Page 1
Understanding the spectacular failure of DNA barcodingin willows (Salix): Does this result from a trans-specificselective sweep?
DIANA M. PERCY,*† GEORGE W. ARGUS,‡ QUENTIN C. CRONK,*† ARON J. FAZEKAS,§PRASAD R. KESANAKURTI ,§ KEVIN S. BURGESS,¶ BRIAN C. HUSBAND,§ STEVEN G.
NEWMASTER,§ SPENCER C.H. BARRETT** and SEAN W. GRAHAM*†
*Department of Botany, University of British Columbia, Vancouver, BC, Canada V6T 1Z4, †Biodiversity Research Centre,
University of British Columbia, Vancouver, BC, Canada V6T 1Z4, ‡Canadian Museum of Nature, PO Box 3443 Stn “D”,
Ottawa, ON, Canada K1P 6P4, §Department of Integrative Biology, University of Guelph, Guelph, ON, Canada N1G 2W1,
¶Department of Biology, Columbus State University, Columbus, GA 31907-5645, USA, **Department of Ecology &
Evolutionary Biology, University of Toronto, 25 Willcocks Street, Toronto, ON, Canada M5S 3B2
Abstract
Willows (Salix: Salicaceae) form a major ecological component of Holarctic floras and
consequently are an obvious target for a DNA-based identification system. We sur-
veyed two to seven plastid genome regions (~3.8 kb; ~3% of the genome) from 71 Salixspecies across all five subgenera, to assess their performance as DNA barcode markers.
Although Salix has a relatively high level of interspecific hybridization, this may not
sufficiently explain the near complete failure of barcoding that we observed: only one
species had a unique barcode. We recovered 39 unique haplotypes, from more than 500
specimens, that could be partitioned into six major haplotype groups. A unique variant
of group I (haplotype 1*) was shared by 53 species in three of five Salix subgenera.
This unusual pattern of haplotype sharing across infrageneric taxa is suggestive of
either a massive nonrandom coalescence failure (incomplete lineage sorting), or of
repeated plastid capture events, possibly including a historical selective sweep of hap-
lotype 1* across taxonomic sections. The former is unlikely as molecular dating indi-
cates that haplotype 1* originated recently and is nested in the oldest major haplotype
group in the genus. Further, we detected significant non-neutrality in the frequency
spectrum of mutations in group I, but not outside group I, and demonstrated a striking
absence of geographical (isolation by distance) effects in the haplotype distributions of
this group. The most likely explanation for the patterns we observed involves recent
repeated plastid capture events, aided by widespread hybridization and long-range
seed dispersal, but primarily propelled by one or more trans-species selective sweeps.
Keywords: chloroplast capture, DNA barcoding, Malpighiales, molecular dating, phylogeogra-
phy, selective sweep
Received 19 December 2012; revision received 29 May 2014; accepted 4 June 2014
Introduction
A partial failure of plastid data to track species bound-
aries is fairly common in phylogenetic plant studies
(Percy et al. 2008; Starr et al. 2009; Hassel et al. 2013).
This is often inferred by observations of strong incon-
gruence between plastid and nuclear gene trees, or dis-
agreements between gene trees and classically defined
species (Xu et al. 2012; Yu et al. 2013). The processes
underlying this incongruence likely contribute to the
somewhat lower success of DNA barcoding markers as
an identification tool in plants vs. animals (Fazekas
et al. 2009), possibly reflecting more slowly evolving
markers, or more problematic levels of hybridization,
Correspondence: Diana M. Percy, Department of Life Sciences,
Natural History Museum, Cromwell Road, London SW7 5BD,
UK, Fax: +44 (0)2079425229; E-mail: [email protected]
© 2014 John Wiley & Sons Ltd
Molecular Ecology (2014) doi: 10.1111/mec.12837
Page 2
introgression or incomplete lineage sorting (Hollings-
worth et al. 2011). Here, we report on patterns of varia-
tion in DNA barcoding markers in the genus Salix. The
spectacular failure of DNA barcoding that we observe
in willows may require an explanation involving trans-
specific selection. This in turn may have important con-
sequences for the interpretation of incongruence in
other taxonomically complex plant groups.
Willows (Salix; Salicaceae), a genus of shrub and tree
species, are a significant component of Holarctic ecosys-
tems (Ager & Phillips 2008; Argus 2010; Myers-Smith
et al. 2011). They are important indicators of riparian
habitats, and many ecological studies include estimates
of the diversity and abundance of willows because of
their contribution to ecosystem function, community
dynamics and assemblage (Myers-Smith et al. 2011).
They are also extensively planted for habitat restoration,
erosion control, and windbreaks, and the physiological
adaptations and ecological resilience of willows make
them valuable species for use in conservation and envi-
ronmental projects (Kuzovkina & Quigley 2005;
Kuzovkina & Volk 2009).
The genus contains ~450 species worldwide, includ-
ing localized and widespread species with extensive cir-
cumpolar distributions. However, Salix species are
notoriously difficult to identify based on morphology.
Many species are vegetatively similar and exhibit sub-
stantial heteroblasty (the production of markedly differ-
ent juvenile and adult leaves; Rechinger 1992; Zotz et al.
2011). Furthermore, Salix populations are dioecious, and
taxonomic keys often require examination of both sta-
minate and pistillate individuals. In many species, flow-
ering (catkin production) occurs before leaf production,
presenting an additional challenge when using keys
that consider both vegetative and reproductive traits.
Collectively, these factors can limit the characters avail-
able to identify individuals sampled at a single period
of development. Finally, hybridization may blur bound-
aries between individual species, and hybrid offspring
can exhibit highly variable morphologies (Mosseler
1990; Hardig et al. 2000).
The development of a molecular identification system
for willows, such as DNA barcoding (CBOL Plant
Working Group 2009), would be a useful tool for
applied and basic research in the genus. The organellar
genomes (plastid or mitochondrial) are currently the
source of choice for plant and animal DNA barcodes.
These genomes are haploid and typically uniparentally
transmitted and have smaller effective population sizes
than those in the diploid nuclear genome (and the dioe-
cious sexual system of Salix would be expected to
reduce effective population size further, Maynard Smith
1978). Loci in organellar genomes therefore undergo
more rapid coalescence, which in turn should reduce
confusion from incomplete lineage sorting and facilitate
species fingerprinting. However, comparative studies
have shown that while many plant groups may be ame-
nable to reasonably precise species identification using
DNA barcoding, others are less so (reviewed in Hol-
lingsworth et al. 2011). Indeed, it is now acknowledged
that plant species are generally harder to barcode than
most animal species. This may reflect various phenom-
ena, including large effective population sizes, periods
of rapid speciation, larger disparity in dispersal rates of
paternal and maternal haploid genomes and higher lev-
els of hybrid speciation or introgression (Fazekas et al.
2009; Hollingsworth et al. 2011).
One or more of these phenomena may have an
impact on the success of DNA barcoding in willows.
Our overall goal in this study was to document the
extent to which plastid DNA barcodes (CBOL Plant
Working Group 2009) can be implemented in Salix. The
more specific aims of this study were to: (i) establish
the extent of species identification that is possible
among western North American Salix using DNA bar-
code markers; (ii) to document haplotype patterns by
sampling widely and deeply (within and among spe-
cies); (iii) to establish a probable diversification time-
scale among plastid haplotypes (e.g. in the context of
historic environmental and climatic shifts in North
America); and (iv) to explore and evaluate hypotheses
for the unusual patterns of molecular diversity that we
observed.
Methods
Specimen sampling
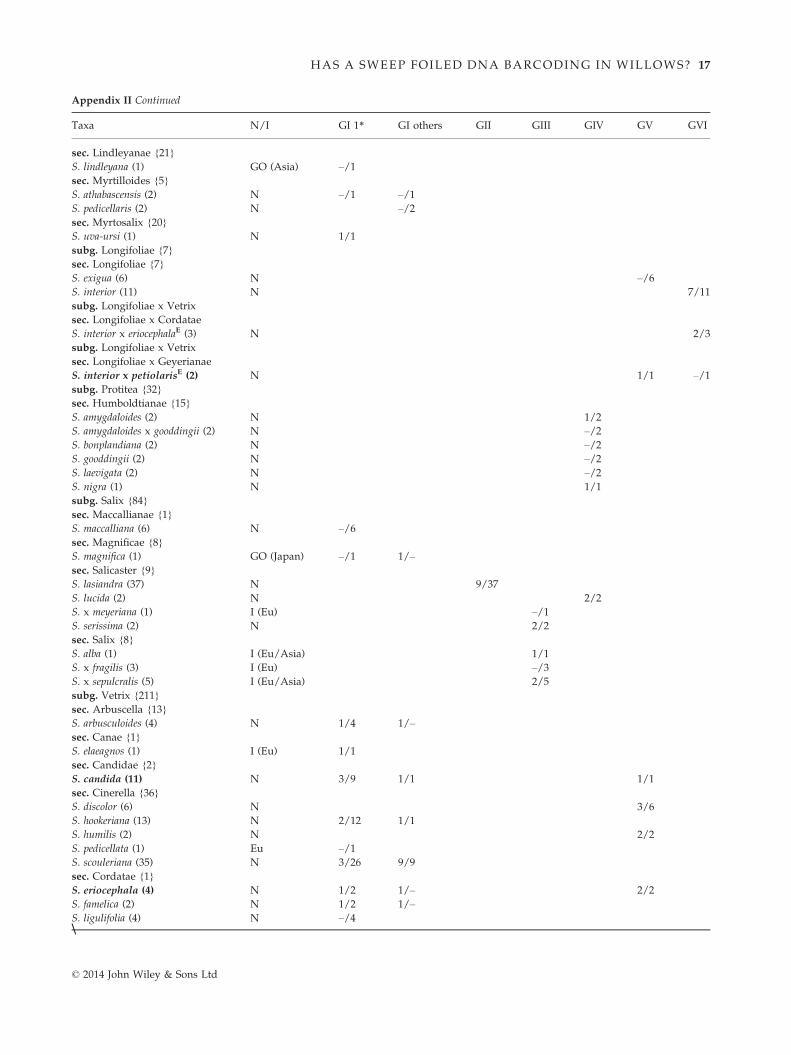
Salix species are subdivided into five subgenera (four
according to Chen et al. 2010) and are mostly found in
arctic, boreal and temperate regions (Argus 2010). Salix
is a large, diverse and widespread genus, and our sam-
pling covers a wide range of the geographic (from
North America, Asia, and Europe) and phenotypic
diversity and includes a fair representation of the broad
taxonomic diversity of the genus, with representatives
from 27 of the ~40 sections (Table 1). Our species sam-
pling focused on western North America, the most spe-
cies-rich region of the North American willow flora. We
sampled extensively within British Columbia, the prov-
ince with the greatest willow diversity in Canada, with
supplemental sampling in adjacent regions (e.g. Yukon,
Alberta) and more widely across northern temperate
regions, including species from central and eastern Can-
ada, the United States, Europe, Mexico and Japan. We
sampled a total of 546 individuals, representing 71 spe-
cies, 10 identified hybrids and an additional 10
individuals that could not be identified to species due
© 2014 John Wiley & Sons Ltd
2 D. M. PERCY ET AL.
Page 3
to vegetative-only voucher material (referred to here as
Salix spp. I-J). The sampling comprises 59% of the 113
species in the Flora of North America (Argus 2010).
Our data set includes 29 species collected in British
Columbia (BC), representing 57% of the willow flora of
the province (BC has 51 native species: 53% of the
native North American willow flora, and 65% of the
native Canadian willow flora). The BC willow flora
includes five introduced species (all are included in this
analysis); none of these appear to be extensively inva-
sive, although a few are naturalized in cultivated situa-
tions such as urban parks.
We typically sampled multiple individuals per species
(a mean of eight individuals per species), with most
(80%) represented by more than two individuals, and
only 12 (eight of these non-native) represented by a sin-
gle individual. Seven of the native, widespread, and
highly variable species were represented by more than
20 individuals, with Salix sitchensis, one of the most
widespread and phenotypically variable species in
western North America, represented by 54 individuals.
Our sampling included several voucher specimens col-
lected between 1946 and 2000; individual herbarium
samples of Salix with well preserved green leaves were
selected, and they were all successfully amplified and
sequenced (the oldest of these were collected in 1946 and
1947). We also sampled herbarium material from five
hybrid willows from a study by Mosseler (1990), derived
from experimental crosses made in 1983. We were able
to sequence all five hybrids (see Appendix II) and the
three parents for rbcL [Salix interior (maternal pistillate
parent); S. eriocephala and S. petiolaris (paternal pollen
donors)], and all of the hybrids and the two paternal
parents for all of the remaining gene regions. Voucher
specimens for new samples are housed at the University
of British Columbia and the Canadian Museum of Nat-
ure, and specimen and collection details are publicly
available on the BOLD database (project: SALIX; www.
boldsystems.org; Ratnasingham & Hebert 2007).
The identity of each individual was carefully checked
(by GWA and DMP), with assignments based on diag-
nostic morphological features of a given species, regard-
less of haplotype (although cases of highly variable
hybrid morphologies are noted below). We also care-
fully checked for sequencing error by resequencing all
individuals of species and hybrids that were placed in
more than one haplotype group, or where hybrids with
the same parents in the experimental crosses did not
carry the same haplotype.
DNA extraction, amplification, sequencing andalignment
We stored field-collected leaf tissue in silica gel and
extracted genomic DNA using a modified CTAB
method (Doyle & Doyle 1987; Rai et al. 2003). For each
individual, we surveyed up to seven plastid loci that
have been considered as candidate DNA barcoding
regions (Fazekas et al. 2008; CBOL Plant Working
Group 2009). These comprised four coding regions
(matK, rbcL, rpoB, rpoC1) and three intergenic spacer
regions (atpF-atpH, psbK-psbI, trnH-psbA). We amplified
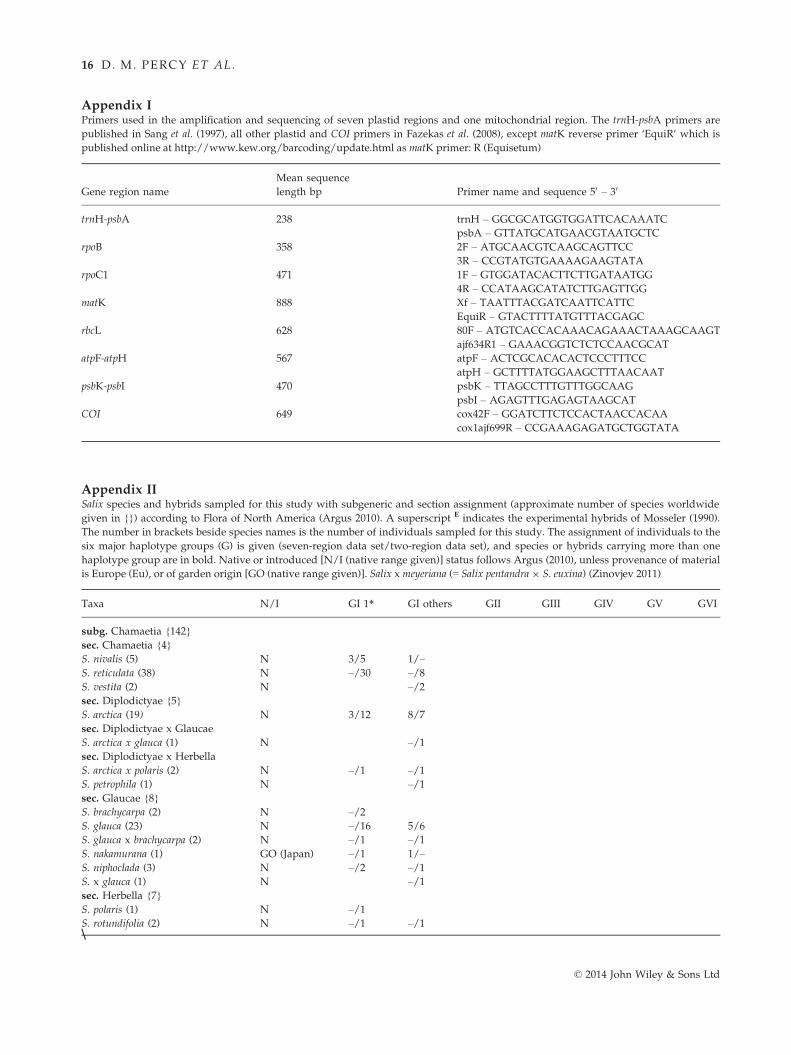
them with published primers (Appendix I) using the
thermocycler and reaction conditions noted in Rai et al.
(2003, 2008), but with the annealing temperature raised
to 53 °C for the trnH-psbA region. Bidirectional
sequencing of amplicons was performed at the Cana-
dian Centre for DNA Barcoding (CCDB, University of
Guelph; Ivanova et al. 2005). Our seven-region analyses
used 145 individuals (39 species; Appendix II). We
compiled a more extensive data set with an additional
401 samples for a total of 546 individuals (56 species)
and sequenced these for the two core plant DNA
barcoding regions, matK and rbcL (CBOL Plant Working
Group 2009) (532 of the 546 individuals were
Table 1 Salix subgenera and sections (according to Flora of
North America, Argus 2010) sampled for this study are shown
(species sampled and number of individuals sampled per
taxon relative to worldwide diversity are given in Appendix II)
with assignment to the six major haploptype groups (G)
Subgenera Section G
Chamaetia Chamaetia I
Diplodictyae I
Glaucae I
Herbella I
Lindleyanae I
Myrtilloides I
Myrtosalix I
Longifoliae Longifoliae V, VI
Protitea Humboldtianae IV
Salix Maccallianae I
Magnificae I
Salicaster II, III, IV
Salix III
Vetrix Arbuscella I
Canae I
Candidae I, V
Cinerella I, V
Cordatae I, V
Fulvae I
Geyerianae I, V
Hastatae I, V
Lanatae I
Mexicanae I
Phylicifoliae I, V
Sitchenses I
Villosae I
Viminella I
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 3
Page 4
completely sequenced for both genes). We also
sequenced cytochrome oxidase I (COI) for 30 samples
(17 species and three hybrids) with the objective of test-
ing a hypothesis that unusual aspects of the evolution-
ary history of the plastid genome (e.g. a selective
sweep) may also be detectable for the mitochondrial
genome (e.g. see Olson & McCauley 2000; Sun et al.
2014). Sequences were assembled in Sequencher 4.7
(Gene Codes Corp, Ann Arbor, MI), and Se-Al (Ram-
baut 1996) and are available from GenBank (see Data
Accessibility section below for GenBank numbers). The
plastid sequences are also archived on the BOLD data-
base (www.boldsystems.org; Ratnasingham & Hebert
2007). Phylogenetic tree descriptions and the aligned
matrices are also available from TreeBase (www.tree-
base.org). We had no detected sequence polymorphisms
suggestive of multiple plastid types (e.g. heteroplasmy)
or instances of DNA inversions. The length variations
and indels were few, and we did not treat these as
independent characters. Alignment was unproblematic,
and we did not need to exclude regions.
Analyses of molecular data
We concatenated the plastid regions for analyses, as
they belong to the same linkage group, and character-
ized major haplotype groups using heuristic maximum
parsimony (MP), distance neighbour-joining (NJ), and
maximum likelihood (ML) analyses in PAUP* (Swofford
2003) and a Bayesian analysis using BEAST (Drummond
& Rambaut 2007), described further below. The major
haplogroups as defined here represent nested sets of
alleles whose phylogenetic relationships to each other
are well supported, and whose relative times of diver-
gence can be determined in a phylogenetic context. We
restrict usage of the term ‘haplotype’ to refer to unique
variants and use haplogroup or ‘Group’ when referring
to sets of closely related haplotypes. The MP analysis
employed 100 random addition replicates and NNI
branch swapping, with MaxTrees set at 500; the NJ
analysis used the Kimura 2-parameter (K2P) and the
BioNJ option (Gascuel 1997), which is an improved NJ
method for large DNA sequences data sets. The ML
analysis considered a subset of taxa representing major
haplotype groups (i.e. unique haplotypes only; see
below) and used DNA substitution models and user-
input model parameters indicated by the Akaike Infor-
mation Criterion (AIC), as implemented in the program
MODELTEST (Posada & Crandall 1998). The ML search
used 10 random addition replicates and NNI branch
swapping. Branch support for major haplotype groups
(Table 2) was assessed using 200 bootstrap replicates
(Felsenstein 1985) in RAXML (Stamatakis 2014). For MP,
we used 10 random addition replicates for each of 200
bootstrap replicates and set MaxTrees to 100; for NJ, we
used 1000 bootstrap replicates.
Maximum parsimony and NJ analyses of the plastid
data were performed for: (i) each gene region indepen-
dently (using the seven-region data set, Appendices II);
(ii) the proposed core plant DNA barcoding combination
(CBOL Plant Working Group 2009) rbcL + matK (using
the extended sampling, Appendix II), both with and
without 13 individuals that were only sequenced for one
of the two regions); (iii) a 145 sample data set in which all
individuals are sequenced for all seven plastid regions
(Appendix II). ML and Bayesian analyses were per-
formed on a reduced version of the rbcL + matK data set
that excluded all identical haplotypes. The resulting 39
unique Salix haplotypes were aligned with sequences
from 15 additional taxa in Salicaceae and Lacistemata-
ceae obtained from GenBank, including representatives
of the genera Populus, Idesia, Poliothyrsis, Flacourtia, Xylo-
sma, Casearia, Lunania, Scyphostegia and Lacistema (see
Appendix III for species and GenBank numbers), and
henceforth, inclusion of all of these taxa (e.g. for the dat-
ing analyses described below) is referred to as a ‘full’ set
of outgroup taxa (we also performed several dating
analyses with a subset of outgroup taxa, see below). A
haplotype network diagram of the seven-region data set
for Salix was produced using Haploviewer (Salzburger
et al. 2011) with the best ML tree topology
(�lnL = �6295.796) with uninformative/missing/ambig-
uous characters removed (see Fig. 1).
To aid in interpreting alternative processes (i.e. geo-
graphical isolation-by-distance effects vs. geographically
widespread horizontal plastid capture) that may explain
patterns observed in our data, we performed two types
of statistical analyses. Although these methods, Mantel
tests and the Tajima’s D test statistic, are typically
employed for within-species analyses, the extensive
hybridization among willow species may support their
applicability to the plastid data. Furthermore, we use
these analyses to look specifically at differences
between group I and the other major haplotype groups.
We used Mantel tests (1000 iterations) performed with
the ‘Isolation-by-Distance Web Service’ (IBDWS v 3.23;
Jensen et al. 2005) to assess the strength of correlation
between geographic distances (GPS coordinate point
data transformed into pairwise distances using the Geo-
graphic Distance Matrix Generator; Ersts 2012) and
plastid genetic distances (uncorrected and K2P dis-
tances obtained from PAUP*) among the North American
individuals sampled on the seven plastid region data
set, either with all taxa or with group I only. Non-North
American samples were excluded to improve the likeli-
hood of detecting any within-continent correlation. To
take into consideration the much larger geographical
distances vs. relatively small genetic distances, we also
© 2014 John Wiley & Sons Ltd
4 D. M. PERCY ET AL.
Page 5
performed the Mantel tests with and without log-trans-
formation of the geographical distances. We used the
Tajima’s D test statistic to assess the frequency spec-
trum of selectively neutral mutations in the plastid data
using DNASP v 5 (Librado & Rozas 2009) with all sites/
mutations using the seven-region and two-gene data
sets, with either a) all taxa, b) group II-VI, or c) group I
only (Table 3). We selected a test of the overall fre-
quency spectrum of polymorphisms rather than a gene-
alogy based approach (e.g. HHT or HCT; Innan et al.
2005) due to the potentially confounding effects of lat-
eral plastid transfer on estimating species boundaries.
