*Authors for correspondence: rdillhoff@evolvingearth.org, tdillhoff@evolvingearth.org, [email protected]ABSTRACT A Late Miocene flora (11.4 ± 0.61 Ma) from Bellevue, Washington, is described using macro- and microfossils to summarize the flora and estimate climate and vegetation coverage. Genera recognized from macrofossils include pteridophytes (Equisetum L., Woodwardia Sm., and Allanto- diopsis Knowlt. & Maxon, an apparent relict from the Eocene Puget Group flora), a rare cupres- saceous conifer, and common angiosperms (Platanus L., Cercidiphyllum Siebold & Zucc., Ulmus L., Fagus L., Hydrangea L., Alnus Mill., Acer L., and Persea Mill.). Including unidentified morpho- types, the macroflora records one bryophyte, six pteridophytes, one gymnosperm, and 24 angio- sperms. Well-preserved fossil pollen from the same sediments reveals a much richer regional flora. Although cupressaceous pollen is the most common element, a diverse record of Pinaceae is re- vealed by the palynoflora, including Abies Mill., Picea A. Dietr., Pinus L., Pseudotsuga Carrière/ Larix Mill., and Tsuga Carrière. In contrast, the Pinaceae are totally absent from the macrofossil record. Additional angiosperm genera identified from pollen include Liquidambar L., Ilex L., cf. Sarcobatus Nees, Carya Nutt., Juglans L., Nyssa L., Fraxinus L., and Salix L. Significant rare elements in the pollen flora include representatives of the Poaceae and Asteraceae. In total the palynomorph assemblage documents more than twice the diversity found in the macroflora. Coexistence analysis based on the pollen assemblage estimates mean annual temperature (MAT) of 11.6°C–16.6°C. Together, the macroflora and palynofloras suggest that Pacific Northwest plant communities—and climates—had not changed substantially since the Middle Miocene. Phyto- liths recovered from the sediments suggest a closed-canopy forest with warm-adapted grasses of the PACMAD (Panicoideae, Arundinoideae, Chloridoideae, Micrairoideae, Aristidoideae, and Danthonioideae) clade in the understory or in forest gaps. Geological and botanical evi- dence indicate that the fossils are preserved in flood overbank deposits associated with widespread Late Miocene conglomerates found throughout western Washington. Key words: geology, Miocene, paleobotany, palynology, phytoliths. CHAPTER 5 THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A.: A WINDOW INTO THE LATE MIOCENE OF THE PACIFIC NORTHWEST Richard M. Dillhoff,* omas A. Dillhoff,* Anthony P. Jijina, and Caroline A. E. Strömberg*
ABSTRACTA Late Miocene flora (11.4 ± 0.61 Ma) from Bellevue, Washington, is described using macro-
and microfossils to summarize the flora and estimate climate and vegetation coverage. Genera recognized from macrofossils include pteridophytes (Equisetum L., Woodwardia Sm., and Allantodiopsis Knowlt. & Maxon, an apparent relict from the Eocene Puget Group flora), a rare cupres-saceous conifer, and common angiosperms (Platanus L., Cercidiphyllum Siebold & Zucc., Ulmus L., Fagus L., Hydrangea L., Alnus Mill., Acer L., and Persea Mill.). Including unidentified morpho-types, the macroflora records one bryophyte, six pteridophytes, one gymnosperm, and 24 angio-sperms. Well-preserved fossil pollen from the same sediments reveals a much richer regional flora. Although cupressaceous pollen is the most common element, a diverse record of Pinaceae is re-vealed by the palynoflora, including Abies Mill., Picea A. Dietr., Pinus L., Pseudotsuga Carrière/Larix Mill., and Tsuga Carrière. In contrast, the Pinaceae are totally absent from the macrofossil record. Additional angiosperm genera identified from pollen include Liquidambar L., Ilex L., cf. Sarcobatus Nees, Carya Nutt., Juglans L., Nyssa L., Fraxinus L., and Salix L. Significant rare elements in the pollen flora include representatives of the Poaceae and Asteraceae. In total the palynomorph assemblage documents more than twice the diversity found in the macroflora. Coexistence analysis based on the pollen assemblage estimates mean annual temperature (MAT) of 11.6°C–16.6°C. Together, the macroflora and palynofloras suggest that Pacific Northwest plant communities—and climates—had not changed substantially since the Middle Miocene. Phyto-liths recovered from the sediments suggest a closed-canopy forest with warm-adapted grasses of the PACMAD (Panicoideae, Arundinoideae, Chloridoideae, Micrairoideae, Aristidoideae, and Danthonioideae) clade in the understory or in forest gaps. Geological and botanical evi-dence indicate that the fossils are preserved in flood overbank deposits associated with widespread Late Miocene conglomerates found throughout western Washington.
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A.: A WINDOW INTO THE LATE MIOCENE OF THE PACIFIC NORTHWEST
Richard M. Dillhoff,* Thomas A. Dillhoff,* Anthony P. Jijina, and Caroline A. E. Strömberg*
05Graham.064_097.indd 64 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 65
We are pleased to participate in the Festschrift honoring Alan Graham on his 80th birthday. Al-an’s early fieldwork included the Middle Miocene sites at Trout and Sucker creeks in eastern Ore-gon. Alan used both macrofossils and palyno-morphs to describe the local and regional flora of southeastern Oregon in the Middle Miocene. We will follow in his footsteps, using macrofossils, the palynoflora, and phytoliths, in addition to sedimentology, to describe the flora, paleoclimate, and paleoenvironment of the Vasa Park locality in the Late Miocene and compare Vasa with other Northwest Miocene occurrences (Fig. 1).
The site is located in a narrow ravine on land owned by a private park on the southern shore of Lake Sammamish in Bellevue, Washington. The majority of sedimentary exposures in the imme-diate vicinity of Vasa Park are the nearshore ma-rine Oligocene sediments of the Blakely Forma-tion and extensive Quaternary glacial deposits (Fig. 2). Tectonic activity along the Seattle fault created small isolated exposures of Miocene con-tinental sediments, including the Vasa Park local-ity (Fig. 3). The Middle Miocene paleobotany of
eastern Washington, eastern Oregon, and adja-cent Idaho has been extensively documented by studies beginning over 100 years ago (Merriam, 1901; Knowlton, 1902, 1926; Chaney & Axel-rod, 1959; Graham, 1963, 1965; Smiley, 1963; Smiley & Rember, 1985). These floras have doc-umented floral evolution and vegetation change in the Pacific Northwest during this important time, characterized by substantial global warm-ing associated with the Mid-Miocene Climatic Optimum (MMCO; Zachos et al., 2001). How-ever, there is comparatively little data from the Late Miocene, especially in a coastal plain setting. As a result, our knowledge of the composition and structure of plant communities during the regional transition to a cooler and more seasonal climate has been extremely limited. Vasa Park provides an important new window into the Late Miocene environment of the Pacific Northwest.
Our data show that the Vasa Park flora is pre-served in flood overbank deposits of a fluvial sys-tem. Large cobble conglomerates bracketing the fossil-bearing sediments record a high-energy en-vironment, and the fossil-bearing sediments point
FIGURE 1. Pacific Northwest Miocene localities mentioned in the text.
05Graham.064_097.indd 65 7/25/14 12:32 PM
66 PALEOBOTANY AND BIOGEOGRAPHY
to a highly disturbed local environment subject to periodic inundation. The macroflora contains many species that grow in regularly disturbed fluvial environments and the most diverse assem-blage of ferns known from a Northwest Miocene locality. The palynoflora records a diverse regional flora containing many elements that are not found in the macroflora. The climate estimate based on the palynoflora is of a warm temperate environ-ment with ample precipitation during the grow-ing season, similar to other Northwest Middle
and Late Miocene deposits. Phytolith analysis shows the environment to have been primarily closed forest with only one horizon indicating a substantial presence of open habitat grasses.
GEOLOGIC SETTING
REGIONAL GEOLOGYThe Puget Lowland, a broad, low-lying region
between the Cascade Mountains to the east and the Olympic Mountains to the west, is part of a
FIGURE 2. Geologic map of Vasa Park locality and surrounding area (Booth et al., 2012).
05Graham.064_097.indd 66 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 67
forearc basin formed in the Early Cenozoic that preserved Paleocene oceanic volcanic rocks and Eocene through Quaternary marine and terrestrial volcanic rocks and sedimentary deposits. Climate changes in the Quaternary resulted in a thick blanket of glacial deposits mantling bedrock. Eocene to Quaternary volcanic and sedimentary rocks lie beneath the glacial deposits in the cen-tral Puget Lowland, and north to south shorten-ing warps these rocks into a series of east-west-trending folds (Wells et al., 1998; Brocher et al., 2001; Booth et al., 2004). The most prominent of these folds in the Seattle region are the New-castle Hills and Gold Mountain anticlines; to-gether these anticlines form a linear, northwest- trending bedrock extending almost 90 km across the central Puget Lowland (Mullineaux, 1970; Booth et al., 2012).
Outcrops of Miocene rocks at Vasa Park owe their exposure to profound tectonism along the north limb of the Newport Hills anticline, local-ized along a thrust fault—the Seattle fault— lying beneath the north limb of the anticline. Blakely et al. (2002), Johnson et al. (1994, 1999), and Lib-erty and Pratt (2008) identify several subparallel fault strands within an east-trending zone along the north side of the Newport Hills anticline using
high-resolution aeromagnetic, geologic mapping, and seismic reflection methods. Rocks within the Newport Hills anticline consist of Eocene non-marine arkosic sandstones with abundant plant fossils, Oligocene marine sandstones and siltstones with nearshore marine and plant fossil horizons, and minor amounts of conglomerate. These are overlain by Miocene nonmarine conglomerate, volcanic sandstone and siltstone, and mudstone, with local peat and tephra layers (Fig. 3). Booth et al. (2012) describe the Miocene deposits as outcrops of sandstone, conglomerate, tuff, and volcaniclastic sandstone above the southwest and southeast shores of Lake Sammamish. Interbeds of lignite contain nearly coherent logs. Bedding is concordant with underlying Oligocene marine deposits.
VASA PARK STRATIGRAPHIC SECTIONA 17-m-thick section at Vasa Park consists of
three main units: a lower conglomeratic unit, a middle fine-grained unit, and an upper con-glomeratic unit (the upper two parts are shown in a measured section on Fig. 4). Conglomerates are generally reddish brown (oxidized), matrix- supported pebble/cobble conglomerates with clast lithologies predominately of dark-colored, fine-
FIGURE 3. Stratigraphy of the Newport Hills anticline in the vicinity of Vasa Park. Redrawn from Liberty and Pratt (2008).
05Graham.064_097.indd 67 7/25/14 12:32 PM
68 PALEOBOTANY AND BIOGEOGRAPHY
grained crystalline rocks and fine-grained andes-itic and felsic volcanic rocks. The middle fine-grained part contains brownish gray tuffaceous siltstone, sandy siltstone, and silty sandstones. Volcanic fragments and tuffaceous material in finer-grained deposits commonly weather to clay. Lamination in finer-grained strata is common. A thick stratum of cross-bedded tuffaceous sand-stone lies near the base of the section.
The stratigraphic position and low degree of lithification of the strata exposed in the creek sec-tion led some previous workers to assign a Qua-ternary age to these deposits (for example, Liesch et al., 1963). However, potassium-argon (K-Ar) ages from one nearby locality range from 9.3 to
14.7 Ma (Yount & Gower, 1991), and subsequent argon-argon (Ar/Ar) laser fusion dating from this same unit yielded an age of 11.40 ± 0.61 Ma (Booth et al., 2012).
Lithologies, sedimentary structures, and plant fossils suggest deposition in an alluvial system is-suing from and between Miocene stratovolcanos located to the east. Conglomerates and interbed-ded sandstones are indicative of deposition in high-energy, nonglacial braided stream channels, and finer-grained sandstone and siltstone were de-posited on the stream’s floodplain. However, strata of fossiliferous silt, clay, and log-bearing lignite indicate that other parts of the deposit are fluvio- lacustrine in origin. The presence of poorly sorted,
FIGURE 4. Stratigraphic section of Vasa Park sedimentary rocks. Burke Museum of Natural History and Culture, University of Washington (UWBM) locality numbers begin with “B”; UWBM sample numbers begin with “PB.”
05Graham.064_097.indd 68 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 69
thin, and interbedded fluvial and lacustrine sed-iments at Vasa Park suggests deposition in a low-relief piedmont environment characterized by floodplains and small lakes (possibly oxbow lakes).
CORRELATIVE UNITSExposures of Middle Miocene conglomerate
and sandstone deposits throughout the Puget Lowland attest to widespread alluvial deposition in the Middle Miocene: the Hammer Bluff For-mation near Enumclaw, Washington; the Mashel Formation near Orting, Washington; the Wilkes Formation near Toledo, Washington; and the Boundary Bay Formation (found in subsurface only) near Bellingham, Washington (Fig. 1). These deposits, like those at Vasa Park, suggest preva-lent low-relief piedmont environments charac-terized by floodplains and small lakes in western Washington, with low hills to the east and west at the present locations of the Cascade and Olympic mountains (e.g., see Mullineaux, 1970). Leopold and Denton (1987) showed that the Columbia Plateau in eastern Washington did not take on its modern character until Pliocene or even Pleisto-cene time, suggesting that a significant moisture barrier at the present-day location of the Cascade Mountains did not exist in the Middle Miocene.
