The Ultrastructure of Spermatozoa of the Squamata (Reptilia) with PhylogeneticConsiderations Barrie G. M. fnutnsox x Zoology Department,University of Queensland, Brisbane, Q 407 2, Australia ABSTRACT Comparativeultrastructure of squamate families is reviewed, with new data for the South African chamaeleon, Bradypodion karroicum. Parsimony analysis is conducted, using Chelonia as the outgroup and branch and bound searching. Two major spermatozoal autapomorphies for the Squamata are extension of the fibrous sheath into the midpiece and (not computed) the paracrystalline subacrosomal cone.Further synapomorphies defining the Squamata sensustrictu are a single perforatorium in place of the two or three of Sphengdontida and Chelonia;loss tlf the endonuclear canal; presenceof sinuous mitochondria (possibly an artefactual parsimony resolution as a columnar form is intuitively preferred);intermitochondrial location of densebodies (mitochondrialtransformations); presence of a well developed epinuclear electron lucent region and,equivocally, arrangement of the dense bodies as periodicrings. A major inference is polyphyly of the 'Sauria', the Scincomorpha and the Scincidae. Sphenomorphus group and egernidskinks show no close relationship to Eugongylus-group skinks which form the sister-group of the pygopodid Lialis. Snakes are the sister-group of the Eugongylus+pygopod clade. Gekkonidae appearto be a relatively plesiomorphicgroup, separated by several families from the Pygopodidae. The Iguania is not a monophyletic assemblage, iguanids andPogona occur in the same clade but Pogona appears to be the sister-taxon of Varanus. Another iguanian,the chamaeleon Bradypodion, bas an unresolved relationship with the gekkonid+snake+pygopodid+Eugongylus-clade. Sphenomorph and egernid skinks form an unresolved cladewith Chalcides and lacertids but linkageof lacertids with the Teiidae in the Lacertoidea is not upheld. Pending further investigations of a largernumber of taxa,these results can only be considered heuristic. nEsuuE L'ultrastructure des spermatozoides des Squamata (Reptilia) et consid6rations phylog6niques Ce chapitre comprendune synthbse de I'ultrastructurecomparde des familles de Squamata, avec des observations nouvelles sur le Cam6l6on d'Afrique du SudBradypodion karroicum, et une analyse de parcimonie utilisant les Chelonia commeout$oup et une recherche par branch and bound.Les deux autapomorphies majeures des spermatozoides pour les Squamata sont I'extensionde la gaine fibreusedansla pidce interm6diaire et (n'intervenant pas dans le calcul) le c6ne subacrosomien paracristallin. D'autres synapomorphies d6finissantles Squamata sensu stictu sont un perforatorium uniquei la placede deux ou trois chezles Sphenodontida et Ies Chelonia; la pertedu canalendonucl6aire; la prdsence de mitochondries sinueuses (ce qui pourrait donner lieu d un artefact de rdsolutioncar la forme en colonneest pr6f6r6e de manidreintuitive); la position intermitochondriale des corps denses (transformations mitochondriales); la pr6sence d'une r6gion 6pinucl6aire claire aux dlectrons bien d6veloppde, et, de manidre6quivoque, la dispositiondes corps denses en anneaux p6riodiques, Une cons6quence majeure est la polyphylie des "Sauria", des Scincomorpha et des Scincidae. Le groupe de Sphenomorphus et les Scincidae ergenides ne montrent pasde relations proches avecles Scincidae du groupe de Eugongylus, qui forment le groupe frbre du pygopodide Lialis. Les Serpentssont le groupe-frdre du clade JAMIESoN, B. G. M., 1995. - The ultrastructure of spermatozoa of the S[uamata (Reptilia).with phylogenetic considerations. 1z: JeMrssoN, B. G. M., Auslo, J., & JusrIlre, J.-L. (eds),Advancbs in Spermatozoal Phylogeny and Taxonomy. Mdm. Mus. natn. Hist. nat.,166: 359-383. Paris ISBN : 2-85653-225-X.
Transcript
The Ultrastructure of Spermatozoa of the Squamata(Reptilia) with Phylogenetic Considerations
Barrie G. M. fnutnsox
x Zoology Department, University of Queensland,Brisbane, Q 407 2, Australia
ABSTRACT
Comparative ultrastructure of squamate families is reviewed, with new data for the South African chamaeleon,Bradypodion karroicum. Parsimony analysis is conducted, using Chelonia as the outgroup and branch and boundsearching. Two major spermatozoal autapomorphies for the Squamata are extension of the fibrous sheath into the midpieceand (not computed) the paracrystalline subacrosomal cone. Further synapomorphies defining the Squamata sensu strictuare a single perforatorium in place of the two or three of Sphengdontida and Chelonia; loss tlf the endonuclear canal;presence of sinuous mitochondria (possibly an artefactual parsimony resolution as a columnar form is intuitivelypreferred); intermitochondrial location of dense bodies (mitochondrial transformations); presence of a well developedepinuclear electron lucent region and, equivocally, arrangement of the dense bodies as periodic rings. A major inference ispolyphyly of the 'Sauria', the Scincomorpha and the Scincidae. Sphenomorphus group and egernid skinks show no closerelationship to Eugongylus-group skinks which form the sister-group of the pygopodid Lialis. Snakes are the sister-groupof the Eugongylus+pygopod clade. Gekkonidae appear to be a relatively plesiomorphic group, separated by severalfamilies from the Pygopodidae. The Iguania is not a monophyletic assemblage, iguanids and Pogona occur in the sameclade but Pogona appears to be the sister-taxon of Varanus. Another iguanian, the chamaeleon Bradypodion, bas anunresolved relationship with the gekkonid+snake+pygopodid+Eugongylus-clade. Sphenomorph and egernid skinks forman unresolved clade with Chalcides and lacertids but linkage of lacertids with the Teiidae in the Lacertoidea is not upheld.Pending further investigations of a larger number of taxa, these results can only be considered heuristic.
nEsuuEL'ultrastructure des spermatozoides des Squamata (Reptilia) et consid6rations phylog6niques
Ce chapitre comprend une synthbse de I'ultrastructure comparde des familles de Squamata, avec des observationsnouvelles sur le Cam6l6on d'Afrique du Sud Bradypodion karroicum, et une analyse de parcimonie utilisant les Cheloniacomme out$oup et une recherche par branch and bound. Les deux autapomorphies majeures des spermatozoides pour lesSquamata sont I'extension de la gaine fibreuse dans la pidce interm6diaire et (n'intervenant pas dans le calcul) le c6nesubacrosomien paracristallin. D'autres synapomorphies d6finissant les Squamata sensu stictu sont un perforatoriumunique i la place de deux ou trois chez les Sphenodontida et Ies Chelonia; la perte du canal endonucl6aire; la prdsence demitochondries sinueuses (ce qui pourrait donner lieu d un artefact de rdsolution car la forme en colonne est pr6f6r6e demanidre intuitive); la position intermitochondriale des corps denses (transformations mitochondriales); la pr6sence d'uner6gion 6pinucl6aire claire aux dlectrons bien d6veloppde, et, de manidre 6quivoque, la disposition des corps denses enanneaux p6riodiques, Une cons6quence majeure est la polyphylie des "Sauria", des Scincomorpha et des Scincidae. Legroupe de Sphenomorphus et les Scincidae ergenides ne montrent pas de relations proches avec les Scincidae du groupe deEugongylus, qui forment le groupe frbre du pygopodide Lialis. Les Serpents sont le groupe-frdre du clade
JAMIESoN, B. G. M., 1995. - The ultrastructure of spermatozoa of the S[uamata (Reptilia).with phylogeneticconsiderations. 1z: JeMrssoN, B. G. M., Auslo, J., & JusrIlre, J.-L. (eds), Advancbs in Spermatozoal Phylogeny andTaxonomy. Mdm. Mus. natn. Hist. nat., 166: 359-383. Paris ISBN : 2-85653-225-X.
resultats peuvent seulement €tre consid6r6s comme treurisuqulJ. d'autres 6tudes sur un plus grand nombre de taxons' ces
There is no comprehensive well-corrobor:AS the utility Of spermatoz1Al ultrastructure aS a SOurCe OI CnaraCrcrs Ior pnyruBeueuu 'm<uJDrD rD
well established 126, Z'l , 3l,32\,JAMI#6N it ot. l}ll and OLIVER et aI. t45l attgmpted to shed
lieht on squamate ;laisification ana pfryfogeni bi a comparative study of spermatozoal
uilrastructure which is reviewed and extended here'The ultrastructure of spermatozoa or spermiogenesis has been studied, though often
cursorily, in the -"loigtoops'of-sqr-amalg rJpiil.es. fidUes studied are: Scincidae [8' 14'2I'
g, ti, tot ,r7,18:451; -Chamaeleonidae [52(spermiogenesis onlyJl'anO'tnlJaccountl: Vaianidae [45]; Gekkonidae 12I,3'1, 461,: and
i$gliiiji?"f ps,-iii.'nbrief account of ifie kinetic a_pparatus of the sperm of Amphislg:ltyahiiiiii (p*tprriiraeiiiaael and Liolaeius weigmanii (i[aanidae) by Sorql]o & CEN6z [5o] is.r".ny "inirt6ritinterest a;d witt not be review;d here. fhe tp?ll1]o?Tl*_tp,:i"^:1"1," 1"".t,r,r,"" ih.rr.rushlv investisated. in terms of taxa examined, than those of other squamates I l, J' ),
6' 20' 21' 24' 33' 45' 46' 491' rides the first description of matureThe present account reviews squamate sperm' pro\
chamaeleoiria rp"r-, ioittr"t*. Soutli African ihamaeieon Bradypodion karroicum' and gives a
preliminary parsimony analYsis.
MATERIALS AND METHODS
Testes and ducts were dissected from a euthanased specimen of Bradypodion karroicum (Chamaeleonidae)'
processing of the tissues was as in t341. A cradistic analysis was performed using the PAUP program of swoFpono [51]
(for details see parsimony analysis).
360 B. G. M.IAMIESON : SQUAMATA (REPTILIA)
RESULTS AND DISCUSSION
C omparativ e sperrn ultrastructureSpermatozoal ultrastructure in the Squamata is summarized in Tables 1 and 2.
tes of the Scincidae include a description of the
Frc. 1. - A generalized spermatozoon (diagrammatic), in longitudinali'ind corresponding transverse sections, of tb
Sphenomorpho. gtoop of the Scincidae (Nangura spinosa,Ctenotus taeniolatus,C' robustus,Anomalopu
virreauxii)ind Egernii-group (Tiliqua scincoides scincoides). Scales of various components are only approxim*.
Reeional zonation-of the acrosome vesicle is shown only for the transverse sections. After [34' 37].