To characterize the timescale of diversification of
plastid markers in Salix, we estimated the ages of the
major haplotype groups based on the matK + rbcL data
set (after reduction to nonidentical sequences). We first
conducted a likelihood ratio test in PAUP*, comparing
the model with and without the molecular clock
enforced to assess whether there was significant rate
heterogeneity in the Salix data, or in Saliceae (Sa-
lix + Populus) with two sets of additional outgroups
(see below). We then performed molecular dating
analyses using a Bayesian approach in BEAST and a max-
imum likelihood approach using r8s (version 1.71, San-
derson 2006). We confirmed that rate heterogeneity
tests in PAUP* (described above) were consistent with
the likelihood ratio test performed in r8s. Our BEAST
analyses estimated mean rates of evolution, and trees
and branch lengths from the data set with the full selec-
tion of outgroup taxa using the following parameters:
substitution model GTR + Γ; clock model relaxed un-
correlated lognormal; tree prior Yule process with uni-
form distribution model; MCMC chain length of 20
million with 25% burn-in (multiple parallel analyses
were run to check for stationarity, chain convergence
and effective sample sizes). We visualized the results in
the BEAST associated programs, TRACER, TREEANNOTATOR
and FIGTREE. The ESS (effective sample size) values for
all parameters estimated were >300. The r8s analyses
used the ML tree recovered using PAUP* (described
above). We ran the r8s analyses with two different out-
groups, one with Poliothyrsis used to root a taxon set
comprising Saliceae + Idesia + Poliothyrsis, and another
with Lacistema used to root a taxon set that included the
full set of outgroup taxa (in each case the outgroup
taxon used to root the tree was pruned before diver-
gence times were estimated; Sanderson 2006). We used
the Langley–Fitch algorithm (LF) (for the taxon set com-
prising Saliceae + Idesia + Poliothyrsis) as these data
were found to satisfy a molecular clock hypothesis. We
used nonparametric rate smoothing (NPRS) and penal-
ized likelihood (PL) methods (Sanderson 1997, 2002),
with either Powell or TN (truncated Newton) algo-
rithms for the full outgroup set, as these data violated a
molecular clock. We established the optimum smooth-
ing value for the PL analysis using the cross validation
option in r8s. For all r8s analyses, we also used the
CheckGradient option as a further confirmation of the
correctness of the selected methods and algorithms.
We calibrated these analyses using several fossils.
Fossil evidence and biogeographic studies suggest a
possible warm temperate origin for Salix in North or
Central America followed by early occupation of ripar-
ian habitats (Collinson 1992; Boucher et al. 2003; Abdol-
lahzadeh et al. 2011). Subsequent range expansion into
cooler northern hemisphere habitats was likely accom-
panied by repeated advances and retreats to refugia
during glacial and interglacial periods (Ager et al. 2010).
Because the leaf characters of salicoid (gland-tipped)
teeth, camptodromous secondary venation, and elliptic,
lanceolate or deltoid shape are not unique to Salix and
Populus (Boucher et al. 2003; Cronk 2005), there are
often problems associated with interpreting fossil mate-
rial. However, the recent identification of the North
American Eocene fossil Pseudosalix (bearing
reproductive structures) as the immediate sister group
to the tribe Saliceae (Salix + Populus; Boucher et al.
2003) has provided a useful additional calibration point
for dating key events in the evolution of the family
Salicaceae, the order Malpighiales, and the origins of
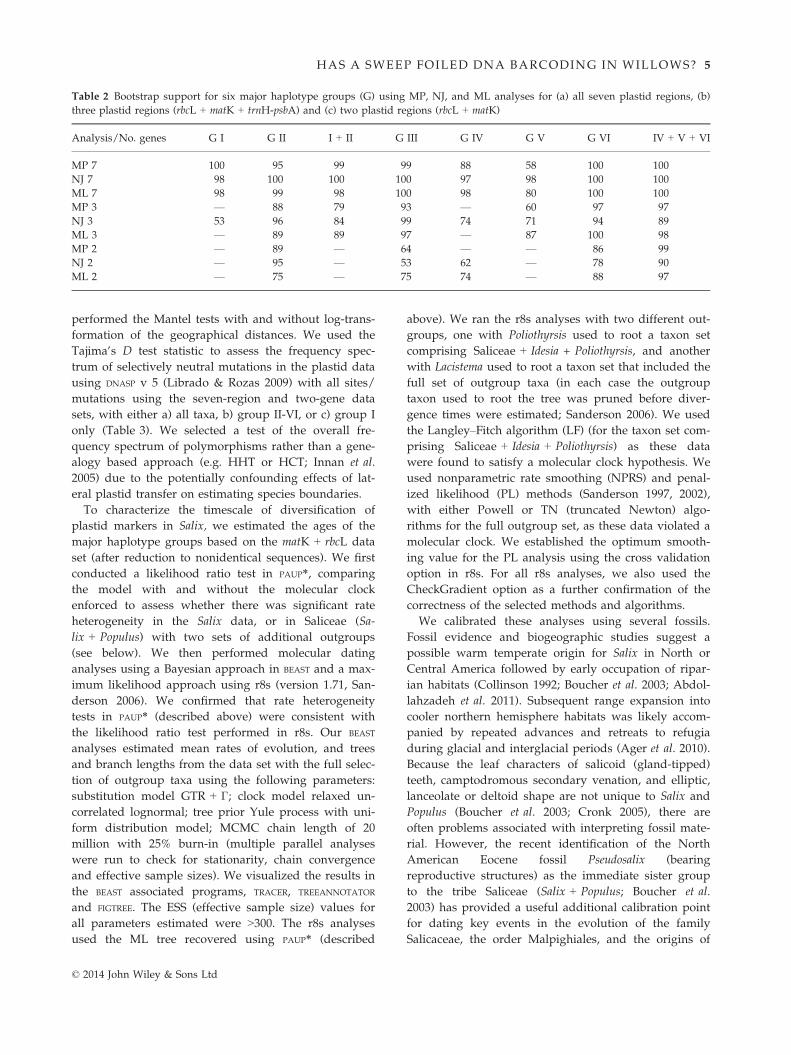
Table 2 Bootstrap support for six major haplotype groups (G) using MP, NJ, and ML analyses for (a) all seven plastid regions, (b)
three plastid regions (rbcL + matK + trnH-psbA) and (c) two plastid regions (rbcL + matK)
Analysis/No. genes G I G II I + II G III G IV G V G VI IV + V + VI
MP 7 100 95 99 99 88 58 100 100
NJ 7 98 100 100 100 97 98 100 100
ML 7 98 99 98 100 98 80 100 100
MP 3 — 88 79 93 — 60 97 97
NJ 3 53 96 84 99 74 71 94 89
ML 3 — 89 89 97 — 87 100 98
MP 2 — 89 — 64 — — 86 99
NJ 2 — 95 — 53 62 — 78 90
ML 2 — 75 — 75 74 — 88 97
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 5
Page 6
modern tropical forests (Davis et al. 2005). The earliest
unequivocal fossils of Populus and the fossil Pseudosalix
are from the early Middle Eocene Green River Forma-
tion of Utah and Colorado, 46–50 Mya (Manchester
et al. 1986; Boucher et al. 2003). These fossils consist of
leafy shoots with unisexual inflorescences, and capsular
infructescences. Earlier leaf records from the Cretaceous
and Palaeocene are thought to combine features of Pop-
ulus, Idesia and Poliothyrsis (Iljinskaya & Chelebaeva
2002; Boucher et al. 2003), and there are plausible
records for Populus from the late Palaeocene (Collinson
1992). There are, however, numerous Eocene records
for Salicaceae s.l., and by this period, Populus seems to
have become well established in North America and
Asia (Sun et al. 2004). It still remains unclear whether
the extant genus Salix was also established in the
Eocene period, despite some reports of Salix fossils
from the Green River Formation (Brown 1934; MacGini-
tie 1969; Boucher et al. 2003). We therefore used two
fossil age constraints: the Middle Eocene (Green River
Formation) Pseudosalix handleyi fossil age of 48 Mya
(Boucher et al. 2003) as a mean node age (in BEAST, stan-
dard deviation 0.5) and minimum age (in r8s) constraint
for the crown clade of Saliceae [Saliceae (Populus + Sa-
lix) vs. Idesia split], and the Casearia-type Late-Middle
Eocene (Panama) pollen age of 37 Mya (Graham 1985)
30
1* (337 individuals)
G III
IV+V+VI
2 (2 individuals) 3 (40 individuals) 4 (3 individuals) 5 (3 individuals)
7 (1 individual) 8 (4 individuals) 9 (1 individual)
10 (1 individual) 11 (1 individual) 12 (1 individual) 13 (2 individuals)
3 spp. 16 spp.
14 (10 individuals) 15 (2 individuals) 16 (3 individuals) 17 (1 individual)
53 spp.
4 spp.
2 spp. 2 spp. 2 spp.
2 spp. 18 (2 individuals) 19 (2 individuals) 20 (1 individual) 21 (3 individuals) 22 (1 individual) 23 (2 individuals) 24 (3 individuals) 25 (39 individuals) 26 (1 individual) 27 (1 individual) 28 (1 individual) 29 (21 individuals)
33 (6 individuals) 34 (6 individuals) 35 (1 individual) 36 (3 individuals) 37 (1 individual) 38 (2 individuals) 39 (2 individuals)
2 spp.
3 spp.
Unique haplotype # (# individuals)
2 spp. 2 spp.
8 spp.
2 spp.
3 spp. 3 spp.
Salix
G I
I+II
Saliceae
G II
G I
G V
G III
G VI
G IV
1*
G VI
G V
G IV
G III
G II
G I
Individuals
G I G II G III G IV G V G VI
7-region 2-region
Salix arbutifolia 6 (1 individual)
ChamaetiaSalixVetrix
Salix
Salix
ProtiteaSalix
LongifoliaeVetrix
Longifoliae
000000
Salix arctica
Salix bebbiana Salix arbutifolia Salix sitchensis
Salix sitchensis Salix calcicola Salix sitchensis Salix alaxensis Salix lasiolepis
Salix cordata
Salix drummondiana
Salix geyeriana
Salix lasiandra Salix exigua Salix petiolaris Salix exigua
Salix discolor
Salix interior
Salix serissima
30 (1 individual)
32 (4 individuals) 31 (9 individuals)
2 spp.
Populus tremuloides Populus tomentosa
Populus nigra
Populus trichocarpa Populus tremula
Salix x meyeriana
Salix x sepulcralis Salix x sepulcralis
Populus balsamifera Populus deltoides
Poliothyrsis sinensis
Flacourtia jangomas Scyphostegia borneensis
Lunania parviflora Casearia spp.
Xylosma congesta
Idesia polycarpa
Lacistema aggregatum – 0.001 substitutions/site
Fig. 1 Maximum clade credibility tree from the BEAST analysis including 39 unique matK + rbcL Salix haplotypes plus sequences
obtained from GenBank for other genera (see Appendix III for details of outgroup taxon sampling). Major haplotype group positions
(Groups I–VI) are shown and bootstrap support is given in Table 2. Inset: haplotype network diagram of the seven-region data set
with major haplotype groups and Salix subgenera (see Table 1 and Appendix II for Salix classification).
© 2014 John Wiley & Sons Ltd
6 D. M. PERCY ET AL.
Page 7
as a mean node age (in BEAST, standard deviation 0.5)
and a fixed-age (in r8s) constraint for the Casearia–Luna-
nia split. Uncertainty in the age of the calibrated node
can be partly accommodated in BEAST by applying a
prior distribution, considering the fossil age as a mini-
mum bound. An underlying assumption in applying
fossil constraints at all is that splits in the gene tree cor-
respond to splits in the species tree. This assumption
may only hold approximately within Salix and relatives.
Using relatively small standard deviations on the distri-
bution priors in BEAST, and minimum/fixed ages in r8s
may be sufficient for our purposes, as we are primarily
interested in understanding the scale and order of gene
tree splits, not the precise dates of these splits.
Results
Characteristics of individual and combined plastidregions
The overall sequencing success among the seven plastid
regions was lowest for matK at 90%, similar to other
comparative studies (Fazekas et al. 2008; Kress et al.
2009). The sequencing success for all other regions was
>95%, with the highest success for rbcL at 98%, and
trnH-psbA at 99%. Sequence characteristics for each of
the seven plastid gene regions (matK, rbcL, rpoC1, rpoB,
trnH-psbA, atpF-atpH, psbK-psbI) and the single mito-
chondrial gene region (COI), including the number of
individuals sequenced, aligned sequence length, mean
sequence length and parsimony informative characters
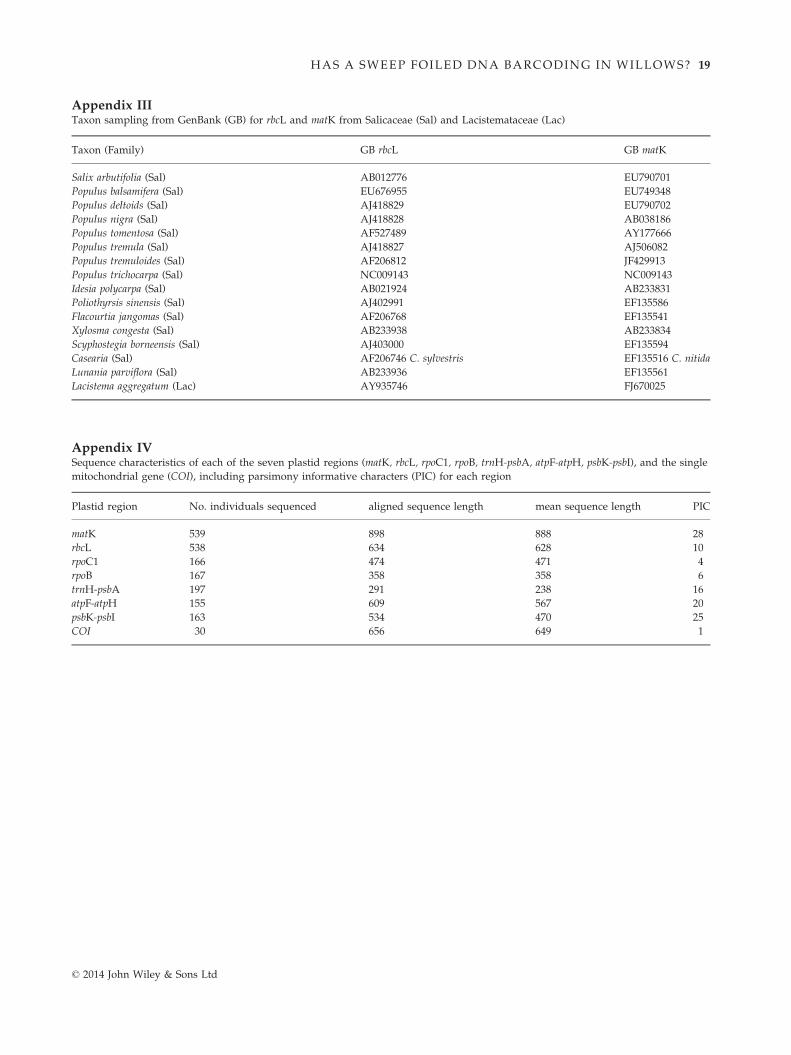
(PIC) for each region are provided in Appendices IV
and V. The matK gene provided the largest number of
phylogenetically informative characters, followed by the
noncoding regions psbK-psbI, atpF-atpH and trnH-psbA
(the latter region is relatively short in Salix; mean
238 bp) and then rbcL. The remaining two coding
regions (rpoB and rpoC1) were the most conserved. We
found only a single nucleotide polymorphism (SNP) in
the COI region, a synapomorphy for two individuals
that are in the same plastid haplotype group (I); other-
wise, the mitochondrial sequences are identical. We
observed six major haplotype groups in various indi-
vidual and combined plastid gene analyses that were in
general moderately to strongly supported by bootstrap
analysis (groups I-VI in Table 2, Fig. 1, Appendix II).
As expected, the seven regions belonging to the same
plastid linkage group yield congruent gene trees, albeit
with greater or lesser degrees of branch support
(Table 2). To illustrate the information present in each
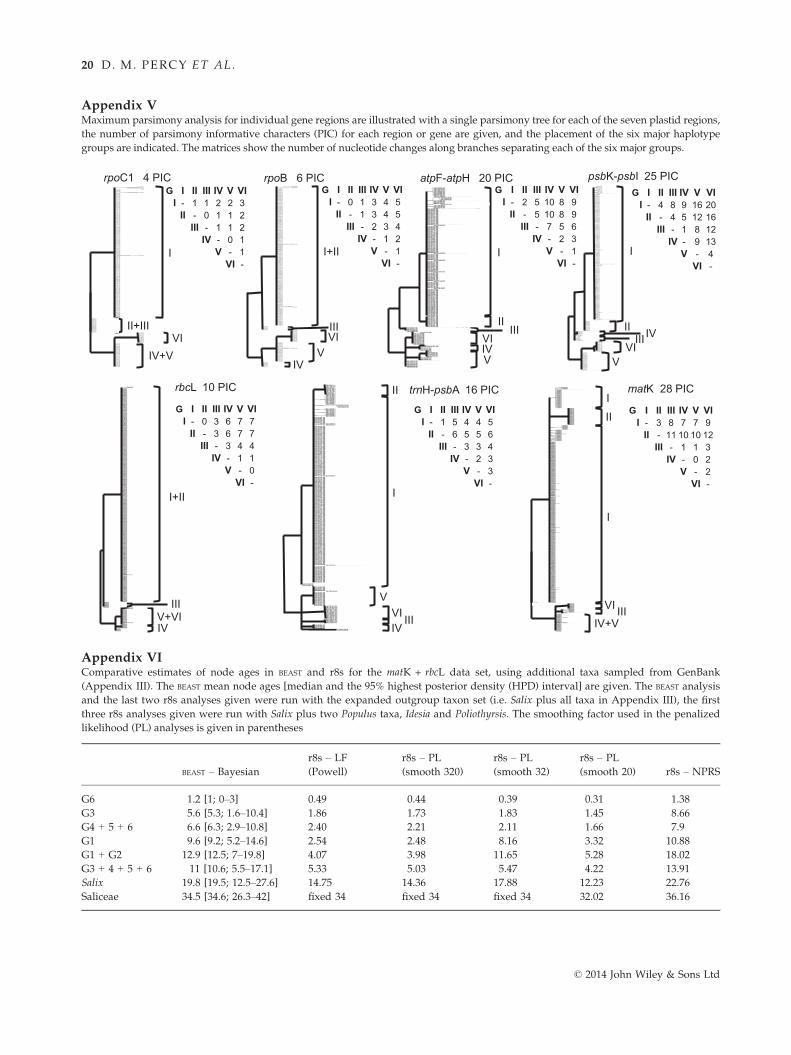
data set, Appendix V shows a single maximum parsi-
mony tree for each of the seven plastid regions analy-
sed separately, the number of parsimony informative
characters (PIC) for each gene region (see also Appen-
dix IV), and pairwise matrices that show the predicted
number of nucleotide substitutions along internal
branches separating the six major haplotype groups.
Not surprisingly, the combined seven-region analysis
(145 individuals; 3798 plastid nucleotides; 109 informa-
tive sites) provides the best support for relationships
among the six haplotype groups, although four to five
of these groups have at least 50% bootstrap support in
analysis of two- and three-region multilocus
barcoding combinations (i.e. matK + rbcL, and
matK + rbcL + trnH-psbA) (Table 2). For all of these
combinations, it is clear that the plastid phylogenies do
not delimit taxonomic species (here based on Argus
2010). Four species in subgenus Vetrix (S. candida, S. cor-
data, S. eriocephala and S. planifolia) belonging to differ-
ent sections, have different individuals that come out in
more than one major haplotype group (in all four cases
from groups I and V; Appendix II), and even within
major haplotype groups, the individuals of a particular
species may not have unique or identical haplotypes
(e.g. group I and haplotype 1*, Appendix II). In the
145-sample, seven-region data set (which comprises 40
Salix species), there are 43 unique haplotypes, and no
less than 52 individuals in 20 species carry the same
variant of haplotype (type 1* in Appendix II; identical
across 3798 bp and seven plastid regions) for some or
all of the individuals sampled for those species.
Expanded sampling of the matK + rbcL data set
The two-gene analysis used a much greater number
of samples (an additional 401 individuals), and
although less well resolved (Table 2), this broader
sampling confirms the overall patterns found in the
seven-region analysis and suggests that the haplotype
groupings based on only two genes are highly repre-
sentative of plastid distributions in willows (Appen-
dix II). With this expanded sampling, the number of
Table 3 Results of the Tajima’s D test statistic (Tajima 1989)
which tests the hypothesis that the frequency spectrum of
mutations is selectively neutral
Data set S Pi Theta Tajima’s D
All taxa, 7 genes 103 0.494 0.625 �0.67 NS
All taxa, 2 genes 31 0.128 0.406 �1.8*G I, 7 genes 38 0.043 0.229 �2.51***G I, 2 genes 15 0.016 0.192 �2.16**G II-VI, 7 genes 79 0.72 0.577 0.9 NS
G II-VI, 2 genes 20 0.383 0.296 0.85 NS
S = number of polymorphic sites; Pi = nucleotide diversity per100 sites; Theta = estimate of mutation rate (2Nel) per 100 sites;Tajima’s D = significance of rejection of neutrality: NS, P > 0.10; *,P < 0.05; **, P < 0.01; ***, P < 0.001.
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 7
Page 8
individuals (337) and species (57 spp.) found with an
identical haplotype (type 1*) surpasses those found
elsewhere in group I (although the lower number of
informative characters in the two-region vs. seven-
region analysis likely also contributes to a higher
number of individuals with identical sequence).
Henceforth, when we refer to haplotype 1*, we mean
in regard to the expanded sampling used for the
two-region analysis, unless otherwise stated, which
incorporates the taxa in the seven-region analysis
(Appendix II). With less sequence data per taxon in
the two-gene analysis, haplotype groups IV, V and VI
are less clearly distinguishable (Table 2, Fig. 1).
The expanded sampling of individuals in the
matK + rbcL analysis provides additional evidence that
divergent haplotypes are present in the four species
already identified as carrying haplotype groups I and
V in the less densely sampled seven-region analysis
(i.e. S. candida, S. cordata, S. eriocephala, and S. planifo-
lia) (Table 1, Appendix II). A subtle difference
occurred for two individuals that had only rbcL
sequence data, when incorporating those individuals
in a combined gene analysis; rbcL by itself lacks suffi-
cient information to differentiate haplotype group I
from II, and group V from VI (Appendix V). Two
other single gene analyses failed to differentiate all
six haplotype groups: rpoC1 with only four informa-
tive characters failed to differentiate group II from III,
and group IV from V; and matK, despite having 28
informative characters, failed to differentiate groups
IV from V (Appendix V). The Asian species, Salix arb-
utifolia, has haplotype group I in our analysis based
on the placement of GenBank sequences for matK
(EU790701) and rbcL (AB012776). This supports other
studies that place this species, previously placed in
the genus Chosenia, within Salix (Ohashi 2001; Chen
et al. 2010; Hardig et al. 2010).
Remarkably, of 71 willow species, only one species, S.
lasiandra, could be barcoded consistently and with confi-
dence using either the seven- or two-region barcode (it
has a distinct and unique haplotype, designated group
II, that has strong bootstrap support, ≥ 95% with seven-
regions; Fig. 1, Table 2, Appendix II). Another species,
S. interior, is the only species in haplotype group VI, and
it groups with most of the hybrid crosses using this spe-
cies as the maternal parent; there is some intra-specific
sequence variability within this haplotype group and
strong support for group VI (bootstrap support 100%
with seven-regions; Fig 1, Table 2, Appendix II).
Evidence from statistical tests
The results from the Mantel tests indicate that there is a
significant correlation between geographic and genetic
distance (the results using either uncorrected or K2P
distances were similar, and we report only the latter
here) using the seven plastid regions when all taxa are
included (P < 0.001, R2 = 0.343), but within group I
alone, there is no such correlation (P = 0.49,
R2 = 0.0002). When the geographical distance axis was
log transformed, the R2 differed (all taxa: R2 = 0.133;
group I: R2 = 0.002), but the significance (or lack) of the
correlation did not change (all taxa: P < 0.001; group I:
P = 0.2). These results add support to the hypothesis
that there is taxonomically indiscriminate and wide-
spread lateral gene transfer and spread of haplotypes,
especially haplotype 1* in group I. Here, we use the
term lateral (or horizontal) transfer to include processes
involving hybridization and introgression.
