This article was downloaded by: [Dalhousie University] On: 03 September 2013, At: 11:24 Publisher: Routledge Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Neurocase: The Neural Basis of Cognition Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/nncs20 The spinning dancer illusion and spontaneous brain fluctuations: An fMRI study Byron Bernal a , Magno Guillen a & Juan Camilo Marquez a a Department of Radiology, Miami Children’s Hospital, Miami, FL, USA Published online: 28 Aug 2013. To cite this article: Neurocase (2013): The spinning dancer illusion and spontaneous brain fluctuations: An fMRI study, Neurocase: The Neural Basis of Cognition, DOI: 10.1080/13554794.2013.826692 To link to this article: http://dx.doi.org/10.1080/13554794.2013.826692 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Transcript
This article was downloaded by: [Dalhousie University]On: 03 September 2013, At: 11:24Publisher: RoutledgeInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Neurocase: The Neural Basis of CognitionPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/nncs20
The spinning dancer illusion and spontaneousbrain fluctuations: An fMRI studyByron Bernala, Magno Guillena & Juan Camilo Marqueza
a Department of Radiology, Miami Children’s Hospital, Miami, FL, USAPublished online: 28 Aug 2013.
To cite this article: Neurocase (2013): The spinning dancer illusion and spontaneous brain fluctuations: An fMRIstudy, Neurocase: The Neural Basis of Cognition, DOI: 10.1080/13554794.2013.826692
To link to this article: http://dx.doi.org/10.1080/13554794.2013.826692
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy, completeness, or suitabilityfor any purpose of the Content. Any opinions and views expressed in this publication are the opinionsand views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy ofthe Content should not be relied upon and should be independently verified with primary sources ofinformation. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands,costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial orsystematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distributionin any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
The spinning dancer illusion and spontaneous brainfluctuations: An fMRI study
Byron Bernal, Magno Guillen, and Juan Camilo Marquez
Department of Radiology, Miami Children’s Hospital, Miami, FL, USA
The brain activation associated with the Spinning Dancer Illusion, a cognitive visual illusion, is not entirelyknown. Inferences from other study modalities point to the involvement of the dorso-parieto-occipital areas inthe spontaneous switchings of perception in other bistable non-kinetic illusions. fMRI is a mature technique usedto investigate the brain responses associated with mental changes. Resting-state fMRI is a novel technique that mayhelp ascertain the effects of spontaneous brain changes in the top-down regulation of visual perception. The pur-pose of this report is to describe the brain activation associated with the subjective illusory changes of perception ofa kinetic bistable stimulus. We hypothesize that there is a relationship between the perception phases with the veryslow cortical spontaneous fluctuations, recently described. A single normal subject who was trained to produce vol-untarily perception phase switches underwent a series of fMRI studies whose blocks were either defined post-hocor accordingly with a predefined timeline to assess spontaneous and voluntarily evoked visual perception switches,respectively. Correlation of findings with resting-state fMRI and independent component analysis of the task serieswas sought. Phases of the rotation direction were found associated with right parietal activity. Independent compo-nent analysis of the task series and their comparison with basal resting-state components suggest that this activity isrelated to one of the very slow spontaneous brain fluctuations. The spontaneous fluctuations of the cortical activitymay explain the subjective changes in perception of direction of the Spinning Dancer Illusion. This observation is aproof-of-principle, suggesting that the spontaneous brain oscillations may influence top-down sensory regulation.
A visual illusion may be defined as a perceptionthat deviates from the underlying reality of theimage (Corney & Lotto, 2007). These “deviations”may be understood as “systematic sensory dis-crepancies from simple measurements with tools(rulers, photometers, clocks, etc.)” (Gregroy, 1997).Visual illusions may be divided accordingly with thecause in two types: physical and cognitive (Gregroy,1997). The former is related to physical causesfrom the object to the retina and include lighteffects, contrast, perspective, refraction, etc. Thelatter is associated with the effect of prior expe-riences (knowledge). In modern neuropsychology,
Address correspondence to Byron Bernal, Department of Radiology, Miami Children’s Hospital, 3100 SW 62nd Ave., Miami, FL33155, USA. (E-mail: [email protected]).
these two opposed data-flow mechanisms are calledbottom-up and top-down processing, respectively.Each of these two types of illusions may fall intofour classes: ambiguity, distortions, paradoxes, andfictions (Gregroy, 1997).
A specific type of cognitive illusion is the illusionin which there are two stable possible interpreta-tions of the stimulus (bistable stimulus) (Kornmeier& Bach, 2006; Poston & Stewart, 1978). A familiarbistable stimulus is the Necker cube, which maybe perceived either as if it were seen from above,exposing the upper face, or from below, exposingthe lower face. In this ambiguous condition, the
brain “chooses” to perceive only one possibility at atime.
The spinning dancer is a bistable illusion that, incontrast to the Necker cube, is based on motion.
The spinning dancer illusion (SDI) is an anima-tion of a rotating silhouette along its major axis inwhich the front and the rear aspects of the imageare the same. Because of this ambiguity, the rota-tion may be experienced by the same subject aseither a clockwise or a counterclockwise rotation.The brain solves the ambiguity by choosing to per-ceive one direction. Often, over time, subjects reportthat the direction switches, or even takes a to-and-fro motion (the illusion an explanation of how thisworks may be seen at http://en.wikipedia.org/wiki/Spinning_Dancer).
The illusion persists despite the angle of view(Pastukhov, Vonau, & Braun, 2012), althoughreversals are more frequent when the viewer focuseson the feet (Owen, Downes, Sahakian, Polkey, &Robbins, 1990), and they are not associated withor are due to habituation (Hock, Schöner, & Voss,1997). Voluntary effort and speed of rotation mayinfluence the frequency of direction changes (Liu,Tzeng, Hung, Tseng, & Juan, 2012). Likewise, vol-untary effort to hold a given direction has beenfound to modify the duration of the phase of thatperception, particularly when attention is avertedsomehow from the core of the perception (Kohler,Haddad, Singer, & Muckli, 2008) or whenever thestimulus factors favored the intended motion phaseto perceive (Suzuki & Peterson, 2000) or even whenattention is focused on the stimulus (Watanabe &Shimojo, 1998).
