition of iNOS and nNOS preventsapoptosis, but not in the level of the control group. The intracellu-lar accumulation of Ca+2 damages mitochondrial functions andactivates phospholipase, protease and endonuclease; moreover,apoptosis has a role in the COX-2 pathway (Brown, 2010). In ourstudy, the inhibitors of iNOS and nNOS prevented apoptosis tosome degree, decreased the caspase-3 activity, and diminishedthe 4-HNE quantity. These decrements would not be caused onlyby iNOS and nNOS, but also by other pathways as well. Since nochange indicative of apoptosis was observed in the retina,parameters such as caspase-3 activity, 4-HNE and nitrite/nitratewere not immunohistochemically evaluated. In our former studyon PD, no immunohistochemical change had been found (Ozsoyet al., 2011).
In patients with PD the visual system is also disturbed (Buttneret al., 1996). Varying with the severity and duration of PD, VEPsshow prolongation of P100 latency and changes in amplitude(Okuda et al., 1995). Also ERG amplitudes are lowered and thresh-old values are increased (Nightingale et al., 1986). Particularly theP100 latency is prolonged, which is identical with the P3 latenciesin animals.
Nitric oxide and sGC are amply present in photoreceptors andthe retina, within Müller, amacrine, bipolar, horizontal and gan-glion cells. nNOS and iNOS are present in the retina. Nitric oxidehas a role in the regulation of light-dependent gap junctions(Vielma et al., 2012; Pang et al., 2010). Nitric oxide modulatesthe phototransduction cascade and neurotransmitter release whileinhibiting DA release.
When the inhibitors of iNOS are nNOS were given, the latenciesof all VEP components were prolonged in relation to the controlgroup. MPTP was found to cause prolongation in the latencies ofVEP components. This result is in agreement with the data in liter-ature (Ozsoy et al., 2011). Following the administration of thenNOS inhibitor, correction was observed in P1, P4, and N4; whileP1 was corrected, P5 was prolonged. Our study shows that in thevisual system the presence MPTP increases the expression of nNOSwhereas its inhibition corrects many components. In the visualsystem nNOS and iNOS induce the production of NO, a significantneurotransmitter and neuromodulator. The data obtained indi-cates that nNOS and iNOS exert their effect in the central nervoussystem rather than in the retina.
It has been concluded that in MPTP-treated mouse models of PDthe visual system is affected and the inhibitors of nNOS and iNOSprovided a protective effect against damage in dopaminergicneurons. Nevertheless, the observation that the VEP values in thetreated groups did not attain the VEP values of the control groupsindicates the likely contribution of other pathways in the involve-ment of the visual system. For elucidation, further studies arerequired.
S. Aras et al. / Neurochemistry International 72 (2014) 48–57 57
Acknowledgments
This study was supported by Akdeniz University ResearchProjects Unit (Project no: 2010.02.0122.007).
References
Andersen, J.K., 2004. Oxidative stress in neurodegeneration: cause or consequence?Nat. Med. 10 (Suppl), S18–S25.
Baltazar, M.T., Dinis-Oliveira, R.J., de Lourdes Bastos, M., Tsatsakis, A.M., Duarte, J.A.,Carvalho, F., 2014. Pesticides exposure as etiological factors of Parkinson’s diseaseand other neurodegenerative diseases-a mechanistic approach. Toxicol. Lett.http://dx.doi.org/10.1016/j.toxlet.2014.01.039 (pii: S0378-4274(14)00059-9).
Baraboi, V.A., 1989. The role of lipid peroxidation in the mechanism of stress. Fiziol.Zh. 35 (5), 85–97.
Bazzu, G., Calia, G., Puggioni, G., Spissu, Y., Rocchitta, G., Debetto, P., et al., 2010.Alpha-Synuclein- and MPTP-generated rodent models of Parkinson’s diseaseand the study of extracellular striatal dopamine dynamics: a microdialysisapproach. CNS Neurol. Disord. Drug Targets. 9 (4), 482–490.