The Tajima’s D test statistic (Tajima 1989) tests the
hypothesis that the frequency spectrum of mutations is
selectively neutral. A significantly negative Tajima’s D
is expected when the data depart from neutral expecta-
tions. Our results using this test support the presence of
strong positive selection located in group I as indicated
by the highly significant negative Tajima’s D results for
this haplotype group (P < 0.001 and P < 0.01 in the
seven-region and two-gene analyses, respectively) and
the contrastingly positive Tajima’s D results when
group I is excluded (Table 3). These results demonstrate
an excess of low-frequency polymorphisms in group I,
which is indicative of a non-random process, such as a
selective sweep of the plastid genome. An alternative
explanation of an ancestral population expansion as the
source of these polymorphisms (Muir & Filatov 2007)
seems improbable, as group I consists of individuals
from many different species distributed across the
Palaearctic.
Molecular dating analyses
Our comparative dating analyses based on the
matK + rbcL reduced data set (i.e. eliminating identical
sequences), contained 54 unique haplotypes (39 Salix
and 15 samples from other genera obtained from Gen-
Bank) and comprised 1550 aligned nucleotides. The age
estimates and order of chronological events for the
divergence of the major haplotype groups are mostly
consistent between the two dating methods that we
employed (Appendix VI shows the age estimates from
the BEAST and r8s analyses with the expanded outgroup
sampling, and the r8s analyses that were run with a
reduced outgroup configuration, see Methods).
Although r8s-based dates are younger than those
obtained from BEAST (the latter shown on Fig. 2), the rel-
ative order of chronological events of clade origins is
the same between methods, and the 95% HPD (highest
posterior density interval) range obtained from the BEAST
© 2014 John Wiley & Sons Ltd
8 D. M. PERCY ET AL.
Page 9
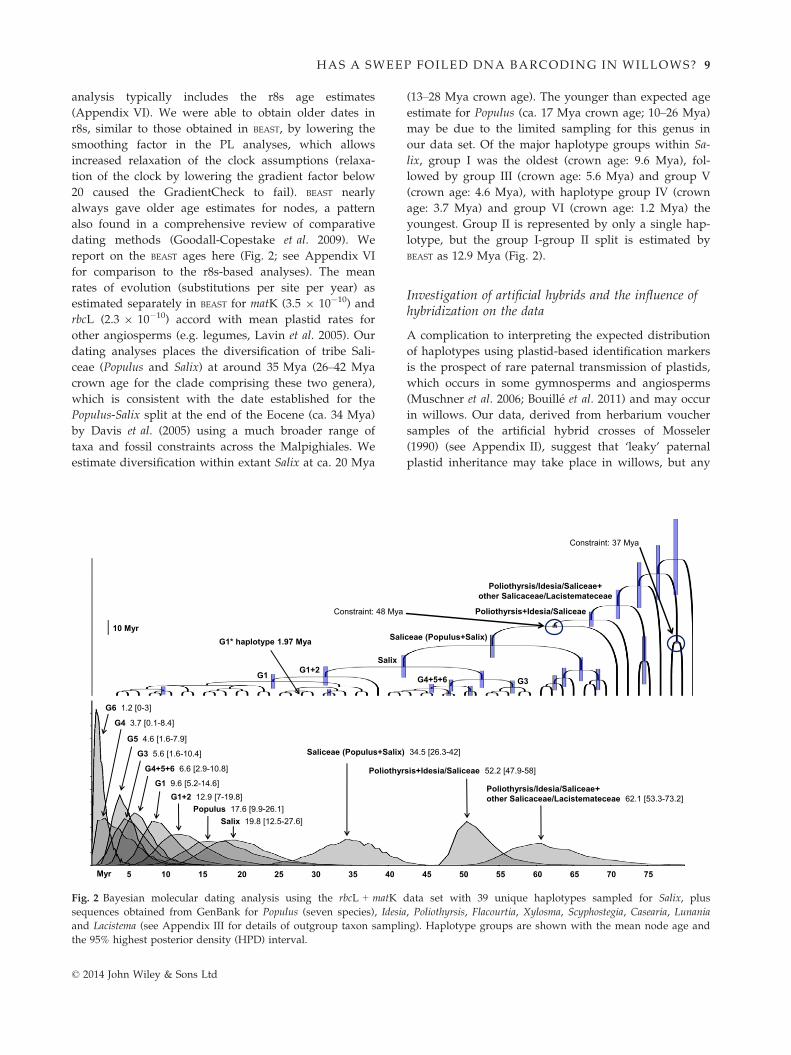
analysis typically includes the r8s age estimates
(Appendix VI). We were able to obtain older dates in
r8s, similar to those obtained in BEAST, by lowering the
smoothing factor in the PL analyses, which allows
increased relaxation of the clock assumptions (relaxa-
tion of the clock by lowering the gradient factor below
20 caused the GradientCheck to fail). BEAST nearly
always gave older age estimates for nodes, a pattern
also found in a comprehensive review of comparative
dating methods (Goodall-Copestake et al. 2009). We
report on the BEAST ages here (Fig. 2; see Appendix VI
for comparison to the r8s-based analyses). The mean
rates of evolution (substitutions per site per year) as
estimated separately in BEAST for matK (3.5 9 10�10) and
rbcL (2.3 9 10�10) accord with mean plastid rates for
other angiosperms (e.g. legumes, Lavin et al. 2005). Our
dating analyses places the diversification of tribe Sali-
ceae (Populus and Salix) at around 35 Mya (26–42 Mya
crown age for the clade comprising these two genera),
which is consistent with the date established for the
Populus-Salix split at the end of the Eocene (ca. 34 Mya)
by Davis et al. (2005) using a much broader range of
taxa and fossil constraints across the Malpighiales. We
estimate diversification within extant Salix at ca. 20 Mya
(13–28 Mya crown age). The younger than expected age
estimate for Populus (ca. 17 Mya crown age; 10–26 Mya)
may be due to the limited sampling for this genus in
our data set. Of the major haplotype groups within Sa-
lix, group I was the oldest (crown age: 9.6 Mya), fol-
lowed by group III (crown age: 5.6 Mya) and group V
(crown age: 4.6 Mya), with haplotype group IV (crown
age: 3.7 Mya) and group VI (crown age: 1.2 Mya) the
youngest. Group II is represented by only a single hap-
lotype, but the group I-group II split is estimated by
BEAST as 12.9 Mya (Fig. 2).
Investigation of artificial hybrids and the influence ofhybridization on the data
A complication to interpreting the expected distribution
of haplotypes using plastid-based identification markers
is the prospect of rare paternal transmission of plastids,
which occurs in some gymnosperms and angiosperms
(Muschner et al. 2006; Bouill�e et al. 2011) and may occur
in willows. Our data, derived from herbarium voucher
samples of the artificial hybrid crosses of Mosseler
(1990) (see Appendix II), suggest that ‘leaky’ paternal
plastid inheritance may take place in willows, but any
G4 3.7 [0.1-8.4]
G5 4.6 [1.6-7.9]
G6 1.2 [0-3]
G3 5.6 [1.6-10.4]
G4+5+6 6.6 [2.9-10.8]
G1 9.6 [5.2-14.6]G1+2 12.9 [7-19.8]
Populus 17.6 [9.9-26.1]Salix 19.8 [12.5-27.6]
Saliceae (Populus+Salix) 34.5 [26.3-42]
Poliothyrsis+Idesia/Saliceae 52.2 [47.9-58]
Poliothyrsis/Idesia/Saliceae+other Salicaceae/Lacistemateceae 62.1 [53.3-73.2]
10 Myr
5 10 15 20 25 30 35 40 45 50 55 60 65 70 75
G1
G1* haplotype 1.97 Mya
G1+2 Salix
Saliceae (Populus+Salix)
Poliothyrsis+Idesia/Saliceae
G4+5+6 G3
Poliothyrsis/Idesia/Saliceae+other Salicaceae/Lacistemateceae
Constraint: 48 Mya
Constraint: 37 Mya
Myr
Fig. 2 Bayesian molecular dating analysis using the rbcL + matK data set with 39 unique haplotypes sampled for Salix, plus
sequences obtained from GenBank for Populus (seven species), Idesia, Poliothyrsis, Flacourtia, Xylosma, Scyphostegia, Casearia, Lunania
and Lacistema (see Appendix III for details of outgroup taxon sampling). Haplotype groups are shown with the mean node age and
the 95% highest posterior density (HPD) interval.
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 9
Page 10
inferences about the degree and mode of inheritance
are limited here because we could only sequence the
individual maternal parent (S. interior) for rbcL, and this
was distinguishable only as an undifferentiated group
V + VI haplotype (providing insufficient information to
place it conclusively in either of these groups). How-
ever, the five hybrid progeny derived from the same
maternal individual and two different paternal individ-
uals are clearly assigned to two different haplotype
groups: either group V or VI; two are in group V [one
interior 9 eriocephala and one interior 9 petiolaris], and
three are in group VI [two interior 9 eriocephala and one
interior 9 petiolaris] see Appendix II). The paternal
S. eriocephala individual is one of the samples of this
species that carries haplotype group I, and the paternal
S. petiolaris individual is in group V. So, although we
cannot place the maternal parent definitively in haplo-
type group V vs. VI, we can at least say that hybrid
progeny derived from the same parental crosses are not
always found in the same haplotype group, supporting
at least occasional biparental inheritance of the plastid
genome. Repeating this experiment on a larger scale
would be needed to clearly demonstrate leaky paternal
inheritance of plastids in willows. We did not encounter
any evidence of heteroplasmy when examining the
DNA sequence chromatograms.
There are 18 taxonomic sections for which multiple
species were sampled (Appendix II), and although the
majority, 11, were found to consistently carry the same
haplotype group among the species sampled in that
section, various sections have at least one species in a
highly divergent major haplotype group (Table 1). Sec-
tion Salicaster within subgenus Salix was the most heter-
ogeneous section with individuals from three different
haplotype groups, including S. lasiandra (group II), S. x
meyeriana (S. pentandra x euxina) and S. serissima (group
III), and S. lucida (group IV). Salix lasiandra and S. luci-
da, once considered to be the same species and still
thought to be closely related based on morphology, are
found in different, highly divergent haplotype groups
(also found in Hardig et al. 2010). In addition to section
Salicaster, six other sections, five within subgenus Vetrix,
were found to have species carrying multiple major
haplotype groups, and in all cases they carried haplo-
type groups I and V. The prevalence for mixed sections
and species to carry these two particular haplotype
groups (groups I and V) appears non-random, and
group I is always more commonly carried by more spe-
cies within a taxonomic section than group V.
Discussion
Several recent studies have attempted to understand
why DNA barcode identification systems may be unable
to assign species in some groups of organisms (Fazekas
et al. 2009; von Cr€autlein et al. 2011; Arca et al. 2012). Wil-
lows have wind and insect-dispersed pollen (Argus 1974;
Vroege & Stelleman 1990), and their seeds may travel
large distances compared to species with more locally
restricted dispersal (Steyn et al. 2004). In principle, there-
fore, the presence of wind pollination in tribe Saliceae
(Populus and Salix; Boucher et al. 2003), combined with
their efficiently wind and water-dispersed seeds should
make them good candidates for barcoding identification
methods. This is because high dispersability of genes
among populations within species should help purge int-
rogressed neutral genes from other species (Currat et al.
2008; Petit & Excoffier 2009; Hollingsworth et al. 2011).
However, our study shows very low species-specific
identity for plastid markers, and so these features of wil-
low biology do not appear to be sufficient to purge for-
eign genes and ensure the success of DNA barcoding in
the genus. Here, we compare expectations and evidence
for phenomena that may contribute to the particularly
poor success of DNA barcoding markers in willows.
Retention of ancestral polymorphisms
Massive coalescence failure (incomplete lineage sorting)
and subsequent haplotype extinction events may explain
widespread sharing of particular haplotypes in willows,
especially haplotype 1* (Fig. 1), via expansions and con-
tractions during a succession of interglacial periods up to
and including after the last glacial maximum. Further-
more, coalescence failure could result from rapid radia-
tion during the early diversification of the genus, and/or
extremely large effective plastid population sizes in indi-
vidual species (however, it is worth noting that both the
haploid nature of the plastid organellar genome and the
dioecious sexual system of Salix decrease the effective
population size and increase the speed and rate of
expected coalescence times compared to nuclear genes or
hermaphroditic species). For this explanation to be tena-
ble, it would require a large number of extinction events
within major taxonomic groups to leave the current dis-
persed but uneven distribution of group I and 1* haplo-
types across the genus. Coalescence failure during the
early diversification of Salix would be expected to exhibit
random sharing or extinction of genotypes among and
across taxon groups. It is difficult to see how subsequent
lineage sorting could have led to the observed pattern of
variation among taxa, as this would require repeated sort-
ing of one variant into terminal taxa. It also seems highly
unlikely that variant 1* would not have diversified and
accumulated variation since the origin of the genus, con-
sidering the deep diversification evident in the dated gene
tree for Salix. The observed pattern of some haplotypes
being limited to within taxonomic groups (Table 1), and
© 2014 John Wiley & Sons Ltd
10 D. M. PERCY ET AL.
Page 11
of other haplotypes being found in many different taxo-
nomic groups, is not consistent with retention of ancestral
polymorphisms across numerous successive speciation
events from the early diversification of Salix, unless sort-
ing and extinction of ancestral polymorphisms occurred
consistently in a non-random fashion. Such non-random-
ness seems highly unlikely. In addition, the argument for
coalescence failure or incomplete lineage sorting is uncon-
vincing, given the dated phylogeny, because the origin of
haplotype 1* occurred recently. We estimated diversifica-
tion of the crown clade of Salix at ca. 20 Mya (13–28 Mya).
The age estimates for the six major haplotype groups in
Salix range from ca. 9 Mya (group I) to 1 Mya (group VI).
Haplotype 1* is the most widespread haplotype and
appears to have originated around 2 Mya, but may be
considerably younger given that the sequences are identi-
cal across many divergent species. It is not possible, given
our data, to determine the relative ages of the haplotype
groups vs. the ages of the species within those groups, but
we can make some assumptions that, where a morpholog-
ically determined taxonomic section is uniquely repre-
sented by a major haplotype group (e.g. Humboldtianae
represented by group IV; Table 1), that this haplotype
groupmay be at least as old as the species within it. In con-
trast, haplotype 1*, within group I, may be considerably
younger than any of the species sharing this haplotype,
which would not support the coalescence-failure
hypothesis.
Horizontal gene transfer and a selective sweephypothesis
If coalescence failure can be ruled out as an explanation
for the large-scale sharing of haplotypes in Salix, the most
obvious remaining explanation for this pattern is that it
reflects repeated capture and spread of plastids across
species and continental barriers. In principle, this is con-
sistent with extensive literature reports of hybridization in
willows, which are primarily within subgenera, but can
also be between subgenera (Argus 2010). However, if this
spread across species boundaries involved neutral genes,
we might expect a more random distribution of shared
haplotypes, although this depends on whether hybridiza-
tion patterns are truly random. Instead we observed an
overwhelming dominance of certain haplotypes, espe-
cially variant 1* (Fig. 1). This hypothesis is difficult to rec-
oncile with the breadth of both taxonomic and geographic
samples with identical or near-identical haplotypes. For
example, willowswith haplotype 1* aremixed phenotypi-
cally, taxonomically (from many different subgenera and
sections) and geographically (from North America and
Europe; Table 1 and Appendix II). The most widely
shared haplotype (type 1*) appears to have a very recent
origin (<2 Myr), and it is nested within one of the oldest
haplotype groups (group I; ca. 9 Myr). Given the large
number of different morphospecies (53 species) sharing
this identical haplotype, and the apparent rapidity and
recency of its spread, it seems doubtful that introgression
(or hybrid speciation) alone can explain the observed pat-
tern of haplotype diversity here.
Several recent studies have suggested that organellar
genomes can undergo selective sweeps in plants and ani-
mals (Muir & Filatov 2007; Lack et al. 2011). We propose
that haplotype 1* may be the result of a trans-specific
selective sweep (seeMuir & Filatov 2007 for an example in
Silene, Caryophyllaceae). The spread of an adaptive plas-
tid may be facilitated by initial plastid capture (Tsitrone
et al. 2003; Kapralov & Filatov 2007); but how well can
selection propel one or more plastid types across multiple
species boundaries? A number of studies have looked at
the spread of advantageous alleles across populations
within species (Gross et al. 2007; Presgraves et al. 2009;
Blackman et al. 2010), and within-species selective sweeps
may contribute to the cohesiveness of species (Morjan &
Rieseberg 2004). But few studies have found evidence for
trans-specific selective sweeps (e.g. Muir & Filatov 2007;
Lack et al. 2011; Brand et al. 2013; Twyford et al. 2013).
Nonetheless, this particular process should carry a detect-
able and testable signature (Muir & Filatov 2007). The Taj-
ima’s D test results (Table 3) support this hypothesis for
Salix. These imply that, for group I at least, introgression
of haplotype 1* is unlikely to have occurred via neutral
processes. Horizontal gene transfer, whether by hybrid-
ization or othermeans, may result in a faster accumulation
of genetic novelties than through mutation alone, and if
selected on also contribute to evolutionary processes
(Muir & Filatov 2007; Lucek et al. 2010; Hudson et al. 2011;
Richards et al. 2011). However, whether hybridization is
actually adaptive remains controversial (Schemske &
Morgan 1990; Mallet 2005; Twyford & Ennos 2012).
Geographically widespread and promiscuous willows,
such as present-day S. candida, S. eriocephala, S. glauca, S.
pedicellaris, S. planifolia, may have aided the transmission
of a plastid type over a wide geographic (and taxo-
nomic) range. An example of a modern-day willow
whose ancestor may have facilitated the initial stages of
plastid transmission is S. pedicellaris (group I). This spe-
cies currently has a wide native geographic range within
North America, forms natural hybrids with at least six
other species in group I, and is known to hybridize with
two of the species that have multiple haplotype groups,
S. candida and S. eriocephala (see Argus 2010; both carry
groups I + V). In addition, S. eriocephala (groups I + V)
forms natural hybrids with at least seven species from
groups I, II, IV, VI (see Argus 2010) and S. glauca has a
Holarctic distribution and forms natural hybrids with at
least eight species in group I. Finally, S. candida and S.
planifolia, both widespread in North America, and both
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 11
Page 12
carrying groups I + V, hybridize naturally with eight or
more species (Argus 2010). The ancestors of these or
other modern-day species may have been similarly pro-
miscuous, and the long-distance seed dispersal typical
of Salix species may further promote plastid spread on a
large geographic scale (Palm�e et al. 2003). In tandem
with a selective sweep, this may explain the lack of geo-
graphical (isolation by distance) effects observed here in
haplotype group I using the Mantel test.
In theory, a putative selective sweep of the plastid gen-
ome should also affect the mitochondrial genome when
both are maternally inherited (Olson & McCauley 2000;
McCauley et al. 2007), but the very different modes and
tempos of evolution in plant organelles make detection of
the same selective forces across both organellar genomes
difficult (Palmer 1990; Soria-Hernanz et al. 2008). Our
mitochondrial data were too invariable to provide evi-
dence for or against a plastid selective sweep. Although
selection on the plastid is feasible due to the number of
functionally important genes encoded by this organelle
(e.g. Kapralov & Filatov 2007), the observed pattern could
also be caused by selection on linkage groups co-inherited
with the plastid. In Salix, these not only comprise the other
cytoplasmically inherited genome, the mitochondrion,
but also recorded cases of cyto-nuclear linkage disequilib-
rium (Latta et al. 2001; Fields et al. 2014). Such patterns of
disequilibrium may result from numerous processes,
including migration, hybridization, drift, but also selec-
tion (Burke et al. 1998; Edmands & Burton 1999). There-
fore, selection on the mitochondrion or a part of the
nucleus that is maternally inherited could be responsible
for the patternwe observe andmay have swept the plastid
by coinheritance.
To date, there are no existing, well sampled and
robustly resolved nuclear phylogenies for the genus Sa-
lix. Two recent studies that include plastid and nuclear
data for a limited number of taxa and gene regions (Har-
dig et al. 2010; Abdollahzadeh et al. 2011) have both
found nonmonophyly of taxa, and in the case of Hardig
et al., incongruence between plastid and nuclear data
and better agreement between the taxonomic classifica-
tion of Salix and nuclear data. Our current understanding
of Salix taxonomy comes from extensive morphological
studies (e.g. Argus 2010) and current molecular data
remains too limited to assist in improving species classifi-
cations (an exception being the synonomization of the
genus Chosenia with Salix). Therefore, addressing the
genetic/phylogenetic extent of species boundaries of Sa-
lix taxa should be a high priority of future research as it
affects our interpretation of when gene transfers are truly
lateral. This is an important caveat of any study, like
ours, that relies on classical species definitions to under-
stand trans-specific sharing of alleles. To further test the
hypothesis of a selective sweep in Salix will require
extensive sampling of taxa and nuclear regions. Addi-
tional data from the nuclear genome is needed to fully
test our hypothesis that the pattern observed here is a
result of a trans-specific selective sweep affecting (at
least) the plastid genome.
Significance of the mode of inheritance
Our analysis of archived material used in experimental
crosses suggests that there is primarily maternal inheri-
tance in willows, but likely accompanied by limited pater-
nal transmission. The material we had access to was not
suitable for definitively answering the mode of plastid
inheritance in Salix, but resolving this issue would be use-
ful for tracking which parent contributed captured orga-
nellar genomes, and for better understanding the
dynamics of gene flow during hybridization. The latter
may differ if the ‘invading’ genome came maternally
(through seeds) or paternally (through pollen). However,
while a selective sweep could be affected by the fine
details of the inheritance mode (e.g. whether seeds or pol-
len travel further), the overall pattern that we observed
could reasonably be expected to occur with either mode of
inheritance. We do not know if introgression is expected
to be more frequent given a predominant mode of orga-
nellar inheritance via pollen or seed.
Conclusions
This study highlights the serious challenges to the use of
plastid data for the barcoding of willows. The only
willow species in our sample that was consistently distin-
guishable using plastid barcoding regions was S. lasian-
dra. The lack of interspecific variation, the occurrence of
multiple species sharing identical haplotypes, combined,
in some cases, with considerable intraspecific variation
and species with multiple divergent haplotypes, all serve
to confound the use of plastid data to identify willow spe-
cies. The willows provide an extreme example of how
DNA barcoding can fail. At the same time, this study
illustrates how the drive to barcode the world’s organ-
isms can lead to insights into, not only the extent of fail-
ure or success expected when barcoding a particular
group, but also the possible evolutionary mechanisms of
this taxon-specific variability (or lack thereof). It is clear
that a reliable barcode for every willow species will never
be achieved using the plastid as the sole source for DNA
markers. Species assignment in the willows using plastid
data is simply not possible for nearly all species, and so
plant DNA barcoding with plastid loci will not be useful
for applications like ecological surveys, identification of
riparian indicator and/or rare species. At most, these
data provide haplotype group assignments that are
nonrandom and have some consistency at the subgeneric
© 2014 John Wiley & Sons Ltd
12 D. M. PERCY ET AL.
Page 13
and sectional levels. The unusual patterns of haplotype
group assignment in willows can likely be explained by a
combination of factors that include interglacial demo-
graphic history of populations, patterns and frequency of
hybridization, and theories of haplotype spread such as
selective sweeps. We have described here the potential
patterns and signature of a trans-specific selective sweep,
in particular the presence of a recently evolved trans-
species haplotype. Past studies that have attributed the
failure of plastid data to track species boundaries to pro-
cesses such as lack of variation, hybridization, introgres-
sion, and incomplete lineage sorting, might also usefully
look at the relative ages of the haplotypes involved to
assess how widespread this phenomenon may be in taxo-
nomically complex plant groups.
Acknowledgements
We are grateful to Genome Canada for financial support
through the Ontario Genomics Institute to the Canadian Bar-
code of Life Network. We thank those who assisted with
sequencing: Maria Kuzmina (University of Guelph) and Doro-
thy Cheung (University of British Columbia). We thank four
anonymous reviewers for help improving the manuscript, and
for useful discussions on population genetics and molecular
dating, we are grateful to Armando Geraldes and William Illes.
For assistance in the field we thank Cecilia Alstrom-Rapaport,
Bruce Bennett, Ying Chang, Dominique Collett, Robert Foottit,
Carl-Eric Granfelt, Jason Grant, Eric Maw, Nancy Moore, Jef-
frey Saarela, John Wardell, and David White.
