Solving Bernstein's Problem: A Proposal forthe Development of Coordinated Movementby Selection
Olaf Sporns and Gerald M. EdelmanThe Neurosciences Institute
SPOHNS, OLAF, AND EDELMAN, GERALD M, Solving Bernstein's Problem: A Proposal for the Devel-opment of Coordinated Movement by Selection. CHILD DEVELOPMENT, 1993, 64, 960-981, Inrecent years, many established concepts in the theory of human motor development have under-gone profound change, and our knowledge has increased greatly. Nevertheless, some outstandingproblems remain unsolved, A centra! problem concerns the redundancy of effective movements,flrst pointed out by N, A, Bernstein, The human motor system is mechanically complex and canmake use of a large number of degrees of freedom. The controlled operation of such a systemrequires a reduction of mechanical redundancy, effectively by reducing the number of degrees offreedom. More recent work has shown that this problem is hard to solve explicitly by computingsolutions to the equations of motion ofthe system. Equally challenging to traditional computa-tional approaches is the fact the motor systems show remarkable adaptability and flexibility inthe presence of changing biomechanical properties of motor organs during development andwhen faced with different environmental conditions or tasks. Solutions to these problems wouldhave a large impact on a variety of issues in child development. In this article, we stress theimportance ofthe somatic selection of neuronal groups in maps for the progressive transformationoi a primary movement repertoire into a set of motor synergies and adaptive action patterns. Wepresent results from computer simulations of a simple motor system that works according to suchselectional principles. This approach suggests a provisional solution to Bernstein's problem andprovides new parameters to guide experimental approaches to the development of sensorimotorcoordination.
A major function of animal and human cial development (Cratty, 1979; Thelen,nervous systems is the coordinated control 1989a),of limb and body movements. Coordinated . , r i j , r i imotor activity enables an animal to explore A satisfactory understanding of early hu-its environment and to sample and attend '"^", development can only be achieved. i- 1- J i • i- 1 f J, within the context of a global theory ot brainto sensory stimuli, and it is essential for its „ , ^ , , ? , j - r
1 lU- j, • u T i,- II u function. Several theories based on lntorma-survival within its econiche. Initially, how- , , i i j T l,
.1 . ui-i,- X- u tion processing have been proposed. In suchever, the motor capabilities of newborn , . * ^ , . i i • j, , , _ , ,u , i , i i - j ^ theories, motor function IS largely viewed ashigher vertebrates are generally inadequate , , ' , >,• 1 i u. 1, , r ,,u i, 1 ? 1 dependent upon cybernetic control mecha-to accomplish any of these vital tasks, , , „ ^ j , i i r i
1.1 u-i .,• J r J nisms and feedback loops of a more or lessand these capabilities undergo profound , ^ , . . T .,u- _•,• i j1 1 J J • J c i intricate nature. In this article, we consider
changes over an extended period of postna- , r , I , , J \ , U I I. , J 1 . T,, , ,4 • r _!• 1 a number of issues related to biologicaltal development. This period is of particular ^ u ,, l • u i, . i , 1 • r i r u movement that pose several serious chal-lnterest in human infants, many of whose , ^ u r l i- r A * 4-U
, , , , . , ^ u • i,u •- u lenges to the formulation of an adequate the-motor abilities at birth appear to be among . . r .. l • r .,•,1 .1 , ji_ j i i U ory in terms oi conventional information pro-
the least developed when compared to those - c r^i. u iir ., _i u i • * u i j ' i- cessing. Some of these challenges are;
of other vertebrate species, A child s contin- °ued progress in achieving precise control of a) Evolutionary changes in the structuremotor functions is an obvious prerequisite and function of the musculoskeletal appara-for its further behavioral, cognitive, and so- tus must be compensated for by accompa-
This work was carried out as part of the Institute Fellows in Theoretical Neurobiologyprogram at The Neurosciences Institute, which is supported by the Neurosciences ResearchFoundation, O,S, is a Charles and Mildred Schnurmacher Fellow, Address correspondence to;Gerald M, Edelman, Tbe Neurosciences Institute, 1230 York Avenue, New York, NY 10021,
nying changes in the nervous system. Dur-ing evolution, genetic changes may affectthe biomechanics of limbs or the morphol-ogy of muscles and tendons in many dififer-ent ways. In order to maintain adequate con-trol by corresponding circuits in the nervoussystem, these circuits must be adjusted in acoordinated manner (for examples, see Edel-man, 1987), Coordinated mutations affectingboth body mechanics and brain structureare, however, unlikely to occur simulta-neously in the same individual (although thetwo domains obviously co-evolve on a largertime scale). This evolutionary problem de-mands a theory of motor function that per-mits for the possibility of rapid alterationsof neural circuits (involving mechanisms ofneural plasticity) in response to peripheralbiomechanical change.
b) All but the most stereotypic move-ments exhibit high degrees of variability,can be executed in a large number of possi-ble ways, are redundantly specified by neu-ral signals, and seem to involve widespreadand mutually overlapping parts of the ner-vous system. While motor learning generallyincreases the precision of movements, evenfully matured motor systems can executemovements in a large number of unpracticedways. The key point is that the number ofexecutable solutions to a given motor taskalways far exceeds the small number oflearned examples. As we have mentioned,the problem posed by motor redundancywas first clearly recognized by N. A. Bern-stein (Bernstein, 1967; Turvey, 1990; Whit-ing, 1984),
c) Motor coordination (particularly inhumans) is not innately specified but devel-ops gradually during postnatal life. Forexample, reaching movements in humaninfants are initially highly imprecise andvariable and lack the characteristic patternsof organization found in the adult (see, e.g..Fetters & Todd, 1987; Thelen & Fisher,1983; Trevarthen, 1974; von Hofsten, 1982).In the course of its development, an individ-ual is able to accommodate great individ-ual variations in neural and biomechanicalstructures. Moreover, many motor systemsremain plastic throughout adult life, ready tocompensate for metric and dynamic changesin the biomechanics of motor organs, includ-ing even the loss of entire limbs.
Although some promising advanceshave been made in the framework of eco-logical psychology and nonlinear systemstheory, these challenges have not been metadequately by cybernetic, kinematic, or in-
Sporns and Edelman 961
formation-processing models. We will con-sider an alternative view that appears todeal more effectively with all these chal-lenges. After briefly discussing the historicalorigins of some key problems of motor de-velopment (especially the problem of redun-dancy in the specification of motor com-mands), we will consider Bernstein's notionof motor synergies as functional units of themotor system. We will then explicitly dis-cuss some limitations and inconsistencies inviewing motor function as governed solelyby feedback control loops, algorithms, orcomputations. In contrast to these ap-proaches, we propose that successful devel-opmental coordination between neuronalactivity and the biomechanics of the muscu-loskeletal system is based on variation andis the result of somatic selective processeswithin brain circuits. Selection acts to matchpossible motor commands to constraintsposed by neural structure and kinematics.We will contrast our proposals with previousideas based on information-processing mod-els and briefly discuss some computer sim-ulations that exemplify the proposed ap-proach and demonstrate its self-consistency,
Bernstein's Problem
Bernstein, in 1935, pointed out that "therelationship between movements and the in-nervational impulses which evoke them isextremely complex and is, moreover, by nomeans univocal" (quoted in Whiting, 1984,p. 77; see also Turvey, 1990), Bernstein rec-ognized that an "unequivocal relationshipbetween impulses and movements does notand cannot exist" (Whiting, 1984, p. 82),partly because any movement is defined bygreatly varying interactions of centrally syn-thesized motor signals and the external forcefield acting on the moving part of the bodyat the periphery. He noted that the designof the vertebrate motor apparatus creates aproblem in the control of body movements,largely because ofthe many sources of inde-terminacy between central command andperipheral movements. The motor organs ofanimals or humans, composed of multiplelinkages and joints and an even greater num-ber of muscles, can attain a large numberof degrees of freedom. This number will ingeneral be far greater than the dimension oftheir workspace, defined as that region ofspace within which the motor organ canmove. This leads to what Bernstein termedredundancy; more than one motor signal canlead to the same trajectory of a given motorsystem; moreover, identical motor signalscan lead to different movements under non-
962 Child Development
identical initial conditions or in the pres-ence of variations in the external force field.Indeed, Bernstein defined motor coordina-tion itself as "the process of mastering re-dundant degrees of freedom of the movingorgan, in other words its conversion to a con-trollable system" (Bernstein, 1967, p. 127),Determining how this conversion processtakes place is what we will refer to as Bern-stein's problem.
Bernstein proposed that the motor appa-ratus is functionally organized into synergiesor classes of movement patterns. He empha-sized the "gestalt" character of individualmovements, finding it difficult to decomposethem into constitutive atomic parts, such asthe activities of individual muscles or rota-tions around a single joint; "A movementnever responds to detailed changes by a
change in its detail; it responds as a wholeto changes in each small part" (in Whiting,1984, p. 84). Quite naturally, repetitions ofmovements produce a class of similar but notidentical trajectories. An example of how thegestalt character of a movement is preservedunder complete reconstruction of its accom-panying muscle activities is given in FigureIA, Many different circles can be describedwith the hand as the arm extends in variousdirections from the body; these movements(forming a functional synergy) can be per-formed without further specific training. Inrelated experiments, Lashley and others no-ticed that "when habitually used motor or-gans are rendered nonfunctional by removalor paralysis, there is an immediate spontane-ous use of other motor systems which hadnot previously been associated with or usedin the performance ofthe activity" (Lashley,
B
nghthand
.ightmirro.(reversed)
, .„ minor(reversed)
7 8 9 10 11 12 13 14
FIG, 1,—A, An equivalence class of movements according to N, A, Bernstein, Movements withtopologically identical trajectories are executed by different sets of motor commands and under partici-pation of different muscle groups. From Bernstein (1967), reproduced with permission, B, Motor equiv-alence according to K, Lashley, This Figure shows "the writing of two individuals, blindfolded, usingright and leit hands in various positions, as well as other muscle groups, unpracticed. In spite ofclumsiness, the general features ofthe writing, individual differences in the forming of letters, and thelike, are characteristically maintained. The mechanics of writing is a sequence of movements in relationto body position, not a set pattern of special groups of muscles " (Lashley, 1942, quoted after Pribram,1969, p, 248), From Pribram, 1969, reproduced with permission, C, "Topology" according to Bernstein(1967), This term is adopted for all qualitative aspects of spatial conRguration or forms of movements,in contrast to quantitative metric aspects. Examples include 1—5, topological class of iive-pointed stars;6, topological class of figure eights with four angles; 7—14, topological class of letters A, From Bernstein(1967), reproduced with permission.
