Page 1
Title 骨細胞の力学刺激感知およびカルシウム応答伝播のバイオメカニクス( Dissertation_全文 )
Author(s) 青沼, 有紀
Citation Kyoto University (京都大学)
Issue Date 2010-03-23
URL https://doi.org/10.14989/doctor.k15320
Right
Type Thesis or Dissertation
Textversion author
Kyoto University
Page 2
骨細胞の力学刺激感知およびカルシウム応答伝播の
バイオメカニクス
青沼 有紀
Page 3
目 次
第 1 章 緒 論 1
1.1 はじめに 1
1.2 骨細胞のバイオメカニクス 2
1.2.1 骨リモデリングと力学的適応 3
1.2.2 骨系細胞による骨組織の代謝活動 4
1.2.3 骨細胞および骨リモデリングにおける骨細胞の役割 6
1.2.4 骨細胞の力学刺激感知特性 8
1.3 本論文の目的 11
1.3.1 目 的 11
1.3.2 医工学的意義 11
1.3.3 本論文の構成 12
第 2 章 骨組織の変形にともなう骨細胞のカルシウム応答 15
2.1 緒 言 15
2.2 実験方法 16
2.2.1 骨基質試料の作製 16
2.2.2 組織内骨細胞の蛍光標識 17
2.2.3 細胞内カルシウムイオンの可視化 18
2.2.4 骨基質試料への変形付与 19
2.2.5 骨基質試料および組織内骨細胞の変形解析 21
2.2.6 組織内骨細胞のカルシウム応答の経時観察 22
i
Page 4
2.3 実験結果 23
2.3.1 断層撮影画像の投影と変形解析領域の設定 23
2.3.2 変形付与にともなう骨基質・骨細胞の変形解析 25
2.3.3 組織内骨細胞のカルシウム応答 26
2.4 考 察 28
2.4.1 組織内骨細胞のライブイメージング実験 28
2.4.2 骨基質の変形にともなう骨細胞の力学刺激応答 29
2.5 結 言 30
第 3 章 骨細胞の局所的構造と力学刺激感知特性 31
3.1 緒 言 31
3.2 実験方法 32
3.2.1 骨細胞の単離 32
3.2.2 マイクロパーティクルの修飾および骨細胞への接着 33
3.2.3 力学刺激の付与 34
3.2.4 細胞内カルシウムイオンの可視化とカルシウム応答の観察 36
3.3 実験結果 37
3.3.1 マイクロパーティクルの骨細胞への接着 37
3.3.2 力学刺激の付与に対するカルシウム応答 39
3.4 考 察 44
3.4.1 マイクロパーティクルを介した局所的な力学刺激の付与 44
3.4.2 細胞内カルシウムイオンの経時変化とカルシウム応答発生メカニズム 48
3.5 結 言 49
ii
Page 5
iii
第 4 章 骨系細胞における細胞応答の細胞間伝播 51
4.1 緒 言 51
4.2 実験方法 52
4.2.1 骨系細胞の単離 52
4.2.2 骨細胞の同定 52
4.2.3 力学刺激の付与 55
4.2.4 近傍細胞の定義 55
4.2.5 細胞内カルシウムイオンの可視化とカルシウム応答の観察 56
4.3 実験結果 56
4.3.1 カルシウム応答の細胞間伝播 56
4.3.2 細胞間伝播の発生率 57
4.4 考 察 59
4.4.1 細胞種と細胞間伝播発生の関係 59
4.4.2 骨組織中における細胞間コミュニケーション 60
4.5 結 言 62
第 5 章 結 論 63
参考文献 67
関連学術論文・学術講演 77
謝 辞 83
Page 6
第 1 章 緒 論
1.1 はじめに
バイオメカニクス (biomechanics) には,生体力学という訳語があてられる.すなわ
ち,生物の構造や,生物の生命活動における,細胞内の生化学反応から個体の行動に
至るまでの諸現象に対して,力学的な解釈を試みる学問分野であると考えることがで
きる.
生物の生命活動は,生体内外において常に生化学的,および,力学的な作用を受け
ている.生物を構成する組織は,これらの生体内外の働きかけに対して,内分泌や代
謝により常に自身を調節・更新させながら組織としての機能を果たすことにより,組
織としての恒常性 (homeostasis) を維持している.
生体組織の恒常性が,組織としての静的な定常状態ではなく,内分泌や代謝という
動的現象の平衡状態により維持されている背景には,生物と,生物を取り巻く環境
(environment) との相互作用が関連している.環境は,「生物の生存に関係する多種類
の外的条件のすべて」として定義される [荒木ら, 1985].すなわち,環境は,生物の
生命活動に対して生化学的,力学的な作用をもたらす空間的時間的総体として存在し,
生命活動の制約要因として機能している.生体組織の恒常性は,生物に絶えず働きか
け続ける環境の刻々の変化に対して常に適応し,組織としての機能を維持させるため
1
Page 7
の性質であると考えられる.
その一方で,生物は,環境の変化に対して主体的に適応し,環境を自らの生存に適
するように変えていくことができる.例えば,人間の場合,衣服の着脱や,住居や冷
暖房機器などを用いた住空間を調えることにより,気温の急激な変化による生命活動
や生活行動への影響を緩和させる.生物は,生物を取り巻く環境を意図的に形成する
ことにより,生物と環境との間に新たな相互作用をなす関係を構築できるのである.
バイオメカニクスの観点から生命現象を理解し,その知見を応用することは,換言
すれば,環境による生物への力学的な働きかけと,生物が環境を生命活動に適した力
学的諸条件を満たすよう改変する行為の相互作用のあり方を検討することである.こ
の学問的な探求と知見の応用の関係が,本質的な生物と環境の相互関係の構築に連関
する点で興味深い.力学的因子を介した生物と環境の相互関係の理解は,バイオメカ
ニクス分野における知見の応用だけでなく,環境科学の観点からの生命現象の理解の
一助にもなると考えられる.
本論文は,生体の支持構造組織である,骨組織の力学的環境に対する適応現象につ
いて,細胞レベルの応答現象の観点から検討したものである.骨は,その構造や機能
の維持において,生物と力学的環境の相互作用の影響を受ける代表的な器官である.
また,力学的負荷にさらされる硬組織としての特性はもちろん,環境に対する適応現
象において細胞レベルの生化学的過程が機能していることから,バイオメカニクスの
分野における重要な研究対象のひとつとなっている.
1.2 骨細胞のバイオメカニクス
骨は,常に組織の代謝を行いながら,骨格構造としての力学的機能を果たしている.
生体組織・器官が,組織としての恒常性を維持しながら動的に構造・機能を変化させ
る現象は,リモデリングと呼ばれる [林ら, 2003].リモデリングは,生体がさらされ
る環境に対する適応現象のひとつとして位置づけられるが,その中でも骨のリモデリ
ングは,重力や荷重などの力学的環境の影響を大きく受けることが知られている [須
田ら, 2007; 林ら, 2003].骨のリモデリングの過程は,骨に存在する骨系細胞の代謝活
2
Page 8
動により制御されている.骨系細胞のひとつである骨細胞は,骨への力学的負荷を刺
激として感知し,骨リモデリングを担う細胞群の活動を調節するメカノセンサーとし
ての役割を果たすと考えられている [Nijweide et al., 1996].これらの骨細胞の力学刺
激感知・伝達メカニズムの解明は,骨リモデリングの理解を深めるとともに,医療・
工学分野における重要な基礎的知見となり得る.
本節では,骨のリモデリング現象と力学的適応,および,骨系細胞による骨リモデ
リング現象の過程を概説する.次に,骨リモデリング現象における骨細胞の役割,お
よび,骨細胞の力学刺激感知特性に関する仮説について,これまでに得られている知
見を述べる.
1.2.1 骨リモデリングと力学的適応
骨リモデリング (bone remodeling) とは,既に形成されている骨組織の一部が吸収
され,新たに再構築される形態的変化のことである.脊椎動物の骨格構造を形成する
骨は,成長を終えた後も骨吸収 (bone resorption) と骨形成 (bone formation) を繰り返
し,常に新しい組織に更新されることにより,その形態や機能を動的に維持している.
骨リモデリングの生理機能として,ミネラルの調節が挙げられる.リン酸カルシウ
ムを主成分とする骨は,副甲状腺や腎臓と同様に,生体内のカルシウム濃度,リン濃
度の調節器官として機能している.成人では,1 日におよそ 300 mg のカルシウム,お
よび 350 mg のリンが,骨吸収により血液などの細胞外液に動員されるとともに,同
量が再石灰化により骨基質に蓄積される [Felig and Frohman, 2001; 須田ら, 2007].骨
リモデリングにともなう物質代謝は,これらのミネラルの出納を通じて,体液中のカ
ルシウム濃度,リン濃度の恒常性を維持している.
また,骨リモデリングは,周囲の力学的環境の変化に対する機能的適応現象として
も知られている.骨は,リモデリングにともなう構造変化により,適応的に身体を支
持する構造体としての機能を維持している.特に,重力や荷重などの生体がさらされ
る力学的環境との間には密接な関係があり,例えば,宇宙空間の微小重力環境下では,
骨への力学的負荷の減少にともない,骨からのカルシウム溶出と骨量減少が促進され
ることが知られている [大島・向井, 2008].微小重力環境に対する骨組織の代謝,な
らびに,カルシウム代謝の適応には長期間を要し,宇宙滞在中は半年以上にわたり適
3
Page 9
応にともなう骨量減少が継続することが報告されている [Nicogossian et al., 1994].こ
れらのことから,骨リモデリングにともなう骨の力学的適応 (mechanical adaptation)
は,骨が支持構造体として,およびミネラルの調節器官としての役割を果たすために
不可欠な機能であると言える.
力学的環境と,骨の構造および機能との関連は,力学的負荷が骨の特徴的な構造の
形成に寄与しているという Galileo Galilei (1564-1642) の見解に端を発し,骨が外部形
状や内部応力の変化に対して,外部形状や内部の骨梁構造を変化させるとする Wolff
の法則 (law of bone transformation, [Wolff, 1892]) によって定義された [Einhorn, 1996].
Wolff の法則は,提唱から 1 世紀以上が経過した現在もなお,骨リモデリングにとも
なう骨の力学的適応の議論において重要な概念のひとつとして位置づけられている.
1.2.2 骨系細胞による骨組織の代謝活動
骨リモデリングにおける骨組織の代謝回転 [林ら, 2003] について述べる.骨リモデ
リングは,エンベロープ (envelope) と呼ばれる骨組織の各表面において,多細胞単位
(basic multicellular unit: BMU) と呼ばれる骨系細胞群の集合単位ごとに行われる
[Parfitt, 1994; 須田ら, 2007].リモデリングの休止状態にあるエンベロープ面は,上層
細胞 (bone lining cells) によって覆われている.骨リモデリングは,この上層細胞に覆
われたエンベロープが活性化され,循環血中の単球マクロファージ (monocyte
macropharge) 系細胞から破骨細胞前駆細胞 (osteoclast progenitor cells) が供給される
ことに始まる.破骨細胞前駆細胞は,骨組織において単核の破骨細胞 (osteoclasts) に
分化した後,付着した骨表面上で複数の細胞が融合することにより多核の破骨細胞と
なり,骨基質の吸収を行う.骨吸収が終了すると,破骨細胞は骨表面から移動し,単
核マクロファージによる骨吸収が行われたのち [須田ら , 2007],間葉系幹細胞
(mesenchymal stem cells) から分化・増殖した骨芽細胞 (osteoblasts) による類骨
(osteoid) の形成が行われる.形成された類骨は,数日を経て石灰化し,骨芽細胞は石
灰化の終了後,一部は骨基質中に埋没して骨細胞 (osteocytes) に分化し,再びエンベ
ロープで休止状態 (上層細胞) となる (Fig. 1-1).
次に,骨リモデリングにおける一連の骨代謝現象を担う上層細胞,骨芽細胞,破骨
細胞の特徴について述べる [須田ら, 2007].
4
Page 10
Fig. 1-1 Schematic of activities of bone cells for bone metabolism on envelope during bone remodeling
([須田ら, 2007] より改変).
上層細胞:上層細胞は,リモデリング休止期に骨組織のエンベロープ上において,
骨表面と骨髄を区画する細胞である.骨リモデリングにともなう骨組織の代謝回転で
は,骨形成相の終期に骨芽細胞が不活性化することによりエンベロープ上に細胞層を
形成することが知られている.
破骨細胞:骨髄中の造血幹細胞に由来する破骨細胞は,骨組織を吸収することによ
り,骨芽細胞とともに骨リモデリングにおいて主要な役割を果たす細胞である.活性
化した破骨細胞は,骨基質への接着側に焦点接着構造を帯状に集積した明帯 (clear
zone) により骨組織を被覆するとともに,その内部に波状縁 (ruffled border) と呼ばれ
る骨吸収領域を形成する.明帯により区画された波状縁部位では,骨表面に対して能
動輸送によるプロトン (H+) と塩素イオン (Cl⁻) の放出が行われるとともに,カテプ
シン K などのタンパク質分解酵素が分泌され,骨基質のヒドロキシアパタイトの溶解
と骨基質タンパクの分解が行われる.
骨芽細胞:間葉系細胞に由来する骨芽細胞は,骨吸収相が終了した骨表面において
骨形成を担う細胞である.骨リモデリングの骨形成相において骨芽細胞が合成・分泌
する有機成分は,骨基質のコラーゲン線維を形成する I 型コラーゲンをはじめとして,
非コラーゲンタンパク質やプロテオグリカンなど,骨基質が含有する有機成分の大部
5
Page 11
分を占めている.これらの有機成分の多くは,カルシウムとの親和性を有し,類骨の
石灰化を調節すると考えられている.
骨リモデリングにおいて,破骨細胞と骨芽細胞の骨代謝活動は,相互に連関する共
役関係 (coupling) にあるとされている.骨芽細胞は,破骨細胞の分化誘導・増殖の必
須因子である M-CSF と RANKL を発現することから,破骨細胞の分化・誘導に重要
な役割を果たしている.一方,破骨細胞が骨吸収を行った場所に骨芽細胞が誘導され
る理由として,血管内皮細胞などの骨芽細胞の前駆細胞が,破骨細胞前駆細胞を供給
した血管系細胞から供給されるメカニズムが提案されており,破骨細胞のリモデリン
グ領域への誘導が骨芽細胞の誘導にも寄与すると考えられている [Parfitt, 2000].
1.2.3 骨細胞および骨リモデリングにおける骨細胞の役割
骨リモデリングにおいて,類骨を形成した骨芽細胞はリモデリングの休止後,大半
が上層細胞となって骨表面に留まるほか,軟骨組織への分化,アポトーシスなどの運
命をたどる [Franz-Odendaal et al., 2006].そのひとつが,リモデリングの過程におい
て形成された類骨中への埋没,および,骨細胞への分化である.骨細胞は,骨組織に
おいて も多く存在する細胞であり,その細胞数は,1 mm3 の皮質骨あたり 25,000 個
存在し,標準的なヒトの骨においては,骨芽細胞の約 10 倍になるとされている [Parfitt,
1994].
骨リモデリングのメカニズムにおいて,骨細胞は,骨への力学的負荷が骨系細胞の
骨代謝活動の制御に結びつく過程で重要な役割を果たすと考えられている.力学的環
境の変化が骨リモデリングの契機となることを考慮すると,骨リモデリングが行われ
る BMU における局所的な力学状態もまた,骨系細胞群の骨代謝活動の調節に寄与す
ると考えられる.骨系細胞の中で唯一,骨基質 (bone matrix) 中に埋没した形で存在
する骨細胞は,硬い骨基質の空隙をぬって多数の細胞突起 (cell processes) を伸ばし,
骨細胞同士,あるいは骨組織表面の骨芽細胞と連結した細胞間ネットワークを形成し
ている [Donahue, 2000; Kamioka et al., 2001] (Fig. 1-2).この骨基質内部のネットワーク
構造は,骨細胞が骨に対する力学的負荷を刺激として感知し,さらに細胞間の刺激情
報伝達に適していることから,骨細胞は,力学的適応にともなう骨代謝現象において,
メカノセンサーとして機能すると考えられている [Burger and Klein-Nulend, 1999].
6
Page 12
Fig. 1-2 Osteocyte in bone matrix (scanning electron micrograph) [Bonewald, 2005].
骨細胞と他の骨系細胞との関係については,骨細胞を選択的に除去した骨組織にお
いて破骨細胞の誘導が起こらず力学的適応性が失われること [Tatsumi et al., 2007],培
養海綿骨試料に対する力学負荷試験により,骨細胞の存在下で骨芽細胞の類骨形成が
促進されること [Takai et al., 2004],骨の変形や損傷による骨細胞死が,骨髄細胞
(bone-marrow cells) からの破骨細胞の分化を促進させること [Kurata et al., 2007;
Kurata et al., 2006] などが実験的に確認されており,力学的環境下における骨細胞の他
の骨系細胞への作用に関する知見が得られつつある.これらの知見とともに,骨細胞
の骨基質中における分布や細胞間ネットワークの存在は,骨リモデリングにおける骨
7
Page 13
細胞の重要な役割を示唆するものでありながら,骨細胞が骨リモデリングを調節する
メカノセンサーとして機能するメカニズムについては,今なお不明な点が多い.その
ため,骨基質における骨細胞の力学刺激感知メカニズムを解明するためには,刺激を
感知する単位機構としての単一の骨細胞における力学刺激感知メカニズムの理解が
重要となる.
1.2.4 骨細胞の力学刺激感知特性
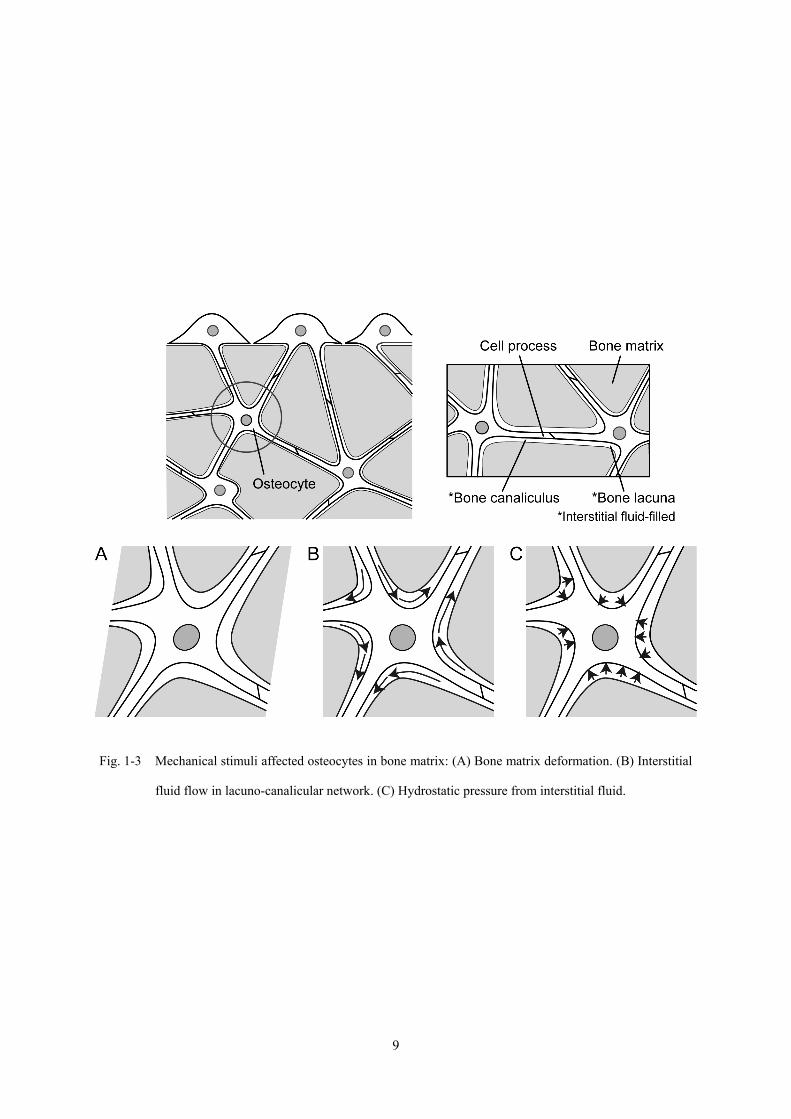
骨細胞の機能に影響を与えると考えられる力学刺激の形態としては,骨組織の変形
にともなう骨基質や骨細胞自身の変形 [Nicolella et al., 2006; 伊藤ら, 2007],骨細胞が
分布する骨小腔―骨細管系内 (lacuno-canalicular system) に生じると考えられる間質
液流 [Cowin et al., 1991; Weinbaum et al., 1994],あるいは,静水圧 [Anderson and Knothe
Tate, 2008; Mishra and Knothe Tate, 2003] などが提案されている (Fig. 1-3).このような
力学刺激に対する骨細胞に関する応答特性の実験的検討は,骨細胞の単離法
[Mikuni-Takagaki et al., 1995; van der Plas and Nijweide, 1992] や骨細胞樹立株 [Kato et
al., 2001; Kato et al., 1997] の確立を経て,in vitro 環境で行われるようになった.これ
までに,骨基質内部において骨細胞に作用するとされる力学刺激に対応した,培養基
質の変形による骨細胞の変形 [You et al., 2000] や,細胞外液の流れによるせん断刺激
[Jacobs et al., 1998; Klein-Nulend et al., 1995a; Klein-Nulend et al., 1995b],細胞に対する
直接刺激 [Vatsa et al., 2006] などの力学刺激の付与に対して,骨細胞が骨形成・骨吸
収を骨系細胞に促す骨代謝調節因子である一酸化窒素 (nitric oxide: NO) [van't Hof and
Ralston, 2001] やプロスタグランジン (prostaglandin: PG) [Raisz et al., 1993] を産生す
ることが報告されている.
8
Page 14
Fig. 1-3 Mechanical stimuli affected osteocytes in bone matrix: (A) Bone matrix deformation. (B) Interstitial
fluid flow in lacuno-canalicular network. (C) Hydrostatic pressure from interstitial fluid.