ADVANCES IN SPERMA'TOZOAL PHYI,OGENTY AND TAXONOMY
materialslructure
prcxmalcentnole
nng structure 4
mitochondrion 4
361
Enilrii*@ilIilIffi
I 0.5.mI l-!------{
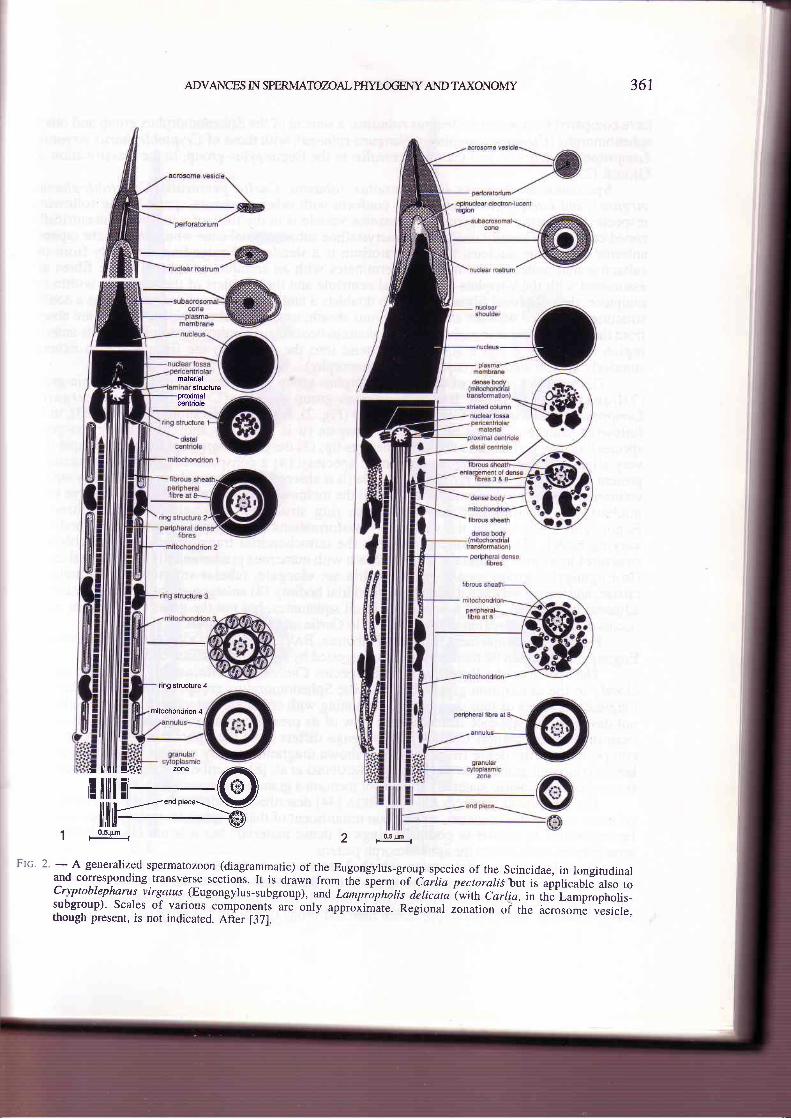
Ftc' 2' - A generalized spermatozoon (diagrammatic) of the Eugongylus-group species of the Scincidae, in longitudinaland corresponding transverse sections. It is drawn from tTre ip...n-or tariia pectoralrj,but is applicable also toCryptoblepharus virSLtus (Eugongylus-subgroup), and Lampipholis delicata (with Carlla, in the Lampropholis-subgroup)' Scales of various components are only upproii.it". Regional ̂ nuti* oi ihe acrosoml vlsicle,rhough present, is not indicated. After [371.
t 0 . 5 u m4 -
362 B. G. M. JAMIESON i SQUAMATA (REPTILIA)
have compared the sperm of Ctenotus robustus, a scincid of the Sphenomorphus group and othersphenomorphs (Ctenotus taeniolatus, Nangura spinosa), with those of Cryptoblepharus virgatus,,Lampropholis delicata, and Carlia pectoralis, in the Eugongylus-group, in the classification ofGREER [23].
Spermatozoa of skinks (e.g. Ctenotus robustus, Carlia pectoralis, Cryptoblepharusvirgatis, and Lampropholis delicata) conform with other squamate sperm in the followingrespects: the sperm are filiform; the acrosome vesicle is in the form of a hollow, concentricallyzoned cone which basally overlies a paracrystalline subacrosomal cone which invests the taperedanterior end of the nucleus; the perforatorium is a slender rod extending anteriorly from thesubacrosomal material; the midpiece terminates with an annulus; peripheral dense fibres areassociated with the 9 triplets of the distal centriole and the doublets of the axoneme within themidpiece; the peripheral fibres adjacent to doublets 3 and 8 are enlarged and each forms a doublestructure associated with the annulated fibrous sheath; usually all nine peripheral fibres are absentfrom the principal piece thoughin l-ampropholis delicata they remain well developed in its anteriorregion and fibres 3 and 8 sometimes extend into the endpiece; the fibrous sheath extendsanteriorly into the midpiece (squamate autapomorphy).
The sperm of species of the Sphenomorphus group (e.9. Ctenotus) and the Egernia-group(Tiliqua) (Fig. 1) differ from Eugongylus-group species (Cryptoblepharus virgatus,Lampropholis delicata and Carlia pectoralis) (Fig. 2), in the classification of GnsBR [23], in thefollowing features: (1) the acrosome is elongate (it is relatively short in Eugongylus-groupspecies); (2) the acrosome is depressed near its tip; (3) the perforatorium is strongly oblique (it isvery slightly oblique in Eugongylus-group species); (4) a conspicuous laminated structure ispresent on each side of the proximal centriole (it is absent, though possibly represented by striatedcolumn(s) in Eugongylus-group species); (5) the midpiece is shorter absolutely and relative to thenucleus; (6) the midpiece has four dense ring structures in longitudinal succession (inEugongylus-group species mitochondrial transformations are scattered irregular dense bodies ofvarying sizes); (7) mitochondria between the mitochondrial transformations form columnarstructures in a circle around the fibrous sheath with numerous predominantly longitudinal cristae(in Eugongylus-group species mitochondria are elongate, tubular structures, with indistinctcristae, and weave between the intermitochondrial bodies); (8) enlargement of the peripheral fibresadjacent to doublets 3 and 8 occurs, as in all squamates, but not the gross enlargement whichoccurs in the anterior region of the axoneme in Carlia and Lampropholis.
From microcomplement fixation of albumin, BAvnRstocK & DotwsLI-AN [4] showed theEugongylus-group to be monophyletic as suggested by sperm ultrastructure [35].
The sperm of the European scincid species Chalcides ocellatus tiligugu [21] conformsclosely to the description given above for the Sphenomorphus group, particularly in having alongitudinal series of four dense rings alternating with columnar mitochondria. An annulus wasnot described but absence is doubtful in view of its presence in all squamates which have beenexamined by the author. Ch. ocellatus tiligugu differs from the Sphenomorphus group in thecomposition of the dense rings which are shown diagrammatically as each being composed oflarge juxtaposed granules in single file. CRRcupINo et al. [8] describe the ring structures as "four
rings of electron-dense material" and do not mention a granular composition.For the scincid Eumeces laticeps, Orre [44] described a midpiece with nine mitochondrial
columns around the axoneme, a condition reminiscent of that in sphenomorphs. These "columns
lie segmented by partial or complete rings of dense material" but it is not clear whether theirarrangement conforms to the sphenomorph pattern.
A suite of character states which JAMIESoN & ScHELTINGA [35] noted for theSphenomorphus and Egernia-groups of the Scincidae .is also seen in the teiid lizard.Cnemidophorus sexlineatus l4l,42l (see below): 1. Anterior @epression of the acrosome. 2. Theconical nuclear fossa containing dense material projecting from the proximal centriole. 3. Tbc
ADVANCES IN SPERMATOZOAL PHYLOGENY AND TAXONOMY 363
p la le
oens(y
lossa
proxrmal cenlriole
'dense body(milochondrial
frtI 'l .
': : barbata (Agamidae)' A diagrammatic representation of the spermatozoon, in longitudinal and--::s transverse sections. After [37].
-^ e'v uvvr'r' ':'tn karroicum (chamaeleonioaej. A diagrammatic representation of the-'. ':drng lransverse sections. Orieinal.
---o-<'r''g!rv rvPrwrv'r4uuu ur tne spermatozoon, in longitudinal
364 B. G. M. JAMIESON : SQUAMATA (REPTILIA)
presence of a laminar structure extending from the pericentriolar apparatus (possibly unilateral inthe teiid, and possibly homologous with the striated column(s) in Eugongylus-group sperm). 4.Presence of four, or in Cnemidophorus five, intermitochondrial rings alternating with columnarmitochondria. 5. The absence of sharply detined nuclear shoulders. 6. The apparent wideseparation of the plasma membrane from the fibrous sheath in the anterior region of the principalpiece. However, features 3 and 6 require confirmation for teiids.
The parsimony analyses (below) which are necessarily preliminary in view of our imperfectknowledge of these and other characters across the spectrum of squamate taxa, strongly suggestthat skinks are not monophyletic and that sphenomorph-egernid and Eugongylus-group skinks aretwo distinct groups of squamates, the Eugongylus-group appearing to be the sister-group ofpygopods (Fig. 10). As snakes form the sister-taxon of these two groups, it would appear that thelegless condition has been independently derived in snakes as compared with pygopods, thoughas a parallelism from an ancestor shared also with Eugongylus-group skinks.
Lacertidae. Spermatozoa of the Lacertidae have been examined ultrastructurally rn Lacertasicula campestris, L. lepida lepida, L. Iaevis, L. viridis and Algyroides alleni [21], L. vivipara[11] and, with reference to development of the sperm head, Podarcis (=[,qsgrta) taurica [7]. Theyare morphologically very similar and resemble those of the sphenomorph-egernid Scincidae inmany respects [21]. The head is curved and depressed [7, ll,2ll. The perforatorium ("apicalgroove") is oblique and the subacrosomal cone is paracrystalline U, 2Il as is typical ofsquamates. The midpiece is distinctive in having only two sets of dense bodies, the first has theappearance of two opaque masses on each side immediately posterior to the nucleus [21] andtermed the "nuclear plate" [11]; this is probably correctly regarded by NEwToN and TRAUTH [42]as the equivalent of the first ring structure in teiid sperm. The second is a large ring (morecertainly equivalent to ring structures of sphenomorph skinks) and questionably regarded as a"chromatoid body" [11], between the mitochondria in the distal third of the midpiece. InL. vivipara, therc are only three tiers of mitochondria in longitudinal section and theintermitochondrial ring lies between mitochondrial tiers 2 and 3 [11]. The appearance and size ofthe dense bodies varies from species to species but not clearly enough to be used as a definingcharacteristic. The internal surface of the posterior ring is indented by numerous niches containingsmall masses of moderately opaque material t211. In the order of 10 mitochondria are illustrated intransverse section for L. lepida [21] but only five in L. vivipara ll1l. Though said to befilamentous [21], the mitochondria appear compact and only a few times longer than wide, witblinear cristae. Peripheral dense fibres at 3 and 8 are enlarged and fused with or adpressed to thefibrous sheath [�ll,2I).In mature sperm no annulus is indicated diagrammatically but in emicrograph a very small, weakly developed annulus appears to be presentl2Il, and it is preseuin the late spermatid [11]. There are two centrioles and the implantation fossa is compact androunded lIl,2ll.