Sporns and Eldelman 963
1933, quoted in Beach, Hebb, Morgan, &Nissen, 1960, p, 239), In other words, "motoracts , , , may be executed immediately withmotor organs which were not associatedwith the act during training" (p, 240; see alsoFig, IB), These reflections gave rise to theconcept of motor equivalence, referring tothe variety of specific muscle contractionsand joint revolutions that produce the sameend result (Hebb, 1949),
Synergies and the Structure ofMovements
Synergies are classes of movement pat-terns involving collections of muscle or jointvariables that act as basic units in the regula-tion and control of movement, Bernstein wasthe first to propose that synergies are usedby the developing nervous system to reducethe number both of controlled parametersand of afferent signals needed to generateand guide an ongoing movement. Accordingto Bernstein, certain synergies are often as-sociated with a particular muscle group andcan therefore be at least partially defined bymorphology and anatomy. Other synergiescan be more clearly related to a given taskand provide a basis for "motor equivalence. "Bernstein also proposed that synergies cap-ture the "topological" rather than metric fea-tures of movements (see Fig. IC), The tem-poral structure of motor programs emergesthrough their parallel or sequential activa-tion. These ideas have been elaborated byothers. In closely related formulations byGelfand (Arshavsky, Celfand, & Orlovsky,1986; Gelfand, Gurfinkel, Tsetlin, & Shik,1971, 1973), syngergies are identified withcentral motor programs that can be triggeredby simple commands, Saltzman (1979) dis-tinguishes between functional synergies andmuscle synergies operating at different lev-els in a hierarchical control structure (Tur-vey, 1977), Functional synergies give rise to,or consist of, a set of motions producedacross a given set of joints. Muscle synergiesare defined by groups of muscles that act asunits and can span many joints. The activa-tion of a functional synergy produces a pat-tern of joint velocities characterized, forexample, by relatively fixed ratios acrossmultiple joints. Most synergies emerge aftersensorimotor training during postnatal de-velopment and underlie such well-practicedactions as walking or reaching for objects.Even after such fundamental synergies haveemerged, however, additional problemsmust be confronted, such as the selection ofthe appropriate functional synergies in agiven situation and the ordering and timing
of muscle contractions and joint anglechanges in ongoing motor activity.
The study of synergies in humans andanimals has led to the identification oi manykinds of invariant movement features. Al-though, in principle, many different strate-gies to execute a movement can be pursued,certain kinematic variables exhibit mutualdependencies and patterned changes. Kine-matic variables have been studied in differ-ent systems and including fixed joint veloc-ity ratios, time profiles of spatial movementparameters, and the relative timing of articu-lator movements during speech production.Examples of fixed joint velocity ratios havebeen described for hand movements (Kots& Syrovegin, 1966) and pointing movements(Soechting & Laquaniti, 1981) in humans,Laquaniti, Soechting, and Terzuolo (1986)showed that, although individual variationsoccur, shoulder and elbow motions are gen-erally tightly coupled. The hand trajectoriesof humans drawing geometrical figures likeellipses and circles in free space (Soechting,Laquaniti, & Terzuelo, 1986) have the sameoverall shape independent ofthe starting po-sition or orientation of the figure (compareFig, IB). In summary, many of the definingcharacteristics of synergies (e.g., the invari-ance of kinematic or muscle variables andthe fact that they cannot be unequivocallydissociated into independent elementarycomponents) have been demonstrated in awide variety of motor systems (Buchanan,Almdale, Lewis, & Rymer, 1986; Ganor &Golani, 1980; Greene, 1982; Lee, 1984;Nashner & McCollum, 1985; Shik & Orlov-sky, 1976; Soechting & Laquaniti, 1989; Viv-iani & McCollum, 1983).
Vigorous efforts to understand these is-sues have been made by workers who haveadopted computational formulations of thekinematics of motion. A brief discussion ofthese approaches will help to identify someof the current problems in this area and setthe stage for a contrasting treatment of theseproblems by selectional theories.
Kinematic Concepts andComputational Strategies
Kinematics.—The variability and flexi-bility of movements reflect the mechanicalredundancy of the musculoskeletal system,A redundant system can be defined as onein which the number of degrees of freedomis larger than the number of independentspatial variables of the mechanical compo-nents. In a mechanically redundant system.
964 Child Development
there is more than one way to generatea given trajectory through the motion ofthe joints involved. According to Saltzman(1979), "the degrees of freedom for a givensystem are the least number of independentcoordinates required to specify the positionsofthe system elements without violating anygeometric constraints." In the case of anopen kinematic chain consisting of multiplelinks with joints that revolve around a singleaxis of rotation (hinge joints), the number ofdegrees of freedom simply equals the num-ber of joints. Some joints of animal and hu-man limbs have more than 1 degree of free-dom; for example, the wrist joint is biaxial,the shoulder joint triaxial. The number ofindependent spatial variables depends alsoin part on the dimension of the workspace.In a two-dimensional (planar) workspace,the position of each link in a kinematic chaincan be specified by its Cartesian coordi-nates. Thus, in a two-dimensional work-space, kinematic chains with more than twolinks are redundant. In a three-dimensionalworkspace, six coordinates must be specifiedfor a given link, three for position and threefor orientation. Typically, the human arm (inits three main joints—wrist, elbow, andshoulder) contains 7 mechanical degrees offreedom and thus represents a redundantsystem in three-dimensional space. If onealso takes body motion or motion of theshoulder girdle into account, the effectivenumber of degrees of freedom grows sig-nificantly (up to 100, according to Turvey,1990).
Roughly speaking, kinematics dealswith the interconversion of joint and spatialvariables. Joint variables may comprise, forexample, a set of joint angles, angular veloci-ties, or angular accelerations, and are oftenexpressed in a polar coordinate system cen-tered at the base ofthe kinematic chain. Spa-tial variables are, for example, the course ofa movement path or the position or velocityof an environmental target and are often ex-pressed in Cartesian coordinates. The directkinematic transform describes the transla-tion of a given set of joint coordinates intoa geometric path of the kinematic chain.While this transform always exists uniquely,the inverse operation (called the inversekinematic transform) is indeterminate if thesystem is redundant. This means that anygiven path can be produced by a variety (po-tentially an infinity) of sets of joint anglechanges. A typical example (easily repro-duced by the reader) is a pointing movementof the arm along a given path. For each con-figuration of start point, end point, and path.
an infinite number of solutions can be car-ried out (see also Fig. IA). Cybernetic theo-ries of motor control and robotics hold thatcomputing the inverse kinematic transformfor a given desired movement path is the keyproblem that motor systems face. Because ofits indeterminacy, the problem is mathemat-ically "ill-posed."
In principle, two different strategies canbe adopted to compute a set of joint anglechanges for the inverse kinematic transform;
a) Decrease the number of independentjoint variables. The number of degrees offreedom that are used in the movement of amultilink chain can be reduced, for example,by freezing one or several of the joints. An-other possibility is to introduce constraintsthat allow the expression of one joint vari-able as a function of another. For example,if elbow movements can be expressed as afunction of shoulder movements, the num-ber of degrees of freedom has effectivelybeen reduced by one. This strategy is re-lated to the formation oi synergies of joint ormuscle ensembles.
b) Increase the number of specified spa-tial variables. In addition to specifying de-sired spatial coordinates for the end pointof the kinematic chain (usually carrying theeffector, e.g., a gripper), spatial coordinatesfor other links of the chain may be intro-duced (e.g., as criteria for optimization of themovement path in terms of length or mini-mal energy consumption). For example,such criteria could involve the requirementthat individual joints rotate as little as possi-ble or that distal joints move more than prox-imal ones.
Both of these strategies work by intro-ducing additional constraints, either on themotor apparatus itself or on the workspace.Let us see how these kinematic concepts canbe put to use in computational approaches.
Computational strategies.—Most com-putational approaches to the problem ofguidance and control of a kinematic chainfollow a number of defined steps. We chooseas an example a robot arm. First, the desiredtrajectory (in Cartesian coordinates) of theeffector attached to the arm is computed.Second, the inverse kinematic transform iscomputed, resulting in a set of joint anglesand joint angle changes that correspond tothe desired effector path. If real motors areinvolved in moving the robot arm, the ade-quate motor torque for each joint must becomputed in a third step.
The second and third steps involve com-puting inverse transforms. In principle,there are two computational strategies thatwill produce solutions for these steps (Hol-lerbach, 1982; see also Loeb, 1983):
a) The joint angle changes and torquesproducing a desired movement path can befound by directly solving the set of dynami-cal equations that describe the behavior ofthe multijointed arm. For realistic motionsof a robot arm, it is important to solve theseequations in real time, that is, all computa-tions have to be done at least as fast as in-coming signals arrive. Computational strate-gies of this class are hampered by the factthat a large number of (often nonlinear)terms has to be taken into account. Theseinclude inertial forces such as link interac-tion, centripetal and Coriolis forces, and avariety of other effects due to gravity. Thedynamical equations for a multijointed armin three-space can thus become extremelycomplicated, and computations of even sim-ple trajectory fragments tend to take signifi-cant computational time. A complete set ofNewtonian equations for position and jointvelocities of a three-jointed arm in three-space contains 1,600 terms and requires13,000 multiplications for its solution (afterRaibert, 1978). The equations to computethe inverse dynamics of motions for a modelwith five degrees of freedom of the upperarm alone can themselves occupy two entirepages of closely spaced text (after Hogan,1988). Attempts have been made to simplifythe systems of equations and associated al-gorithms sufficiently, to allow fast and reli-able computation (Hollerbach, 1980; Luh,Walker, & Paul, 1980). The continual in-crease in computer power is likely to makethis strategy more feasible for many applica-tions, but its biological relevance remainsmarginal.
b) A second class of solutions to the in-verse dynamics problem involves the re-trieval from memory (in the form of a "look-up table") of the desired joint torques for agiven desired movement ofthe multijointedarm (first introduced by Raibert, 1978; Rai-bert & Horn, 1978). However, the amount ofmemory needed is very large and increasesdramatically with the dimensionality andcomplexity of the task. Furthermore, a pre-cise index is needed for retrieval, and thestored items may have to be updated in-stantly, particularly if the mechanical con-figuration or mass of a kinematic chainchanges abruptly (as happens when a load ispicked up).