9
Page 15
メカノトランスダクション (mechanotransduction),すなわち,力学刺激が生化学シグ
ナルに変換される過程における初期応答のひとつとして,細胞内のカルシウム応答
(calcium-signaling response) が挙げられる.カルシウム応答は,細胞分裂や増殖,遺伝
子発現など,種々の細胞機能の上流調節因子として機能することが知られている
[Berridge et al., 1998].カルシウム応答を含め,カルシウムイオンを必要とする細胞応
答の発現の多くは,細胞における各種の応答機構に先行して発生する.例えば,骨系
細胞において,力学刺激の付与にともなう細胞内へのカルシウムイオン流入を物理的
に阻害すると,遺伝子の発現 [Chen et al., 2000; You et al., 2001a] やプロスタグランジ
ンの一種である PGE2 の産生 [Ajubi et al., 1999] ,またアクチン細胞骨格 (actin
cytoskeleton)の再構成 [Chen et al., 2000] が阻害されることが報告されている,また,
細胞膜の局所変形にともなう骨細胞のカルシウム応答は,細胞間連絡構造を介して隣
接する細胞に拡散伝達されることが報告されており [Gu et al., 2006; Yellowley et al.,
2000].カルシウム応答の細胞間伝達は,骨細胞が骨基質中で形成する細胞間ネット
ワーク構造を介した情報伝達,および骨細胞をはじめとする骨系細胞間の同調のメカ
ニズムとの関連が示唆されている [Guo et al., 2006].これらのことより,力学刺激に
対する骨系細胞のカルシウムイオンの動態を観察・測定することは,力学刺激に対す
る初期応答能を評価する方法として有効である.
10
Page 16
1.3 本論文の目的
1.3.1 目 的
骨リモデリングにともなう骨組織の代謝回転は,実質的には,破骨細胞による骨吸
収,ならびに,骨芽細胞による骨形成によって実現しており,骨代謝現象における骨
細胞の直接的な関与は不明な点が多い.しかしながら,第 1.1.3 項で述べたように,
骨細胞の存在は,骨リモデリングが正常に機能するために不可欠な存在であるととも
に,骨細胞が骨基質内で形成する細胞間ネットワーク構造は,力学的適応にともなう
骨リモデリングの調節メカニズムを説明できるものである.骨基質中の骨細胞が力学
刺激を感知し,細胞間ネットワークを介して刺激情報が伝達されることにより,正常
な骨リモデリングが実現すると仮定すると,骨細胞の力学応答特性を細胞レベルの応
答現象の観点から検討することは,骨リモデリング現象への理解を深める上で重要で
ある.
そこで,本論文では,骨基質中の骨細胞の力学刺激の感知・伝達特性に着目し,骨
基質中における骨細胞の力学応答特性,および,細胞間ネットワーク構造が,骨リモ
デリング現象に果たす役割を解明することを目指して,力学刺激の付与に対する初期
応答としてのカルシウム応答挙動を実験的に検討・評価することを目的とした.
1.3.2 医工学的意義
重力や荷重などの力学的環境の影響により骨が変形する現象 [Cowin, 2000],すな
わち,骨の力学的適応能は,臨床においては,例えば,骨折後に癒合した骨が自然に
矯正される事例や,長期間のベッドレスト状態が廃用性の骨萎縮を併発させる症例
[Greenleaf, 1984] などとして,古くから観察されてきた.その一方で,リハビリテー
ションや歯科矯正などの臨床領域では,このような骨の力学的適応能を積極的に応用
する試みも行われてきた.これらの方法は,骨や歯に対して意図的に力学的負荷を与
える処置により,骨や,歯を支持する歯槽骨の形態変化を促し,短期間での骨量の回
復や矯正を試みるものである [Lin and Lane, 2006; Ren et al., 2003; 山本, 2008].
骨細胞の力学刺激応答特性を明らかにすることは,骨の力学的適応現象に対する理
解を深めるとともに,先に挙げたリハビリテーションや歯科矯正術を効率的に実施す
11
Page 17
るための指針策定,臨床医療技術をはじめとする医工学分野への技術的応用のための
基礎的知見につながる.骨折骨の固定,骨の欠損部治療における固定器や人工置換
材・補填材などの導入は,骨組織に新たな力学的環境をもたらし,骨の力学的適応に
ともなうリモデリングの結果,骨の変形を生じさせることが予測される.例えば,骨
折骨の固定の場合,過度に強固な固定は,骨の支持構造体としての機能を失わせ骨の
骨質を低下させることから,骨折部を癒合させてもとの状態に戻すという本来の目的
が果たせなくなる可能性が生じ得る.骨と力学的環境との相互作用に関する知見に基
づき,骨の変形の抑止や適応的なリモデリングを実現する適切な荷重の付与条件を予
測・評価することができれば,骨再生技術の構築・実用化に当たって,強度や生体適
合性の観点に加えて,残存骨を含めた骨組織維持のための設計指針の確立に寄与でき
ると考えられる.
1.3.3 本論文の構成
本論文では,まず,第 2 章において,骨組織に埋没した形で存在する骨細胞につい
て,骨組織への変形付与に対するカルシウム応答を観察し,骨細胞の力学刺激に対す
る応答現象を検証した.次に,第 3 章では,刺激を感知する単位機構となる単一骨細
胞が,骨への力学的負荷を刺激として感知するメカニズムについて,力学刺激の付与
に対するカルシウム応答発生メカニズムを検討した.さらに,第 4 章では,骨細胞を
含む骨系細胞間で行われるカルシウム応答の細胞間伝播を観察し,骨組織内において
形成される細胞間ネットワークが,骨系細胞間の力学刺激情報伝達に果たす役割を検
討した.これらの実験的検討により,骨組織内における骨細胞の力学刺激応答特性,
および,細胞間ネットワークが骨リモデリング現象に果たす役割を明らかにすること
を目指した.
第 2 章では,骨基質内の骨細胞の力学刺激感知特性を実験的に検討した.骨組織に
対する変形の付与と,それに対する組織内骨細胞の生化学的応答の経時的観察を可能
とする ex vivo 実験系を確立し,骨組織と骨細胞の変形計測,および,それにともな
う組織内骨細胞のカルシウム応答を観察した.
第 3 章では,骨細胞の力学刺激感知特性において細胞形状が果たす役割に着目し,
骨細胞の局所部位における力学刺激感知特性を検討した.骨細胞に対して定量制御し
12
Page 18
た局所的な力学刺激を付与する in vitro 実験系を確立し,細胞の局所部位に対する力
学刺激付与実験を行い,カルシウム応答の発生に要する力学刺激量を検討した.また,
実験結果より得られた骨細胞の局所的な力学刺激感知特性について,骨基質内の骨細
胞における力学刺激感知メカニズムを考察した.
第 4 章では,骨細胞の力学刺激感知・伝達特性に対して,骨細胞が骨組織内におい
て形成する細胞間ネットワークがもたらす細胞間コミュニケーションの機能を検討
した.骨組織中に存在する骨細胞と,骨組織表面に分布する骨系細胞との間に生じる
力学刺激の付与に対するカルシウム応答の伝播現象を観察し,骨細胞を含む骨系細胞
における力学刺激伝達メカニズムを,骨組織中の骨系細胞の分布状態を踏まえて考察
した.
後に,第 5 章では,得られた結果を結論としてまとめ,骨の基礎科学,および,
医工学的観点から,その意義について論じた.
13
Page 20
第 2 章 骨組織の変形にともなう骨細胞のカルシウム応答
2.1 緒 言
骨細胞は,骨への力学的負荷を刺激として感知し,その情報を周囲の細胞に伝達す
るメカノセンサーとしての機能を有すると考えられている [Nijweide et al., 1996].し
かしながら,骨細胞は,周囲を骨基質に覆われた形で存在するため,実際の骨組織内
における骨細胞の力学刺激応答特性を検証することは困難である.このため,骨細胞
の力学刺激に対する応答特性は,骨組織から単離した骨細胞や骨細胞樹立株を用いた,
in vitro 実験系で検討されることが多い.
骨細胞は,硬い組織に埋没した形で存在するため,骨芽細胞や破骨細胞など,骨の
表面で骨代謝を担う細胞とは異なる力学環境下に置かれている.骨組織中の骨細胞が,
骨への力学的負荷を感知し,これらの細胞の骨代謝活動の調節を行っていると考える
と,骨細胞の力学応答特性の検討は,組織内環境下で実施することが望まれる.そこ
で,近年では,骨基質中に存在する ex vivo の状態における骨細胞の力学応答特性を
明らかにする試みが行われている.これまでに,骨組織を用いた ex vivo 実験系 [Mann
et al., 2006; Takai et al., 2004],組織を模した三次元ゲルを用いた in vitro 実験系 [Kurata
et al., 2006] などを用いた検討において,組織への力学的負荷や損傷に対する骨細胞の
生存率やアポトーシス,またシグナル因子の産生や骨芽細胞・破骨細胞との相互作用
15
Page 21
が明らかにされている.これらの報告では,組織化学的な検討に加えて,カルセイン
(Calcein) や DAPI (Ethidium homodimer-1,4',6-diamidino-2-phenylindole) を用いた骨組
織内部の骨細胞の蛍光観察による細胞の生死判定を行い,骨細胞の三次元的な分布状
態や,組織への力学的負荷に対する組織内骨細胞の生化学的挙動の観察を試みている.
しかしながら,これらの蛍光観察は,いずれも力学的負荷が付与されていない状態,
もしくは負荷後の組織に対して行われており,力学実験の 中における組織内骨細胞
の生化学的挙動を観察対象としていない.骨細胞を含む組織を用いた実験系において,
力学的負荷に対する骨細胞の生化学的応答の経時的観察が可能になれば,組織内骨細
胞の力学応答メカニズムへの理解がより深まると考えられる.
そこで本章は,骨組織に対する変形の付与と,それに対する組織内骨細胞の生化学
的応答の経時的観察を顕微鏡ステージ上で可能とする実験系の確立を目指した.ニワ
トリ胚頭蓋冠から調製した骨基質試料に対して変形を与え,組織内の単一の骨細胞の
変形,および初期応答としてのカルシウム応答を観察した.骨組織への変形付与に対
する組織内骨細胞のカルシウム応答の発生を観察することにより,組織内骨細胞の力
学刺激感知特性を確認することを目的とした.
2.2 実験方法
2.2.1 骨基質試料の作製
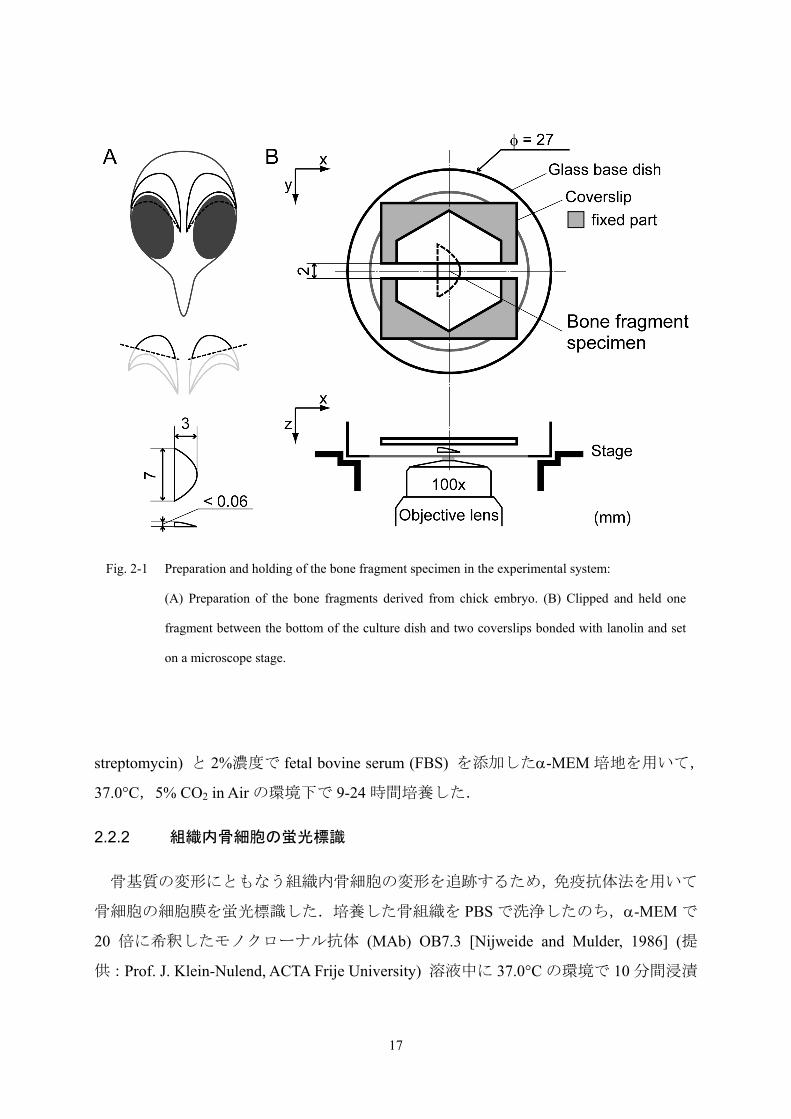
実験には,13 日齢ニワトリ胚より採取した頭蓋冠を用いた (Fig. 2-1A).ニワトリ
胚頭蓋冠は,厚さ約 60 μm の組織内部に骨細胞のネットワークが形成されており
[Kamioka et al., 2001],光学顕微鏡での透過光観察が可能である.採取した頭蓋冠は,
組織表面の骨膜および骨膜中の骨芽細胞を除去するため,Bone isolation buffer (BIB)
[Hefley, 1987]を溶媒とする濃度 1 mg/ml,温度 37.5°C の collagenase 溶液中で 30 分間
振盪培養した.また,骨組織に変形を付与するため,0.1% BSA を添加した Dulbecco's
phosphate buffered saline (DPBS) を溶媒とする温度 37.5°C の 0.5 mM ethylene etraacetic
acid (EDTA) 溶液中で 15 分間培養することにより,組織の表層領域に脱灰処理を施し,
組織を柔軟化した.各試薬により処理した頭蓋冠は,抗生物質 (ampicillin および
16
Page 22
Fig. 2-1 Preparation and holding of the bone fragment specimen in the experimental system:
(A) Preparation of the bone fragments derived from chick embryo. (B) Clipped and held one
fragment between the bottom of the culture dish and two coverslips bonded with lanolin and set
on a microscope stage.
streptomycin) と 2%濃度で fetal bovine serum (FBS) を添加したα-MEM 培地を用いて,
37.0°C,5% CO2 in Air の環境下で 9-24 時間培養した.
2.2.2 組織内骨細胞の蛍光標識
骨基質の変形にともなう組織内骨細胞の変形を追跡するため,免疫抗体法を用いて
骨細胞の細胞膜を蛍光標識した.培養した骨組織を PBS で洗浄したのち,α-MEM で
20 倍に希釈したモノクローナル抗体 (MAb) OB7.3 [Nijweide and Mulder, 1986] (提
供:Prof. J. Klein-Nulend, ACTA Frije University) 溶液中に 37.0°C の環境で 10 分間浸漬
17
Page 23
させた.その後,DPBS で洗浄し,2 次抗体として Alexa Fluor 546 goat anti-mouse IgG
(Invitrogen) を作用させることにより,免疫抗体法による蛍光標識を施した.MAb
OB7.3は,骨細胞の細胞膜貫通タンパク質 Phexに特異的である [Westbroek et al., 2002]
ことから,蛍光標識により骨細胞の同定を同時に行うことができる.
2.2.3 細胞内カルシウムイオンの可視化
骨基質の変形に対する骨細胞のカルシウム応答を観察するため,細胞内カルシウム
イオンを可視化し,力学刺激に対する細胞内カルシウムイオン濃度 ([Ca2+]i) の変動
を蛍光輝度の変化として観察した.本研究では,蛍光特性の異なる 2 種類のカルシウ
ムイオン蛍光指示薬を細胞内に導入し,蛍光輝度の輝度比をとることにより[Ca2+]iを
評価する Ratiometry 法 [Grynkiewicz et al., 1985] を用いた.
カルシウムイオン蛍光指示薬の導入は,2 次抗体のα-MEM 希釈培地に Ca2+蛍光指
示薬 Fluo-4 AM (Invitrogen) を 5 μM,Fura Red AM (Invitrogen) を 10 μM,および溶解
補助剤として Cremophore EL (Sigma-Aldrich) を 0.01%の濃度で添加し,培養した実験
試料を 37.0°C の環境で 15 分間培養することにより行った.その後,DPBS で洗浄し,
CO2 independent medium (Invitrogen) に入れ替えたものを実験に用いた.
18
Page 24
2.2.4 骨基質試料への変形付与
組織内の骨細胞に蛍光標識およびカルシウムイオン蛍光指示薬を導入した骨組織
は,Fig. 2-1B に示すように,2 枚のカバーガラスを用いて押さえ,その上からラノリ
ン,シリコンゴムを用いて押さえることにより φ = 27 mm Glass base dish (Iwaki) に固
定した.
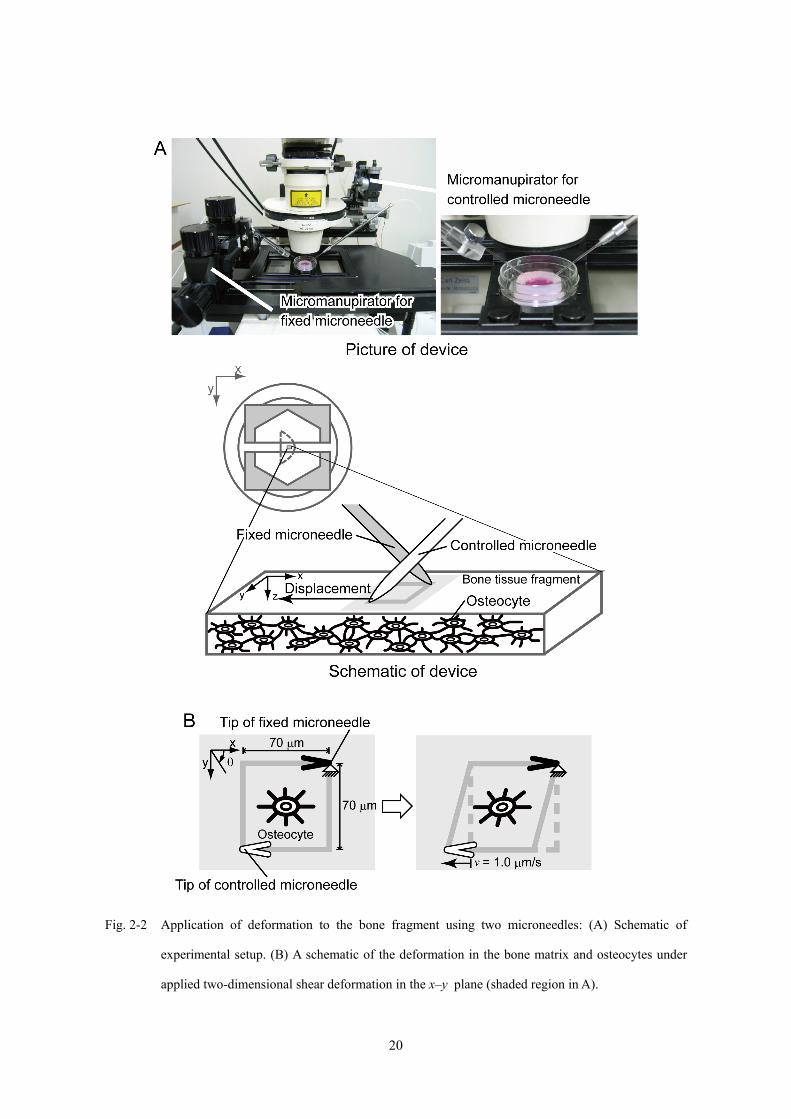
骨基質への変形付与の概要図を Fig. 2-2 に示す.骨基質への変形は,先端直径を約 φ
= 2 μm に加工した 2 本のガラス製マイクロニードルを用いて,Fig. 2-2A に示すよう
に骨組織の観察面 (x-y 平面) 内にせん断変形を与えた.マイクロニードルは,直径 φ
= 1.0 mm のガラス管をマイクロピペットプラー (MC-10, Narishige) を用いて加熱延
伸することにより作製し,研磨機 (EG-40, Narishige) を用いて,先端直径がおよそ 2-5
μm となるよう,軸方向に対する垂直面を研磨した.2 本のマイクロニードルをそれ
ぞれ顕微鏡ステージに設置した 3 次元水圧式マイクロマニピュレーター (可動側:
ONW135, 固定側:MC-35A, Narishige) に装着し,初期のニードル先端位置を,一辺
70 μm の正方形領域の対角に配置した.一方のマイクロニードルの位置を固定したま
ま,もう一方のマイクロニードルの先端を v = 1.0 μm/s の一定速度で x 軸の負の方向
(左方向) に水平移動させた.これにより,2 本のニードル先端間の骨組織に対して,
Fig. 2-2B に示すような x-y 面内の 2 次元的なせん断変形を与えた.
19
Page 25
Fig. 2-2 Application of deformation to the bone fragment using two microneedles: (A) Schematic of
experimental setup. (B) A schematic of the deformation in the bone matrix and osteocytes under
applied two-dimensional shear deformation in the x–y plane (shaded region in A).
20
Page 26
2.2.5 骨基質試料および組織内骨細胞の変形解析
骨基質変形時の骨細胞の変形を観察するため,変形付与前後の骨基質の変形状態を
微分干渉観察画像として,また,骨細胞の細胞形状を蛍光観察画像としてそれぞれ取
得した.骨基質の微分干渉観察画像と細胞膜の蛍光観察画像に対して,組織の厚さ方
向 (z 方向) の画像情報を取得した.注目する骨細胞の細胞形状が も明瞭に現れる
観察面を観察基準面 (z = 0.0 μm) とし,z = -7.5 ~ 7.0 μm の領域における断層撮影画像
を,微分干渉観察 (骨基質) および赤色 (細胞膜) の観察波長帯域 (585 nm Long Pass)
を用いた蛍光観察により,それぞれ 0.5 μm 間隔で 30 枚取得した.取得した画像群は,
PC 上で重ね合わせることにより,3 次元的に分布する骨基質内部の骨小腔構造,およ
び骨細胞の細胞形状を,1 枚の x-y 平面に投影した画像としてそれぞれ再構築した.
上記のようにして変形付与前後の断層撮影画像より得られた,微分干渉観察および
蛍光観察の投影画像の組に対し,画像解析ソフトウェア Flow-vec32 (ライブラリー)
による画像相関法 [小林ら, 2004] を適用し,せん断変形による骨基質および骨細胞の
変位場を求めた.
本解析では,画像相関法の解析精度を向上させるため,1 pixel を超える分解能での
変位分布を求めることが可能なサブピクセル解析手法 [小林ら, 2004] を用いた.すな
わち,画像相関を取る一組の画像について,1 枚目の画像のある pixel に対して,2 枚
目の画像の中でもっとも相関の強い pixel とその両隣の pixel での相関関数を求め,そ
れら 3 点を通る 2 次関数から相関関数が極大となる位置を求めた.