Teiidae. The subacrosomal rod (putative perforatorium) has been shown tn CnemidophoroIemniscatus lemniscatus to develop from a subacrosomal granule [19]. The spermatozoon d
FIG. 5. - Bradypodion kanoicum (Chamaeleonidae). A, B, C: Longitudinal sections (LS) of acrosome andregion of nucleus. D' E: LS of posterior region of nucleus and anterior region of midpiece. F-M:transverse sections through F-J, the acrosome, K, the nucleus, L, the midpiece and M, the principal piea- tcentriolar region and midpiece. 0: LS posterior region of midpiecq,-including annulus, and anterior rGltprincipal piece. a = acrosome vesicle; an = annulus; db = dense body (mitochondrial transformation): & =cent r io le ; e t=e lec t ron lucentspace; fs= f ib roussheath ; m=mi tochondr ia ; n -nuc leus ;n f=nuc lear fq ,perforatorium; pc = proximal centriole; pf = peripheral dense fibre (coarse fibre); pm = plasma memhrg, Isubacrosomal cone. Original. Scale bar I gm,
ADVANCES IN SPERMATOZOAL PHYLOGENY AND TAXONOMY
p
et
3C
nr
365
fs
M
366 B. G. M. JAMIESON : SQUAMATA (REPTILIA)
C. sexlinealzs has been the subject of detailed description 141,421, augmented here by referenceto published micrographs. A suite of characters shared with the Sphenomorphus and Egernia-group Scincidae is discussed under that family, above.
In C. sexlineatus, the acrosome caps and invests the anterior I pm of the nucleus whichforms a pointed nuclear rostrum. The acrosome is depressed and spatulate, as in Sphenomorphusand Egernia-group scincids. Although an acrosome vesicle is recognized, no subacrosomal coneis described. Instead, a large vesicle and a more basal small vesicle are described posterior to themain part of the acrosome vesicle. From a micrograph there seems some possibility that the largeand small vesicles represent subacrosomal material. It appears that a basal extension of theacrosome vesicle envelops this material, as in typical squamate and, indeed, tetrapodspermatozoa. This interpretation is supported by presence of an electron dense connectivebetween the posterior acrosome and the cell membrane. A well developed rodlike perforatorium,composed of longitudinal fibres, possibly in helical Tnay, is present. It does not appear to beoblique but is unusual in being strongly eccentric. A narrow epinuclear electron lucent zone isvisible anterior to the rostrum. The nucleus is curved and strongly condensed and has a low,conical, basal implantation fossa. The absence of sharply defined nuclear shoulders at the base ofthe rostrum is a resemblance to the above-mentioned scincids. The midpiece consists of five tiers,usually, of shortly columnar mitochondria separated by intermitochondrial dense "ring structures"and terminating with an annulus, giving the formula rsl/mil, rs2lmi2, rs3/mi3, rs4/mi4, rs5/m6,an. Each mitochondrial set consists of 8 to l0 (usually 9) mitochondria arranged around thcaxoneme. The axoneme is surrounded by the fibrous sheath from rs3 to the posterior end of thcprincipal piece,leaving a 0.7 pm endpiece. The A subtubules of the 9+2 axoneme are filled wit[dense material. A transverse section of the distal centriole (basal body) shows coarse fibrElenveloping the triplets, extending both peripherally and internally to each triplet, and, asdisplaced clockwise into the inter-triplet space. Dense material is associated with the twosinglets. Coarse fibres are absent from illustrated transverse sections of the principalLongitudinal extracellular tubules surround the nucleus and flagellum in the ductus deferens.suggested presence of a laminar structure extending from the pericentriolar apparatus, equito the bilateral structure in scincids [35], requires confirmation.
Iguanidae. The account by FunmRI1221, based chiefly on three species of theCupriguanus scapulatus, Phymaturus palluma and Liolaemus austromendocinus, ts shere, using the terminology employed in the present account. The general structurespermatozoon is much as described for skinks, with the following details. The acrorcircular in cross section; the single, well developed perforatorium appears to have asquare, tip and to lack a basal plate but this requires confirmation. An endonuclear canal irThe subacrosomal cone ("inner cap") is paracrystalline and surrounds a pointed nuclertwhich is preceded by an epinuclear electron lucent region. The nucleus iscondensed chromatin, and has a shallow basal fossa which houses the anteriorproximal centriole; this is at an angle to the distal centriole. There are distinct nuclearapparently intermediate in shape between the sharp and rounded forms. The midpiece bshort, and, with a length of 7 pm, is much shorter than the head. Although the miowhich are long, thin and numerous, are arranged in a regular circlet around the arcfibrous sheath, they form helices, with a regular arrangement in C. scapulatus ag|lL. austromendocinus. They have a very fine calibre tn L. austromindocinus.reminiscent of those of snake sperm. The quantity and arrangement ofintermitochondrial bodies are characteristic of each of the three species. Inmitochondrial sleeve is subdivided into six long sections, separated by contiguousof opaque material arranged in a ring which is sometimes nottompleteiy closed Ais present at the beginning of the mitochondrial sleeve and a seventh, muchconsidered to be the annulus) closes the sleeve, giving the formula, in the system
ADVANCESINSPERMATOZOALPHYLOGENYANDTAXONOMY 367
TRauTr 1421,of rsl/mil, rs2/mi2,rs3/mi3, rs4/mi4,rs5/mi5, rs6/mi6,an. In cupriguanus,thedense material i, $o.t".scal1ty and'does not seem to form closed rings. In Liolaemus, thedensebodies form small dark plales which are few in-nu-u"r and are distributed without order.Development of nine peripheral axonemal fibres is limiied to the rrrori i"gion of the midpieceanterior to rhe fibrouslheattr. wittrin the fibrous,h.;ih ;h;;;#;;;: Lnarged fibres andthese are joined to the sheath. These are said to lie aiJoubilr:'";;'i ffiii. micrographs anddiagram indicate that they are in the usual poritlo" uAJ"r*t to doublets 3 and g.Anolidae' The account of spermiogenesis in itre so-called American chamaeleo n, Anoliscarolinensis (Anolidae) t10l revials no-characters wnlcn can conclusively be ascribed to themature spermatozoon. However, the spermatid has the uruur u"ioro-"1;;i;i" and subacrosomalmaterial investing the nuclear rostrurir; a condenseo, cuived nucleus; ;l;;, dome shaped butapically pointed iirplantation i";;;;;d uproii-ur.inirior. tilted relative to the disral centriole.The fibrous sheath is posterior to the annulus but whetheiit rut"i"*tlnJi i"i" ir," midpiece is notknown, though likely.Tropiduridae' Accounts of spermiogenesis in Tropidurus. torquatus fr2,l3l yield littleinformation with regard to the matui. spermatozoon. Thsfoll;;i"g i"-i":itii derived from textand micrographs. As usual for squamaies, the u.roro-" u"ri"r" iJr"a"iryi"g subacrosomalcone ensheath a tapered anterior'extension or trre nucreus (nuclear rort*"i;. smooth nuclearshoulders support the posterior end of the ;.;;r;;;:Th; ir " r.rg" r"""ri within the nuclearrostrum, near it tip' The spenn head does not appear to be dep-ressed.ihe very strort midpiece hasonlv three gvres-of mito;hondria. iie statem-enr thar rh;Tib;;;; ;;th ll rouno only in theprincipal piece, except for a few dense- spots around the annulus (unlike any other knownsquamate sperm) is to be doubted as the obsbrvation is noi derived r.om a -ature spernatozoon.Peripheral dense fibres are limited to a short r;il;i;;ar the short neck cylinder which liesbetween the proximal centriole and the base of irr" *.i"r;.'Ni"irpl; ;;;pl'"r-ic membranesaround the acrosome tend to disappg.ar by;;u;ity;#;;r rhose in snak6s ulno pygopods appearto persist. No evidence of dense 6ooies or ring it i,.iri", i, provided [13].Agamidae' In,the Agamidae, differeniiation of the sperm head has been described forAsama stetlio [2], uromaitvx phitbyt tllll slr;;;";rylit tbt ini ii6i il;. adramitana trSland, for the nuclear m-anchetle, A. osA;;idi.-uiii"*i;{;;;"of the spenn tail has been describedin uromnstvx phitqj!\71_.-rhe entiie sperm has ue"n oiscitb"df"; pi';;i;Z)iuorotasl Gig. 3).The mature spermatozoon of Ag-ama stellio iltuiiiatea by AL-HAJr et al. [2] conformsclosely in ultrastructure to that of the "":t*tlIii'iis"i) ii\*"[45]. Flattening of the acrosome isagain seen [2], as in A. adramitana [r8],dtil6T#"fd;.,G;i;#;;;i;il is not supporredby micrographs' A well defined narrow acrosbmal cortex is present (see also tg, 1gl). Frompublished accounts [37], division of the acrosome into cortex and medulli "un 6" claimed not onlyfor the Agamidae but also for Lacertidae, Teiidae, ah;;;Gonidae, varanidae and all families ofthe Serpentes' This subdivision is noi apparent in Gekkonidae but it is possible that layerinsdescribed for sphenomorph skinks t34l and the anterioi;;;;6;;;i.;";#; i" rhe Eugongylustgroup skink Lampropholis delican ua n th! pyggp.i-iirtx_uurtonis t37l is equivalent. Thewide zone between the acrosoma cortex and tlrg leiro'ril;;* [2, 1g] is clearly the subacrosomalcone' Late spermiogenic stages of A. agama t9l6onfirmitr".iti.rr* ill#rJ.tion but flattenedacrosome and wide, distinci nuclear slouldirs ;hi;ir;;]" pogona barbata,are intermediatebetween the concavg, angular shoulders of, for i"rfi;,bugongylus-group scincids, and therounded shoulders of sphJnomomhs.
with resard to.aTfinities of the sperm of Pogona barbata,it shares a larger number ofcharacter statis with.the fn* ,t"iir;""s gouldiithun *itr, othbr nori-agulid ,quu- ate taxail#?*1ffi!!:il*1"d. rhose atso commorito the,ph;il;;{pii-i.,riir:r.#i"ih1sse*orage have
368 B. G. M. JAMIESON : SQUAMATA (REPTILIA)
A varanid-agamid relationship does not appear to have been suggested on the basis ofsomatic .morphology (s9e review by RmeenL 147)). Of the three noteworthy spermatozoalsimilarities of-depressed acrosome, alternating, intermitochondrial rings and lhe'not entirelysimilar basal plates, o.nly the knob-like form of the basal plate appears td be an agamid-varanidsynapomorphy and it is homoplasic with Eugongylus-groups skinki. Nevertheless,lhe validity ofrecognizing a varanid-agamid relationship deserves further consideration.
A number of features shared between Pogona barbata and sphenomorph skinks, Tiliquascincoides (Egernia-group)., Cnemidophorus sexlineatus (Teiidae) and. Varanur appear, onintuitive consideration and in parsimony analysis, to be symplesiomorphic for these taxa: thesingle, pointed perforatorium and lineai cristae (plesiomoiphjc for squimates); the epinuclearelectron lucent zone, the elongate, cylindrical nucleus (plesibmorphic ior tetrapods); absence ofmultilaminar membtanes around the midpiece (presence has rbcently been recognized as asimilarity and po_ssible synapomorphy of snakes and pygopods [37,'and this acEount]; andextension of the fibrous sheath into the midpiece (autapomorphy of squamates).
