Page 1
This article was downloaded by: [Memorial University of Newfoundland]On: 18 July 2014, At: 08:07Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Plant NutritionPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/lpla20
Tolerance of Lisianthus to High Ammonium Levels inRockwool CultureRosalinda Mendoza-Villarreala, Luis A. Valdez-Aguilara, Alberto Sandoval-Rangela, ValentínRobledo-Torresa & Adalberto Benavides-Mendozaa
a Departamento de Horticultura, Universidad Autónoma Agraria Antonio Narro, Saltillo,Coah, MéxicoAccepted author version posted online: 27 May 2014.
To cite this article: Rosalinda Mendoza-Villarreal, Luis A. Valdez-Aguilar, Alberto Sandoval-Rangel, Valentín Robledo-Torres &Adalberto Benavides-Mendoza (2014): Tolerance of Lisianthus to High Ammonium Levels in Rockwool Culture, Journal of PlantNutrition, DOI: 10.1080/01904167.2014.920379
To link to this article: http://dx.doi.org/10.1080/01904167.2014.920379
Disclaimer: This is a version of an unedited manuscript that has been accepted for publication. As a serviceto authors and researchers we are providing this version of the accepted manuscript (AM). Copyediting,typesetting, and review of the resulting proof will be undertaken on this manuscript before final publication ofthe Version of Record (VoR). During production and pre-press, errors may be discovered which could affect thecontent, and all legal disclaimers that apply to the journal relate to this version also.
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Page 2
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 1
Tolerance of Lisianthus to High Ammonium Levels in Rockwool Culture
Rosalinda Mendoza-Villarreal, Luis A. Valdez-Aguilar, Alberto Sandoval-Rangel, Valentín
Robledo-Torres, Adalberto Benavides-Mendoza
Departamento de Horticultura, Universidad Autónoma Agraria Antonio Narro, Saltillo, Coah.,
México
Address correspondence to Luis A. Valdez-Aguilar: Departamento de Horticultura, Universidad
Autónoma Agraria Antonio Narro, Saltillo, Coah., México. E-mail: [email protected]
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 3
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 2
ABSTRACT
Nitrogen (N) by form of nutrition, ammonium (NH4+) or nitrate (NO3
–), affects metabolic and
physiological processes of plants. In general, a high proportion of N in the NH4+ form results in
poor growth of plants. Nonetheless, a number of species exhibit optimum growth when high
levels of NH4+ are provided. In the present study, lisianthus [Eustoma grandiflorum (Raf.) Shinn]
was grown in rockwool cultures and irrigated with nutrient solutions containing 15 mM N with
varying proportions of NH4+ and NO3
–. The results showed that an increase in NH4+-N form
increased plant height, number of flowers and leaves, leaf area, and shoot, stem, and leaf dry
weight. The proportion of NH4+ also affected leaf concentration of phosphorus, potassium (K),
calcium (Ca), and magnesium (Mg), although leaf N concentration was unaffected. Potassium
leaf concentration was higher when a low proportion of NH4+ was supplemented in the nutrient
solution; however, plants exhibited a decrease in leaf K concentration and a decrease in leaf Ca
as the proportion of NH4+-N increased. Shoot dry weight was higher with low leaf K whereas
high leaf Ca was associated with high shoot dry weight. Net photosynthesis rate was higher in
plants irrigated with solutions containing 75% of total N in NH4+ form than in those irrigated
with solutions of 0 or 25%. The results suggest that lisianthus can tolerate high levels of NH4+,
probably associated with a higher assimilation of Ca.
Keywords: Nitrate:ammonium ratio, Nitrogen nutrition, Nutrient solution, Soilless culture
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 4
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 3
INTRODUCTION
The form by which nitrogen (N) is absorbed by plants, ammonium (NH4+) or nitrate (NO3
–),
markedly affects metabolic and physiological processes (Gerendás et al., 1997). This response is
due partially to the effect of the uptake of different N forms on cytosol and rhizosphere acidity,
which in turn affects availability of nutrients essential for plant growth and development. In
general, a high proportion of N supplemented as NH4+ results in poor growth of most of plants
species; however, contrasting results have been reported. Gaiad et al. (2006) indicated that, with
NH4+ nutrition, seedlings of mate plant (Ilex paraguariensis St. Hil.) exhibited increased leaf
area, leaf number, and net photosynthesis; nonetheless, in two cultivars of strawberry plants
(Fragaria × ananassa Duch), a supplement of NH4+ at 75% of total N resulted in decreased leaf
area, leaf fresh weight, and leaf dry weight (Tabatabaei et al., 2006).