Sporns and Edelman 965
What both approaches have in commonis that they attempt to solve the inverse kine-matics problem directly by computation.Neither strategy makes use of synergies asfunctional components of the motor ensem-ble or ofthe dynamic and associative proper-ties of neural architectures in order to con-strain the number of possible solutions.Neural network models have been claimedto take some of these issues into account intheir approach to the inverse kinematicsproblem.
Neural network models,—Computa-tional approaches to motor control havebeen enriched by a recent resurgence of in-terest in neural network models (e.g., Ber-kinblit, Celfand, & Feldman, 1986; Bullock& Crossberg, 1988; Hinton, 1984; Kawato,Furukawa, & Suzuki, 1987; Kawato, Isobe,Maeda, & Suzuki, 1988; Kuperstein, 1988;Massone & Bizzi, 1989). Much ofthe appealof these models appears to have arisen fromtheir resemblance to real brain structures, aresemblance that is nonetheless often onlysuperficial. Most network models share acommon feature with more conventionalcomputational approaches. The movementof a kinematic chain is determined by a "de-sired trajectory." This is either precomputedand presented to the model as a desired out-put vector to which network behavior mustconverge (Kawato et al., 1987, 1988), or it isrepresented internally (computed from sen-sory input) and then serves to guide an on-going movement to its goal (Bullock &Crossberg, 1988; Hogan, 1988).
Hinton's (1984) iterative model for thegeneration of reaching movements containsrepresentations of joint angles and positionsin addition to a "desired vector" from thetip of the arm to the target. Joint angles areupdated in a parallel fashion while the de-sired vector is minimized or shortened. Hin-ton notes that elimination of interactions be-tween individual joints will frequently leadto grossly suboptimal trajectories. To rem-edy this situation, he proposes that, undercertain conditions, synergies act as computa-tional subroutines.
Bullock and Grossberg (1988) have de-scribed a model of arm movements calledthe vector-integration-to-end-point model. Atarget position command specifies where thearm is intended to move, and an indepen-dently generated second command specifiesthe movement speed. An arm trajectory isthen computed from a present position vec-tor and a difference vector that is continu-ously updated and that specifies the differ-
966 Child Development
ence between present and target position.The present position vector (in connectionwith the difference vector) determines anoutgoing motor signal leading to motion.The model essentially depends on the con-tinuous updating of the difference vector,presumably by sensory feedback signalssampled during motion. A model requiringconstant updating may be inadequate forrapid movements such as saccades or ballis-tic reaching.
Ritter, Martinetz, and Schulten (1989)have constructed a model containing neuralmaps that learns to position an arm undervisual feedback. Ultimately, the arm moveswith high accuracy to a target position. Theauthors found that, if some subregions ofspace are covered more frequently than oth-ers, the sensorimotor map adapts by recruit-ing more neuronal units in that area. An-other model of sensorimotor coordinationhas been developed by Kuperstein (1988;see also Grossberg & Kuperstein, 1986; Ku-perstein, 1991). The arm used in their modelhas 5 degrees of freedom and operates inthree-dimensional space. During reaching,the arm travels instantly from one equilib-rium position to another (that is, joint anglesare linearly proportional to muscle activity).During the first stage of training, the "eyes"of a robot are fixed on the target by anunspecified mechanism. The arm is allowedrepetitively to position itself by randommotions from a standard starting position.During these movements, kinesthetic eyeposition signals and target vision are sam-pled and correlated with the random motorsignal in simulated neural networks. Thediscrepancies between actual postures andthe ones computed from the sensory infor-mation are split into respective componentsfor individual sets of muscles and the synap-tic weights in the networks are changed inorder to minimize this discrepancy. Aftertraining ofthe system, the random generatoris removed and a visual target can then drivethe individual joints, using the maps con-structed in the first stage.
In summary, most of the existing net-work models require both the setting of adesired path and the specification of fairlyelaborate feedback signals, such as differ-ence or error vectors, in order to converge toa desired mode of action. However, thesefeedback signals are often not specified inbiological terms. While it has certainly beenshown in many experiments (e.g., Bauer &Held, 1975; Held & Bauer, 1974) that sen-sory input is essential for ongoing purpose-
ful movement as well as motor learning, thepredominant view of learning as conver-gence to a previously determined or pre-scribed state, and thus the instructive natureof such inputs, is far from proven.
Dynamic systems theory.—Recently, at-tempts have been made to link the emer-gence of coordinated movements to con-cepts of nonlinear systems theory. Thisapproach was originated by Kelso (for re-view see Kelso & Tuller, 1984; Schoner &Kelso, 1988), as well as Kugler and Turvey(1987; see also Kugler, Kelso, & Turvey,1982). According to this approach, coordi-nated movement is produced by many inter-acting elements with potentially many de-grees of freedom, constituting a nonlinearsystem which can attain a certain numberof dynamic states. For example, rhythmicswimming motions can be described by rela-tively few parameters (such as frequencyand amplitude), that is, are characterized byonly a few degrees of freedom, even thoughmany neurons, muscles, and bones may beinvolved in their generation. The interest inrhythmic movements of this kind dates backto Erich von Hoist (1937); recent work onrhythmic movements in animals and humansviewed in the context of dynamic systemstheory includes the studies by Schmidt,Beek, Treffner, and Turvey (1991); Schonerand Kelso (1988); Thelen, Skala, and Kelso(1987); and Ulrich (1989). More recently,Thelen has proposed that developmentmight be understood as a temporal sequenceof coordinative modes or attractor states(Thelen, 1990). The transition from one stateto another would be under the control ofrelatively few developmental control pa-rameters; the emergence of coordinated legmovements in infants has been investigatedfrom this perspective (Thelen, 1989b).
Many basic concepts (such as the impor-tance of nonlinear self-organizing inter-actions within a complex system) of thesedynamical theories are consistent with se-lectionism (see Kelso & Tuller, 1984). Likeselectionism, dynamical systems theory re-jects instructionism and an algorithmic treat-ment of learning and motor development.However, selectionism differs in that it isexplicitly based on neuroanatomy and neu-rophysiology and explicitly recognizes theimportance of analyzing the interactions ofmultiple levels of organization (such as neu-ral circuits or limb biomechanics). In partic-ular, dynamical systems theory by itself doesnot identify specific neural mechanisms ofdeveloping and mature motor systems.
Sporns and Edelman 967
The next section contains a broad out-line of our proposal to abandon the instruc-tive or computational paradigm and look atthe problem from a new selectional per-spective.
An Approach to Solving Bernstein'sProblem by Selection
There is overwhelming evidence thatthe emergence of coordinated movements isintimately tied both to the growth of themusculoskeletal system and to the develop-ment ofthe brain. Thus, neural developmentand learning cannot be considered outsideof their biomechanical context. A key theo-retical issue is how changes in brain cir-cuitry controlling muscles and joints becomematched to simultaneously occurring devel-opmental changes at the periphery (the in-terface between the musculoskeletal systemand the environment ofthe organism). Howcan coordination emerge from the manycomponents and interactions of such a sys-tem? A promising approach is to considervarious aspects of this question in terms ofpopulation thinking (Mayr, 1959). The the-ory of neuronal group selection (Edelman,1978, 1987, 1989, 1993) is a consistent at-tempt to apply population thinking to thefunctioning of the nervous system and pro-vides the basis for the present proposal.
Neuronal group selection.—The theoryof neuronal group selection places great em-phasis on the structural variability of braincircuitry. During development, neuronal cir-cuits are not precisely wired at the level ofmicroanatomy. Thus, the brain contains rep-ertoires of variant circuits (structural vari-ability) that can give rise to many differentoutputs (dynamic variability). Those variantcircuits form neuronal groups, local collec-tives of several hundreds to thousands ofmore strongly interconnected neurons thattend to share functional properties and todischarge in a temporally correlated fashion.The postulated neuronal groups are consid-ered to be the basic functional units or unitsof selection. They have recently been exper-imentally identified in several cerebral corti-cal regions (Gray & Singer, 1989).
In the cortex, neuronal groups are ar-ranged in neural maps, representing, for ex-ample, visual space or the body surface.While these neural maps are often function-ally segregated and occupy circumscribedregions of the cortex, they are anatomicallycoupled through long-range connections.Virtually all of these connections are recip-
rocal. Recursive reciprocal signaling (calledreentry) between neuronal groups in distantsensory and motor regions gives rise to newdynamical properties, including temporalcorrelations (Sporns, Tononi, & Edelman,1993; Tononi, Sporns, & Edelman, 1992a,1992b). In general, reentry is the key opera-tion that serves to integrate the activities ofthe multiple sensory and motor areas of thebrain.