Chamaeleonidae. A preliminary study of the spermatozoon of the rare South Africanchamaeleon, Bradypodion karroicum (Figs 4, 5)j reveals close similarity, on intuitiveconsideration, to the spermatozoa of the Agamidae (see above). The acrosome is sharplyattenuated but is depressed in one plane, Ttie acrosome vesicle is divisible into cortex andmedulla. Within the acrosome medulla the subacrosomal space encloses a perforatorium in theform of a naffow cylinder which tapers to a blunt point antbriorly and is crbss striated. A basalplate., questionably present in Pog.oia barbata, is a6sent. The subicrosomal cone, occupying thebasal two thirds of the 1""g14 of the acrosome, is conspicuously paracryStalline. 'fhe cyiindr"ical,electron dense nucleus has. slight, smoothly rounded shbulders inh a long, slightly .urv6d nuclearpoint (rostrum) which projects, almost to ihe tip of the subacrosomal coie uriA iJ preceded by anelectron lucent epinuc.lear 1egi.on. There is a broad basal nuclear fossa which, in Uelng polirteOanteriorly_,resembles that of sphenomorph skinks more closely than the shallow rounded iossa ofPogona. The proximal centriole, consistlng of nine triplets,I6s at right angles to the long axis ofthe sperm with its anterior region within the nuclear fossa (implantaTion dssa); a n*.o*"densityanterolateral to this centriole app.ears to continue to the suminit of the fossa. As in agamids, Ldistinct lamellar structure is nofdiscernible. As rn Pogona,the distal centriole, forming"ttr" Uuruibody of the axoneme, is elongate but similarly dols not extend into the iibro.rs iheath. Intransverse section, of^the midpiece through the fibrous sheath approximately 10 circularmitochondrial profiles.form a single circlet ar-ound the sheath. In longitudinal sectiois, tfre pioniesare again mostly circular but may show elongation consistent with a-spiral arrangement and this isparticularly tryg of the mitochondria in thelhort region of the midpiece antefror to the fibroussheath. Small intermitochondrial dense bodies sforadically interrupt the mitochondria intransverse and longitudinal sections. Dense rings of ihe type se-en in sphenomorph skinks are notprese^nt. A.large dense granule is visible in some mitochondria'and there is evidence oftransformation of mitochondria into intermitochondrial dense bodies. The midpiece is moOeiateiy9.l-ongatg and terminates at a distinct if small annulus. In the midpiece the axon6me has ninr coarsefibres; those at 3 and.-8- are enlarged, appear double, and are adpressed to the fibrous sheath. Nocoarse fibres are visible in the principal piece, posterior to th; mitochondrial midpie.". Fo; ;considerable distance posteriorio the annulus a-wide band of cytoplasm surrounds the fibroussheath as in Pogona barbata andVaranus ggLllQji [45] but also in Eugongylus, spfr"norno.pfr, unJegernid group skinks [35] and in the gekkonid HetZronotia binoei l3il.
In the parsimony g3lylis (Fig. 10) chamaeleonids have an unresolved position, withgekkonids, at the base of the Eugongylus-group-pygopodid-snake clade. Their risual iguanianrelationship or placement in the Agamidae is not uphi:i"a.^
Varanidae.In view of the fact that only one varanid has been examined for spermatozoal
I
ADVANCES IN SPERMAIOZOAL PHYI,OGENY AND TAXONOMY 369
pertoratorial baseplate
tlange ol thesubacrosomal
cone
pericentriolardense material
centriolestructure 1
distal centriole
structure 2
peripheralfibre
structure
fibrous sheath
enlargedperipheral fibre
a t 3
ililllli'll[ annulus
principal piece
llll lll llll--- endpiece 1.0 pmlllllllF-endpiece
F----u'-{
Frc. 6. - Varanus gouldii ftavirufus (Varanidae). A diagrammatic representation of the spermatozoon, in longitudinal and
corresponding transverse sections. After [45].Flc.7.- Heteronotia binoei. (Gekkonidae). Adiugtu.-utic representation of ihe spermatozoon, in longitudinal and
corresponding transverse sections. After [37].
370 B. G. M. JAMIESON : SQUAMATA (REPTILIA)
ultrastructure (Fig. 6), the following comparison with the sperm of other squamates can beconsidered only a most preliminary indication of affinities of the family. A suite of character stateshas been described for the spenn of Varanus gouldii flavirufus [45] which is known elsewhere insquamates in the Sphenomorphus and Egernia (Tiliqua) groups of the Scincidae [37] and in theteiid lizard, Cnemidophorus sexlineatus l42l although some are also seen in agamids (e.g.Pogona barbata t45l). This suite includes the following states. 1. Anterior depression of theacrosome (also in agamids). 2. The conical nuclear fossa containing dense material projectingfrom the proximal centriole. 3. The presence of a laminar structure extending from thepericentriolar apparatus (possibly unilateral only in the teiid) although this is less definitelylaminated tn V. gouldii. 4. Presence of four intermitochondrial rings alternating with columnarmitochondria. The rings differ in V. gouldii in being composed of many large granules incontrast with the solid condensed structure in the sphenomorph-Egernia-group and inCnemidophorus. The difference is lessened, however, by supposed constitution of the rings inthe scincid Chalcides ocellatus from a single circlet of large granules [21]. (An alternation ofmitochondria and albeit incomplete rings occurs in also in P. barbata).5. The rounded rather thanangular nuclear shoulders are apomorphic relative to the "sharp" or angular shoulders ofSphenodon l32l and primitive frogs [36]; a feature occurring only in the Eugongylus-groupskinks t3ll. (P. barbata shows an intermediate condition.) 6. The wide separation of the plasmamembrane from the fibrous sheath in the anterior region of the principal piece (also inP. barbata), although this has not been demonstrated with certainty for Cnemidophorus.
In the parsimony analysis (Fig. 10) the alternation of ring structures with mitochondrialcolumns computes as an ambiguous basic synapomorphy of the squarnates as a whole, rather thanmerely of the iguanid through sphenomorph assemblage. Flattening of the acrosome computes asthe sole basal synapomorphy of this assemblage, independently developed in Bradypodion.Consideration may be given, however, to the possibility that flattening is basic to squamates andhas been lost in the snake+pygopodid+Eugongylus-group clade.
Gekkonidae. In the Gekkonidae, FURTERI [21] briefly described the sperm of Lygodactyluspicturatus and made reference to Hemidactylus frenatus, H. mabouia, andTarentola mauritanicamauritanica; PurtrwS & AsA [46] described formation of the midpiece in Sphaerodactyluscinereus and JeumsoN e/ al.l37l described the sperm of Heteronotia binoei.
In the spermatozoa of H. binoei (Fig. 7) no epinuclear electron-lucent region has beenobserved (computing as a loss); nuclear shoulders are smooth, as in sphenomorph skinks;mitochondria are large and discrete, arranged in a circle around the fibrous sheath, withintervening mitochondrial transformations, and extend longitudinally as slender columns. Afeature not known in skinks is indentation of the median surfaces of the mitochondria at intervalsby triangular dense bodies which are perhaps always longitudinally interconnected.
The sperm of Lygodactylus picturatus, Tarentola mauritanica and Hemidactylus frenatusl2ll arc generally similar to those of H. binoei, but the amount of intermitochondrial material(dense bodies or putative mitochondrial transformations of the present work) is greater inLygodactylus picturatus and is present in decreasing amounts tn Tarentola mauritanica andHemidactylus frenatus respectively. The stellate arrangement of dense bodies, seen in crosssection of the midpiece, in T. mauritanica closely resembles that in H. binoei. A stopper likeelectron dense perforatorial plate, seen in H. binoei, is illustrated for Z. mauritanica-and, lessclearly, L. picturatus. A micrograph of the midpiece of Sphaerodactylus cinereus [46] shows alongitudinal series of four columnar mitochondria on each side (said to totaI20 mitochondria forthe midpiece) alternating with dense bodies, of comparable length, and the small annulus. In theabsence of transverse sections it is difficult to compare this arrangement with that of othergekkonids but the columnar form of the mitochondria is reminiscent of that in Heteronotia.
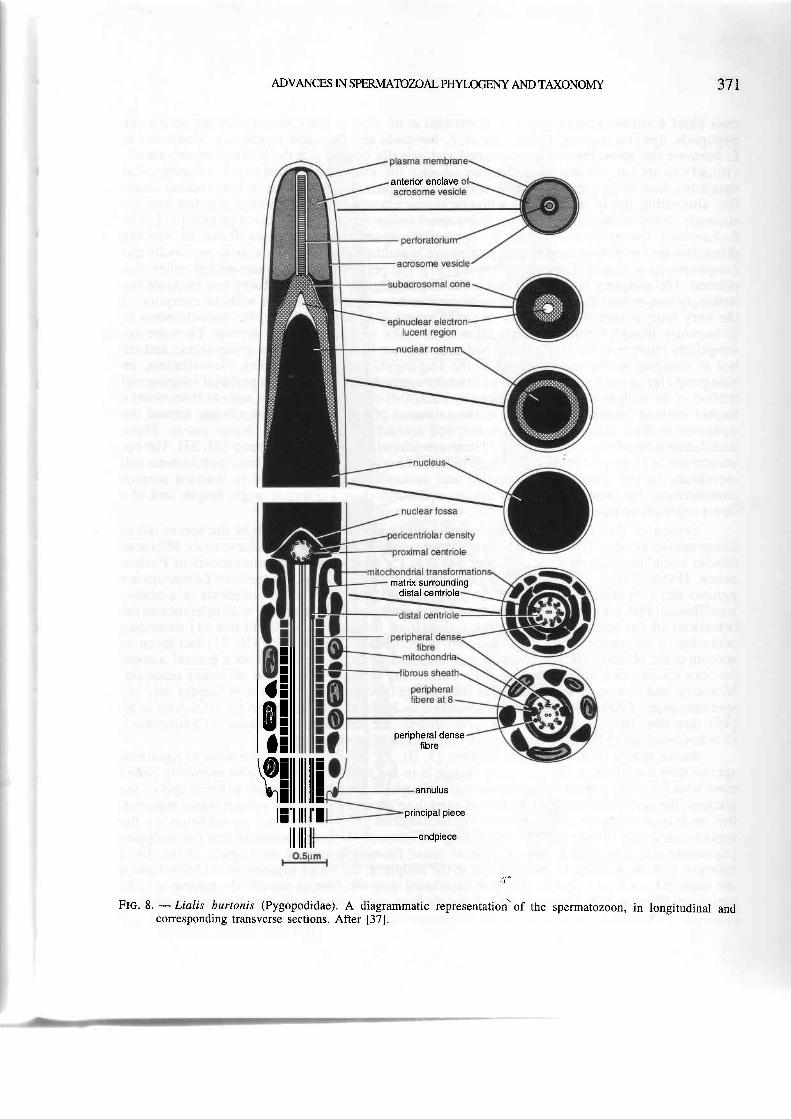
Pygopodidae. The spermatozo a of Lialis burtonis (Fig:. 8) are again like those of scincids in
371ADVANCES IN SPERMA TOZOAL PTIYI'GENY AND TAXONOMY
annutus
principal piece
peripheral densefibre
ililt!il t'i
rilTI I
!ilril
!llrilT I Ir l t
rtl
!tl] lT I
l .l@| l\@q
I
anterior enclave
matrix surroundingdistal centriole
ll lll ll_-endpiece
Flc.8. -Lialis burtonis (Pygopodidae). A diagrammatic representation'of the spermatozoon, in longitudinal andcorresponding transverse sections. After [37].