The negative effects of high NH4+ proportions on plant growth have been associated with
acidification in the root zone (Walch-Liu et al., 2000; Claussen and Lenz, 1999), an action that
affected growth due to direct high acidity effects or reductions in cations uptake, such as
potassium (K) (Arnozis et al., 1988). Excessive NH4+ nutrition also has been associated with
plant hormones status since the biosynthesis of ethylene and polyamines are triggered under such
conditions; severe changes in the levels of aminoacids and organic acids such as citrate have
been also observed (Britto and Kronzucker, 2002; Zhang and Rengel, 1999; Gerendás et al.,
1997; Redinbaugh and Campbell, 1993; Barker and Corey, 1991).
Lisianthus [Eustoma grandiflorum (Raf.) Shinn] is a species that was introduced recently
in the market of ornamental plants (Harbaugh, 2007). Native to the arid and semiarid regions of
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 5
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 4
south United States (Halevy and Kofranek, 1984) and north México (Harbaugh, 2007), its
cultivation has extended worldwide. Little research has been conducted though as to the
nutritional demands of lisianthus. It has been reported that increasing N concentration in the
nutrient solution from 0 to 400 mg L-1 resulted in improved growth in lisianthus (Freet et al.,
1988) whereas concentration from 250 to 300 mg L-1 results in higher chlorophyll concentration
and photosynthesis rate (Marchese et al., 2005). However, as there are not reports defining the
optimum proportion of N form, the present study was conducted to determine the effect of the
proportion of NH4+ and NO3
– in nutrient solutions on growth, internal macronutrient status, and
photosynthetic rate on lisianthus cultivated in rockwool.
MATERIALS AND METHODS
The study was conducted in a greenhouse located in Saltillo, Coah., México (25° 27 North
Latitude, 101° 02 West Longitude, 1610 meters above sea level) and equipped with automatic
control of temperature. Average daytime photosynthetically active radiation (PAR) registered
throughout the experiment was 190.5 µmol m–2 s–1; maximum PAR, observed from 13:00 to
15:00, was 498.5 µmol m–2 s–1. Average maximum and minimum temperatures recorded were de
28.9 °C and 19.9°C, respectively.
Seeds of lisianthus [Eustoma grandiflorum Raf. (Shinn)] Echo Blue were sown on
sphagnum peat in 200 cell-rigid plastic trays on Feb. 26, 2008. Non-bolting seedlings of 4-cm
height and four leaf pairs were transplanted on May 8, 2008, in rockwool slabs of 100 × 20 × 7.5
cm. A total of 14 seedlings were planted on each slab and allowed to establish for one week
previous to treatment imposition.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 6
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 5
The treatments consisted of six nutrient solutions with a total N concentration of 15 mM.
Nutrient solutions were prepared at 75%, 50%, 37.5%, 25%, 12.5%, and 0% NH4+-N by varying
the amounts of ammonium sulfate, calcium nitrate, ammonium nitrate, ammonium chloride, and
potassium nitrate. The remaining N to complete the 15 mM was supplemented with NO3–-N. The
nutrient solutions contained also 1.6 mM phosphorus (P), 6.2 mM K, 6 mM calcium (Ca), 2.8
mM magnesium (Mg), and 0.1-0.2 mM sulfate. Micronutrients were supplemented at the
following concentrations: 5 ppm iron, 0.05 ppm zinc, 0.02 ppm copper, 0.7 ppm manganese, and
0.5 ppm boron. The pH of the nutrient solutions was adjusted to 6.8 with phosphorus acid
(H3PO4) previous preparation and average electrical conductivity was 2.7 dS m–1. Irrigation with
the nutrient solutions started on May 14, 2008 applying 2.5 L per slab three times a week;
leaching nutrient solution was not retrieved.