SDI has not been studied with EEG, PET, MEG,or fMRI as of yet. Several non-neuroimaging stud-ies have tackled distinct aspects of the bistableeffects based either on static pictures (Petersik& McDill, 1981; Ta’eed, Ta’eed & Wright, 1988)or on simple motion illusions (Kuriki, Ashida,Murakami, & Kitaoka, 2008). The most frequentvisual bistable illusions studied have been theNecker cube and the spinning wheel illusions.Perceptual changes in bistable stimulus with staticstimulus have been found associated with right infe-rior parietal electrical activity (Britz, Landis, &Michel, 2009). In an fMRI study, subjects werescanned while observing a standard Necker cube,a disambiguated cube (erasing the hidden lines),and a pair of planes corresponding to the frontand posterior faces of the cube. From these exper-iments, the authors found premotor and parietal
areas during the Necker cube shift perception, areasalso found in mental processes associated withimage manipulation (Inui et al., 2000). In an event-related fMR study, voluntary shifts of attention andperceptual configuration have been associated withactivity in the posterior parietal cortex, a part ofthe attentional network (Slotnick & Yantis, 2005).In addition to these reports that consistently involveposterior dorsal parietal structures, there are otherreports pointing to the right frontal lobe and theinferior occipito-temporal areas (Lapeyre-Mestreet al., 1997; Pastukhov & Braun, 2007; Pitts,Martínez, Stalmaster, Nerger, & Hillyard, 2009;Shen et al., 2009).
Hsieh et al. studied the brain activation producedby the rebound motion illusion, a type of kineticbistable illusion elicited when viewing bars alternat-ing between white and black on a gray background(Hsieh, Caplovitz, & Tse, 2006). The stimulus isbistable because the subject’s perception alternatesbetween one of flashing bars (true) and the reboundmotion illusion, where the bars appear to moveback and forth. They found activation of the humanmotion processing area associated with shiftingbetween modalities of perception. The motion pro-cessing area is located in the dorsal visual streamat the level of the posterior parietal cortex. Thearea seems to be equally involved in switching,independent of the type of stimulus, since it hasalso been found with a different bistable illusion,called the dynamic dot quartet (an illusion con-sisting of the perception of motion of four blockslocated in the corners of a frame, which may be per-ceived as moving vertically or horizontally (Sterzer,Eger, & Kleinschmidt, 2003)), and the spinningwheel (Sterzer, Russ, Preibisch, & Kleinschmidt,2002). Occipital activation has also been describedin bistable stimulus but this seems more stimulusspecific (Freeman, Sterzer, & Driver, 2012).
In 2009, Raemaekers and coworkers reportedanother kinetic bistable illusion study with fMRI.They mapped the brain of subjects while look-ing at repeating or alternating rotational possibil-ities of a spinning sphere illusion (Raemaekers,van der Schaaf, van Ee, & van Wezel, 2009).Alternating perception was associated with the acti-vation of dorsal parietal and frontal areas, sup-plementary motor area (SMA), cerebellum, andinsula. An interesting finding of this group isthe individual bias of perceiving in a determinedway—the greater the bias, the smaller the activationobtained.
Despite the limited number of studies, the rela-tionship between brain activations associated withsustained perception of a bistable stimulus and theneural mechanisms involved in spontaneous switch-ing of perception is not yet well understood. It hasbeen found though that slow oscillations throughthe dorsal attentional network initiate spontaneousperceptual shifts (Ozaki et al., 2012). Dorso-fronto-parietal and putaminal activation for “selecting aparticular orientation” has been described in anfMRI study with a set of five static bistable stimuli(Raz, Lamar, Buhle, Kane, & Peterson, 2007).
These variations of perception may be related tothe spontaneous very slow oscillations of the basalbrain activity, which have been recently describedwith data-driven analysis of long BOLD-sensitivesequences with subjects at rest. The technique isknown as resting-state-fMRI. With this technique,data-driven computer algorithms (e.g., IndependentComponent Analysis) may extract brain areas show-ing high temporal or spatial correlation. Typically,these independent areas or “components” oscillateat very low frequencies, between 0.01 and 0.1 Hz.Of all cortical components, two have been describedinvolving ipsilateral connectivity between dorsalareas of the frontal and parietal lobes. For a reviewof state of the art discussion on resting-state fMRI,the reader is advised to read the paper “Resting-state brain networks: literature review and clinicalapplications” by Rosazza and Minati (2011).
The purpose of the present study was to demon-strate proof-of-principle that the brain activationrelated to the sustained perception (phase) of bothdirections on the SDI is correlated with sponta-neous brain activity of the kind recently describedin resting-state fMRI. To achieve our goal, we lever-aged a subject capable of choosing and sustaining atwill the direction of rotation and experiencing spon-taneous shifting of direction as any non-trainedvolunteer.
To our knowledge, this is the first study of fMRIon SDI, despite the popularity and striking volumeof publications on the Internet.
MATERIALS AND METHODS
Subject and acquisition sequences
The volunteer is one of the co-authors (JCM), a30-year-old right-handed male, who experiencedfrequent spontaneous changes in rotation direction
of SDI and who has also been trained to perceiveat will a specific turning direction of the image.For this purpose, JCM was instructed to focus hisattention on the dancer’s feet and mentally choosea given aspect (front or back) of the silhouettewhen the lower feet were at the midline. Lookingat the dancer’s feet has been described to increasethe switching of perception as mentioned above.This technique was repeated in a number of ses-sions, after which the subject was able to switch therotation direction at will.
The MRI acquisitions were performed on a 1.5 Tscanner (Philips Intera series A Scanner). Therewere a total of 5 runs, one structural and fourecho planar image (EPI) acquisitions sensitive tothe BOLD effect (Forster et al., 1998).
The first run was a pre-task resting-state fMRIacquisition, where the subject was instructed tolie down with his eyes closed and thinking of hisbreathing. The purpose of this was to determinethe basal brain activity by characterizing its neu-ral components. In the second run, the subjectwas presented with a spinning dancer animationprojected at 30 fps. In this run, the subject wasinstructed to indicate when a spontaneous change ofspinning direction was observed. The subject wasinstructed to indicate the initial direction of thespinning, utilizing the left or right index finger toindicate a counterclockwise or clockwise rotation,respectively. Subsequent changes in the directionof the spinning were prompted by a single rightindex movement. The number of time points cor-responding to each direction change was registered.With this sequence, a block design was obtained inwhich each perceived direction becomes a regressorto determine the brain activation associated with aspecific perception of direction, since the remain-der of the stimuli (luminance, intensity, brightness,contrast, frames per second, motor responses, etc.)is controlled. In the third acquisition (VoluntarilyControlled Perception), the subject is instructed toperceive a desired direction of spinning by request,in a block design of 40 seconds per cycle. Once thesubject is prompted for a specific spinning direc-tion, the subject reports if the desired shift tookplace by moving slightly the right index finger, andthat specific time point is recorded. This sequencecould be contrasted with the prior one to seek forcommonality and thus control for aspects differentfrom perception (motivation, switching triggering,monitoring). Moreover, it may serve to validatethe findings of the block analysis of the second
Dow
nloa
ded
by [
Dal
hous
ie U
nive
rsity
] at
11:
24 0
3 Se
ptem
ber
2013
4 BERNAL, GUILLEN, MARQUEZ
sequence. Finally, a post-task resting-state sequencewas acquired after all the sequences were com-pleted, in an attempt to assess the stability of theendogenous components.