References
Abdollahzadeh A, Kazempour Osaloo S, Maassoumi AA (2011)
Molecular phylogeny of the genus Salix (Salicaceae) with an
emphasize to its species in Iran. Iranian Journal of Botany, 17,
244–253.
Ager TA, Phillips RL (2008) Pollen evidence for Late Pleisto-
cene Bering land and bridge environments from Norton
Sound, northeastern Bering Sea, Alaska. Arctic, Antarctic, and
Alpine Research, 40, 451–461.
Ager TA, Carrara PE, McGeehin JP (2010) Ecosystem develop-
ment in the Girdwood area, south-central Alaska, following
late Wisconsin glaciation. Canadian Journal of Earth Sciences,
47, 971–985.
Arca M, Hinsinger DD, Cruaud C et al. (2012) Deciduous trees
and the application of universal DNA barcodes: a case study
on the circumpolar Fraxinus. PLoS ONE, 7, e34089.
Argus GW (1974) An experimental study of hybridization and
pollination in Salix (willows). Canadian Journal of Botany, 52,
1613–1619.
Argus GW (2010) Salix. In: Flora of North America Editorial
Committee, eds. 1993 + . Flora of North America North of
Mexico. 16 + vols. Flora of North America Association, New
York and Oxford. Vol. 7, 23–162.
Blackman BK, Strasburg JL, Michaels SD et al. (2010) The role
of recently derived FT paralogs in sunflower domestication.
Current Biology, 20, 629–635.
Boucher LD, Manchester SR, Judd WS (2003) An extinct genus
of Salicaceae based on twigs with attached flowers, fruits
and foliage from the Eocene Green River Formation of Utah
and Colorado. American Journal of Botany, 90, 1389–1399.Bouill�e M, Senneville S, Bousquet J (2011) Discordant mtDNA
and cpDNA phylogenies indicate geographic speciation and
reticulation as driving factors for the diversification of the
genus Picea. Tree Genetics and Genomes, 7, 469–484.Brand CL, Kingan SB, Wu L et al. (2013) A selective sweep
across species boundaries in Drosophila. Molecular Biology and
Evolution, 30, 2177–2186.
Brown RW (1934) The recognizable species of the Green River
flora. United States Geological Survey Professional Paper, 185-C,
45–77.Burke JM, Voss TJ, Arnold ML (1998) Genetic interactions and
natural selection in Louisiana Iris hybrids. Evolution, 52,
1304–1310.
CBOL Plant Working Group (2009) A DNA barcode for land
plants. Proceedings of the National Academy of Sciences, USA,
106, 12794–12797.Chen J-H, Sun H, Wen J et al. (2010) Molecular phylogeny of
Salix L. (Salicaceae) inferred from three chloroplast datasets
and its systematic implications. Taxon, 59, 29–37.
Collinson ME (1992) The early fossil history of Salicaceae: a
brief review. Proceedings of the Royal Society of Edinburgh, 98B,
155–167.von Cr€autlein M, Korpelainen H, Pietil€ainen M et al. (2011)
DNA barcoding: a tool for improved taxon identification and
detection of species diversity. Biodiversity and Conservation,
20, 373–389.
Cronk QCB (2005) Plant eco-devo: the potential of poplar as a
model organism. New Phytologist, 166, 39–48.
Currat M, Ruedi M, Petit RJ et al. (2008) The hidden side of
invasions: massive introgression by local genes. Evolution, 62,
1908–1920.Davis CC, Webb CO, Wurdack KJ et al. (2005) Explosive radia-
tion of Malpighiales supports a mid-cretaceous origin of
modern tropical rain forests. The American Naturalist, 165,
E36–E65.Doyle JJ, Doyle JL (1987) A rapid isolation procedure for small
quantities of fresh leaf tissue. Phytochemical Bulletin, 19, 11–15.Drummond AJ, Rambaut A (2007) BEAST: Bayesian evolutionary
analysis by sampling trees. BMC Evolutionary Biology, 7, 214.
Edmands S, Burton RS (1999) Cytochrome C oxydase activity
in interpopulation hybrids of a marine copepod: a test of
nuclear-nuclear or nuclear-cytoplasmic coadaptation. Evolu-
tion, 53, 1972–1978.Ersts PJ (2012) Geographic Distance Matrix Generator (version
1.2.3). American Museum of Natural History, Center for Bio-
diversity and Conservation. Available from http://biodiver-
sityinformatics.amnh.org/open_source/gdmg. Accessed on
29 July 2012.
Fazekas AJ, Burgess KS, Kesanakurti PR et al. (2008) Multiple
multilocus DNA barcodes from the plastid genome discrimi-
nate plant species equally well. PLoS ONE, 3, e2802.
Fazekas AJ, Kesanakurti PR, Burgess KS et al. (2009) Are plant
species harder to discriminate than animal species using
DNA barcoding markers? Molecular Ecological Resources, 9,
130–139.Felsenstein J (1985) Confidence limits on phylogenies: an
approach using the bootstrap. Evolution, 39, 783–791.
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 13
Page 14
Fields PD, McCauley DE, McAssey EV et al. (2014) Patterns of
cyto-nuclear linkage disequilibrium in Silene latifolia: genomic
heterogeneity and temporal stability. Heredity, 112, 99–104.
Gascuel O (1997) BIONJ: an improved version of the NJ algo-
rithm based on a simple model of sequence data. Molecular
Biology and Evolution, 14, 685–695.Goodall-Copestake WP, Harris DJ, Hollingsworth PM (2009)
The origin of a mega-diverse genus Begonia (Begoniaceae)
using alternative datasets, calibrations and relaxed clock
methods. Botanical Journal of the Linnean Society, 159, 363–380.Graham A (1985) Studies in neotropical paleobotany. IV. The
Eocene communities of Panama. Annals of the Missouri Botan-
ical Garden, 72, 504–534.
Gross BL, Turner KG, Rieseberg LH (2007) Selective sweeps in
the homoploid hybrid species Helianthus deserticola: evolution
in concert across populations and across origins. Molecular
Ecology, 16, 5246–5258.
Hardig TM, Brunsfeld SJ, Fritz RS et al. (2000) Morphological
and molecular evidence for hybridization and introgression
in a willow (Salix) hybrid zone. Molecular Ecology, 9, 9–24.Hardig TM, Anttila CK, Brunsfeld SJ (2010) A phylogenetic
analysis of Salix (Salicaceae) based on matK and ribosomal
DNA sequence data. Journal of Botany, ID 197696, 12.
Hassel K, Segreto R, Ekrem T (2013) Restricted variation in plant
barcoding markers limits identification in closely related bryo-
phyte species.Molecular Ecology Resources, 13, 1047–1057.Hollingsworth PM, Graham SW, Little DP (2011) Choosing and
using a plant DNA barcode. PLoS ONE, 6, e19254.
Hudson AG, Vonlanthen P, Seehausen O (2011) Rapid parallel
adaptive radiations from a single hybridogenic ancestral
population. Proceedings of the Royal Society B-Biological Sci-
ences, 278, 58–66.
Iljinskaya IA, Chelebaeva AI (2002) A new fossil genus Utkholo-
kia combining leaf characters of Populus (Salicaceae) with
those of Idesia and Poliothyrsis (Flacourtiaceae). Botanicheskii
Zhurnal, 87, 101–110.
Innan H, Zhang K, Marjoram P et al. (2005) Statistical tests of the
coalescentmodel basedon thehaplotype frequencydistribution
andthenumberofsegregatingsites.Genetics,169, 1763–1777.Ivanova NV, DeWaard JR, Hajibabaei M et al. (2005) Protocols for
high volume DNA barcoding. http://www.dnabarcoding.ca/
(accessed 20March 2014).
Jensen JL, Bohonak AJ, Kelley ST (2005) Isolation by distance,
web service. BMC Genetics 6, 13. http://ibdws.sdsu.edu/
(accessed 20 March 2014).
Kapralov MV, Filatov DA (2007) Widespread positive selection
in the photosynthetic Rubisco enzyme. BMC Evolutionary
Biology, 7, 73.
Kress WJ, Erickson DL, Jones FA et al. (2009) Plant DNA bar-
codes and a community phylogeny of a tropical forest
dynamics plot in Panama. Proceedings of the National Academy
of Sciences, USA, 106, 18621–18626.
Kuzovkina YA, Quigley MF (2005) Willows beyond wetlands:
uses of Salix L. species for environmental projects. Water, Air
and Soil Pollution, 162, 183–204.Kuzovkina YA, Volk TA (2009) The characterization of willow
(Salix L.) varieties for use in ecological engineering applica-
tions: co-ordination of structure, function and autecology.
Ecological Engineering, 35, 1178–1189.Lack JB, Nichols RD, Wilson GM et al. (2011) Genetic signa-
ture of reproductive manipulation in the phylogeography of
the bat fly, Trichobius major. Journal of Heredity, 102, 705–
718.
Latta RG, Linhart YB, Mitton JB (2001) Cytonuclear disequilib-
rium and genetic drift in a natural population of ponderosa
pine. Genetics, 158, 843–850.
Lavin M, Herendeen P, Wojciechowski MF (2005) Evolutionary
rates analysis of Leguminosae implicates a rapid diversifica-
tion of lineages during the Tertiary. Systematic Biology, 54,
530–549.
Librado P, Rozas J (2009) DNASP v5: A software for comprehen-
sive analysis of DNA polymorphism data. Bioinformatics, 25,
1451–1452.Lucek K, Roy D, Bezault E et al. (2010) Hybridization between
distant lineages increases adaptive variation during a biolog-
ical invasion: stickleback in Switzerland. Molecular Ecology,
19, 3995–4011.MacGinitie HD (1969) The Eocene Green River flora of north-
western Colorado and northeastern Utah. University of Cali-
fornia Publications in Geological Sciences, 83, 1–203.
Mallet J (2005) Hybridization as an invasion of the genome.
Trends in Ecology and Evolution, 20, 229–237.
Manchester SR, Dilcher DL, Tidwell WD (1986) Interconnected
reproductive and vegetative remains of Populus (Salicaceae)
from the Middle Eocene Green River Formation, northeast-
ern Utah. American Journal of Botany, 73, 156–160.
Maynard Smith J (1978) The Evolution of Sex, p. 236. Cambridge
University Press, Cambridge, UK.
McCauley DE, Sundby AK, BaileyMF et al. (2007) Inheritance of chlo-
roplast DNA is not strictly maternal in Silene vulgaris (Caryophylla-
ceae): evidence from experimental crosses and natural populations.
American Journal of Botany, 94, 1333–1337.Morjan CL, Rieseberg LH (2004) How species evolve collec-
tively: implications of gene flow and selection for the spread
of advantageous alleles. Molecular Ecology, 13, 1341–1356.
Mosseler A (1990) Hybrid performance and species crossability
relationships in willows (Salix). Canadian Journal of Botany,
68, 2329–2338.Muir G, Filatov D (2007) A selective sweep in the chloroplast
DNA of dioecious Silene (Section Elisanthe). Genetics, 177,
1239–1247.
Muschner VC, Lorenz-Lemke AP, Vecchia M et al. (2006) Dif-
ferential organellar inheritance in Passiflora’s (Passifloraceae)
subgenera. Genetica, 128, 449–453.Myers-Smith IH, Forbes BC, Wilmking M et al. (2011) Shrub
expansion in tundra ecosystems: dynamics, impacts and
research priorities. Environmental Research Letters, 6, 045509.
Ohashi H (2001) Salicaceae of Japan. Scientific Report, Tohoku
University, 4th Series Biology, 40, 269–396.
Olson MS, McCauley DE (2000) Linkage disequilibrium and
phylogenetic congruence between chloroplast and mitochon-
drial haplotypes in Silene vulgaris. Proceedings of the Royal
Society, Biological Sciences, 267, 1801–1808.
Palm�e AE, Semerikov V, Lascoux M (2003) Absence of geo-
graphical structure of chloroplast DNA variation in sallow,
Salix caprea L. Heredity, 91, 465–474.Palmer JD (1990) Contrasting modes and tempos of genome evo-
lution in land plant organelles. Trends in Genetics, 6, 115–120.Percy DM, Garver AM, Wagner WL et al. (2008) Progressive
island colonization and ancient origin of Hawaiian Metrosid-
eros (Myrtaceae). Proceedings of the Royal Society, B-Biological
Sciences, 275, 1479–1490.
© 2014 John Wiley & Sons Ltd
14 D. M. PERCY ET AL.
Page 15
Petit RJ, Excoffier L (2009) Gene flow and species delimitation.
Trends in Ecology & Evolution, 24, 386–393.Posada D, Crandall KA (1998) MODELTEST: testing the model of
DNA substitution. Bioinformatics, 14, 817–818.Presgraves DC, G�erard PR, Cherukuri A, Lyttle TW (2009)
Large-scale selective sweep among segregation distorter
chromosomes in African populations of Drosophila melanogas-
ter. PLoS Genetics, 5, e1000463.
Rai HS, O’Brien HE, Reeves PA et al. (2003) Inference of higher-
order relationships in the cycads from a large chloroplast data
set.Molecular Phylogenetics and Evolution, 29, 350–359.
Rai HS, Reeves PA, Peakall R et al. (2008) Inference of higher-
order conifer relationships from a multilocus plastid data
set. Botany, 86, 658–669.Rambaut A (1996) Se-Al: Sequence Alignment Editor, Version 2.
Oxford University, Oxford.
Ratnasingham S, Hebert PDN (2007) BOLD: The Barcode of Life
Data System. Molecular Ecology Notes, 7, 355–364.Rechinger KH (1992) Salix taxonomy in Europe – problems,
interpretations, observations. Proceedings of the Royal Society
of Edinburgh, 98B, 1–12.
Richards TA, Soanes DM, Jones MDM et al. (2011) Horizontal
gene transfer facilitated the evolution of plant parasitic
mechanisms in the oomycetes. Proceedings of the National
Academy of Sciences, USA, 108, 15258–15263.
Salzburger W, Ewing G, Haeseler A (2011) The performance
of phylogenetic algorithms in estimating haplotype gene-
alogies with migration. Molecular Ecology, 20, 1952–1963.
Sanderson MJ (1997) A nonparametric approach to estimating
divergence times in the absence of rate constancy. Molecular
Biology and Evolution, 14, 1218–1231.Sanderson MJ (2002) Estimating absolute rates of molecular
evolution and divergence times: a penalized likelihood
approach. Molecular Biology and Evolution, 19, 101–109.
Sanderson MJ (2006) r8s version 1.71. Analysis of rates (‘r8s’)
of evolution. Section of Evolution and Ecology. Univeristy
of California, Davis, http://loco.biosci.arizona.edu/r8s/
(accessed 15 January 2012).
Sang T, Crawford DJ, Stuessy TF (1997) Chloroplast DNA phy-
logeny, reticulate evolution and biogeography of Paeonia
(Paeoniaceae). American Journal of Botany, 84, 1120–1136.Schemske DW, Morgan MT (1990) The evolutionary signifi-
cance of hybridization in Eucalyptus. Evolution, 44, 2150–2151.Soria-Hernanz DF, Braverman JM, Hamilton MB (2008) Parallel
rate heterogeneity in chloroplast and mitochondrial genomes
of Brazil nut trees (Lecythidaceae) is consistent with lineage
effects. Molecular Biology and Evolution, 25, 1282–1296.Stamatakis A (2014) RAXML Version 8: A Tool for Phylogenetic
Analysis and Post-Analysis of Large Phylogenies. In Bioinformat-
ics, 2014, open access.
Starr JR, Naczi RF, Chouinard BN (2009) Plant DNA barcodes
and species resolution in sedges (Carex, Cyperaceae). Molecu-
lar Ecology Resources, Suppl. s1, 151–163.Steyn EMA, Smith GF, Van Wyk AE (2004) Functional and tax-
onomic significance of seed structure in Salix mucronata (Sal-
icaceae). Bothalia, 34, 53–59.
Sun B, Yan D, Xie S, Cong P, Xin C, Yun F (2004) Palaeogene
fossil Populus leaves from Lanzhou Basin and their palaeocli-
matic significance. Chinese Science Bulletin, 49, 1494–1501.Sun Y, Abbott RJ, Li L et al. (2014) Evolutionary history of pur-
ple cone spruce Picea purpurea in the Qinghai-Tibet Plateau:
homoploid hybrid origin and Pleistocene expansion. Molecu-
lar Ecology, 23, 343–359.Swofford DL (2003) PAUP*: Phylogenetic Analysis Using Parsi-
mony (*and Other Methods), Version 4. Sinauer, Sunderland,
Massachusetts.
Tajima F (1989) Statistical method for testing the neutral mutation
hypothesis by DNA polymorphism. Genetics, 123, 585–595.
Tsitrone A, Kirkpatrick M, Levin DA (2003) A model for chlo-
roplast capture. Evolution, 57, 1776–1782.
Twyford AD, Ennos RA (2012) Next-generation hybridization
and introgression. Heredity, 108, 179–189.
Twyford AD, Kidner CA, Harrison N et al. (2013) Population
history and seed dispersal in widespread Central American
Begonia species (Begoniaceae) inferred from plastome-derived
microsatellite markers. Botanical Journal of the Linnean Society,
171, 260–276.Vroege PW, Stelleman P (1990) Insect and wind pollination in Salix
repens L. and Salix caprea L. Israel Journal of Botany, 39, 125–132.Xu B, Wu N, Gao XF et al. (2012) Analysis of DNA sequences
of six chloroplast and nuclear genes suggests incongruence,
introgression, and incomplete lineage sorting in the evolu-
tion of Lespedeza (Fabaceae). Molecular Phylogenetics and Evo-
lution, 62, 346–358.
Yu WB, Huang PH, Li DZ et al. (2013) Incongruence between
nuclear and chloroplast DNA phylogenies in Pedicularis sec-
tion Cyathophora (Orobanchaceae). PLoS ONE, 8, e74828.
Zinovjev AG (2011) Salix x meyeriana (= Salix pentandra x S. eux-
ina) – a forgotten willow in Eastern North America. Phytotax-
a, 22, 57–60.Zotz G, Wilhelm K, Becker A (2011) Heteroblasty – a review.
The Botanical Review, 77, 109–151.
D.P., Q.C., S.G. and S.B. planned and wrote the first draft
of the manuscript. All authors contributed to subsequent
revisions and editing of the manuscript. The following
authors contributed to specimen sampling and identifica-
tion (D.P., G.A., Q.C., A.F., P.K., K.B., B.H., S.N.); lab
work and sequencing (D.P., A.F., P.K., K.B.); combining
molecular genetics and ecological theory (Q.C., B.H.).
Data accessibility
DNA sequences: Genbank accessions KM001945-
KM002483 (matK), KM002817-KM003354 (rbcL),
KM002484-KM002650 (rpoB), KM002651-KM002816
(rpoC1), KM003552-KM003706 (atpF-atpH), KM003707-
KM003869 (psbK-psbI), KM003355-KM003551 (trnH-
psbA), KM001915-KM001944 (COI).
Final DNA sequence assembly: aligned sequences
available from BOLD database and GenBank.
Phylogenetic data: TreeBASE Study accession no.
TB2:S15979.
Specimen information, sampling locations, and DNA
sequences and alignments uploaded to Dryad (doi:10.
5061/dryad.qf6t5).
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 15
Page 16
Appendix IPrimers used in the amplification and sequencing of seven plastid regions and one mitochondrial region. The trnH-psbA primers are
published in Sang et al. (1997), all other plastid and COI primers in Fazekas et al. (2008), except matK reverse primer ‘EquiR’ which is
published online at http://www.kew.org/barcoding/update.html as matK primer: R (Equisetum)
Gene region name
Mean sequence
length bp Primer name and sequence 50 – 30
trnH-psbA 238 trnH – GGCGCATGGTGGATTCACAAATC
psbA – GTTATGCATGAACGTAATGCTC
rpoB 358 2F – ATGCAACGTCAAGCAGTTCC
3R – CCGTATGTGAAAAGAAGTATA
rpoC1 471 1F – GTGGATACACTTCTTGATAATGG
4R – CCATAAGCATATCTTGAGTTGG
matK 888 Xf – TAATTTACGATCAATTCATTC
EquiR – GTACTTTTATGTTTACGAGC
rbcL 628 80F – ATGTCACCACAAACAGAAACTAAAGCAAGT
ajf634R1 – GAAACGGTCTCTCCAACGCAT
atpF-atpH 567 atpF – ACTCGCACACACTCCCTTTCC
atpH – GCTTTTATGGAAGCTTTAACAAT
psbK-psbI 470 psbK – TTAGCCTTTGTTTGGCAAG
psbI – AGAGTTTGAGAGTAAGCAT
COI 649 cox42F – GGATCTTCTCCACTAACCACAA
cox1ajf699R – CCGAAAGAGATGCTGGTATA
Appendix IISalix species and hybrids sampled for this study with subgeneric and section assignment (approximate number of species worldwide
given in {}) according to Flora of North America (Argus 2010). A superscript E indicates the experimental hybrids of Mosseler (1990).