How do these ideas relate to populationthinking? Repertoires of variant neuronalgroups are subject to somatic selection (i.e.,selection occurring during the lifetime oftheorganism). This selection occurs for particu-lar groups when their activation in a givencontext matches given environmental andinternal constraints as compared to compet-ing groups. For example, particular groupsmay be selected for their contribution to per-ceptual categorization or to motor tasks. Themechanism of selection in the nervous sys-tem is synaptic change (by a variety of de-tailed mechanisms) leading to the selectiveamplification or diminution of neuronalgroup responses. The selection of neuronalgroups ultimately allows the discriminationand categorization of sensoiry inputs and theintegration of sensory and motor processesto yield adaptive behavior. This integrationdepends upon higher-order relationships.For example, dynamically coupled sets ofneuronal groups in a variety of functionallysegregated maps linked by reentrant con-nections that are active for a given sensori-motor task can constitute a global mapping(see Edelman, 1989). By its nature, a givenglobal mapping involves widespread anddistributed regions ofthe brain.
Neuronal group selection by no meansrelies purely on mechanisms of eliminationor regression, as has been implied by others(Purves, 1988). Simple regression (i.e., over-production of neural connections and subse-quent elimination of "inappropriate" ones indevelopment) may occur in some isolated in-stances, but is not general enough to accountfor either somatic or evolutionary selection(Edelman, 1987, 1989). Instead, the theoryof neuronal group selection stresses the im-portance of both stabilization of selectedneuronal circuitry (including the possibilityof generating new connections, under favor-able circumstances) and balanced compe-tition.
Selection in sensorimotor develop-ment.—How do these notions apply to Bern-stein's problem? We propose that the de-velopment of sensorimotor coordination
968 Child Development
proceeds in three concurrent steps: (a) thespontaneous generation during develop-ment of a variety of movements forming abasic movement repertoire; (b) developmentof the ability to sense the effects of variousmovements in the environment, eventuallyallowing neural selection to be guided byadaptive value (for a discussion oi value, seebelow); and (c) the actual selection of move-ments in various global mappings under theconstraint of adaptive value. Selection in thenervous system is mediated mainly via syn-aptic change, resulting (on average) in thestabilization of those brain circuits that sup-port adaptive (or goal-directed) movements.Somatic selection in the nervous system andmotor action at the periphery are thus subtlyintertwined through the action of globalmappings. Successive selection events willresult in a progressive modification of agiven movement repertoire and will thusshape the ongoing motor activity of the or-ganism. On the other hand, an emergingmovement repertoire (occurring as a resultof changes in biomechanics or environmen-tal demands) will also drive selection toyield consistent and adaptive coordination.
A natural consequence of selection isthe emergence of synergic relationships be-tween members of the motor ensemble. In-deed, the formation of synergies by selectioneffectively replaces the need to solve theproblem of inverse kinematics by computa-tional means. Instead of having explicitlyand accurately to compute a desired trajec-tory in order to reach a goal, animals selectthose purposeful movements from a diverserepertoire that fulfill adaptive constraints.After selection, synergies will emerge thatare formed by the amplification of move-ments that are favored in a given task andcontext. Thus, a model based on somatic se-lective processes eliminates the need explic-itly to compute systems of nonlinear differ-ential equations containing many kinematicand dynamic terms. Rather, we suggest that,through spontaneous and exploratory motoractivity, the organism discovers possible"solutions" that satisfy the task constraintsas well as internal value. The variability anddegeneracy that are so vexing in computa-tional or kinematic approaches are in facta necessary prior condition fbr successfulselection. The synergic, "topological" and"gestalt" character of movements emergeunder a given set of mechanical and neuralconstraints because movements are selectedas whole patterns, without being decom-posed into parts that are subject to separate
analysis and adjustment by instructive learn-ing algorithms.
A selectional scheme also allows preex-istent or preferred movement patterns to bereadily integrated with subsequent motorlearning and adaptation. There is ample evi-dence that organisms are born with a consid-erable set of intrinsically specified motionpatterns (presumably the result of evolution-arily selected circuits in certain parts of thebrain). Such characteristic movement pat-terns can even be observed at embryonicstages (Bradley & Bekoff, 1990; Smotherman& Robinson, 1988); indeed, prenatal motilitymay be important for the anatomical speci-fication of neuromuscular connection pat-terns (Coghill, 1929; Hamburger, 1970).Preexistent patterns impose considerableconstraints on the basic movement reper-toire of an organism and on subsequent mo-tor development. The selective history of anorganism is critically influenced by suchpreexistent patterns which can guide thetransformation of the primary movementrepertoire into a set of adaptive action pat-terns. These issues bear upon that of value.
Value.—Inasmuch as a selectional sys-tem operates upon preexistent diversity anddoes not have a specific program for mostrefined movements, the question of con-straints upon selection arises. According tothe theory of neuronal group selection(Edelman, 1989), these constraints are pro-vided by value systems already specifiedduring embryogenesis as the result of evolu-tionary selection upon the phenotype. In thecase of movements, such constraints appearin neuroanatomy as specific circuitry signal-ing via particular neurotransmitters. Suchvalue circuits are prespecified by evolution-ary selection rather than derived from expe-rience. The crucial features of such valueschemes are their anatomical simplicity andthe diffuseness of their effects on their targetnetworks; these features sharply distinguishvalue schemes from the instructive agentsand error feedback employed in most learn-ing algorithms. In general, value circuitscannot themselves lead to the variety ofhighly articulated movements necessary fordevelopment and survival. In this view, incontrast to instructive views, the coordina-tion of movement occurs ex post facto as aresult of somatic selection upon value thathas been established by evolutionary se-lection.
Value systems involved in sensorimotordevelopment receive sensory inputs that en-
able them to respond to actions or events inthe environment by increasing or decreasingtheir level of neuronal activity. Due to theirevolved anatomical structure, the level of re-sponse of such value systems is related tosimple criteria of saliency or adaptiveness.For example, a reaching movement estab-lishing tactile contact with an object may re-sult in increased neuronal firing in the valuesystem, thus signaling the adaptiveness ofthe movement. This value system might re-ceive sensory inputs from tactile receptorslocated in the skin.
In order to be effective on sensorimotordevelopment, value systems must be abledifferentially to affect synaptic changes incircuitry controlling movement. For exam-ple, increased activity in a value system caninfluence synaptic changes involving a largemajority of those neurons that participatedin a successful (adaptive) reaching move-ment. As a result of a positive change (givenpositive value) in the underlying circuitry,this adaptive movement is more likely to beexecuted in the future. It is important tostress that the action of value systems is bothdiffuse and probabilistic. Because outputfibers of value systems branch widely overextended regions ofthe cerebral cortex, theyinfiuence synaptic modification in largeparts of the nervous system. Given this dif-f'use effect of value, there can be no localdetermination ofthe contribution of individ-ual neurons or synapses to a given move-ment; instead, value acts on whole popula-tions of neurons. Therefore, value does notact deterministically but probabilistically.While on average (over time) selection uponvalue will lead to increasingly adaptive be-havior, on individual trials some members ofthe neuronal populations may exhibit "un-desirable" synaptic changes (driving the sys-tem away from a more adaptive state). Suchongoing effects may serve an important func-tion in maintaining variability as a basis forfurther selection. We will give a specific anddetailed example of how a value system actsin the section on computer simulations.
An important issue concerning valuehas to do with the specificity or generalityof value systems. It seems unlikely that aseparate value system exists for each motortask that an organism might carry out duringits lifetime. Many different levels of pheno-typic constraints act to define value systems.These include morphology itself, the acqui-sition of new neurotransmitters, interactionsbetween cortical and midbrain homeostaticsystems, and hormonal changes. There is no
Sporns and Edelman 969
simple relation between the mapping ofmotor control systems and combinations ofthese different constraints on value. Giventhe fact that value is evolutionarily selectedfor in the species and varies with differentspecies, no simple one-to-one correspon-dence between value and a given motor sys-tem would be expected. Clearly, since motorresponses are achieved differently in differ-ent individuals during ontogeny, their possi-ble patterns are much more numerous thancould be arranged by evolutionary selection.We would expect that different value sys-tems are activated for a variety of function-ally interrelated tasks, that different valuesystems might interact, or that hierarchies ofspecificity might exist. It is consistent withthis general view that, as compared to spe-cific acquired cortical responses and theirinteractions, there seem to be relativelyfew value-related "saliency" systems in thebrain and that these systems project to verywidespread and diverse regions of the cere-bral cortex. Examples are the hypothalamus,the locus coeruleus, the raphe nuclei, andthe cholinergic basal forebrain system. Howspecificity of learning is achieved throughthe combined action of such systems withthe cortex is a deep problem that requiresmore attention. We believe that the issue ofvalue constraints and their number presentsone of the greatest future challenges to se-lectional theories of brain function. A possi-ble insight may be gained by consideringdifferent combinatorial arrangements of sys-tems of value as they act on different motorarrangements.
Selection from a movement reper-toire.—It is evident from the above outlineof a selectional model that the formation ofsynergies requires not only the interactionof the organism with the environment butalso a means by which the adaptive valueof a movement can be assessed and in turninfiuence future movements. A schematic di-agram illustrating the selectional principlefor coordination of movement is given inFigure 2. We assume for simplicity that amovement such as a simple gesture is char-acterized by values of a set of n joint vari-ables <!)„. The set of all possible combina-tions of these n joint variables thus forms ann-dimensional movement space M (for sim-plicity n = 2 in Fig. 2). Initially, the subsetof movements that are available to the organ-ism is only constrained by the mechanics ofits motor ensemble and preexistent move-ment structures, but is basically uncon-strained by experience in the environment.
970 Child Development
TIME 1 TIME 2 TIME 3
Primary MovementRepertoire
Preexistent MovementPattern
Developing MovementRepertoire
FIG, 2,—Schematic diagram of a developing movement repertoire contained in a movement spaceM. A single movement within the space is specified by a combination ofthe movement variable cbi andi>2; it is represented as a small dot. The dot density represents the frequency with which movementsare executed in a particular region of M. The three frames depict different temporal stages. At the leftis the primary movement repertoire containing several preexistent (or "innate') movement patterns.The shape of the movement repertoire evolves with time to include previously unoccupied regions ofM or to exclude others. Hatched regions define movement patterns that correspond to a given task.Movements within these regions 9 meet with positive adaptive value. As a result, their frequencyincreases. With time, due to changing environmental and biomechanical constraints, both the move-ment repertoire and the regions 9 will continue to change shape (see middle and right).