All the fMRI sequences were echo-planar gra-dient imaging BOLD-sensitive sequences. Theparameters for the EPI sequence acquisition wereas follows: 200 time points; TR = 2000 ms, TE =60 ms, FA = 90◦, transversal in-plane voxel size 3.75× 3.75 mm, field of view (FOV) of 240 mm, andresolution 64 × 64 × 14. Axial cuts of 5 mm withno gap were taken in an inferior to superior man-ner using an 8-channel head sense coil. To allowco-registration of activation results, an anatomical3D T1 sequence was also acquired using voxel size1 × 1 × 2 mm, resolution 288 × 288 × 160.
Post-processing
The fMRI data sets were processed using theFMRIB software library version 4.1.9 (FSL soft-ware by Oxford University). Data pre-processingconsisted of motion detection and correction usingthe MCFLIRT algorithm (Jenkinson, Bannister,Brady, & Smith, 2002); brain extraction usingBET (Smith, 2002); spatial smoothing using theGaussian kernel of FWHM 7 mm; and high-passtemporal filtering (Gaussian-weighted least squarefitting (LSF) straight line fitting, with sigma =100 s).
The task-related series were post-processed usingFEAT (FMRI Expert Analysis Tool version 5.98),generating Z (Gaussianized T/F) statistic imagesthresholded using clusters determined by Z >
2.3 and a (corrected) cluster significance threshold
of p = .01 (Friston et al., 1994). In addition tothis approach, an independent component analysisof the spontaneous-observation dataset was per-formed using MELODIC version 3.10. (Jenkinson& Smith, 2001; Jenkinson et al., 2002; Rowe &Hoffmann, 2006). The resting-state series were pro-cessed only using MELODIC.
Specific post-processing
(1) Spontaneous SDI observation and VoluntarilyControlled Perception.The timetable, derived from the patient report,was utilized to generate a block design regres-sor accounting for epochs in which a given sus-tained rotational perception was experiencedby the subject. Two contrasts were applied inthe model to characterize the on and off statesin the paradigm. Notice that each epoch couldbe considered either “on” or “off” with respectto the opposite contrast. The observed direc-tional change has to be perceived for morethan 3 time points (6 s), to be considered asa real change; otherwise, the previous rotationdirection was kept.
The model for sustained observation of rota-tion direction can be observed in Figure 1.
(2) Independent Component Analysis (ICA).ICA was performed to the resting-state acqui-sitions and to the spontaneous spinning dancerobservation series. Neural components weresubjectively selected by consensus of twoauthors (BB and MG) with 3 years of experi-ence in resting-state fMRI, taking into accountcluster frequency domain (between 0.01 and0.1 Hz) and intraparenchymal localization.
Figure 1. Block design fMRI modeled from the spontaneous-perception phases of SDI. For this model, the “on” contrast correspondsto the counterclockwise direction since the starting perception phase was clockwise; the reverse contrast corresponds to the clockwisedirection.
Dow
nloa
ded
by [
Dal
hous
ie U
nive
rsity
] at
11:
24 0
3 Se
ptem
ber
2013
THE ORIGIN OF THE SPINNING DANCER ILLUSION 5
For the post-processing of the spontaneousobservation series, the information provided by theGLM files design.mat and design.con, which con-tain model definition and contrasts, respectively,were used to define a confound regressor of thepost-statistical analysis to ascertain the correlationbetween the ICA components and the providedmodel.
The objective of the ICA analysis on the taskseries was determining whether the perception ofa given spinning direction was associated witha brain activation distinct to the independentcomponents or intrinsic spontaneous networks asdetected in the first series or not. ICA serieswere post-processed with a 10% brain/backgroundthreshold (single-session ICA analysis, 50% thresh-old of IC maps). The ICA of the spontaneousspinning dancer observation was also correlatedwith the model regressor derived from the con-structed block design of the spontaneous rotationchanges. Significant correlation of the componentsto the regressor was accepted when a value ofF greater than 3.5 and p greater than .001 wereobtained.
Each final activation result was co-registeredwith the subject anatomical image using FLIRT5.5. and normalized to the WFU Pick Atlas(Wake Forest University School of Medicine)(Maldjian, Laurienti, Kraft, & Burdette, 2003),to allow segmentation in Brodmann areas (BA).All sequences and templates were uploaded intoAMIDE software (http://amide.sourceforge.net).Overlay of functional maps and BA regions wereperformed to determine brain activation localiza-tion in BA. To contrast and compare activationsbetween (1) the spontaneous and voluntarily con-trolled perception and (2) the spontaneous activationand the ICA components derived from the pre-task resting-state sequence, their maps were over-laid utilizing the same tool. The second contrastwas simplified by masking the dataset contain-ing the activation of the spontaneous perceptiontask with the 3D-activation cluster or region ofinterest (ROI) derived from the parieto-occipitalcomponent of the resting state. The activation out-side of the ROI was set automatically to zero,leaving just the activation that was included inthe ROI. This heuristic approach was privilegedover a parametric analysis of the comparisons,owing to the presence of multiple regions of inter-est that would imply unnecessary high complexdescription and troublesome interpretation of theresults.
RESULTS
The subject went through all the tasks withoutany overt or subjective stress. He was able to com-plete all the sequences and experience enough shiftsin direction to perform further analysis. Duringthe spontaneous perception run, the subject expe-rienced 90 time-points as counterclockwise and110 as clockwise perception. During voluntarilycontrolled perception, the subject was able to shiftdirection in all attempts during the next 2–4 s, asexpected.