The number in brackets beside species names is the number of individuals sampled for this study. The assignment of individuals to the
six major haplotype groups (G) is given (seven-region data set/two-region data set), and species or hybrids carrying more than one
haplotype group are in bold. Native or introduced [N/I (native range given)] status follows Argus (2010), unless provenance of material
is Europe (Eu), or of garden origin [GO (native range given)]. Salix x meyeriana (= Salix pentandra 9 S. euxina) (Zinovjev 2011)
Taxa N/I GI 1* GI others GII GIII GIV GV GVI
subg. Chamaetia {142}sec. Chamaetia {4}S. nivalis (5) N 3/5 1/–
S. reticulata (38) N –/30 –/8
S. vestita (2) N –/2sec. Diplodictyae {5}S. arctica (19) N 3/12 8/7
sec. Diplodictyae x Glaucae
S. arctica x glauca (1) N –/1sec. Diplodictyae x Herbella
S. arctica x polaris (2) N –/1 –/1S. petrophila (1) N –/1
sec. Glaucae {8}S. brachycarpa (2) N –/2
S. glauca (23) N –/16 5/6
S. glauca x brachycarpa (2) N –/1 –/1
S. nakamurana (1) GO (Japan) –/1 1/–S. niphoclada (3) N –/2 –/1
S. x glauca (1) N –/1sec. Herbella {7}S. polaris (1) N –/1S. rotundifolia (2) N –/1 –/1\
© 2014 John Wiley & Sons Ltd
16 D. M. PERCY ET AL.
Page 17
Taxa N/I GI 1* GI others GII GIII GIV GV GVI
sec. Lindleyanae {21}S. lindleyana (1) GO (Asia) –/1sec. Myrtilloides {5}S. athabascensis (2) N –/1 –/1S. pedicellaris (2) N –/2
sec. Myrtosalix {20}S. uva-ursi (1) N 1/1
subg. Longifoliae {7}sec. Longifoliae {7}S. exigua (6) N –/6S. interior (11) N 7/11
subg. Longifoliae x Vetrix
sec. Longifoliae x Cordatae
S. interior x eriocephalaE (3) N 2/3
subg. Longifoliae x Vetrix
sec. Longifoliae x Geyerianae
S. interior x petiolarisE (2) N 1/1 –/1
subg. Protitea {32}sec. Humboldtianae {15}S. amygdaloides (2) N 1/2
S. amygdaloides x gooddingii (2) N –/2
S. bonplandiana (2) N –/2S. gooddingii (2) N –/2
S. laevigata (2) N –/2
S. nigra (1) N 1/1
subg. Salix {84}sec. Maccallianae {1}S. maccalliana (6) N –/6
sec. Magnificae {8}S. magnifica (1) GO (Japan) –/1 1/–
sec. Salicaster {9}S. lasiandra (37) N 9/37
S. lucida (2) N 2/2
S. x meyeriana (1) I (Eu) –/1
S. serissima (2) N 2/2
sec. Salix {8}S. alba (1) I (Eu/Asia) 1/1
S. x fragilis (3) I (Eu) –/3
S. x sepulcralis (5) I (Eu/Asia) 2/5
subg. Vetrix {211}sec. Arbuscella {13}S. arbusculoides (4) N 1/4 1/–
sec. Canae {1}S. elaeagnos (1) I (Eu) 1/1
sec. Candidae {2}S. candida (11) N 3/9 1/1 1/1
sec. Cinerella {36}S. discolor (6) N 3/6
S. hookeriana (13) N 2/12 1/1
S. humilis (2) N 2/2
S. pedicellata (1) Eu –/1S. scouleriana (35) N 3/26 9/9
sec. Cordatae {1}S. eriocephala (4) N 1/2 1/– 2/2
S. famelica (2) N 1/2 1/–S. ligulifolia (4) N –/4\
Appendix II Continued
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 17
Page 18
Taxa N/I GI 1* GI others GII GIII GIV GV GVI
S. lutea (2) N –/2
S. prolixa (13) N 1/11 –/2sec. Fulvae {7}S. bebbiana (34) N 7/24 4/8
sec. Geyerianae {4}S. geyeriana (2) N –/1 –/1S. lemmonii (2) N –/2
S. petiolaris (4) N 4/4
sec. Hastatae {25}S. arizonica (3) N –/2 –/1S. barclayi (32) N 7/24 4/5
S. boothii (8) N –/5 1/3
S. commutata (3) N 1/3 1/–
S. cordata (5) N –/2 1/1 –/2S. eastwoodiae (2) N –/1 –/1
S. farriae (2) N –/2S. monticola (3) N –/2 –/1
S. myricoides (2) N –/1 –/1S. myrtillifolia (4) N 2/2 –/2
S. pseudomonticola (7) N –/7 2/–S. pseudomyrsinites (6) N 2/6 1/–
S. pyrifolia (2) N 1/2
sec. Lanatae {5}S. calcicola (2) N –/2
S. richardsonii (5) N –/3 –/2sec. Mexicanae {5}S. irrorata (2) N –/2S. lasiolepis (4) N –/2 –/2
sec. Phylicifoliae {11}S. drummondiana (18) N –/14 1/3
S. planifolia (8) N –/6 –/1 –/1S. pulchra (2) N –/2
sec. Sitchenses {4}S. jepsonii (3) N –/3
S. sitchensis (54) N 7/45 8/9
sec. Villosae {6}S. alaxensis (8) N 1/7 –/1S. barrattiana (7) N 1/5 3/8
sec. Villosae x Phylicifoliae
S. alaxensis x drummondiana (1) N –/1
sec. Viminella {13}S. viminalis (1) Eu –/1 1/–
Appendix II Continued
© 2014 John Wiley & Sons Ltd
18 D. M. PERCY ET AL.
Page 19
Appendix IIITaxon sampling from GenBank (GB) for rbcL and matK from Salicaceae (Sal) and Lacistemataceae (Lac)
Taxon (Family) GB rbcL GB matK
Salix arbutifolia (Sal) AB012776 EU790701
Populus balsamifera (Sal) EU676955 EU749348
Populus deltoids (Sal) AJ418829 EU790702
Populus nigra (Sal) AJ418828 AB038186
Populus tomentosa (Sal) AF527489 AY177666
Populus tremula (Sal) AJ418827 AJ506082
Populus tremuloides (Sal) AF206812 JF429913
Populus trichocarpa (Sal) NC009143 NC009143
Idesia polycarpa (Sal) AB021924 AB233831
Poliothyrsis sinensis (Sal) AJ402991 EF135586
Flacourtia jangomas (Sal) AF206768 EF135541
Xylosma congesta (Sal) AB233938 AB233834
Scyphostegia borneensis (Sal) AJ403000 EF135594
Casearia (Sal) AF206746 C. sylvestris EF135516 C. nitida
Lunania parviflora (Sal) AB233936 EF135561
Lacistema aggregatum (Lac) AY935746 FJ670025
Appendix IVSequence characteristics of each of the seven plastid regions (matK, rbcL, rpoC1, rpoB, trnH-psbA, atpF-atpH, psbK-psbI), and the single
mitochondrial gene (COI), including parsimony informative characters (PIC) for each region
Plastid region No. individuals sequenced aligned sequence length mean sequence length PIC
matK 539 898 888 28
rbcL 538 634 628 10
rpoC1 166 474 471 4
rpoB 167 358 358 6
trnH-psbA 197 291 238 16
atpF-atpH 155 609 567 20
psbK-psbI 163 534 470 25
COI 30 656 649 1
© 2014 John Wiley & Sons Ltd
HAS A SWEEP FOILED DNA BARCODING IN WILLOWS? 19
Page 20
Appendix VMaximum parsimony analysis for individual gene regions are illustrated with a single parsimony tree for each of the seven plastid regions,
the number of parsimony informative characters (PIC) for each region or gene are given, and the placement of the six major haplotype
groups are indicated. The matrices show the number of nucleotide changes along branches separating each of the six major groups.
Appendix VIComparative estimates of node ages in BEAST and r8s for the matK + rbcL data set, using additional taxa sampled from GenBank
(Appendix III). The BEAST mean node ages [median and the 95% highest posterior density (HPD) interval] are given. The BEAST analysis
and the last two r8s analyses given were run with the expanded outgroup taxon set (i.e. Salix plus all taxa in Appendix III), the first
three r8s analyses given were run with Salix plus two Populus taxa, Idesia and Poliothyrsis. The smoothing factor used in the penalized
likelihood (PL) analyses is given in parentheses
BEAST – Bayesian
r8s – LF
(Powell)
r8s – PL
(smooth 320)
r8s – PL
(smooth 32)
r8s – PL
(smooth 20) r8s – NPRS
G6 1.2 [1; 0–3] 0.49 0.44 0.39 0.31 1.38
G3 5.6 [5.3; 1.6–10.4] 1.86 1.73 1.83 1.45 8.66
G4 + 5 + 6 6.6 [6.3; 2.9–10.8] 2.40 2.21 2.11 1.66 7.9
G1 9.6 [9.2; 5.2–14.6] 2.54 2.48 8.16 3.32 10.88
G1 + G2 12.9 [12.5; 7–19.8] 4.07 3.98 11.65 5.28 18.02
G3 + 4 + 5 + 6 11 [10.6; 5.5–17.1] 5.33 5.03 5.47 4.22 13.91
Salix 19.8 [19.5; 12.5–27.6] 14.75 14.36 17.88 12.23 22.76
Saliceae 34.5 [34.6; 26.3–42] fixed 34 fixed 34 fixed 34 32.02 36.16
G I II III IV V VII - 1 1 2 2 3II - 0 1 1 2III - 1 1 2IV - 0 1V - 1VI -
rpoC1 4 PIC
I
II+III
IV+VVI
rpoB 6 PICG I II III IV V VII - 0 1 3 4 5II - 1 3 4 5III - 2 3 4IV - 1 2V - 1VI -
I+II
IIIVI
V
SALIX118-08 07-EM-0583 Salix bebbianaSALIX348-08 07-GWA-14380 Salix bebbianaSALIX384-08 75-UBC-155629 Salix alaxensis
SALIX385-08 78-UBC-162474 Salix alaxensisSALIX390-08 73-UBC-149089 Salix arbusculoides
SALIX392-08 69-UBC-149365 Salix arbusculoidesSALIX137-08 07-DPQC-0017 Salix arcticaSALIX140-08 07-DPQC-0020 Salix arcticaSALIX141-08 07-DPQC-0024 Salix arcticaSALIX142-08 07-DPQC-0025 Salix arcticaSALIX144-08 07-DPQC-0028 Salix arcticaSALIX150-08 07-DPQC-0034 Salix arcticaSALIX155-08 07-DPQC-0039 Salix arcticaSALIX167-08 07-DPQC-0055 Salix arcticaSALIX170-08 07-DPQC-0058 Salix arcticaSALIX380-08 61-UBC-119656 Salix arcticaSALIX383-08 73-UBC-152112 Salix arcticaSALIX151-08 07-DPQC-0035 Salix barclayiSALIX153-08 07-DPQC-0037 Salix barclayiSALIX160-08 07-DPQC-0048 Salix barclayiSALIX162-08 07-DPQC-0050 Salix barclayiSALIX163-08 07-DPQC-0051 Salix barclayiSALIX164-08 07-DPQC-0052 Salix barclayiSALIX165-08 07-DPQC-0053 Salix barclayiSALIX171-08 07-DPQC-0059 Salix barclayiSALIX194-08 07-UBCBG-037155-1003-2004 Salix barclayiSALIX199-08 07-UBCBG-native3 Salix barclayiSALIX139-08 07-DPQC-0019 Salix barrattianaSALIX156-08 07-DPQC-0040 Salix barrattianaSALIX173-08 07-DPQC-0061 Salix barrattianaSALIX381-08 43-UBC-65549 Salix barrattianaSALIX382-08 73-UBC-149084 Salix barrattianaSALIX005-08 07-JMS-1059 Salix bebbianaSALIX008-08 07-JMS-1094 Salix bebbianaSALIX113-08 07-EM-0315 Salix bebbianaSALIX115-08 07-EM-0472 Salix bebbianaSALIX119-08 07-EM-0596 Salix bebbianaSALIX121-08 07-EM-0628 Salix bebbianaSALIX122-08 06-DPQC-S1 Salix bebbianaSALIX129-08 07-DPQC-0009 Salix bebbianaSALIX134-08 07-DPQC-0014 Salix bebbianaSALIX198-08 07-UBCBG-037023-1003-2003 Salix bebbianaSALIX200-08 07-UBCBG-34916-5555-99 Salix bebbianaSALIX323-08 07-GWA-14349 Salix bebbianaSALIX197-08 07-UBCBG-037007-1003-2003 Salix boothiiSALIX120-08 07-EM-0618 Salix candidaSALIX312-08 07-GWA-14337 Salix candidaSALIX313-08 07-GWA-14338 Salix candidaSALIX315-08 07-GWA-14340 Salix candidaSALIX143-08 07-DPQC-0027 Salix commutataSALIX161-08 07-DPQC-0049 Salix commutataSALIX342-08 07-GWA-14371 Salix cordataSALIX343-08 07-GWA-14372 Salix cordataSALIX135-08 07-DPQC-0015 Salix drummondianaSALIX191-08 07-UBCBG-013854-0013-1976 Salix elaeagnosSALIX219-08 07-CAN-532085 Salix eriocephalaSALIX334-08 07-GWA-14363 Salix eriocephalaSALIX309-08 07-GWA-14334 Salix famelicaSALIX310-08 07-GWA-14335 Salix famelicaSALIX007-08 07-JMS-1084 Salix glaucaSALIX168-08 07-DPQC-0056 Salix glaucaSALIX174-08 07-DPQC-0062 Salix glaucaSALIX388-08 46-UBC-61323 Salix glauca
SALIX389-08 47-UBC-60831 Salix glaucaSALIX386-08 56-UBC-58897 Salix glauca x brachycarpaSALIX387-08 56-UBC-58933 Salix glauca x brachycarpaSALIX024-08 07-JMS-1297 Salix hookerianaSALIX189-08 07-DPQC-UBCFarm5 Salix hookerianaSALIX192-08 07-UBCBG-037096-1003-2004 Salix hookerianaSALIX341-08 07-GWA-14370 Salix hookerianaSALIX195-08 07-UBCBG-023877-0368-1984 Salix magnificaSALIX377-08 07-WAGNER-MYR1 Salix myrtillifoliaSALIX196-08 07-UBCBG-26352-117-87 Salix nakamuranaSALIX138-08 07-DPQC-0018 Salix nivalisSALIX149-08 07-DPQC-0033 Salix nivalisSALIX157-08 07-DPQC-0041 Salix nivalisSALIX158-08 07-DPQC-0042 Salix nivalisSALIX148-08 07-DPQC-0032 Salix prolixaSALIX117-08 07-EM-0579 Salix pseudomonticolaSALIX314-08 07-GWA-14339 Salix pseudomonticolaSALIX316-08 07-GWA-14342 Salix pseudomonticolaSALIX116-08 07-EM-0475 Salix pseudomyrsinitesSALIX378-08 07-WAGNER-PS1 Salix pseudomyrsinitesSALIX379-08 07-WAGNER-PS2 Salix pseudomyrsinitesSALIX347-08 07-GWA-14379 Salix pyrifoliaSALIX003-08 07-JMS-1050 Salix scoulerianaSALIX006-08 07-JMS-1081 Salix scoulerianaSALIX014-08 07-JMS-1141 Salix scoulerianaSALIX015-08 07-JMS-1142 Salix scoulerianaSALIX017-08 07-JMS-1211 Salix scoulerianaSALIX025-08 07-JMS-1298 Salix scoulerianaSALIX026-08 07-JMS-1299 Salix scoulerianaSALIX128-08 07-DPQC-0008 Salix scoulerianaSALIX133-08 07-DPQC-0013 Salix scoulerianaSALIX145-08 07-DPQC-0029 Salix scoulerianaSALIX146-08 07-DPQC-0030 Salix scoulerianaSALIX179-08 07-DPQC-MI3 Salix scoulerianaSALIX180-08 07-DPQC-MI4 Salix scoulerianaSALIX010-08 07-JMS-1123 Salix sitchensisSALIX012-08 07-JMS-1132 Salix sitchensisSALIX018-08 07-JMS-1212 Salix sitchensisSALIX023-08 07-JMS-1281 Salix sitchensisSALIX123-08 06-DPQC-S2 Salix sitchensisSALIX124-08 07-DPQC-0001A Salix sitchensisSALIX126-08 07-DPQC-0002 Salix sitchensisSALIX136-08 07-DPQC-0016 Salix sitchensisSALIX166-08 07-DPQC-0054 Salix sitchensisSALIX172-08 07-DPQC-0060 Salix sitchensisSALIX182-08 07-DPQC-MINavChan Salix sitchensisSALIX186-08 07-DPQC-UBCFarm2 Salix sitchensisSALIX202-08 07-DPQC-VanIs1 Salix sitchensisSALIX203-08 07-DPQC-VanIs2 Salix sitchensisSALIX206-08 07-DPQC-WhistRd2 Salix sitchensisSALIX209-08 07-DPQC-WhistRd5 Salix sitchensisSALIX210-08 07-DPQC-WhistRd6 Salix sitchensisSALIX205-08 07-DPQC-WhistRd1 Salix sp.SALIX207-08 07-DPQC-WhistRd3 Salix sp.SALIX208-08 07-DPQC-WhistRd4 Salix sp.SALIX184-08 07-PMP-20885 Salix uvaursiSALIX204-08 07-DPQC-vimEur ope Salix viminalisSALIX391-08 73-UBC-146342 Salix glauca
SALIX201-08 07-DMP-UniGuelph Salix albaSALIX011-08 07-JMS-1124 Salix lasiandraSALIX013-08 07-JMS-1133 Salix lasiandraSALIX131-08 07-DPQC-0011 Salix lasiandraSALIX169-08 07-DPQC-0057 Salix lasiandraSALIX175-08 07-DPQC-natJerBeach Salix lasiandraSALIX178-08 07-JW-lasiand2 Salix lasiandraSALIX181-08 07-DPQC-MIBenBay Salix lasiandraSALIX185-08 07-DPQC-UBCFarm1 Salix lasiandraSALIX193-08 07-UBCBG-015012-0268-1974 Salix lasiandraSALIX327-08 07-GWA-14353 Salix serissimaSALIX328-08 07-GWA-14354 Salix serissimaSALIX176-08 07-DPQC-weepJerBeach Salix x sepulcralis
SALIX183-08 07-PMP-20866 Salix exiguaSALIX112-08 07-JMS-0981 Salix interiorSALIX317-08 07-GWA-14343 Salix interiorSALIX331-08 07-GWA-14360 Salix interiorSALIX336-08 07-GWA-14365 Salix interiorSALIX337-08 07-GWA-14366 Salix interiorSALIX338-08 07-GWA-14367 Salix interiorSALIX212-08 07-CAN-483931 Salix interior x eriocephalaSALIX217-08 07-CAN-532065 Salix interior x eriocephalaSALIX215-08 07-CAN-532056 Salix interior x eriocephala
SALIX320-08 07-GWA-14346 Salix amygdaloidesSALIX332-08 07-GWA-14361 Salix amygdaloidesSALIX340-08 07-GWA-14369 Salix candidaSALIX344-08 07-GWA-14375 Salix cordataSALIX114-08 07-EM-0359 Salix discolorSALIX319-08 07-GWA-14345 Salix discolorSALIX321-08 07-GWA-14347 Salix discolorSALIX329-08 07-GWA-14355 Salix discolorSALIX339-08 07-GWA-14368 Salix discolorSALIX345-08 07-GWA-14376 Salix eriocephalaSALIX346-08 07-GWA-14377 Salix eriocephalaSALIX127-08 07-DPQC-0003 Salix exiguaSALIX330-08 07-GWA-14356 Salix humilisSALIX335-08 07-GWA-14364 Salix humilisSALIX214-08 07-CAN-483950 Salix interior x petiolarisSALIX324-08 07-GWA-14350 Salix lucidaSALIX326-08 07-GWA-14352 Salix lucidaSALIX333-08 07-GWA-14362 Salix nigraSALIX213-08 07-CAN-483942 Salix petiolarisSALIX318-08 07-GWA-14344 Salix petiolarisSALIX322-08 07-GWA-14348 Salix petiolarisSALIX325-08 07-GWA-14351 Salix petiolaris
0.05 changes
SALIX380-08 61-UBC-119656 Salix arcticaSALIX310-08 07-GWA-14335 Salix famelica
SALIX384-08 75-UBC-155629 Salix alaxensisSALIX385-08 78-UBC-162474 Salix alaxensisSALIX390-08 73-UBC-149089 Salix arbusculoidesSALIX392-08 69-UBC-149365 Salix arbusculoidesSALIX137-08 07-DPQC-0017 Salix arcticaSALIX140-08 07-DPQC-0020 Salix arcticaSALIX141-08 07-DPQC-0024 Salix arcticaSALIX142-08 07-DPQC-0025 Salix arcticaSALIX144-08 07-DPQC-0028 Salix arcticaSALIX150-08 07-DPQC-0034 Salix arcticaSALIX155-08 07-DPQC-0039 Salix arcticaSALIX167-08 07-DPQC-0055 Salix arcticaSALIX170-08 07-DPQC-0058 Salix arcticaSALIX383-08 73-UBC-152112 Salix arcticaSALIX151-08 07-DPQC-0035 Salix barclayiSALIX153-08 07-DPQC-0037 Salix barclayiSALIX160-08 07-DPQC-0048 Salix barclayiSALIX162-08 07-DPQC-0050 Salix barclayiSALIX163-08 07-DPQC-0051 Salix barclayiSALIX164-08 07-DPQC-0052 Salix barclayiSALIX165-08 07-DPQC-0053 Salix barclayiSALIX171-08 07-DPQC-0059 Salix barclayiSALIX194-08 07-UBCBG-037155-1003-2004 Salix barclayiSALIX199-08 07-UBCBG-native3 Salix barclayiSALIX139-08 07-DPQC-0019 Salix barrattianaSALIX156-08 07-DPQC-0040 Salix barrattianaSALIX173-08 07-DPQC-0061 Salix barrattianaSALIX381-08 43-UBC-65549 Salix barrattianaSALIX382-08 73-UBC-149084 Salix barrattianaSALIX005-08 07-JMS-1059 Salix bebbianaSALIX008-08 07-JMS-1094 Salix bebbianaSALIX113-08 07-EM-0315 Salix bebbianaSALIX115-08 07-EM-0472 Salix bebbianaSALIX118-08 07-EM-0583 Salix bebbianaSALIX119-08 07-EM-0596 Salix bebbianaSALIX121-08 07-EM-0628 Salix bebbianaSALIX122-08 06-DPQC-S1 Salix bebbianaSALIX129-08 07-DPQC-0009 Salix bebbianaSALIX134-08 07-DPQC-0014 Salix bebbianaSALIX198-08 07-UBCBG-037023-1003-2003 Salix bebbianaSALIX200-08 07-UBCBG-34916-5555-99 Salix bebbianaSALIX323-08 07-GWA-14349 Salix bebbianaSALIX348-08 07-GWA-14380 Salix bebbianaSALIX197-08 07-UBCBG-037007-1003-2003 Salix boothiiSALIX120-08 07-EM-0618 Salix candidaSALIX312-08 07-GWA-14337 Salix candidaSALIX313-08 07-GWA-14338 Salix candidaSALIX315-08 07-GWA-14340 Salix candidaSALIX143-08 07-DPQC-0027 Salix commutataSALIX161-08 07-DPQC-0049 Salix commutataSALIX342-08 07-GWA-14371 Salix cordata