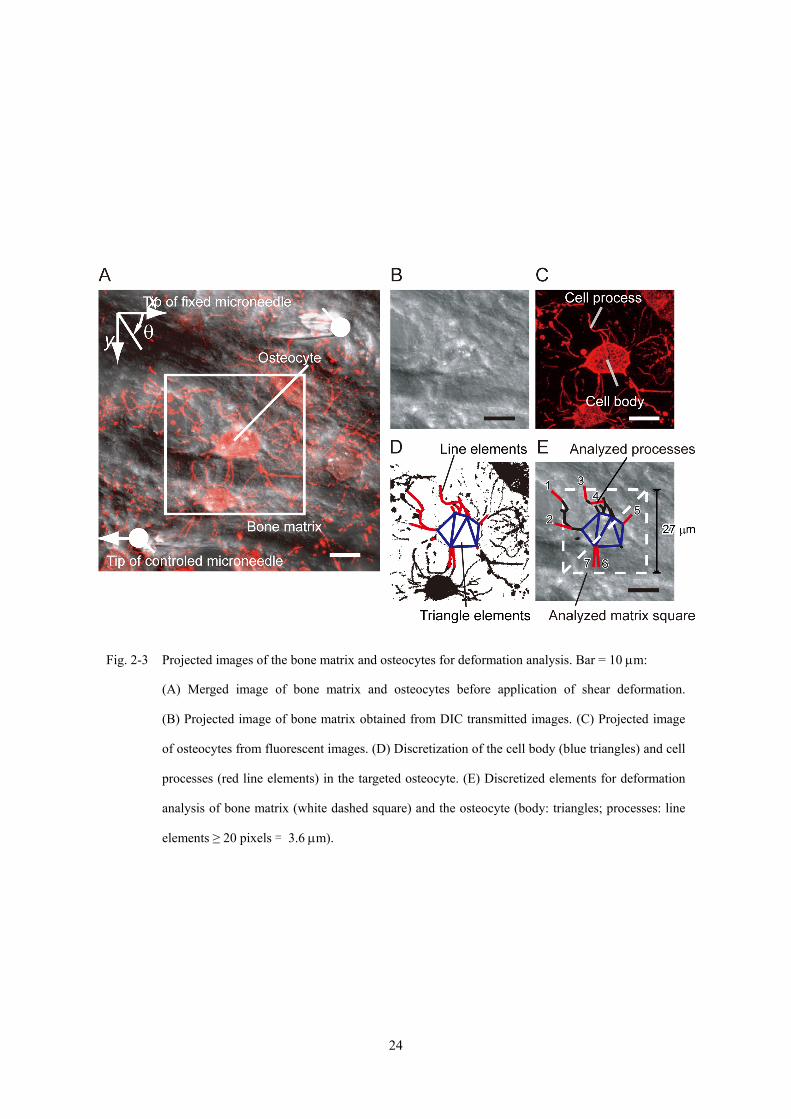
取得した組織断層撮影画像から作成した投影像,および,投影像から抽出・離散化
した骨基質・骨細胞の解析領域を Fig. 2-3 に示す.骨基質のひずみは,骨細胞周辺領
域の平均的なひずみとして求めた.すなわち,細胞体重心位置を中心とした一辺 150
pixel (27 μm) の正方領域を,ふたつの三角形領域に分割し (Fig. 2-3E 破線),領域頂点
の変位よりそれぞれの領域内のひずみを求め,それらの平均ひずみを求めた.一方,
骨細胞の細胞体部分と細胞突起部分を投影画像の組織内骨細胞の蛍光観察画像から
定義し (Fig. 2-3D),それぞれの部位におけるひずみを求めた.細胞突起のひずみは,
細胞突起を線分の集合とみなし (Fig. 2-3D 赤実線),各線分の端点の変位より変形前
後の細胞突起の長さ変化を求め,細胞突起長手方向のひずみを計算した.また,細胞
体のひずみは,細胞突起を表す線分の細胞体側起点を結ぶ多角形領域を分割した三角
21
Page 27
形要素 (Fig. 2-3D 青実線) に対して,各頂点の変位から求めた変位勾配より三角形の
ひずみを算出し,さらに,三角形要素の面積比に応じた重みづけ平均により,細胞体
全体 (多角形領域) の平均ひずみを求めた.
2.2.6 組織内骨細胞のカルシウム応答の経時観察
骨基質の変形に対する組織内骨細胞のカルシウム応答の観察には,共焦点レーザー
走査型蛍光顕微鏡 (LSM510, Carl Zeiss) を用い,対物レンズは 100 倍油浸レンズを使
用した.変形付与中の組織内骨細胞のカルシウムイオン濃度変化を観察するため,ふ
たつの Ca2+蛍光指示薬を 488 nm の波長光で同時に励起したのち,変形付与中の骨細
胞の蛍光画像を 0.25 sec 間隔で取得した.観察波長帯域は,緑色 (505-530 nm Band
Pass) と赤色 (560 nm Long Pass) とし,Fluo-4 (516 nm peak) の蛍光と Fura Red (599
nm peak) の蛍光をそれぞれ異なるチャンネルで検出した.取得した蛍光画像は,8 bit
(256 階調) の輝度画像として PC に記録した.
22
Page 28
2.3 実験結果
2.3.1 断層撮影画像の投影と変形解析領域の設定
マイクロニードルを用いて与えた x-y 面内の二次元せん断変形にともなう骨基質・
骨細胞の変形を定量的に評価するため,変形付与前後における断層撮影画像を x-y 平
面に投影して重ね合わせた画像を作成した.厚み 14.5 μm の観察領域に対して,撮影
した骨基質と骨細胞のそれぞれ 30 枚の断層撮影画像を 1 枚の画像に重ね合わせた
(Fig. 2-3A).これにより,骨基質中に存在する骨小腔などの構造が強調された画像 (Fig.
2-3B),および,三次元的に細胞突起を伸展する骨細胞の全体形状を二次元的に再現
した画像 (Fig. 2-3C) をそれぞれ取得することができた.
作成した骨基質・骨細胞の投影図を用いて,変形付与にともない骨基質・骨細胞に
生じる変形を解析するため,それぞれの画像に対して解析領域を設定した.骨細胞の
変形解析には,変形付与前後それぞれの画像において,骨細胞の細胞突起と細胞体を,
それぞれ区分的な線分および三角形要素に離散化し,各部位における変形付与前後の
各節点の変位を追跡した (Fig. 2-3D).骨基質の変形解析には,変形付与前の画像に対
し,細胞重心を中心とする正方領域を設定し,変形付与前後の画像に対して各頂点の
変位を追跡した (Fig. 2-3E).
23
Page 29
Fig. 2-3 Projected images of the bone matrix and osteocytes for deformation analysis. Bar = 10 μm:
(A) Merged image of bone matrix and osteocytes before application of shear deformation.
(B) Projected image of bone matrix obtained from DIC transmitted images. (C) Projected image
of osteocytes from fluorescent images. (D) Discretization of the cell body (blue triangles) and cell
processes (red line elements) in the targeted osteocyte. (E) Discretized elements for deformation
analysis of bone matrix (white dashed square) and the osteocyte (body: triangles; processes: line
elements ≥ 20 pixels = 3.6 μm).
24
Page 30
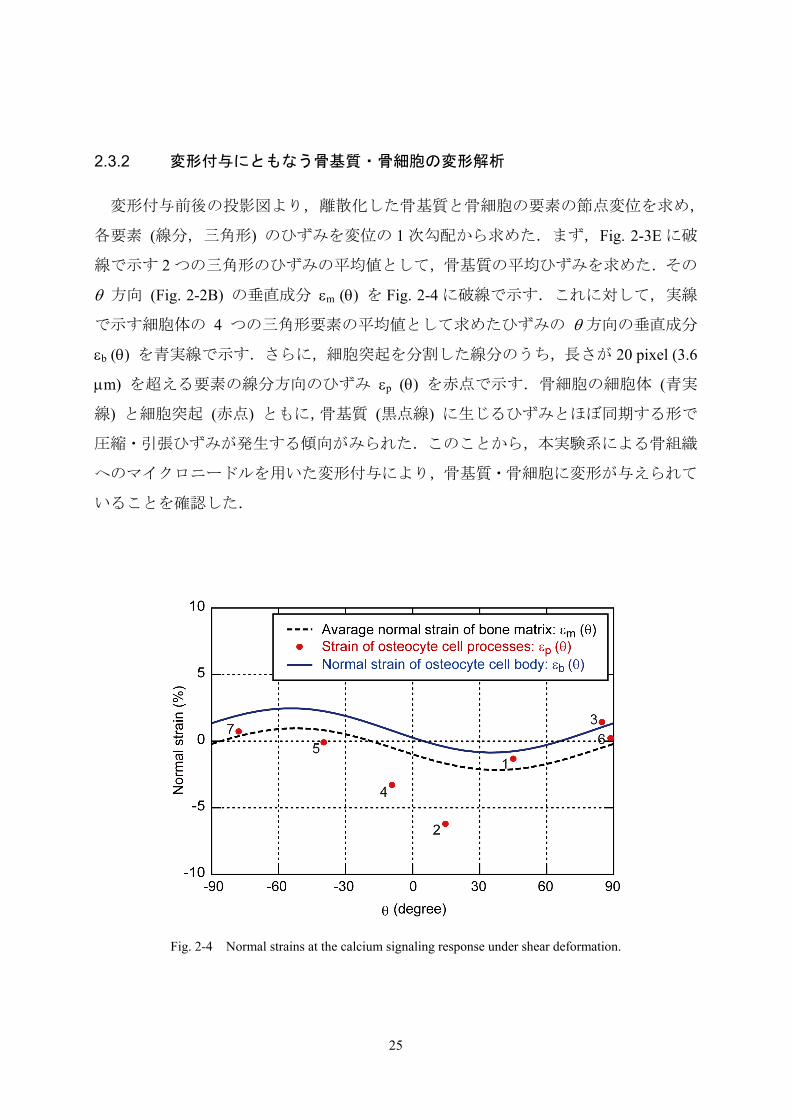
2.3.2 変形付与にともなう骨基質・骨細胞の変形解析
変形付与前後の投影図より,離散化した骨基質と骨細胞の要素の節点変位を求め,
各要素 (線分,三角形) のひずみを変位の 1 次勾配から求めた.まず,Fig. 2-3E に破
線で示す 2 つの三角形のひずみの平均値として,骨基質の平均ひずみを求めた.その
θ 方向 (Fig. 2-2B) の垂直成分 εm (θ) を Fig. 2-4 に破線で示す.これに対して,実線
で示す細胞体の 4 つの三角形要素の平均値として求めたひずみの θ 方向の垂直成分
εb (θ) を青実線で示す.さらに,細胞突起を分割した線分のうち,長さが 20 pixel (3.6
μm) を超える要素の線分方向のひずみ εp (θ) を赤点で示す.骨細胞の細胞体 (青実
線) と細胞突起 (赤点) ともに,骨基質 (黒点線) に生じるひずみとほぼ同期する形で
圧縮・引張ひずみが発生する傾向がみられた.このことから,本実験系による骨組織
へのマイクロニードルを用いた変形付与により,骨基質・骨細胞に変形が与えられて
いることを確認した.
Fig. 2-4 Normal strains at the calcium signaling response under shear deformation.
25
Page 31
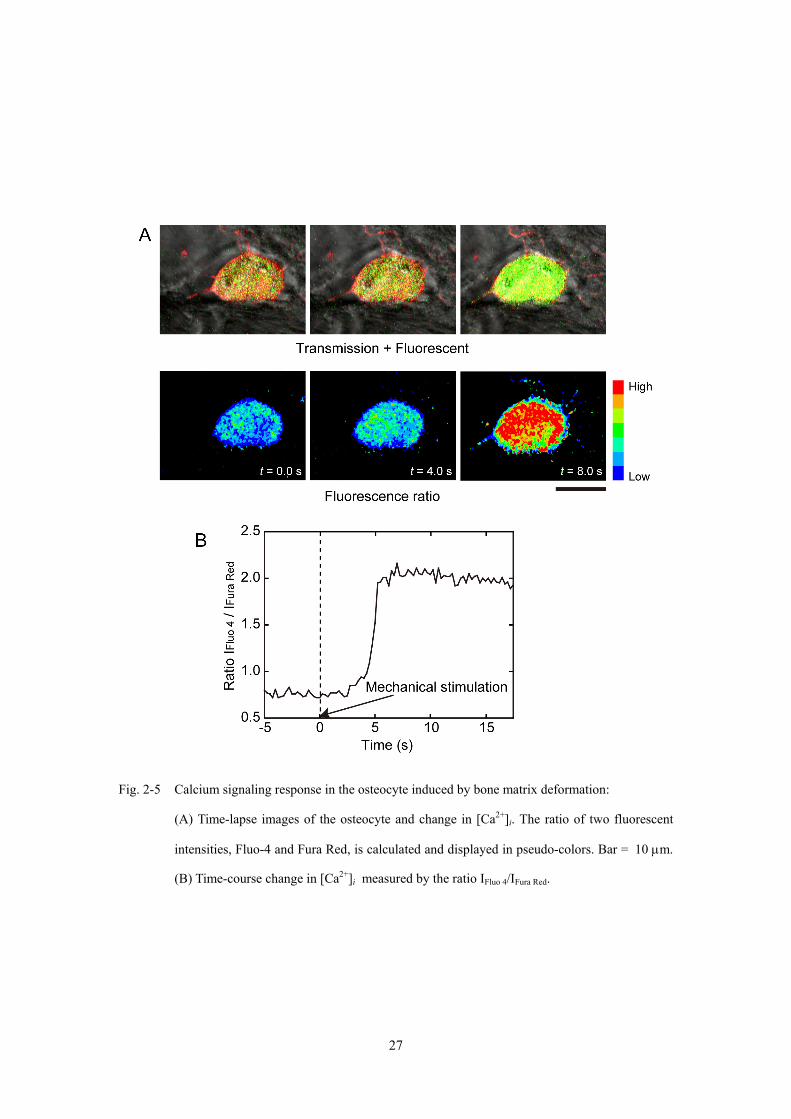
2.3.3 組織内骨細胞のカルシウム応答
骨組織に変形を付与した際の経時観察より,骨基質内における骨細胞のカルシウム
応答を観察した (Fig. 2-5).骨組織への変形付与中において,焦点面のずれおよび視
野からの大きな逸脱はみられなかった (Fig. 2-5A).また,2 種類の蛍光指示薬の輝度
比により示された [Ca2+]i より,変形付与中の細胞内カルシウム濃度の急激な変化が
観察された (Fig. 2-5B).このように,本実験系により,骨組織への変形にともなう骨
基質内の骨細胞のカルシウム応答発生を確認することができた.
26
Page 32
Fig. 2-5 Calcium signaling response in the osteocyte induced by bone matrix deformation:
(A) Time-lapse images of the osteocyte and change in [Ca2+]i. The ratio of two fluorescent
intensities, Fluo-4 and Fura Red, is calculated and displayed in pseudo-colors. Bar = 10 μm.
(B) Time-course change in [Ca2+]i measured by the ratio IFluo 4/IFura Red.
27
Page 33
2.4 考 察
本研究では,培養骨組織への変形付加に対する組織内骨細胞の応答を,直接ライブ
イメージングにより初めて定量的に観察した.すなわち,これまで,カルシウム応答
に代表される力学刺激に対する速い応答と,組織および細胞の変形とを,ex vivo 環境
において同時に観察対象とする検討は行われていなかった.これは,組織に対する変
形付与にともない,細胞スケールの観察領域が,顕微鏡の焦点面,あるいは顕微鏡の
視野から大きく外れるなどの技術的な難しさに起因する.骨組織への力学的負荷に対
する骨細胞の応答観察例としては,力学刺激の付与後の細胞生存率やシグナル因子の
産生を組織化学的に観察・分析した例が報告されている.例えば,Takai ら [2004] や
Mann ら [2006] は,骨組織を用いた培養実験系により,培養骨組織中の骨細胞の生存
率やアポトーシスに及ぼす力学的負荷の影響について検討している.一方,組織の変
形や細胞に対する力学刺激は与えていないが,組織内骨細胞の生化学的応答をライブ
イメージングで経時観察を試みた例も報告されている.例えば,Ishihara ら [2008] は,
副甲状腺ホルモン (PTH) による細胞間ギャップ結合 (GJIC: Gap Junctional
Intercellular Communication) の 制 御 を , FRAP (Fluorescence Recovery After
Photobleaching) 法を用いて確認している.これらの実験に対して,本研究では,培養
骨組織への変形付与にともなう組織内骨細胞の変形,および,細胞内の生化学的応答
を直接ライブイメージングにより観察することができた.
2.4.1 組織内骨細胞のライブイメージング実験
本研究では,2 本のガラス製マイクロニードルを用い,顕微鏡ステージ上で骨基質
試料の観察面に対して平行な面内でせん断変形を付与する新たな実験系を提案した.
この手法を用いて,骨組織に対して二次元的なせん断変形を付与することにより,組
織内骨細胞に対しても同様なせん断変形が与えられることを確認した.また,骨組織
への変形付与中,観察面および観察視野の大きなずれは生じず,組織内骨細胞の形状
変化やカルシウム応答のライブイメージング観察が可能となった.これにより,骨基
質の変形にともない生じる骨細胞のひずみの大きさや変形速度と様々な生化学的応
答との関係など,組織内骨細胞の力学刺激に対する応答特性を ex vivo 環境において
28
Page 34
定量的に評価するための基礎的な実験を確立した.
本実験系では,変形領域の中心に存在する単一の骨細胞の変形および細胞応答を観
察した.観察面に対して二次元的な面内せん断変形を付与する本手法は,同一焦点面
であれば,変形付与中の骨組織と周囲の複数の骨細胞の力学応答挙動の同時観察が可
能となる.また,観察面に対して垂直方向の分解能を持つ観察系 [Adachi et al., 2008]
を併用することにより,三次元的な細胞間応答伝播挙動の観察も可能になる.このよ
うに,本研究で提案した骨組織に対する力学刺激付与方法は,今後,組織内の細胞間
コミュニケーションなど,ネットワークを形成する複数の細胞の応答特性を評価する
実験系へと拡張することが可能である.
また,本研究では,組織内骨細胞のカルシウム応答を観察するため,細胞内に 2 種
類のカルシウムイオン蛍光指示薬を導入し,それぞれの蛍光輝度の比を取ることによ
り,[Ca2+]i の定量的に評価する Ratiometry 法を用いた.Ratiometry 法は,本来は in vitro
観察における [Ca2+]i の定量評価法として提案された手法である [Grynkiewicz et al.,
1985].蛍光輝度の比を取ることにより細胞厚さに由来する見かけ上の蛍光輝度の変
化を打ち消す本手法は,本実験系のように,厚みのある骨基質試料に対して変形を付
与し,組織中の細胞を蛍光観察する場合においても,組織内骨細胞の 3 次元的な分布
や,変形中の細胞厚さ変化に由来する蛍光輝度変化の影響を低減させ,組織内骨細胞
の [Ca2+]i の評価において有効であることを確認した.
2.4.2 骨基質の変形にともなう骨細胞の力学刺激応答
骨組織中の骨細胞の構造的特徴として,細胞骨格アクチンフィラメント構造
[Tanaka-Kamioka et al., 1998] や,接着に関連するビンキュリン [Kamioka et al., 2006]
(vinculin),パキシリン [Vatsa et al., 2008] (paxilin) などのタンパク質の局在が挙げられ
る.これらの細胞内構造物は,細胞の力学刺激の受容において重要な役割を果たすと
考えられている[Wang Y. et al., 2007; You et al., 2004].したがって,組織内骨細胞の細
胞内構造物の発現部位を蛍光可視化した上で,本実験システムを用いて力学的刺激に
対する生化学的シグナル応答を経時観察することにより,組織内骨細胞の力学応答の
発生部位や細胞内・細胞間伝播などの応答現象,また,応答発生時の骨細胞の力学状
態などの詳細な検討が可能になると考えられる.
29
Page 35
30
2.5 結 言
本章では,組織内骨細胞の力学刺激感知特性を検討するため,骨組織に対する変形
付与と,組織内骨細胞の生化学的応答の経時的観察を可能とする実験系を構築し,骨
組織と骨細胞の変形,および,それにともなう骨細胞のカルシウム応答を観察した.
その結果,生体から取り出して作製した骨基質試料を用いた ex vivo 実験系において,
骨組織への変形付与にともなう組織内骨細胞のカルシウム応答を初めてライブイメ
ージングにより直接観察した.また,骨細胞が骨基質の内部において,骨への力学的
負荷を刺激として感知することを実験的に確認した.この結果より,骨組織内部に分
布する骨細胞において,骨への力学的負荷を刺激として感知するメカニズムが機能し
ている可能性が示された.
Page 36
第 3 章 骨細胞の局所的構造と力学刺激感知特性
3.1 緒 言
前章の検討により,骨組織内の骨細胞が,骨への力学的負荷を刺激として感知する
ことを実験的に確認した.組織内骨細胞に作用する力学的刺激は,骨基質が形成する
複雑な力学的環境の影響を受ける.例えば,骨小腔に微視的かつ局所的に生じるひず
みは,巨視的なレベルで骨に生じるひずみよりも有意に大きいことが,実験的検討と
計算機シミュレーションによりそれぞれ示されている [Bonivtch et al., 2007; Nicolella
et al., 2006].これらの報告は,in vitro 実験系において,細胞の応答に要するひずみ
(1~3%) が,生理的レベルで骨に生じるひずみ ( 大 0.2%) を大きく上回る [Burger
and Klein-Nulend, 1999] 事実を説明する論拠のひとつとして挙げられる.
一方,細胞体から多数の細胞突起を伸長させた,骨細胞自身の特徴的な細胞形態も
また,組織内骨細胞の力学刺激感知特性に重要な役割を果たすと考えられている.骨
細胞の細胞膜には,骨細胞が細胞外基質 (extracellular matrix: ECM) に接着するための
焦点接着構造 (focal adhesive structure) に特徴的な分布がみられることが知られてい
る.焦点接着構造の構成要素であり,細胞内のアクチンフィラメントと細胞膜を縦貫
するインテグリン受容体 (integrin receptor) とを連結するビンキュリンは,骨基質中
の骨細胞の細胞突起突出部に局在して分布する [Kamioka et al., 2006].このインテグ
31
Page 37
リンと焦点接着構造の分布形態は,骨細胞の細胞膜に対する変形・ひずみの局所的な
集中を引き起こすことが考えられる.また,プロテオグリカンなど,骨細管内部に存
在する細胞突起と骨細管内壁との牽引構造やインテグリンの分布が細胞突起のひず
みを増幅するという仮説 [Han et al., 2004; Wang Y. et al., 2007] では,流れによって引
き起こされる細胞周囲基質の牽引力が,骨細胞の細胞突起を変形させ,細胞膜上の接
着構造や細胞内に力学的刺激を伝達すると考えられている.