TASKS
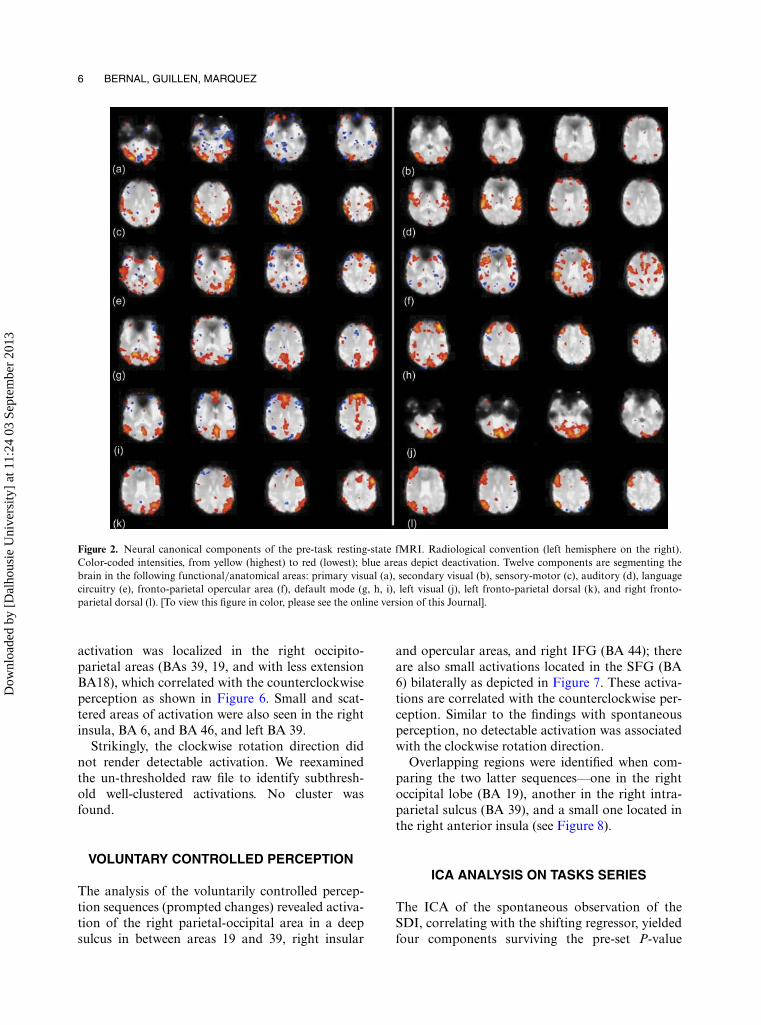
Several neural canonical components were iden-tified in the pre-task resting-state sequence asdepicted in Figure 2. The following can be identi-fied: primary and secondary visual, sensory motor,auditory, language, left occipital, default mode(posterior and anterior), and 2 asymmetric dorsalcomponents with connectivity between parietal andipsilateral frontal lobes (images k and l in Figure 2).Figure 3 displays a bar plot of the different con-tributions of each of the components found in theindependent component analysis of the pre-taskresting-state series.
The post-task resting-state fMRI showed thefollowing components: visual, auditory, salience,default mode, posterior cingulate gyrus, and againthe dorsal asymmetric parietal frontal observed inthe first resting-state sequence in Figure 4. Of noteis the stability of the dorso-fronto-parietal asym-metric components (images k and l in Figure 2,the same as i and j in Figure 4), and auditory (dand d) and default mode. Interestingly, the typicaldefault mode is less stable, although still recogniz-able (images h and i in Figure 2, also g and h inFigure 4). In Figure 5, a bar plot depicts the dif-ferent contributions of the components found inthe post-task brain state independent componentanalysis.
SPONTANEOUS OBSERVATION OF THESPINNING DANCER
The spontaneous observation task produced18 valid switches, for 10 blocks for condition 1(clockwise) and 9 blocks for condition 2 (coun-terclockwise). The minimum/maximum blocktime-point dimension ratio was 4/23. The main
Figure 2. Neural canonical components of the pre-task resting-state fMRI. Radiological convention (left hemisphere on the right).Color-coded intensities, from yellow (highest) to red (lowest); blue areas depict deactivation. Twelve components are segmenting thebrain in the following functional/anatomical areas: primary visual (a), secondary visual (b), sensory-motor (c), auditory (d), languagecircuitry (e), fronto-parietal opercular area (f), default mode (g, h, i), left visual (j), left fronto-parietal dorsal (k), and right fronto-parietal dorsal (l). [To view this figure in color, please see the online version of this Journal].
activation was localized in the right occipito-parietal areas (BAs 39, 19, and with less extensionBA18), which correlated with the counterclockwiseperception as shown in Figure 6. Small and scat-tered areas of activation were also seen in the rightinsula, BA 6, and BA 46, and left BA 39.
Strikingly, the clockwise rotation direction didnot render detectable activation. We reexaminedthe un-thresholded raw file to identify subthresh-old well-clustered activations. No cluster wasfound.
VOLUNTARY CONTROLLED PERCEPTION
The analysis of the voluntarily controlled percep-tion sequences (prompted changes) revealed activa-tion of the right parietal-occipital area in a deepsulcus in between areas 19 and 39, right insular
and opercular areas, and right IFG (BA 44); thereare also small activations located in the SFG (BA6) bilaterally as depicted in Figure 7. These activa-tions are correlated with the counterclockwise per-ception. Similar to the findings with spontaneousperception, no detectable activation was associatedwith the clockwise rotation direction.
Overlapping regions were identified when com-paring the two latter sequences—one in the rightoccipital lobe (BA 19), another in the right intra-parietal sulcus (BA 39), and a small one located inthe right anterior insula (see Figure 8).
ICA ANALYSIS ON TASKS SERIES
The ICA of the spontaneous observation of theSDI, correlating with the shifting regressor, yieldedfour components surviving the pre-set P-value
Dow
nloa
ded
by [
Dal
hous
ie U
nive
rsity
] at
11:
24 0
3 Se
ptem
ber
2013
THE ORIGIN OF THE SPINNING DANCER ILLUSION 7
Figure 3. Contributions of each component as identified by the blind source separation performed by independent component analysison the pre-task resting-state sequence. Note that since 3 default mode components were identified, the individual contributions of eachone were added. The value displayed for each component corresponds to the % Explained Variance observed on the rest-state seriesaccounted for each component.
Figure 4. Neural canonical components of the post-task resting-state fMRI. Same conventions as in Figure 2. Ten neural componentswere identified: visual (a), posterior cingulate gyrus (b), motor dorso-lateral (c), auditory-frontolateral (d), salient (e), auditory (f),frontal opercular (g), default mode (h), left fronto-parietal dorsal (i), and right fronto-parietal dorsal (j). Notice the reproducibility ofthe dorsal components (i and j) when compared to pre-task resting-state fMRI. [To view this figure in color, please see the online versionof this Journal].
