Abstract: To study the thermostability of Nattokinase (subtilisin NAT, NK), three double mutant plasmids (pET-28a-NKG61C/S98C, pET-28a-NKT22C/S87C, pET-28a-NKS24C/S87C) were constructed by site-directed mutagenesis. Target enzymes were detected using SDS-PAGE and disulfide bond formation was detected using Western blotting analysis. Thermostability was tested by rates of inactivation at certain temperature. The results showed that disul-fide bond was not formed within two cysteines and the thermosta-bility of three double mutants was not increased compared with the wild-type NK. The thermostability of NK performed in Ca2+ was stronger than in ethylenediaminetetraacetic acid (EDTA). But when the temperature reached 62 ℃, the enzymes rapidly dena-tured and inactivated even in the presence of Ca2+. Although the thermostability of mutants was not increased, this study shows a tendency of improving thermostability of NK in protein engineer-ing. Key words: nattokinase; subtilisin; thermostability CLC number: Q 816 Received date:2013-11-28 Foundation item: Supported by the National Natural Science Foundation of China (30670464, 20873092, 30800190) and Science and Technology Project of Wuhan (200960323115) Biography: WENG Meizhi, female, Competent technician, Ph. D., research direction: biochemistry and molecular biology. E-mail: [email protected] † To whom correspondence should be addressed. E-mail: [email protected]. cn
0 Introduction
Nattokinase (NK, formerly designated subtilisin NAT), a cysteine-free alkaline serine protease, is a potent fibri-nolytic enzyme produced from Bacillus subtilis var. natto and was first found in the traditional Japanese soy-bean food natto[1]. NK is encoded by the aprN gene and synthesized in a precursor form, in which a signal pep-tide and a propeptide are attached to the N-terminus of the mature domain, with a molecular mass of 27.7×103 [2]. As a member of the subtilisin family of serine protease, NK has the same conserved catalytic triad (Asp32, His64, and Ser221) [2, 3]. gene sequence of NK is homologous to other members of the subtilisin family, such as subtilisin Carlsberg, subtilisin BPN′ , subtilisin E and subtilisin amylosacchariticus [1]. Recently, the crystal structure of NK at 0.174 nm resolution was determined by Yanagi-sawa and the report showed that three-dimensional structures of NK is almost identical with subtilisin E from Bacillus subtilis DB104 [4].
NK has been extensively studied not only for the insight into the mechanism of enzyme catalysis, but also for its commercial applications. NK has strong fi-brinolytic activity and may be useful for improving circulation and reducing the risk of thrombosis [1, 5]. NK not only degrades fibrin in thrombi but also cleaves plasminogen activator inhibitor type 1[6]. Previous studies have shown that oral administration of NK pro-duced a gradual enhancement of fibrinolytic activity in the plasma [7]. In addition, oral administration of NK could be considered as a cardiovascular disease (CVD)
Wuhan University Journal of Natural Sciences 2014, Vol.19 No.3 230
nutraceutical by decreasing plasma levels of fibrinogen, factor Ⅶ, and factor Ⅷ [8]. All these therapeutic ef-fects make it a potential oral drug for the treatment of cardiovascular disease.
For broadening the proteins’ commercial and medical applications, protein engineering has been used to enhance the stability of enzymes and the en-hancement of thermostability is one of the most im-portant aims. Several strategies have been undertaken for this purpose, such as introduction of Cys residue(s) or disdulfide bond(s) into protein molecules by site- directed mutagenesis [9-11]. We previously used the method of molecular dynamic simulation (MD) and constructed a 3D model and probed the significance of hydrogen bonds in the catalytic center of NK with the method of site-directed mutagenesis [12, 13]. Moreover, we have obtained two mutants whose oxidative stabil-ity were greatly increased compared with the wild- type NK [14] and we also obtained a NK whose fibri-nolytic activity was increased [15].