This primary movement repertoire is thuslargely unstructured—with the exception ofpreexistent synergic relationships betweenthe relevant movement variables (()„, At theonset of sensorimotor experience, the organ-ism is confronted with demands from the en-vironment, while at the same time it is sub-ject to its own value systems. For example,during spontaneous motor activity, somemovements will help to accomplish a taskbetter than others. In our scheme, suchmovements, often primarily defined by con-straints in the environment (as in a spatialreaching task), but also by constraints im-posed by the geometry of the kinematicchain, might form one or more subsets 6within the movement space M (Fig, 2), Be-cause of mechanical redundancy for eachgiven task, these subsets may span extensiveregions in M.
Somatic selection in the nervous systemresults from the competitive strengtheningof neural connections (synapses) involved inthe generation of "successful" movements(e.g., those involved in touching an object orbringing it to the center of the visual field).Those giving rise to other movements areweakened or left unchanged, depending onthe particular synaptic mechanism that isinvoked. As a result of selective synapticchange, movements that help to accomplish
the task become more probable on averagethan others. Because of individual variationsin the biomechanics of motor organs, theirprogressive structural and dynamical changeduring development, and the unpredictablenature of environmental demands, these"desirable " subsets 6 will appear quite dif-ferent, not just for different species, but alsofor different coexisting individuals of thesame species. Thus, there are narrow limitsfor "preprogrammed" or "hard-wired" neu-ral control of movements; the organism mustrely both on spontaneous (though not neces-sarily random) motor activity and the subse-quent selection of those movements that si-multaneously match environmental demandand internal value.
In this scheme, synergies control move-ments, forming different task-related subsets8, The set of movements comprising func-tional synergies is often degenerate (Edel-man, 1987) in that it contains movementsthat are functionally related (they all accom-plish the same task) but are structurally non-isomorphic (they utilize different joint vari-ables or muscles groups to different extents,and are controlled by different sets of neu-ronal groups). The degenerate structure offunctional synergies provides a basis for mo-tor equivalence. By allowing fiexibility (Par-tridge, 1986) and by forming a diverse sub-
strate for further selective events, a richdegenerate structure yields evolutionary ad-vantages to an organism.
Experiments in many areas of neurosci-ence and psychology are needed to substan-tiate and refine the key points of the pro-posal briefly described here. But it is alsoimportant to show how the proposed mecha-nism actually can work. One way to showits overall feasibility and self-consistency isto simulate on a supercomputer a structur-ally and functionally realistic motor systemworking on selectional principles.
Computer Simulations
For several years, we have pursued anapproach to the understanding of higherbrain functions called synthetic neural mod-eling (for a review, see Reeke, Finkel,Sporns, & Edelman, 1990; Reeke, Sporns, &Edelman, 1990), This approach attempts toaddress the complex problem of integrativebrain function by using computers to carryout large-scale simulations of neuronal cir-cuits acting together with phenotypic organsthat are embedded in an environment. In thesimulations discussed in this section, we re-quire that afferent sensory signals (comingfrom visual or kinesthetic receptors) be sam-pled both during and after the occurrence ofa movement, A second requirement is thatan intrinsic bias or value must exist favoringmodes of behavior that are more adaptivethan others; an example of such a favoredaction might be reaching out and touchingan object. The first instantiation of an auton-omous "artificial creature," Darwin III, con-sists of a complex nervous system, an arm,and an eye. It behaves in a two-dimensionalenvironment containing sensory stimuli,Darwin III is able to track objects visuallyand reach for them; visual and tactile sen-sory inputs are combined to allow complexperceptual categorizations of a large varietyof stimulus objects. For more detailed de-scriptions of all of Darwin Ill's components,see Reeke, Finkel, Sporns, and Edelman(1990) and Reeke, Sporns, and Edelman(1990), Here we focus exclusively on its mo-tor functions,
Darwin III contains a four-jointed armmoved by four sets of independent agonist-antagonist muscle pairs operating in two-dimensional space. The system thus has sur-plus degrees of freedom and is mechanicallyredundant. The basic idea behind the modelis that, through experience, gestures or ges-tural components (corresponding to syner-
Sporns and Edelman 971
gies) are selected from a relatively unstruc-tured primary repertoire of movements. Themodel has two main functional compo-nents (see Fig. 3): a module correspondingroughly to the cerebral cortex, where motorsignals eventually leading to gestural mo-tions are generated, and a module corre-sponding roughly to the cerebellum, whichcorrelates these motor signals with currentsensory inputs and smooths the responsesby eliminating components that are inappro-priate. Hand and object vision as well as asimple form of kinesthesia and light touchprovide the sensory signals utilized by thesystem. The different signals from these mo-dalities are transformed through mappedand unmapped interactions of separate neu-ronal networks present in the two modulesinto motor signals that move the arm to newpositions. It must be understood that, whilecomponents of the model perform functionssimilar to those in real brains (and are namedanalogically), they are not modeled in ex-plicit detail. The model is simple as com-pared to real vertebrate brains; Darwin IIIessentially contains a total of 50,000 neuronsand 620,000 connections, organized in about50 neuronal repertoires.
The "motor cortex" generates patternsof activity corresponding to primary gesturalmotions through a combination of spontane-ous activity (triggered by a component ofGaussian noise) and by responses to sensoryinputs from vision and kinesthetic signalsfrom the arm. This activity pattern is selec-tively remodeled ("sculpted") under the in-fluence of inhibitory connections comingfrom the "cerebellum," Ultimately, the "cor-tical" module sends out connections to foursets of motor neurons (one for each joint) or-ganized in extensor/flexor pairs.
The "cerebellar" networks receive sen-sory inputs (vision and kinesthesia) as wellas activity driving the current primary ges-ture emanating from the "motor cortex." Thenetworks respond to specific combinationsof the actual positions of the arm and targetand to combinations of activity patterns cor-responding to primary gestures. Their inhib-itory infiuence (negative selection) on pri-mary gestures as they are generated in themotor cortex serves to select or "filter out"those gestures that are appropriate.
Both "motor cortex" and "cerebellum"in Darwin III are topographically ordered inthat they contain groups of units that primar-ily connect in an ordered sequence to single
972 Child Development
FIG 3 —Schematic diagram ofthe neural circuits controlling Darwin Ill's four-jointed arm system.For detailed explanation see text. Main modules are labeled "MOTOR CORTEX, CEREBELLUM"(with GR = granule cells and PK = Purkinje cells), and VALUE, Inputs to modules are visual (V) andkinesthetic (K), Value-dependent modification of connections to and from the cerebellum is mdicatedby dashed lines.
joint extensors or flexors. This arrangementis consistent with anatomical and physiolog-ical evidence for such motor maps. Intrinsicconnections in the "motor cortical" network(not shown in Fig. 3) are biased to favor thespread of activity from units representingproximal to more distal joints. Gestural mo-tions therefore tend to unfold proximodis-tally, a useful preexistent characteristic forthe large-amplitude reaching movementsunder study in the simulations. Although acertain degree of topography is present inthe system from the beginning, there is noexplicit representation of either "move-ments," "muscles," or "joints" within themotor cortex. (What is represented in actualmotor cortex is still an open and muchdebated question; see Humphrey, 1986;Lemon, 1988.)
Initially, the connectivity of the reach-ing system of Darwin III is uninstructedwith respect to appropriate directions andamplitudes of movement. Coordinated mo-tions of sets of joints occur only later, aftergestural components have been selectedfrom spontaneous movements by amplifica-tion of those synaptic populations that giverise to such gestures. To detect and signalthe consequences of such successful motor
activity, neuronal circuitry instantiating anintrinsic value scheme is required. In Dar-win Ill's reaching system, this circuitry con-sists of a set of neurons responding morestrongly as the moving hand approaches thevicinity of the visually foveated target ob-ject. These neurons receive topographicallymapped visual inputs from two separateneural repertoires responsive, respectively,to objects in the environment and to thehand of the automaton itself. Each of theseinputs arborizes in an overlapping fashionover the surface of the value network, andthresholds are arranged such that correlatedactivity in both inputs is required for a vigor-ous response. The responses of the valueunits thus increase, independent ofthe abso-lute position of the target, as the hand ap-proaches the target and as the degree ofoverlap in the topographically mapped in-puts increases.
The activity in the value repertoire istransmitted to parts of both the "motor corti-cal" and "cerebellar" networks and servesselectively (but only probabilistically) tostrengthen or weaken populations of syn-apses. After repeated amplification, appro-priate activity patterns in the cerebellumwill arise as soon as a target appears in the
visual field and before a gesture is initiated.Its inhibitory connections terminating in themotor cortex are thus available for "filtering"and "sculpting" gestures as they happen.