Dow
nloa
ded
by [
Dal
hous
ie U
nive
rsity
] at
11:
24 0
3 Se
ptem
ber
2013
8 BERNAL, GUILLEN, MARQUEZ
Figure 5. Contributions of each component generated by the independent component analysis during the post-task resting-statesequence. Note the reduction of the contribution of default mode components in brain activity.
Figure 6. Brain activation related to counterclockwise perception of SDI. [To view this figure in color, please see the online version ofthis journal].
Figure 7. Brain activation related to counterclockwise perception of SDI, from the series with spinning direction voluntarily controlledby the subject. Main areas of activation are located in the right occipito-parietal region and the right insula. Some small dorsal premotorcortex and right frontobasal activations are present as well. [To view this figure in color, please see the online version of this Journal].
threshold. The highest correlation corresponded tocomponent #13 (F = 7.4; P = .0007), as shown inFigure 9.
This component is similar to the dorsal parieto-frontal asymmetric components (component i inFigure 2, and component j in Figure 4), which alsoinvolves BA 19 and 39. The second componentcorresponds to a left-hemisphere-dominant com-ponent (component #24, F = 6.33; P = .0021),
involving frontobasal, frontolateral, and opercularareas, along with left parietal and left SMA.This component suggests language circuitry. Thethird component (component #33, F = 6.22, P =.002) exhibits areas of activation common to theanterior and posterior default mode combined.The fourth component (component #6, F = 6.67,P = .004) depicts areas of activation in BA 46, 19,and 18.
Dow
nloa
ded
by [
Dal
hous
ie U
nive
rsity
] at
11:
24 0
3 Se
ptem
ber
2013
THE ORIGIN OF THE SPINNING DANCER ILLUSION 9
Figure 8. Common brain areas of activation between sponta-neous and “controlled” perception of counterclockwise rotationof SDI. Radiological convention. Spontaneous perception iscoded from light yellow to green; “controlled” perception iscoded from orange to red. [To view this figure in color, pleasesee the online version of this Journal].
DISCUSSION
In this paper, we report the fMRI findings ofbistable visual illusion in a subject who presentedfrequent and relatively stable changes in perceptionof the phases, and the odd ability to switch easily atwill from one phase of perception to the other. Thebistable paradigm used was the “spinning dancer”,with which we attempt to identify the brain areasinvolved in the phenomenon utilizing a combina-tion of two fMRI techniques: task-related blockdesign fMRI and resting-state ICA. We hypothe-size that the brain areas involved in the perceptionphase (the brain mechanisms associated with a
sustained type of perception in a bistable stimulus)have a correlation with the very slow spontaneousbrain activity.
We describe for the first time the brain activa-tion of BA 19 and 39 as a function of sustainedperception (phase) in a kinetic bistable stimulus,and as related to one of the networks conform-ing the components of the very slow spontaneousbrain fluctuations. This association occurred onlyin the right side, in keeping with the anticlock-wise perception. The involvement of BA 19 and39 areas in the sustained perception of a phaseof the SDI is not surprising since the functionof these areas in tracking of motion patterns(Deutschländer et al., 2002; Dieterich, Bauermann,Best, Stoeter, & Schlindwein, 2007; Dupont, Orban,De Bruyn, Verbruggen, & Mortelmans, 1994),visual attention to shapes (Le, Pardo, & Hu,1998), selective attention to visual targets (Larsson,Landy, & Heeger, 2006), and visual mental imagery(Knauff, Mulack, Kassubek, Salih, & Greenlee,2002) have been well established. More relevantis the specific right-side involvement in the analy-sis of stereoscopic stimulus (Fortin, Ptito, Faubert,& Ptito, 2002), required to discriminate depth,which is an important factor in the generationof SDI.
Our results are in agreement with Kamitaniand Tong (2006), who found that primary andsecondary visual cortices attend and decode spe-cific motion directions. In their experiment withfMRI and motion targets, they were able to predict
Figure 9. Independent component analysis of the spontaneous perception series (component #13), correlated to the block design regres-sor. Upper row: activations correlated to the regressor (green line in graph). Lower row: Time course plot of peak of intensity voxel andmodel fit (regressor). Conventions and color code intensities are the same as in prior images. The circuitry demonstrated corresponds toa default mode component with mild asymmetry to the right hemisphere. Notice the marked coincidence of the posterior right area ofactivation with the location of areas depicted in Figures 4–6. [To view this figure in color, please see the online version of this Journal].
Dow
nloa
ded
by [
Dal
hous
ie U
nive
rsity
] at
11:
24 0
3 Se
ptem
ber
2013
10 BERNAL, GUILLEN, MARQUEZ
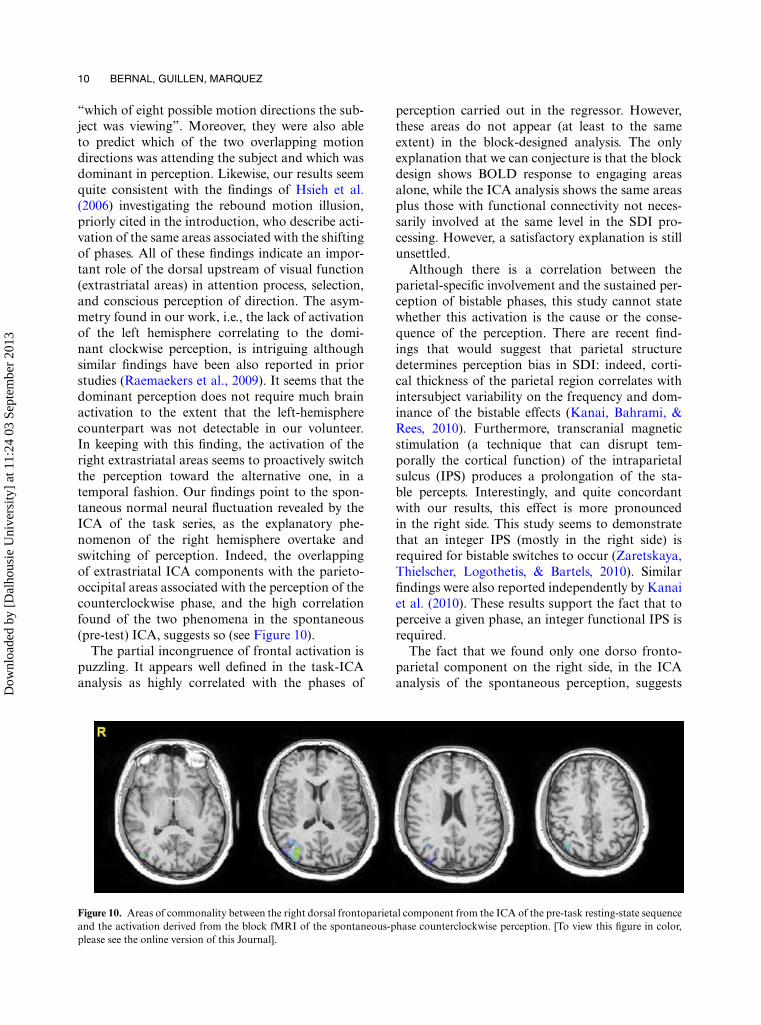
“which of eight possible motion directions the sub-ject was viewing”. Moreover, they were also ableto predict which of the two overlapping motiondirections was attending the subject and which wasdominant in perception. Likewise, our results seemquite consistent with the findings of Hsieh et al.(2006) investigating the rebound motion illusion,priorly cited in the introduction, who describe acti-vation of the same areas associated with the shiftingof phases. All of these findings indicate an impor-tant role of the dorsal upstream of visual function(extrastriatal areas) in attention process, selection,and conscious perception of direction. The asym-metry found in our work, i.e., the lack of activationof the left hemisphere correlating to the domi-nant clockwise perception, is intriguing althoughsimilar findings have been also reported in priorstudies (Raemaekers et al., 2009). It seems that thedominant perception does not require much brainactivation to the extent that the left-hemispherecounterpart was not detectable in our volunteer.In keeping with this finding, the activation of theright extrastriatal areas seems to proactively switchthe perception toward the alternative one, in atemporal fashion. Our findings point to the spon-taneous normal neural fluctuation revealed by theICA of the task series, as the explanatory phe-nomenon of the right hemisphere overtake andswitching of perception. Indeed, the overlappingof extrastriatal ICA components with the parieto-occipital areas associated with the perception of thecounterclockwise phase, and the high correlationfound of the two phenomena in the spontaneous(pre-test) ICA, suggests so (see Figure 10).