Here, we study the effect of Cys residue(s) or di-sulfide bond(s) on the thermostability of NK by site- directed mutagenesis. Two cysteines were introduced in the cysteine-free NK molecule and three double mutants (G61C/S98C, T22C/S87C and S24C/S87C) were obtained. The thermostabilities of enzymes were determined by their rates of thermal inactivation. This study should be meaningful for broadening the appli-cation of NK for commercial and medical purposes.
1 Materials and Methods
1.1 Materials Escherichia coli BL21 (DE3) and the pET- 28a
vector were purchased from Novagen. The substrate suc-AAPF-pNA was from Sigma (St. Louis, MO, U.S.A). All enzymes for DNA manipulations were purchased from Takara Shuzo Co. Ltd., and the reaction conditions were recommended by the supplier. Ni-NTA resin was purchased from Invitrogen and DEAE Sepharose Fast Flow was purchased from Pharmacia Biotech (Uppsala, Sweden). The BCA protein assay reagent kit was pur-chased from Pierce. Mouse Anti-His-tag Monoclonal Antibody and HRP-Goat anti-mouse IgG (H+L) were purchased from GenScript USA Inc. (Nanjing, China). Coomassie Brilliant Blue G-250 was from Fluka. All other chemicals and reagents were of the highest quality and obtained from standard commercial sources. 1.2 Construction of Plasmids
The plasmid pET-28a-aprN [14], which carries the NK gene (aprN), was used as the template for the construction of double-mutant plasmids (pET-28a-NKG61C/S98C, pET-28a- NKT22C/S87C, pET-28a-NKS24C/S87C) by means of site-di-rected mutagenesis. Site-directed mutagenesis was per-formed by the overlap extension method [16]. Oligonu-cleotide sequences used for site-directed mutagenesis were shown in Table 1. The sequences of all constructs were verified by DNA sequencing.
Table 1 Synthetic oligonucleotides used for site-directed mutagenesis
Gene type Oligonucleotide sequences
Wild-type Forward: 5′-CGCTGAATTCGTGAGAAGCAAAAAATTGTGG-3′, EcoR I site
Reverse: 5′-CTGGAGCTCTT TTGTGCAGCTGCTTGTACGTT-3′, Sac I site
Mutants
G61C 5′-AACCCATACCAGGACTGCAGTTCTCACGGTACG-3′
S98C 5′-CAGTAAAAGTGCTTGATTGTACAGGAAGCGGCCAA-3′
S87C 5′-CTGGGCGTAGCGCCATGCGCATCATTATATGCAG-3′
T22C 5′-CACTCTCAAGGCTACTGTGGCTCTAACGTAAAAG-3′
S24C 5′-CAAGGCTACACAGGCTGTAACGTAAAAGTAGCTG-3′ For mutants, only the forward mutation site oligonucleotides were shown and base changes from the encode gene of the wild-type NK were underlined
1.3 Expression of aprN Gene and Purification of Wild-type NK and Mutants
The expression of aprN gene and purification of wild-type NK and mutants were performed as previ-ously described[14]. The purity of enzymes was checked using SDS-PAGE and disulfide bond was confirmed
using Western blotting analysis. SDS-PAGE was per-formed on a 15% (m︰V) polyacrylamide separating gel with a 5% (m︰V) stacking layer according to the pro-cedure of Laemmli[17], and gel was then stained with Coomassie Brilliant Blue G-250. Western blotting analysis of purified proteins was taken as previously
WENG Meizhi et al : Thermostability of Subtilisin Nattokinase Obtained by …
231
described[14]. After purifying to homogeneity, the con-centration of proteins was determined by the BCA pro-tein assay reagent kit. 1.4 Determination of Thermal Inactivation
The thermostabilities of enzymes were tested by their rates of inactivation at indicated temperatures (50 ℃, 55 and 62 ) in the presence of 1 mmol/L ℃ ℃
EDTA or CaCl2· Remaining activity after heating was determined against suc-AAPF-pNA in 100 mmol/L Tris/HCl (pH 8.60), 4% (V︰V) DMSO equilibrated at (25±0.2) ℃ and was expressed as the percentage of a nontreated enzyme control. The purified enzymes were either treated or not treated with 10 mmol/L dithiothreitol (DTT) at room temperature for 30 min. Each reaction includes 0.7 μmol/L enzyme and the samples were incu-bated in a thermostatic circulating water bath equili-brated at the indicated temperature. At the designated time intervals (0, 5, 10, 15, 20, 25, 30 min), 60 μL ali-quot was removed and quickly transferred to a 25 ℃
water bath. Then, a 50 μL aliquot was removed and re-sidual activity was assayed against suc-AAPF-pNA. Re-sidual activity is expressed as a percentage of a nontreated enzyme control.