Harvesting was performed from Aug. 18 to Sept. 3 when plants had three flowers fully
open. Plant height was measured at harvest time as well as the diameter at the base of the main
stem, the number of leaves and flowers per plant, leaf area (LI-3100 Area Meter; LI-COR
Biosciences, Lincoln, NE, USA), and dry weight of leaves, flowers, and stems. Shoot dry weight
was calculated adding the partial weights of stems, flowers, and leaves. Plant parts were bagged
and placed in an oven at 70ºC for three days previous to weighing. Previous to drying, leaves
were washed twice in distilled water; once dry, the leaves were ground to pass a 20-mesh screen.
Ground samples were analyzed for N concentration with the Kjeldahl procedure (Chapman and
Pratt, 1961) and phosphorus (P), potassium (K), calcium (Ca), and magnesium (Mg)
concentrations were determined on acid digests [sulfuric acid (H2SO4):perchloric acid (HClO4)
(2:1ml)] (Alcántar and Sandoval-Villa, 1999) with inductively coupled plasma spectrometry
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 7
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 6
(ICP-AES VARIAN, model Liberty; Varian Medical Systems, Palo Alto, CA, USA). On Aug. 5,
2008, net photosynthetic rate was measured on fully expanded young leaves in plants of selected
treatments (75%, 25%, and 0% NH4+) with an infrared gas analyzer (Li-6200; LI-COR
Biosciences, Lincoln, NE, USA).
Treatments were distributed in a randomized complete block design with four
replications. Each replication consisted on one rockwool slab with 14 plants uniformly
distributed. Analysis of data was with analysis of variance, trend analysis (linear, quadratic, and
cubic effects), and mean comparison with Tukey’s procedure; SAS version 8.02 (SAS Institute,
Inc. 2001, Cary, NC, USA) was used to process the data. Regression models were calculated
when associations between growth parameters or nutrient concentrations were observed.
RESULTS
Increasing NH4+-N form enhanced plant growth since lisianthus resulted with larger plant height
and increased number of flowers and leaves, leaf area, and shoot, stems, and leaf dry weight
(Table 1). The effect of NH4+ was predominantly lineal in most of these growth parameters and
the plants irrigated with solutions containing 50% or 75% of total N as NH4+ exhibited
significant enhancing effects compared to the other levels.
The proportion of NH4+ in the nutrient solution significantly affected leaf concentration
of P, K, Ca, and Mg, although leaf N concentration was unaffected (Table 2). The effect on leaf
P, Ca, and Mg concentration fitted a quadratic and/or cubic trend, whereas for leaf K
concentration the trend was lineal. At low NH4+ proportions in the nutrient solution, 0% or
12.5%, leaves exhibited a low concentration of P, Ca, and Mg; however, increasing the
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 8
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 7
proportion of NH4+ to moderate levels, 12.5% to 37.5%, resulted in a significant increase in the
concentration of these nutrients. Higher NH4+ proportions, 50% to 75%, were associated with a
decrease in the concentration of leaf P, Ca, and Mg concentrations. Potassium leaf concentration
was higher when a low proportion of NH4+ was supplemented in the nutrient solution; however,
as the proportion of NH4+ was increased, plants exhibited a significant decrease in leaf K
concentration.
Leaf K concentration was associated with leaf Ca concentration. An increasing leaf K
concentration was associated with a decrease in leaf Ca with a quadratic trend (Figure 1). Leaf
concentration of K or Ca, was associated contrastingly with growth of lisianthus since shoot dry
weight was high with low leaf K (Figure 2) whereas high leaf Ca was associated with high shoot
dry weight (Figure 3).