以上に述べたように,骨細胞の細胞突起は,骨細胞の力学的刺激受容において重要
な部位であると考えられる.しかしながら,力学的刺激に対する骨細胞の細胞突起部
位の意義を直接的に示した検討はこれまでに行われていない.したがって,その重要
性を検討するための手段として,骨組織より単離した骨細胞において,細胞突起と細
胞体の力学応答能を比較することは,非常に重要な意味を持つと考える.
本章では,骨組織から単離・培養した単一の初代培養細胞における,局所的な力学
刺激感知特性を検討した.特に,骨細胞の細胞体部位および細胞突起部位の違いに着
目し,それぞれの部位の直接変形に対する初期応答としてのカルシウム応答を観察し
た.マイクロパーティクルにニワトリ骨細胞に特異的な MAb OB7.3 を修飾し,骨細
胞の細胞膜に付着させることにより骨細胞を同定するとともに.マイクロニードルを
用いた微視的な変形負荷を,細胞膜に付着させたマイクロパーティクルを介して付与
した.この実験系を用いて,マイクロパーティクルの変位を計測することにより,骨
細胞がカルシウム応答に至る際の微視的な変形負荷の大きさを計測し,骨細胞の局所
部位における力学刺激感知特性を検討するとともに,力学刺激の入力に対するカルシ
ウム応答発生のメカニズムを考察した.
3.2 実験方法
3.2.1 骨細胞の単離
13 日齢ニワトリ胚の頭蓋冠より,Kamioka ら [2004] の手法を一部改変したものを
用いて骨細胞を単離した.まず採取した頭蓋冠を 2 mm 角程度の大きさに剪刀で裁断
し,骨組織表面の骨芽細胞などの細胞群を骨膜ごと除去するために,Bone isolation
32
Page 38
buffer [Hefley, 1987] を溶媒とする 1 mg/ml collagenase type I (Sigma-Aldrich) 溶液中に
おいて,37.5°C で 30 分間振盪培養した.次に,骨組織を脱灰し,コラーゲン線維と
ともに骨組織の細胞を露出させるため,骨組織片を,DPBSを溶媒とする0.5 mM EDTA
に 0.1% BSA を添加した溶液中において,37.5°C で 15 分間の振盪培養を 2 度行った.
ここまでの作業では,骨組織片を各溶液中で振盪培養する前に,DPBS による骨組織
片の洗浄を毎回行った.その後,露出した骨組織中の細胞をコラーゲン線維から解放
するため,再度 1 mg/ml collagenase type I 溶液中において 37.5°C で 30 分間振盪培養し,
コラーゲンを酵素消化した.
振盪後,溶液の遠心分離を行い,上澄みを除いた沈殿物を抗生物質 (ampicillin およ
び streptomycin),および,2% fetal bovine serum (FBS, Sigma-Aldrich) を添加したα-MEM
(Invitrogen) 中に分散させた.この細胞を含む液体培地中から骨細胞を分離するため,
液体培地を孔径 φ = 10.0 μm のメンブレンフィルタ(Millipore) で濾過し,濾液として
フィルタを通過した骨細胞を含む液体培地を得た.単離した骨細胞は,φ = 35 mm の
Glass bottom dish (MatTek) に 7.5×103-2.0×104 cells/dish の細胞密度で播種し,1 時間
の培養後,培地を交換し,15-18 時間培養した.
遠心分離は,すべて 1200 rpm で 5 分間,骨細胞の培養は,温度 37°C,湿度 100%,
5% CO2 in Air の環境下でそれぞれ行った.また,細胞を播種した Glass bottom dish は,
20 μg/ml poly-D-lysine (Sigma-Aldrich),および 20 μg/ml fibronectin (ヒト血漿由来,
Chemicon Int.) で二重にコーティングしたものを用いた.
3.2.2 マイクロパーティクルの修飾および骨細胞への接着
マイクロパーティクル (φ = 1.0 μm, Polysciences, Inc.) を,20 μg/ml polyethylene
glycol (Wako Pure Chemical Industries) 溶液に懸濁させ,37°C の環境に 60 分間浸漬さ
せコーティングを施した.これにより,マイクロパーティクルを骨細胞の細胞膜に付
着させることが可能である.さらに,OB7.3 のコーティングを施したマイクロパーテ
ィクルが細胞に付着することにより,単離細胞中の骨細胞を同定することが可能とな
る.OB7.3 のコーティングを施したマイクロパーティクルは,後述のカルシウムイオ
ン蛍光指示薬導入時の培地中に懸濁させ,単離骨細胞を 37°C の環境で 15 分間培養す
ることにより,骨細胞の細胞膜にマイクロパーティクルを付着させた.
33
Page 39
OB7.3 のマイクロパーティクルのコーティング,および,マイクロパーティクルの
骨細胞への接着を確認するため,骨細胞にマイクロパーティクルを付着させた後,固
定処理を施し,細胞ならびにマイクロパーティクルを観察した.マイクロパーティク
ルを付着させた骨細胞を DPBS で洗浄し,3% paraformaldehyde (Wako) 溶液に室温環
境下で 10 分間静置して固定したのち,Alexa Fluor 546 goat anti-mouse IgG (Invitrogen)
溶液に室温環境下で 10 分間静置し,OB7.3 を蛍光標識した.標識後,DPBS で洗浄し,
細胞に付着していないマイクロパーティクルを除去したものを観察試料とした.また,
マイクロパーティクルの細胞膜に対する接着状態を確認するため,2% Glutaraldehyde
で固定した試料により電界放射型走査電子顕微鏡 (FE-SEM, S-4700, Hitachi
High-Technologies) 用の観察試料を作成し,細胞表面の観察を行った.
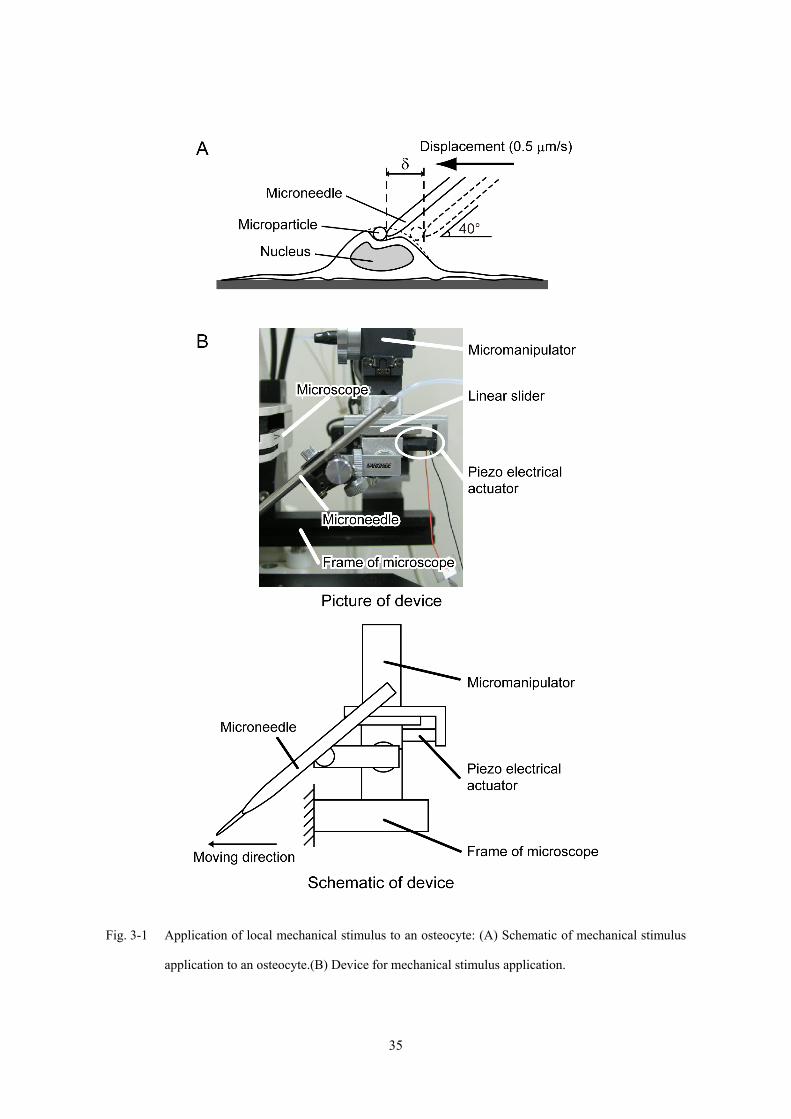
3.2.3 力学刺激の付与
単一の骨細胞に局所的な力学刺激を付与するため,Fig. 3-1A に示すようにガラス製
マイクロニードルを用いて細胞膜に付着させたマイクロパーティクルを介して細胞
膜に変形を与えた.マイクロニードルは,直径 φ = 1.0 mm のガラス管をマイクロピ
ペットプラー (MC-10, Narishige) を用いて加熱延伸することにより作製し,マイクロ
フォージ (MF-830, Narishige) を用いて先端部を溶融し直径約 φ = 2.0 μm の滑らかな
状態に加工した.顕微鏡に取り付けた三次元液圧式マニピュレーター (MM-188NE,
Narishige) に,dish 底面との角度を 40°としてマイクロニードルを固定した.
力学刺激の付与に用いたマイクロマニピュレーターの写真および模式図を Fig.
3-1B に示す.本研究では,定量的な力学刺激の付与に対するカルシウム応答発生を検
討するため,マイクロニードルの変位速度を制御することにより,付与する力学刺激
量を制御した.変位速度は,Sato ら [2007] による,マイクロニードルを固定したマ
イクロマニピュレーターにリニアスライダーとピエゾアクチュエータ (P802.10,
Physik Instrumente) を設置した装置 (Fig. 3-1B) を用いて,ピエゾアクチュエータに電
圧を印加することにより制御した.マイクロニードルの変位速度は,観察視野と,デ
ィッシュ培養の際に突起を除いた細胞長が直径 10-20 μm である骨細胞の寸法を考慮
し, v = 0.5 μm/s で一定とした.
34
Page 40
Fig. 3-1 Application of local mechanical stimulus to an osteocyte: (A) Schematic of mechanical stimulus
application to an osteocyte.(B) Device for mechanical stimulus application.
35
Page 41
細胞膜の変形付与は,骨細胞の細胞膜に付着させたマイクロパーティクルを介在さ
せることにより行った.その際,骨細胞全体の形状におけるマイクロパーティクルの
付着位置,および局所変形付与部位の位相差観察を行った.時間間隔 0.25 s の連続観
察画像を取得し,マイクロニードルによるマイクロパーティクルの移動が確認された
瞬間を,細胞に対する局所的な力学刺激の付与開始時間 t = 0.0 s とした.本研究では,
力学刺激の付与時間にともなうマイクロパーティクルの移動距離を,骨細胞に対する
力学刺激量として定義した.力学刺激の付与に対してカルシウム応答が観察された場
合,力学刺激の付与時間 t よりマイクロパーティクルの移動距離に対応するマイクロ
ニードルの移動距離 δ を求め,それぞれの試行について記録した.
3.2.4 細胞内カルシウムイオンの可視化とカルシウム応答の観察
骨細胞への蛍光指示薬の導入は,骨細胞を,α-MEM に Fluo-4 AM (Invitrogen) を 6
μM , Fura Red AM (Invitrogen) を 8 μM ,および溶解補助剤 Cremophor EL
(Sigma-Aldrich)を 0.01%の濃度でそれぞれ添加した培地で,37°C の環境で 15 分間培養
することにより行った.その後,DPBS で洗浄し,培地の pH を安定化させるために
CO2 Independent medium (Invitrogen) に入れ替えたものを実験に用いた.
局所変形付与に対するカルシウム応答の観察には,共焦点レーザー走査型蛍光顕微
鏡(LSM 510, Carl Zeiss) を用いた.第 2 章の手法と同様に,ふたつのカルシウムイオ
ン蛍光指示薬を 488 nmのレーザーで同時に励起した後,緑色 (505 - 530 nm Band Pass)
と赤色(560 nm Long Pass) のふたつの観察波長帯域を用いて,それぞれ Fluo-4 (516 nm
peak) の蛍光と Fura Red (599 nm peak) の蛍光を異なるチャンネルで検出した.取得し
た蛍光画像は,8 bit (256 階調) の輝度画像として PC に記録した.ふたつのカルシウ
ムイオン蛍光指示薬の蛍光輝度の輝度比を細胞内カルシウムイオン濃度 ([Ca2+]i) の
相対値とするため,観察で記録した輝度画像において,緑色蛍光 (Fluo-4) の輝度値
を赤色蛍光 (Fura Red) の輝度値で除算し,輝度比による 8 bit 画像を作成した.画像
作成および画像処理には,ImageJ 1.38 (NIH Image) を用いた.
カルシウム応答発生の判断には,局所変形付与後の [Ca2+]i の相対値が,局所変形
付与前の値の 1.5 倍以上に達した場合,カルシウム応答が発生したと定義する先行研
究 [Hung et al., 1995] の基準を用いた.細胞体部位と細胞突起部位,それぞれの変形
36
Page 42
付与部位ごとにカルシウム応答の発生率,およびカルシウム応答発生時の力学刺激量
を調べ,カルシウム応答の発生率に関してフィッシャーの正確確率検定,力学刺激量
に関してスチューデントの t 検定による統計的有意差検定をそれぞれ行った.
3.3 実験結果
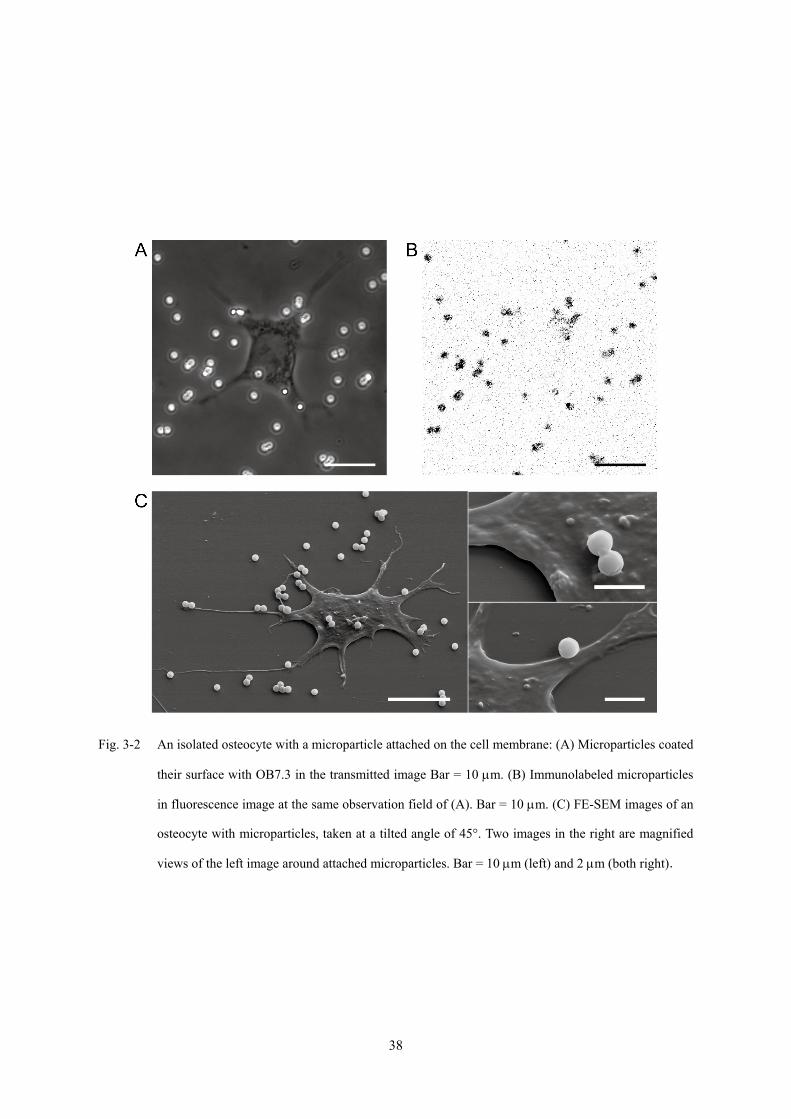
3.3.1 マイクロパーティクルの骨細胞への接着
MAb OB7.3 をコーティングしたマイクロパーティクルを Alexa Fluor 546 goat
anti-mouse IgG で蛍光標識した際の観察画像を Fig. 3-2A, 3-2B に示す.透過光による
観察画像 (Fig. 3-2A) において,DPBS による洗浄の後も骨細胞の表面にマイクロパ
ーティクルが観察され,マイクロパーティクルが骨細胞に付着していることが確認で
きる.また,マイクロパーティクルが,蛍光観察画像 (Fig. 3-2B) において蛍光標識
されていることが確認できる.これより,マイクロパーティクルが確かに OB7.3 でコ
ーティングされていることが確認できる.
骨細胞にマイクロパーティクルを付着させた試料を FE-SEM を用いて観察した画
像を Fig. 3-2C に示す.OB7.3 をコーティングしたマイクロパーティクルが,細胞体部
位,および細胞突起部位に接着している様子が確認できる.
37
Page 43
Fig. 3-2 An isolated osteocyte with a microparticle attached on the cell membrane: (A) Microparticles coated
their surface with OB7.3 in the transmitted image Bar = 10 μm. (B) Immunolabeled microparticles
in fluorescence image at the same observation field of (A). Bar = 10 μm. (C) FE-SEM images of an
osteocyte with microparticles, taken at a tilted angle of 45°. Two images in the right are magnified
views of the left image around attached microparticles. Bar = 10 μm (left) and 2 μm (both right).
38
Page 44
3.3.2 力学刺激の付与に対するカルシウム応答
骨細胞の局所部位に対して,マイクロニードルを用いてマイクロパーティクルを介
した力学刺激を付与した際の[Ca2+]i の変化を,細胞体部位に付与した細胞の例 (Fig.
3-3A ~ 3-3D)と細胞突起部位に付与した細胞の例 (Fig. 3-3E ~ 3-3H) に分けて,Fig. 3-3
にそれぞれ示す.Fig. 3-3A および Fig. 3-3E の,透過光観察画像と蛍光観察画像の重
ね合わせ画像において,白矢印で示されたマイクロパーティクルに対して,図の右方
向から速度 v = 0.5 μm/s で移動させてきたマイクロニードルを用いて変位を与えた.
力学刺激の付与開始時刻 (t = 0 s) における細胞内に導入した 2 種類のカルシウムイ
オン蛍光指示薬の輝度比を Fig. 3-3B および Fig. 3-3F に,力学刺激付与開始後の経時
画像を Fig. 3-3C および Fig. 3-3G にそれぞれ示す.白矢頭で示されるパーティクルの
変位による力学刺激の付与に対し,細胞体部位,および,細胞突起部位ともに [Ca2+]i
の上昇が観察され,細胞突起部位への力学刺激付与 (Fig. 3-3G) においては,マイク
ロパーティクルの近傍よりカルシウム応答が発生し,細胞体部位や他の細胞突起に波
及する様子が確認できる.
Fig. 3-3A,および,Fig. 3-3E,それぞれにおいて四角で囲まれたふたつの領域 (I, II)
について, [Ca2+]i の経時変化を Fig. 3-3D および Fig. 3-3H にそれぞれ示す.細胞体部
位に対する力学刺激では,経時観察画像 (Fig. 3-3C) より [Ca2+]i の上昇が細胞体全体
に波及する様子は確認できるものの,ゆるやかな濃度上昇のため,0.25 s ごとの画像
取得時間では明瞭なカルシウム応答の細胞内伝播は確認されなかった.一方,細胞突
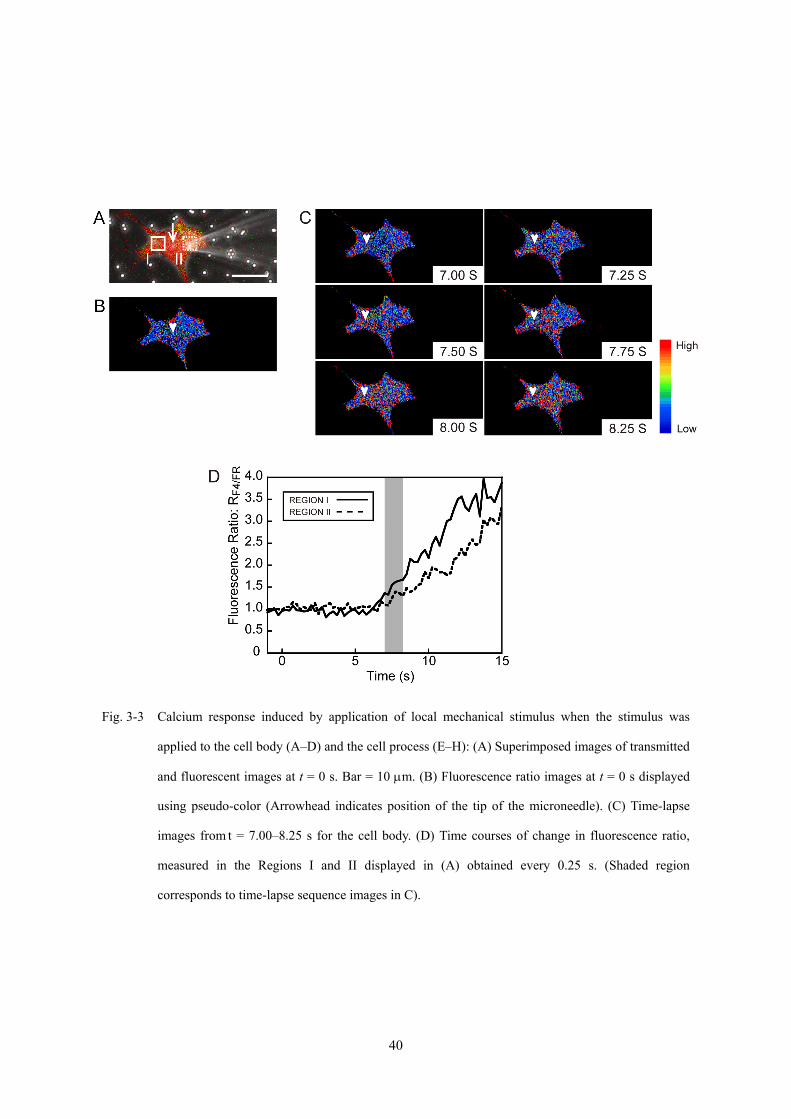
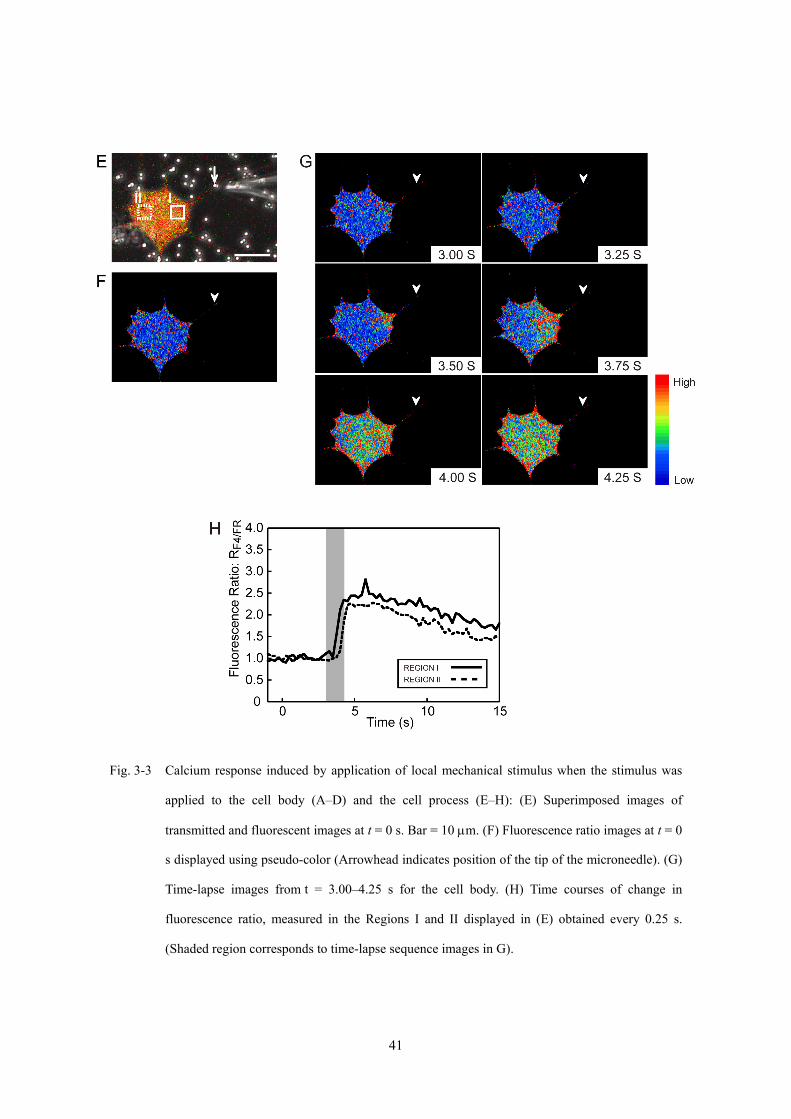
起部位に対する力学刺激では,力学刺激を付与した細胞突起により近い領域 (Region
I) で t = 3.75 s に急激なカルシウムイオン濃度の上昇が開始し,t = 5.75 s にピークに
達したのち,時間とともに減少した (Fig. 3-3H).一方,Region I から 10 μm 離れた細
胞突起から遠い領域 (Region II) では,カルシウムイオン濃度の上昇が,Region I にお
けるカルシウムイオン濃度の上昇から 0.25 s 遅れて発生していた.これらのことより,
カルシウム応答が力学刺激の付与部位近傍より発生し,1.00 s 以内に細胞全体に波及
していることがわかる.