372 B. G. M. JAMIESON : SQUAMATA (REPTILIA)
their chief features, as was noted by HenoIN G et aI. I25l in their preliminary report for the
pygopoOs Aprasia' repens, Delma iincta, L. burtoni{ ald Pygopyi ,lepidopus. However, in
T.Turtontt the ucroro*e is fore-shortened and apically domed and the perforatorium extends
"ittourrv to its tip; nuclear shoulders are abseni; the-mitochondria are small sub-spheroidal
rtt.r.t rt""r, four oi five in a transverse section, and appear very numerous in longitudinal single
iiG, att"rnating singly or in groups with one or more dense bodies, with evidence that_they-are
sinuous; densE Uoiies also"form an interrupted collar around the distal centriole [37]. The
L. burtonis sperm shares a suite of apparently apgmgrphic character states of varying
distinctivenesiwith Eugongylus-group ski^nks (Cryptobtepharus virgatus, Carlia pectoralis and
faipropholis delicataifn.V arias follows I3JJ. The perforatorium is square-ended rather than
poin'i"a. The midpiece'is elongate, that in the Eugongylus-group species being less elongate but
itri6ngfft,ong". fhu.r that of rih"no*o.phs or otlier inv_estigatecl squamates with the exception of
itr" ".ii lonfmitochondrial^sheath of snake.sperm [33,-451. Hbwever, the mitochondria in
L. burt'onis, ihough forming a single layer as in snake sperm, are not as narrow' fngY are not
m"g"f*fy intersp"ersed witlithe deise bodies as they ar9 in the Eugongylus-group skinks.and are
not"as elongate as the mitochondria of the Fugongylus-group alq snakes. Nevertheless, an
elongated ,igrugconfiguration much as in snakeiperm is illustrated in a superficial.longitudinal;;;tifi oi tfrE *Tdpi""""of Aprasia repens by HanorNG et al. tzl) anlthese authors have noted a
further striking similarity tb snakes-in the existence of a multilaminar membrane around the
midpiece in tlieir material of L. burtonis, and around the flagellumin Delma tincta. These
multilaminar membranes had previously been considered unique to snake spefln l21,33l.The cg-
o."urr"nr" of elongate, tubular, zigzagledmitochondria an{ an, albeit'transient, multilaminar cell
membrane in the-sperm of pygopodias and snakes is here considered to warrant seriousconsideration that thise two grbulsbf legless squamates share a common origin despite lack of adirect sister-group relationship in the cladistic analysis (Figs 10' 11).
Serpentes. AusrtN [3] gave a detailed account of the fine structure of the spelry.tail inLampropZlfis getulus, Colibaeiconstrictor, Drymarchon,c_orais, Crotalus adamanteus, Micrurus
futvius and Cbnstricitor sp., BoISSoN & MATTEI [5, 6] described spermiogenesis in Pyth.on"sebae. HAMTLT9N & FawcerT l24l gave details of the neck and midpiece in I'ampropeltis
getulus and Constrictor constrictor, Sntre et al. l49l described pgelmioggnesis in Coluber"viridtflavius, PHTLLTpS & Ase [46] described the formation of-the midpiece with reference to the
behaviour of the annulus in Masticophis flagellum flagellum and AFzsLtuS [1] describedocclusion of microtubules in Liophis miliaris. Nevertheless, only FunmRI l20,2ll had given anaccount of the ultrastructure of the entire spermatozoon (giving in the latter work a g_eneral accountfor four species of Colubridae, Coluber viridiflavius viridifl.avius, Natrix tesselata tesselata,N. natrix, and Coronella austriaca, and one species of Viperidae, Vipera aspis aspis) until thespermatozoon of Nerodia sipedon was described by JaumsoN & KOEHLER [33]. Or-tvEF- et al.
Snalie sperm (Flg. 9) present features 13,20,21,24,33, 45,461 common to s,quamatespern: they are filiform; the acrosome vesicle is in the form of a hollow, concentrically zonedcbne whicl basally overlies a subacrosomal cone which invests the tapered anterior end of thenucleus; the perfoiatorium is a slender rod extending anteriorly from the subacrosomal material:the midpiec-e contains dense bodies (mitochondrial transformations) in addition to themitochondria; the fibrous sheath surrounding the axoneme extends anteriorly into the midpiece(squamate autapomorphy); nine peripheral dense fibres accompany_.the triplets of the distdcentriole and the doubleti of the aionerne in the midpiece; the fibres adjacent to doublets 3 and 8are enlarged, each as a double structure associated with thd fibrous sheath; the endpiece lacksperipheral fibres and the fibrous sheath. A poorly developed'lstopper-like" perforatorial base plate
ADVANCESINSPEKIuA-TOZOALPHYLOGEI\TANDTAXONOMY
connecting piece to/ peripheral tibre
oense
distalcentriole
multilaminar
enlarged peripheralfibre at 8'--
313
fibrous sheath.
. mitochondrial,transtormation
!il||tTill
\llFl---", 6\1 n,,m
Piece-(\:t
F,c. 9. - A diagrammatic representation of the spermatozoon of the Serpentes, in longitudinal and correspondingtransverse sections. After [45].
374 B. G. M. JAMIESON : SQUAMATA (REPTILIA)
in the colubrid Nerodia sipedon, unknown in other snakes, is presumably homoplasic with that ofgekkonids. An electron-lucent space caps the nuclear point in the snakes Boiga inegularis andStegonotus cucullatus as in some other squamate orders 133, 451but is poorly developed.
Less widespread is a suite of apparently apomorphic characters states of snake sperm [33,451 shared with Eugongylus-group skinks (Cryptoblepharus virgatus, Carlia pectoralis andInmpropholis delicata) and pygopodids. The shared states are as follows. The midpiece is greatlyelongated in snakes (mitochondrial sheath of FURIBRI I}ID, that in the Eugongylus-groupspecies, and pygopodids but also in gekkonids, being less elongate but strikingly longer than thatof sphenomorph skinks or other investigated squamates. The mitochondria in at least somepygopodids [25] formzigzagged tubes as in snakes. They are not inegularly interspersed with thedense bodies as they are in the Eugongylus-group skinks. A multilaminar membrane is seen insome pygopods as in snakes (see above). JAMIESoN et al. [37] stated that these similarities ofsnake, pygopodid and Eugongylus-group sperm warranted further investigation with a view todetermining the degree of homoplasy as against synapomorphy. Individually these charactersappear in some other squamates. Thus we have seen that extracellular tubules occur in immatureteiid sperm and that multilaminar membranes invest the acrosome in immature tropidurid spenn.Possibly both types of structure are a normal feature of developing squamate sperm. Twoapomorphies of snakes and pygopodids, the multilaminar membrane and the tubular zigzaggedmitochondria are striking, and intuitively were considered synapomorphic resemblances.However, multilaminar membranes computed as basal not only to these two groups but also toEugongylus-group skinks, in which they were computed as lost, and sinuous mitochondriaparsimoniously (but not, perhaps, plausibly) computed as a basic squanlate feature.
The evidence, albeit uncertain, for transformation of extracellular tubules into multilaminarmembranes in Boiga irregularis possibly represents a stage in production of the membranes.Conversion of microtubules into membrane-like laminate appendages of testicular spenn is knownin the Lepidoptera (see review in Jamieson t30l). It has been suggested that the multiple layers ofmembranes provide a source of endogenous phospholipid that could be utilized as a source ofenergy for motility [24].
The dense collar (termed the neck cylinder by AustN [3]) is considered to be homologouswith the intermitochondrial dense bodies 124,451which in turn have been shown in skinks (e.g.Cryptoblepharus virgatus) to be derived from mitochondrial3Tl. The dense element in the centralaxis -of the proximal centriole, seen in Aspidites, is also illustrated and described for Lampropeltisgetulus l24l and is stated to be regularly found in mammalian sperm. They [24] also observe thepresence, unknown in other vertebrates, of extracellular microtubules, observed here in at leastAspidites melanocephalus and Boiga irregularis. With regard to the axoneme, they remind us thatenlargement and prolongation of the peripheral fibres 3 and 8 in snakes (as,-we note, in allsquamates) is the reverse of the situation in the mammalian sperm tail where these are the smallestfibres and terminate first. The density associated with one of the two central singlets 13,241,Uggg!!-tg be unique to snake spenn, has since been demonstrated for the skinks Naigura spinoiap!], filiqua scincoides and less certainly Carlia pectoralis [35], the gekko Heteronotia'binoei[37] and the teiid Cnemidophorus sexlineatus [42] and may well be moie widely demonstrated insquamates yhgn favourable sections of the centriole are obtained. Dense material filling the Asubtubules in the posterior portion of the axoneme has been demonstrated for doublets I:2, 5,6or L,2,5, 6 and 7 in Liophis miliaris [Il.
Parsimony analysisA po{eriori to intuitive consideration of comparative spermatozoal ultrastructure, with some
inferred relationships, which has been detailed above, preliriinary parsimony analyses have been
ADVANCES IN SPERMATOZOAL PHYLOGEI\IY AND TAXONOMY 375
Tesls 1' - Ultrastructural characters of squamate sperm used in parsimony analysis. character states numberedparentheses are not transformation series as polarity was detirmined by use of an outgroup. The first statenevertheless considered plesiomorphic.
lnis
Character States
I Acrosome TS2 Perforatorial base plate3 Perforatorial tio4 Perforatoria, number5 Epinuclear lucent zone6 Midpiece7 Mitochondria in TS8 Mitochondria, shape
(0) circular (l) depressed,(0) absent oi lnOistinct (t) noUru" (2) stopperlike,(0) pointed (1) square ended,(0) two or more (1) one,(0) absent (1) poorly developed (2) well developed,(0) short (1) moderately long (2) very long,(0) regular circlet (l) not regular (2) intermediate,(0) rounded (1) columnar (2) sinuous tubes (3) .intermediaterounded-columnar',(0) intramitochondrial (1) regular rings (2) scattered (3) linearseries (4) stellate spiral (5) 2 groups,(0) not applicable (1) solid (2) granular (3) single file granules,(0) concentric (1) linear,(0) sharp (1) rounded (2) absent,(0) elongate (l) stout,(0) present (1) absent,(0) enlarged (1) grossly enlarged anteriorly,(0) absent (l) presenr,(0) not in midpiece (1) in midpiece
performgd.using the PAUP program. The characters employed are those listed in Table 1 and thestates of these characters are given in Table 2 whicli piouioei ;;ililt of ultrastructurediscussed in this account.
A branch and bound *Tch was performed for the total data matrix (Table 2). The followineoPtio.ns Ye-t-e--TPtje4: addition sequbnce: furthest; 1 trei held at "."it-;;p]*tft';"ilil!addition; MULPARS option in effeit; steepest descent option not in effecC iriitial;p[;ffi;a;unknown (compute via stepwise); branch'es having -uil-o- length ,"io.ottupiJ,j to i;16polytomies; no topological constraints; trees u-nrooted; multi-"state taxa iriterpr"t..,i--u,polymorphism; character-state optimizaiion acceleraied iransformutio"llccTRAN). Allcharacters were treated as unordered.