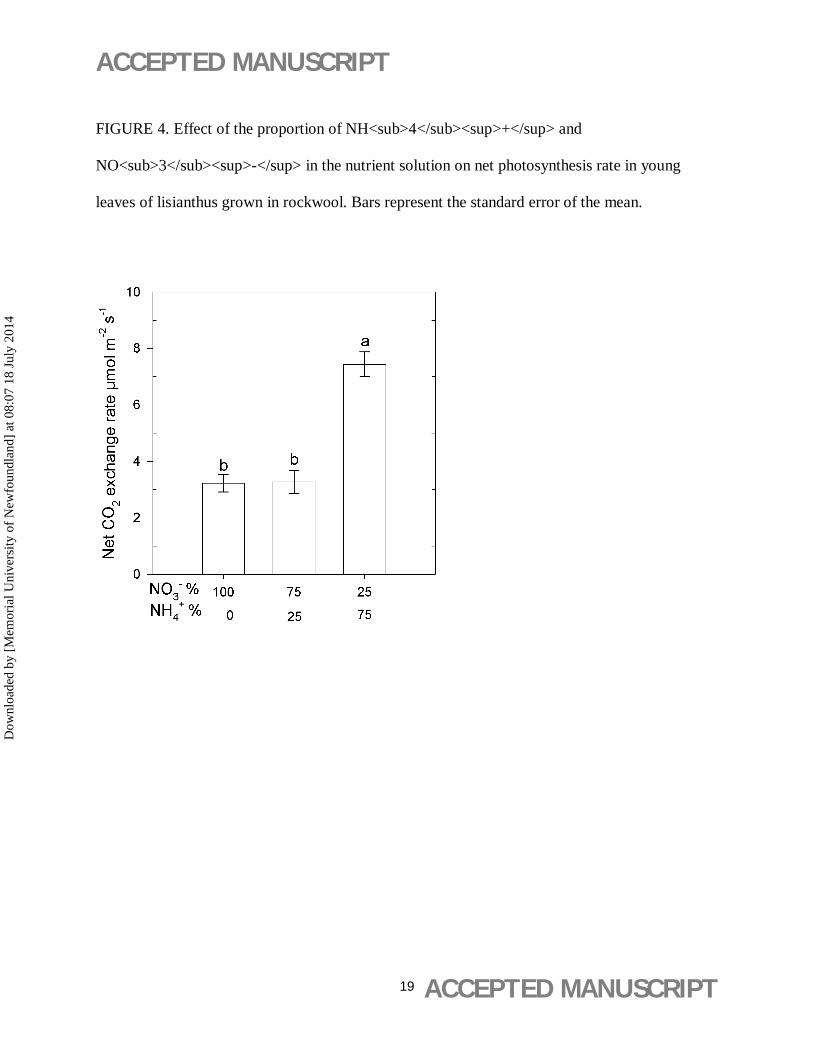
Net photosynthesis rate was affected by the proportion of NH4+ in the nutrient solution.
Plants irrigated with solutions containing 75% of total N as NH4+ exhibited higher
photosynthesis rate than plants that received NH4+ at proportions of 25% or lower (Figure 4).
DISCUSSION
It is accepted widely that NO3– or mixtures of NO3
– with low proportions of NH4+ are the best
form for N supplementation in plants due to the toxic effect of a N nutrition based on high
proportions of NH4+ (Chen et al., 2005; Dong et al., 2004; Shen et al., 2003). Nonetheless, the
results of the present study indicated that growth of lisianthus is enhanced under N nutrition
based on high proportions of NH4+ mixed with low proportions of NO3
–. The tolerance of
lisianthus to high NH4+ rates also was detected at physiological levels since net photosynthesis
was higher when plants were irrigated with solutions containing a high proportion of NH4+.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 9
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 8
Comparable results have been reported in other plant species; in mate plant (I. paraguariensis St.
Hil.) an increase in leaf area, leaf number and net photosynthesis was detected when N was
supplemented exclusively in NH4+ form (Gaiad et al., 2006). Similarly, candelabra tree
(Araucaria angustifolia Bertol.) exhibited improved growth when NH4+ was provided compared
to exclusively NO3– fed plants, which exhibited a lower chlorophyll and N concentration in
young leaves than plant receiving NH4+ nutrition (Garbin and Dillenburg, 2008). In common
heather plants [Calluna vulgaris (L.) Hull] optimum growth was observed when NH4+ was the
sole N form supplemented (De Graaf et al., 1998).
In the present study, N form had no effect on leaf N concentration. This result suggests
that N assimilation in lisianthus was a function of total external N concentration since it was
maintained constant in all the nutrient solutions, and it was not dependent on the form on which
N was supplemented. However, N form did affect the assimilation of other plant nutrients.