As a biologically based system, DarwinIll's reaching system offers several distinctadvantages over neural network models,such as that described by Kuperstein (1988,1991): (a) No instructive learning algorithmis used to adjust synaptic weights. Instead,Darwin Ill's networks are subject to selec-tive amplification depending on a valuescheme (no error vector is used), and synap-tic changes simultaneously affect neuronsrelated to all components of the movement.On the other hand, Kuperstein's model re-quires the determination of an error vectorand the splitting of this vector into compo-nents used to adjust synapses related to spe-cific joints. While Kuperstein's calculation ofthe error vector is accomplished by a compu-tational subroutine, value schemes in Dar-win III are implemented as specialized net-works that are anatomically and functionallyintegrated with the rest of the model, (b)Kuperstein's model operates as a look-up ta-ble of movements arranged in a sensorimo-tor map; it does not use synergies to reducethe number of degrees of freedom. In fact,its 5-degrees-of-freedom arm is nonredun-dant in three-dimensional space. The net-work architecture does not allow for interac-tions between individual joints, (c) Eachindividual reaching movement in DarwinIII evolves and changes in time and is elabo-rated over multiple time steps. This allowsthe evaluation of the shape of arm trajecto-ries, a very important characteristic of hu-man reaching movements. In contrast, areaching movement in Kuperstein's modelconsists of only one step from the initial po-sition to the end point; this excludes the pos-sibility of characterizing trajectories, (d) Incontrast to Kuperstein's model, Darwin IIIdoes not operate in distinct training and per-formance modes. Instead, the developmentof synergies by selective amplification andactual performance ofthe arm are concurrentprocesses; it is not necessary (as in Ku-perstein's model) to connect or disconnectcomponents of the model when switchingfrom training to testing,
Darwin Ill's reaching movements maybe examined by looking at a short sequenceof simulation cycles (Fig. 4) or by consider-ing diagrams showing plots of bundles of tra-jectories (Fig. 5). The large variance amongindividual motions that exists initially (Fig.5A) is progressively reduced to a narrow en-velope of motions, most of which point to-
Sporns and Edelman 973
ward the object (Fig. 5B). This result resem-bles data shown by Georgopoulos, Kalaska,and Massey (1981, see especially their Fig,3), who have studied the geometry of point-ing movements in rhesus monkeys duringlearning, (Of course, this resemblance doesnot by itself indicate that the underlyingneural processes are identical,)
More detailed observation of singlejoints in Darwin III (e.g., "elbow" and"shoulder") reveals that, after training,changes in joint angles are no longer inde-pendent of each other (Fig, 6), Instead, jointsact together in more or less stable ratios and,as a result of this synergic interaction, theeffective number of degrees of freedom isreduced. Such relationships exist only intrained movements (Fig, 6B), but are absentbefore training and selective amplification(Fig, 6A), We have observed linkages be-tween proximal as well as distal joints andbetween joints that are not immediately ad-jacent to each other (data not shown). Jointrelations may be linear or nonlinear, varyingfrom example to example, reflecting the dif-ferent requirements of parts of the work-space, of arm geometry and neural struc-tures, as well as differences in the individualhistory of selectional events.
The complex anatomical organization ofthe system does not allow precalculation ofthe relevant neural pathways for a given"desired" movement. Given the mechanicalredundancy of Darwin Ill's arm, there aremultiple degenerate means for each trajec-tory to reach a given end point. We havepointed out above that one way to solveBernstein's problem (and to reduce the num-ber of degrees of freedom in a multilink ki-nematic chain) is to group muscles or jointstogether to form synergies. In the presentmodel, selection constrains the envelopeof possible motions; as a result, synergiesemerge. Thus, selection guarantees execu-tion of one or another ofthe numerous solu-tions that lead to a successful outcome. Notethat Darwin Ill's reaching system does notwork by determining a desired trajectory; infact, trajectories are not explicitly repre-sented anywhere within the system. Thus,the model embodied in Darwin III mayserve as a first working example to illustratethe benefits of a selectional strategy in creat-ing synergies,
A system designed to execute a numberof different synergies must incorporate neu-ral maps that allow the selection of the ap-propriate synergy for a given sensory con-text. Synthetic neural modeling allows one
ENVIRONMENT
I 1—
[ 0
0
0
o
SR'0'
'K
VALUE VALUE0 '
•Oo.
BENVIRONMENT
FIG, 4,—Display of Darwin Ill's four-jointed reaching system on three consecutive cycles A, B,and C, At each cycle Darwin Ill's environment and the state of its neuronal repertoires (with circlesrepresenting active neurons) are displayed. The neuronal repertoires, plotted as planar maps of neu-ronal units, are labeled as follows: VALUE, "value system"; HV, "hand vision"; TH, touch"; WD,"world"; MC, "motor cortex"; IN, "intermediate layer ofthe motor cortex"; SG, "spinal ganglia"; KE,"kinesthesia"; IO, "inferior olive"; GR, "granule cells"; PK, "Purkinje cells," In addition to object(dark square) and visual field, the environment display contains a four-jointed arm, anchored at theijottom of the environment. Repertoire WD reports the position of the target object to MC and GR,networks representing motor cortex and granule cells of the cerebellum, respectively. In addition tothis visual input, MC and GR receive subthreshold inputs from KE, a network that reports kinestheti-cally the positions ofthe four joints. Notice how the activity in KE varies systematically with changingarm position. The pattern of activity in MC is transmitted to IO, which sends a number of strongexcitatory connections to a repertoire, PK, representing Purkinje cells. Units in IO only fire if positivevalue occurred in the previous cycle (as happens in C); value acts as a modulatory signal that allowsactivity in MC to be transmitted to PK ("gating"). There are also a great number of initially weak
974
64
0 64
X-Position (Pixels)
64
Fic:, 5,—Examples of paths taken by the tip ofthe four-jointed arm as it reaches for square objectspresented at the marked locations in the input array (compare Fig, 4), Left and right panels in A andB display data sampled before and after 1,440 cycles of training, respectively. Each panel displays 480cycles (,30 trials), recorded while synaptic amplification was frozen, A and JB give examples of motionsstarting at different initial conditions. The initial set of joint angles was 60, 100, 200, and 90 degreesfor A, and 25, 140, 200, and 90 degrees for B (see Fig, 6 inset). From Reeke and Sporns (1990),reproduced with permission.
connections between GR and PK; these connections, as well as descending inhibitory fibers from PKterminating on IN, are modifiable. The PK inhibitory fibers act to select appropriate motor signals forgestures that are generated in MC and transmitted to the motor neurons, SG, through IN, Thus it isthe pattern of active units in IN that determines which path the arm will describe. The IN activitypattern in A determines that the arm will move toward the target and is the result of a combination ofexcitatory infiuences from MC, as well as inhibitory action of PK units, PK units can be activated bystrengthened connections from GR alone, and the IO network becomes unnecessary after training iscompleted. On impact on the target (B and C), touch-sensitive neurons in a sensory sheet tied to thetip of the arm, TH, become active and guide the subsequent tracing of the edges of the object (notshown), A value-sensitive network (VALUE) receives topographically mapped inputs from both thetarget visual area WD, as well as from a repertoire sensitive to the position ofthe hand (HV), VALUEfires more vigorously when the hand approaches the target (compare A vs, B); the glohal output of thisrepertoire is used to modulate activity in IO and amplify cormections between WD and MC, Theconnections between GR and PK, as well as those between PK and IO, are modified without theparticipation of value, but modification can only occur if the IO network is active and fires PK units.The inhibitory action of PK units results in freezing of the second joint while the first joint flexes A,On impact gross arm motion is inhibited B, and an additional motion of the third and fourth jointestablishes full contact C (for more complete information see Reeke, Finkel, Sporns, & Edelman, 1990),From Reeke, Sporns, and Edelman (1989), reproduced with permission,
975
Sporns and Edelman 977
to explore the issue of mapping a sensoryinput .space onto a map of synergies (or ges-tures) by incorporating different neural andphenotypic structures into any given model.Indeed, we have designed a second motorsystem for Darwin III with more sophis-ticated neural maps and have been ableto achieve reliable and accurate reachingmovements over the entire input space (for adetailed description, see Sporns, 1990). Themechanics ofthe arm were simplified by re-ducing the number of joints to two. Whileworking according to the same selective par-adigm used to create synergies in the modelwith the four-jointed arm, the model incor-porating neural maps exceeded the perfor-mance of the previous model in several re-spects. The model was able to provide foraccurate reaching movements from an ar-bitrary starting position to any target posi-tion within the workspace. Unlike the firstmodel, such reaching movements could beperformed at the same time the eye wasmoving, for example, while saccading to avisual object (Fig. 7). Furthermore, themodel was able to perform over a wide rangeof changes in arm size or metric without ex-ternal adjustments of structural or dynamicparameters in the nervous system. A suddenchange in the biomechanics of the arm ledto an initial decrease in performance accu-
racy. Continuing somatic selection of cir-cuits, however, provided a sufficient basisfor progressive adaptive change. This exam-ple provides a demonstration that develop-mental and evolutionary changes in the bio-mechanics of motor organs can becompensated for by accompanying somaticselective changes in the nervous structurescontrolling their action. This ability is oneofthe strongest arguments for somatic selec-tion in the brain (Edelman, 1987).
In conclusion, the computer simulationspresented in this article show the self-consistency of an approach based on se-lectional principles. Taken together withexperimental evidence on sensorimotordevelopment (e.g., Ulrich, 1989; see also thereview by Thelen, 1989a), they make a casefor the role of selection in the emergence ofmotor synergies. According to our hypoth-esis, selection provides the superveningprinciple that transforms a complex andheterogeneous system into one that is charac-terized by the coordinated action of its parts.
DiscussionExperimental evidence, theoretical con-
siderations, and modeling studies suggestthat somatic selection plays a key role inhigher brain function and in the develop-
100
MM0)uu(A
0)
u
I
3000 6000 9000 12000
Cycle NumberFIG. 7.—Training curves of oculomotor and reaching systems for a simultaneous simulation of a
moving eye and two-jointed arm. Because of simultaneous training, visual signals used in the reachingsystem change constantly as the eye moves. As compared to simulations without simultaneous eyemovements (data not shown), progress in training the arm is somewhat delayed, but eventually ahigh performance level is reached and maintained. This shows that the selective training paradigm isindependent of the details of the simulation and the way sensory inputs are provided to the neuronalrepertoires. Absolute performance limits are 100% for the oculomotor and about 80% for the reachingsystem. From Reeke, Sporns, and Edelman (1990), reproduced with permission.