The branch and bound analysis produced 240 most parsimonious trees from which a strictqtg' 11) a1d a50vo.m.aj9ri!y iute (nig. 10) consenruilr." was computed. These had thefollowing characteristics in both analysel:-tree ir-rg-m = +j; .onristencyindtx = O.l;j;rr"-"pf"rvindex = 0.302: retention index = 0.839; rescale? "onrirt"n.y inde'x = 0.64i. Character statechange-s are included in Fig. 10.If the search was made with multi-state taxa scored as uncertainty (to allow for the equivocalcondition of the mitochondrial derivatives, charac ter 9, ii iogonal inlo*ti"ul number of treesand topolo8.Y yere obtained but the tree was^three stepi strortir, with consisten-y index = 0.750;homoplasy index = o.250; retention index = 0.839; ."r'.ul"d .onsistenry inJ"i = 9.629.It must be stressed that the strict consensus tree (Fig. I 1;, agreed.br;ly with the majorityru19 grger.differing only in placing cnemidophoru, oitir.iu,,,.level as ttre remainJ;;ilr'Jilaand Lialis on the same levrel as the carliaind snake "ruao. These l*" p"ivl"-ies correspondwith the only percentages below r00vo on the ma;ority;i";;"".
316 B. G. M. JAMIESON : SQUAMATA (REPTILIA)
TABLE 2. - Comparative ultrastructure of spermatozoa of Squamata, Sphenodon punctatus and Chelonia' (Input datamatrix).
Taxon Characters Reference11111111
t23456'7 890!23456'7
CheloniaSphenodon punctatusCtenotus robustusChalcides ocellatusLacertidaeCnemidophorus sexlineatusTiliqua scincoidesCarlia pectoralisLampropholis delicataHeteronotia binoeiLy go dac ty lus pic turatusLialis burtonisPogona barbata
Discussion of the phylogramsThe majority rule phylogram (Fig. l0) indicates those similarities of spermatozoal
ultrastructure within the Squamata, and relative to Sphenodon and the Chelonia, which aresynapomorphic for the respective nodes. These similarities, as in all spermiocladistic studies,must reflect similar fertilization biology but also phylogenetic, i.e. genetic, constraints.Spermatozoal morphology is here considered sufficiently conservative to reflect at least closephylogenetic relationship as it can be expected to be subject to stabilizing selection [22]. Theevidence of many other studies (see, for instance , 126, 27 , 31, 32) and references in [34]) showsthat, although caution is required in their interpretation, such spermiocladistic phylograms holdsignificant phylogenetic information. Bearing in mind the necessity for caution and also thepressing need for inclusion of reliable data from a much larger sample of taxa, and confirmationof some data from the present sample, many of the following conclusions drawn from thephylogram can be regarded only as heuristic. Findings which both intuitively and by computationare unequivocal are stressed. Because of limitations of space, references to the literature willchiefly be confined to the excellent review of squamate classification by RIEPPEL [47].
Squamata. The Squamata, which are monophyletic in all most parsimonious trees in theanalysis, are apomorphic relative to the Sphenodontida and Chelonia in the following featureswhich are synapomorphies defining the Squamata s. strict. Sphenodon and the Chelonia areequally plesiomorphic and indistinguishable on the charactef,s employed and therefore referencewill chiefly be made to Sphenodon.It should be stressed, however, that Sphenodon is not here
FIG. 10. - Branch and Bound 50 Vo majofity rule consensus phylogram for 240 equally most parsimonious trees, forcharacter states of chelonian, sphenodontid and squamate sperm ultrastructure listed in Tabies I and 2. Settings asin text. Tree length = 43; consistency index = 0.767; homoplasy index = 0.302; retention index = 0.839; r"sJd.dconsistency index = 0.644. Numbers in circles are the percentage of trees supporting the node to which theyattach. All other nodes were supported in 100 Ea of the tees.
regarded as the immediate sister-group of the Squamata sensu strictu but as a very basal amniotein a sphenodgntidan Jing?g" whiCh possible predates emergence of the Crocodilia, Aves, and, itappears, the Mamm alia [26, 32].
. The s-quamltq s.ynapgmo.rphies_follow: 1. Possession of a single perforatorium in place ofthe two or three of Sphenodontida; 2. Loss of the endonuclear canal.-Thd wholly epinuclear ratherlhqn parjly endoluclear.perforatorium is a notable difference of the Squamaia irom Chelonia,Sphenodontida, Crocodilia_, non-passerine birds, and also basal lissam^phibians (urodeles andA.scaphus). 3. Presence of the (well developed) epinuclear electron-lucerit region. 4. presence ofsinuous mitochondria (possibly an artefactual^parsimony resolution as a"columnar form isintqitively prefened). 5. linear cristae, not the subspherical
-mitochondria, with concentric cristae
of.S-ph-enodon. 6. Dense bodies (mitochondrial transformations) intermitochondrial, in contrisiwith the intramitochondrial.dense body in Sphen-odory, ald (e_quivocally) forming regular rings.Although the intermitochondrial position rya! weil be basic tlail squamii"s, on" tfiignT qr.w tf;"ithe.arrangement in regular rings is in their ground plan. 7. Ehe fibrous sheath extindi inio themidpiece. This is a most significant and convincing synapq-morphy and autapomorphy of the
FIc' 11. - Branch and Bound 50 Vo strict consensus phylogram for 240 equally most parsimonious trees, for characterstates of chelonian,^sphenodontid and squamate 'sperm ultrastructure listed in Tables I and 2. Settings, length andindices as in Fis. 10.
Squamata and is not seen in the Sphenodontid,a 126, 321. 8. Development of rounded nuclearshoulders is basic to squamates. In addilion, though nol includea iir ttre parsimony unuf'rir,squamates are diagnosed by the paracrystalline substructure of the subacrosoriral cone.... . . Jh.te squamate spermatozoal autapomorphies constitute a striking endorsement of the-ligttV corroborated" monophyly of the Squamita for which evidence isieviewed by Rmrrpit471.
'Sauria'- The Sauria is the third group- of the Squamata, the other two being the$mphisbaenia and the Serpentes. Whereal the latter two groups appear to be monophyietic,4IEPIF_L [47] stated that monophyly of the Sauria could ndt ue esti6fished and that the'nu*"should be dropped or, in effect, brbadened to equate with Squamutu.if the evidence of thespermatozoal phylograms (Figs 10, 11) be accepted, the view tirat Sauria (in the strict sense of'lrzarcls' rather than squamates in general) are not monophyletic is emphatically endorsed as some:f:tiii: q",I*?gly Eugongylus-group skinks and pygbp6dids, and,iess clos"ely, gekkonids andme ciamaeleontd) aqpear.more closely related to snakes than they are to othei 'lizards'. Thus,Sauria is_only monophyletic when it is,-indeed, equated with Squaniata anA ii inttudes lizards andsnakes. The positio.l .o.f the amphisbaenians remains to be investigated when spermatozoalsamples become available.
.Scincomorqha. Monophyly of_the Scincomorpha, though well supported by somaticpgr.phology,-has been considered to deserve further criticat 919iy t+Zl. rtr'e'gro"p ir"g"n.ditheld to include the Xantusioidea, Lacertoide_a (Lacertidae, Teiidae unOby-"Spntfiafmlaaeiantthe.Scincoidea (Scincidae and Cordylidae). Spermatologically @igs io, lil it.'s"i"."-"tlit " i,diphyletic or possibly polyphyleti-c. Thi,s is drie to the tictthittfrJscincoicieu,irpr.r"nted in theparsimony analysis by the Scincidae only, are themselves at least Oipfrvf"tio tjolubty U""uui" it "Eugongylus-group skinks associate with the Serpentes, nygopoii,ia",-drru-u"reonidae andGekkonidae
ADVANCESINSPERMA,TOZOALPTM,GEI.{YANDTAXONOMY 37g
. . RtBppEL (e.g' [47]) has repeatedly questioned the monophyly of the scincidae and hisdoubts are underlined spermatdlogicaily. ^lttof trre eugongyiui-gtoup ,kinks (carlia andLampropholis) constirute the sister-lroup'of the eygopoOi?*i'unO tf;i pfgopoOiO_iG"ilyt,;;_clade is in turn the sister-group of thE serpent"s in'tfie'maiority ;G i;;":(F:gi rol. However, thepygopodid forms a trichotomv with the'Eugongylus-gr6up ano serpenies"thd", in the strictconsensus tree (Fig. l1).
. The apparenl, synaPoTorp.hiel joining the Carlia through Serpentes clades are weak. Theyare.loss of 1!9 regular mitochondriat Lirctetlano, iquiuo.aiy,"o""Jdp*"rt "iri**-oJ"J f"^oi!',and of multilaminar membranes. of these, the lait two conditions are not seen in Carlia and,Lampropholis. These two genera are linkedig theptggp;dto-u/i""i "p"tii"iirries. Two of these,the square-ended perforaiorium and alteration .ir ftr6 nucleus to a ^stout
iorm appear strongsynapomorphies, while two. are.equivocal, acquisition or a m991ik; b"il;iut" Crrorn6praric *itf;Pogoryc andvaranu) lld {.evelopment of shdrp il.G;rhoulders.The severance of the Eugongylus-group front oth..rtinr.r merits further investigation usingnon-spennatozoal characters. The ipermatoioal synapomorphies d;ii"irg i-t. SuSo.ngyrus_groupare striking' They are the characteristic developrnJnt-o'ficutt"leo dense bodies, romung more thanone layer around the axoneme; the fact that co'arse nui"r i *o 8 ;;;il;ly-eritargeo anteriorly inaddition to the usual enlargement in squamates; and, equivocally, ,"ulrrioo'to no murtilaminar cellmembranes.
.. Th".Position of the Lacertidae and Teiidae is poorly resolved (Fig. 10), and further data onthese families are rqquired. Linkage of the fidid;-"rd reiidae lir tfiei6iertoidea [47] is notupheld. A teiid relationship to S9 ier*i*g (;f*rlt;;r inrttitffi;; nitffio."d. rn the strictconsensus tree (Fig. 11), lacertids, ihe teiid Qryeryidgnhorus, andthe skinks Cienotus, chalcides,and ri.liqua.form an unresolved polytomy with the Fi joii-voronus clade.Ho*.u"r, in the 50vomajority rule tree !lig. to)'.gr? eintdophdrus is less ctoie to the other r.in.o-orphs than they areto each other. Intuitive considerations had plaqed teiias near qph""A;{pil (Z;;; otus) and.egernid(Tiliqua) skinks [37] but an especiallv ctosiiel"ii""rr"p-ir rot upheld in this analysis.Lacertid spenn have noibeen exami:red by the aJttoi uut'thi;;l"di" Jutu for the family,while placing them with the other, non-nugon"gyi;r-r.iti.o-orprrs, ao'noiaipresent ally themmore closely to any scincid or with the teild. ifreii rynupolo.phies are arrangement of densebodies into two sroups and, of uncertain uirioity, iJu*i"r to a poorly developed epinuclearelectron-lucent re-sion. As recoroeAi;h;.;;#ii"r r.&* above, the arrangement of densebodies in lacertidiappears to be unusually divtrse 6oi ir-i"u"r strongly similar to the ctenotuscondition.