Leaves of lisianthus resulted with increased P, Ca, and Mg when the proportion of NH4+ was
from 12.5% to 50% of total N.
Plants fed with NH4+ are reported to exhibit a K deficiency attributed to competition for
uptake sites (Hoopen et al., 2010; Hess et al., 2006) due to the similarities in both cations (Xu et
al., 2002; Wang et al., 1996). In the present study, lisianthus showed a lineal decrease in leaf K
concentration as NH4+ in the nutrient solution was increased, especially when its proportion was
from 50% to 75% of total N. The decrease in K leaf concentration was from 23% to 36% at these
NH4+ proportions, compared to plants that received only NO3
– N. Nonetheless, in spite of the
decrease in leaf K concentration, plants did not exhibit K deficiency symptoms and were able to
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 10
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 9
reach optimum growth, suggesting that interactions with other nutrients may have resulted
positive for growth.
An excessive proportion of NH4+ has been reported to cause a decrease in Ca
concentration in plant tissues due to the antagonism between both cations (Siddiqi et al., 2002).
Nonetheless, lisianthus did not exhibit a decrease in leaf Ca concentration at high proportions of
NH4+ in the nutrient solution, but an increase in leaf Ca accumulation was detected at moderate
NH4+ proportions. This response may be related to the interaction of Ca with other nutrients,
specifically K, since an increase in leaf Ca was associated with a decrease in leaf K
concentration. Bar-Tal and Pressman (1996) reported similar results in tomato since there was a
decrease in tissue Ca concentration as external K was increased. The negative interaction
between K and Ca was confirmed by the association between a low leaf K and a high leaf Ca
concentration with a high shoot biomass accumulation by lisianthus plants and, in general, with
improved plant growth. Likewise, the higher net photosynthetic rate observed in plants irrigated
with solutions of high NH4+ proportions suggests that lisianthus was in adequate physiological
status so that plants were able to produce higher biomass.
The results observed in the present study suggest that lisianthus can tolerate high levels of
NH4+, probably due to an increase in calcium concentration. Further research is recommended to
assess the possibility of achieving higher tolerance of lisianthus to even higher NH4+ proportions
by increasing Ca concentrations in the nutrient solution or foliar spays.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 11
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 10
REFERENCES
Arnozis, P.A., J.A. Nelemans, and G.R. Findenegg. 1988. Phospoenolpyruvate carboxylate
activity in plants grown with either NO3– or NH4
+ as inorganic nitrogen source. Journal
of Plant Physiology 132:23–27.
Alcántar, G.G., and M. Sandoval-Villa. 1999. Chemical analysis of plant tissues manual.
Sociedad Mexicana de la Ciencia del Suelo. México. In Spanish.
Bar-Tal, A., and E. Pressman. 1996. Root restriction and potassium and calcium solution
concentrations affect dry-matter production, cation uptake, and blossom-end rot in
greenhouse tomato. Journal of the American Society for Horticultural Science 121:649–
655.
Barker, A.V., and K.A. Corey. 1991. Interrelations of ammonium toxicity and ethylene action in
tomato. HortScience 26:177–180.
Britto, D., and H. Kronzucker. 2002. NH4+ toxicity in higher plants: a critical review. Journal of
Plant Physiology 159:567–584.
Chapman, H.D., and P.F. Pratt. 1961. Methods of Analysis for Soils, Plants and Waters. University of California,
Division of Agricultural Sciences: Riverside, CA.
Chen, W., J.K. Lou, and Q.R. Shen. 2005. Effect of NH4+-N/ NO3
–-N ratios on growth and some
physiological parameters of chinese cabbage cultivars. Pedosphere 15:310–318.
Claussen, W., and F. Lenz. 1999. Effect of ammonium or nitrate nutrition on net photosynthesis
growth and activity of the enzymes nitrate reductase and glutamine synthetase in
blueberry, raspberry and strawberry. Plant and Soil 208:95–102.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 12
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 11
De Graaf, M.C.C., R. Bobbink, G.M. Jan, J.G.M. Roelofs, and P.J.M. Verbeek. 1998.
Differential effects of ammonium and nitrate on three heathland species. Plant Ecology
135:185–196.