39
Page 45
Fig. 3-3 Calcium response induced by application of local mechanical stimulus when the stimulus was
applied to the cell body (A–D) and the cell process (E–H): (A) Superimposed images of transmitted
and fluorescent images at t = 0 s. Bar = 10 μm. (B) Fluorescence ratio images at t = 0 s displayed
using pseudo-color (Arrowhead indicates position of the tip of the microneedle). (C) Time-lapse
images from t = 7.00–8.25 s for the cell body. (D) Time courses of change in fluorescence ratio,
measured in the Regions I and II displayed in (A) obtained every 0.25 s. (Shaded region
corresponds to time-lapse sequence images in C).
40
Page 46
Fig. 3-3 Calcium response induced by application of local mechanical stimulus when the stimulus was
applied to the cell body (A–D) and the cell process (E–H): (E) Superimposed images of
transmitted and fluorescent images at t = 0 s. Bar = 10 μm. (F) Fluorescence ratio images at t = 0
s displayed using pseudo-color (Arrowhead indicates position of the tip of the microneedle). (G)
Time-lapse images from t = 3.00–4.25 s for the cell body. (H) Time courses of change in
fluorescence ratio, measured in the Regions I and II displayed in (E) obtained every 0.25 s.
(Shaded region corresponds to time-lapse sequence images in G).
41
Page 47
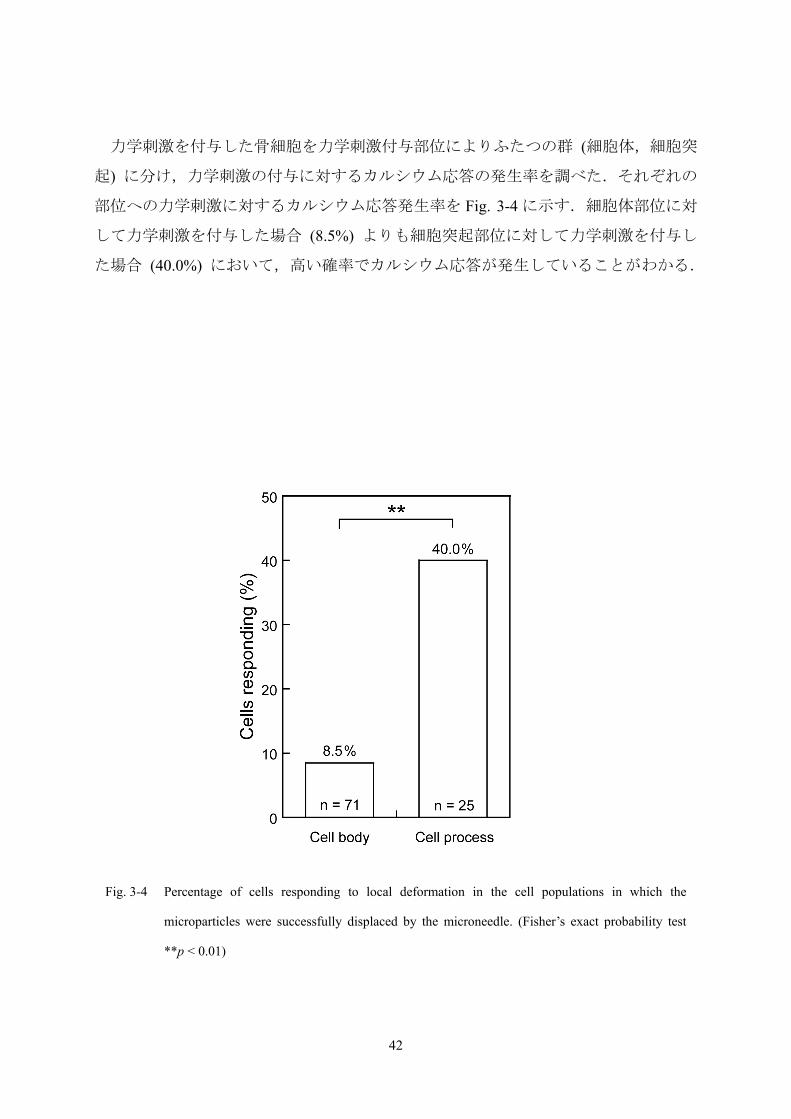
力学刺激を付与した骨細胞を力学刺激付与部位によりふたつの群 (細胞体,細胞突
起) に分け,力学刺激の付与に対するカルシウム応答の発生率を調べた.それぞれの
部位への力学刺激に対するカルシウム応答発生率を Fig. 3-4 に示す.細胞体部位に対
して力学刺激を付与した場合 (8.5%) よりも細胞突起部位に対して力学刺激を付与し
た場合 (40.0%) において,高い確率でカルシウム応答が発生していることがわかる.
Fig. 3-4 Percentage of cells responding to local deformation in the cell populations in which the
microparticles were successfully displaced by the microneedle. (Fisher’s exact probability test
**p < 0.01)
42
Page 48
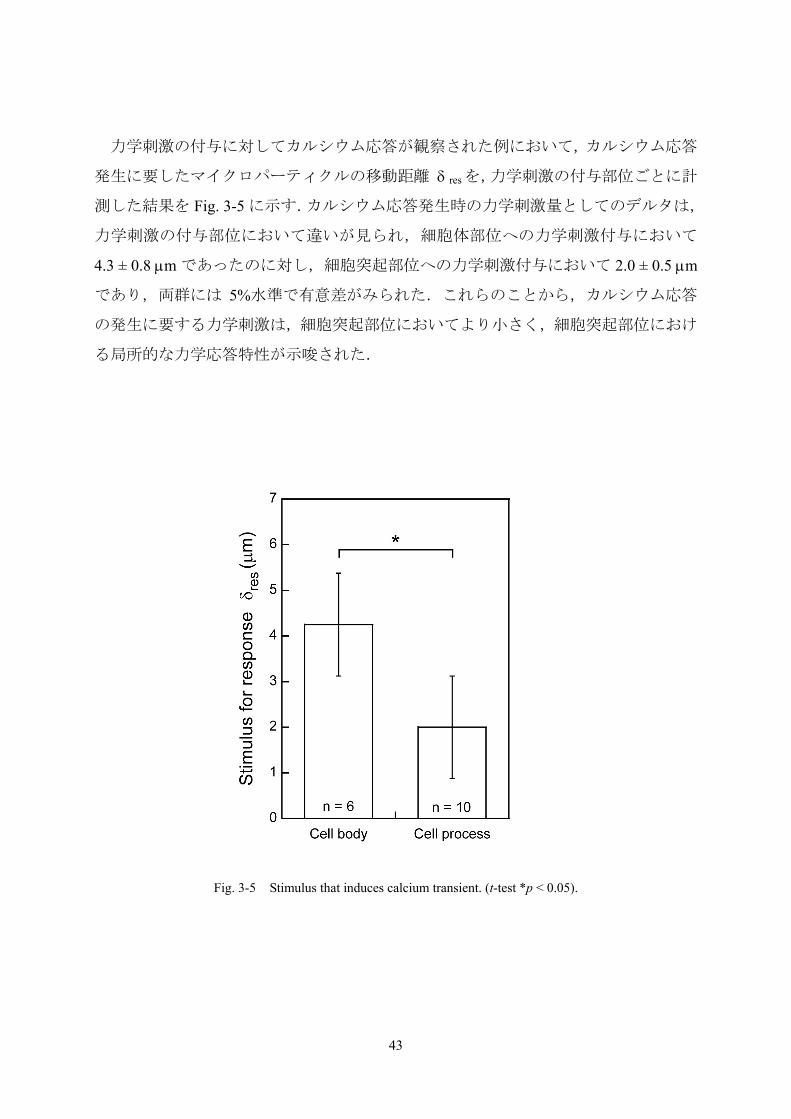
力学刺激の付与に対してカルシウム応答が観察された例において,カルシウム応答
発生に要したマイクロパーティクルの移動距離 δ resを,力学刺激の付与部位ごとに計
測した結果を Fig. 3-5 に示す.カルシウム応答発生時の力学刺激量としてのデルタは,
力学刺激の付与部位において違いが見られ,細胞体部位への力学刺激付与において
4.3 ± 0.8 μm であったのに対し,細胞突起部位への力学刺激付与において 2.0 ± 0.5 μm
であり,両群には 5%水準で有意差がみられた.これらのことから,カルシウム応答
の発生に要する力学刺激は,細胞突起部位においてより小さく,細胞突起部位におけ
る局所的な力学応答特性が示唆された.
Fig. 3-5 Stimulus that induces calcium transient. (t-test *p < 0.05).
43
Page 49
3.4 考 察
本研究では,マイクロパーティクルを介した骨細胞への局所的な力学刺激の付与に
対する細胞体部位と細胞突起部位のカルシウム応答特性を検討した.骨基質中の骨細
胞は,骨小腔と呼ばれる空隙に分布し,骨細管に多数の細胞突起を伸長させている.
したがって,骨細胞の細胞体と細胞突起は,それぞれ力学状態の異なる環境下におか
れていることが考えられる.このような配置の相違は,骨への力学的負荷によっても
たらされる力学的刺激が,骨細胞の部位によって異なる大きさで付与される可能性を
意味している.Anderson ら [2005] は,この可能性について計算機シミュレーション
により検討しており,骨への力学的負荷がもたらす細胞外液の移動が,骨細管におい
て高いせん断応力を発生させることを示している.また,Nicolella ら [2006] は,画
像相関法を用いた手法により,生理的な骨組織のひずみが,骨小腔近傍で局所的に増
幅されることを報告している.これらのことより,骨組織内の骨細胞は,骨小腔から
骨細管にかけての部位において局所的に増大した力学的刺激を感知すると考えられ
ている.その一方で,骨細胞自身の特徴的な細胞形状がもたらす組織内骨細胞の力学
刺激感知特性に関しては,組織観察画像に基づく計算機シミュレーションによる検討
が行われている [Han et al., 2004; Wang Y. et al., 2007] ものの,実験的検討によりその
意義を示す研究は行われていなかった.本研究で観察された,細胞体部位と細胞突起
部位における異なるカルシウム応答特性は,骨細胞の特徴的な細胞形状を形成してい
る細胞内構造が,骨細胞の局所的な力学刺激感知メカニズムに寄与していることが考
えられる.
3.4.1 マイクロパーティクルを介した局所的な力学刺激の付与
本研究では,単一の骨細胞に対して定量的に局所的な力学刺激を付与するため,骨
細胞の細胞膜に抗体でコーティングしたマイクロパーティクルを付着させ,一定速度
に制御したマイクロニードルを用いてマイクロパーティクルに変位を与える実験系
を確立した.この手法により,骨細胞に対して局所的な力学刺激を付与するとともに,
抗体による骨細胞の同定を行った.実験に用いたマイクロパーティクルには,MAb
OB7.3 のコーティングを施した.MAb OB7.3 は,ニワトリの骨細胞の細胞膜に存在す
44
Page 50
る膜貫通タンパク質 Phex に特異的に結合することが知られている [Westbroek et al.,
2002].したがって,MAb OB7.3 コーティングを施したマイクロパーティクルを用い
ることにより,パーティクルが結合した単離細胞を骨細胞と同定することができる.
また,MAb OB7.3 の骨細胞の細胞膜に対する強い結合力は,初期の骨細胞単離法にお
いて,磁性マイクロパーティクルを用いた panning法にも用いられている [van der Plas
and Nijweide, 1992].したがって,骨細胞の細胞膜に接着した MAb OB7.3 コーティン
グを施したマイクロパーティクルをマイクロニードルを用いて移動させることによ
り,骨細胞に対して,アクチン細胞骨格などの細胞内構造物に間接的な変形を付与す
ることが可能となった.
力学刺激の付与に対するカルシウム応答の発生は,マイクロパーティクルを介さず
に,マイクロニードルを用いて細胞膜に直接変形を付与する方法によっても確認され
ている.しかしながら,マイクロニードルの先端部は,細胞膜に接着していないため,
変位中のマイクロニードルが細胞膜上で滑る状態が生じる [Adachi et al., 2008; Sato et
al., 2007].そこで,本研究では,細胞膜の変位量を定量的に評価するため,細胞膜に
直接接着させたマイクロパーティクルを用いた.マイクロパーティクルの移動に用い
たマイクロニードルの変位は,一定速度に制御されており,またマイクロニードルの
変位は,マイクロパーティクルを介して細胞膜の変位に反映されることから,マイク
ロニードルの変位を計測することにより,細胞に対する局所的な力学刺激量を定量化
することが可能となった.
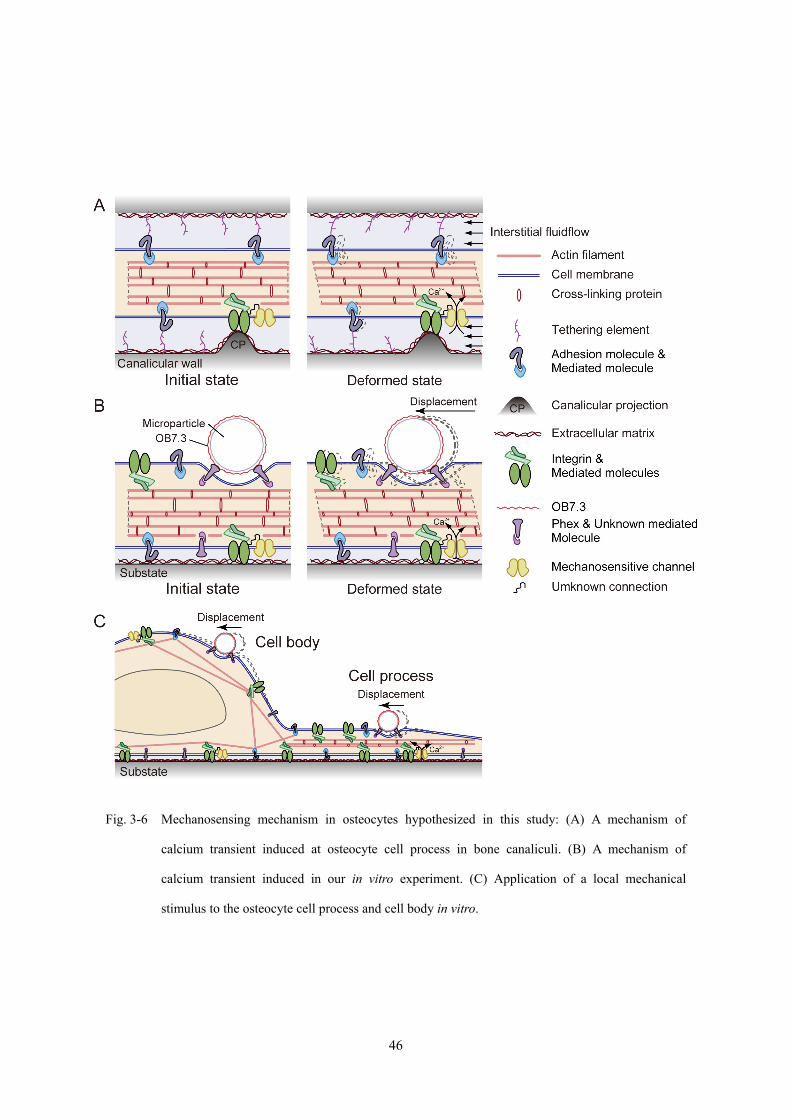
本研究の実験系の確立にあたって想定した,骨細胞の力学刺激受容メカニズムを
Fig. 3-6 に示す.このメカニズムは,細胞体部位や細胞突起部位への力学刺激が,力
学刺激受容 (MS) チャネル [Hayakawa et al., 2008] (mechanosensitive channel) が分布
するとされる焦点接着構造近傍に付与されることにより,力学刺激がアクチン細胞骨
格などの構造物を介して細胞内に伝達されるというものである (Fig. 3-6A).本研究で
は,力学刺激の付与に用いたマイクロパーティクルの変位量に対応するマイクロニー
ドルの変位量 δ を,骨細胞に対する力学刺激量として定義した.このことは,骨細
胞が受容する力学的刺激の形態として,Bonewald [2006] や Wang ら [Wang Y. et al.,
2007] が提案するように,焦点接着構造などの細胞膜上の固定支持点の相対的な摂動
(変位) が果たす役割を検討するためである.
45
Page 51
Fig. 3-6 Mechanosensing mechanism in osteocytes hypothesized in this study: (A) A mechanism of
calcium transient induced at osteocyte cell process in bone canaliculi. (B) A mechanism of
calcium transient induced in our in vitro experiment. (C) Application of a local mechanical
stimulus to the osteocyte cell process and cell body in vitro.
46
Page 52
上記のような骨細胞の力学刺激受容メカニズムを想定する場合,本研究で実際に用
いた実験系は,細胞膜上の焦点接着構造近傍に力学刺激を付与するという目的を十分
に果たしているとは言えない.前述したように,本研究の実験系においてマイクロパ
ーティクルの修飾に用いた MAb OB7.3 は,骨細胞の細胞膜に存在する膜貫通タンパ
ク質 Phex に特異的に結合する.しかしながら,Phex は,アクチン細胞骨格などの細
胞内構造物との連携が不明である.一方,本研究で用いた手法と同様の形態の力学刺
激を付与する実験系として,マイクロパーティクルにフィブロネクチンを修飾する方
法が挙げられる.フィブロネクチンは,細胞外基質を形成する主要な糖タンパクのひ
とつであり,細胞が基質に接着する際の足場としての役割を担う.フィブロネクチン
のコーティングを施したマイクロパーティクルは,膜貫通型タンパク質であるインテ
グリン受容体に接着し,インテグリンを介して細胞骨格構造との連携 [Ingber, 1991]
を形成すると考えられる.したがって,後に詳述する骨細胞の力学刺激受容メカニズ
ムを考慮すれば,本研究の実験系においても,フィブロネクチンを修飾したマイクロ
パーティクルを用いることが望ましい.
しかしながら,本研究において,骨細胞に対してフィブロネクチンのコーティング
を施したマイクロパーティクルの接着を試みたところ,細胞体および細胞突起の辺縁
領域に接着していることが観察されたが,細胞体部位の頂端側の細胞膜への接着はほ
とんど観察されなかった.このような細胞の局所部位における異なる接着性は,細胞
体部位と細胞突起部位における焦点接着構造の分布や,接着特性の違いに由来すると
考えられる.したがって,本研究では,細胞体部位と細胞突起部位の力学刺激受容特
性の違いを検討するにあたって,細胞体部位と細胞突起部位の両方にマイクロパーテ
ィクルが接着する MAb OB7.3 をコーティングした方法を用いた.MAb OB7.3 を用い
た抗原抗体反応は,骨細胞の細胞膜全体を標識することから [Kamioka et al., 2006],
本研究の目的である局所部位への力学刺激受容特性の比較検討が可能である.
また,本研究の実験系では,マイクロパーティクルに対する定量的な変位を圧電制
御したマイクロニードルを用いて付与したが,細胞に付与する力の計測は行っていな
い.骨細管内の細胞突起においては,細胞突起に分布する焦点接着斑を形成するイン
テグリンに対して pN レベルの力が作用することが理論モデルにおいて示されている
[Wang Y. et al., 2007].したがって,in vitro 環境下で pN レベルの力を計測する実験が
47
Page 53
可能になれば,骨細胞の力学刺激受容特性の理解において非常に有効な手段となり得
る.