2 Results and Discussion
2.1 Construction of Recombinant Plasmids, Expression of aprN Gene and Purification of Wild-type NK and Mutants
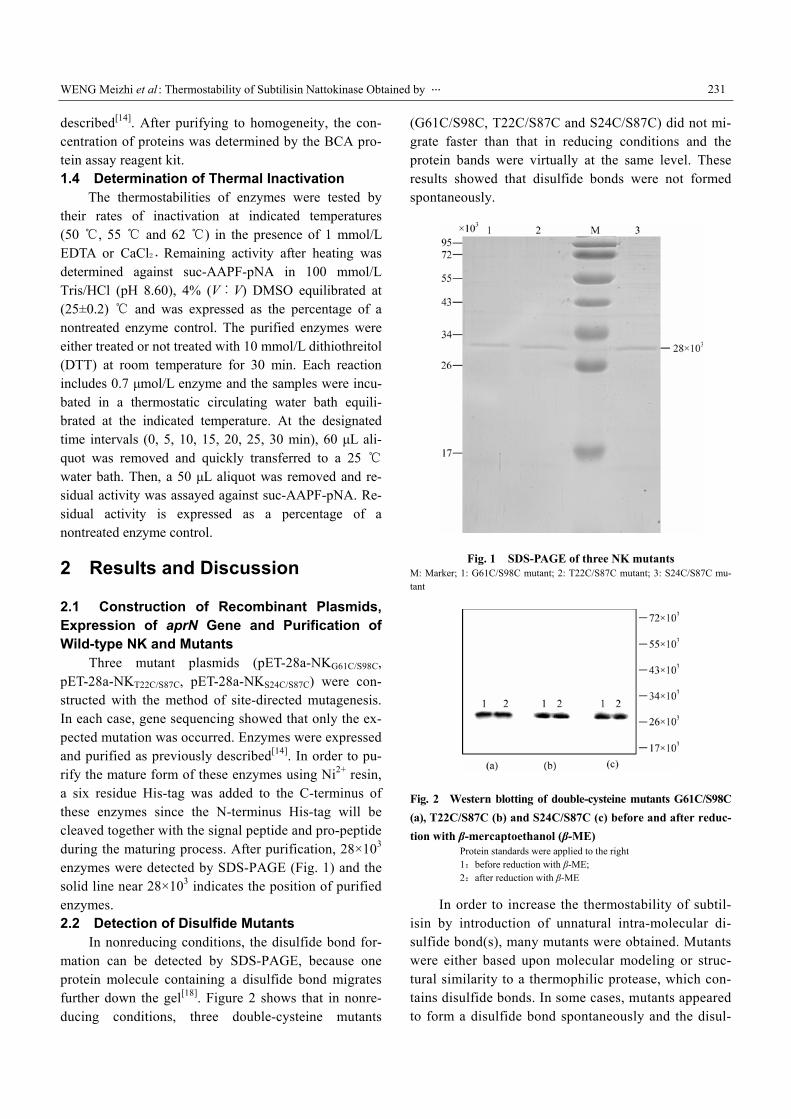
Three mutant plasmids (pET-28a-NKG61C/S98C, pET-28a-NKT22C/S87C, pET-28a-NKS24C/S87C) were con-structed with the method of site-directed mutagenesis. In each case, gene sequencing showed that only the ex-pected mutation was occurred. Enzymes were expressed and purified as previously described[14]. In order to pu-rify the mature form of these enzymes using Ni2+ resin, a six residue His-tag was added to the C-terminus of these enzymes since the N-terminus His-tag will be cleaved together with the signal peptide and pro-peptide during the maturing process. After purification, 28×103 enzymes were detected by SDS-PAGE (Fig. 1) and the solid line near 28×103 indicates the position of purified enzymes. 2.2 Detection of Disulfide Mutants
In nonreducing conditions, the disulfide bond for-mation can be detected by SDS-PAGE, because one protein molecule containing a disulfide bond migrates further down the gel[18]. Figure 2 shows that in nonre-ducing conditions, three double-cysteine mutants
(G61C/S98C, T22C/S87C and S24C/S87C) did not mi-grate faster than that in reducing conditions and the protein bands were virtually at the same level. These results showed that disulfide bonds were not formed spontaneously.