Dong, C.X., Q.R. Shen, and G. Wang. 2004. Tomato growth and organic acid changes in
response to partial replacement of NO3 -N by NH4
+-N. Pedosphere 14:159–164.
Freet, J.J., J.W. Kelly, B.K. Harbaugh, and M. Roh. 1988. Optimizing nitrogen and calcium
nutrition of lisianthus. Communications in Soil Science and Plant Analysis 19:13–24.
Gaiad, S., M. Rakocevic, and C.B. Reissmann. 2006. N sources affect growth, nutrient content,
and net photosynthesis in mate (Ilex paraguariensis St. Hil.). Brazilian Archives of
Biology and Technology 49:689–697.
Garbin, M.L., and L.R. Dillenburg. 2008. Effects of different nitrogen sources on growth,
chlorophyll concentration, nitrate reductase activity and carbon and nitrogen distribution
in Auracaria angustifolia. Brazilian Society of Plant Physiology 20:295–303.
Gerendás, J., Z. Zhu, R. Bendixen, R., Ratcliffe, and B. Sattelmacher. 1997. Physiological and
biochemical processes related to ammonium toxicity in higher plants. Journal of Plant
Nutrition and Soil Science 160:239–251.
Halevy, A., and A.M. Kofranek. 1984. Evaluation of Lisianthus as a new flower crop.
HortScience 19:845–847.
Harbaugh, B.K. 2007. Lisiantus. Eustoma grandiflorum. In: Flower Breeding and Genetics, ed.
N.O. Anderson, pp 645–663. Springer Verlag: Dordrecht, The Netherlands.
Hess, D.C., W. Lu, J.D. Rabinowitz, and D. Botstein. 2006. Ammonium toxicity and potassium
limitation in yeast. Public Library of Science Biology 4:351–355.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 12
Hoopen, F., T.A. Cuin, P. Pedas, J.N. Hegelund, S. Shabala, J.K. Schjoerring, and T.P. Jahn.
2010. Competition between uptake of ammonium and potassium in barley and
Arabidopsis roots: molecular mechanisms and physiological consequences. Journal of
Experimental Botany 61:2303–2315.
Marchese, J.A., I. Katz, A.P. Sousa, and J.D. Rodrigues. 2005. Gas exchange in lisianthus plants
(Eustoma grandiflorum) submitted to different doses of nitrogen. Photosynthetica
43:303–305.
Redinbaugh, M.G., and W.H. Campbell. 1993. Glutamine-synthetase and ferredoxin-dependent
glutamate synthase expression in the maize (Zea mays) root primary response to nitrate.
Evidence for an organ-specific response. Plant Physiology 101:1249–1255.
SAS Institute, Inc. 2001. SAS/STAT software changes and enhancements through release 8.0.2.
SAS Institute Inc., Cary, NC.
Shen, Q.R., L. Tang, Y.C. Xu. 2003. A review on the behavior of nitrate in vacuoles of plants.
Acta Pedologica Sinica (in Chinese) 40:465–470.
Siddiqi, M.Y., B. Malhotra, X. Min, and A.D.M. Glass. 2002. Effects of ammonium and
inorganic carbon enrichment on growth and yield of a hydroponic tomato crop. Journal
of Plant Nutrition and Soil Science 165:191–197.
Tabatabaei, S.J., L.S. Fatemi, and E. Fallahi. 2006. Effect of ammonium:nitrate ratio on yield,
calcium concentration, and photosynthesis rate in strawberry. Journal of Plant Nutrition
29:1273–1285.
Walch-Liu, P., G. Neumann, F. Bangerth, and C. Engels. 2000. Rapid effects of nitrogen form on
leaf morphogenesis in tobacco. Journal of Experimental Botany 51:227–237.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 14
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 13
Wang, M.Y., M.Y. Siddiqi, and A.D.M. Glass. 1996. Interaction between K+ and NH4+: effects
on ion uptake by rice roots. Plant Cell and Environment 19:1037–1046.
Xu, G., S. Wolf, and U. Kafkafi. 2002. Ammonium on potassium interaction in sweet pepper.
Journal of Plant Nutrition 25:719–734.