Iguania' Traditionally these include the Iguanidae, Anolidae, Tropiduridae, Agamidae andchamaeleonidae' The Iguania does not emerge.as a monophyletic grgup in the present analysis(Figq f 0' ll)' Pogona gioups not with a1 igullian;il;ifr varanus in -a
clade which otherwiseconsists of non-Eugongylui scincomorphr"*hil" igu;rd;--id;h";Gri";;h sister-group ofthis clade. The oledlorritirpti. ri"iw "l^thr rgu*id?";;il;r to agamids (references in RrsppsrlaTl) is endorsed but here in u p*ufty[ri.,j;ti;ilip. ^--
The chamaeleon Bradypbaioin is severed from ihe Iguanidae and is basal in the gekkonid-snake-pvgopodid-Eueofqvlus-sroup claae in iil;;j;;fty.rute rrig.-iol-ano strict (Fig. tl)consensus trees in tl-re present analysis. It may u" n6i.o," rror"u"rJtrrut'in-ll*, of patristicdistance, the nearest taxon to the 6hamaeleoi ir, r"r"iirreless, the lguanidae. on intuitiveconsideration (see comparative account ugo'"i, G rli;';iil;;;il;;j.;?;#; closely similarto that of the aeamid in manv respects out-trr.-.in"d;;ffi1*.ri"Jrr""iii5L an imporrantfifferengg,from-agamias undl"ritriieh a resemblan."lo iguunids,-are also seen in the carlia-pygopodid-snake clade. Much furtlier.inuertigation oi Jp"r-utozoa, and comparison withmorp$.loqical- characters, is necessary iriguani""-;i"ti;hips are to-be elucidated.The knob-likebasal
Plut: uppgqs to"be the ;b;t;up1*orprry of varanus and pogonabutis homoplasic with that in eailiaiia r"ipiii:iitii.rv uruqv\
380 B, c. M. JAMIESoN : SeUAMATA (REITILIA)
Anguimorpha. Anguimorphs are conventionally divided into the Anguioidea and theVaranoidea [47]. Anguioidea have yet to be examined for sperm ultrastructure. In the comparativestudy above, the similarity of the Varanus spenn to that of Pogona has been described. From thecomparative study, varanid sperm resemble those of agamids, sphenomorph and egernid skinks,and teiids in the depressed acrosome and intermitochondrial rings; they further resemble agamidsand also (homoplasically) Eugongylus-group skinks in possessing a knob-like perforatorial baseplate. Intuitively, as in the analysis (Fig. 10), the varanid is part of a sphenomorph-egernid-agamid assemblage. It is noteworthy that although a close relationships of varanids with scincidsdoes not appear to have been previously suggested, brain data have been considered to indicatethat the Teiidae are most closely related to the Varanidae [43].
A very close relationship between varanids and snakes which has formerly been postulated(see references in 145,47D is not supported by spermatozoal ultrastructure. Varanus spefin differfrom those of snakes in the short midpiece, columnar mitochondria in a regular circlet, denseintermitochondrial bodies forming regular rings and absence of multilaminar membranes. It isexpected that varanid relationships will come nearer to resolution when more than one species ofeach family is represented. A unique varanid apomorphy is replacement of the solid condition ofthe intermitochondrial rings, also seen in Ctenotus, Cnemidophorus,Tiliqua, Pogona andiguanids, with a granular condition.
Thus, the relationships of varanids remain enigmatic despite the parsimony analyses but aclose relationship to snakes is not upheld.
Gekkota. Monophyly of the Gekkota, comprising the two extant families Gekkonidae andPygopodidae has been considered well corroborated (references in REppsL t47D. However, themajority rule consensus tree (Fig. 10) represents the Pygopodidae, exemplified by Lialis burtonis,as the sister-taxon of the Eugongylus-group skinks (for synapomorphies see Scincomorphaabove). In the strict consensus tree (Fig. 1 l), Lialis forms a trichotomy with these skinks and thesnakes. In the parsimony analysis (Figs 10, 11) the gekkonids form a monophyletic clade which,however, has an unresolved relationship with the chamaeleon, at the base of thesnake+pygopodid+Eugongylus-group clade. The only unequivocal synapomorphy for thecomposite clade, is development of a moderately long midpiece which becomes very long insnakes. No unique synapomorphies are apparent between the sperm of gekkonids and pygopodswhich would support the special relationship between these two families suggested by Kruce[38, 39, 40] who from a cogent cladistic study of general morphology has concluded [39] that thePygopodidae should be placed within the Gekkonidae. This departure from somaticmorphological evidence requires fuither investigation from sperm of larger numbers of taxa.
A relatively primitive position of the Gekkonidae in the 'Sauria' proposed by some workers1471, as opposed to the Pygopodidae, is supported by both consensus phylograms. Gekkos havesomewhat generalized scincid spenn but a stellate arrangement of dense bodies in the midpiecedistinguishes at least some of them. Heteronotia and Lygodactylus are sister-taxa in l}OVo of thetrees. Computed synapomorphies of gekkonids (Heteronotia and Lygodactylus) arc the stopper-like perforatorial base, the columnar condition of the mitochondria, supposedly derivedsecondarily from a sinuous condition; and, ambiguously, the stellate mitochondrial derivativeswhich compute as arising from a scattered condition such as is seen in Eugongylus-group skinks.
Serpentes. The origin of snakes, on the grounds of somatic morphology, continues to be anenigma [47]. However, the spermiocladistic analysis indicates origin of snakes from an ancestor,which must have been skink-like, shared with Eugongylus-group skinks (Carlia andLampropholis) and with pygopodids. The pygopodid has a sister-group relationship with theEugongylus-group_ skinks in the majority rule tree (Fig. 10) but forms anunresolved tiichotomywith these and snakes in the strict consensus tree (Fig- 1l). At present this origin of snakes mustIgTaE mgrely an heuristic finding but spermatozoa do provide a serpent apomorphy (see alsoI33,45D shared only with the pygopodid, presence of multilaminar sperm cell-membranes. These
ADVANCES IN SPERMATOZOAL PFTLOGEI\ry AND TAXONOMY 381
membranes appear much better developed in snake sperm which differ further, and-uniquely,inthe immense elbngation of the midpiece as a mitochondrial sheath around most of the lengthof theaxoneme. That the multilaminar membranes have been lost in Eugongylus-group skinks, ascomputed, may be questioned. If not, presence would be a snake-Pygopod synapomorphy.Further snake apomorptries appear to be reduction of the epinuclear electron-lucent regior_r andloss of a perforatorial base phe. Snake spenn also have a greater development of extracellulartubules (not coded) than is known in any other squamate.
With regard to groups not included in the analyses, tropidurid sperm are insufficientlyknown for determination of phylogenetic affinities but the three gyres of mitochondria are also alacertid characteristic; supposed failure of the fibrous sheath to penetrate the midpiece (thesquamate autapomorphy) requires confirmation.-
Investigations of sperm of additionaltaxa are required if suggested relationships throughoutthe Squamata are to be adequately tested.
ACKNOWLEDGEMENTSI am grateful to Mrs Lina DADDow, David Scsslrwc.q, Chris TUDGE, and Simon Olrvpn for excellent assistance and
to Chris TuDGE and Tom GonnrNcs for printing the micrographs. Professor Alan HoocsoN and Dr. Rick BenNano kindlysupplied a resin block of Bradypodior? sperm. This work was partly supported by an Australian Research Council grant.
REFERENCES
1. Arzerrus, B. A., 1981. - Electron dense microtubules in the animal sperm tail. Journal of SubmicroscopicCyto logy, l3 :199-207. :
2. AI-HAJJ, H., JANAKAT, S. & Ml.Hr4ouo, F., 1987. - Electron microscopic study of the sperm head differentiation inthe lizard Agama stellio. Canadian Journal of Zoology,65: 2959-2968.
3 . Ausrnq, C. R., 1965. - Fine structure of the snake sperm tail, Journal of Ultrastucture Research,12: 452-462.4. BAVERSTocK, P. R. & DoNNst-r.eN, S. C., 1990. - Molecular evolution in Australian dragons and skinks: a progress
report. Memoirs of the Queensland Museum,29: 323-331.5. BoISSoN, C. & Merret, X., 1965. - Sur la spermiogenEse de Python sebae (Gmelin) 6tudi6e au microscope
6lectronique. Compies Rendus des S4ances de la Soci4td de Biologie de l'Ouest Africain, S: 1192-1194.6. BorssoN, C. & Marrsr,X., 1966. - La spermiogendse de Python sebae, Gmelin, observ6e au microscope
6lectronique. Annale s des Science s Naturelles, Zoolo gie, 8: 363-390.7 . BurLER, R. D. & GasnI, M. S., 1984. - Structure and development of the sperm head in the lizard Podarcis (=
Lac e rt a) t auric a. J ournal of U ltr a s t ruc ture Re s e arc h, 88 : 261 -27 4.8, C,q,ncuprNo, M., CoRso, G. & P.lLa, M., 1989. - Spermiogenesis in Chalcides ocellatus ti l igugu (Gmelin)
(Squamata, Scincidae): an electron microscope sfidy. Bollettino di Zoologia,56: 119-124.9 . CsenNtnn, M., BoISSoN, C. & M,qrrsr, X., 1967. - Ultrastructure de la manchette chromatique dans la spermatide
d'Agama agama (Agamidae, Lacertilien). Comptes Rendus de la Soci4td de Biologie de l'Ouest Af 'cain, 16ll.888 -890 .
10. Clenr, A. W., 1967. - Some aspects of spermiogenesis in a Iizard. American Journal of Anatomy,l2l:369-4Q0.11. CounrnNs, J.L. & DEPEIGES, A., 1985. - Spermiogenesis of Lacerta vivipara. Journal of Ultrastructure Research,
90: 203-220.12. le Cnuz HOFLUNG, M. A. & nl Cnuz LlNorv, C., 1978. - The fine structure of nuclei during spermiogenesis in the
13. Da Cnuz-LaNona, C. & De Cnuz-HOFLtrrc, M. A., 1977. - Electron microscope study of lizard spermiogenesis inTropidurus torquatus. Caryolo gia, 30: 15l-162.
14. DpslewI, G. Y., 1992. - Studies on the ultrastructure of the spermiogenesis of Saudian reptiles 4. The sperm headdifferentiation in Chalcides ocellatus. Histology and Histochemistry,T (C):331-347.
15. DEHLAwI, G. Y. & ISMAIL, M. F., 1990. - Studies on the ultrastructure of the spermiogenesis of Saudian reptiles 1-The sperm head differentiation in Uromastyx philbyi. Proceedings of the Zoological Society of the ArabianRepublic of Egypt,2l: 79-89.
16. Dpslawr, G. Y. & Isv.q.n, M. F., 1991. - Studies on the ultrastructure bf the spermiogenesis of Saudian reptiles 3.The sperm head differentiation in Stenodactylus selvini. Egyptian Journal of Histology, 14: 71-80.
382 B. G. M. JAMIESON : SQUAMATA (REPTILIA)
l7 . DeHr-Rwr, G. Y., Isue[, M. F. & SALEH, A. M., 1990. - Studies on the ultrastructure of the spermiogenesis ofSaudian reptiles 2- The sperm tail differentiation in Uromastyx philbyi. Proceedings of the Zoological Societyof the Arabian Republic of Egypt,2l:91-101.
18. DnnLRwr, G. Y., ISMAIL, M. F., HAMDI, S. A. & JAMJooM, M. B., 1992. - Ultrastructure of spermiogenesis ofSaudian reptiles. 6. The sperm head differentiation in Agama adramitana. Archives of Andrology,2S: 223-234.
19' DplCoNrp,8., 1976. - The subacrosomal granule and its evolution during spermiogenesis in a lizard. Cell andTissue Research, l7 l: 483-498.
20. FuRIsnr, P., 1965. - Prime osservazioni al microscopio elettronico sulla ultrastruttura dello spermatozoo di Viperaaspis aspis. Bollettino della Societd Italiana di Biologia Sperimentale,4l: 478-480.