The partial incongruence of frontal activation ispuzzling. It appears well defined in the task-ICAanalysis as highly correlated with the phases of
perception carried out in the regressor. However,these areas do not appear (at least to the sameextent) in the block-designed analysis. The onlyexplanation that we can conjecture is that the blockdesign shows BOLD response to engaging areasalone, while the ICA analysis shows the same areasplus those with functional connectivity not neces-sarily involved at the same level in the SDI pro-cessing. However, a satisfactory explanation is stillunsettled.
Although there is a correlation between theparietal-specific involvement and the sustained per-ception of bistable phases, this study cannot statewhether this activation is the cause or the conse-quence of the perception. There are recent find-ings that would suggest that parietal structuredetermines perception bias in SDI: indeed, corti-cal thickness of the parietal region correlates withintersubject variability on the frequency and dom-inance of the bistable effects (Kanai, Bahrami, &Rees, 2010). Furthermore, transcranial magneticstimulation (a technique that can disrupt tem-porally the cortical function) of the intraparietalsulcus (IPS) produces a prolongation of the sta-ble percepts. Interestingly, and quite concordantwith our results, this effect is more pronouncedin the right side. This study seems to demonstratethat an integer IPS (mostly in the right side) isrequired for bistable switches to occur (Zaretskaya,Thielscher, Logothetis, & Bartels, 2010). Similarfindings were also reported independently by Kanaiet al. (2010). These results support the fact that toperceive a given phase, an integer functional IPS isrequired.
The fact that we found only one dorso fronto-parietal component on the right side, in the ICAanalysis of the spontaneous perception, suggests
Figure 10. Areas of commonality between the right dorsal frontoparietal component from the ICA of the pre-task resting-state sequenceand the activation derived from the block fMRI of the spontaneous-phase counterclockwise perception. [To view this figure in color,please see the online version of this Journal].
Dow
nloa
ded
by [
Dal
hous
ie U
nive
rsity
] at
11:
24 0
3 Se
ptem
ber
2013
THE ORIGIN OF THE SPINNING DANCER ILLUSION 11
rather a causal relationship between the sponta-neous brain fluctuation and the perception. Hadit been the other way around, we would haveobserved two similar components dissociated intime, an effect that we did not have: one pertainingto the spontaneous brain fluctuations (canonicalcomponent) and the other correlated to the per-ception of the subject. This inference seems to besupported by the fact that this spontaneous andasymmetric activity occurs at frequencies that sta-tistically have been found to be close to the meanfor bistable changes. Frequency of shifting of thespinning dancer studied in a group of subjects hasa mean of 3.68 per minutes (Troje & McAdam,2010), i.e., 0.061 Hz—a number strikingly close tothe mean of the very low spontaneous fluctuations,characterized between 0.01 and 0.1 Hz (Fransson,2005). This indicates that for a dissociated compo-nent like the fronto-parietal component, it shouldbe turned on 2–4 times per minute. Thus, corticalspontaneous fluctuations and switching of percep-tion of the kinetic bistable images share the samefrequency domain.
LIMITATIONS OF OUR WORK AND POSSIBLEPITFALLS
Since our work is based on only one subject, itcannot be generalized for the normal population.Nevertheless, we took advantage of being able toexamine an educated volunteer who has the abil-ity of presenting several spontaneous switchingslong enough to allow a block design paradigm.More importantly, this volunteer developed theability to switch and sustain the phase of percep-tion at will, allowing more detailed explorationwith conventional fMR methods as a sort ofintrasession/intersubject validation of the results.We are confident of the validity of our results, asthey seem to be widely supported by previous find-ings from other techniques exploring similar visualillusions.
The method of exploring a specific function inonly one subject demands a very reliable individ-ual, in particular in those tasks where the out-put is entirely subjective. The authors believe thatalthough there is a quite important limitation ofhaving only one subject in the study, it does notinvalidate the findings provided a solid methodol-ogy is applied and the subject is trustable. At least,this study may serve as a guide for further studies in
groups aimed to verify, or further characterize, ourfindings.
The motor response may add cognitive noise.However, since this response is equally present forprompting the on and off epochs, we believe itseffect is cancelled out and should be of no sig-nificance. Actually, we did not obtain hand motoractivation in our processing.
The partial variability of the resting-state brainfluctuations poses some questions about their ori-gin and significance; however, it does not seem toimpact the assumptions on the presence, frequency,and dissociation of the occipito-parieto-frontalcomponents that we have linked to the explana-tion of SDI. Indeed, these components are the same“RSN4” component described recently as one ofthe seven more stable components in a study ofintra- and intersubject stability of the resting-statenetworks (Bellec, Rosa-Neto, Lyttelton, Benali, &Evans, 2010).