Middle to Late Miocene siltstones, sandstones, and conglomerates of the Astoria and Montesano formations outcrop west of Olympia, Washing-ton, and fossils within each formation demon-strate deposition in shallow-water marine environ-ments (Rau, 1967). The location of these marine deposits of similar age as nonmarine deposits found in the Puget Lowland suggest that the Mid-dle to Late Miocene shoreline probably lay west of present-day Olympia.
MATERIALS AND METHODS
The Vasa Park locality occurs in a steep canyon that starts at the I-90 freeway and ends near the southwestern shore of Lake Sammamish in King
County, Washington, located in T24N R5E. The exposure consists of approximately 17 m of mud-stone, siltstones, and sandstone bracketed by massive beds of coarse, well-rounded basaltic conglomerate that has been documented across the Puget Sound region in association with the Seattle fault (see above; Blakely et al., 2002). Con-glomerate layers often contain woody debris up to 50 cm in length, whereas leaves are preserved in fine-grained sandstones. Plant fossils (Figs. 5–7) typically occur along bedding planes separated by 1- to 5-cm sections of barren rock. The macro-flora locality is designated as Burke Museum of Natural History and Culture, University of Wash-ington locality UWBM 6816 (see Figs. 8 and 9 for microflora locality numbers within the section). Macrofossils were morphotyped using methods from the Manual of Leaf Architecture (Ellis et al., 2009).
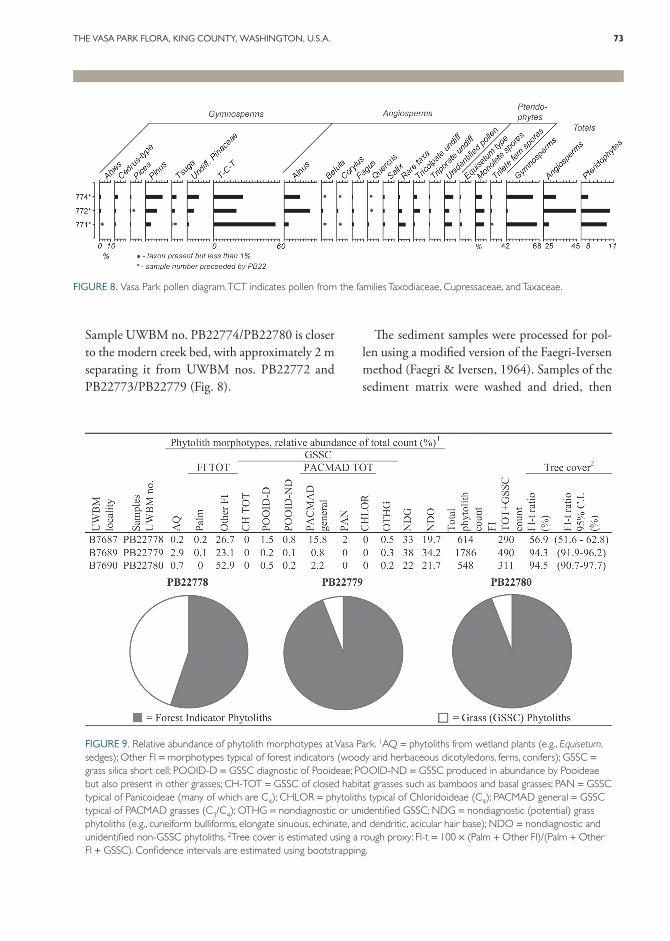
Sediment samples (Figs. 8, 9) for pollen and phytolith analysis were collected from two main horizons. We did not attempt to collect incre-mental samples from the entire section, which consists largely of non-fossiliferous (based on preliminary sampling) sandstone; rather, in an attempt to provide an overview of the microfloral assemblages of the deposit, samples were collected from fine-grained exposures that are more likely to contain palynomorphs and phytoliths. Incre-mental stratigraphic sampling would be further complicated at this site because it is heavily vege-tated, thus there is not a continuous exposure of the sediments. Two sediment samples (resulting in pollen or phytolith sample/specimen UWBM nos. PB22777 and PB22771/PB22778; see Fig. 9) were collected from a layer 0–75 cm below the leaf-bearing layer in gray and tan massive mud-stone with oxidation and fine rooting sediment. Another four samples (resulting in pollen or phy-tolith sample/specimen UWBM nos. PB22772, PB22773/PB22779, and PB22774/PB22780; Figs. 8, 9) were collected from a single, fine-grained siltstone leaf-bearing exposure between 11.5 and 12 m from the base of the section.
05Graham.064_097.indd 69 7/25/14 12:32 PM
70 PALEOBOTANY AND BIOGEOGRAPHY
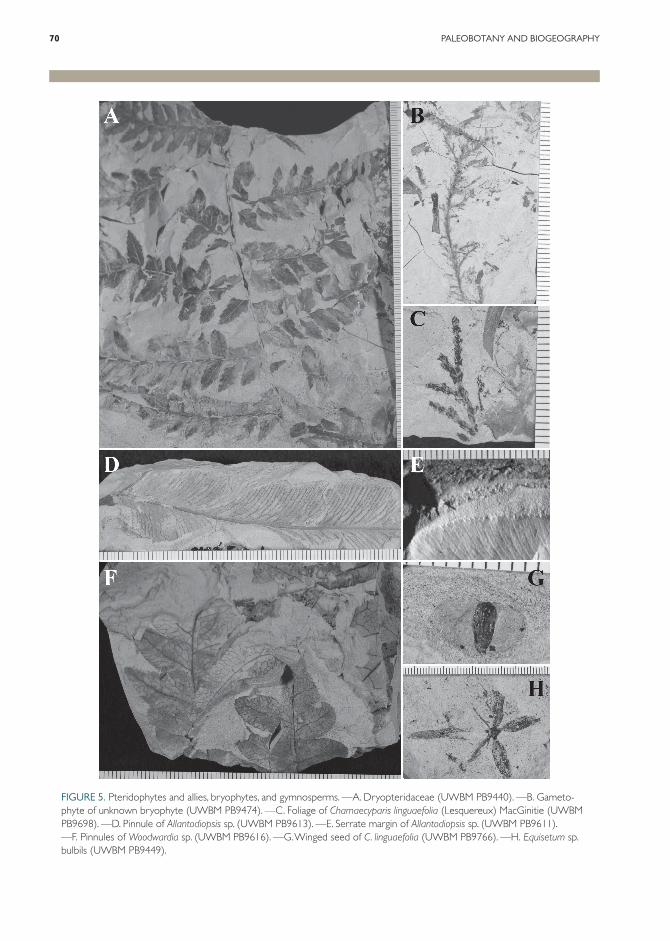
FIGURE 5. Pteridophytes and allies, bryophytes, and gymnosperms. —A. Dryopteridaceae (UWBM PB9440). —B. Gameto-phyte of unknown bryophyte (UWBM PB9474). —C. Foliage of Chamaecyparis linguaefolia (Lesquereux) MacGinitie (UWBM PB9698). —D. Pinnule of Allantodiopsis sp. (UWBM PB9613). —E. Serrate margin of Allantodiopsis sp. (UWBM PB9611). —F. Pinnules of Woodwardia sp. (UWBM PB9616). —G. Winged seed of C. linguaefolia (UWBM PB9766). —H. Equisetum sp. bulbils (UWBM PB9449).
05Graham.064_097.indd 70 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 71
FIGURE 6. Vasa Park angiosperms. —A. Leaf of Acer sp. (UWBM PB89436). —B. Samara of Acer sp. (UWBM PB9961). —C. Leaf of Cercidiphyllum crenatum (Unger) Brown (UWBM PB9964). —D. Leaf of Fagus washoensis LaMotte (UWBM PB9538). —E. Leaf of Persea pseudocarolinensis Lesquereux (UWBM PB9960). —F. Leaf of cf. Gossypium sp. (UWBM PB9961). —G. Leaf of Smilax magna Chaney (UWBM PB9732).
05Graham.064_097.indd 71 7/25/14 12:32 PM
72 PALEOBOTANY AND BIOGEOGRAPHY
FIGURE 7. Vasa Park angiosperms continued. —A. Leaf of Platanus dissecta Lesquereux (UWBM PB9440). —B. Detail of A showing teeth. —C. Leaf of Aesculus sp. (UWBM PB9448). —D. Hydrangea bendirei (Ward) Knowlton (UWBM PB9944). —E. Detail of C showing teeth. —F. Infructescence of Alnus sp. (UWBM PB9605). —G. Leaf of Alnus sp. (UWBM PB9955). —H. Leaf of Pauwlonia columbiana Smiley (UWBM PB9588).
05Graham.064_097.indd 72 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 73
Sample UWBM no. PB22774/PB22780 is closer to the modern creek bed, with approximately 2 m separating it from UWBM nos. PB22772 and PB22773/PB22779 (Fig. 8).
The sediment samples were processed for pol-len using a modified version of the Faegri-Iversen method (Faegri & Iversen, 1964). Samples of the sediment matrix were washed and dried, then
FIGURE 8. Vasa Park pollen diagram. TCT indicates pollen from the families Taxodiaceae, Cupressaceae, and Taxaceae.
FIGURE 9. Relative abundance of phytolith morphotypes at Vasa Park. 1AQ = phytoliths from wetland plants (e.g., Equisetum, sedges); Other FI = morphotypes typical of forest indicators (woody and herbaceous dicotyledons, ferns, conifers); GSSC = grass silica short cell; POOID-D = GSSC diagnostic of Pooideae; POOID-ND = GSSC produced in abundance by Pooideae but also present in other grasses; CH-TOT = GSSC of closed habitat grasses such as bamboos and basal grasses; PAN = GSSC typical of Panicoideae (many of which are C4); CHLOR = phytoliths typical of Chloridoideae (C4); PACMAD general = GSSC typical of PACMAD grasses (C3/C4); OTHG = nondiagnostic or unidentified GSSC; NDG = nondiagnostic (potential) grass phytoliths (e.g., cuneiform bulliforms, elongate sinuous, echinate, and dendritic, acicular hair base); NDO = nondiagnostic and unidentified non-GSSC phytoliths. 2Tree cover is estimated using a rough proxy: FI-t = 100 × (Palm + Other FI)/(Palm + Other FI + GSSC). Confidence intervals are estimated using bootstrapping.
05Graham.064_097.indd 73 7/25/14 12:32 PM
74 PALEOBOTANY AND BIOGEOGRAPHY
ca. 3 g of shale were crushed with a mortar and pestle and treated with 1 M hydrochloric acid (HCl) to remove carbonates. The samples were then rinsed with deionized water followed by an overnight soak in cold 48% hydrofluoric acid to remove silicates. The remaining material was rinsed twice in 1N HCl followed by two addi-tional rinses with deinoized water to remove the acid residuals. After rinsing, the samples were washed through a 185-µm mesh Nitex filter to remove large particles, then through a 10-µm mesh Nitex filter to remove fine particles. The sample fraction remaining on the 10-µm mesh filter was treated by acetolysis to remove excess organics. Slides were prepared from the processed sample by mounting in glycerin jelly treated with 2% safranin dye. Of the four samples prepared for pollen analysis, one (UWBM PB22773) pre-served only an insignificant amount of pollen and spores. The other three samples contained suffi-cient pollen and spores to allow detailed counting and analysis (Figs. 10–12).
Pollen slides were viewed at ×400 magnifica-tion under an optical light microscope to identify and photograph the different palynomorphs, and then counts of at least 500 grains were made for each sample to obtain relative abundances for the palynoflora. Pollen and spores were identified based on available literature (e.g., Wodehouse, 1935; Erdtman, 1952, 1957) and by comparison with modern pollen slides from the Jane Gray collection at the Burke Museum of Natural His-tory and Culture, University of Washington.
Paleoclimate was estimated from the pollen assemblage using the coexistence approach devel-oped by Mosbrugger and Utescher (1997). This method provides quantitative climate data based on the assumption that fossil taxa will have simi-lar climatic requirements as their modern nearest living relative. After fossil pollen from the Vasa Park site was identified to the genus or family level as appropriate, quantitative climatic data for the modern representatives of these taxa were ob-tained from the Palaeoflora database (<http://www
.palaeoflora.de>) and analyzed using the ClimStat program (Heinemann, 1998–1999). This program determines the zone where the climate require-ments for the nearest living relatives of identified taxa overlap and provides a quantitative range for the analyzed parameters, including mean annual temperature, cold month mean temperature, warm month mean temperature, mean annual precipi-tation, wet month precipitation, dry month pre-cipitation, and warm month precipitation.
In addition to the samples collected for both pollen and phytolith extraction, three sediment samples from other fine-grained paleosols (entisol; Soil Survey Staff, 2006) levels, as marked by root traces, were also collected, because fossil soils have been shown to contain the highest concentra-tions of phytoliths (Strömberg, 2004). Phytolith extraction was carried out following methods modified from Strömberg (2003, 2005). Ap-proximately 1.2 g of sediment was treated with concentrated (12 M) HCl to remove carbonates, sieved through a 250-µm sieve to remove coarse particles, and treated with heated Schultze’s solu-tion to remove organic compounds. Clay parti-cles were removed after deflocculation by sieving through a 53-µm sieve and repeated centrifuging. Finally, biogenic silica was isolated using heavy liquid (ZnBr+HCl+H2O). From the extracted silica, multiple fixed microscope slides were pre-pared for counting and analysis using an optical light microscope under ×1000 magnification with immersion oil.