SALIX135-08 07-DPQC-0015 Salix drummondianaSALIX191-08 07-UBCBG-013854-0013-1976 Salix elaeagnosSALIX219-08 07-CAN-532085 Salix eriocephalaSALIX334-08 07-GWA-14363 Salix eriocephalaSALIX309-08 07-GWA-14334 Salix famelicaSALIX007-08 07-JMS-1084 Salix glaucaSALIX168-08 07-DPQC-0056 Salix glaucaSALIX174-08 07-DPQC-0062 Salix glaucaSALIX388-08 46-UBC-61323 Salix glaucaSALIX389-08 47-UBC-60831 Salix glaucaSALIX386-08 56-UBC-58897 Salix glauca x brachycarpaSALIX387-08 56-UBC-58933 Salix glauca x brachycarpaSALIX024-08 07-JMS-1297 Salix hookerianaSALIX189-08 07-DPQC-UBCFarm5 Salix hookerianaSALIX192-08 07-UBCBG-037096-1003-2004 Salix hookerianaSALIX341-08 07-GWA-14370 Salix hookerianaSALIX011-08 07-JMS-1124 Salix lasiandraSALIX013-08 07-JMS-1133 Salix lasiandraSALIX131-08 07-DPQC-0011 Salix lasiandraSALIX169-08 07-DPQC-0057 Salix lasiandraSALIX175-08 07-DPQC-natJerBeach Salix lasiandraSALIX178-08 07-JW-lasiand2 Salix lasiandraSALIX181-08 07-DPQC-MIBenBay Salix lasiandraSALIX185-08 07-DPQC-UBCFarm1 Salix lasiandraSALIX193-08 07-UBCBG-015012-0268-1974 Salix lasiandraSALIX195-08 07-UBCBG-023877-0368-1984 Salix magnificaSALIX377-08 07-WAGNER-MYR1 Salix myrtillifoliaSALIX196-08 07-UBCBG-26352-117-87 Salix nakamuranaSALIX138-08 07-DPQC-0018 Salix nivalisSALIX149-08 07-DPQC-0033 Salix nivalisSALIX157-08 07-DPQC-0041 Salix nivalisSALIX158-08 07-DPQC-0042 Salix nivalisSALIX148-08 07-DPQC-0032 Salix prolixaSALIX117-08 07-EM-0579 Salix pseudomonticolaSALIX314-08 07-GWA-14339 Salix pseudomonticolaSALIX316-08 07-GWA-14342 Salix pseudomonticolaSALIX116-08 07-EM-0475 Salix pseudomyrsinitesSALIX378-08 07-WAGNER-PS1 Salix pseudomyrsinitesSALIX379-08 07-WAGNER-PS2 Salix pseudomyrsinitesSALIX347-08 07-GWA-14379 Salix pyrifoliaSALIX003-08 07-JMS-1050 Salix scoulerianaSALIX006-08 07-JMS-1081 Salix scoulerianaSALIX014-08 07-JMS-1141 Salix scoulerianaSALIX015-08 07-JMS-1142 Salix scoulerianaSALIX017-08 07-JMS-1211 Salix scoulerianaSALIX025-08 07-JMS-1298 Salix scoulerianaSALIX026-08 07-JMS-1299 Salix scoulerianaSALIX128-08 07-DPQC-0008 Salix scoulerianaSALIX133-08 07-DPQC-0013 Salix scoulerianaSALIX145-08 07-DPQC-0029 Salix scoulerianaSALIX146-08 07-DPQC-0030 Salix scoulerianaSALIX179-08 07-DPQC-MI3 Salix scoulerianaSALIX180-08 07-DPQC-MI4 Salix scoulerianaSALIX010-08 07-JMS-1123 Salix sitchensisSALIX012-08 07-JMS-1132 Salix sitchensisSALIX018-08 07-JMS-1212 Salix sitchensisSALIX023-08 07-JMS-1281 Salix sitchensisSALIX123-08 06-DPQC-S2 Salix sitchensis
SALIX124-08 07-DPQC-0001A Salix sitchensisSALIX126-08 07-DPQC-0002 Salix sitchensisSALIX136-08 07-DPQC-0016 Salix sitchensisSALIX166-08 07-DPQC-0054 Salix sitchensisSALIX172-08 07-DPQC-0060 Salix sitchensisSALIX182-08 07-DPQC-MINavChan Salix sitchensisSALIX186-08 07-DPQC-UBCFarm2 Salix sitchensisSALIX202-08 07-DPQC-VanIs1 Salix sitchensisSALIX203-08 07-DPQC-VanIs2 Salix sitchensisSALIX206-08 07-DPQC-WhistRd2 Salix sitchensisSALIX209-08 07-DPQC-WhistRd5 Salix sitchensisSALIX210-08 07-DPQC-WhistRd6 Salix sitchensisSALIX205-08 07-DPQC-WhistRd1 Salix sp.SALIX207-08 07-DPQC-WhistRd3 Salix sp.SALIX208-08 07-DPQC-WhistRd4 Salix sp.SALIX184-08 07-PMP-20885 Salix uvaursiSALIX204-08 07-DPQC-vimEur ope Salix viminalisSALIX391-08 73-UBC-146342 Salix glauca
SALIX201-08 07-DMP-UniGuelph Salix albaSALIX327-08 07-GWA-14353 Salix serissimaSALIX328-08 07-GWA-14354 Salix serissimaSALIX176-08 07-DPQC-weepJerBeach Salix x sepulcralis
SALIX183-08 07-PMP-20866 Salix exiguaSALIX112-08 07-JMS-0981 Salix interiorSALIX317-08 07-GWA-14343 Salix interiorSALIX331-08 07-GWA-14360 Salix interiorSALIX336-08 07-GWA-14365 Salix interiorSALIX337-08 07-GWA-14366 Salix interiorSALIX338-08 07-GWA-14367 Salix interiorSALIX212-08 07-CAN-483931 Salix interior x eriocephalaSALIX217-08 07-CAN-532065 Salix interior x eriocephalaSALIX216-08 07-CAN-532063 Salix interior x petiolarisSALIX215-08 07-CAN-532056 Salix interior x eriocephala
SALIX340-08 07-GWA-14369 Salix candidaSALIX343-08 07-GWA-14372 Salix cordataSALIX344-08 07-GWA-14375 Salix cordataSALIX114-08 07-EM-0359 Salix discolorSALIX319-08 07-GWA-14345 Salix discolor
SALIX321-08 07-GWA-14347 Salix discolorSALIX329-08 07-GWA-14355 Salix discolorSALIX339-08 07-GWA-14368 Salix discolorSALIX345-08 07-GWA-14376 Salix eriocephalaSALIX346-08 07-GWA-14377 Salix eriocephalaSALIX127-08 07-DPQC-0003 Salix exiguaSALIX330-08 07-GWA-14356 Salix humilisSALIX335-08 07-GWA-14364 Salix humilisSALIX214-08 07-CAN-483950 Salix interior x petiolarisSALIX213-08 07-CAN-483942 Salix petiolarisSALIX318-08 07-GWA-14344 Salix petiolarisSALIX322-08 07-GWA-14348 Salix petiolarisSALIX325-08 07-GWA-14351 Salix petiolaris
SALIX320-08 07-GWA-14346 Salix amygdaloidesSALIX332-08 07-GWA-14361 Salix amygdaloidesSALIX324-08 07-GWA-14350 Salix lucidaSALIX326-08 07-GWA-14352 Salix lucidaSALIX333-08 07-GWA-14362 Salix nigra
0.05 changes
SALIX197-08 07-UBCBG-037007-1003-2003 Salix boothiiSALIX376-08 07-GWA-14436A Salix eastwoodiaeSALIX090-08 07-JMS-0942 Salix hookerianaSALIX226-08 07-BAB-023 Salix alaxensisSALIX239-08 07-BAB-088 Salix alaxensisSALIX250-08 07-BAB-197 Salix alaxensisSALIX261-08 07-BAB-230 Salix alaxensisSALIX263-08 07-BAB-232 Salix alaxensisSALIX264-08 07-BAB-237 Salix alaxensisSALIX384-08 75-UBC-155629 Salix alaxensisSALIX385-08 78-UBC-162474 Salix alaxensisSALIX534-08 08-GWA-14451 Salix alaxensis x drummondianaSALIX060-08 07-JMS-1354 Salix arbusculoidesSALIX229-08 07-BAB-030 Salix arbusculoidesSALIX390-08 73-UBC-149089 Salix arbusculoidesSALIX392-08 69-UBC-149365 Salix arbusculoidesSALIX037-08 07-JMS-1330 Salix arcticaSALIX038-08 07-JMS-1333 Salix arcticaSALIX039-08 07-JMS-1334 Salix arcticaSALIX049-08 07-JMS-1343 Salix arcticaSALIX137-08 07-DPQC-0017 Salix arcticaSALIX140-08 07-DPQC-0020 Salix arcticaSALIX141-08 07-DPQC-0024 Salix arcticaSALIX142-08 07-DPQC-0025 Salix arcticaSALIX144-08 07-DPQC-0028 Salix arcticaSALIX150-08 07-DPQC-0034 Salix arcticaSALIX154-08 07-DPQC-0038 Salix arcticaSALIX155-08 07-DPQC-0039 Salix arcticaSALIX167-08 07-DPQC-0055 Salix arcticaSALIX170-08 07-DPQC-0058 Salix arcticaSALIX234-08 07-BAB-047A Salix arcticaSALIX235-08 07-BAB-047B Salix arcticaSALIX383-08 73-UBC-152112 Salix arcticaSALIX232-08 07-BAB-033 Salix arctica x glaucaSALIX240-08 07-BAB-096A Salix arctica x polarisSALIX247-08 07-BAB-186 Salix arctica x polarisSALIX297-08 07-CEG-27 Salix arizonicaSALIX298-08 07-CEG-26 Salix arizonicaSALIX510-08 08-CEG-12 Salix arizonicaSALIX057-08 07-JMS-1351 Salix athabascensisSALIX517-08 08-GWA-14434 Salix athabascensisSALIX040-08 07-JMS-1335 Salix barclayiSALIX041-08 07-JMS-1336 Salix barclayiSALIX047-08 07-JMS-1341 Salix barclayiSALIX048-08 07-JMS-1342 Salix barclayiSALIX050-08 07-JMS-1344B Salix barclayiSALIX053-08 07-JMS-1347 Salix barclayiSALIX151-08 07-DPQC-0035 Salix barclayiSALIX152-08 07-DPQC-0036 Salix barclayiSALIX153-08 07-DPQC-0037 Salix barclayiSALIX160-08 07-DPQC-0048 Salix barclayiSALIX162-08 07-DPQC-0050 Salix barclayiSALIX163-08 07-DPQC-0051 Salix barclayiSALIX164-08 07-DPQC-0052 Salix barclayiSALIX165-08 07-DPQC-0053 Salix barclayiSALIX171-08 07-DPQC-0059 Salix barclayiSALIX194-08 07-UBCBG-037155-1003-2004 Salix barclayiSALIX199-08 07-UBCBG-native3 Salix barclayiSALIX227-08 07-BAB-025 Salix barclayiSALIX238-08 07-BAB-082 Salix barclayiSALIX260-08 07-BAB-227 Salix barclayiSALIX266-08 07-BAB-246 Salix barclayiSALIX270-08 07-BAB-267 Salix barclayiSALIX272-08 07-BAB-269 Salix barclayiSALIX274-08 07-BAB-272 Salix barclayiSALIX279-08 07-BAB-287 Salix barclayiSALIX287-08 07-BAB-313 Salix barclayiSALIX404-08 07-BAB-024 Salix barclayiSALIX443-08 08-DPQC-0095A Salix barclayiSALIX444-08 08-DPQC-0095B Salix barclayiSALIX454-08 08-DPQC-0107 Salix barclayiSALIX455-08 08-DPQC-0108 Salix barclayiSALIX458-08 08-DPQC-0112 Salix barclayiSALIX139-08 07-DPQC-0019 Salix barrattianaSALIX156-08 07-DPQC-0040 Salix barrattianaSALIX173-08 07-DPQC-0061 Salix barrattianaSALIX246-08 07-BAB-168 Salix barrattianaSALIX382-08 73-UBC-149084 Salix barrattianaSALIX522-08 08-GWA-14437A Salix barrattianaSALIX004-08 07-JMS-1056 Salix bebbianaSALIX005-08 07-JMS-1059 Salix bebbianaSALIX008-08 07-JMS-1094 Salix bebbianaSALIX063-08 07-JMS-1362 Salix bebbianaSALIX066-08 07-JMS-1365 Salix bebbianaSALIX113-08 07-EM-0315 Salix bebbianaSALIX115-08 07-EM-0472 Salix bebbianaSALIX118-08 07-EM-0583 Salix bebbianaSALIX119-08 07-EM-0596 Salix bebbianaSALIX121-08 07-EM-0628 Salix bebbianaSALIX129-08 07-DPQC-0009 Salix bebbianaSALIX134-08 07-DPQC-0014 Salix bebbianaSALIX198-08 07-UBCBG-037023-1003-2003 Salix bebbianaSALIX200-08 07-UBCBG-34916-5555-99 Salix bebbianaSALIX221-08 07-BAB-014 Salix bebbianaSALIX248-08 07-BAB-193 Salix bebbianaSALIX253-08 07-BAB-200 Salix bebbianaSALIX275-08 07-BAB-273 Salix bebbianaSALIX323-08 07-GWA-14349 Salix bebbianaSALIX348-08 07-GWA-14380 Salix bebbianaSALIX414-08 08-DPQC-0066 Salix bebbianaSALIX417-08 08-DPQC-0069 Salix bebbianaSALIX419-08 08-DPQC-0071 Salix bebbianaSALIX424-08 08-DPQC-0076 Salix bebbianaSALIX428-08 08-DPQC-0080 Salix bebbianaSALIX432-08 08-DPQC-0084 Salix bebbianaSALIX440-08 08-DPQC-0092 Salix bebbianaSALIX442-08 08-DPQC-0094 Salix bebbianaSALIX463-08 08-DPQC-0118 Salix bebbianaSALIX472-08 08-DPQC-0127 Salix bebbianaSALIX513-08 08-CEG-15 Salix bebbianaSALIX514-08 08-CEG-16 Salix bebbianaSALIX065-08 07-JMS-1364 Salix boothiiSALIX293-08 07-CEG-19 Salix boothiiSALIX355-08 07-GWA-14387 Salix boothiiSALIX356-08 07-GWA-14388 Salix boothiiSALIX360-08 07-GWA-14395 Salix boothiiSALIX515-08 08-CEG-17 Salix boothiiSALIX516-08 08-GWA-14433 Salix boothiiSALIX526-08 08-GWA-14443 Salix brachycarpaSALIX527-08 08-GWA-14444 Salix brachycarpaSALIX532-08 08-GWA-14449 Salix calicolaSALIX533-08 08-GWA-14450 Salix calicolaSALIX395-08 08-ISQC-CanIs Salix canariensisSALIX120-08 07-EM-0618 Salix candidaSALIX243-08 07-BAB-130 Salix candidaSALIX259-08 07-BAB-226 Salix candidaSALIX312-08 07-GWA-14337 Salix candidaSALIX313-08 07-GWA-14338 Salix candidaSALIX315-08 07-GWA-14340 Salix candidaSALIX481-08 08-DPQC-0136 Salix candidaSALIX485-08 08-DPQC-0141 Salix candidaSALIX569-09 09-DPQC-11 Salix candidaSALIX573-09 09-DPQC-26 Salix candidaSALIX143-08 07-DPQC-0027 Salix commutataSALIX161-08 07-DPQC-0049 Salix commutataSALIX529-08 08-GWA-14446 Salix commutataSALIX342-08 07-GWA-14371 Salix cordataSALIX400-08 07-DW-6000 Salix cordataSALIX402-08 07-DW-6002 Salix cordataSALIX054-08 07-JMS-1348 Salix drummondianaSALIX059-08 07-JMS-1353 Salix drummondianaSALIX064-08 07-JMS-1363 Salix drummondianaSALIX135-08 07-DPQC-0015 Salix drummondianaSALIX357-08 07-GWA-14390 Salix drummondianaSALIX359-08 07-GWA-14393 Salix drummondianaSALIX430-08 08-DPQC-0082 Salix drummondianaSALIX435-08 08-DPQC-0087 Salix drummondianaSALIX457-08 08-DPQC-0111 Salix drummondianaSALIX466-08 08-DPQC-0121 Salix drummondianaSALIX470-08 08-DPQC-0125 Salix drummondianaSALIX478-08 08-DPQC-0133 Salix drummondianaSALIX520-08 08-GWA-14437 1 Salix drummondianaSALIX521-08 08-GWA-14437 2 Salix drummondianaSALIX528-08 08-GWA-14445 Salix drummondianaSALIX530-08 08-GWA-14447 Salix drummondianaSALIX574-09 07-GWA-14436B Salix drummondianaSALIX375-08 07-GWA-14431 Salix eastwoodiaeSALIX191-08 07-UBCBG-013854-0013-1976 Salix elaeagnosSALIX219-08 07-CAN-532085 Salix eriocephalaSALIX334-08 07-GWA-14363 Salix eriocephalaSALIX309-08 07-GWA-14334 Salix famelicaSALIX310-08 07-GWA-14335 Salix famelicaSALIX524-08 08-GWA-14439 Salix farriaeSALIX570-09 09-DPQC-13 Salix farriaeSALIX109-08 07-JMS-0974 Salix geyerianaSALIX511-08 08-CEG-13 Salix geyerianaSALIX007-08 07-JMS-1084 Salix glaucaSALIX168-08 07-DPQC-0056 Salix glaucaSALIX174-08 07-DPQC-0062 Salix glaucaSALIX224-08 07-BAB-017 Salix glaucaSALIX286-08 07-BAB-299 Salix glaucaSALIX361-08 07-GWA-14396 Salix glaucaSALIX362-08 07-GWA-14397 Salix glaucaSALIX368-08 07-GWA-14424 Salix glaucaSALIX374-08 07-GWA-14430 Salix glaucaSALIX388-08 46-UBC-61323 Salix glaucaSALIX389-08 47-UBC-60831 Salix glaucaSALIX441-08 08-DPQC-0093 Salix glaucaSALIX471-08 08-DPQC-0126 Salix glaucaSALIX486-08 08-DPQC-0142 Salix glaucaSALIX487-08 08-DPQC-0143 Salix glaucaSALIX459-08 08-DPQC-0113 Salix glauca villosaSALIX460-08 08-DPQC-0114 Salix glauca villosaSALIX464-08 08-DPQC-0119 Salix glauca villosaSALIX465-08 08-DPQC-0120 Salix glauca villosaSALIX480-08 08-DPQC-0135 Salix glauca villosaSALIX531-08 08-GWA-14448 Salix glauca villosaSALIX386-08 56-UBC-58897 Salix glauca x brachycarpaSALIX387-08 56-UBC-58933 Salix glauca x brachycarpaSALIX024-08 07-JMS-1297 Salix hookerianaSALIX069-08 07-JMS-0865 Salix hookerianaSALIX074-08 07-JMS-0885 Salix hookerianaSALIX078-08 07-JMS-0900 Salix hookerianaSALIX098-08 07-JMS-0956 Salix hookerianaSALIX100-08 07-JMS-0958 Salix hookerianaSALIX102-08 07-JMS-0960 Salix hookerianaSALIX103-08 07-JMS-0961 Salix hookerianaSALIX189-08 07-DPQC-UBCFarm5 Salix hookerianaSALIX192-08 07-UBCBG-037096-1003-2004 Salix hookerianaSALIX341-08 07-GWA-14370 Salix hookerianaSALIX399-08 08-DPsitchSB Salix hookerianaSALIX290-08 07-CEG-15 Salix irrorataSALIX291-08 07-CEG-16 Salix irrorataSALIX364-08 07-GWA-14411 Salix jepsoniiSALIX365-08 07-GWA-14413 Salix jepsoniiSALIX370-08 07-GWA-14426 Salix jepsoniiSALIX011-08 07-JMS-1124 Salix lasiandraSALIX013-08 07-JMS-1133 Salix lasiandraSALIX028-08 07-JMS-1309 Salix lasiandraSALIX031-08 07-JMS-1314 Salix lasiandraSALIX055-08 07-JMS-1349 Salix lasiandraSALIX056-08 07-JMS-1350 Salix lasiandraSALIX067-08 07-JMS-0863 Salix lasiandraSALIX077-08 07-JMS-0899 Salix lasiandraSALIX080-08 07-JMS-0905 Salix lasiandraSALIX082-08 07-JMS-0923 Salix lasiandraSALIX088-08 07-JMS-0937 Salix lasiandraSALIX091-08 07-JMS-0943 Salix lasiandraSALIX106-08 07-JMS-0966 Salix lasiandraSALIX131-08 07-DPQC-0011 Salix lasiandraSALIX169-08 07-DPQC-0057 Salix lasiandraSALIX175-08 07-DPQC-natJerBeach Salix lasiandraSALIX178-08 07-JW-lasiand2 Salix lasiandraSALIX181-08 07-DPQC-MIBenBay Salix lasiandraSALIX185-08 07-DPQC-UBCFarm1 Salix lasiandraSALIX193-08 07-UBCBG-015012-0268-1974 Salix lasiandraSALIX251-08 07-BAB-198 Salix lasiandraSALIX262-08 07-BAB-231 Salix lasiandraSALIX296-08 07-CEG-25 Salix lasiandraSALIX351-08 07-GWA-14383 Salix lasiandraSALIX393-08 08-DPYC-02 Salix lasiandraSALIX406-08 07-CEG-17 Salix lasiandraSALIX423-08 08-DPQC-0075 Salix lasiandraSALIX425-08 08-DPQC-0077 Salix lasiandraSALIX426-08 08-DPQC-0078 Salix lasiandraSALIX434-08 08-DPQC-0086 Salix lasiandraSALIX438-08 08-DPQC-0090 Salix lasiandraSALIX445-08 08-DPQC-0096 Salix lasiandraSALIX447-08 08-DPQC-0100 Salix lasiandraSALIX449-08 08-DPQC-0102 Salix lasiandraSALIX476-08 08-DPQC-0131 Salix lasiandraSALIX477-08 08-DPQC-0132 Salix lasiandraSALIX292-08 07-CEG-18 Salix lasiolepisSALIX371-08 07-GWA-14427 Salix lasiolepisSALIX372-08 07-GWA-14428 Salix lasiolepisSALIX353-08 07-GWA-14385 Salix lemmoniiSALIX354-08 07-GWA-14386 Salix lemmoniiSALIX288-08 07-CEG-13 Salix ligulifoliaSALIX289-08 07-CEG-14 Salix ligulifoliaSALIX500-08 08-CEG-03A Salix ligulifoliaSALIX501-08 08-CEG-03B Salix ligulifoliaSALIX396-08 08-DPSlind Salix lindleyanaSALIX350-08 07-GWA-14382 Salix luteaSALIX352-08 07-GWA-14384 Salix luteaSALIX479-08 08-DPQC-0134 Salix maccallianaSALIX483-08 08-DPQC-0139 Salix maccallianaSALIX484-08 08-DPQC-0140 Salix maccallianaSALIX488-08 08-DPQC-0144 Salix maccallianaSALIX489-08 08-DPQC-0145 Salix maccallianaSALIX535-08 08-GWA-14452 Salix maccallianaSALIX195-08 07-UBCBG-023877-0368-1984 Salix magnificaSALIX299-08 07-CEG-28 Salix monticolaSALIX300-08 07-CEG-29 Salix monticolaSALIX512-08 08-CEG-14 Salix monticolaSALIX401-08 07-DW-6001 Salix myricoidesSALIX403-08 07-DW-6003 Salix myricoidesSALIX223-08 07-BAB-016 Salix myrtillifoliaSALIX258-08 07-BAB-224 Salix myrtillifoliaSALIX277-08 07-BAB-276 Salix myrtillifoliaSALIX377-08 07-WAGNER-MYR1 Salix myrtillifoliaSALIX196-08 07-UBCBG-26352-117-87 Salix nakamuranaSALIX236-08 07-BAB-051 Salix niphocladaSALIX268-08 07-BAB-250 Salix niphocladaSALIX269-08 07-BAB-251 Salix niphocladaSALIX138-08 07-DPQC-0018 Salix nivalisSALIX149-08 07-DPQC-0033 Salix nivalisSALIX157-08 07-DPQC-0041 Salix nivalisSALIX158-08 07-DPQC-0042 Salix nivalisSALIX523-08 08-GWA-14438 Salix nivalisSALIX271-08 07-BAB-268 Salix pedicellarisSALIX273-08 07-BAB-270 Salix pedicellarisSALIX363-08 07-GWA-14409 Salix petrophilaSALIX058-08 07-JMS-1352 Salix planifoliaSALIX220-08 07-BAB-013 Salix planifoliaSALIX249-08 07-BAB-194 Salix planifoliaSALIX366-08 07-GWA-14416 Salix planifoliaSALIX367-08 07-GWA-14417 Salix planifoliaSALIX422-08 08-DPQC-0074 Salix planifoliaSALIX461-08 08-DPQC-0115 Salix planifoliaSALIX285-08 07-BAB-298 Salix polarisSALIX061-08 07-JMS-1360 Salix prolixaSALIX148-08 07-DPQC-0032 Salix prolixaSALIX421-08 08-DPQC-0073 Salix prolixaSALIX427-08 08-DPQC-0079 Salix prolixaSALIX429-08 08-DPQC-0081 Salix prolixaSALIX436-08 08-DPQC-0088 Salix prolixaSALIX439-08 08-DPQC-0091 Salix prolixaSALIX450-08 08-DPQC-0103 Salix prolixaSALIX452-08 08-DPQC-0105 Salix prolixaSALIX469-08 08-DPQC-0124 Salix prolixaSALIX475-08 08-DPQC-0130 Salix prolixaSALIX491-08 08-DPQC-0148 Salix prolixaSALIX492-08 08-DPQC-0149 Salix prolixaSALIX117-08 07-EM-0579 Salix pseudomonticolaSALIX267-08 07-BAB-247 Salix pseudomonticolaSALIX314-08 07-GWA-14339 Salix pseudomonticolaSALIX316-08 