Our results are of importance in differentfields beyond those of neuropsychology andneurophysiology of the top-down regulation ofvisual perception. It also sheds lights on the neu-ral bases of intersubject variability of perceptionand its meaning, the implications in interhumanvariability of subjectivity, and the appreciation ofmoral and ethical values, as they are studied by newavenues of human knowledge as neurophilosophy(Churchland & Churchland, 2002) and the neuro-biology of consciousness (da Silva, 2004; Young &Pigott, 1999).
CONCLUSIONS
Our results show that the phase of perception inkinetic bistable stimulus is related to the sponta-neous activation of a circuitry involving the rightfronto-parietal dorsal areas and that this circuitryswitches spontaneously with the same frequency asthe subject experiences the perception switches.
Original manuscript received 7 December 2012Revised manuscript received 28 Jun 2013Revised manuscript accepted 8 July 2013
First published online 24 August 2013
REFERENCES
Bellec, P., Rosa-Neto, P., Lyttelton, O. C., Benali, H., &Evans, A. C. (2010). Multi-level bootstrap analysis ofstable clusters in resting-state fMRI. NeuroImage, 51,1126–1139. doi: 10.1016/j.neuroimage.2010.02.082
Churchland, P. S., & Churchland, P. M. (2002). Neuralworlds and real worlds. Nature Reviews. Neuroscience,11, 903–907. doi:10.1038/nrn958
Corney, D., & Lotto, R. B. (2007). What are lightnessillusions and why do we see them? Plos ComputationalBiology, 3, 1790–1800.
da Silva, F. H. (2004). Contribution to a neurophysiologyof consciousness. Supplements to ClinicalNeurophysiology, 57, 645–655.
Deutschländer, A., Bense, S., Stephan, T., Schwaiger, M.,Brandt, T., & Dieterich, M. (2002). Sensory systeminteractions during simultaneous vestibular and visualstimulation in PET. Human Brain Mapping, 16(2),92–103. PubMed PMID: 11954059.
Dieterich, M., Bauermann, T., Best, C., Stoeter, P., &Schlindwein, P. (2007). Evidence for cortical visualsubstitution of chronic bilateral vestibular failure(an fMRI study). Brain, 130(8), 2108–2116. doi:10.1093/brain/awm130
Dupont, P., Orban, G. A., De Bruyn, B., Verbruggen,A., & Mortelmans, L. (1994). Many areas in thehuman brain respond to visual motion. Journal ofNeurophysiology, 72(3), 1420–1424.
Forster, B. B., MacKay, A. L., Whittall, K. P., Kiehl,K. A., Smith, A. M., Hare, R. D., & Liddle, P.F. (1998). Functional magnetic resonance imaging:The basics of blood-oxygen-level dependent (BOLD)imaging. Canadian Association Radiology Journal, 49,320–329.
Fortin, A., Ptito, A., Faubert, J., & Ptito, M.(2002). Cortical areas mediating stereopsis in thehuman brain: A PET study. NeuroReport, 13(6),895–898.
Fransson, P. (2005). Spontaneous low-frequencyBOLD signal fluctuations: An fMRI investigationof the resting-state default mode of brain func-tion hypothesis. Human Brain Mapping, 1, 15–29.doi:10.1002/hbm.20113
Freeman, E. D., Sterzer, P., & Driver, J. (2012).fMRI correlates of subjective reversals in ambiguousstructure-from-motion. Journal of Vision, 12, 35–40.doi:10.1167/12.6.35
Friston, K. J., Holmes, A. P., Worsley, K. J., Poline, J. P.,Frith, C. D., & Frackowiak, R. S. J. (1994). Statisticalparametric maps in functional imaging: A general lin-ear approach. Human Brain Mapping, 2, 189–210. doi:10.1002/hbm.460020402
Gregroy, R. L. (1997). Knowledge in perception and illu-sion. Philosophical Transactions of the Royal SocietyB, 352, 1121–1128.
Hock, H. S., Schöner, G., & Voss, A. (1997). The influ-ence of adaptation and stochastic fluctuations onspontaneous perceptual changes for bistable stimuli.Perception & Psychophysics, 59, 509–522.
Hsieh, P. J., Caplovitz, G. P., & Tse, P. U. (2006).Bistable illusory rebound motion: Event-relatedfunctional magnetic resonance imaging of percep-tual states and switches. NeuroImage, 32, 728–739.doi:10.1016/j.neuroimage.2006.03.047
Inui, T., Tanaka, S., Okada, T., Nishizawa, S., Katayama,M., & Konishi, J. (2000). Neural substrates fordepth perception of the Necker cube; a func-tional magnetic resonance imaging study in humansubjects. Neuroscience Letters, 282, 145–148. doi:10.1016/S0304-3940(00)00899-5
Jenkinson, M., Bannister, P., Brady, M., & Smith, S.(2002). Improved optimization for the robust andaccurate linear registration and motion correctionof brain images. NeuroImage, 17, 825–841. doi:10.106/nimg.2002.1132
Jenkinson, M., & Smith, S. A. (2001). Global optimi-sation method for robust affine registration of brainimages. Medical Image Analysis, 5, 143–156. doi:10.1016/S1361–8415(01)00036–6
Kamitani, Y., & Tong, F. (2006). Decoding seen andattended motion directions from activity in the humanvisual cortex. Current Biology, 16, 1096–1102. doi:10.1016/j.cub.2006.04.003
Kanai, R., Bahrami, B., & Rees, G. (2010). Humanparietal cortex structure predicts individual differ-ences in perceptual rivalry. Current Biology, 20(18),1626–1630. doi:10.1016/j.cub.2010.07.027
Knauff, M., Mulack, T., Kassubek, J., Salih, H. R.,& Greenlee, M. W. (2002). Spatial imagery indeductive reasoning: A functional MRI study. BrainResearch Cognitive Brain Research, 13, 203–212.doi:10.1016/S0926-6410(01)00116-1
Kohler, A., Haddad, L., Singer, W., & Muckli, L.(2008). Deciding what to see: The role of inten-tion and attention in the perception of appar-ent motion. Vision Research, 48, 1096–1106. doi:10.1016/j.visres.2007.11.020
Kornmeier, J., & Bach, M. (2006). Bistable perception—along the processing chain from ambiguous visualinput to a stable percept. International Journal ofPsychophysiology, 62, 345–349.