Zhang, X-K., and Z. Rengel. 1999. Gradients of pH and ammonium and phosphorus
concentration between the banded fertilizer and wheat roots. Australian Journal of
Agricultural Research 50:365–373.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 15
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 14
Table 1. Effect of varying proportions of NH4+ in the nutrient solution on growth parameters of
lisianthus grown in rockwool with N concentration maintained at 15 mM
% in nutrient solution
Dry weight g
NH4+ NO3
–
Number of
flowers
Plant height
cm
Leaf area cm2
Stem diameter
mm
Number of
leaves Leaf Flower Stem Shoot
0 100 12b 73.9ab 459d 3.98 47d 1.60d 1.92 2.60c 6.14c 12.5 87.5 15b 77.1a 690c 4.40 67c 2.20cd 2.34 3.55bc 8.11bc 25 75 14b 75.5a 825abc 4.56 72c 2.60bc 2.26 3.83ab 8.71b 37.5 62.5 14b 69.8b 718bc 4.31 81bc 2.47bc 2.02 3.19bc 7.69bc 50 50 16b 79.0a 874ab 4.61 95b 3.09ab 2.69 4.07ab 9.86ab 75 25 22a 74.1ab 970a 4.88 118a 3.67a 3.06 4.71a 11.45a
Anova *** * *** NS *** *** NS * ** Lineal *** NS *** * *** *** * ** ***
Quadratic NS NS NS NS NS NS NS NS NS Cubic * NS * NS NS NS NS NS NS
Means followed by the same letter indicate non-significant difference according to Tukey´s test
with P=0.05.
NS, *, **, *** non-significant and significant at P<0.05, 0.01, and 0.001, respectively, according
to F test
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 16
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 15
Table 2. Effect of varying proportions of NH4+ in the nutrient solution on nitrogen (N),
phosphorus (P), potassium (K), calcium (Ca), and magnesium (Mg) concentration in leaves of
lisianthus grown in rockwool with N concentration maintained at 15 mM
% in nutrient solution N P K Ca Mg NH4
+ NO3– mmol kg-1
0 100 2429 97cd 928a 120b 361b 12.5 87.5 2386 159ab 835ab 204a 395ab 25 75 2571 184a 832ab 166ab 426ab 37.5 62.5 2429 79d 860ab 141b 447a 50 50 2293 120bc 594c 182ab 379ab 75 25 2279 137b 702bc 173ab 371ab
Anova NS *** ** ** * Lineal NS NS *** NS NS
Quadratic NS * NS NS ** Cubic NS *** NS * NS
Means followed by the same letter indicate non-significant difference according to Tukey´s test
with P=0.05.
NS, *, **, *** non-significant and significant at P<0.05, 0.01, and 0.001, respectively, according
to F test
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 17
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 16
FIGURE 1. Association between internal concentration of K and Ca in leaves of lisianthus
grown in rockwool in response to varying proportions of NH<sub>4</sub><sup>+</sup> and
NO<sub>3</sub><sup>-</sup> in the nutrient solutions. Bars represent the standard error of the
mean.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 18
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 17
FIGURE 2. Association between leaf concentration of K and shoot dry mass accumulation at
experiment termination in plants of lisianthus grown in rockwool in response to varying
proportions of NH<sub>4</sub><sup>+</sup> and NO<sub>3</sub><sup>-</sup> in the
nutrient solutions. Bars represent the standard error of the mean.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 19
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 18
FIGURE 3. Association between leaf concentration of Ca and shoot dry mass accumulation at
experiment termination in plants of lisianthus grown in rockwool in response to varying
proportions of NH<sub>4</sub><sup>+</sup> and NO<sub>3</sub><sup>-</sup> in the
nutrient solutions. Bars represent the standard error of the mean.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4
Page 20
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 19
FIGURE 4. Effect of the proportion of NH<sub>4</sub><sup>+</sup> and
NO<sub>3</sub><sup>-</sup> in the nutrient solution on net photosynthesis rate in young
leaves of lisianthus grown in rockwool. Bars represent the standard error of the mean.
Dow
nloa
ded
by [
Mem
oria
l Uni
vers
ity o
f N
ewfo
undl
and]
at 0
8:07
18
July
201
4