07-GWA-14342 Salix pseudomonticolaSALIX468-08 08-DPQC-0123 Salix pseudomonticolaSALIX474-08 08-DPQC-0129 Salix pseudomonticolaSALIX482-08 08-DPQC-0137 Salix pseudomonticolaSALIX116-08 07-EM-0475 Salix pseudomyrsinitesSALIX245-08 07-BAB-167 Salix pseudomyrsinitesSALIX278-08 07-BAB-286 Salix pseudomyrsinitesSALIX378-08 07-WAGNER-PS1 Salix pseudomyrsinitesSALIX379-08 07-WAGNER-PS2 Salix pseudomyrsinitesSALIX405-08 07-BAB-344 Salix pseudomyrsinitesSALIX225-08 07-BAB-018 Salix pulchraSALIX281-08 07-BAB-291 Salix pulchraSALIX347-08 07-GWA-14379 Salix pyrifoliaSALIX418-08 08-DPQC-0070 Salix pyrifoliaSALIX536-09 08-CAR-001 Salix reticulataSALIX537-09 08-CAR-002 Salix reticulataSALIX538-09 08-CAR-003 Salix reticulataSALIX539-09 08-CAR-004 Salix reticulataSALIX540-09 08-CAR-005 Salix reticulataSALIX541-09 08-CAR-006 Salix reticulataSALIX542-09 08-CAR-007 Salix reticulataSALIX044-08 07-JMS-1338 Salix reticulataSALIX045-08 07-JMS-1339 Salix reticulataSALIX046-08 07-JMS-1340 Salix reticulataSALIX233-08 07-BAB-039 Salix reticulataSALIX244-08 07-BAB-165 Salix reticulataSALIX282-08 07-BAB-292 Salix reticulataSALIX543-09 08-CAR-008 Salix reticulataSALIX544-09 08-CAR-009 Salix reticulataSALIX545-09 08-CAR-010 Salix reticulataSALIX546-09 08-CAR-011 Salix reticulataSALIX547-09 08-CAR-012 Salix reticulataSALIX548-09 08-CAR-013 Salix reticulataSALIX550-09 08-CAR-015 Salix reticulataSALIX551-09 08-CAR-016 Salix reticulataSALIX552-09 08-CAR-017 Salix reticulataSALIX553-09 08-CAR-018 Salix reticulataSALIX554-09 08-CAR-019 Salix reticulataSALIX555-09 08-CAR-020 Salix reticulataSALIX556-09 08-CAR-021 Salix reticulataSALIX557-09 08-CAR-022 Salix reticulataSALIX558-09 08-CAR-023 Salix reticulataSALIX559-09 08-CAR-024 Salix reticulataSALIX560-09 08-CAR-025 Salix reticulataSALIX561-09 08-CAR-026 Salix reticulataSALIX562-09 08-CAR-027 Salix reticulataSALIX563-09 08-CAR-028 Salix reticulataSALIX564-09 08-CAR-029 Salix reticulataSALIX565-09 08-CAR-030 Salix reticulataSALIX566-09 08-CAR-031 Salix reticulataSALIX567-09 08-CAR-032 Salix reticulataSALIX568-09 08-CAR-033 Salix reticulataSALIX228-08 07-BAB-029 Salix richardsoniiSALIX230-08 07-BAB-031 Salix richardsoniiSALIX231-08 07-BAB-032 Salix richardsoniiSALIX255-08 07-BAB-209 Salix richardsoniiSALIX284-08 07-BAB-294 Salix richardsoniiSALIX237-08 07-BAB-072 Salix rotundifoliaSALIX242-08 07-BAB-108 Salix rotundifoliaSALIX001-08 07-JMS-1037 Salix scoulerianaSALIX002-08 07-JMS-1038 Salix scoulerianaSALIX003-08 07-JMS-1050 Salix scoulerianaSALIX006-08 07-JMS-1081 Salix scoulerianaSALIX014-08 07-JMS-1141 Salix scoulerianaSALIX015-08 07-JMS-1142 Salix scoulerianaSALIX017-08 07-JMS-1211 Salix scoulerianaSALIX025-08 07-JMS-1298 Salix scoulerianaSALIX026-08 07-JMS-1299 Salix scoulerianaSALIX033-08 07-JMS-1316 Salix scoulerianaSALIX034-08 07-JMS-1317 Salix scoulerianaSALIX052-08 07-JMS-1346 Salix scoulerianaSALIX062-08 07-JMS-1361 Salix scoulerianaSALIX070-08 07-JMS-0869 Salix scoulerianaSALIX071-08 07-JMS-0871 Salix scoulerianaSALIX076-08 07-JMS-0897 Salix scoulerianaSALIX085-08 07-JMS-0926 Salix scoulerianaSALIX086-08 07-JMS-0927 Salix scoulerianaSALIX087-08 07-JMS-0929 Salix scoulerianaSALIX107-08 07-JMS-0967 Salix scoulerianaSALIX128-08 07-DPQC-0008 Salix scoulerianaSALIX130-08 07-DPQC-0010 Salix scoulerianaSALIX133-08 07-DPQC-0013 Salix scoulerianaSALIX145-08 07-DPQC-0029 Salix scoulerianaSALIX146-08 07-DPQC-0030 Salix scoulerianaSALIX147-08 07-DPQC-0031 Salix scoulerianaSALIX179-08 07-DPQC-MI3 Salix scoulerianaSALIX180-08 07-DPQC-MI4 Salix scoulerianaSALIX222-08 07-BAB-015 Salix scoulerianaSALIX257-08 07-BAB-211 Salix scoulerianaSALIX280-08 07-BAB-288 Salix scoulerianaSALIX358-08 07-GWA-14392 Salix scoulerianaSALIX409-08 07-JMS-0898B Salix scoulerianaSALIX502-08 08-CEG-04 Salix scoulerianaSALIX503-08 08-CEG-05 Salix scoulerianaSALIX009-08 07-JMS-1119 Salix sitchensisSALIX010-08 07-JMS-1123 Salix sitchensisSALIX012-08 07-JMS-1132 Salix sitchensisSALIX018-08 07-JMS-1212 Salix sitchensisSALIX021-08 07-JMS-1259 Salix sitchensisSALIX022-08 07-JMS-1260 Salix sitchensisSALIX023-08 07-JMS-1281 Salix sitchensisSALIX029-08 07-JMS-1310 Salix sitchensisSALIX030-08 07-JMS-1313 Salix sitchensisSALIX032-08 07-JMS-1315 Salix sitchensisSALIX035-08 07-JMS-1319 Salix sitchensisSALIX051-08 07-JMS-1345 Salix sitchensisSALIX068-08 07-JMS-0864 Salix sitchensisSALIX072-08 07-JMS-0878 Salix sitchensis
SALIX073-08 07-JMS-0884 Salix sitchensisSALIX075-08 07-JMS-0891 Salix sitchensisSALIX079-08 07-JMS-0901 Salix sitchensisSALIX083-08 07-JMS-0924 Salix sitchensisSALIX084-08 07-JMS-0925 Salix sitchensisSALIX089-08 07-JMS-0941 Salix sitchensisSALIX092-08 07-JMS-0948 Salix sitchensisSALIX093-08 07-JMS-0949 Salix sitchensisSALIX094-08 07-JMS-0952 Salix sitchensisSALIX095-08 07-JMS-0953 Salix sitchensisSALIX097-08 07-JMS-0955 Salix sitchensisSALIX099-08 07-JMS-0957 Salix sitchensisSALIX101-08 07-JMS-0959 Salix sitchensisSALIX104-08 07-JMS-0962 Salix sitchensisSALIX105-08 07-JMS-0965 Salix sitchensisSALIX110-08 07-JMS-0977 Salix sitchensisSALIX123-08 06-DPQC-S2 Salix sitchensisSALIX124-08 07-DPQC-0001A Salix sitchensisSALIX126-08 07-DPQC-0002 Salix sitchensisSALIX132-08 07-DPQC-0012 Salix sitchensisSALIX136-08 07-DPQC-0016 Salix sitchensisSALIX166-08 07-DPQC-0054 Salix sitchensisSALIX172-08 07-DPQC-0060 Salix sitchensisSALIX182-08 07-DPQC-MINavChan Salix sitchensisSALIX186-08 07-DPQC-UBCFarm2 Salix sitchensisSALIX202-08 07-DPQC-VanIs1 Salix sitchensisSALIX203-08 07-DPQC-VanIs2 Salix sitchensisSALIX206-08 07-DPQC-WhistRd2 Salix sitchensisSALIX209-08 07-DPQC-WhistRd5 Salix sitchensisSALIX210-08 07-DPQC-WhistRd6 Salix sitchensisSALIX394-08 08-DPYC-03 Salix sitchensisSALIX408-08 07-JMS-0895 Salix sitchensisSALIX415-08 08-DPQC-0067 Salix sitchensisSALIX437-08 08-DPQC-0089 Salix sitchensisSALIX446-08 08-DPQC-0097 Salix sitchensisSALIX448-08 08-DPQC-0101 Salix sitchensisSALIX451-08 08-DPQC-0104 Salix sitchensisSALIX453-08 08-DPQC-0106 Salix sitchensisSALIX456-08 08-DPQC-0110 Salix sitchensisSALIX473-08 08-DPQC-0128 Salix sitchensisSALIX096-08 07-JMS-0954 Salix sp.SALIX108-08 07-JMS-0968 Salix sp.SALIX205-08 07-DPQC-WhistRd1 Salix sp.SALIX207-08 07-DPQC-WhistRd3 Salix sp.SALIX208-08 07-DPQC-WhistRd4 Salix sp.SALIX407-08 07-JMS-0887 Salix sp.SALIX410-08 07-PMP-21007 Salix sp.SALIX413-08 08-DPQC-0065 Salix sp.SALIX420-08 08-DPQC-0072 Salix sp.SALIX571-09 09-DPQC-17 Salix sp.SALIX572-09 09-DPQC-25 Salix sp.SALIX184-08 07-PMP-20885 Salix uvaursiSALIX518-08 08-GWA-14435 Salix vestitaSALIX519-08 08-GWA-14436 Salix vestitaSALIX204-08 07-DPQC-vimEur ope Salix viminalisSALIX043-08 07-JMS-1337B Salix x glaucaSALIX467-08 08-DPQC-0122 Salix bebbianaSALIX391-08 73-UBC-146342 Salix glaucaSALIX416-08 08-DPQC-0068 Salix lasiandraSALIX462-08 08-DPQC-0117 Salix prolixaSALIX201-08 07-DMP-UniGuelph Salix albaSALIX036-08 07-JMS-1320 Salix pentandraSALIX397-08 08-DPsmallSB Salix sepulcralisSALIX398-08 08-DPweepSB Salix sepulcralisSALIX327-08 07-GWA-14353 Salix serissimaSALIX328-08 07-GWA-14354 Salix serissimaSALIX016-08 07-JMS-1206 Salix x rubensSALIX020-08 07-JMS-1254 Salix x rubensSALIX027-08 07-JMS-1303 Salix x rubensSALIX019-08 07-JMS-1213 Salix x sepulcralis
SALIX081-08 07-JMS-0912 Salix x sepulcralisSALIX176-08 07-DPQC-weepJerBeach Salix x sepulcralis SALIX254-08 07-BAB-201 Salix interiorSALIX265-08 07-BAB-242 Salix interiorSALIX317-08 07-GWA-14343 Salix interiorSALIX525-08 08-GWA-14440 Salix interiorSALIX340-08 07-GWA-14369 Salix candidaSALIX343-08 07-GWA-14372 Salix cordataSALIX344-08 07-GWA-14375 Salix cordataSALIX114-08 07-EM-0359 Salix discolorSALIX319-08 07-GWA-14345 Salix discolorSALIX321-08 07-GWA-14347 Salix discolorSALIX329-08 07-GWA-14355 Salix discolorSALIX339-08 07-GWA-14368 Salix discolorSALIX431-08 08-DPQC-0083 Salix discolorSALIX345-08 07-GWA-14376 Salix eriocephalaSALIX346-08 07-GWA-14377 Salix eriocephalaSALIX127-08 07-DPQC-0003 Salix exiguaSALIX183-08 07-PMP-20866 Salix exiguaSALIX349-08 07-GWA-14381 Salix exiguaSALIX373-08 07-GWA-14429 Salix exiguaSALIX490-08 08-DPQC-0147 Salix exiguaSALIX499-08 08-CEG-02 Salix exiguaSALIX504-08 08-CEG-06 Salix exiguaSALIX330-08 07-GWA-14356 Salix humilisSALIX335-08 07-GWA-14364 Salix humilisSALIX112-08 07-JMS-0981 Salix interiorSALIX331-08 07-GWA-14360 Salix interiorSALIX336-08 07-GWA-14365 Salix interiorSALIX337-08 07-GWA-14366 Salix interiorSALIX338-08 07-GWA-14367 Salix interiorSALIX212-08 07-CAN-483931 Salix interior x eriocephalaSALIX217-08 07-CAN-532065 Salix interior x eriocephalaSALIX214-08 07-CAN-483950 Salix interior x petiolarisSALIX216-08 07-CAN-532063 Salix interior x petiolarisSALIX213-08 07-CAN-483942 Salix petiolarisSALIX318-08 07-GWA-14344 Salix petiolarisSALIX322-08 07-GWA-14348 Salix petiolarisSALIX325-08 07-GWA-14351 Salix petiolarisSALIX433-08 08-DPQC-0085 Salix planifoliaSALIX218-08 07-CAN-532084 Salix interiorSALIX215-08 07-CAN-532056 Salix interior x eriocephalaSALIX320-08 07-GWA-14346 Salix amygdaloidesSALIX506-08 08-CEG-08 Salix amygdaloides x goodingiiSALIX507-08 08-CEG-09 Salix amygdaloides x goodingiiSALIX508-08 08-CEG-10 Salix gooddingiiSALIX509-08 08-CEG-11 Salix gooddingiiSALIX295-08 07-CEG-24 Salix laevigataSALIX294-08 07-CEG-23 Salix laevigataSALIX324-08 07-GWA-14350 Salix lucidaSALIX326-08 07-GWA-14352 Salix lucidaSALIX333-08 07-GWA-14362 Salix nigraSALIX411-08 07-PMP-21162 Salix sp.SALIX412-08 07-PMP-21311 Salix sp.
0.01 changes
rbcL 10 PIC
G I II III IV V VII - 0 3 6 7 7II - 3 6 7 7III - 3 4 4IV - 1 1V - 0VI -
I+II
IIIV+VIIV
SALIX011-08 07-JMS-1124 Salix lasiandraSALIX013-08 07-JMS-1133 Salix lasiandraSALIX131-08 07-DPQC-0011 Salix lasiandraSALIX169-08 07-DPQC-0057 Salix lasiandraSALIX175-08 07-DPQC-natJerBeach Salix lasiandraSALIX178-08 07-JW-lasiand2 Salix lasiandraSALIX181-08 07-DPQC-MIBenBay Salix lasiandraSALIX185-08 07-DPQC-UBCFarm1 Salix lasiandraSALIX193-08 07-UBCBG-015012-0268-1974 Salix lasiandraSALIX168-08 07-DPQC-0056 Salix glaucaSALIX536-09 08-CAR-001 Salix reticulataSALIX538-09 08-CAR-003 Salix reticulataSALIX553-09 08-CAR-018 Salix reticulataSALIX556-09 08-CAR-021 Salix reticulataSALIX559-09 08-CAR-024 Salix reticulataSALIX566-09 08-CAR-031 Salix reticulata
SALIX174-08 07-DPQC-0062 Salix glaucaSALIX202-08 07-DPQC-VanIs1 Salix sitchensisSALIX203-08 07-DPQC-VanIs2 Salix sitchensis
SALIX540-09 08-CAR-005 Salix reticulataSALIX543-09 08-CAR-008 Salix reticulataSALIX123-08 06-DPQC-S2 Salix sitchensisSALIX129-08 07-DPQC-0009 Salix bebbianaSALIX542-09 08-CAR-007 Salix reticulataSALIX206-08 07-DPQC-WhistRd2 Salix sitchensisSALIX210-08 07-DPQC-WhistRd6 Salix sitchensis
SALIX384-08 75-UBC-155629 Salix alaxensisSALIX385-08 78-UBC-162474 Salix alaxensisSALIX390-08 73-UBC-149089 Salix arbusculoidesSALIX392-08 69-UBC-149365 Salix arbusculoidesSALIX137-08 07-DPQC-0017 Salix arctica
SALIX140-08 07-DPQC-0020 Salix arcticaSALIX141-08 07-DPQC-0024 Salix arctica
SALIX142-08 07-DPQC-0025 Salix arcticaSALIX144-08 07-DPQC-0028 Salix arcticaSALIX150-08 07-DPQC-0034 Salix arcticaSALIX155-08 07-DPQC-0039 Salix arcticaSALIX167-08 07-DPQC-0055 Salix arcticaSALIX170-08 07-DPQC-0058 Salix arcticaSALIX380-08 61-UBC-119656 Salix arcticaSALIX383-08 73-UBC-152112 Salix arcticaSALIX151-08 07-DPQC-0035 Salix barclayiSALIX153-08 07-DPQC-0037 Salix barclayiSALIX160-08 07-DPQC-0048 Salix barclayiSALIX162-08 07-DPQC-0050 Salix barclayiSALIX163-08 07-DPQC-0051 Salix barclayiSALIX164-08 07-DPQC-0052 Salix barclayiSALIX165-08 07-DPQC-0053 Salix barclayiSALIX171-08 07-DPQC-0059 Salix barclayiSALIX194-08 07-UBCBG-037155-1003-2004 Salix barclayiSALIX199-08 07-UBCBG-native3 Salix barclayiSALIX156-08 07-DPQC-0040 Salix barrattianaSALIX173-08 07-DPQC-0061 Salix barrattianaSALIX381-08 43-UBC-65549 Salix barrattianaSALIX005-08 07-JMS-1059 Salix bebbianaSALIX008-08 07-JMS-1094 Salix bebbianaSALIX113-08 07-EM-0315 Salix bebbianaSALIX115-08 07-EM-0472 Salix bebbianaSALIX118-08 07-EM-0583 Salix bebbianaSALIX119-08 07-EM-0596 Salix bebbianaSALIX121-08 07-EM-0628 Salix bebbianaSALIX122-08 06-DPQC-S1 Salix bebbianaSALIX134-08 07-DPQC-0014 Salix bebbianaSALIX198-08 07-UBCBG-037023-1003-2003 Salix bebbianaSALIX200-08 07-UBCBG-34916-5555-99 Salix bebbianaSALIX323-08 07-GWA-14349 Salix bebbianaSALIX348-08 07-GWA-14380 Salix bebbianaSALIX197-08 07-UBCBG-037007-1003-2003 Salix boothiiSALIX120-08 07-EM-0618 Salix candidaSALIX312-08 07-GWA-14337 Salix candidaSALIX313-08 07-GWA-14338 Salix candidaSALIX315-08 07-GWA-14340 Salix candidaSALIX143-08 07-DPQC-0027 Salix commutata
SALIX161-08 07-DPQC-0049 Salix commutataSALIX342-08 07-GWA-14371 Salix cordataSALIX135-08 07-DPQC-0015 Salix drummondianaSALIX191-08 07-UBCBG-013854-0013-1976 Salix elaeagnosSALIX219-08 07-CAN-532085 Salix eriocephalaSALIX334-08 07-GWA-14363 Salix eriocephalaSALIX309-08 07-GWA-14334 Salix famelicaSALIX310-08 07-GWA-14335 Salix famelicaSALIX007-08 07-JMS-1084 Salix glaucaSALIX388-08 46-UBC-61323 Salix glaucaSALIX389-08 47-UBC-60831 Salix glaucaSALIX386-08 56-UBC-58897 Salix glauca x brachycarpaSALIX387-08 56-UBC-58933 Salix glauca x brachycarpaSALIX024-08 07-JMS-1297 Salix hookerianaSALIX189-08 07-DPQC-UBCFarm5 Salix hookerianaSALIX192-08 07-UBCBG-037096-1003-2004 Salix hookerianaSALIX341-08 07-GWA-14370 Salix hookerianaSALIX195-08 07-UBCBG-023877-0368-1984 Salix magnificaSALIX377-08 07-WAGNER-MYR1 Salix myrtillifoliaSALIX196-08 07-UBCBG-26352-117-87 Salix nakamuranaSALIX138-08 07-DPQC-0018 Salix nivalisSALIX149-08 07-DPQC-0033 Salix nivalisSALIX157-08 07-DPQC-0041 Salix nivalisSALIX158-08 07-DPQC-0042 Salix nivalisSALIX148-08 07-DPQC-0032 Salix prolixaSALIX117-08 07-EM-0579 Salix pseudomonticolaSALIX314-08 07-GWA-14339 Salix pseudomonticolaSALIX316-08 07-GWA-14342 Salix pseudomonticolaSALIX116-08 07-EM-0475 Salix pseudomyrsinitesSALIX378-08 07-WAGNER-PS1 Salix pseudomyrsinitesSALIX379-08 07-WAGNER-PS2 Salix pseudomyrsinitesSALIX347-08 07-GWA-14379 Salix pyrifoliaSALIX537-09 08-CAR-002 Salix reticulataSALIX539-09 08-CAR-004 Salix reticulataSALIX541-09 08-CAR-006 Salix reticulataSALIX544-09 08-CAR-009 Salix reticulataSALIX545-09 08-CAR-010 Salix reticulataSALIX546-09 08-CAR-011 Salix reticulataSALIX547-09 08-CAR-012 Salix reticulataSALIX548-09 08-CAR-013 Salix reticulataSALIX550-09 08-CAR-015 Salix reticulataSALIX551-09 08-CAR-016 Salix reticulataSALIX552-09 08-CAR-017 Salix reticulataSALIX554-09 08-CAR-019 Salix reticulataSALIX555-09 08-CAR-020 Salix reticulataSALIX557-09 08-CAR-022 Salix reticulataSALIX558-09 08-CAR-023 Salix reticulataSALIX560-09 08-CAR-025 Salix reticulataSALIX561-09 08-CAR-026 Salix reticulataSALIX562-09 08-CAR-027 Salix reticulataSALIX563-09 08-CAR-028 Salix reticulataSALIX564-09 08-CAR-029 Salix reticulataSALIX565-09 08-CAR-030 Salix reticulataSALIX567-09 08-CAR-032 Salix reticulataSALIX568-09 08-CAR-033 Salix reticulataSALIX003-08 07-JMS-1050 Salix scoulerianaSALIX006-08 07-JMS-1081 Salix scoulerianaSALIX014-08 07-JMS-1141 Salix scoulerianaSALIX015-08 07-JMS-1142 Salix scoulerianaSALIX017-08 07-JMS-1211 Salix scoulerianaSALIX025-08 07-JMS-1298 Salix scoulerianaSALIX026-08 07-JMS-1299 Salix scoulerianaSALIX128-08 07-DPQC-0008 Salix scoulerianaSALIX133-08 07-DPQC-0013 Salix scoulerianaSALIX145-08 07-DPQC-0029 Salix scoulerianaSALIX146-08 07-DPQC-0030 Salix scoulerianaSALIX179-08 07-DPQC-MI3 Salix scoulerianaSALIX180-08 07-DPQC-MI4 Salix scoulerianaSALIX010-08 07-JMS-1123 Salix sitchensisSALIX012-08 07-JMS-1132 Salix sitchensisSALIX018-08 07-JMS-1212 Salix sitchensisSALIX023-08 07-JMS-1281 Salix sitchensis
SALIX124-08 07-DPQC-0001A Salix sitchensisSALIX126-08 07-DPQC-0002 Salix sitchensisSALIX136-08 07-DPQC-0016 Salix sitchensisSALIX166-08 07-DPQC-0054 Salix sitchensisSALIX172-08 07-DPQC-0060 Salix sitchensisSALIX182-08 07-DPQC-MINavChan Salix sitchensisSALIX186-08 07-DPQC-UBCFarm2 Salix sitchensisSALIX209-08 07-DPQC-WhistRd5 Salix sitchensisSALIX205-08 07-DPQC-WhistRd1 Salix sp.SALIX207-08 07-DPQC-WhistRd3 Salix sp.SALIX208-08 07-DPQC-WhistRd4 Salix sp.SALIX184-08 07-PMP-20885 Salix uvaursiSALIX204-08 07-DPQC-vimEurope Salix viminalisSALIX391-08 73-UBC-146342 Salix glaucaSALIX549-09 08-CAR-014 Salix reticulata
SALIX139-08 07-DPQC-0019 Salix barrattianaSALIX382-08 73-UBC-149084 Salix barrattiana
SALIX340-08 07-GWA-14369 Salix candidaSALIX343-08 07-GWA-14372 Salix cordataSALIX344-08 07-GWA-14375 Salix cordataSALIX114-08 07-EM-0359 Salix discolor
SALIX319-08 07-GWA-14345 Salix discolorSALIX329-08 07-GWA-14355 Salix discolorSALIX339-08 07-GWA-14368 Salix discolorSALIX345-08 07-GWA-14376 Salix eriocephalaSALIX346-08 07-GWA-14377 Salix eriocephalaSALIX330-08 07-GWA-14356 Salix humilisSALIX335-08 07-GWA-14364 Salix humilisSALIX214-08 07-CAN-483950 Salix interior x petiolarisSALIX213-08 07-CAN-483942 Salix petiolarisSALIX318-08 07-GWA-14344 Salix petiolarisSALIX322-08 07-GWA-14348 Salix petiolarisSALIX325-08 07-GWA-14351 Salix petiolaris