3.4.2 細胞内カルシウムイオンの経時変化とカルシウム応答発生メカニズム
本研究の in vitro 実験系において,細胞膜上のマイクロパーティクルの変位を骨細
胞の内部に伝達すると考えられる接着構造は,in vivo 環境において骨細胞が骨細管の
内壁に対して形成する接着構造に対応している (Fig. 3-6A, 3-6B).骨基質内部の骨細
胞は,連結構造 (tethering elements) を介在して骨細管の内壁に接している [Wang Y. et
al., 2007; You et al., 2004] ことや,細胞体部位や細胞突起の基部に焦点接着構造が局在
していること [Kamioka et al., 2006; Vatsa et al., 2008] が報告されている,
組織内骨細胞において想定される,接着構造を介した力学刺激受容メカニズムの模
式図を Fig. 3-6A に示す.このメカニズムでは,You ら [2001b] や Wang [Wang Y. et al.,
2007] らの観察結果に基づいて,骨細管内に存在する骨細胞の細胞突起が,連結構造
によって骨細管内壁に牽引されており,骨細管内壁に存在する突出部位とインテグリ
ンなどを介した焦点接着構造を形成している.You ら [2001b],Han ら [2004],Wang
ら [Wang Y. et al., 2007] が考察しているように,骨への力学的負荷によって骨細管の
軸方向に生じる間質液の流れが,細胞突起と骨細管内壁の連結構造を介して細胞突起
を牽引するとき,固定支持点である焦点接着構造に相対的な変位が生じ,細胞内の介
在分子とアクチン細胞骨格を介して力学刺激を細胞内に伝達することにより,焦点接
着構造の近傍に存在する MS チャネルを活性化する.すなわち,細胞膜と骨細管内壁
との間に生じる相対的な摂動によってもたらされる,接着構造に対する局所的な力学
刺激が,MS チャネルを活性化するというものである.
同様に,本研究の実験系における力学刺激の付与に対するカルシウム応答は,Fig.
3-6B に図示したメカニズムによって説明することができる.Fig. 3-6B における,骨
細胞の細胞膜に接着したマイクロパーティクルの変位は,Fig. 3-6A で示した間質液の
流れが連結構造を介して細胞突起を牽引するメカニズムに対応しており,マイクロパ
ーティクルの変位により,細胞膜上の焦点接着構造に力学刺激の入力が生じていると
考えられる.
本章では,マイクロパーティクルを介した骨細胞への局所的な力学刺激の付与に対
48
Page 54
し,細胞体部位と細胞突起部位で異なるカルシウム応答特性を観察した.このことは,
Fig. 3-6C に示すように,骨細胞において細胞体部位よりも細胞突起部位に高い密度で
アクチンフィラメントが局在していること [Tanaka-Kamioka et al., 1998; 青沼, 2007]
が寄与していると考えられる.すなわち,マイクロパーティクルの変位にともなう固
定支持点に対する引張変形 [Adachi et al., 2008; Sato et al., 2007] が,固定支持点を形成
している焦点接着構造に牽引されている細胞内構造物を介して,力学刺激として伝達
されると考えられる.
3.5 結 言
本研究では,骨細胞の局所部位における力学刺激感知特性を検討するため,単離初
代培養骨細胞に対して局所的に定量的な力学刺激を付与する in vitro 実験系を構築し,
力学刺激の付与に対する骨細胞のカルシウム応答を観察した.その結果,骨細胞が細
胞突起部位において鋭敏な力学刺激感知特性を示す結果を得た.本研究で得られた結
果は,骨細胞の力学刺激感知メカニズムに対する細胞形状の寄与を示唆するものであ
り,単一の単離骨細胞における力学応答メカニズムの検討において,細胞形状による
局所的な力学刺激感知特性の差異を示したものである.
49
Page 56
第 4 章 骨系細胞における細胞応答の細胞間伝播
4.1 緒 言
前章における骨細胞の局所部位における力学刺激感知特性の検討では,骨細胞への
力学刺激の付与に対し,カルシウム応答が力学刺激の付与部位の近傍から発生し,そ
の後,細胞全体にカルシウム応答が波及する様子が観察された (Fig. 3-3C, [青沼,
2007]).骨細胞が感知した力学刺激情報は,種々の細胞機能を活性化させるとともに,
骨基質内部の骨小腔‐骨細管系からなる空隙構造や,細胞突起により構成された細胞
間ネットワークを介して骨表面に伝達すると考えられる [Aarden et al., 1994].例えば,
Ishihara ら [2008] は,ニワトリ胚頭蓋冠の骨基質中において,骨細胞‐骨芽細胞間に
ギャップ結合が存在し,さらに,このギャップ結合を介して,細胞間の物質移動が行
われていることを報告している.また,Vatsa ら [2007] は,骨細胞株 MLO-Y4 間の
NO の細胞間伝播を観察したほか,Yellowley ら [2000] は,骨細胞株 MLO-Y4 から骨
芽細胞株 MC3T3-E1 へのカルシウム応答の細胞間伝播を確認している.これらのこと
から,力学的負荷にともなう骨系細胞の応答,および,細胞間の情報伝達現象を理解
するためには,これらのシグナル分子の動態を観察する方法が有効である.
骨細胞が骨基質中においてメカノセンサーとして機能し,感知された刺激情報に応
じて骨表面における骨芽細胞・破骨細胞の活動が調節されると考えると,骨基質内部
51
Page 57
から骨表面に向かう細胞間ネットワークを介した情報の流れが存在することが推測
される,そのため,細胞間の伝達特性を明らかにすることは,骨リモデリングにおけ
る重要な知見を与えるものと期待される.そこで本章では,ニワトリ胚から単離した
骨細胞と主に骨芽細胞を多く含む骨表面細胞 (bone surface cells: BSC) が混在した in
vitro 実験系において,力学刺激に対するカルシウム応答の細胞間伝播を観察し,骨に
おける細胞間コミュニケーションについて考察した.
4.2 実験方法
4.2.1 骨系細胞の単離
13 日齢ニワトリ胚の頭蓋冠より,Kamioka ら [2004] の手法を一部改変して骨系細
胞を単離した.まず,裁断した頭蓋冠を,第 3 章における骨細胞の単離方法に従い,
collagenase 溶液および EDTA 溶液中で振盪培養したのち,細胞を含む液体培地を骨細
胞の単離時よりも孔径を大きくしたφ = 11.0 μm のナイロンネットフィルタ
(Millipore) で濾過し,濾液としてフィルタを通過した骨系細胞を含む液体培地を得た.
濾過後,濾液の遠心分離を行い,上澄みを除いた沈殿物を,0.5% NaCl 溶液と培地の
1:1 混合液に分散させ,液体培地中に含まれていた血液細胞などを破壊した.その後,
培地を加えて混合液を希釈し,遠心分離を行い,骨系細胞を沈殿物として得た.骨系
細胞は,φ = 35 mm glass bottom dish (MatTek) に 2.0×104 - 8.0×104 cells/dish の細胞密
度で播種し,1 時間の培養後,培地を交換し,その後 15-18 時間培養した.
4.2.2 骨細胞の同定
単離培養した骨系細胞を,骨細胞 (Ocy) とそれ以外の bone surface cells (BSC) に分
類するため,骨細胞の細胞膜貫通タンパク質 Phex に特異的なモノクローナル抗体
(MAb) OB7.3 [Nijweide and Mulder, 1986] を用いた免疫組織化学染色 (標識) を行った.
α-MEM で 20 倍に希釈した MAb OB7.3 (kindly provided by Prof. J. Klein-Nulend, ACTA
Frije University) を一次抗体,Alexa Fluor 546 goat anti-mouse IgG (Invitrogen) を二次抗
体としてそれぞれ用いることにより,骨細胞を特異的に蛍光標識した.顕微鏡での蛍
52
Page 58
光観察において,細胞に導入したカルシウムイオン蛍光指示薬 (後述) の蛍光が観察
された生細胞のうち,Alexa Fluor 546 (OB7.3) の蛍光が観察された細胞を Ocy,観察
されなかった細胞を BSC としてそれぞれ同定した.
免疫抗体法による蛍光標識を行った観察画像を,Fig. 4-1A に示す.画像上のすべて
の細胞において,別チャンネルにてカルシウムイオン蛍光指示薬の蛍光を観察してお
り,これらの細胞が生細胞であることを確認している.ここで,Fig. 4-1B に示すよう
に,力学刺激を与える細胞を視野の中央にとらえ,この細胞を Cell #0 としてラベル
し,周囲の細胞に対しても個別の番号を付けた.Fig. 4-1A 中の矢頭で示される細胞に
おいて,Alexa Fluor 546 の蛍光が確認できることから,これらの細胞を Ocy と同定し,
その他の生細胞を BSC とした.
53
Page 59
Fig. 4-1 Observation of the calcium signaling response to applied mechanical stimulus and its propagation
to NCs: (A) Immunolabeling of osteocytes. Mechanical stimulus was applied to Cell #0 Bar = 20
μm. (B) NCs numbered in increasing order of the distance Li. (C) Schematic illustration of
distance Li and minimum distance lijmin. (D) Cell types based on immunolabeling and judgment of
NCs based on distances, Li and lijmin.
54
Page 60
4.2.3 力学刺激の付与
単一の骨系細胞に力学刺激を付与するため,ガラス製マイクロニードルを用いて細
胞に対し直接変形を与えた.マイクロニードルの作成方法は第 3 章の方法に従った.
作製したマイクロニードルは,顕微鏡に取り付けた三次元液圧式マニピュレーター
(Narishige) に,ディッシュ底面との角度を 40°として固定した.マイクロニードルの
先端を,ディッシュ底面から 1 μm の高さで静止させ,その状態から細胞体の中心付
近まで v = 5.0 μm/s の速度で水平移動させることにより,細胞に直接力学刺激を付与
した.
4.2.4 近傍細胞の定義
カルシウム応答の細胞間伝播現象 [Newman, 2001; Yellowley et al., 2000] は,細胞間
のギャップ結合を経由した IP3 [Sanderson et al., 1994] の拡散や細胞外における ATP
[Jørgensen et al., 1997; Newman, 2001] 等の活性物質の拡散によって発生する.したが
って,これらの活性物質の伝播経路や,細胞間の距離,伝播する細胞の密度により,
直接刺激を受けた細胞から間接的な (二次的な) 細胞間伝播が生じることが考えられ
る.そこで本研究では,力学刺激の付与に対して発生した初期のカルシウム応答が近
傍する細胞に伝播する現象のみに着目して骨系細胞間のカルシウム応答伝播特性を
検討するため,力学刺激を付与した細胞に近接する細胞を観察および考察の対象とし
た.
骨系細胞の観察画像 (Fig. 4-1A) 上の細胞に対して,まず,力学刺激を付与した細
胞から Fig. 4-1B に示す周囲の各細胞 Cell #i (i = 1, n) までの細胞間距離 Li を求めた.
ここで,#i は,Liの値が小さい順に番号を付した.次に,各細胞 Cell #i について,他
の細胞 Cell #j (i ≠ j, j = 0, n) との細胞間距離 lijをそれぞれ求めた(Fig. 4-1C).ここで,
各細胞 Cell #i において,lijの 小値 lijmin が lij
min = li0 = Liとなるとき,すなわち 近接
細胞が Cell #0 のとき,Cell #i を NC と定義した.なお,細胞間距離 Li,lij は,位相差
顕微鏡による観察画像より,各細胞の細胞膜輪郭を抽出し,それらの間の 短距離を
計測して求めた.Fig. 4-1A の観察例においては,lijmin = Liとなった Cell #i は,Cell #1,
#2, #3, #4 であり,これらを近傍細胞 (Neighboring cell: NC) と定義し,本研究ではこ
55
Page 61
の NC のみを観察の対象とした (Fig. 4-1D).
4.2.5 細胞内カルシウムイオンの可視化とカルシウム応答の観察
本研究では,第 2 章,および,第 3 章の手法と同様に,細胞内に蛍光特性の異なる
2 種類のカルシウムイオン蛍光指示薬 Fluo-4 (Invitrogen),Fura Red (Invitrogen) を導入
することにより,細胞内カルシウムイオン (Ca2+) を可視化した.第 3 章と同様の条
件を用いて細胞内に試薬を導入し,蛍光顕微鏡観察において,検出された両試薬の蛍
光輝度値の比を取得することにより (Ratiometry 法),細胞内カルシウムイオン濃度
([Ca2+]i) の経時変化を観察した.本研究では,力学刺激を付与した細胞においてカル
シウム応答が発生した場合,NC の蛍光輝度比の変化について考察した.力学刺激を
付与した細胞におけるカルシウム応答の判断には,第 3 章と同様に Hung [Hung et al.,
1995] の基準を用いた.その際,NC においても同様の基準を用いてカルシウム応答
の発生が確認された場合,力学刺激を付与した細胞と NC 間においてカルシウム応答
の伝播が発生したと判断した.力学刺激を付与した細胞,および NC の細胞種の組み
合わせごとに,カルシウム応答の伝播発生率を求め,各群間に対して Fisher の正確確
率検定 (p < 0.01) による有意差検定を行った.
4.3 実験結果
4.3.1 カルシウム応答の細胞間伝播
力学刺激を付与した細胞,および,4.2.4 節で定義した NC を対象として,力学刺激
に対する細胞のカルシウム応答,および,周囲の細胞に対するカルシウム応答伝播を
観察した.結果を Fig. 4-2 に示す.Cell #0 に対する力学刺激の付与開始 (t = 0.0 s) か
ら 5.0 s 後,Cell #0 (Ocy)において,細胞内に導入した Fluo-4 (green)の蛍光輝度の上昇
および Fura Red (red) の蛍光輝度の下降がみられ,Fig. 4-2A に示すように,細胞内に
その変化が広がる様子が観察された (t = 3.5 s).また,力学刺激の付与開始から 10.0 s
後には,Cell #1,#3,#4 (いずれも BSC) において蛍光輝度の変化が観察された.一
方,Cell #2 (Ocy) においては蛍光輝度の変化は観察されなかった (Fig. 4-2A).
56
Page 62
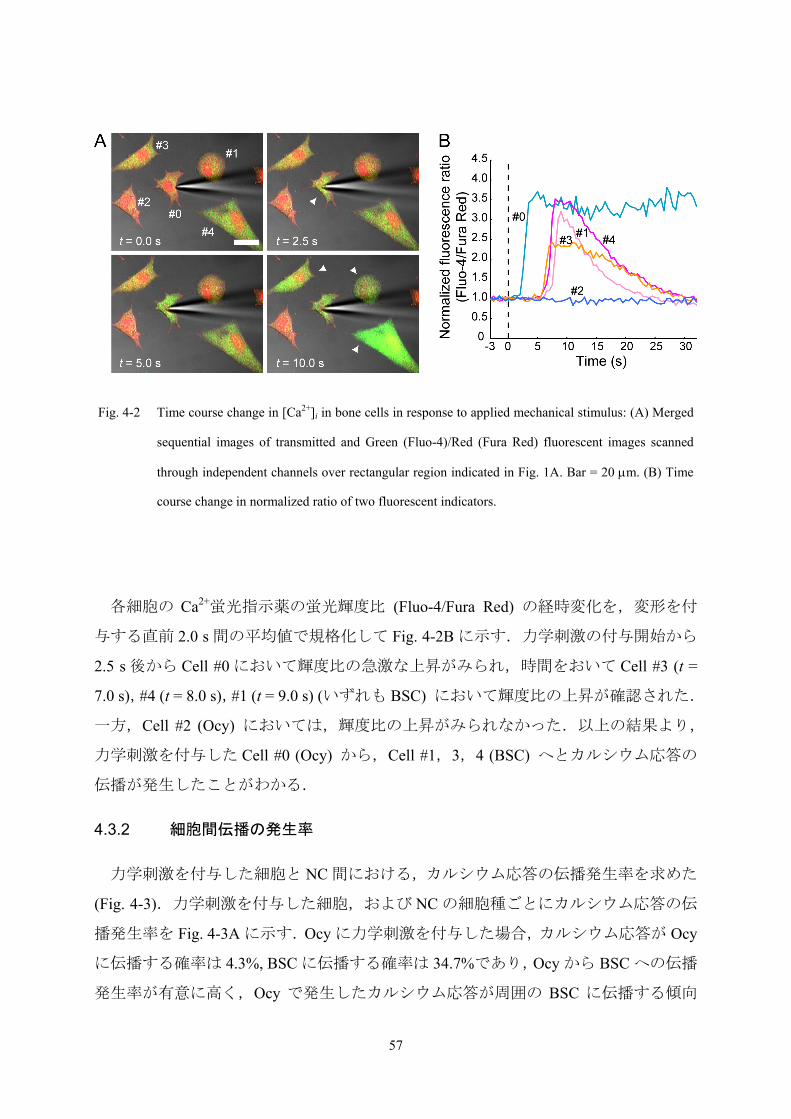
Fig. 4-2 Time course change in [Ca2+]i in bone cells in response to applied mechanical stimulus: (A) Merged
sequential images of transmitted and Green (Fluo-4)/Red (Fura Red) fluorescent images scanned
through independent channels over rectangular region indicated in Fig. 1A. Bar = 20 μm. (B) Time
course change in normalized ratio of two fluorescent indicators.
各細胞の Ca2+蛍光指示薬の蛍光輝度比 (Fluo-4/Fura Red) の経時変化を,変形を付
与する直前 2.0 s 間の平均値で規格化して Fig. 4-2B に示す.力学刺激の付与開始から
2.5 s 後から Cell #0 において輝度比の急激な上昇がみられ,時間をおいて Cell #3 (t =
7.0 s),#4 (t = 8.0 s),#1 (t = 9.0 s) (いずれも BSC) において輝度比の上昇が確認された.
一方,Cell #2 (Ocy) においては,輝度比の上昇がみられなかった.以上の結果より,
力学刺激を付与した Cell #0 (Ocy) から,Cell #1,3,4 (BSC) へとカルシウム応答の
伝播が発生したことがわかる.
4.3.2 細胞間伝播の発生率
力学刺激を付与した細胞と NC 間における,カルシウム応答の伝播発生率を求めた
(Fig. 4-3).力学刺激を付与した細胞,および NC の細胞種ごとにカルシウム応答の伝
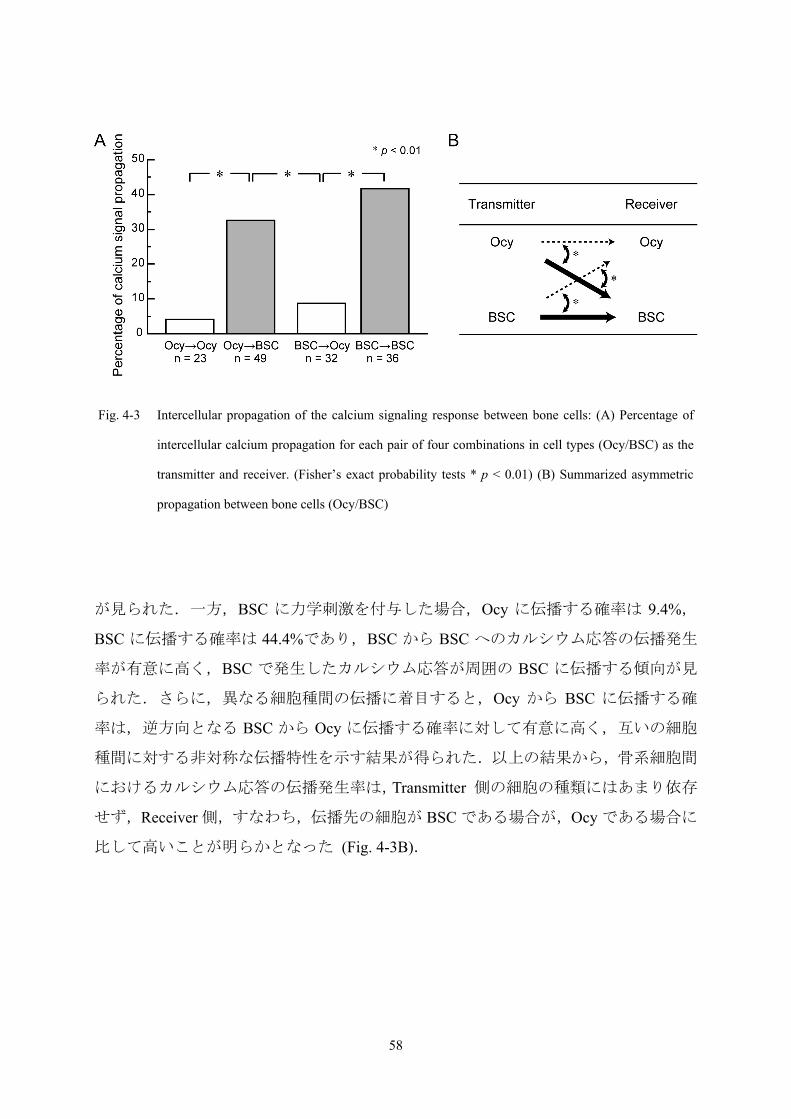
播発生率を Fig. 4-3A に示す.Ocy に力学刺激を付与した場合,カルシウム応答が Ocy
に伝播する確率は 4.3%, BSC に伝播する確率は 34.7%であり,Ocy から BSC への伝播
発生率が有意に高く,Ocy で発生したカルシウム応答が周囲の BSC に伝播する傾向
57
Page 63
Fig. 4-3 Intercellular propagation of the calcium signaling response between bone cells: (A) Percentage of
intercellular calcium propagation for each pair of four combinations in cell types (Ocy/BSC) as the
transmitter and receiver. (Fisher’s exact probability tests * p < 0.01) (B) Summarized asymmetric
propagation between bone cells (Ocy/BSC)
が見られた.一方,BSC に力学刺激を付与した場合,Ocy に伝播する確率は 9.4%,
BSC に伝播する確率は 44.4%であり,BSC から BSC へのカルシウム応答の伝播発生
率が有意に高く,BSC で発生したカルシウム応答が周囲の BSC に伝播する傾向が見
られた.さらに,異なる細胞種間の伝播に着目すると,Ocy から BSC に伝播する確
率は,逆方向となる BSC から Ocy に伝播する確率に対して有意に高く,互いの細胞
種間に対する非対称な伝播特性を示す結果が得られた.以上の結果から,骨系細胞間
におけるカルシウム応答の伝播発生率は,Transmitter 側の細胞の種類にはあまり依存
せず,Receiver 側,すなわち,伝播先の細胞が BSC である場合が,Ocy である場合に
比して高いことが明らかとなった (Fig. 4-3B).
58
Page 64
4.4 考 察
4.4.1 細胞種と細胞間伝播発生の関係 細胞種と細胞間伝播発生の関係
本研究において観察された,骨系細胞間におけるカルシウム応答の伝播現象におい
て,BSC に力学刺激を付与し発生したカルシウム応答が骨細胞に伝播する確率に対し
て,骨細胞に力学刺激を付与し発生したカルシウム応答が BSC に伝播する確率が,
有意に高いという結果が得られた.このことにより,骨表面に近い層に分布する BSC
と骨基質内部に存在する骨細胞間の細胞間伝播が,骨細胞から BSC に対して非対称
に,すなわち,骨基質内部から骨表面方向に向かって一方向的に行われている可能性
が示唆された (Fig. 4-4).