In samples considered sufficiently productive (UWBM nos. PB22778, PB22779, PB22780), at least 250 (290–490) diagnostic phytoliths (see definition below) were classified along with non-diagnostic phytoliths, to provide statistically ro-bust counts for vegetation analysis (Strömberg, 2009), resulting in total counts of 548 to 1786 (Fig. 5). Rare phytolith morphotypes were noted to establish the presence of certain taxa, but not factored into the counts. Other biogenic silica, in-cluding diatoms, sponge spicules, and chrysophyte cysts, were noted for presence but not quantified.
05Graham.064_097.indd 74 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 75
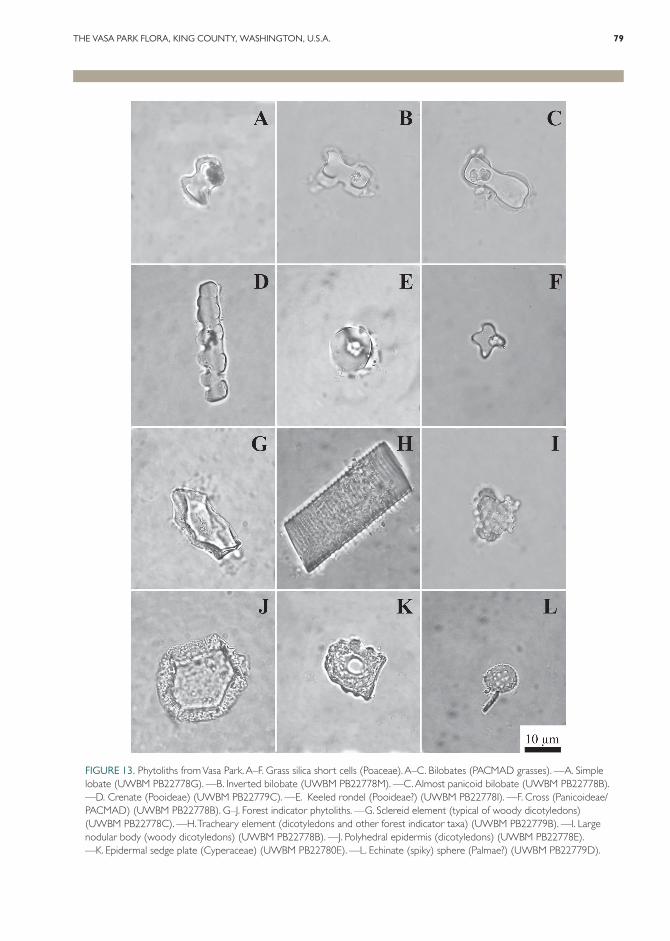
The fossil phytoliths were classified according to a scheme outlined in Strömberg (2004, 2005) and Strömberg et al. (2007a, 2007b) based on the literature (e.g., Piperno, 1988, 2006) and on description and analysis of a modern reference collection (Strömberg, 2003, unpublished data). Morphotypes were grouped into the following major categories: (1) forest indicator phytoliths (FI TOT), forming primarily in palms, woody or herbaceous dicotyledons, conifers, and ferns (Fig. 13G–J, L); (2) grass silica short cells (GSSC), found only in grasses, including forms indicative of closed-habitat grasses, such as bambusoid and basal grasses, and open-habitat grasses, such as pooid and PACMAD (Panicoideae, Arundinoi-deae, Chloridoideae, Micrairoideae, Aristidoideae, and Danthonioideae) grasses (Duvall et al., 2007; Aliscioni et al., 2012) (Fig. 13A–F); (3) phyto-liths indicative of taxa typically growing in wet-lands (AQ), including Equisetum L. and sedges (Fig. 13K); and (4) various nondiagnostic and unclassified forms.
Diagnostic phytoliths (FI TOT and GSSC) were used for reconstruction of vegetation type (e.g., forest, grassland) following Strömberg (2005) and Strömberg et al. (2007a, 2007b, 2013); specifically, types and abundances of forest in-dicator forms relative to grass silica short cells, as well as the composition of the GSSC assem-blages, were examined. The so-called FI-t ratio (FI TOT/(FI TOT + GSSC)%) provided a rough estimate of tree cover; bootstrap 95% confidence intervals (unconditional case, using the total count as the sample size) for this metric was estimated using the software Resampling Stats (Simon, 1997). FI-t does not directly translate to percent tree cover, but a comparison of FI-t through time or space can allow inference of trends in openness through time (Strömberg, 2004, 2005, 2009).
Grass community composition was inferred by analyzing relative abundances of different grass types within the GSSC assemblages. Using boot-strapping methods (Resampling Stats), the fossil samples were statistically compared to grasses in a
modern reference collection to account for over-lap in morphotype production among major grass groups (see Strömberg, 2005, for detailed meth-ods). Specifically, because the samples contained high relative abundance of PACMAD grasses they were tested to determine if pooid grasses may also have contributed to the GSSC assemblages (Strömberg 2005, 2009). In addition, the pres-ence of particularly diagnostic morphotypes (cer-tain crenate GSSC) was documented. Proximity to water is indicated by relative frequency of sedge phytoliths, as well as semiquantitative estimates of relative abundance of diatoms, chrysophyte cysts, and sponge spicules (Strömberg, 2005).
RESULTS
FLORISTIC STUDYTaxa identified from macrofossil (Figs. 5–7)
and palynomorph remains (Figs. 10–12) are sum-marized in Table 1.
Pteridophytes and allies (Figs. 5, 11).Bryophyta. The vegetative gametophyte of
an unknown bryophyte (Fig. 5B) is preserved at Vasa Park.
Lycopodiaceae. Lycopodium L. spores (Fig. 11A) are uncommon but present in all samples.
Selaginellaceae. Selaginella P. Beauv. spores with intact endospore (Fig. 11C) occur rarely in the pollen and spore assemblage.
Equisetaceae. Equisetum L. sp. (Fig. 5H). Compressions of small woody underground bul-bils arranged radially in groups of five are rela-tively common at Vasa Park. A few specimens representing nodal sheaths from fertile shoots have also been recognized (not figured). This rep-resents the first confirmed occurrence of Equisetum macrofossils in the Miocene of Washington. The specimen figured as an underground stem of Equisetum from the Middle Miocene Latah For-mation by Knowlton (1926: plate IX, fig. 1) is a stem with attached Nordenskioldia Heer fruits. An Equisetum rhizome and stem were found at the
05Graham.064_097.indd 78 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 79
FIGURE 13. Phytoliths from Vasa Park. A–F. Grass silica short cells (Poaceae). A–C. Bilobates (PACMAD grasses). —A. Simple lobate (UWBM PB22778G). —B. Inverted bilobate (UWBM PB22778M). —C. Almost panicoid bilobate (UWBM PB22778B). —D. Crenate (Pooideae) (UWBM PB22779C). —E. Keeled rondel (Pooideae?) (UWBM PB22778I). —F. Cross (Panicoideae/PACMAD) (UWBM PB22778B). G–J. Forest indicator phytoliths. —G. Sclereid element (typical of woody dicotyledons) (UWBM PB22778C). —H. Tracheary element (dicotyledons and other forest indicator taxa) (UWBM PB22779B). —I. Large nodular body (woody dicotyledons) (UWBM PB22778B). —J. Polyhedral epidermis (dicotyledons) (UWBM PB22778E). —K. Epidermal sedge plate (Cyperaceae) (UWBM PB22780E). —L. Echinate (spiky) sphere (Palmae?) (UWBM PB22779D).
05Graham.064_097.indd 79 7/25/14 12:32 PM
80 PALEOBOTANY AND BIOGEOGRAPHY
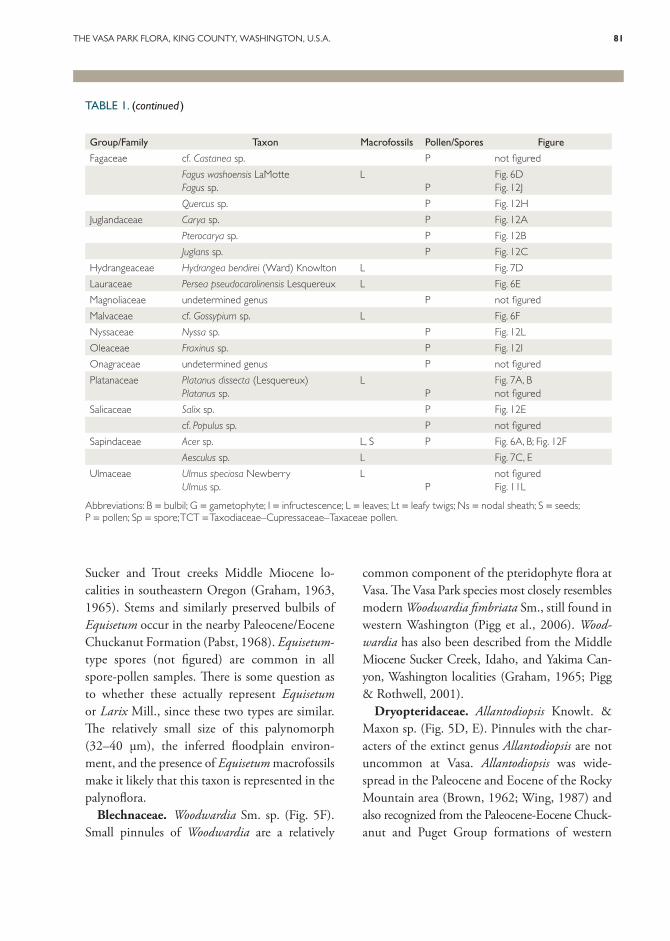
TABLE 1. Vasa Park combined flora from macrofossils and palynomorphs.
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 81
Sucker and Trout creeks Middle Miocene lo-calities in southeastern Oregon (Graham, 1963, 1965). Stems and similarly preserved bulbils of Equisetum occur in the nearby Paleocene/Eocene Chuckanut Formation (Pabst, 1968). Equisetum- type spores (not figured) are common in all spore-pollen samples. There is some question as to whether these actually represent Equisetum or Larix Mill., since these two types are similar. The relatively small size of this palynomorph (32–40 µm), the inferred floodplain environ-ment, and the presence of Equisetum macrofossils make it likely that this taxon is represented in the palynoflora.
Blechnaceae. Woodwardia Sm. sp. (Fig. 5F). Small pinnules of Woodwardia are a relatively
common component of the pteridophyte flora at Vasa. The Vasa Park species most closely resembles modern Woodwardia fimbriata Sm., still found in western Washington (Pigg et al., 2006). Woodwardia has also been described from the Middle Miocene Sucker Creek, Idaho, and Yakima Can-yon, Washington localities (Graham, 1965; Pigg & Rothwell, 2001).
Dryopteridaceae. Allantodiopsis Knowlt. & Maxon sp. (Fig. 5D, E). Pinnules with the char-acters of the extinct genus Allantodiopsis are not uncommon at Vasa. Allantodiopsis was wide-spread in the Paleocene and Eocene of the Rocky Mountain area (Brown, 1962; Wing, 1987) and also recognized from the Paleocene-Eocene Chuck-anut and Puget Group formations of western
Abbreviations: B = bulbil; G = gametophyte; I = infructescence; L = leaves; Lt = leafy twigs; Ns = nodal sheath; S = seeds; P = pollen; Sp = spore; TCT = Taxodiaceae–Cupressaceae–Taxaceae pollen.
05Graham.064_097.indd 81 7/25/14 12:32 PM
82 PALEOBOTANY AND BIOGEOGRAPHY
Washington (Pabst, 1968; Wolfe, 1968). Wolfe (1968) differentiated A. pugetensis Wolfe based pri-marily on unbranched secondary veins. The Vasa species typically has branched secondaries, or occasionally twice branched, as seen in A. erosa (Lesquereux) Pabst.
The phylogenetic position of Allantodiopsis among the ferns is uncertain (Collinson, 2001). The association of the single specimen of fertile material associated with the common sterile foli-age that is the basis for the genus name has been questioned (McIver & Basinger, 1993). Similar foliage originally assigned to Allantodiopsis from the Eocene of Oregon was determined based on the cuticle to be an extinct cycad with fernlike foliage, Eostangeria barthel (Kvaček & Manches-ter, 1999). In that genus secondaries sometimes reunite at the margin, forming loops, but second-aries in the Vasa specimens do not. The Vasa Park occurrence dramatically extends the known strati-graphic occurrence of Allantodiopsis.
In addition to Allantodiopsis, foliage of a second member of the family, morphotype VP003 (Fig. 5A), is recognized as a member of the Dryopteri-daceae. In the absence of fertile material it is not possible to assign this material to a genus; it could represent a bipinnate Polystichum Roth or Dryopteris Adans. The pinnule morphology of the fossil resembles modern P. dudleyi Maxon found in coastal California. Spores of the Dryopteri-daceae type (Fig. 11D) are also present in the palynoflora.
Pteridophyte spores are diverse and constitute a significant percentage of all palynological sam-ples. Monolete spores without preserved perine of the Laevigatosporites Ibrahim type (not figured) are common throughout the deposit. In addition, there are four distinct types with well-preserved perine; three of these are figured (Fig. 11D–F). Trilete spores are less common but still diverse, with at least two types without preserved perine (Fig. 11B), and three distinct types with pre-served perine.