Betarbet, R., Sherer, T.B., Greenamyre, J.T., 2002. Animal models of Parkinson’sdisease. BioEssays 24 (4), 308–318.
Bodis-Wollner, I., Yahr, M.D., 1978. Measurements of visual evoked potentials inParkinson’s disease. Brain 101 (4), 661–671.
Boo, Y.C., Tressel, S.L., Jo, H., 2007. An improved method to measure nitrate/nitritewith an NO-selective electrochemical sensor. Nitric Oxide 16 (2), 306–312(PMCID: 1850282).
Brown, G.C., 2010. Nitric oxide and neuronal death. Nitric Oxide 23 (3), 153–165.Brzozowski, M.J., Alcantara, S.L., Iravani, M.M., Rose, S., Jenner, P., 2011. The effect of
nNOS inhibitors on toxin-induced cell death in dopaminergic cell lines dependson the extent of enzyme expression. Brain Res. 1404, 21–30.
Bugnon, O., Schaad, N.C., Schorderet, M., 1994. Nitric oxide modulates endogenousdopamine release in bovine retina. Neuroreport 5 (4), 401–404.
Castagnoli, K.P., Steyn, S.J., Petzer, J.P., Van der Schyf, C.J., Castagnoli Jr., N., 2001.Neuroprotection in the MPTP Parkinsonian C57BL/6 mouse model by acompound isolated from tobacco. Chem. Res. Toxicol. 14 (5), 523–527.
Celesia, G.G., 1984. Evoked potential techniques in the evaluation of visual function.J. Clin. Neurophysiol. 1 (1), 55–76.
Date, I., Felten, D.L., Felten, S.Y., 1990. Long-term effect of MPTP in the mouse brainin relation to aging: neurochemical and immunocytochemical analysis. BrainRes. 519 (1–2), 266–276.
Di Matteo, V., Pierucci, M., Benigno, A., Crescimanno, G., Esposito, E., Di Giovanni, G.,2009. Involvement of nitric oxide in nigrostriatal dopaminergic systemdegeneration: a neurochemical study. Ann. N. Y. Acad. Sci. 1155, 309–315.
Djamgoz, M.B., Cunningham, J.R., Davenport, S.L., Neal, M.J., 1995. Nitric oxideinhibits depolarization-induced release of endogenous dopamine in the rabbitretina. Neurosci. Lett. 198 (1), 33–36.
Du, Y., Ma, Z., Lin, S., Dodel, R.C., Gao, F., Bales, K.R., et al., 2001. Minocyclineprevents nigrostriatal dopaminergic neurodegeneration in the MPTP model ofParkinson’s disease. Proc. Natl. Acad. Sci. U.S.A. 98 (25), 14669–14674 (PMCID:64739).
Eberhardt, O., Schulz, J.B., 2003. Apoptotic mechanisms and antiapoptotic therapy inthe MPTP model of Parkinson’s disease. Toxicol. Lett. 139 (2–3), 135–151.
Guo, S., Yan, J., Yang, T., Yang, X., Bezard, E., Zhao, B., 2007. Protective effects of greentea polyphenols in the 6-OHDA rat model of Parkinson’s disease throughinhibition of ROS-NO pathway. Biol. Psychiatry 62 (12), 1353–1362.
Gutteridge, J.M., 1994. Biological origin of free radicals, and mechanisms ofantioxidant protection. Chem. Biol. Interact. 91 (2–3), 133–140.
Hattoria, N., Wanga, M., Taka, H., Fujimura, T., Yoritaka, A., Kubo, S., et al., 2009.Toxic effects of dopamine metabolism in Parkinson’s disease. ParkinsonismRelat. Disord. 15 (Suppl 1), S35-8.
Hesslinger, C., Strub, A., Boer, R., Ulrich, W.R., Lehner, M.D., Braun, C., 2009.Inhibition of inducible nitric oxide synthase in respiratory diseases. Biochem.Soc. Trans. 37 (Pt 4), 886–891.