Fig. 1 SDS-PAGE of three NK mutants M: Marker; 1: G61C/S98C mutant; 2: T22C/S87C mutant; 3: S24C/S87C mu-tant
Fig. 2 Western blotting of double-cysteine mutants G61C/S98C
(a), T22C/S87C (b) and S24C/S87C (c) before and after reduc-
tion with β-mercaptoethanol (β-ME) Protein standards were applied to the right 1:before reduction with β-ME; 2:after reduction with β-ME
In order to increase the thermostability of subtil-isin by introduction of unnatural intra-molecular di-sulfide bond(s), many mutants were obtained. Mutants were either based upon molecular modeling or struc-tural similarity to a thermophilic protease, which con-tains disulfide bonds. In some cases, mutants appeared to form a disulfide bond spontaneously and the disul-
Wuhan University Journal of Natural Sciences 2014, Vol.19 No.3 232
fide bond enhanced the thermostability[19]. However, the thermostabilities of some mutants were not dra-matically improved by introducing a designed disul-fide bridge[9] and no disulfide bond was detected in some mutants[11], indicating that finding the proper amino acid sites of enzymes to form disulfide bond is not a clear-cut task. 2.3 Thermal Stability of NK Mutants
Thermal stability is an important property for en-zymes used in industrial processes. To test the autolytic stability of enzymes, the rates of thermal inactivation were measured at indicated temperatures in the presence of Ca2+ or EDTA. The half-life values (t1/2) were deter-mined by measuring catalytic activity after incubation at indicated temperatures for various time periods and it was the average of at least two separated determinations. For G61C/S98C mutant, the stability was nearly equal to the wild-type enzyme at 50 ℃ in presence of 1 mmol/L
EDTA, and the half-life (t1/2) was 3 min. At 55 ℃, the rate of thermal inactivation of the G61C/S98C mutant in pres-ence of 1 mmol/L Ca2+ was found to be 5-6 times longer than that in presence of 1 mmol/L EDTA. However, at 62 ℃, the rates of thermal inactivation were significantly low even in presence of 1 mmol/L Ca2+ and the half-life (t1/2) was 3 min for both the G61C/S98C mutant and wild-type NK (Fig.3). For T22C/S87C and S24C/S87C mutants, the stability was nearly equal to the wild-type NK in the presence of 1 mmol/L EDTA at 55 ℃ and the half-life (t1/2) was 3 min (Fig. 4). When the enzymes were treated with 10 mmol/L DTT for reduction, the mutants (G61C/S98C, T22C/S87C and S24C/S87C) had a half-life nearly equivalent to that of the wild-type NK and the half-life (t1/2) was 3 min. Moreover, the stability of the three mutants was almost the same regardless of whether or not they were pretreated with 10 mmol/L DTT for reduction.
Fig. 3 Kinetics of thermal inactivation of wild-type NK and G61C/S98C mutant at different temperature (a) 50 ℃; (b) 55 ℃; (c) 62 ℃
Values are the average of three independent experiments with standard deviation
Fig. 4 Kinetics of thermal inactivation of wild-type NK, T22C/S87C and S24C/S87C mutants treated without DTT (a)or with DTT (b) at 55 ℃
Values are the average of three independent experiments with standard deviation
Introduction of Cys residue(s) or disulfide bond(s) has been demonstrated to increase the thermostability of enzymes by previous studies. Some mutants increased enzyme thermostability [11, 20], while some decreased [9].