本研究において観察された,骨系細胞間におけるカルシウム応答の伝播現象におい
て,BSC に力学刺激を付与し発生したカルシウム応答が骨細胞に伝播する確率に対し
て,骨細胞に力学刺激を付与し発生したカルシウム応答が BSC に伝播する確率が,
有意に高いという結果が得られた.このことにより,骨表面に近い層に分布する BSC
と骨基質内部に存在する骨細胞間の細胞間伝播が,骨細胞から BSC に対して非対称
に,すなわち,骨基質内部から骨表面方向に向かって一方向的に行われている可能性
が示唆された (Fig. 4-4).
Fig. 4-4 Biased cellular communication in propagation of the calcium signaling response between
BSCs or from Ocy to BSC
59
Page 65
また,骨系細胞間におけるカルシウム応答の伝播は,力学刺激を付与した細胞が骨
細胞,BSC いずれの場合においても,伝播先が BSC である場合において,有意に高
い確率で発生した.このことから,骨系細胞におけるカルシウム応答の伝播特性が,
伝播先の細胞 (Receiver) の細胞種に大きく依存していることが考えられる.骨基質中
において骨細胞が形成する細胞間ネットワークは,骨への力学的負荷を刺激として感
知した骨細胞が,その情報を周囲の細胞へ伝達する経路として機能すると考えられて
いる [Yellowley et al., 2000].骨細胞間のカルシウム応答伝播の発生率が,BSC 間,お
よび骨細胞から BSC へのカルシウム応答伝播に比して低いことは,骨系細胞間の力
学刺激の受容伝達において,BSC 間,および骨細胞から BSC への細胞応答伝播が重
要な役割を果たしていることが考えられる.
4.4.2 骨組織中における細胞間コミュニケーション
骨組織中において,BSC は骨組織の表層に,骨細胞は骨基質内部に埋没してそれぞ
れ分布している.したがって,本研究で観察された異なる骨系細胞種間の応答に見ら
れる非対称性は,主に骨表面に近い層に分布する骨細胞において機能していると考え
ることができる.一方,骨基質内において骨細胞が感知する力学刺激のひとつと考え
られている,骨小腔‐骨細管の空隙を満たす間質液の流れは,骨梁に力学的負荷が作
用した場合,骨梁の中立軸付近よりも表層に近い領域ほど大きくなることが理論的に
示されている [Kameo et al., 2008].このことから,骨組織表面層の骨系細胞は,骨の
リモデリングにおける骨細胞の力学刺激感知と,骨表面の骨芽細胞への刺激情報伝達
の過程に適した分布をとっていることが示唆される.
本研究では,力学刺激の付与に対するカルシウム応答の細胞間伝播を in vitro 環境
で観察した.実際の骨組織においては,骨細胞は骨小腔‐骨細管の空隙に三次元的に
分布しており,細胞の分布状態および細胞がおかれる微小環境は,本研究における実
験環境とは大きく異なる.このことは,本研究の in vitro 環境における実験と,実際
の骨組織中 (in vivo 環境) において,細胞間の物質移動や,それにともなう細胞応答
特性が異なる可能性を示唆している.本研究で観察対象としたカルシウム応答の細胞
間伝播は,刺激を付与した細胞からの活性物質 (IP3,ATP など) の拡散により発生す
る [Jørgensen et al., 1997; Newman, 2001].細胞周囲の空間 (peripheral space) が限定さ
60
Page 66
れている骨組織内の in vivo 環境下における蛍光色素の拡散は,in vitro 環境下の 62%
に制限されるという実験値が報告されている [Wang L.Y. et al., 2005] ことから,in
vivo 環境下でのカルシウム応答を誘起する伝達物質の拡散,および細胞間伝播特性も
また,本研究で得られた結果と比較して応答発生率,応答発生時間が大きく変化する
ことが考えられる.そのため,これらを考慮し得る新たな実験系の構築が望まれる.
in vitro 環境において in vivo の骨系細胞の分布状態を二次元的に再現する 1 つの方
法として,マイクロパターニング [Goffin et al., 2006] を用いた細胞間ネットワークの
制御 [Guo et al., 2006] が挙げられる.培養基盤に細胞の接着領域と非接着領域を配置
することにより,任意の細胞間距離を持つ二次元的な細胞間ネットワークを作成する
ことができる.このほか,フィルタを挟んで BSC と骨細胞を共培養したモデル [Taylor
et al., 2007] を用いることにより,骨表面に近い層の骨系細胞の分布と細胞間ネットワ
ークを擬似的に再現した実験系の構築が可能になると考える.
カルシウム応答の細胞間伝播には,細胞間のギャップ結合を経由した IP3 の拡散や
細胞外における ATP 等の活性物質の拡散など,複数の機序が存在する [Jørgensen et
al., 2000].力学刺激を付与した細胞で発生したカルシウム応答が接触する近くの細胞
に伝播する場合,ギャップ結合などの細胞間の連絡構造を介して伝達物質が輸送され,
Receiver 側の細胞内 Ca2+ストアからの放出や細胞外からの Ca2+流入を誘起するメカニ
ズム [Jiang et al., 2007] が考えられる.一方,本研究では,力学刺激を付与した細胞
との接触がみられない,細胞膜間が離れている細胞においてもカルシウム応答伝播が
観察された.このような離れた細胞間のカルシウム応答伝播は,伝達物質としての
ATP の拡散 [Newman, 2001] によると考えられる.本研究で観察された,カルシウム
応答の細胞間伝播は,これら複数の細胞間伝播機序を区別せずに示している.細胞の
ATP 受容特性は細胞種によって異なることが考えられることから,細胞間伝播の各機
序における伝達経路を阻害したうえで,BSC 間,骨細胞間,BSC と骨細胞間の伝播の
発生率を比較することにより,伝播特性のより詳細な検討が可能になると考える.
61
Page 67
62
4.5 結 言
本章では,力学刺激を感知した骨細胞が刺激情報を周囲の細胞に伝達する特性に関
して,骨細胞が形成する細胞間ネットワーク構造を介した細胞間コミュニケーション
の機能を検討した.骨組織中に存在する骨細胞と,骨組織表層において骨代謝に関わ
る骨表面細胞間に発生するカルシウム応答の伝播は,力学刺激を付与する細胞によら
ず,骨表面細胞に伝播する場合において高い確率で発生した.この結果より,骨系細
胞間の細胞応答の伝播現象は,骨細胞から骨表面細胞に対して,すなわち,骨組織の
内部から骨組織表面層に向かって一方向的に行われている可能性が示唆された.
Page 68
第 5 章 結 論
骨基質中に埋没した形で存在する骨細胞は,骨への力学的負荷を刺激として感知し,
骨基質内部で形成する細胞間ネットワークを介して,周囲の細胞に刺激情報を伝達す
るメカノセンサーとしての役割を果たすと考えられている.力学的適応にともなう骨
リモデリング過程においては,この骨細胞の力学刺激感知・伝達特性が,骨組織の代
謝を行う骨系細胞群の活動を調節することにより,骨の組織としての恒常性が維持さ
れていると考えられる.骨細胞の力学刺激に対する応答特性を検討することは,骨リ
モデリングのメカニズムの詳細の解明に加えて,医工学分野への応用における基礎的
知見となり得る点で重要である.
これまでに,骨細胞への力学刺激応答特性の実験的検討として,骨細胞の機能に影
響を与え得る力学刺激の形態や,それらの力学刺激の付与に対する骨代謝調節因子の
産生が報告され,骨細胞のおかれる力学的環境と,他の骨系細胞への調節作用との関
係が明らかにされてきた.このような骨細胞と他の骨系細胞の生化学因子を介した相
互作用を実現させているのは,カルシウム応答をはじめとした,力学刺激を生化学シ
グナルに変換するメカノトランスダクション機構である.したがって,力学的適応に
よる骨リモデリングにおける骨細胞の力学応答特性を解明するためには,骨基質にお
ける骨細胞の力学刺激感知・伝達メカニズムを,細胞レベルの応答現象の観点から検
討する必要がある.
63
Page 69
本論文では,骨基質中に存在する骨細胞の力学刺激の感知・伝達特性に着目し,骨
細胞の力学刺激応答特性,および,細胞間ネットワーク構造が,骨リモデリング現象
に果たす役割を明らかにすることを目指して,骨細胞への力学刺激に対する初期応答
としてのカルシウム応答挙動を実験的に検討した.
第 2 章では,骨基質内骨細胞の力学応答特性を検討するため,骨組織に対する変形
の付与と,それに対する組織内骨細胞の生化学的応答の経時的観察を可能とする ex
vivo 実験系を確立し,骨組織と骨細胞の変形計測,および,それにともなう組織内骨
細胞のカルシウム応答を観察した.本研究で確立した実験系においては,ニワトリ胚
より採取した頭蓋冠より作製した骨基質試料に対し,2 本のガラス製マイクロニード
ルを用いた面内せん断変形を付与する方法を用いることにより,骨基質内の骨細胞へ
の力学実験を可能とした.骨基質と細胞の変形は,変形付与前後の骨基質の微分干渉
画像と骨細胞の細胞膜を免疫標識した蛍光観察画像を用いて,画像相関法により定量
的に評価した.また,骨基質への変形付与中に,カルシウムイオン蛍光指示薬を導入
した骨細胞の経時的観察を行った.その結果,骨組織への変形付与にともなう骨組織
内骨細胞の変形,および,カルシウム応答の発生を初めて観察することに成功した.
この結果より,骨組織への力学的負荷に対し,組織内骨細胞において力学刺激感知機
構が機能していることを確認した.
第 3 章では,第 2 章の検討で観察された骨組織内骨細胞の力学刺激感知特性に関し
て,骨細胞が骨への力学的負荷を刺激として感知するメカニズムの in vitro 検討を行
った.特に,組織内骨細胞の力学刺激感知特性において骨細胞の特徴的な細胞形態が
果たす役割に着目し,骨細胞の局所部位における力学刺激感知特性を検討した.ニワ
トリ胚頭蓋冠より単離した単一の骨細胞に対して,定量的に制御した力学刺激を付与
する in vitro 実験系を確立し,細胞応答としてのカルシウム応答を観察することによ
り,骨細胞の局所的な力学刺激感知特性を検討した.ここでは,モノクローナル抗体
OB7.3 で修飾したマイクロパーティクルを骨細胞の細胞膜に付着させ,ガラス製マイ
クロニードルを用いてマイクロパーティクルを移動させることにより,骨細胞に局所
的な力学刺激を付与した.その結果,力学刺激の付与に対し,カルシウム応答が力学
刺激の付与部位の近傍から発生し,その後,細胞全体に波及する様子が観察された.
また,細胞突起においては,細胞体に比べて,高い確率,かつ,より小さい刺激量に
64
Page 70
よってカルシウム応答が発生した.このことは,骨細胞の細胞突起における鋭敏な力
学刺激感知特性を示すものであり,骨細胞の力学刺激応答メカニズムに対する細胞形
態と内部の細胞骨格構造の寄与を示すものである.
第 4 章では,第 3 章の検討において観察された力学刺激の付与に対する骨細胞の応
答が,周囲の細胞に伝達される特性,すなわち,骨細胞が形成する細胞間ネットワー
ク構造を介した細胞間コミュニケーションの機能を検討した.ニワトリ胚頭蓋冠より
単離・培養した骨細胞を含む骨系細胞にガラス製マイクロニードルを用いて力学刺激
を付与し,発生したカルシウム応答が周囲の細胞に伝播する様子を観察した.その結
果,骨組織中に存在する骨細胞と,骨組織表層において骨代謝に関わる骨表面細胞間
に発生するカルシウム応答の伝播は,力学刺激を付与した細胞種を問わず,骨表面細
胞に対して高い確率で伝播した.このことは,骨系細胞間の細胞応答の伝播現象が,
骨細胞から骨表面細胞に対して,すなわち,骨組織の内部から骨組織表面層に向かっ
て一方向的に行われている可能性を示しており,骨リモデリングにおける骨細胞の指
向性のある力学刺激伝達特性の存在が示唆された.
以上のように,本論文では,ex vivo / in vitro 実験系を用いた検討により,骨組織内
骨細胞の力学刺激感知特性,および,骨細胞の力学刺激に対する細胞形態に依存した
力学刺激感知特性を明らかにするとともに,骨細胞と他の骨系細胞間における力学刺
激伝達において非対称的な細胞間コミュニケーションが機能していることを示した.
本論文における,バイオメカニクスの手法を用いた骨細胞の力学刺激感知・伝達特
性の実験的検討,および,評価は,骨組織内骨細胞の力学刺激に対する応答の細胞レ
ベルでの素過程を示すものである.骨細胞の力学応答特性の検討に関しては,これま
でに,骨組織内において骨細胞に作用する力としてさまざまな力学刺激の形態が提案
され,それらの力学刺激に対する骨細胞の応答が in vitro 実験系を用いた検討により
報告されている.しかしながら,骨細胞が骨組織への力学的負荷を刺激として感知し,
他の骨系細胞に伝達することにより骨リモデリングを調節するメカノセンサーとし
て機能する,その一連のメカニズムについて,実験的手法により検討された報告はな
かった.本論文で示した結果は,これらの報告にみられる骨組織内骨細胞への力学的
負荷が,骨細胞の骨代謝調節機能を発現させるためのメカノトランスダクション機構,
ならびに,骨系細胞に対する刺激情報の伝達機構に関する新たな知見を提供するもの
65
Page 71
66
である.
骨の力学的適応現象に対する理解を,医工学分野における将来的な技術的応用につ
なげるためには,骨リモデリングに関与する各骨系細胞間の相互的な制御メカニズム
を解明するための分子生物学的アプローチに加えて,骨系細胞の生理的反応の場とな
る骨組織の力学的環境条件の解明が重要となる.本論文で得られた結果を踏まえて,
骨組織内骨細胞の力学刺激応答能と骨基質内部の力学状態の解明につなげるために
は,ex vivo 環境下,または,骨組織内骨細胞の三次元的分布と細胞間ネットワークを
再現した in vitro 環境下における実験的検討が望まれる.
さらに,細胞を生命活動の機能単位として扱う場合,本論文で得られた,単一の骨
細胞の力学刺激感知機構,および,骨系細胞間における刺激情報の伝達機構に関する
知見は,骨代謝現象のメカニズム解明にとどまらず,メカノバイオロジーの観点から
の多細胞生物における細胞の秩序ある生命活動の理解に寄与するものであると考え
る.本論文における一連の実験的検討は,細胞内におけるシグナル伝達,遺伝子発現,
代謝などの諸現象が,同種・異種の細胞間で組織化され,高次組織の形成に関与する
メカニズムの理解を目指すための手法,および,骨の力学的適応能に見られるような,
細胞と力学的環境の相互作用が組織形成において機能するメカニズムの理解を目指
すための手法として普遍化することができる.本論文における実験的検討の枠組みを,
細胞種,組織によらない形で体系化することにより,生体を形成する階層構造を考慮
したシステムバイオメカニクスの基礎的知見につながることが期待される.
Page 72
参考文献
Aarden EM, Burger EH, Nijweide PJ: Function of osteocyte in bone, Journal of Cellular
Biochemistry, 55-3, pp. 287-299 (1994).
Adachi T, Sato K, Higashi N, Tomita Y, Hojo M: Simultaneous observation of calcium
signaling response and membrane deformation due to localized mechanical stimulus in single
osteoblast-like cells, Journal of the Mechanical Behavior of Biomedical Materials, 1-1, pp.
43-50 (2008).
Ajubi NE, Klein-Nulend J, Alblas MJ, Burger EH, Nijweide PJ: Signal transduction pathways
involved in fluid flow-induced PGE2 production by cultured osteocytes, The American
Journal of Physiology, 276-1 Pt 1, pp. E171-178 (1999).
Anderson EJ, Kaliyamoorthy S, Iwan J, Alexander D, Knothe Tate ML: Nano-microscale
models of periosteocytic flow show differences in stresses imparted to cell body and
processes, Annals of biomedical engineering, 33-1, pp. 52-62 (2005).
Anderson EJ, Knothe Tate ML: Idealization of pericellular fluid space geometry and
dimension results in a profound underprediction of nano-microscale stresses imparted by fluid
drag on osteocytes, Journal of Biomechanics, 41-8, pp. 1736-1746 (2008).
Berridge MJ, Bootman MD, Lipp P: Calcium - a life and death signal, Nature, 395-6703, pp.
645-648 (1998).
Bonewald LF: Generation and function of osteocyte dendritic processes, Journal of
Musculoskeletal & Neuronal Interactions, 5-4, pp. 321-324 (2005).
67
Page 73
Bonewald LF: Mechanosensation and transduction in osteocytes, Bonekey Osteovision, 3-10,
pp. 7-15 (2006).
Bonivtch AR, Bonewald LF, Nicolella DP: Tissue strain amplification at the osteocyte lacuna:
a microstructural finite element analysis, Journal of Biomechanics, 40-10, pp. 2199-2206
(2007).
Burger EH, Klein-Nulend J: Mechanotransduction in bone - role of the lacuno-canalicular
network, FASEB Journal, 13 Suppl, pp. S101-112 (1999).
Chen NX, Ryder KD, Pavalko FM, Turner CH, Burr DB, Qiu J, Duncan RL: Ca2+ regulates
fluid shear-induced cytoskeletal reorganization and gene expression in osteoblasts, American
Journal of Physiology. Cell Physiology, 278-5, pp. C989-997 (2000).
Cowin SC: How is a tissue built?, Journal of Biomechanical Engineering, 122-6, pp. 553-569
(2000).
Cowin SC, Moss-Salentijn L, Moss ML: Candidates for the mechanosensory system in bone,
Journal of Biomechanical Engineering, 113-2, pp. 191-197 (1991).
Donahue HJ: Gap junctions and biophysical regulation of bone cell differentiation, Bone,
26-5, pp. 417-422 (2000).
Einhorn TA: Biomechanics of bone, in: Bilezikian JP, Raisz LG, Rodan GA (Eds.), Principle
of Bone Biology, Academic Press, (1996).
Felig P, Frohman LA (Eds), Endocrinology and metabolism, 4th edition, McGraw-Hill (2001).
Franz-Odendaal TA, Hall BK, Witten PE: Buried alive: how osteoblasts become osteocytes,
Developmental Dynamics, 235-1, pp. 176-190 (2006).
68
Page 74
Goffin JM, Pittet P, Csucs G, Lussi JW, Meister JJ, Hinz B: Focal adhesion size controls
tension-dependent recruitment of alpha-smooth muscle actin to stress fibers, The Journal of
Cell Biology, 172-2, pp. 259-268 (2006).
Greenleaf JE: Physiological responses to prolonged bed rest and fluid immersion in humans,
Journal of applied physiology: respiratory, environmental and exercise physiology, 57-3, pp.
619-633 (1984).
Grynkiewicz G, Poenie M, Tsien RY: A new generation of Ca2+ indicators with greatly
improved fluorescence properties, J Biol Chem, 260-6, pp. 3440-3450 (1985).
Gu G, Nars M, Hentunen TA, Metsikko K, Väänänen HK: Isolated primary osteocytes express
functional gap junctions in vitro, Cell and Tissue Research, 323-2, pp. 263-271 (2006).
Guo XE, Takai E, Jiang X, Xu Q, Whitesides GM, Yardley JT, Hung CT, Chow EM,
Hantschel T, Costa KD: Intracellular calcium waves in bone cell networks under single cell
nanoindentation, Molecular & cellular biomechanics, 3-3, pp. 95-107 (2006).
Han Y, Cowin SC, Schaffler MB, Weinbaum S: Mechanotransduction and strain amplification
in osteocyte cell processes, Proceedings of the National Academy of Sciences of the United
States of America, 101-47, pp. 16689-16694 (2004).
Hayakawa K, Tatsumi H, Sokabe M: Actin stress fibers transmit and focus force to activate
mechanosensitive channels, Journal of Cell Science, 121-Pt 4, pp. 496-503 (2008).
Hefley TJ: Utilization of FPLC-purified bacterial collagenase for the isolation of cells from
bone, Journal of Bone and Mineral Research, 2-6, pp. 505-516 (1987).
Hung CT, Pollack SR, Reilly TM, Brighton CT: Real-time calcium response of cultured bone
cells to fluid flow, Clincal Orthopaedics and Related Research, -313, pp. 256-269 (1995).
69
Page 75
Ingber D: Integrins as mechanochemical transducers, Curr Opin Cell Biol, 3-5, pp. 841-848
(1991).
Ishihara Y, Kamioka H, Honjo T, Ueda H, Takano-Yamamoto T, Yamashiro T: Hormonal, pH,
and calcium regulation of Connexin 43-mediated dye transfer in osteocytes in chick calvaria,
Journal of Bone and Mineral Research, 23-3, pp. 350-360 (2008).
Jørgensen NR, Geist ST, Civitelli R, Steinberg TH: ATP- and gap junction-dependent
intercellular calcium signaling in osteoblastic cells, The Journal of Cell Biology, 139-2, pp.
497-506 (1997).
Jørgensen NR, Henriksen Z, Brot C, Eriksen EF, Sørensen OH, Civitelli R, Steinberg TH:
Human osteoblastic cells propagate intercellular calcium signals by two different mechanisms,
Journal of Bone and Mineral Research, 15-6, pp. 1024-1032 (2000).
Jacobs CR, Yellowley CE, Davis BR, Zhou Z, Cimbala JM, Donahue HJ: Differential effect
of steady versus oscillating flow on bone cells, Journal of Biomechanics, 31-11, pp. 969-976
(1998).
Jiang JX, Siller-Jackson AJ, Burra S: Roles of gap junctions and hemichannels in bone cell
functions and in signal transmission of mechanical stress, Frontiers in Bioscience, 12, pp.
1450-1462 (2007).
Kameo Y, Adachi T, Hojo M: Transient response of fluid pressure in a poroelastic material
under uniaxial cyclic loading, Journal of the mechanics and physics of solids, 56-5, pp.
1794-1805 (2008).
Kamioka H, Honjo T, Takano-Yamamoto T: A three-dimensional distribution of osteocyte
processes revealed by the combination of confocal laser scanning microscopy and differential
interference contrast microscopy, Bone, 28-2, pp. 145-149 (2001).
70
Page 76
Kamioka H, Sugawara Y, Honjo T, Yamashiro T, Takano-Yamamoto T: Terminal
differentiation of osteoblasts to osteocytes is accompanied by dramatic changes in the
distribution of actin-binding proteins, Journal of Bone and Mineral Research, 19-3, pp.