(Lesquereux) MacGinitie (Fig. 5C, G). Chamaecyparis Spach macrofossils have been identified from foliage, seeds, and a possible ovulate cone. The genus has also been identified from the Mid-dle Miocene at Clarkia, Idaho (Smiley & Rember, 1985) and Trout Creek in southeastern Oregon (Graham, 1963). MacGinitie (1933) identified this taxon at Trout Creek and later identified it as C. linguaefolia (MacGinitie, 1953). The Vasa Park species is consistent with the Trout Creek occurrence. Chamaecyparis was also identified in coeval sediments from the nearby Late Miocene Hammer Bluff Formation (Mullineaux, 1970).
Metasequoia Hu & W. C. Cheng/Sequoia Endl. type (Fig. 10F). Rare cupressaceous pollen grains with papillae are found in all samples. The over-all morphology is most similar to Metasequoia/ Sequoia, but other cupressaceous genera are known to have papillae, including Taxodium Rich. and Glyptostrobus Endl. All four of these genera have been reported as macrofossils from the Miocene of the Pacific Northwest (Chaney & Axelrod, 1959; Graham, 1965; Smiley & Rember, 1985). In the absence of co-occurring macrofossils from Vasa Park, it is difficult to assign this pollen type to a particular genus.
The palynoflora also contains a large propor-tion of cupressaceous Taxodiaceae–Cupressaceae– Taxaceae (TCT) pollen (Fig. 10D).
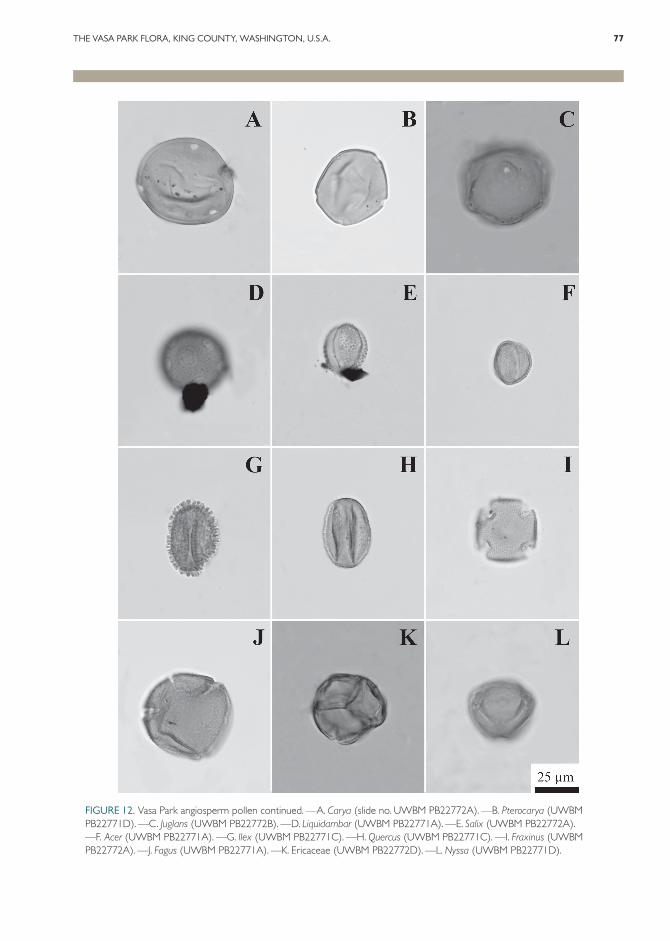
Pinaceae. Abies Mill. (Fig. 10I). Two distinct types of Abies pollen have been noted in the pal-ynoflora. Abies pollen is rare, but present in all pollen-bearing samples. The first type is quite large (155–187 µm) and is the one most often en-countered. The figured type is smaller and is sim-ilar to modern A. amabilis Douglas ex J. Forbes, which is extant in the Pacific Northwest.
Cedrus Trew sp. (Fig. 10H). Cedrus pollen is a minor element in all samples from Vasa Park. This type is widespread in the Miocene of the Pacific Northwest, although to our knowledge
05Graham.064_097.indd 82 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 83
there have been no reports of confirmed Cedrus macrofossils.
Picea A. Dietr. (Fig. 10A). There are two types of Picea pollen found in the Vasa Park palynoflora: one is particularly large at up to 170 µm, while the other is smaller (90–97 µm). Picea is well rep-resented throughout the Cenozoic of the Pacific Northwest and is widely reported in the Miocene.
Pinus L. sp. (Fig. 10B). Pinus is a common el-ement of the Vasa Park palynoflora, contributing between 5% and 15% of the total pollen count.
Pseudotsuga Carrière/Larix Mill. type (Fig. 10E). Pollen of Pseudotsuga and Larix is very similar, thus they are reported as a single palyno-morph. The large size of the grains (101–109 µm) indicates that they most likely represent Pseudotsuga. Pseudotsuga macrofossils are not common in the fossil record of the region, but foliage and cones have been reported from the Miocene of Oregon (Chaney & Axelrod, 1959; Graham, 1965). Recently, Pseudotsuga has been confirmed from the Middle Miocene of Washington based on well-preserved wood from the Ginkgo Petri-fied Forest (Wheeler & Dillhoff, 2009). To our knowledge, there have been no confirmed reports of Larix macrofossils from the Miocene of the Pacific Northwest.
Tsuga Carrière (Fig. 10C). Tsuga pollen is rep-resented by two distinct types. One type is simi-lar to modern T. heterophylla (Raf.) Sarg., which is extant in the Pacific Northwest. Similar pollen has been figured from the Clarkia deposit of Idaho (Gray, 1985). The second type is super-ficially similar to T. canadensis (L.) Carrière, but under high magnification it has visible spines, which are lacking in T. canadensis. This type may be more closely related to some of the modern Asiatic species of Tsuga, which are similar in mor-phology and have small spines.
Incertae sedis. There are two bisaccate pollen types (Fig. 10G) similar to those reported as Podocarpus Persoon or Podocarpidites Cookson ex Couper from other Cenozoic floras of the Pacific
Northwest (Barnett & Fisk, 1980; Reininck- Smith & Leopold, 2005). Zetter et al. (2011) state that pollen of this type may also represent Cathaya Chun & Kuang, and that the pollen can only be reliably distinguished using features visible under SEM. Because of this uncertainty, these types are reported as Podocarpaceae/Cathaya. There have been no confirmed macrofossils of Podocarpus reported from the Cenozoic of the Pacific Northwest, while cones and foliage with pre-served cuticle of Cathaya were described from the Middle Miocene Clarkia deposit of western Idaho (Kvaček & Rember, 2000).
Angiosperms, Monocotyledons.Liliaceae. Several pollen grains attributed to
the Liliaceae (Fig. 11O) were found at Vasa Park. These are rare, but present in both the upper and lower sampling horizons. Similar pollen was re-ported from the Miocene Palouse Falls locality of Washington, but not figured (Barnett & Fisk, 1980).
Poaceae. Several well-preserved grains of grass pollen (Fig. 11I) have been identified from all sample horizons at Vasa Park. This type is rare, but the presence of grasses at Vasa Park is corrob-orated by the phytolith analysis.
Potamogetonaceae. Rare pollen of Potamogeton L. (Fig. 11G) is present at Vasa Park.
Smilacaceae. Smilax magna Chaney (Fig. 6G). Smilax magna is preserved as rare examples at Vasa and was also present in the Middle Mio-cene Clarkia (Smiley & Rember, 1985), Mascall (Chaney and Axelrod, 1959), and Trout Creek (Graham, 1963) localities. It was also present in the Late Miocene Ellensburg Formation (Smiley, 1963). Chaney and Axelrod (1959) argued for designating all Northwest Miocene occurrences of Smilax L. by the epithet magna because species could not be reliably separated on leaf characters alone. The nearby Middle Eocene Puget Group deposits record the first known occurrence of Smilax in western Washington (Wolfe, 1968).
05Graham.064_097.indd 83 7/25/14 12:32 PM
84 PALEOBOTANY AND BIOGEOGRAPHY
Typhaceae. Pollen monads of the Typha L./Sparganium L. type (Fig. 11H) are found from the upper sampling horizon at Vasa Park. No macrofossils have been found.
Angiosperms, Dicotyledons.Altingiaceae. Liquidambar L. sp. (Fig. 12D).
Pollen grains of Liquidambar occur rarely at the site. No macrofossils have been found. Liquidambar is a common element in the Middle Mio-cene floras of the Pacific Northwest, and today has disjunct distribution in Asia and eastern North America and Central America.
Aquifoliaceae. Ilex L. sp. (Fig. 12G). Ilex is uncommon, but found in all pollen samples. No macrofossils have been found.
Asteraceae. Uncommon but distinctive grains of pollen from the Asteraceae (Fig. 11N) are pres-ent in samples from the upper horizon.
Betulaceae. Alnus Mill. sp. (Fig. 7G, F). Com-mon leaves and rare infructescences document the presence of Alnus at Vasa Park. Overlapping leaf morphologies preclude the separation of Alnus from Betula L. based on leaf characters alone. At Vasa Park the presence of Alnus repro-ductive structures and the overwhelming prepon-derance of Alnus over Betula pollen support an identification of Alnus for the leaves. The species A. relata (= A. relatus (Knowlton) Brown) is re-corded from the nearby coeval Mashel Formation (Walters, 1964) and Alnus pollen was also recov-ered from the Hammer Bluff Formation in Renton (Mullineaux, 1970). Alnus is common in most Northwest deposits from the Early Eocene onwards, and five species occur today in Wash-ington and Oregon (Franklin & Dyrness, 1988). The leaf characters of the Vasa Park Alnus corre-spond closely to modern A. sinuata (Regel) Rydb., which occurs in western Washington.
Several genera of Betulaceae are represented in the palynoflora, including Alnus (Fig. 11K), Betula (Fig. 11J), Corylus L. (not figured), and Ostrya Scop./Carpinus L. (not figured) type. Alnus is par-ticularly abundant, represented by 4-, 5-, 6-, and
7-pore forms (the 4- and 5-pore forms are pre-dominant). It comprises 4% of the pollen count in the lower sampling horizon, and up to 24% of the counts in the upper horizons, where it is a dominant type. The other genera of the Betulaceae are minor contributors to the overall palynoflora.
Cercidiphyllaceae. Cercidiphyllum crenatum (Unger) Brown (Fig. 6C). Cercidiphyllum crenatum is a common macrofossil species recovered at Vasa Park, as well as from the Middle Miocene in the Mascall Formation and at Clarkia (Chaney & Axelrod, 1959; Smiley & Rember, 1985). It is recorded from the Late Miocene Ellensburg Formation (Smiley, 1963) as well. Wolfe (1968) identified C. piperoides (Lesquereux) LaMotte from Middle Eocene Puget Group sediments in King County.
Rare grains of Cercidiphyllum Siebold & Zucc. type pollen (not figured) are found in the lower sampling horizon. These are consistent with mod-ern pollen of Cercidiphyllum, and are similar to fossil pollen from the Miocene of Oregon identi-fied as Cercidiphyllum by Gray (1958); however, the grains are folded and thus their identification is not certain.
Chenopodiaceae. Three pollen grains similar to that of the Chenopodiaceae (Fig. 11M) were identified from the Vasa Park palynoflora. Simi-lar pollen has been reported from other Miocene sites in the Pacific Northwest (Chaney & Axel-rod, 1959; Graham, 1965; Barnett & Fisk, 1980; Smiley & Rember, 1985). Graham (1965) notes that pollen of the Chenopodiaceae is morpho-logically similar to the Amaranthaceae and thus difficult to separate. Barnett and Fisk (1980) re-corded their specimens as Chenopodiaceae, cf. Sarcobatus Nees. The Vasa Park specimens are small in size (20–25 µm), have relatively few pores, and bear a strong resemblance to modern S. vermiculatus (Hook.) Torr. It should be noted that Sarcobatus was removed from the Chenopodiaceae/Amaranthaceae clade and assigned to the mono-generic family Sarcobataceae by Behnke (1997).We refer to the Vasa Park pollen type as cf. Che-
05Graham.064_097.indd 84 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 85
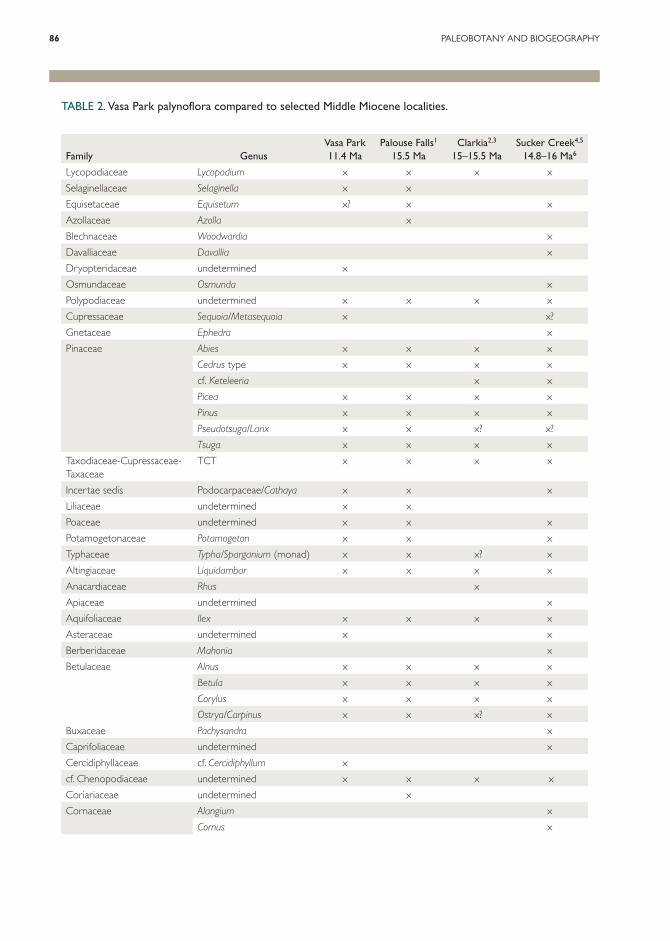
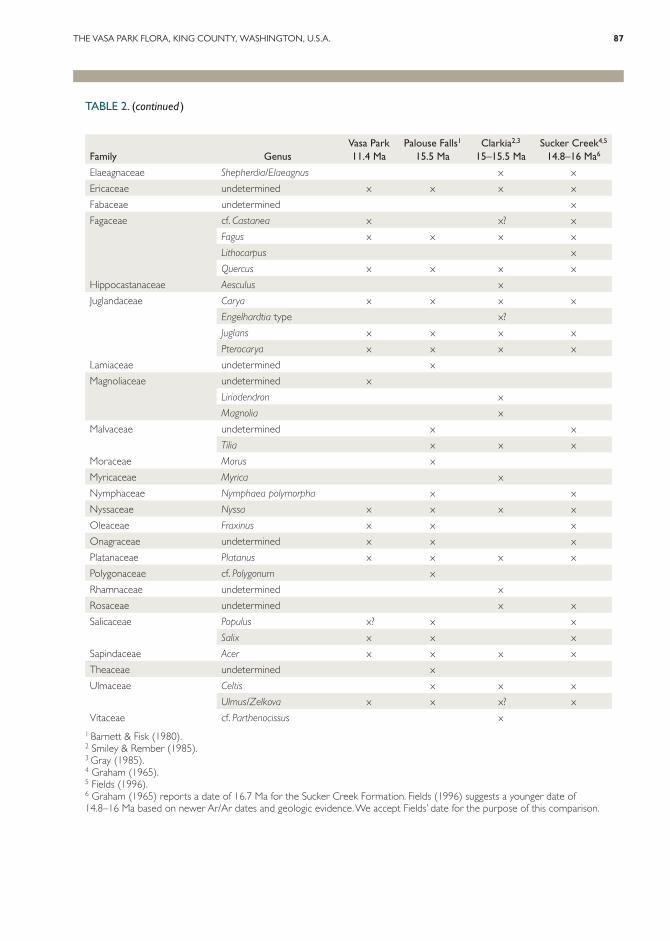
nopodiaceae to allow comparison with the previ-ously described palynofloras in Table 2.