Kuriki, I., Ashida, H., Murakami, I., & Kitaoka, A.(2008). Functional brain imaging of the RotatingSnakes illusion by fMRI. Journal of Vision, 8(10),1–10. doi:10.1167/8.10.16
Lapeyre-Mestre, M., Gary, J., Machelard-Roumagnac,M., Bonhomme, C., Bugat, R., & Montastruc, J. L.(1997). Incidence and cost of adverse drug reactions ina French cancer institute. European Journal of ClinicalPharmacology, 53, 19–22.
Larsson, J., Landy, M. S., & Heeger, D. J. (2006).Orientation-selective adaptation to first- andsecond-order patterns in human visual cortex.Journal of Neurophysiology, 95, 862–881. doi:10.1152/jn.00668.2005
Le, T. H., Pardo, J. V., & Hu, X. (1998). 4 T-fMRIstudy of nonspatial shifting of selective attention:Cerebellar and parietal contributions. Journal ofNeurophysiology, 79, 1535–1548.
Liu, C. H., Tzeng, O. J., Hung, D. L., Tseng, P., &Juan, C. H. (2012). Investigation of bistable percep-tion with the “silhouette spinner”: Sit still, spin thedancer with your will. Vision Research, 60, 34–39. doi:10.1016/j.visres.2012.03.005
Maldjian, J. A., Laurienti, P. J., Kraft, R. A., &Burdette, J. H. (2003). An automated methodfor neuroanatomic and cytoarchitectonic atlas-based
Dow
nloa
ded
by [
Dal
hous
ie U
nive
rsity
] at
11:
24 0
3 Se
ptem
ber
2013
THE ORIGIN OF THE SPINNING DANCER ILLUSION 13
interrogation of fmri data sets. NeuroImage, 19,1233–1239. (WFU Pickatlas, version 3.0.4.)
Owen, A. M., Downes, J. J., Sahakian, B. J., Polkey, C. E.,& Robbins, T. W. (1990). Planning and spatial work-ing memory following frontal lobe lesions in man.Neuropsychologia, 28, 1021–1034.
Ozaki, T. J., Sato, N., Kitajo, K., Someya, Y.,Anami, K., Mizuhara, H., . . . Yamaguchi, Y. (2012).Traveling EEG slow oscillation along the dor-sal attention network initiates spontaneous percep-tual switching. Cognitive Neurodynamics, 6, 185–198.doi:10.1007/s11571-012-9196-y
Pastukhov, A., & Braun, J. (2007). Perceptual reversalsneed no prompting by attention. Journal of Vision, 7,1–17.
Pastukhov, A., Vonau, V., & Braun, J. (2012). Believablechange: Bistable reversals are governed by phys-ical plausibility. Journal of Vision, 12, 17–20.doi:10.1167/12.1.17
Petersik, J. T., & McDill, M. (1981). A new bistablemotion illusion based upon “kinetic optical occlu-sion”. Perception, 10, 563–572.
Pitts, M. A., Martínez, A., Stalmaster, C., Nerger, J. L.,& Hillyard, S. A. (2009). Neural generators of ERPslinked with Necker cube reversals. Psychophysiology,46, 694–702. doi: 10.1111/j.1469–8986.2009.00822.x
Poston, T., & Stewart, I. (1978). Nonlinear model-ing of multistable perception. Behavioral Science, 23,318–334.
Raemaekers, M., van der Schaaf, M. E., van Ee,R., & van Wezel, R. J. (2009). Widespread fMRIactivity differences between perceptual statesin visual rivalry are correlated with differencesin observer biases. Brain Research, 3, 161–171.doi:10.1016/j.brainres.2008.11.078
Raz, A., Lamar, M., Buhle, J. T., Kane, M. J., &Peterson, B. S. (2007). Selective biasing of a specificbistable-figure percept involves fMRI signal changesin frontostriatal circuits: A step toward unlockingthe neural correlates of top-down control and self-regulation. The American Journal of Clinical Hypnosis,50, 137–156.
Rosazza, C., & Minati, L. (2011). Resting-statebrain networks: Literature review and clinicalapplications. Neurological Sciences, 32, 773–785. doi:10.1007/s10072-011-0636-y
Rowe, D. B., & Hoffmann, R. G. (2006). Multivariatestatistical analysis in fMRI. IEEE Engineering inMedicine and Biology Magazine, 25, 60–64.
Shen, L., Zeng, Z. L., Huang, P. Y., Li, Q., Mu, J., Huang,X. Q., . . . Xie, P. (2009). Temporal cortex participatesin spontaneous perceptual reversal. NeuroReport, 20,647–651. doi: 10.1097/WNR.0b013e32832974a8
Slotnick, S. D., & Yantis, S. (2005). Common neu-ral substrates for the control and effects ofvisual attention and perceptual bistability. BrainResearch Cognitive Brain Research, 24, 97–108. doi:10.1016/j.cogbrainres.2004.12.008
Smith, S. M. (2002). Fast robust automated brain extrac-tion. Human Brain Mapping, 17, 143–155.
Sterzer, P., Eger, E., & Kleinschmidt, A. (2003).Responses of extrastriate cortex to switchingperception of ambiguous visual motion stimuli.NeuroReport, 14, 2337–2241.
Sterzer, P., Russ, M. O., Preibisch, C., & Kleinschmidt,A. (2002). Neural correlates of spontaneousdirection reversals in ambiguous apparentvisual motion. NeuroImage, 15, 908–916.doi:10.1006/nimg.2001.1030
Suzuki, S., & Peterson, M. A. (2000). Multiplicativeeffects of intention on the perception of bistableapparent motion. Psychological Science, 11,202–209.
Ta’eed, L. K., Ta’eed, O., & Wright, J. E. (1988).Determinants involved in the perception of the Neckercube: An application of catastrophe theory. BehavioralScience, 33, 97–115.
Troje, N., & McAdam, M. (2010). The viewing-from-above bias and the silhouette illusion. i-Perception, 1,143–148.
Watanabe, K., & Shimojo, S. (1998). Attentional modula-tion in perception of visual motion events. Perception,27, 1041–1054.
Young, G. B., & Pigott, S. E. (1999). Neurobiologicalbasis of consciousness. Archives of Neurology,56, 153–157. doi: 10-1001/pubs.ArchNeurol.-ISSN-0003-9942-56-2-nnr7524
Zaretskaya, N., Thielscher, A., Logothetis, N.K., & Bartels, A. (2010). Disrupting parietalfunction prolongs dominance durations in binoc-ular rivalry. Current Biology, 20, 2106–21111.doi:10.1016/j.cub.2010.10.046