Jenner, P., 1998. Oxidative mechanisms in nigral cell death in Parkinson’s disease.Mov. Disord. 13 (Suppl 1), 24–34.
Ji, C., Amarnath, V., Pietenpol, J.A., Marnett, L.J., 2001. 4-Hydroxynonenal inducesapoptosis via caspase-3 activation and cytochrome c release. Chem. Res.Toxicol. 14 (8), 1090–1096.
Kalgutkar, A.S., Castagnoli Jr., N., 1992. Synthesis of novel MPTP analogs as potentialmonoamine oxidase B (MAO-B) inhibitors. J. Med. Chem. 35 (22), 4165–4174.
Kastner, A., Herrero, M.T., Hirsch, E.C., Guillen, J., Luquin, M.R., Javoy-Agid, F., et al.,1994. Decreased tyrosine hydroxylase content in the dopaminergic neurons ofMPTP-intoxicated monkeys: effect of levodopa and GM1 ganglioside therapy.Ann. Neurol. 36 (2), 206–214.
Kidd, P.M., 2000. Parkinson’s disease as multifactorial oxidative neurodegeneration:implications for integrative management. Altern. Med. Rev. 5 (6), 502–529.
Kishimoto, J., Tsuchiya, T., Emson, P.C., Nakayama, Y., 1996. Immobilization-inducedstress activates neuronal nitric oxide synthase (nNOS) mRNA and protein inhypothalamic–pituitary–adrenal axis in rats. Brain Res. 720 (1–2), 159–171.
Krukoff, T.L., 1999. Central actions of nitric oxide in regulation of autonomicfunctions. Brain Res. Brain Res. Rev. 30 (1), 52–65.
Kucukatay, V., Hacioglu, G., Savcioglu, F., Yargicoglu, P., Agar, A., 2006. Visual evokedpotentials in normal and sulfite oxidase deficient rats exposed to ingestedsulfite. Neurotoxicology 27 (1), 93–100.
Langheinrich, T., Tebartz van Elst, L., Lagreze, W.A., Bach, M., Lucking, C.H., Greenlee,M.W., 2000. Visual contrast response functions in Parkinson’s disease: evidencefrom electroretinograms, visually evoked potentials and psychophysics. Clin.Neurophysiol. 111 (1), 66–74.
Levecque, C., Elbaz, A., Clavel, J., Richard, F., Vidal, J.S., Amouyel, P., et al., 2003.Association between Parkinson’s disease and polymorphisms in the nNOS andiNOS genes in a community-based case-control study. Hum. Mol. Genet. 12 (1),79–86.
Lima, M.G., Maximino, C., Matos Oliveira, K.R., Brasil, A., Crespo-Lopez, M.E., BatistaEde, J., Rocha, F.A., Picanço-Diniz, D.L., Herculano, A.M., 2014. Nitric oxide as aregulatory molecule in the processing of the visual stimulus. Nitric Oxide 36,44–50.
Neal, M.J., Cunningham, J.R., Matthews, K.L., 1999. Release of endogenous ascorbicacid preserves extracellular dopamine in the mammalian retina. Invest.Ophthalmol. Vis. Sci. 40 (12), 2983–2987.
Nightingale, S., Mitchell, K.W., Howe, J.W., 1986. Visual evoked cortical potentialsand pattern electroretinograms in Parkinson’s disease and control subjects. J.Neurol. Neurosurg. Psychiatry 49 (11), 1280–1287 (PMCID: 1029077).
O’Donnell, V.B., Chumley, P.H., Hogg, N., Bloodsworth, A., Darley-Usmar, V.M.,Freeman, B.A., 1997. Nitric oxide inhibition of lipid peroxidation: kinetics ofreaction with lipid peroxyl radicals and comparison with alpha-tocopherol.Biochemistry 36 (49), 15216–15223.