Ericaceae. Rare tetrads of ericaceous pollen (Fig. 12K) were found in sample UWBM PB22772. These were not detected in any other samples.
Fagaceae. Fagus washoensis LaMotte (Fig. 6D). Common leaves at Vasa Park match the charac-ters of F. washoensis. This species was recorded in the Middle Miocene Mascall Formation (Chaney & Axelrod, 1959), at Trout and Sucker creeks (Graham, 1963), and Fagus L. also occurs at Clarkia (Smiley & Rember, 1985). The species is also recorded in Late Miocene deposits of the Ellensburg Formation (Smiley, 1963) and the nearby coeval Hammer Bluff Formation (Mul-lineaux, 1970).
The Fagaceae is also represented in the pollen assemblage by cf. Castanea Mill., Fagus sp., and two types of Quercus L. The Castanea type (not figured) is only represented by a few pollen grains. Fagus (Fig. 12J) and Quercus (Fig. 12H) are more common, especially from the lowest sampling horizon.
Hydrangeaceae. Hydrangea bendirei (Ward) Knowlton (Fig. 7D). Several well-preserved leaves of Hydrangea L. have been found at Vasa Park. The species was widespread in the Middle Mio-cene, occurring in the Latah (Knowlton, 1926; Brown, 1936), Mascall (Knowlton, 1902; Chaney & Axelrod, 1959), and Trout Creek and Sucker Creek (Graham, 1963) deposits.
Juglandaceae. The Juglandaceae is represented by pollen of Carya Nutt. (Fig. 12A), Juglans L. (Fig. 12C), and Pterocarya Kunth (Fig. 12B). Juglans is quite rare, with only two pollen grains found in the 16 slides examined for this study. Carya and Pterocarya are more common, but still compose less than 1% of the pollen in all of the samples that were counted. All three of these taxa are commonly reported in the palynofloras of the Pacific Northwest.
Lauraceae. Persea pseudocarolinensis Lesquereux (Fig. 6E). Persea pseudocarolinensis was a wide-spread species first recorded in King County from
the Middle Eocene Puget Group (Wolfe, 1968). This species is also found at all of the well-known Northwest Middle Miocene localities and the Late Miocene Ellensburg flora (Smiley, 1963). Persea lanceolata (Berry) Brown was identified by Wolfe (Walters, 1964) from the nearby coeval Mashel Formation, but no description or figure was provided. That species was separated from P. pseudocarolinensis based on a narrow-leaved morphotype of Persea found in the Latah flora.
Magnoliaceae. Only two pollen grains of the Magnoliaceae (not figured) have been identified from Vasa Park. No macrofossils have been identified.
Malvaceae. Cf. Gossypium L. sp. (Fig. 6F). Common leaves at Vasa Park have morphology similar to modern G. barbadense L. In both cases, leaves are palmate, 3- to 5-lobed, entire, lobes with acuminate tips, actinodromous with an ob-tuse to reflex cordate base, brochidodromous sec-ondaries, percurrent tertiaries, and an irregular reticulate fabric. Gossypol glands observable on modern leaves are possibly present in the fossil but equivocal. The stellate hairs present on mod-ern Gossypium leaves are not observed in the fossil. Graham (1963) described G. arnoldii Graham from Trout Creek based on leaf morphology, but no other Gossypium fossils have been described prior to the Pleistocene. The genus Gossypium has been estimated to have originated approximately 12.5 Ma (Wendel et al., 2010).
Nyssaceae. Nyssa L. pollen (Fig. 12L) is present but rare at Vasa Park. This genus was widespread in the Pacific Northwest during the Miocene.
Oleaceae. Rare examples of Fraxinus L. pollen (Fig. 12I) are present in the Vasa Park assem-blage. No macrofossils have been found.
Onagraceae. Only two grains of distinctive pollen (not figured) from the Onagraceae were noted at Vasa Park; both were from the upper sample horizon.
Platanaceae. Platanus dissecta Lesquereux (Fig. 7A, B). This widespread species has been found in the Northwest Middle Miocene at Clarkia
05Graham.064_097.indd 85 7/25/14 12:32 PM
86 PALEOBOTANY AND BIOGEOGRAPHY
TABLE 2. Vasa Park palynoflora compared to selected Middle Miocene localities.
Family GenusVasa Park 11.4 Ma
Palouse Falls1
15.5 MaClarkia2,3
15–15.5 MaSucker Creek4,5
14.8–16 Ma6
Lycopodiaceae Lycopodium x x x x
Selaginellaceae Selaginella x x
Equisetaceae Equisetum x? x x
Azollaceae Azolla x
Blechnaceae Woodwardia x
Davalliaceae Davallia x
Dryopteridaceae undetermined x
Osmundaceae Osmunda x
Polypodiaceae undetermined x x x x
Cupressaceae Sequoia/Metasequoia x x?
Gnetaceae Ephedra x
Pinaceae Abies x x x x
Cedrus type x x x x
cf. Keteleeria x x
Picea x x x x
Pinus x x x x
Pseudotsuga/Larix x x x? x?
Tsuga x x x x
Taxodiaceae-Cupressaceae- Taxaceae
TCT x x x x
Incertae sedis Podocarpaceae/Cathaya x x x
Liliaceae undetermined x x
Poaceae undetermined x x x
Potamogetonaceae Potamogeton x x x
Typhaceae Typha/Sparganium (monad) x x x? x
Altingiaceae Liquidambar x x x x
Anacardiaceae Rhus x
Apiaceae undetermined x
Aquifoliaceae Ilex x x x x
Asteraceae undetermined x x
Berberidaceae Mahonia x
Betulaceae Alnus x x x x
Betula x x x x
Corylus x x x x
Ostrya/Carpinus x x x? x
Buxaceae Pachysandra x
Caprifoliaceae undetermined x
Cercidiphyllaceae cf. Cercidiphyllum x
cf. Chenopodiaceae undetermined x x x x
Coriariaceae undetermined x
Cornaceae Alangium x
Cornus x
05Graham.064_097.indd 86 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 87
TABLE 2. (continued )
Family GenusVasa Park 11.4 Ma
Palouse Falls1
15.5 MaClarkia2,3
15–15.5 MaSucker Creek4,5
14.8–16 Ma6
Elaeagnaceae Shepherdia/Elaeagnus x x
Ericaceae undetermined x x x x
Fabaceae undetermined x
Fagaceae cf. Castanea x x? x
Fagus x x x x
Lithocarpus x
Quercus x x x x
Hippocastanaceae Aesculus x
Juglandaceae Carya x x x x
Engelhardtia type x?
Juglans x x x x
Pterocarya x x x x
Lamiaceae undetermined x
Magnoliaceae undetermined x
Liriodendron x
Magnolia x
Malvaceae undetermined x x
Tilia x x x
Moraceae Morus x
Myricaceae Myrica x
Nymphaceae Nymphaea polymorpha x x
Nyssaceae Nyssa x x x x
Oleaceae Fraxinus x x x
Onagraceae undetermined x x x
Platanaceae Platanus x x x x
Polygonaceae cf. Polygonum x
Rhamnaceae undetermined x
Rosaceae undetermined x x
Salicaceae Populus x? x x
Salix x x x
Sapindaceae Acer x x x x
Theaceae undetermined x
Ulmaceae Celtis x x x
Ulmus/Zelkova x x x? x
Vitaceae cf. Parthenocissus x1 Barnett & Fisk (1980).2 Smiley & Rember (1985).3 Gray (1985).4 Graham (1965).5 Fields (1996).6 Graham (1965) reports a date of 16.7 Ma for the Sucker Creek Formation. Fields (1996) suggests a younger date of 14.8–16 Ma based on newer Ar/Ar dates and geologic evidence. We accept Fields’ date for the purpose of this comparison.
05Graham.064_097.indd 87 7/25/14 12:32 PM
88 PALEOBOTANY AND BIOGEOGRAPHY
(Smiley & Rember, 1985), Mascall (Chaney & Axelrod, 1959), Sucker and Trout creeks (Gra-ham, 1965), and in the Late Miocene Ellensburg Formation (Smiley, 1963). The species was also recorded in the nearby coeval Mashel Formation (Walters, 1964), and Wolfe (1968) included Platanus L. in the flora of the Middle Eocene Puget Group in King County. Platanus pollen is a rare constituent of the Vasa Park palynoflora.
Salicaceae. This family is represented by pol-len of Salix L. (Fig. 12E) and possibly Populus L. (not figured). Salix pollen is relatively common, making up slightly over 1% of the assemblage in each sample that was counted. The cf. Populus pollen is rare, with only two grains identified. Gray (1985) notes that Populus pollen has low preservation potential, and thus its presence in the fossil record is sporadic.
Sapindaceae. Acer L. (Fig. 6A, B). Well- preserved leaves and samaras of Acer are com-mon at Vasa Park. The leaves range widely in size from a laminar length of 5–20 cm. They are shal-lowly lobate, 3- or 5-lobed, with a finely serrate margin. The leaves most closely resemble A. glabroides Brown, although A. glabroides is usually 3-lobed. The samaras found at Vasa Park do not match those associated with A. glabroides by pre-vious authors (Brown, 1936; Wolfe & Tanai, 1987). The samaras found at Vasa closely re-semble samaras of A. medianum Knowlton. Wolfe and Tanai (1987) argued for an association between samaras of A. medianum and leaves of A. columbianum Chaney & Axelrod based on the co-occurence of leaves of A. columbianum with samaras of A. medianum at eight of the 14 known localities where A. medianum samaras occur. But A. glabroides is also found at many of the same localities.
Multiple species of Acer are present at each of the Middle Miocene localities of the Pacific Northwest (Knowlton, 1902, 1926; Brown, 1936; Chaney & Axelrod, 1959; Graham, 1963; Smiley & Rember, 1985). In many cases the actual num-ber of species is overstated because isolated leaves
and samaras cannot be reliably associated with each other. Acer was also found at both of the coeval western Washington Late Miocene locali-ties of the Mashel and Hammer Bluff formations (Walters, 1964; Mullineaux, 1970).
Two types of Acer pollen (Fig. 12F) are also found at Vasa Park, although they are both rare.
Aesculus L. sp. (Fig. 7C, E). A few leaflets of Aesculus have been found at Vasa Park. Aesculus has also been identified at Clarkia (Smiley & Rember, 1985) but has not been recognized at other Pacific Northwest Miocene localities.
Scrophulariaceae. Paulownia columbiana Smi-ley (Fig. 7H). Distinctive palmate leaves with prominent tertiaries and a round to obtuse apex were described as P. columbiana by Smiley (1961) from the Late Miocene Ellensburg Formation and compared to extant P. tomentosa (Thunb.) Steud. Paulownia tomentosa has a wider range of leaf morphologies, including lobed leaves, while P. columbiana leaves are not lobed. Unlobed leaves of P. tomentosa and P. columbiana are entire, except for a few small teeth found near the leaf base. Excellent examples have been collected at Vasa Park. Paulownia Siebold & Zucc. is also found in the Middle Miocene at Clarkia (Smiley & Rem-ber, 1985), and Knowlton (1926) figured a Latah specimen as Acer (plate XXVIII, fig. 1) that likely represents this taxon. He also illustrates a Paulownia capsule (plate XXIX, fig. 12) that he de-scribed as Carpites paulownia Knowlton. Paulownia columbiana was also identified by Wolfe in the coeval Mashel Formation (Walters, 1964). Wolfe figures a leaf with the same characteristics from the Puget Group (Wolfe, 1968: plate 5, fig. 1) which he described as Macaranga pugetensis.