In addition, the thermostability of subtilisin E was en-hanced by single-Cys mutant, replacing Serl61 with Cys161 [10]. The data presented here seemed that neither of the mutants (G61C/S98C, T22C/S87C and S24C/S87C)
WENG Meizhi et al : Thermostability of Subtilisin Nattokinase Obtained by …
233
increased the thermal stability. The reason for such a result reported here is not entirely clear. Sometimes this can be rationalized by the long distance between these two Cys residues.
The total kinetic thermal stability of a protein molecule can be separated into a contribution due to the binding of Ca2+ and an intrinsic contribution of the polypeptide chain[21]. Masilamani showed that calcium ion improved the thermal stability of β-1,3-1,4-gluca-nase from Bacillus subtilis SU40 [22] and a low concen-tration of CaCl2 increased the thermostability of Dextran glucosidase from Streptococcus mutans (SmDG) [23]. In addition, Ca2+ can be as a stabilizer to enhanced pro-duction of thermostable MBP-fused heparinase I [24]. The results here showed that Ca2+ also increased the thermal stability of NK. The enhanced stability by Ca2+
was probably related to calcium binding sites in NK. However, the thermostability of G61C/S98C mutant was lower than that of the wild-type NK in the presence of Ca2+ at 55 ℃. The reason for this sometimes can be rationalized by the deleterious effects of the parent mu-tations.
3 Conclusion
In order to increase the thermostability of NK, three double-cysteine mutant plasmids (pET-28a- NKG61C/S98C, pET-28a-NKT22C/S87C, pET-28a-NKS24C/S87C) were constructed by site-directed mutagenesis and target enzymes were obtained. Although mutants re-ported here did not increase the thermostability, the work demonstrated that it may be possible to find thermostability amino acid substitutions in NK. In ad-dition, the data presented here showed that the ther-mostability of NK performed in presence of Ca2+ was 5-6 times longer than that in presence of EDTA at the same temperature. But when the temperature reached 62 , the enzymes rapidly denatured and ina℃ ctivated even in presence of Ca2+. Increasing thermostability has significant benefit in stabilizing enzymes used in industrial processes and engineering the stronger thermostability substitutions is worth further studying and is currently in progress.
[1] Sumi H, Hamada H, Tsushima H, et al. A novel fibrinolytic
enzyme (nattokinase) in the vegetable cheese Natto; A typi-
cal and popular soybean food in the Japanese diet [J]. Ex-
perientia, 1987, 43(10): 1110-1111.
[2] Nakamura T, Yamagata Y, Ichishima E. Nucleotide sequence of
the subtilisin NAT gene, aprN, of Bacillus subtilis (natto) [J].
Bioscience, Biotechnology, and Biochemistry, 1992, 56 (11):
1869-1871.
[3] Carter P, Wells J A. Dissecting the catalytic triad of a serine
protease [J]. Nature, 1988, 332(6164): 564-568.
[4] Yanagisawa Y, Chatake T, Naito S, et al. X-ray structure
determination and deuteration of nattokinase [J]. Journal of Synchrotron Radiation, 2013, 20 (6): 875-879.
[5] Tai M W, Sweet B V. Nattokinase for prevention of throm-
bosis [J]. American Journal of Health-System Pharmacy,
2006, 63(12): 1121-1123.
[6] Urano T, Ihara H, Umemura K, et al. The profibrinolytic
enzyme subtilisin NAT purified from bacillus subtilis
cleaves and inactivates plasminogen activator inhibitor type
1 [J]. Journal of Biological Chemistry, 2001, 276(27):
24690- 24696.
[7] Sumi H, Hamada H, Nakanishi K, et al. Enhancement of the
fibrinolytic activity in plasma by oral administration of nat-