2I' FuRIrnr, P., 1970. - Sperm morphology of some reptiles: Squamata and Chelonia. 1n..B. Baccerrr,ComparativeSpermatology. Rome, Accademia Nazionale dei Lincei: 115-131.
22. FURIERI, P., 1974. - Spermi e spermatogenesi in alcuni iguanidi argentini. Rivista di Biologia,6T:233-279.23. Gnesn, A.8., 1979. - A phylogenetic subdivision of Australian skinks. Records of the Australian Museum,32:
339-37 | .24. HAMILTON, D. W. & Fewcprr, D. W., 1968. - Unusual features of the neck and middle-piece of snake spermatozoa.
Journal of Ultrastructure Research, 23: 8l-9j .25. Hanotuc, H. R., ApuN, K. P. & MAZUR, M., 1994, - Sperm ultrastructure and phylogeny in repti les: the
Pygopodidae. ftl: M. BnanlEv & J. CuuvrNs, Seventh International Symposium on Spermatology, Program andMini-Poster Abstracts. Cairns, North eueensland: 5.35-5.36
26' Hnelv, J. M. & JAMIESON, B. G. M., 1992. - Ultrastructure of the spermatozoon of the tuatara (Sphenodonpunctatus) and its relevance to the relationships of the Sphenodontida. Philosophical Transactions of the RoyalSociety of London 8,335: 193-205.
27. HBer-v, J. M. & JAMIESoN, B. G. M., 1994. - The ultrastructure of spermatogenesis and epididymal spermatozoa ofthe tuatara Sphglqdon punctatus (Sphenodontida, Amniota). Philosophical lransactions of the Royal Societyof London 8,344: 187-199.
28. HurcHINSoN, M. N. & DONNELLAN, S. C., 1993. - Biogeography and phylogeny of the Squamata. In: C. J. GLAsBy,G. J. B. Ross & P. L. BEESLEY, Fauna of Australia. Vol 2A Amphibia- and Reptilii. Australian GovernmentPublishing Service, Canberra: 210-220.
29. ISMAIL, M. F. & DEHLAWI, G., 1992. - Studies on the ultrastructure of the spermiogenesis of Saudian reptiles 5 - Thesperm tail differentiation in Chalcides ocellatus. Histology and Histochemistry,T (C): 2ll-222.
30. JaurnsoN, B. G. M., 1987. - The Ultrastructure and Phylogeny of Insect Spermatopa. Cambridge (U. K),Cambridge University Press.
31. JRturrsoN, B. G' M., 1991. - Fish Evolution and Systematics: Evidence from Spermatozoa. Cambridge (U. K),Cambridge University Press.
32. JeN'rrssoN, B. G. M. & Hp,ql-v, J. M., 1992. - The phylogenetic position of the tuatara Sphenodon (Sphenodontida,Amniota), as indicated by cladistic analysis of the ultrastructure of spermatozoa. Philosophical Tiansactions ofthe Royal Society of London B, 335: 20j-219.
33. JAMIESON, B. G. M. & KoprnEn, L. 1994. - The ultrastructure of the spermatozoon of the Northern Water Snake,Nerodia sipedon (Colubridae, Serpentes) with phylogenetic consideiations. Canadian Journal of Zootogy,T2:1648-1652.
34. JAMIESON, B' G. M. & ScHsI-rtNcA, D. M., 1993. - The ultrastructure of spermatozoa of Nangura spinosa(Scincidae, Reptilia). Memoirs of the eueensland Museum,34: 169_179.
35' Je,lrrnsoN, B. G. M. & ScHEI-rlNcl, D. M., 1994. * The ultrastructure of spermatozoa of the Australian skinks,Ctenotus taeniolatus,Carlia pectoralis andTiliqua scincoides scincoidesjscincidae, Reptilia). Memoirs of theQueensland Museum, 37: l8l-193.
36. JAMIESON, B. G. M.' Lep, M. S. Y. & LoNc, K., lgg3. - Ultrastructure of the spermatozoon of the internallyfertilizing frog Ascaphus truei (Ascaphidae: Anura: Amphibia) with phylogenetic considerations.H erpe t olo gic a, 49l. 52-65.
37 . JevIEsoN' B. G. M., OLIVER, S. C. & Scsslrntca, D. M., 1995. - The ultrastructure of spermatozoa of Squamata. I.Scincidae, Gekkonidae and Pygopodidae (Reptilia). Acta Zoologica (Stockholm.), (In-press).
38' Klluce' A. G., 1967. - Higher taxonomic categories of gekkonid lizards and their evolution. Bulletin of theAmerican Museum of Natural History,l35: l_59.
39. Kt-ucn, A. G., 1983. - Cladistic relationships among gekkonid lizards. Copeia, 1983:465-475.40' Klucs, A. G., 1987. - Cladistic relationships in the Gekkonoidea (Sqr:amata, Sauria). Miscellaneous publications
of the Museum of Zoology, University of Michigan,173: l_54.
4 1 .
4 2 .
43.
44 .
4 5 .
46 .
4 7 .
4 8 .
49 .
5 0 .
5 1 .
52.
ADVANCES IN SPERMATOZOAL PHYLOGENY AND TAXONOMY 383
NEwroN, W. D. & TRAUTH, S. E., 1990. - Sperm morphology and ultrastructure in Cnemidophorus sexlineatusSauria Teiidae. American Zoologist,30: 44A.
NEwToN, D. W. & Tneurn, S. 8., 1992. - Ultrastructure of the spermatozoon of the lizard Cnemidophoruss exlineatus (Sauria: Teiidae), H erpetolo gica, 48: 330-343.
Nonrucurr, R. G. & BurLER, A. B., 1978. - Forebrain and midbrain organization in lizards and its phylogeneticsignificance. 1z; N. Gnnshlssnc & P. MAcLEAN, Behavior and Neurology of Lizards. Rockville, Maryland,NIMH Publication: 11-64.
OKIA, N. O., 1990. - The ultrastructure of the developing sperm tail middle piece in the skink Eumeces laticeps(Lacertilia: Scincidae). Proceedings of the XIIth Intemational Congress for electron Microscopy - ElectonMicroscopy,3: 84-85.
Or-rvsn, S. C., JeutnsoN, B. G. M. & ScHELTINGA, D. M., 1995. - The ultrastructure of the spermatozoa ofSquamata. II. Agamidae, Varanidae, Colubridae, Elapidae, and Boidae (Reptilia). Herpetologica, (In Press).
Pnrr.lrps, D. M. & Asl, C, S., 1993. - Strategies for formation of the midpiece. In: B. BAccErrI, ComparativeSpermatology 20 Years A,fer. New York, Raven Press: 997-1000.
RIeppBI-, O., 1988. - The classification of the Squamata. In: M. J. BENroN, The Plrylogeny and Classification of theTetrapods. Oxford, Clarendon Press: 261-293.
SAITA, 4., CoMAzzr, M. & Pennorre, E., 1988a. - New data at the E. M. on the spermiogenesis of lguanadelicatissima Laurent involving comparative significance. Acta Embryologiae et Morphologiae Experimentalis(New Series), 9: 105-1 14.
Serra, A., COMAZZL M. & PERRorrA, 8., 1988b. - Ulteriori osservazioni al M. E sulla spermiogenesi di unserpente: Coluber viridiflavus (Lacepede) in riferimento ad elementi comparativi nella spermiogenesi dei rettili.Atti della Accademia Nazionale dei Lincei. Rendiconti. Classe di Scienze Fisiche, Matematiche e Naturali,LXXXII: 137-143.
Sorslo, J. R, & TnunLLo-CsNOz, O., 1958. - Electron microscope study of the kinetic apparatus in animal spermcells. Zeitschrift filr Zellfurschung, 48: 565-601.
Swoppoxo, D., 1993. - PAUP: Phylogenetic analysis using parsimony, version 3.0s. Computer programdistributed by the Illinois Natural History Survey, Champaign, Illinois.
Tuznr, O. & Bouncer, R., 1973. - Recherches ultrastructurales sur la spermiogenBse de Chamaeleo senegalensisOudin 1802. Bulletin Biologique de l.a France et de Ia Belgique,l0Tl. 195-212.
Ultrastructure of Spermatozoa of Australian Blindsnakes,Ramphotyphlops spp. (Typhlopidae, Squamata):First Observations on the Mature Spermatozoon
of Scolecophidian Snakes
H. Ronnie Hl,notNG * 'F*{<, Ken P. Apurv ** & Maria MAZUR *
x Institute of Environmental Studies,University of New South Wales, Sydney, 2052, NSW, Australia
** Western Australian Museum, Francis Street, Perth, 6000, Western Australia, Australiaxxx Honorary Research Associate at the Department of Anatomy and Histology, University of Sydney
ABSTRACT
The structure of mature, epididymidal spermatozoa of three species of Australian blindsnakes, Ramphotyphlops spp.(Typhlopidae, Squamata) was investigated by a combination of scanning and transmission electron microscopy. Specialattention was paid to features of potential phylogenetic significance, in the hope that aspects of sperm morphologymight shed light on the evolutionary affinities of this highly specialized and enigmatic group of reptiles. Spermatozoa ofRamphotyphlopr spp. share many features in common with other squamates (lizards and snakes), but display somestriking features not known in any other taxa. Special derived features shared by Ramphotyphlops spp. and otherSquamata include: a paracrystalline subacrosomal cone; dense intermitochondrial "rings" or "plaques"; a relativelyinconspicuous annulus; an anteriorly extensive fibrous sheath which penetrates the midpiece; and the absence of anendonuclear canal. Additional derived features which may be shared exclusively by Ramphotyphlops spp. and other snakesinclude: the great elongation of the midpiece; the forward prolongation of the fibrous sheath to the level of the neck; thepresence of a distinct "neck cylinder"; the extremely elongate form of the mitochondria; and the presence of a multi-layered midpiece plasma membrane. These cladistic interpretations lend support to the current taxonomic placement ofTyphlopidae within Squamata, and specifically within Serpentes. Several highly distinctive features of Ramphotyphlopsspermatozoa are identified, the most striking being the extreme length and regular zig-zagging arrangement of themitochondria.
nssuN4EUlt rast ructure des spermatozoides des serpents aveugles austra l iens, Ramphotyphlops spp.(Typhlopidae, Squamata) : premibres observat ions sur le spermatozoide mf i r des SerpentsS c o l e c o p h i d i e n s .
La structure des spermatozoides m0rs dpididymaires de trois espdces de serpents aveugles australiens. Ramphotyphlopsspp. (Typhlopidae; Squamata) a 6tE 6tudi6e en microscopie 6lectronique d balayage et i transmission. Une attentionparticulibre a 6td port6e aux d6tails d'importance phylog€nique potentielle, dans I'espoir que des caract€ristiques de lamorphologie du spermatozoide puissent 6clairer les affinit6s 6volutives de ce groupe de Reptiles, trbs 6volu6s et
HARDING, H. R., AplrN, K. P., & M^qzun, M., 1995. - Ultrastructurg-of spermatozoa of australian blindsnakes,Ramphotyphlops spp. (Typhlopidae, Squamata): first observations on the mature spermatozoon of scolecophidiansnakes. 1n: J.qvrssoN, B. G. M., Ausro, J., & JusrrNs, J.-L. (eds), Advances ih Spermatozoal Phylogeny and Taxonomy.Mdm. Mus. natn. Hist. nat.. 166: 385-396. Paris ISBN : 2-85653-225-X.