471-478 (2004).
Kamioka H, Sugawara Y, Murshid SA, Ishihara Y, Honjo T, Takano-Yamamoto T: Fluid shear
stress induces less calcium response in a single primary osteocyte than in a single osteoblast:
implication of different focal adhesion formation, J Bone Miner Res, 21-7, pp. 1012-1021
(2006).
Kato Y, Boskey A, Spevak L, Dallas M, Hori M, Bonewald LF: Establishment of an osteoid
preosteocyte-like cell MLO-A5 that spontaneously mineralizes in culture, Journal of Bone
and Mineral Research, 16-9, pp. 1622-1633 (2001).
Kato Y, Windle JJ, Koop BA, Mundy GR, Bonewald LF: Establishment of an osteocyte-like
cell line, MLO-Y4, Journal Bone and Mineral Research, 12-12, pp. 2014-2023 (1997).
Klein-Nulend J, Semeins CM, Ajubi NE, Nijweide PJ, Burger EH: Pulsating fluid flow
increases nitric oxide (NO) synthesis by osteocytes but not periosteal fibroblasts--correlation
with prostaglandin upregulation, Biochemical and Biophysical Research Communication,
217-2, pp. 640-648 (1995a).
Klein-Nulend J, van der Plas A, Semeins CM, Ajubi NE, Frangos JA, Nijweide PJ, Burger
EH: Sensitivity of osteocytes to biomechanical stress in vitro, FASEB Journal, 9-5, pp.
441-445 (1995b).
Kurata K, Fukunaga T, Matsuda J, Higaki H: Role of mechanically damaged osteocytes in the
initial phase of bone remodeling, International Journal of Fatigue, 29-6, pp. 1010-1018
(2007).
71
Page 77
Kurata K, Heino TJ, Higaki H, Väänänen HK: Bone marrow cell differentiation induced by
mechanically damaged osteocytes in 3D gel-embedded culture, Journal of Bone and Mineral
Research, 21-4, pp. 616-625 (2006).
Lin JT, Lane JM: Rehabilitation of the older adult with an osteoporosis-related fracture,
Clinics in Geriatric Medicine, 22-2, pp. 435-447 (2006).
Mann V, Huber C, Kogianni G, Jones D, Noble B: The influence of mechanical stimulation on
osteocyte apoptosis and bone viability in human trabecular bone, Journal of Musculoskeletal
& Neuronal Interactions, 6-4, pp. 408-417 (2006).
Mikuni-Takagaki Y, Kakai Y, Satoyoshi M, Kawano E, Suzuki Y, Kawase T, Saito S: Matrix
mineralization and the differentiation of osteocyte-like cells in culture, Journal of Bone and
Mineral Research, 10-2, pp. 231-242 (1995).
Mishra S, Knothe Tate ML: Effect of lacunocanalicular architecture on hydraulic conductance
in bone tissue: Implications for bone health and evolution, The Anatomical Record. Part A,
Discoveries in Molecular, Cellular and Evolutionaly Biology, 273-2, pp. 752-762 (2003).
Newman EA: Propagation of intercellular calcium waves in retinal astrocytes and Muller cells,
The Journal of Neuroscience, 21-7, pp. 2215-2223 (2001).
Nicogossian AE, Sawin CF, Huntoon CL: Overall physiologic response to space flight, in:
Nicogossian AE, Huntoon CL, Pool SL (Eds.), Space physiology and medicine, 3rd edition,
Lea & Febiger, (1994).
Nicolella DP, Moravits DE, Gale AM, Bonewald LF, Lankford J: Osteocyte lacunae tissue
strain in cortical bone, Journal of Biomechanics, 39-9, pp. 1735-1743 (2006).
Nijweide PJ, Burger EH, Klein-Nulend J, van der Plas A: The osteocyte, in: Bilezikian JP,
Raisz LG, Rodan GA (Eds.), Principles of Bone Biology, Academic Press, (1996).
72
Page 78
Nijweide PJ, Mulder RJ: Identification of osteocytes in osteoblast-like cell cultures using a
monoclonal antibody specifically directed against osteocytes, Histochemistry, 84-4-6, pp.
342-347 (1986).
Parfitt AM: Osteonal and hemi-osteonal remodeling: the spatial and temporal framework for
signal traffic in adult human bone, Journal of Cellular Biochemistry, 55-3, pp. 273-286
(1994).
Parfitt AM: The mechanism of coupling: a role for the vasculature, Bone, 26-4, pp. 319-323
(2000).
Raisz LG, Pilbeam CC, Fall PM: Prostaglandins - Mechanisms of action and Reguration of
production in bone, Osteoporosis International, 3, pp. S136-S140 (1993).
Ren Y, Maltha JC, Kuijpers-Jagtman AM: Optimum force magnitude for orthodontic tooth
movement: a systematic literature review, The Angle Orthodontist, 73-1, pp. 86-92 (2003).
Sanderson MJ, Charles AC, Boitano S, Dirksen ER: Mechanisms and function of intercellular
calcium signaling, Molecular and Cellular Endocrinology, 98-2, pp. 173-187 (1994).
Sato K, Adachi T, Ueda D, Hojo M, Tomita Y: Measurement of local strain on cell membrane
at initiation point of calcium signaling response to applied mechanical stimulus in osteoblastic
cells, Journal of Biomechanics, 40-6, pp. 1246-1255 (2007).
Takai E, Mauck RL, Hung CT, Guo XE: Osteocyte viability and regulation of osteoblast
function in a 3D trabecular bone explant under dynamic hydrostatic pressure, Journal of Bone
and Mineral Research, 19-9, pp. 1403-1410 (2004).
Tanaka-Kamioka K, Kamioka H, Ris H, Lim SS: Osteocyte shape is dependent on actin
filaments and osteocyte processes are unique actin-rich projections, Journal of Bone and
Mineral Research, 13-10, pp. 1555-1568 (1998).
73
Page 79
Tatsumi S, Ishii K, Amizuka N, Li M, Kobayashi T, Kohno K, Ito M, Takeshita S, Ikeda K:
Targeted ablation of osteocytes induces osteoporosis with defective mechanotransduction,
Cell Metabolism, 5-6, pp. 464-475 (2007).
Taylor AF, Saunders MM, Shingle DL, Cimbala JM, Zhou Z, Donahue HJ: Mechanically
stimulated osteocytes regulate osteoblastic activity via gap junctions, American Journal of
Physiology, Cell Physiology, 292-1, pp. C545-552 (2007).
van't Hof RJ, Ralston SH: Nitric oxide and bone, Immunology, 103-3, pp. 255-261 (2001).
van der Plas A, Nijweide PJ: Isolation and purification of osteocytes, Journal of Bone and
Mineral Research, 7-4, pp. 389-396 (1992).
Vatsa A, Mizuno D, Smit TH, Schmidt CF, MacKintosh FC, Klein-Nulend J: Bio imaging of
intracellular NO production in single bone cells after mechanical stimulation, Journal of Bone
and Mineral Research, 21-11, pp. 1722-1728 (2006).
Vatsa A, Semeins CM, Smit TH, Klein-Nulend J: Paxillin localisation in osteocytes-Is it
determined by the direction of loading?, Biochemical and Biophysical Research
Communications, pp. (2008).
Vatsa A, Smit TH, Klein-Nulend J: Extracellular NO signalling from a mechanically
stimulated osteocyte, Journal of Biomechanics, 40 Suppl 1, pp. S89-95 (2007).
Wang LY, Wang YL, Han YF, Henderson SC, Majeska RJ, Weinbaum S, Schaffler MB: In situ
measurement of solute transport in the bone lacunar-canalicular system, Proceedings of the
National Academy of Sciences of the United States of America, 102-33, pp. 11911-11916
(2005).
Wang Y, McNamara LM, Schaffler MB, Weinbaum S: A model for the role of integrins in
flow induced mechanotransduction in osteocytes, Proceedings of the National Academy of
74
Page 80
Sciences of the United States of America, 104-40, pp. 15941-15946 (2007).
Weinbaum S, Cowin SC, Zeng Y: A model for the excitation of osteocytes by mechanical
loading-induced bone fluid shear stresses, Journal of Biomechanics, 27-3, pp. 339-360 (1994).
Westbroek I, De Rooij KE, Nijweide PJ: Osteocyte-specific monoclonal antibody MAb
OB7.3 is directed against Phex protein, Journal of Bone and Mineral Research, 17-5, pp.
845-853 (2002).
Wolff J (Eds), Das Gesetz der Transformation der Knochen, Hirschwald (1892).
Yellowley CE, Li Z, Zhou Z, Jacobs CR, Donahue HJ: Functional gap junctions between
osteocytic and osteoblastic cells, Journal of Bone and Mineral Research, 15-2, pp. 209-217
(2000).
You J, Reilly GC, Zhen X, Yellowley CE, Chen Q, Donahue HJ, Jacobs CR: Osteopontin gene
regulation by oscillatory fluid flow via intracellular calcium mobilization and activation of
mitogen-activated protein kinase in MC3T3-E1 osteoblasts, Journal of Biological Chemistry,
276-16, pp. 13365-13371 (2001a).
You J, Yellowley CE, Donahue HJ, Zhang Y, Chen Q, Jacobs CR: Substrate deformation
levels associated with routine physical activity are less stimulatory to bone cells relative to
loading-induced oscillatory fluid flow, Journal of Biomechanical Engineering, 122-4, pp.
387-393 (2000).
You L, Cowin SC, Schaffler MB, Weinbaum S: A model for strain amplification in the actin
cytoskeleton of osteocytes due to fluid drag on pericellular matrix, Journal of Biomechanics,
34-11, pp. 1375-1386 (2001b).
You L, Weinbaum S, Cowin SC, Schaffler MB: Ultrastructure of the osteocyte process and its
pericellular matrix, The Anatomical Record. Part A, Discoveries in Molecular, Cellular, and
75
Page 81
76
Evolutionary Biology, 278-2, pp. 505-513 (2004).
青沼有紀: 局所変形負荷に対する単離骨細胞のシグナル応答, 神戸大学大学院修士学
位論文 (2007).
荒木峻, 沼田眞, 和田攻 編, 環境科学辞典, 東京化学同人 (1985).
伊藤慎一, 田中基嗣, 安達泰治, 上岡寛, 山本照子, 北條正樹: 骨基質内における骨細
胞の変形および細胞応答の観察, 日本機械学会第 22 回バイオエンジニアリング講演
会講演論文集, 06-55, pp. 102-103 (2007).
大島博, 向井千秋: 有人飛行とベッドレスト研究における骨代謝, Clinical Calcium,
18-9, pp. 1245-1253 (2008).
小林敏雄, 佐賀徹雄, 植村知正: PIV 画像処理による流速計測: 可視化情報学会 編,
PIV と画像解析技術, 朝倉書店, (2004).
須田立雄, 小澤英浩, 高橋榮明, 田中栄, 中村浩彰, 森諭史 編, 新 骨の科学, 医歯薬
出版 (2007).
林紘三郎, 安達泰治, 宮崎浩: 骨のリモデリングのバイオメカニクス: 日本エム・イー
学会 編, 生体組織・細胞のリモデリングのバイオメカニクス, コロナ社, (2003).
山本照子: 歯科矯正とメカニカルストレス, Clinical Calcium, 18-9, pp. 1254-1263 (2008).
Page 82
関連学術論文・学術講演
A. 学術論文
A-1. Taiji Adachi, Yuki Aonuma, Shin-ichi Ito, Mototsugu Tanaka, Masaki Hojo,
Teruko Takano-Yamamoto, Hiroshi Kamioka
Osteocyte calcium signaling response to bone matrix deformation
Journal of Biomechanics No. 42, Vol. 15, pp. 2507-2512 (2009) (第 2 章)
A-2. Taiji Adachi, Yuki Aonuma, Mototsugu Tanaka, Masaki Hojo, Teruko Takano-Yamamoto,
Hiroshi Kamioka
Calcium response in single osteocytes to locally applied mechanical stimulus: Differences in
cell process and cell body
Journal of Biomechanics No. 42, Vol. 12, pp. 1989-1995 (2009). (第 3 章)
A-3. Taiji Adachi, Yuki Aonuma, Keisuke Taira, Masaki Hojo, Hiroshi Kamioka
Asymmetrical intercellular communication among bone cells: Propagation of calcium
signaling response
Biochemical and Biophysical Research Communications No. 389, Vol. 3, pp. 495-500 (2009).
(第 4 章)
B. Proceedings
B-1. Mototsugu Tanaka, Yuki Aonuma, Taiji Adachi, Hiroshi Kamioka, Teruko Takano-Yamamoto,
Masaki Hojo
Observation of Calcium Response of Isolated Osteocyte to Localized Mechanical Stimulus
The 12th International Conference on Biomedical Engineering, Proceedings Vol. 12, 4 pages,
(Singapore), (2005.12.7-10).
77
Page 83
B-2. Yuki Aonuma, Taiji Adachi, Mototsugu Tanaka, Masaki Hojo, Teruko Takano-Yamamoto,
Hiroshi Kamioka
Site-dependence of mechanosensitivity in isolated osteocytes
The 13th International Conference on Biomedical Engineering, Proceedings Vol. 13, pp.
2000–2004 (Singapore), (2008.12.3-6).
C. Abstract(国際学会)
C-1. Mototsugu Tanaka, Yuki Aonuma, Taiji Adachi, Hiroshi Kamioka, Teruko Takano-Yamamoto,
Masaki Hojo
Calcium response in isolated chick osteocytes and osteoblasts to direct deformation
The 5th World Congress of Biomechanics, Journal of Biomechanics, Vol. 39, Supplement 1,
pp. S448-449, (Munich), (2006.7.31-8.4).
C-2. Yuki Aonuma, Taiji Adachi, Mototsugu Tanaka, Masaki Hojo, Teruko Takano-Yamamoto,
Hiroshi Kamioka
Mechanosensitivity of a single osteocyte: Difference in cell process and cell body
Proceedings of the Third Asian Pacific Conference on Biomechanics, Journal of
Biomechanical Science and Engineering, Vol. 2, Suppl. 1, pp. S165 (Tokyo), (2007.11.5-8).
C-3. Taiji Adachi, Yuki Aonuma, Shin-ichi Ito, Mototsugu Tanaka, Masaki Hojo,
Teruko Takano-Yamamoto, Hiroshi Kamioka
Observation of calcium signaling response of osteocytes to bone matrix deformation
Third Switzerland-Japan workshop on Biomechanics 2009 (Switzerland), (2009.9.1-4).
78
Page 84
D. 学術講演(国内学会)
D-1. 青沼有紀,田中基嗣,安達泰治,矢野澄雄,北條正樹
単離骨細胞の力学刺激に対する応答観察
日本機械学会 2005 年関西学生会学生員卒業研究発表講演会,講演前刷集pp. 13-11,
京都,(2005.3.17-19).
D-2. 田中基嗣,青沼有紀,安達泰治,上岡 寛,山本照子,北條正樹,
力学刺激に対する単離骨細胞の応答観察
日本機械学会 2005 年年次大会講演論文集,No. 05-1,Vol. 6,pp. 119-120,調布,
(2005.9.19-22).
D-3. 田中基嗣,青沼有紀,安達泰治,上岡 寛,山本照子,北條正樹,
メカノセンサーとしての骨細胞における力学刺激応答のin vitro観察
日本複合材料学会第 30 回複合材料シンポジウム講演論文集,pp. 255-256,松山,
(2005.10.19-21).
D-4. 青沼有紀,田中基嗣,安達泰治,上岡 寛,山本照子,矢野澄雄,北條正樹
局所変形負荷に対する単離骨細胞のカルシウム応答
日本機械学会第 16 回バイオフロンティア講演会,日本機械学会講演論文集No. 05-53,
pp. 7-8,草津,(2005.11.10-11).
D-5. 田中基嗣,青沼有紀,伊藤慎一,安達泰治,上岡 寛,山本照子,北條正樹
骨細胞ネットワークによるメカノセンサーシステムの力学応答観察
日本材料学会第 35 回FRPシンポジウム講演論文集,pp. 157-161,京田辺,(2006.3.16-17).
D-6. 青沼有紀,田中基嗣,安達泰治,上岡 寛,山本照子,矢野澄雄,北條正樹
単一の単離骨細胞における局所変形による力学刺激とカルシウム応答の関係
日本機械学会 2006 年年次大会講演論文集No. 06-1,Vol. 5,pp. 263-264,熊本,
(2006.9.18-22).
D-7. 青沼有紀,田中基嗣,安達泰治,上岡 寛,山本照子,矢野澄雄,北條正樹
マイクロパーティクルを用いた直接変形負荷に対する単離骨細胞のカルシウム応答
79
Page 85
日本機械学会第 19 回バイオエンジニアリング講演会,日本機械学会講演論文集No.
06-65,pp. 106-107,仙台,(2007.1.7-8).
D-8. 青沼有紀,安達泰治,田中基嗣,上岡 寛,山本照子,北條正樹
単離骨細胞の細胞突起/細胞体の力学的刺激に対するカルシウム応答の違い
第 26 回日本骨代謝学会学術集会,講演論文集pp. 215,大阪,(2007.7.19-21).
D-9. 青沼有紀,安達泰治,田中基嗣,上岡 寛,山本照子,北條正樹,
骨基質の変形にともなう骨細胞の変形およびカルシウム応答の観察
日本機械学会第 20 回バイオエンジニアリング講演会,日本機械学会講演論文集No.
07-49,pp. 337-338,東京,(2008.1.25-26).
D-10. 平 圭佑,青沼有紀,安達泰治,北條正樹,上岡 寛
単離骨系細胞間のカルシウム応答伝播特性
日本機械学会 2008 年関西学生会学生員卒業研究発表講演会,講演前刷集pp. 2-13,吹
田,(2008.3.17).
D-11. 安達泰治,青沼有紀,田中基嗣,北條正樹,山本照子,上岡 寛
単離骨細胞における局所的な力学刺激応答特性
第 31 回バイオレオロジー学会年会,日本バイオレオロジー学会誌,Vol. 22,Suppl.,
pp. 31,東京,(2008.6.5-6).
D-12. 平 圭佑,青沼有紀,安達泰治,北條正樹,上岡 寛
力学刺激に対する単離骨系細胞のカルシウム応答と細胞間伝播
日本機械学会第 19 回バイオフロンティア講演会,日本機械学会講演論文集No. 08-30,
pp. 171-172, 八王子,(2008.9.24-25).
D-13. 青沼有紀,安達泰治,田中基嗣,北條正樹,山本照子,上岡 寛
骨基質の変形に対する骨細胞のカルシウム応答観察
第 26 回日本骨代謝学会学術集会,講演論文集pp. 173,大阪,(2008.10.29-31).
80
Page 86
D-14. 平 圭佑,青沼有紀,安達泰治,北條正樹,上岡 寛
単離骨系細胞間におけるカルシウム応答伝播の非対称性
日本機械学会 2009 年年次大会,講演論文集No. 09-1,pp. 21-22,盛岡,(2009.9.13-16).
D-15. 青沼有紀,平 圭佑,安達泰治,北條正樹,上岡 寛
力学刺激に対する単離骨系細胞間の情報伝達特性に関する考察
日本機械学会第 20 回バイオエンジニアリング講演会,日本機械学会講演論文集No.
09-55,pp. 240,岡山,(2010.1.9-10).
81
Page 88
謝 辞
本研究の遂行にあたり,懇切なる御指導,また研究全般にわたる貴重な御助言を賜
りました京都大学大学院工学研究科の北條正樹教授に心より感謝の意を表します.ま
た,終始懇切な御指導と御鞭撻を賜りました京都大学大学院工学研究科の安達泰治准
教授に深甚なる謝辞を捧げます.
また,京都大学大学院工学研究科の富田直秀教授には,本論文の作成に際して,御
多忙中のところ有益な御助言と御教示を賜りました.厚く御礼申し上げます.
本論文の作成にあたり,丁寧な御指導,御助言を賜りました金沢工業大学工学部の
田中基嗣講師に深く感謝いたします.また,日々の研究活動において,有益な御助言,
御討論を賜りました京都大学大学院工学研究科の井上康博助教に,心より御礼申し上
げます.
東北大学大学院歯学研究科の山本照子教授,岡山大学大学院医歯薬学総合研究科の
上岡寛准教授,菅原康代助教,石原嘉人博士には,本研究の遂行における実験に際し
て,骨細胞研究の概論と単離法の御教授にとどまらず,貴重な御助言を頂きました.
ここに厚く御礼申し上げます.
ACTA Frije University の Jenneke Klein-Nulend 教授,Cornelius M. Semeins 氏,Rommel
G. Bacabac 博士には,本研究で用いたモノクローナル抗体 OB 7.3 を御提供頂き,また
京都に御滞在の際には骨細胞研究に関する有益な御討論の機会を頂きました.深く感
謝の意を表します.
本研究の共同研究者として,ともに研究を遂行させていただいた伊藤慎一氏 (現帝
人株式会社),平圭佑氏 (京都大学大学院工学研究科) に深く感謝いたします.また,
細胞培養や顕微鏡観察法をはじめとする基本的な実験手技から細胞生物学の教授に
至るまで,幅広い御指導を頂きました,理化学研究所の須長純子氏,電子顕微鏡を用
いた細胞観察において丁寧な御指導と御助言を頂きました,京都大学大学院医学研究
科総合解剖センターの幸田晴康氏,古田敬子氏に心より御礼申し上げます.
83
Page 89
84
徳島大学大学院ソシオテクノサイエンス研究部の佐藤克也講師には,著者が神戸大
学発達科学部在籍時,本研究の立ち上げにあたって,細胞に対する力学実験の指導を
はじめ,丁寧な御指導を頂きました.ここに深く感謝いたします.
神戸大学大学院人間発達環境学研究科の矢野澄雄教授には,著者が神戸大学発達科
学部,および神戸大学大学院総合人間科学研究科在籍時,人間の生活環境を構成する
力学的環境の諸問題という広範な研究対象から,本研究遂行のための動機付け,医工
学的意義に至るにあたって,懇切な御指導を賜りました.心より感謝の意を表します.
Okeyo Kennedy Omondi 氏,島田義孝氏をはじめとする,京都大学大学院工学研究
科機械理工学専攻適応材料力学研究室の諸氏とは,ともに研究生活を過ごし,研究の
ことにとどまらず有意義な時間を過ごさせて頂きました.記してここに謝意を表しま
す.
後に,博士後期課程の進学に御理解を頂き,研究生活を通して御支援を頂いた両
親,祖母,姉,兄に 大限の感謝の意を表します.また,研究活動において,家族と
変わらぬ御配慮を頂きました友人・知人の皆様に,深く感謝いたします.