978 Child Development
ment of coordinated movement. In this arti-cle, we have reconsidered Bernstein's prob-lem within the framework of a selectionaltheory of the brain. We have argued thatan appropriate theory of selection appearscloser to the biological data than a purelycomputational treatment ofthe inverse kine-matics problem. Coordinated movement ispossible without explicit computations ofdesired movement paths. A selectional the-ory also accounts for synergies as functionalunits of the motor ensemble and as activecomponents in sensorimotor development.Synergies emerge from selective events ascoordinative structures (Kelso & Tuller,1984; Thelen, 1988; Thelen, Kelso & Fogel,1987; Ulrich, 1989) that are able to reducethe numbers of degrees of freedom in a com-plex multicomponent motor system. Themain advantage of a selectional theory is thatit accounts for the instantaneous adaptabilityof the motor ensemble and its associatedneural circuitry in response to biomechani-cal and environmental changes. This adapt-ability is not conferred upon the system byinstructive procedures imposed from with-out. Instead, adaptation results from the dy-namic interplay ofthe motor ensemble withthe environment constrained by internalvalue systems. Such a view readily accountsfor individual developmental change.
As we stated earlier, for selection to beeffective in a motor system, three require-ments must be met; (a) There must be a pri-mary movement repertoire of sufficient rich-ness and variability, (b) Movements musthave differential effects in the environmentwhich can be sensed by their originator, (c)Mechanisms must exist that allow sensoryinputs reporting the consequences of move-ments to act differentially within the ner-vous system, such that (on average) thosemovements that simultaneously satisfy envi-ronmental constraints and evolved internalvalue constraints will become more likelythan others. All of these requirements areeasily met by actual biological motor sys-tems. In contrast, there is little direct evi-dence that the nervous system precomputesdesired trajectories, computes comparisonsbetween actual movements and desiredones, or uses explicit error signals to adjustindividual components of the motor controlsystem and minimize future error.
Finally, several points that have notbeen considered in detail deserve at leastsome brief emphasis. The intricate interplaybetween sensory inputs and motor action inthe formation of gestural components and
synergies highlights only one aspect of thedeep interrelation between action and per-ception. Motion is fundamental for percep-tual categorization, one of the most basic ofneural and cognitive processes (Edelman,1987). For example, motion is crucial in thedetection of spatially correlated object fea-tures, and it naturally ensures the continuityof successive sensory inputs referring to asingle object. Body and eye movements areessential for the development of fundamen-tal visual cognitive concepts such as theunity, boundedness, and persistence of ob-jects (Spelke, 1990), and the constructionduring development of spatial maps ofthe environment. The importance of self-generated movements in such a variety offunctions is recognized by the theory of neu-ronal group selection. Coordinated move-ment is taken to be not just peripheral out-put, but an active ingredient in perceptualand cognitive processes, both during devel-opment and in adult function.
References
Arshavsky, Y. I., Gelfand, I. M., & Orlovsky, G. N.(1986). Cerebellum and rhythmical move-ments. Berlin; Springer.
Bauer, J., & Held, R. (1975). Comparison of visu-ally guided reaching in normal and deprivedinfant monkeys. Journal of ExperimentalPsychology; Animal Behavior Processes, 1,298-308.
Beach, F. A., Hebb, D. O., Morgan, C. T., &: Ni.s-.sen, H. W. (1960). The neuropsychology ojLashley. New York; McGraw-Hill.
Berkinblit, M. V., Gelfand, I. M., & Feldman,A. G. (1986). Model ofthe control ofthe move-ments of a multijoint limb. Biophysics, 31,142-153.
Bernstein, N. A. (1967). The coordination and reg-ulation of movements. Oxford; Pergamon.
Bradley, N. S., & Bekoff, A. (1990). Developmentof coordinated movement in chicks; I. Tem-poral analysis of hindlimb muscle synergiesat embryonic days 9 and 10. DevelopmentalPsychobiology, 23, 763-782.
Buchanan, T. S., Almdale, D. P. J., Lewis, J. L., &Rymer, W. Z. (1986). Gharacteristics of syner-gic relations during isometric contractions ofhuman elbow muscles./ourna/ ofNeurop/ujs-iology, 56, 1225-1241.
Bullock, D., & Grossberg, S. (1988). Neural dy-namics of planned arm movements; Emer-gent invariants and speed-accuracy propertiesduring trajectory formation. Psychological Re-view, 95, 49-90.
Goghill, G. E. (1929). Anatomy and the problem
Sporns and Edelman 979
of behavior. Cambridge; Cambridge Univer-sity Press.
Cratty, B. J. (1979). Perceptual and motor devel-opment in infants and children. EnglewoodCliffs, NJ; Prentice-Hall.
Edelman, G. M. (1978). Group selection and pha-sic re-entrant signalling; A theory of higherbrain function. In G. M. Edelman & V. B.Mountcastle (Eds.), The mindful brain (pp.51-100), Cambridge, MA; MIT Press.
Edelman, G. M. (1987). Neural Darwinism. NewYork; Basic.
Edelman, G. M. (1989). The remembered present.New York; Basic.
Edelman, G. M. (1993). Neural Darwinism; Selec-tion and reentrant signaling in higher brainfunction. Neuron, 10, 115-125.
Fetters. L., & Todd, J. (1987). Quantitative assess-ment of infant reaching movements. Journalof Motor Behavior, 19, 147-166.
Ganor, I., & Golani, I. (1980). Coordination andintegration in the hindleg step cycle of therat; Kinematic synergies. Brain Research,195, 57-67.
Gelfand, I. M., Gurfinkel, V. S., Tsetlin, M. L.,& Shik, M. L. (1971). Some problems in theanalysis of movements. In I. M. Gelfand, V. S.Gurfinkel, S. V. Eomin, & M. L. Tsetlin(Eds.), Models of the structural-functionalorganization of certain biological systems(pp. 329-345), Gambridge, MA; MIT Press.
Gelfand, I. M., Gurflnkel, V. S., Tsetlin, M. L., &Shik, M. L. (1973). Certain problems in theinvestigation of movement. In M. L. Tsetlin(Ed.), Automata theory and modeling of bio-logical systems. New York; Academic Press.
Georgopoulos A. P., Kalaska, J. F., & Massey, J. T.(1981). Spatial trajectories and reaction timeof aimed movements; Effects of practice, un-certainty, and change in target location. Jo«r-riat of Neurophysiology, 46, 725-743.
Gray, C. M., & Singer, W. (1989). Stimulus-specific neuronal oscillations in orientationcolumns of cat visual cortex. Proceedings ofthe National Academy of Sciences (USA), 86,1698-1702.
Greene, P. H. (1982). Why is it easy to controlyour arms? Journal of Motor Behavior, 14,260-286.
Grossberg, S., & Kuperstein, M. (1986). Neural dy-namics of adaptive sensory-motor control.Amsterdam; New Holland.
Hamburger, V. (1970). Embryonic motility in ver-tebrates. In F. O. Schmitt (Ed.), The neurosci-ences; Second study program (pp. 141—151).New York; Rockefeller University Press.
Hebb, D. O. (1949). The organization of behavior;A neuropsychological theory. New York;Wiley.
Held, R., & Bauer, J. A., Jr. (1974). Development
of sensorially-guided reaching in infant mon-keys. Brain Research, 71, 265-271.
Hinton, G. (1984). Parallel computations for con-trolling an arm. Journal of Motor Behavior,16, 171-194.
Hogan, N. (1988). Planning and execution ofmultijoint movements. Canadian Journal ofPhysiology and Pharmacology, 66, 508-517.
Hollerbach, J.M. (1980). A recursive Lagrangianformulation of manipulator dynamics and acomparative study of dynamics formulationcomplexit>'. IEEE Transactions Systems Manand Cybernetics, SMC-10, 730-736.
Hollerbach, J. M. (1982). Computers, brains andthe control of movement. Trends in Neurosci-ences, 5, 189-192.
Hoist, E. von (1937). Vom Wesen der Ordnung imZentralnervensystem. Naturwissenschaften,25, 625-631, 641-647.
Humphrey, D. R. (1986). Representation of move-ments and muscles within the primate pre-central motor cortex; Historical and currentperspectives. Federation Proceedings, 45,2687-2699.
Kawato, M., Furukawa, K., & Suzuki, R. (1987). Ahierarchical neural-network model for controland learning of voluntary movement. Biologi-cal Cybernetics, 57, 169-185.
Kawato, M., Isobe, M., Maeda, Y., & Suzuki, R.(1988). Coordinates transformation and lean-ing control for visually-guided voluntarymovement with iteration; A Newton-likemethod in a function space. Biological Cyber-netics, 59, 161-177.
Kelso, J. A. S., & Tuller, B. (1984). A dynamicalbasis for action systems. In M. S. Gazzaniga(Ed.), Handbook of cognitive neuroscience(pp. 321-3.56). New York and London;Plenum.
Kots, Y. M., & Syrovegin, A. V. (1966). Fixed setof variants of interaction ofthe muscles of twojoints used in the execution of simple volun-tary movements. Biophysics, 11, 1212-1219.
Kugler, P. N., Kelso, J. A. S., & Turvey, M. T.(1982). On the control and coordination of nat-urally developing systems. Im J. A. S. Kelso &J. E. Clark (Eds.), The development of controland coordination (pp. 5—78), New York;Wiley.
Kugler, P. N., & Turvey, M. T. (1987). Informa-tion, natural law, and the self-assembly ofrhythmic movement. Hillsdale, NJ; Erlbaum.
Kuperstein, M. (1988). Neural model of adaptivehand-eye coordination for single postures.Science,239, 1308-1311.
Laquaniti, F., Soechting, J. F., & Terzuolo, S. A.(1986). Path constraints on point-to-point arm
980 Child Development
movements in three-dimensional space. Neu-roscience, 17, 313-324.
Lashley, K. S. (1933). Integrative function ofthecerebral cortex. Physiological Reviews, 13,1-42.
Lashley, K. S. (1942). The problem of cerebral or-ganization in vision. In H. Kluever (Ed.), Bio-logical symposia; Vol. 7, Visual mechanisms(pp. 301-322). Lancaster; Cattell.
Lee, W. A. (1984). Neuromotor synergies as a basisfor coordinated intentional action. Journal ofMotor Behavior, 16, 135-170.