Ogawa, N., 1997. Dopamine neurotransmission and treatments for Parkinson’sdisease in the molecular biology era. Eur. Neurol. 38 (Suppl 1), 2–5.
Okuda, B., Tachibana, H., Kawabata, K., Takeda, M., Sugita, M., 1995. Visual evokedpotentials (VEPs) in Parkinson’s disease: correlation of pattern VEPsabnormality with dementia. Alzheimer Dis. Assoc. Disord. 9 (2), 68–72.
O’Mahony, D., Rowan, M., Feely, J., O’Neill, D., Walsh, J.B., Coakley, D., 1993.Parkinson’s dementia and Alzheimer’s dementia: an evoked potentialcomparison. Gerontology 39 (4), 228–240.
Ozsoy, O., Tanriover, G., Derin, N., Uysal, N., Demir, N., Gemici, B., et al., 2011. Theeffect of docosahexaenoic acid on visual evoked potentials in a mouse model ofParkinson’s disease: the role of cyclooxygenase-2 and nuclear factor kappa-B.Neurotox. Res. 20 (3), 250–262.
Pang, J.J., Gao, F., Wu, S.M., 2010. Light responses and morphology of bNOS-immunoreactive neurons in the mouse retina. J. Comp. Neurol. 518 (13), 2456–2474 (PMCID: 3000797).
Peachey, N.S., Ball, S.L., 2003. Electrophysiological analysis of visual function inmutant mice. Doc. Ophthalmol. 107 (1), 13–36.
Przedborski, S., Jackson-Lewis, V., Yokoyama, R., Shibata, T., Dawson, V.L., Dawson,T.M., 1996. Role of neuronal nitric oxide in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced dopaminergic neurotoxicity. Proc. Natl.Acad. Sci. U.S.A. 93 (10), 4565–4571 (PMCID: 39317).
Qi, Z., Miller, G.W., Voit, E.O., 2014. Rotenone and paraquat perturb dopaminemetabolism: a computational analysis of pesticide toxicity. Toxicology 315, 92–101.
Sener, H.O., Akbostanci, M.C., Yucesan, C., Dora, B., Selcuki, D., 2001. Visual evokedpotentials in Parkinson’s disease-correlation with clinical involvement. Clin.Neurol. Neurosurg. 103 (3), 147–150.
Smeyne, M., Smeyne, R.J., 2002. Method for culturing postnatal substantia nigra asan in vitro model of experimental Parkinson’s disease. Brain Res. Brain Res.Protoc. 9 (2), 105–111.
Soulere, L., Queneau, Y., Doutheau, A., 2007. An expeditious synthesis of 4-hydroxy-2E-nonenal (4-HNE), its dimethyl acetal and of related compounds. Chem. Phys.Lipids 150 (2), 239–243.
Tieu, K., Ischiropoulos, H., Przedborski, S., 2003. Nitric oxide and reactive oxygenspecies in Parkinson’s disease. IUBMB Life 55 (6), 329–335.
Tran, M.H., Yamada, K., Olariu, A., Mizuno, M., Ren, X.H., Nabeshima, T., 2001.Amyloid beta-peptide induces nitric oxide production in rat hippocampus:association with cholinergic dysfunction and amelioration by inducible nitricoxide synthase inhibitors. FASEB J. 15 (8), 1407–1409.
Vielma, A.H., Retamal, M.A., Schmachtenberg, O., 2012. Nitric oxide signaling in theretina: what have we learned in two decades? Brain Res. 1430, 112–125.
Wichmann, T., DeLong, M.R., 2003. Pathophysiology of Parkinson’s disease: theMPTP primate model of the human disorder. Ann. N. Y. Acad. Sci. 991, 199–213.
Yokoyama, H., Kuroiwa, H., Kasahara, J., Araki, T., 2011. Neuropharmacologicalapproach against MPTP (1-methyl-4-phenyl-1,2,3,6- tetrahydropyridine)-induced mouse model of Parkinson’s disease. Acta Neurobiol. Exp. (Wars) 71(2), 269–280.