Ulmaceae. Ulmus speciosa Newberry (not fig-ured). Ulmus L. is another widespread genus found at most of the Middle and Late Miocene localities in the Pacific Northwest. Knowlton (1926) included U. speciosa in the Latah flora, and Chaney and Axelrod (1959) also identified it in the Mascall flora, but described a second species, U. paucidentata, based on smaller leaf size
05Graham.064_097.indd 88 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 89
and slender shape. That species was included in the Ellensburg Formation flora (Smiley, 1963), but the specimens at Vasa correspond best with the description of U. speciosa. This species was re-ported by Wolfe in the coeval Mashel Formation (Walters, 1964). Ulmus was also reported from the nearby Late Miocene Hammer Bluff For-mation (Mullineaux, 1970). Ulmus oregoniana Knowlton was recognized in the Middle Eocene Puget Group flora (Wolfe, 1968). Ulmaceous pollen (Fig. 11L) has been found in 4-, 5-, and 6-pored forms, although it is rare.
POLLEN RESULTSThe pollen in the three counted samples was
generally abundant and well preserved. In the in-terest of showing the most significant aspects of the palynoflora, the pollen diagram (Fig. 8) pre-sents only those taxa that contributed at least 1% of the total counts. Conifer pollen is dominant in all samples, with TCT being the major contribu-tor. Pinus also makes a strong showing, and Tsuga is important from the layers near the top of the exposure. Among angiosperms, Alnus is by far the major contributor, and is the single most common palynomorph in sample UWBM no. PB22772. Pteridophytes are an important constituent of all samples, making up approximately 8%–10% of the total counts. One aspect of the flora that is not clear from the pollen diagram is the high spe-cies richness in all three samples. We were able to distinguish over 100 distinct palynomorphs, with high species richness in all groups: pteridophytes, conifers, and angiosperms. Those types that were identifiable to the family or genus level are pre-sented in Table 2.
Because the macroflora assemblage was not suf-ficiently well preserved or diverse to allow a cli-mate estimate based on techniques using nearest living relatives or leaf morphology, we used the coexistence approach (Mosbruger & Utescher, 1997) based on the fossil pollen assemblage. Pol-len is adequate but not ideal for this purpose, since the coexistence approach works best with
fossils identified to the genus or species level. This allows for a better determination of possible nearest living relatives for a given fossil taxon and thus tighter constraints on the paleoclimate esti-mates. Since pollen is typically only identified at the family or genus level, the climatic resolution provided by the analysis is lower. The results of the coexistence analysis are as follows:
Mean annual temperature: 11.6°C–16.6°CCold month mean temperature: –0.3°C–2.9°CWarm month mean temperature: 23.0°C–
PHYTOLITH RESULTSThe extracted phytolith assemblages were
deemed well preserved, due to the presence of fine ornamentation on the grass silica short cells and the generally unaltered texture of the phyto-liths, despite some etching of larger bodies, and thus considered appropriate for quantitative phy-tolith analysis (Strömberg, 2003; Strömberg et al. 2007b). Although sample UWBM no. PB22779 had relatively low phytolith productivity, UWBM nos. PB22778 and PB22780 had extremely low productivity (13 slides had to be examined to achieve a sufficient count).
UWBM no. PB22778 has a relatively high abundance of FI-t phytoliths at 57% (95% CI: 52%–63%), including anticlinal epidermal cells, helical tracheary elements, and nodular bodies. A single palm phytolith was also noted, as well as rare sedge forms. UWBM no. PB22779 has a very high FI-t ratio (94%; 95% CI: 92%–96%), and contained forms such as nodular bodies and polyhedral epidermis, as well as a single palm phytolith. Similarly, UWBM no. PB22780 has a high FI-t ratio (94%; 95% CI: 91%–98%), with FI-TOT forms made up primarily by nodular bodies.
05Graham.064_097.indd 89 7/25/14 12:32 PM
90 PALEOBOTANY AND BIOGEOGRAPHY
In all three samples, GSSC assemblages are dominated by PACMAD forms (60%–86% of GSSC), represented by, for example, simple lo-bates, inverted bilobates, and near panicoid bilo-bates, with a smaller component of pooid mor-photypes (11%–24% of GSSC). Closed habitat GSSC forms were not observed in any of the samples. Bootstrapping tests indicate that the rel-ative abundance distribution of different GSSC morphotypes could be explained by PACMAD grasses alone; however, the occurrence of highly diagnostic pooid forms (e.g., crenates) in all three samples suggests that pooids were present in low abundances as well.
Sedge phytoliths were found in abundance in UWBM no. PB22778 and are rare in nos. PB22779 and PB22780. Other biosilica types potentially indicative of wetlands (diatoms, etc.) were relatively rare.
DISCUSSION
The macroflora and palynoflora provide de-tailed floristic information compared to phytoliths (except within Poaceae) and will be considered together for reconstructing the plant community composition at Vasa Park. They complement each other because of the localized nature of the macrofossil assemblage versus the more regional sampling provided by windborne pollen. Among the pteridophytes and allies, a rich macroflora consisting of a bryophyte, Equisetum, and five fern morphotypes is reinforced by a diverse spore record. In the case of gymnosperms, macrofossils are extremely rare, but pollen documents a rich flora of at least a dozen taxa, consistent with other regional Miocene palynofloras (see Table 2). The angiosperm macroflora is not very diverse com-pared to Northwest Middle Miocene localities (see Table 3), but this likely represents taphonomic bias. Flood overbank deposits, such as those found at Vasa Park, typically preserve mostly autoch-thonous plant macrofossils while many lacustrine deposits record allochthonous material, some-
times on a regional scale (Behrensmeyer & Hook, 1992). Most of the described Middle Miocene sites represent lacustrine deposition and are much more diverse than the Vasa Park flora. The Late Miocene record from the Tyrrell site in the El-lensburg flora (Table 3) is the only one directly comparable to Vasa Park, representing collections from a single site of fluvial deposition.
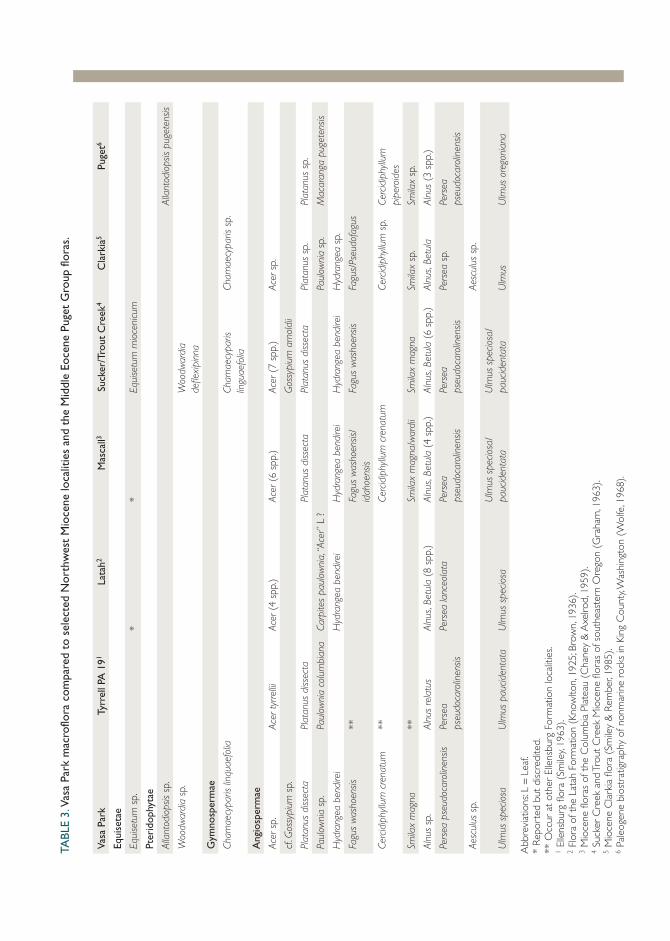
The identified macroflora at Vasa Park consists of elements common to all of the well-studied Middle Miocene sites from the interior of north-west North America (see Table 3). At the genus and often the species level, Acer, Alnus, Fagus, Hydrangea, Persea, Platanus, Smilax, and Ulmus are common to virtually all localities. Only a few of the genera recognized at Vasa Park are rare at other localities, namely Equisetum (likely because of taphonomic bias), Aesculus, and Allantodiopsis sp. Five distinctive fern morphotypes have been found at Vasa Park, which is a much more diverse fern component than other Northwest Miocene occurrences. We hypothesize that this difference is also likely due to taphonomic bias linked to facies type.
Comparison of Vasa Park to other Northwest Middle and Late Miocene sites shows a striking stasis in the floras. Ninety-five percent of the identified macrofossils from Vasa occur in at least one of the Middle Miocene sites, and all of the morphotypes that can be resolved to the species level were first described from Middle Miocene localities. The Middle Eocene Puget flora, which occurs within a few miles of Vasa Park, includes 50% of the macroflora found at Vasa Park at the genus level.
Vasa Park gymnosperms are primarily known from the palynoflora. The Pinaceae are represented by Abies (two types), Cedrus, Picea (two types), Pinus, Pseudotsuga/Larix, and Tsuga (two types). In addition, there are several bisaccate palyno-morphs, which we were unable to identify at the genus level. Pinus is the most common type, con-stituting approximately 5%–15% of the pollen in the counted samples. The Pseudotsuga/Larix type
05Graham.064_097.indd 90 7/25/14 12:32 PM
TABL
E 3.
Vas
a Pa
rk m
acro
flora
com
pare
d to
sel
ecte
d N
orth
wes
t M
ioce
ne lo
calit
ies
and
the
Mid
dle
Eoce
ne P
uget
Gro
up fl
oras
.
Vasa
Par
kTy
rrel
l PA
191
Lata
h2M
asca
ll3Su
cker
/Tro
ut C
reek
4C
lark
ia5
Puge
t6
Equi
seta
e
Equi
setu
m s
p.
**
Equi
setu
m m
ioce
nicu
m
Pter
idop
hyta
e
Alla
ntod
iops
is sp
.
Alla
ntod
iops
is pu
gete
nsis
Woo
dwar
dia
sp.
W
oodw
ardi
a de
flexi
pinn
a
Gym
nosp
erm
ae
Cham
aecy
paris
linq
uaef
olia
Ch
amae
cypa
ris
lingu
aefo
liaCh
amae
cypa
ris s
p.
Ang
iosp
erm
ae
Acer
sp.
Acer
tyrre
lliiAc
er (
4 sp
p.)
Acer
(6
spp.
)Ac
er (
7 sp
p.)
Acer
sp.
cf. G
ossy
pium
sp.
G
ossy
pium
arn
oldi
i
Plat
anus
diss
ecta
Plat
anus
diss
ecta
Pl
atan
us d
issec
taPl
atan
us d
issec
taPl
atan
us s
p.Pl
atan
us s
p.
Paul
owni
a sp
.Pa
ulow
nia
colu
mbi
ana
Carp
ites
paul
owni
a, “A
cer”
L ?
Paul
owni
a sp
.M
acar
anga
pug
eten
sis
Hyd
rang
ea b
endi
rei
H
ydra
ngea
ben
dire
iH
ydra
ngea
ben
dire
iH
ydra
ngea
ben
dire
iH
ydra
ngea
sp.
Fagu
s w
asho
ensis
**
Fagu
s w
asho
ensis
/id
ahoe
nsis
Fagu
s w
asho
ensis
Fagu
s/Ps
eudo
fagu
s
Cerc
idip
hyllu
m c
rena
tum
**
Cerc
idip
hyllu
m c
rena
tum
Ce
rcid
iphy
llum
sp.
Cerc
idip
hyllu
m
pipe
roid
es
Smila
x m
agna
**
Smila
x m
agna
/war
dii
Smila
x m
agna
Smila
x sp
.Sm
ilax
sp.
Alnu
s sp
.Al
nus
rela
tus
Alnu
s, Be
tula
(8
spp.
)Al
nus,
Betu
la (
4 sp
p.)
Alnu
s, Be
tula
(6
spp.
)Al
nus,
Betu
laAl
nus
(3 s
pp.)
Pers
ea p
seud
ocar
olin
ensis
Pers
ea
pseu
doca
rolin
ensis
Pers
ea la
nceo
lata
Pers
ea
pseu
doca
rolin
ensis
Pers
ea
pseu
doca
rolin
ensis
Pers
ea s
p.Pe
rsea
ps
eudo
caro
linen
sis
Aesc
ulus
sp.
Aesc
ulus
sp.
Ulm
us s
pecio
saU
lmus
pau
ciden
tata
Ulm
us s
pecio
saU
lmus
spe
ciosa
/pa
ucid
enta
taU
lmus
spe
ciosa
/pa
ucid
enta
taU
lmus
Ulm
us o
rego
nian
a
Abb
revi
atio
ns: L
= L
eaf.
* Re
port
ed b
ut d
iscre
dite
d.**
Occ
ur a
t ot
her
Elle
nsbu
rg F
orm
atio
n lo
calit
ies.