Lemon, R. (1988). The output map ofthe primatemotor cortex. Trends in Neurosciences, 11,501-506.
Loeb, G. E. (1983). Finding common ground be-tween robotics and physiology. Trends inNeurosciences, 6, 203—204.
Luh, J. Y. S., Walker, M. W., & Paul, R. P. C.(1980). On-line computational scheme for me-chanical manipulators. Journal of DynamicSystems, Measurement, and Control, 102,69-76.
Massone, L., & Bizzi, E. (1989). A neural networkmodel for limb trajectory formation. Biologi-cal Cybernetics, 61, 417-425.
Mayr, E. (1959). Darwin and the evolutionary the-ory in biology. In B. J. Meggers (Ed.), Evo-lution and anthropology; A centennialappraisal (pp. 1-10), Washington, DC; An-thropological Society of Washington.
Nashner, L. M., & McColIum, G. (1985). The orga-nization of human postural movements; A for-mal basis and experimental synthesis. Behav-ioral and Brain Sciences, 8, 135—172.
Partridge, L. D. (1986). Frogs solve Bernstein'sproblem. Behavioral and Brain Sciences, 9,619-620.
Pribram, K. H., Ed. (1969). Perception and action:Selected readings. Harmondsworth; Penguin.
Purves, D. (1988). Body and brain; A trophic the-ory of neural connections. Cambridge, MA;Harvard University Press.
Raibert, M. H. (1978). A model for sensorimotorcontrol and learning. Biological Cybernetics,29, 29-36.
Raibert, M. H., & Horn, B. K. P. (1978). Manipula-tor control using the configuration spacemethod. Industrial Robot, 5, 69—73.
Reeke, G. N., Jr., Finkel, L. H., Sporns, O., &Edelman, G. M. (1990). Synthetic neuralmodelling; A multi-level approach to braincomplexity. In G. M. Edelman, W. E. Gall, &W. M. Cowan (Ed.s.), Signal and sense; Localand global order in perceptual maps (pp.607-707). New York; Wiley.
Reeke, G. N., Jr., & Sporns, O. (1990). Selectionistmodels of perceptual and motor systems andimplications for functionalist theories of brainfunction. Physica D, 42, 347-364.
Reeke, G. N., Jr., Sporns, O., & Edelman, G. M.(1989). Synthetic neural modeling; Com-parisons of population and connectionistapproaches. In R. Pfeifer, Z. Schreter, F.Fogelman-Soulie, & L. Steels (Eds.), Connec-tionism in perspective (pp. 113-119). Amster-dam; Elsevier.
Reeke, G. N., Jr., Sporns, O., & Edelman, G. M.(1990). Synthetic neural modeling; The "Dar-win" series of recognition automata. Proceed-ings ofthe IEEE, 78, 1498-1530.
Ritter, H. J., Martinetz, T. M., & Schulten, K. J.(1989). Topology-conserving maps for learn-ing visuo-motor-coordination. Neural Net-works, 2, 159-168.
Saltzman, E. (1978). Levels of sensorimotor repre-sentation. Journal of Mathematical Biology,20, 91-163.
Schmidt, R. C , Beek, P. J., Treffner, P. J., & Tur-vey, M. T. (1991). Dynamical substructure ofcoordinated rhythmic movements. Journal ofExperimental Psychology; Human Perceptionand Performance, 17, 635—651.
Schoner, G., & Kelso, J. A. S. (1988). Dynamicpattern generation in behavioral and neuralsystems. Science, 239, 1513-1520.
Shik, M. L., & Orlovsky, G. N. (1976). Neurophys-iology of locomotor automatism. Physiologi-cal Review, 56, 465-501.
Smotherman, W. P., & Robinson, S. R. (1988). Be-havior of the fetus. Caldwell, NJ; Telford.
Soechting, J. F., & Laquaniti, F. (1981). Invariantcharacteristics of a pointing movement inmAn. Journal of Neuroscience, 1, 710-720.
Soechting, J. F., & Laquaniti, F. (1989). An assess-ment of the existence of muscle synergiesduring load perturbations and intentionalmovements of the human arm. ExperimentalBrain Research, 74, 535-548.
Soechting, J. F., Laquaniti, F., & Terzuolo, C. A.(1986). Coordination of arm movements inthree-dimensional space. Sensorimotor map-ping during drawing movement. Neurosci-ence, 17,295-311.
Spelke, E. S. (1990). Origins of visual knowledge.In D. N. Osherson, S. M. Kosslyn, & J. M.HoOerbach (Eds.), Visual cognition and ac-tion (Vol. 2, pp. 99-127). Cambridge, MA;MIT Press.
Sporns, O. (1990). Synthetic neural modeling;Computer simulations of perceptual and mo-tor systems. Ph.D. thesis. Rockefeller Uni-versity.
Sporns, O., Tononi, G., & Edelman, G. M. (1993).Temporal correlations and integrative func-tions ofthe brain. Manuscript submitted forpublication.
Thelen, E. (1988). On the nature of developingmotor systems and the transition from prena-tal to postnatal life. In W. P. Smotherman &
Sporns and Edelman 981
S. R. Robinson (Eds.), Behavior of the fetus(pp. 207-224), Caldwell, NJ; Telford.
Thelen, E. (1989a). The (re)discovery of motor devel-opment; Leaming new things from an old field.Developmental Psychology, 25, 946—949.
Thelen, E. (1989b). Evolving and dissolving syn-ergies in the development of leg coordina-tion. In S. A. Wallace (Ed.), Perspectives onthe coordination of movement (pp. 259—281).Amsterdam; Elsevier.
Thelen, E., & Fisher, D. M. (1983). From sponta-neous to instrumental behavior; Kinematicanalysis of movement changes during veryearly learning. Child Development, 54,129-140.
Thelen, E., Kelso, J. A. S., & Fogel, A. (1987).Self-organizing systems and infant motor de-velopment. Developmental Review, 7, 39—65.
Thelen, E., Skala, K., & Kelso, J. A. S. (1987). Thedynamic nature of early coordination; Evi-dence from bilateral leg movements in younginfants. Developmental Psychology, 23, 179-186.
Tononi, G., Sporns, O., & Edelman, G. (1992a).The problem of neural integration; Inducedrhythms and short-term correlations. In E. Ba-sar & T. H. Bullock (Eds.), Induced rhythmsin the brain (pp. 367-395). Boston; Birk-hauser.
Tononi, G., Sporns, O., & Edelman, G. (1992b).Reentry and the problem of integrating multi-ple cortical areas; Simulation of dynamic inte-gration in the visual system. Cerebral Cortex,2, 310-335.
Trevarthen, C. (1974). The psychobiology ofspeech development. Neurosciences Re-search Program Bulletin, 12, 570-585.
Turvey, M. T. (1977). Preliminaries to a theory ofaction with reference to vision. In R. Shaw& J. Bransford (Eds.), Perceiving, acting, andknowing; Toward an ecological psychology(pp. 211-265). Hillsdale, NJ; Erlbaum.
Turvey, M. T. (1990). Coordination. AmericanPsychologist, 45, 938-953.
Ulrich, B. D. (1989). Development of steppingpatterns in human infants; A dynamical sys-tems perspective./ournaZ of Motor Behavior,21, 392-408.
Viviani, P., & McCollum, G. (1983). The relationbetween linear extent and velocity in drawingmovements. Neuroscience, 10, 211-218.
von Hofsten, C. (1982). Eye-hand coordination inthe newborn. Developmental Psychology, 18,450-461.
Whiting, H. T. A., Ed. (1984). Human motoractions. Bernstein reassessed. Amsterdam;North-Holland.














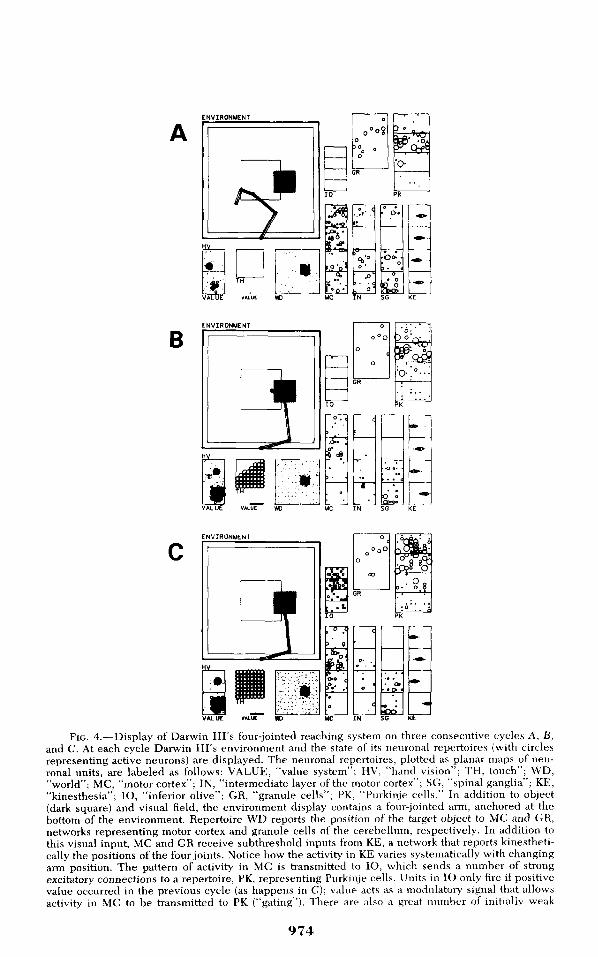

























![Bernstein's Theorem · minimal k-cones in Rk+l would imply Bernstein's theorem for minimal graphs in Rk+2. In 1966, Almgren [A] proved that there exist no singular cones in R4. These](https://static.documents.pub/doc/80x56/5f13a577587f4a74b061b3b5/bernsteins-theorem-minimal-k-cones-in-rkl-would-imply-bernsteins-theorem-for.jpg)