1 El
lens
burg
flor
a (S
mile
y, 19
63).
2 Fl
ora
of t
he L
atah
For
mat
ion
(Kno
wlto
n, 1
925;
Bro
wn,
193
6).
3 M
ioce
ne fl
oras
of t
he C
olum
bia
Plat
eau
(Cha
ney
& A
xelro
d, 1
959)
.4
Suck
er C
reek
and
Tro
ut C
reek
Mio
cene
flor
as o
f sou
thea
ster
n O
rego
n (G
raha
m, 1
963)
. 5
Mio
cene
Cla
rkia
flor
a (S
mile
y &
Rem
ber,
1985
).6
Pale
ogen
e bi
ostr
atig
raph
y of
non
mar
ine
rock
s in
Kin
g C
ount
y, W
ashi
ngto
n (W
olfe
, 196
8).
05Graham.064_097.indd 91 7/25/14 12:32 PM
92 PALEOBOTANY AND BIOGEOGRAPHY
is relatively rare, whereas the remainder contrib-ute a small but significant percentage of the pollen in each sample. Coniferous pollen is often interpreted as a slope or montane element (e.g., Chaney & Axelrod, 1959; Smiley & Rember, 1985), but it should be noted that Abies, Picea, Pinus, Pseudotsuga, Thuja L., and Tsuga are all pres-ent in the extant coastal lowland forests of the Pacific Northwest, as well as at higher elevations.
The abundance of Alnus pollen suggests a local source. Modern Alnus is a common riparian tree type and a pioneer species in disturbed habitats. The relative abundance of fern spores can also be associated with disturbance (Walker & Sharpe, 2010), and Salix is a common riparian shrub, while Nyssa is associated with swamps and wet-lands. The macrofossil records of abundant ferns and pioneer angiosperms such Alnus and Acer, coupled with evidence provided by the palyno-flora, are consistent with a disturbance or early successional flora that formed in a fluvial, flood-plain environment. The presence of fine siltstone sediments and aquatic taxa such as Typha/Sparganium and Potamogeton in the pollen record indi-cate that there were also some quiet water areas in the vicinity.
Table 2 shows a comparison of the Vasa Park palynoflora with several other Miocene sites from the Pacific Northwest. Even though these sites span a distance of hundreds of kilometers and a time period of several million years, the similari-ties are striking. Of the 64 total taxa listed in the table, 36% are common to all of the floras and 51% are found in at least three out of four floras, indicating that there was a widespread and stable regional forest assemblage during the Miocene. Vasa Park occurred during the global cooldown that followed the Mid-Miocene Climatic Opti-mum (Zachos et al., 2001), but it is clear that no significant floral turnover had yet occurred in the Pacific Northwest by this time.
Fifty percent of the genera found at Vasa Park are no longer extant in northwest North America. They are found in eastern North American and
Eurasian forests with a temperate climate and year-round rainfall. The coexistence approach analysis suggests a similar climate at Vasa Park during the Miocene, with the mean annual tem-perature in a range of 11.6°C–16.6°C, limited frost in the winter (cold month mean tempera-ture, –0.3°C to 2.9°C), moderate to high precipi-tation (mean annual precipitation, 641–1577 mm), and adequate rainfall during the growing season (warm month precipitation, 89–175 mm). This is consistent with climate interpretations for other Miocene floras from the Northwest, and the Vasa Park climate analysis, coupled with the high de-gree of taxonomic overlap between the Miocene palynofloras, supports the hypothesis that the regional climate did not change significantly during the 4- to 5-million-year time frame repre-sented by these floras. Graham (1999) uses evi-dence from a number of fossil sites in the western United States to show that a cooling climate and dry summer conditions started to set in after the Mid-Miocene Climatic Optimum, beginning in southwestern North America and the continen-tal interior. The dry summer conditions would have resulted in the loss of many deciduous hardwoods that require adequate summer rain-fall. According to this analysis and consistent with our data, the Pacific Northwest did not experi-ence these effects until later, and Graham (1999) states that the warm temperature and wet sum-mer climate conditions persisted in the region into the Early Pliocene.
Because the phytolith assemblages were gen-erally well preserved in Vasa Park deposits, we assume that they reflect deposition from plants growing on the floodplain, rather than phytoliths transported by water or wind from surrounding highlands (Osterrieth et al., 2009). The overall low abundances of sedge phytoliths and other bi-osilica (e.g., diatoms, sponge spicules) commonly associated with lakes and rivers further indicate that the assemblages represent deposition in a soil, rather than in water, such as in an oxbow lake. All extracted samples from Vasa Park had very low
05Graham.064_097.indd 92 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 93
concentrations of phytoliths compared to typi-cal Cenozoic sediments (Strömberg, unpublished data). This suggests that the horizons collected, even those that had clear root traces, did not re-main stable soil surfaces for very long. Of the three samples that were quantitatively analyzed for phytoliths, UWBM nos. PB22779 and PB22780 both point to closed habitats (forest) with a low abundance of PACMAD grasses and rare pooids. Since these samples were collected from the same horizon, their similarity indicates little spatial het-erogeneity in vegetation, at least over the small distance sampled at Vasa Park. The higher abun-dance of sedges in UWBM no. PB22779 suggests that this sample reflects a microhabitat closer to water. Assemblage UWBM no. PB22778 appears to represent a somewhat more open microhabitat, with a mix of trees and forbs, mainly PACMAD grasses and infrequent pooids. The presence of palm phytoliths in two assemblages opens up the intriguing possibility that palms persisted in the Pacific Northwest until the early Late Miocene; the youngest previously described palms from the region are pollen from the Middle Miocene Succor Creek flora of the Oregon-Idaho border (Taggart & Cross, 1990), and phytoliths from the late Middle and early Late Miocene of south-western Montana (Strömberg, 2005; Smith et al., 2012; Strömberg et al., 2012). In addition, palm phytoliths have been recovered from the Middle Miocene Mascall Formation of Oregon (R. Dunn & C. Strömberg, unpublished data). However, because of the very low abundance of palm phy-toliths at Vasa Park, the fact that other monocot-yledons produce phytoliths that can look similar to palm phytoliths (e.g., orchids; Piperno, 1988), and the lack of co-occurring pollen attributable to Arecaceae, the identification of palms at Vasa Park has to remain tentative.
In all, phytolith assemblages point to a mostly forested floodplain at Vasa Park. The abundance of PACMADs in grass communities could be consistent with warmer climates, and could also point to mesophytic PACMAD grasses growing
close to water (e.g., Arundo L.; see Strömberg et al., 2007a). The specific PACMAD morphotypes that dominate GSSC assemblages (e.g., so-called almost panicoid bilobates) are similar to those produced by Arundo. However, as noted above, other lines of evidence (diatoms, etc.; Fig. 9) are not congruent with an interpretation of these as-semblages as mirroring wetland vegetation. Palms, if present, would also support the pollen-based inference that climate was warmer than today in Washington during the early Late Miocene. This reconstructed vegetation differs from roughly coeval phytolith assemblages from sites further inland, in western Montana. Here, assemblages appear to reflect open habitats dominated by cool-temperate pooids (Strömberg et al., 2012). Similarly, faunal and paleosols data have been in-terpreted as pointing to savanna or woodland habitats for the early Late Miocene of eastern Oregon (Shotwell, 1970; Retallack, 2004). These differences may be a result of a difference in re-gional climate (e.g., rainfall) between western Washington and areas further east, but it cannot be ruled out that it is a consequence of sampling different, local edaphic conditions.
CONCLUSIONS
Applying a range of geological and paleobotan-ical approaches, we are able to more completely describe the flora recorded at Vasa Park than by using single lines of evidence. Geological study not only provides a “big picture” view of the sed-imentary system that created the deposit but also informs us about taphonomic filtering and the age of fossil-bearing sediments. The macroflora mostly records localized plant communities adapted to regular disturbance close to fluvial channels and is not diverse enough to estimate climate. The palynoflora adds a regional perspective of likely conifer-dominated upland floras and the much more species-rich surrounding hardwood forests. It also provides us with a regional climate esti-mate. The phytolith assemblage informs us about
05Graham.064_097.indd 93 7/25/14 12:32 PM
94 PALEOBOTANY AND BIOGEOGRAPHY
forest coverage and grass ecology in the flora re-corded at Vasa Park. In all, these lines of evidence indicate that plant communities in western Wash-ington differed little from Middle to Late Mio-cene floras from around the Pacific Northwest, and that the regional climate had remained mild and humid since the Middle Miocene. Unlike coeval sites from the northern Rocky Mountains and Great Plains, open habitats dominated by cool- and dry-adapted pooid grasses appear not to have been important in the Pacific Northwest during this time. These patterns suggest that the Pacific Northwest may have served as a climatic refugium amidst dropping global temperatures and increasing seasonality during the Late Mio-cene (Zachos et al., 2001).
ACKNOWLEDGMENTSWe thank the Vasa Park Homeowners Associa-
tion for access to their property and permission to collect the specimens used in this study. We particularly thank Susan Winkowski for her help securing locality access. We thank Jonathan Hager, who first discovered fossils here and found many of the important specimens used in this study. Jonathan would like to acknowledge the assistance and encouragement of Wesley C. Wehr, former affiliate curator of paleobotany at the Burke Mu-seum of Natural History and Culture, University of Washington. Brian Sherrod of the U.S.G.S. provided the geological setting and figures but was unable to participate as an author. Thank you for your contribution, Brian. We also thank Regan Dunn, Don Hopkins, and the rest of the Strömberg lab for helping with field work, fossil collecting, and measuring of the section. R. M. D. thanks Steven Manchester, Bill Rember, Kathleen Pigg, David Greenwood, and Jonathan Wendel for their assistance in macrofossil identification. T. A. D. thanks Estella B. Leopold and Stephanie Zaborac for their help with palynological studies and Torsten Utescher for his assistance with the coexistence approach climate analysis. A. P. J.
thanks Brian Buchwitz, Marijana Šurković Davis, and the Strömberg lab for mentoring and help. Funding was provided by the Evolving Earth Foundation to R. M. D. and T. A. D.; the Mary Gates Foundation, a UW-HHMI Integrative Re-search Internship Program Scholarship, a Geo-logical Society of America Travel Grant to A. P. J., and a National Science Foundation grant (EAR-0819910) to C. A. E. S.; and the Burke Museum of Natural History and Culture, University of Washington.
LITERATURE CITEDAliscioni, S., H. L. Bell, G. Besnard, P. A. Christin,
J. T. Columbus, M. R. Duvall, E. J. Edwards, L. Gi-ussani, K. Hasenstab-Lehman, K. W. Hilu, T. R. Hodkinson, A. L. Ingram, E. A. Kellogg, S. Ma-shayekhi, O. Morrone, C. P. Osborne, N. Salamin, H. Schaefer, E. Spriggs, S. A. Smith & F. Zuloaga. 2012. New grass phylogeny resolves deep evolu-tionary relationships and discovers C4 origins. New Phytol. 193: 304–312.
Barnett, J. & L. H. Fisk. 1980. Palynology and paleo-ecology of a sedimentary interbed in the Yakima Basalt (Miocene), Palouse Falls, Washington. Northw. Sci. 54: 259–278.
Behnke, H. D. 1997. Sarcobataceae: A new family of Caryophyllales. Taxon 46: 495–507.
Behrensmeyer, A. K. & R. W. Hook. 1992. Paleoen-vironmental contexts and taphonomic modes. Pp. 15–93 in Behrensmeyer et al. (editors), Terrestrial Ecosystems through Time: Evolutionary Paleoecol-ogy of Terrestrial Plants and Animals. University of Chicago Press, Chicago.
Blakely, R. J., R. E. Wells, C. S. Weaver & S. Y. Johnson. 2002. Location, structure, and seismicity of the Seattle fault zone, Washington—Evidence from aeromagnetic anomalies, geologic mapping, and seismic-reflection data. Geol. Soc. Amer. Bull. 114: 169–177.
Booth, D., K. Troost & J. Hagstrum. 2004. Deforma-tion of Quaternary strata and its relationship to crustal folds and faults, south-central Puget Low-land, Washington State. Geology 32: 505–508.
Booth, D. B., T. J. Walsh, K. G. Troost & S. A. Shimel. 2012. Geologic Map of the East Half of the Bel-levue South 7.5′ × 15′ Quadrangle, Issaquah Area, King County, Washington: U.S. Geological Survey Scientific Investigations Map 3211, scale 1:24,000.
05Graham.064_097.indd 94 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 95
Brocher, T., T. Parsons, R. Blakely, M. Fisher, R. Wells & N. Christensen. 2001. Upper crustal structure in Puget Lowland, Washington: Results from the 1998 seismic hazards investigation in Puget Sound. J. Geophys. Res. 106: 541–564.
Brown, R. W. 1936. Additions to some fossil floras of the western United States. U.S. Geol. Survey Prof. Paper 186-J: 163–206.
Brown, R. W. 1962. Paleocene Flora of the Rocky Mountains and Great Plains. U.S. Geol. Survey Prof. Paper 375. U.S. Govt. Print. Off., Washington, D.C.
Chaney, R. W. & D. I. Axelrod. 1959. Miocene Floras of the Columbia Plateau. Carnegie Institution of Washington 617. Carnegie Institution, Washington, D.C.
Collinson, M. E. 2001. Cainozoic ferns and their dis-tribution. Brittonia 53: 173–235.
Duvall, M. R., J. I. Davis, L. G. Clark, J. D. Noll, D. H. Goldman & J. G. Sánchez-Ken. 2007. Phy-logeny of the grasses (Poaceae) revisited. Aliso 23: 237–247.
Ellis, B., D. C. Daly, L. J. Hickey, K. R. Johnson, J. D. Mitchell, P. Wilf & S. L. Wing. 2009. Manual of Leaf Architecture. The New York Botanical Garden Press, New York.
Erdtman, G. 1952. Pollen Morphology and Plant Taxonomy, Angiosperms. Almqvist & Wiksell, Stockholm.
Erdtman, G. 1957. Pollen and Spore Morphology/Plant Taxonomy. Gymnospermae, Pteridophyta, Bryophyta (Illustrations). Almqvist & Wiksell, Stockholm.
Faegri, K. & J. Iversen. 1964. Textbook of Pollen Anal-ysis. Hafner Publishing Co., New York.
Fields, P. F. 1996. The Succor Creek Flora of the Mid-dle Miocene Sucker Creek Formation, Southwestern Idaho and Eastern Oregon: Systematics and Paleo-ecology. [Unpubl.] Ph.D. Dissertation, Michigan State University, East Lansing.
Franklin, J. F. & C. T. Dyrness. 1988. Natural Vegeta-tion of Oregon and Washington, rev. ed. Oregon State University Press, Corvallis.
Graham, A. 1963. Systematic revision of the Sucker Creek and Trout Creek Miocene floras of southeast-ern Oregon. Amer. J. Bot. 50: 921–936.
Graham, A. 1965. The Sucker Creek and Trout Creek Miocene Floras of Southeastern Oregon. Kent State University Research Ser. IX. Kent State University Press, Kent, Ohio.
Graham, A. 1999. Late Cretaceous and Cenozoic His-tory of North American Vegetation. Oxford Univer-sity Press, Oxford.
Gray, J. 1958. Plant Microfossils from the Miocene of the Columbia Plateau, Oregon. [Unpubl.] Ph.D. Dissertation, University of California, Berkeley.
Gray, J. 1985. Interpretation of co-occurring megafos-sils and pollen: A comparative study with Clarkia as an example. Pp. 185–244 in C. J. Smiley, A. E. Lev-iton & M. Berson (editors), Late Cenozoic History of the Pacific Northwest. American Association for the Advancement of Science, San Francisco.
Heinemann, A. 1998–1999. ClimStat version 1.02. University of Tübingen, Tübingen, Germany.
Johnson, S. Y., C. J. Potter & J. M. Armentrout. 1994. Origin and evolution of the Seattle fault and Seattle basin, Washington. Geology 22: 71–74.
Johnson, S. Y., S. V. Dadisman, J. R. Childs & W. D. Stanley. 1999. Active tectonics of the Seattle Fault and central Puget Sound, Washington: Implications for earthquake hazards. G.S.A. Bull. 111: 1042– 1053.
Knowlton, F. H. 1902. Fossil Flora of the John Day Basin, Oregon. U.S. Geol. Survey Bull. 204. U.S. Govt. Print. Off., Washington, D.C.
Knowlton, F. H. 1926. Flora of the Latah Formation of Spokane, Washington, and Coeur D’Alene, Idaho. U.S. Geol. Survey Prof. Paper 140-A: 17–55.
Kvaček, Z. & S. R. Manchester. 1999. Eostangeria barthel (extinct Cycadales) from the Paleogene of West-ern North America and Europe. Int. J. Pl. Sci. 160: 621–629.
Kvaček, Z. & W. C. Rember. 2000. Shared Miocene conifers of the Clarkia flora and Europe. Acta Univ. Carol., Geol. 44: 75–85.
Leopold, E. B. & M. F. Denton. 1987. Comparative age of grassland and steppe east and west of the northern Rocky Mountains. Ann. Missouri Bot. Gard. 74: 841–867.
Liberty, L. M., & T. L. Pratt. 2008. Structure of the eastern Seattle fault zone, Washington State: New insights from seismic reflection data. Bull. Seismo-logical Soc. Amer. 98: 1681–1695.
Liesch, B. A., C. E. Price & K. L. Walters. 1963. Ge-ology and ground water resources of northwestern King County, Washington. Wash. State Water Sup-ply Bull. 20: 1–24.
MacGinitie, H. D. 1933. The Trout Creek flora of southeastern Oregon. Pp. 21–68 in Carnegie Insti-tution of Washington 416, Contributions to Pale-ontology. Carnegie Institution, Washington, D.C.
MacGinitie, H. D. 1953. Fossil Plants of the Florissant Beds, Colorado. Carnegie Institution of Washing-ton 599. Carnegie Institution, Washington, D.C.
05Graham.064_097.indd 95 7/25/14 12:32 PM
96 PALEOBOTANY AND BIOGEOGRAPHY
McIver, E. E. & J. F. Basinger. 1993. Flora of the Ra-venscrag Formation (Paleocene), southwestern Sas-katchewan, Canada. Paleontographica Canadiana 10: 1–167.
Merriam, J. C. 1901. A contribution to the geology of the John Day basin. Univ. Calif. Bull. Dept. Geol. II(9): 269–314.
Mosbrugger, V. & T. Utescher. 1997. The coexistence approach: A method for quantitative reconstructions of Tertiary terrestrial palaeoclimate data using plant fossils. Palaeogeogr. Palaeoclimatol. Palaeoecol. 134: 61–86.
Mullineaux, D. R. 1970. Geology of the Renton, Auburn and Black Diamond Quadrangles, King County, Washington. Geol. Soc. Prof. Paper 674: 24–26.
Osterrieth, M., M. Madella, D. Zurro & M. F. Alva-rez. 2009. Taphonomical aspects of silica phytoliths in the loess sediments of the Argentinean Pampas. Quatern. Int. 193: 70–79.
Pabst, M. B. 1968. The Flora of the Chuckanut For-mation of Northwestern Washington, The Equise-tales, Filicales and Coniferales. Univ. Calif. Publ. Geol. Sci. 76: 1–60.
Pigg, K. B. & G. W. Rothwell. 2001. Anatomically preserved Woodwardia virginica (Blechnaceae) and a new filicalean fern from the Middle Miocene Ya-kima Canyon flora of central Washington, USA. Amer. J. Bot. 74: 458–461.
Pigg, K. B., M. L. DeVore & W. C. Wehr. 2006. Fili-calean Ferns from the Tertiary of Western North America: Osmunda L. (Osmundaceae : Pteridophyta), Woodwardia SM. (Blechnaceae : Pteridophyta) and Onocleoid forms (Filicales : Pteridophyta). Fern Gaz. 17: 279–286.
Piperno, D. R. 1988. Phytolith Analysis, an Archaeo-logical and Geological Perspective. Academic Press, San Diego.
Piperno, D. R. 2006. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists, 1st ed. AltaMira Press, New York.
Rau, W. W. 1967. Geology of the Wynoochee Valley [15-minute] quadrangle, Grays Harbor County, Washington, Bull. 56, Wash. Division of Geology and Earth Res., 1–51, scale 1:62,500.
Reininck-Smith, L. M. & E. B. Leopold. 2005. Warm climate in the Late Miocene of the south coast of Alaska and the occurrence of Podocarpaceae pollen. Palynology 29: 205–262.
Retallack, G. J. 2004. Late Miocene climate and life on land in Oregon within a context of Neogene
global change. Palaeogeogr. Palaeoclimatol. Palaeo-ecol. 214: 97–123.
Shotwell, J. A. 1970. Pliocene mammals of southeast Oregon and adjacent Idaho. Bull. Museum Nat. Hist., Univ. Oregon 17: 1–103.
Simon, J. L. 1997. Resampling: The New Statistics, 2nd ed. Resampling Stats, Inc., Arlington, Virginia.
Smiley, C. J. 1961. A record of Paulownia in the Ter-tiary of North America. Amer. J. Bot. 48: 175–179.
Smiley, C. J. 1963. The Ellensburg Flora of Washing-ton. Univ. Calif. Publ. Geol. Sci. 35: 159–276.
Smiley, C. J. & W. C. Rember. 1985. The composition of the Miocene Clarkia flora. Pp. 95–112 in C. J. Smiley, A. E. Leviton & M. Berson (editors), Late Cenozoic History of the Pacific Northwest. Pacific Division of the American Association for the Ad-vancement of Science, San Francisco.
Smith, K. J., C. A. E. Strömberg, N. D. Sheldon & S. Y. Smith. 2012. Vegetation reconstruction of the Middle Miocene in southwest Montana using phy-tolith and stable isotope analyses. G.S.A. Abstracts with Programs 44(7): 291.
Soil Survey Staff. 2006. Keys to Soil Taxonomy, 10th ed., U.S. Department of Agriculture, Natural Re-sources Conservation Services, Washington, D.C.
Strömberg, C. A. E. 2003. The origin and spread of grass-dominated ecosystems during the Tertiary of North America and how it relates to the evolution of hypsodonty in equids. [Unpubl.] Ph.D. Thesis, University of California, Berkeley.
Strömberg, C. A. E. 2004. Using phytolith assem-blages to reconstruct the origin and spread of grass- dominated habitats in the great plains of North America during the Late Eocene to Early Miocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 207: 239– 275.
Strömberg, C. A. E. 2005. Decoupled taxonomic ra-diation and ecological expansion of open-habitat grasses in the Cenozoic of North America. Proc. Natl. Acad. Sci. U.S.A. 102: 11980–11984.
Strömberg, C. A. E. 2009. Methodological concerns for analysis of phytolith assemblages: Does count size matter? Quatern. Int. 193: 124–140.
Strömberg, C. A. E., E. M. Friis, M. Liang, L. Werdelin & Y. Zhang. 2007a. Paleoecology of an Early-Middle Miocene lake in China: Preliminary interpretations based on phytoliths from the Shanwang Basin. Ver-tebrata PalAsiatica 45: 145–169.
Strömberg, C. A. E., L. Werdelin, E. M. Friis & G. Saraç. 2007b. The spread of grass-dominated habitats in Turkey and surrounding areas during the
05Graham.064_097.indd 96 7/25/14 12:32 PM
THE VASA PARK FLORA, KING COUNTY, WASHINGTON, U.S.A. 97
Strömberg, C. A. E., N. D. Sheldon, S. Y. Smith, J. M. Cotton & J. M. M. Hamer. 2012. Multi-proxy (phytoliths, stable isotopes, trace fossils) high-reso-lution record of Late Miocene ecosystem variability. G.S.A. Abstracts with Programs 44(7): 265.
Strömberg, C. A. E., R. E. Dunn, R. H. Madden, M. J. Kohn & A. A. Carlini. 2013. Decoupling the spread of grasslands from the evolution of grazer- type herbivores in South America. Nat. Communica-tions 4: article no. 1478. doi:10.1038/ncomms2508.
Taggart, R. E. & A. T. Cross. 1990. Plant successions and interruptions in Miocene volcanic deposits, Pa-cific Northwest. G.S.A. Special Papers 244: 57–68.
Walker, L. R. & J. M. Sharpe. 2010. Ferns, distur-bance and succession. Pp. 177–219 in J. Mehltreter, L. R. Walker & J. M. Sharpe (editors), Fern Ecol-ogy. Cambridge University Press, Cambridge.
Walters, K. L. 1964. Mashell Formation of Southwest-ern Pierce County, Washington. Geol. Survey Bull. 1224-A: A55–A59.
Wells, R., C. Weaver & R. Blakely. 1998. Fore-arc mi-gration in Cascadia and its neotectonic significance. Geology 26: 759–762.
Wendel, J. F., C. L. Brubaker & T. Seelanan. 2010. The origin and evolution of Gossypium. Pp. 1–18 in J. M. Stewart, D. Oosterhuis, J. J. Heitholt & J. R. Mauney (editors), Physiology of Cotton. Springer Verlag, Dordrecht.
Wheeler, E. A. & T. A. Dillhoff. 2009. The Middle Miocene Wood Flora of Vantage, Washington, USA. IAWA Journal, Supplement 7. International Associ-ation of Wood Anatomists, Leiden, Netherlands.
Wing, S. L. 1987. Eocene and Oligocene floras and vegetation of the Rocky Mountains. Ann. Missouri Bot. Gard. 74: 748–784.
Wodehouse, R. P. 1935. Pollen Grains, Their Struc-ture, Identification and Significance in Science and Medicine, 1st ed. McGraw-Hill Book Company, Inc., New York.
Wolfe, J. A. 1968. Paleogene Biostratigraphy of Non-marine Rocks in King County, Washington. Geol. Survey Prof. Paper 571: 1–29.
Wolfe, J. A. & T. Tanai. 1987. Systematics, phylogeny and distribution of Acer (maples) in the Cenozoic of Western North America. J. Fac. Sci. Hokkaido Univ., Ser. 4, Geol. Mineral. 22: 1–246.
Yount, J. C. & H. D. Gower. 1991. Bedrock geologic map of the Seattle 30 × 60 quadrangle, Washington. U.S. Geological Survey, scale 1:100,000.
Zachos, J., M. Pagani, L. Sloan, E. Thomas & K. Bil-lups. 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292 (5517): 686–693.
Zetter, R., M. J. Farabee, K. B. Pigg, S. R. Manchester, M. L. DeVore & M. D. Nowak. 2011. Palynoflora of the Late Paleocene silicified shale at Almont, North Dakota, USA. Palynology 2011: 1–33. doi: 10.1080/01916122.2010.50164.