SALIX127-08 07-DPQC-0003 Salix exiguaSALIX183-08 07-PMP-20866 Salix exiguaSALIX112-08 07-JMS-0981 Salix interiorSALIX317-08 07-GWA-14343 Salix interiorSALIX331-08 07-GWA-14360 Salix interiorSALIX336-08 07-GWA-14365 Salix interiorSALIX337-08 07-GWA-14366 Salix interiorSALIX338-08 07-GWA-14367 Salix interiorSALIX212-08 07-CAN-483931 Salix interior x eriocephalaSALIX217-08 07-CAN-532065 Salix interior x eriocephalaSALIX216-08 07-CAN-532063 Salix interior x petiolarisSALIX201-08 07-DMP-UniGuelph Salix albaSALIX327-08 07-GWA-14353 Salix serissimaSALIX328-08 07-GWA-14354 Salix serissima
SALIX176-08 07-DPQC-weepJerBeach Salix x sepulcralisSALIX332-08 07-GWA-14361 Salix amygdaloidesSALIX324-08 07-GWA-14350 Salix lucidaSALIX326-08 07-GWA-14352 Salix lucidaSALIX333-08 07-GWA-14362 Salix nigra
SALIX320-08 07-GWA-14346 Salix amygdaloides0.1 changes
G I II III IV V VII - 1 5 4 4 5II - 6 5 5 6III - 3 3 4IV - 2 3V - 3VI -
trnH-psbA 16 PIC
I
II
VVI IIIIV
SALIX003-08 07-JMS-1050 Salix scoulerianaSALIX014-08 07-JMS-1141 Salix scoulerianaSALIX015-08 07-JMS-1142 Salix scoulerianaSALIX025-08 07-JMS-1298 Salix scoulerianaSALIX026-08 07-JMS-1299 Salix scoulerianaSALIX145-08 07-DPQC-0029 Salix scoulerianaSALIX146-08 07-DPQC-0030 Salix scoulerianaSALIX136-08 07-DPQC-0016 Salix sitchensisSALIX174-08 07-DPQC-0062 Salix glauca
SALIX202-08 07-DPQC-VanIs1 Salix sitchensisSALIX203-08 07-DPQC-VanIs2 Salix sitchensis
SALIX383-08 73-UBC-152112 Salix arcticaSALIX179-08 07-DPQC-MI3 Salix scoulerianaSALIX180-08 07-DPQC-MI4 Salix scouleriana
SALIX197-08 07-UBCBG-037007-1003-2003 Salix boothiiSALIX316-08 07-GWA-14342 Salix pseudomonticolaSALIX219-08 07-CAN-532085 Salix eriocephalaSALIX378-08 07-WAGNER-PS1 Salix pseudomyrsinites
SALIX385-08 78-UBC-162474 Salix alaxensisSALIX390-08 73-UBC-149089 Salix arbusculoidesSALIX392-08 69-UBC-149365 Salix arbusculoidesSALIX137-08 07-DPQC-0017 Salix arcticaSALIX140-08 07-DPQC-0020 Salix arcticaSALIX141-08 07-DPQC-0024 Salix arcticaSALIX142-08 07-DPQC-0025 Salix arcticaSALIX144-08 07-DPQC-0028 Salix arcticaSALIX150-08 07-DPQC-0034 Salix arcticaSALIX155-08 07-DPQC-0039 Salix arcticaSALIX167-08 07-DPQC-0055 Salix arcticaSALIX170-08 07-DPQC-0058 Salix arcticaSALIX380-08 61-UBC-119656 Salix arcticaSALIX151-08 07-DPQC-0035 Salix barclayi
SALIX153-08 07-DPQC-0037 Salix barclayiSALIX162-08 07-DPQC-0050 Salix barclayiSALIX163-08 07-DPQC-0051 Salix barclayiSALIX164-08 07-DPQC-0052 Salix barclayiSALIX165-08 07-DPQC-0053 Salix barclayiSALIX171-08 07-DPQC-0059 Salix barclayiSALIX194-08 07-UBCBG-037155-1003-2004 Salix barclayiSALIX199-08 07-UBCBG-native3 Salix barclayiSALIX139-08 07-DPQC-0019 Salix barrattianaSALIX156-08 07-DPQC-0040 Salix barrattianaSALIX173-08 07-DPQC-0061 Salix barrattianaSALIX381-08 43-UBC-65549 Salix barrattianaSALIX382-08 73-UBC-149084 Salix barrattianaSALIX005-08 07-JMS-1059 Salix bebbianaSALIX008-08 07-JMS-1094 Salix bebbianaSALIX115-08 07-EM-0472 Salix bebbianaSALIX118-08 07-EM-0583 Salix bebbianaSALIX121-08 07-EM-0628 Salix bebbianaSALIX122-08 06-DPQC-S1 Salix bebbianaSALIX134-08 07-DPQC-0014 Salix bebbianaSALIX198-08 07-UBCBG-037023-1003-2003 Salix bebbianaSALIX200-08 07-UBCBG-34916-5555-99 Salix bebbianaSALIX323-08 07-GWA-14349 Salix bebbianaSALIX348-08 07-GWA-14380 Salix bebbiana
SALIX120-08 07-EM-0618 Salix candidaSALIX312-08 07-GWA-14337 Salix candidaSALIX313-08 07-GWA-14338 Salix candidaSALIX315-08 07-GWA-14340 Salix candidaSALIX143-08 07-DPQC-0027 Salix commutataSALIX161-08 07-DPQC-0049 Salix commutata
SALIX342-08 07-GWA-14371 Salix cordataSALIX135-08 07-DPQC-0015 Salix drummondianaSALIX191-08 07-UBCBG-013854-0013-1976 Salix elaeagnosSALIX334-08 07-GWA-14363 Salix eriocephalaSALIX309-08 07-GWA-14334 Salix famelicaSALIX310-08 07-GWA-14335 Salix famelicaSALIX007-08 07-JMS-1084 Salix glaucaSALIX168-08 07-DPQC-0056 Salix glaucaSALIX388-08 46-UBC-61323 Salix glaucaSALIX389-08 47-UBC-60831 Salix glaucaSALIX386-08 56-UBC-58897 Salix glauca x brachycarpaSALIX387-08 56-UBC-58933 Salix glauca x brachycarpaSALIX024-08 07-JMS-1297 Salix hookerianaSALIX189-08 07-DPQC-UBCFarm5 Salix hookerianaSALIX192-08 07-UBCBG-037096-1003-2004 Salix hookerianaSALIX341-08 07-GWA-14370 Salix hookerianaSALIX195-08 07-UBCBG-023877-0368-1984 Salix magnificaSALIX377-08 07-WAGNER-MYR1 Salix myrtillifolia
SALIX196-08 07-UBCBG-26352-117-87 Salix nakamuranaSALIX138-08 07-DPQC-0018 Salix nivalisSALIX149-08 07-DPQC-0033 Salix nivalisSALIX157-08 07-DPQC-0041 Salix nivalisSALIX158-08 07-DPQC-0042 Salix nivalisSALIX148-08 07-DPQC-0032 Salix prolixa
SALIX314-08 07-GWA-14339 Salix pseudomonticolaSALIX116-08 07-EM-0475 Salix pseudomyrsinitesSALIX379-08 07-WAGNER-PS2 Salix pseudomyrsinitesSALIX347-08 07-GWA-14379 Salix pyrifoliaSALIX006-08 07-JMS-1081 Salix scoulerianaSALIX128-08 07-DPQC-0008 Salix scoulerianaSALIX133-08 07-DPQC-0013 Salix scoulerianaSALIX010-08 07-JMS-1123 Salix sitchensisSALIX012-08 07-JMS-1132 Salix sitchensisSALIX018-08 07-JMS-1212 Salix sitchensisSALIX023-08 07-JMS-1281 Salix sitchensisSALIX124-08 07-DPQC-0001A Salix sitchensisSALIX126-08 07-DPQC-0002 Salix sitchensisSALIX166-08 07-DPQC-0054 Salix sitchensisSALIX182-08 07-DPQC-MINavChan Salix sitchensisSALIX186-08 07-DPQC-UBCFarm2 Salix sitchensisSALIX206-08 07-DPQC-WhistRd2 Salix sitchensisSALIX209-08 07-DPQC-WhistRd5 Salix sitchensisSALIX210-08 07-DPQC-WhistRd6 Salix sitchensisSALIX205-08 07-DPQC-WhistRd1 Salix sp.SALIX207-08 07-DPQC-WhistRd3 Salix sp.SALIX208-08 07-DPQC-WhistRd4 Salix sp.SALIX184-08 07-PMP-20885 Salix uvaursiSALIX204-08 07-DPQC-vimEurope Salix viminalisSALIX391-08 73-UBC-146342 Salix glaucaSALIX011-08 07-JMS-1124 Salix lasiandraSALIX013-08 07-JMS-1133 Salix lasiandraSALIX131-08 07-DPQC-0011 Salix lasiandraSALIX169-08 07-DPQC-0057 Salix lasiandraSALIX175-08 07-DPQC-natJerBeach Salix lasiandraSALIX178-08 07-JW-lasiand2 Salix lasiandraSALIX181-08 07-DPQC-MIBenBay Salix lasiandraSALIX185-08 07-DPQC-UBCFarm1 Salix lasiandraSALIX193-08 07-UBCBG-015012-0268-1974 Salix lasiandra
SALIX201-08 07-DMP-UniGuelph Salix albaSALIX327-08 07-GWA-14353 Salix serissimaSALIX328-08 07-GWA-14354 Salix serissimaSALIX176-08 07-DPQC-weepJerBeach Salix x sepulcralis
SALIX183-08 07-PMP-20866 Salix exiguaSALIX112-08 07-JMS-0981 Salix interiorSALIX317-08 07-GWA-14343 Salix interiorSALIX331-08 07-GWA-14360 Salix interiorSALIX336-08 07-GWA-14365 Salix interiorSALIX337-08 07-GWA-14366 Salix interiorSALIX338-08 07-GWA-14367 Salix interiorSALIX212-08 07-CAN-483931 Salix interior x eriocephalaSALIX217-08 07-CAN-532065 Salix interior x eriocephalaSALIX216-08 07-CAN-532063 Salix interior x petiolarisSALIX215-08 07-CAN-532056 Salix interior x eriocephala
SALIX340-08 07-GWA-14369 Salix candidaSALIX345-08 07-GWA-14376 Salix eriocephalaSALIX346-08 07-GWA-14377 Salix eriocephalaSALIX335-08 07-GWA-14364 Salix humilisSALIX213-08 07-CAN-483942 Salix petiolarisSALIX325-08 07-GWA-14351 Salix petiolaris
SALIX319-08 07-GWA-14345 Salix discolorSALIX339-08 07-GWA-14368 Salix discolorSALIX330-08 07-GWA-14356 Salix humilis
SALIX324-08 07-GWA-14350 Salix lucidaSALIX326-08 07-GWA-14352 Salix lucida
SALIX320-08 07-GWA-14346 Salix amygdaloidesSALIX332-08 07-GWA-14361 Salix amygdaloidesSALIX333-08 07-GWA-14362 Salix nigra
SALIX344-08 07-GWA-14375 Salix cordataSALIX321-08 07-GWA-14347 Salix discolorSALIX329-08 07-GWA-14355 Salix discolorSALIX214-08 07-CAN-483950 Salix interior x petiolarisSALIX318-08 07-GWA-14344 Salix petiolarisSALIX322-08 07-GWA-14348 Salix petiolaris
0.1 changes
G I II III IV V VII - 2 5 10 8 9II - 5 10 8 9III - 7 5 6IV - 2 3V - 1VI -
atpF-atpH 20 PIC
IIIVI
V
II
I
SALIX140-08 07-DPQC-0020 Salix arcticaSALIX142-08 07-DPQC-0025 Salix arcticaSALIX388-08 46-UBC-61323 Salix glaucaSALIX113-08 07-EM-0315 Salix bebbianaSALIX219-08 07-CAN-532085 Salix eriocephalaSALIX138-08 07-DPQC-0018 Salix nivalisSALIX172-08 07-DPQC-0060 Salix sitchensis
SALIX384-08 75-UBC-155629 Salix alaxensisSALIX385-08 78-UBC-162474 Salix alaxensisSALIX390-08 73-UBC-149089 Salix arbusculoidesSALIX392-08 69-UBC-149365 Salix arbusculoidesSALIX137-08 07-DPQC-0017 Salix arcticaSALIX141-08 07-DPQC-0024 Salix arcticaSALIX144-08 07-DPQC-0028 Salix arcticaSALIX150-08 07-DPQC-0034 Salix arcticaSALIX155-08 07-DPQC-0039 Salix arcticaSALIX167-08 07-DPQC-0055 Salix arcticaSALIX170-08 07-DPQC-0058 Salix arcticaSALIX380-08 61-UBC-119656 Salix arcticaSALIX383-08 73-UBC-152112 Salix arcticaSALIX151-08 07-DPQC-0035 Salix barclayiSALIX153-08 07-DPQC-0037 Salix barclayiSALIX160-08 07-DPQC-0048 Salix barclayiSALIX162-08 07-DPQC-0050 Salix barclayiSALIX163-08 07-DPQC-0051 Salix barclayiSALIX164-08 07-DPQC-0052 Salix barclayiSALIX165-08 07-DPQC-0053 Salix barclayiSALIX171-08 07-DPQC-0059 Salix barclayiSALIX194-08 07-UBCBG-037155-1003-2004 Salix barclayiSALIX199-08 07-UBCBG-native3 Salix barclayiSALIX139-08 07-DPQC-0019 Salix barrattianaSALIX156-08 07-DPQC-0040 Salix barrattianaSALIX173-08 07-DPQC-0061 Salix barrattianaSALIX381-08 43-UBC-65549 Salix barrattianaSALIX382-08 73-UBC-149084 Salix barrattianaSALIX005-08 07-JMS-1059 Salix bebbianaSALIX008-08 07-JMS-1094 Salix bebbianaSALIX115-08 07-EM-0472 Salix bebbianaSALIX118-08 07-EM-0583 Salix bebbianaSALIX119-08 07-EM-0596 Salix bebbianaSALIX121-08 07-EM-0628 Salix bebbianaSALIX122-08 06-DPQC-S1 Salix bebbianaSALIX129-08 07-DPQC-0009 Salix bebbianaSALIX134-08 07-DPQC-0014 Salix bebbianaSALIX198-08 07-UBCBG-037023-1003-2003 Salix bebbianaSALIX200-08 07-UBCBG-34916-5555-99 Salix bebbianaSALIX323-08 07-GWA-14349 Salix bebbianaSALIX348-08 07-GWA-14380 Salix bebbianaSALIX197-08 07-UBCBG-037007-1003-2003 Salix boothiiSALIX120-08 07-EM-0618 Salix candidaSALIX312-08 07-GWA-14337 Salix candidaSALIX313-08 07-GWA-14338 Salix candidaSALIX315-08 07-GWA-14340 Salix candidaSALIX143-08 07-DPQC-0027 Salix commutataSALIX161-08 07-DPQC-0049 Salix commutataSALIX342-08 07-GWA-14371 Salix cordata
SALIX135-08 07-DPQC-0015 Salix drummondianaSALIX191-08 07-UBCBG-013854-0013-1976 Salix elaeagnosSALIX334-08 07-GWA-14363 Salix eriocephalaSALIX309-08 07-GWA-14334 Salix famelicaSALIX310-08 07-GWA-14335 Salix famelicaSALIX007-08 07-JMS-1084 Salix glaucaSALIX168-08 07-DPQC-0056 Salix glaucaSALIX174-08 07-DPQC-0062 Salix glaucaSALIX389-08 47-UBC-60831 Salix glaucaSALIX386-08 56-UBC-58897 Salix glauca x brachycarpaSALIX387-08 56-UBC-58933 Salix glauca x brachycarpaSALIX024-08 07-JMS-1297 Salix hookerianaSALIX192-08 07-UBCBG-037096-1003-2004 Salix hookeriana
SALIX341-08 07-GWA-14370 Salix hookerianaSALIX195-08 07-UBCBG-023877-0368-1984 Salix magnifica
SALIX377-08 07-WAGNER-MYR1 Salix myrtillifoliaSALIX196-08 07-UBCBG-26352-117-87 Salix nakamuranaSALIX149-08 07-DPQC-0033 Salix nivalisSALIX157-08 07-DPQC-0041 Salix nivalisSALIX158-08 07-DPQC-0042 Salix nivalisSALIX148-08 07-DPQC-0032 Salix prolixaSALIX117-08 07-EM-0579 Salix pseudomonticolaSALIX314-08 07-GWA-14339 Salix pseudomonticolaSALIX316-08 07-GWA-14342 Salix pseudomonticolaSALIX116-08 07-EM-0475 Salix pseudomyrsinitesSALIX378-08 07-WAGNER-PS1 Salix pseudomyrsinitesSALIX379-08 07-WAGNER-PS2 Salix pseudomyrsinitesSALIX347-08 07-GWA-14379 Salix pyrifoliaSALIX003-08 07-JMS-1050 Salix scoulerianaSALIX006-08 07-JMS-1081 Salix scoulerianaSALIX014-08 07-JMS-1141 Salix scoulerianaSALIX015-08 07-JMS-1142 Salix scoulerianaSALIX017-08 07-JMS-1211 Salix scoulerianaSALIX025-08 07-JMS-1298 Salix scoulerianaSALIX026-08 07-JMS-1299 Salix scoulerianaSALIX128-08 07-DPQC-0008 Salix scoulerianaSALIX133-08 07-DPQC-0013 Salix scoulerianaSALIX145-08 07-DPQC-0029 Salix scoulerianaSALIX146-08 07-DPQC-0030 Salix scoulerianaSALIX179-08 07-DPQC-MI3 Salix scoulerianaSALIX180-08 07-DPQC-MI4 Salix scoulerianaSALIX010-08 07-JMS-1123 Salix sitchensisSALIX012-08 07-JMS-1132 Salix sitchensisSALIX018-08 07-JMS-1212 Salix sitchensisSALIX023-08 07-JMS-1281 Salix sitchensisSALIX123-08 06-DPQC-S2 Salix sitchensisSALIX124-08 07-DPQC-0001A Salix sitchensisSALIX126-08 07-DPQC-0002 Salix sitchensisSALIX136-08 07-DPQC-0016 Salix sitchensisSALIX166-08 07-DPQC-0054 Salix sitchensisSALIX182-08 07-DPQC-MINavChan Salix sitchensisSALIX186-08 07-DPQC-UBCFarm2 Salix sitchensisSALIX202-08 07-DPQC-VanIs1 Salix sitchensisSALIX203-08 07-DPQC-VanIs2 Salix sitchensisSALIX206-08 07-DPQC-WhistRd2 Salix sitchensis
SALIX209-08 07-DPQC-WhistRd5 Salix sitchensisSALIX210-08 07-DPQC-WhistRd6 Salix sitchensisSALIX205-08 07-DPQC-WhistRd1 Salix sp.SALIX207-08 07-DPQC-WhistRd3 Salix sp.SALIX208-08 07-DPQC-WhistRd4 Salix sp.SALIX184-08 07-PMP-20885 Salix uvaursi
SALIX204-08 07-DPQC-vimEur ope Salix viminalisSALIX391-08 73-UBC-146342 Salix glauca
SALIX011-08 07-JMS-1124 Salix lasiandraSALIX013-08 07-JMS-1133 Salix lasiandraSALIX131-08 07-DPQC-0011 Salix lasiandraSALIX169-08 07-DPQC-0057 Salix lasiandraSALIX175-08 07-DPQC-natJerBeach Salix lasiandraSALIX178-08 07-JW-lasiand2 Salix lasiandraSALIX181-08 07-DPQC-MIBenBay Salix lasiandraSALIX185-08 07-DPQC-UBCFarm1 Salix lasiandraSALIX193-08 07-UBCBG-015012-0268-1974 Salix lasiandra
SALIX332-08 07-GWA-14361 Salix amygdaloidesSALIX324-08 07-GWA-14350 Salix lucidaSALIX326-08 07-GWA-14352 Salix lucidaSALIX333-08 07-GWA-14362 Salix nigraSALIX201-08 07-DMP-UniGuelph Salix alba
SALIX320-08 07-GWA-14346 Salix amygdaloidesSALIX327-08 07-GWA-14353 Salix serissimaSALIX328-08 07-GWA-14354 Salix serissima
SALIX176-08 07-DPQC-weepJerBeach Salix x sepulcralisSALIX183-08 07-PMP-20866 Salix exiguaSALIX112-08 07-JMS-0981 Salix interiorSALIX331-08 07-GWA-14360 Salix interiorSALIX336-08 07-GWA-14365 Salix interiorSALIX337-08 07-GWA-14366 Salix interiorSALIX338-08 07-GWA-14367 Salix interiorSALIX212-08 07-CAN-483931 Salix interior x eriocephalaSALIX217-08 07-CAN-532065 Salix interior x eriocephalaSALIX215-08 07-CAN-532056 Salix interior x eriocephala
SALIX317-08 07-GWA-14343 Salix interiorSALIX340-08 07-GWA-14369 Salix candidaSALIX343-08 07-GWA-14372 Salix cordataSALIX114-08 07-EM-0359 Salix discolorSALIX319-08 07-GWA-14345 Salix discolorSALIX321-08 07-GWA-14347 Salix discolorSALIX329-08 07-GWA-14355 Salix discolorSALIX339-08 07-GWA-14368 Salix discolorSALIX345-08 07-GWA-14376 Salix eriocephalaSALIX346-08 07-GWA-14377 Salix eriocephalaSALIX330-08 07-GWA-14356 Salix humilisSALIX335-08 07-GWA-14364 Salix humilisSALIX214-08 07-CAN-483950 Salix interior x petiolarisSALIX213-08 07-CAN-483942 Salix petiolarisSALIX318-08 07-GWA-14344 Salix petiolarisSALIX322-08 07-GWA-14348 Salix petiolarisSALIX325-08 07-GWA-14351 Salix petiolaris
0.1 changes
psbK-psbI 25 PICG I II III IV V VII - 4 8 9 16 20II - 4 5 12 16III - 1 8 12IV - 9 13V - 4VI -
I
II IVIIIVI
V
SALIX197-08 07-UBCBG-037007-1003-2003 Salix boothiiSALIX376-08 07-GWA-14436A Salix eastwoodiaeSALIX090-08 07-JMS-0942 Salix hookerianaSALIX234-08 07-BAB-047A Salix arcticaSALIX240-08 07-BAB-096A Salix arctica x polarisSALIX137-08 07-DPQC-0017 Salix arcticaSALIX141-08 07-DPQC-0024 Salix arcticaSALIX170-08 07-DPQC-0058 Salix arcticaSALIX232-08 07-BAB-033 Salix arctica x glaucaSALIX057-08 07-JMS-1351 Salix athabascensisSALIX160-08 07-DPQC-0048 Salix barclayiSALIX165-08 07-DPQC-0053 Salix barclayiSALIX270-08 07-BAB-267 Salix barclayiSALIX272-08 07-BAB-269 Salix barclayiSALIX063-08 07-JMS-1362 Salix bebbianaSALIX119-08 07-EM-0596 Salix bebbianaSALIX122-08 06-DPQC-S1 Salix bebbianaSALIX323-08 07-GWA-14349 Salix bebbianaSALIX424-08 08-DPQC-0076 Salix bebbianaSALIX440-08 08-DPQC-0092 Salix bebbianaSALIX356-08 07-GWA-14388 Salix boothiiSALIX532-08 08-GWA-14449 Salix calicolaSALIX481-08 08-DPQC-0136 Salix candidaSALIX007-08 07-JMS-1084 Salix glaucaSALIX361-08 07-GWA-14396 Salix glaucaSALIX368-08 07-GWA-14424 Salix glaucaSALIX387-08 56-UBC-58933 Salix glauca x brachycarpaSALIX401-08 07-DW-6001 Salix myricoidesSALIX269-08 07-BAB-251 Salix niphocladaSALIX363-08 07-GWA-14409 Salix petrophilaSALIX061-08 07-JMS-1360 Salix prolixaSALIX436-08 08-DPQC-0088 Salix prolixaSALIX282-08 07-BAB-292 Salix reticulataSALIX548-09 08-CAR-013 Salix reticulataSALIX554-09 08-CAR-019 Salix reticulataSALIX557-09 08-CAR-022 Salix reticulataSALIX558-09 08-CAR-023 Salix reticulataSALIX562-09 08-CAR-027 Salix reticulataSALIX563-09 08-CAR-028 Salix reticulataSALIX231-08 07-BAB-032 Salix richardsoniiSALIX237-08 07-BAB-072 Salix rotundifoliaSALIX126-08 07-DPQC-0002 Salix sitchensisSALIX415-08 08-DPQC-0067 Salix sitchensisSALIX446-08 08-DPQC-0097 Salix sitchensisSALIX453-08 08-DPQC-0106 Salix sitchensisSALIX043-08 07-JMS-1337B Salix x glaucaSALIX011-08 07-JMS-1124 Salix lasiandraSALIX013-08 07-JMS-1133 Salix lasiandraSALIX028-08 07-JMS-1309 Salix lasiandraSALIX031-08 07-JMS-1314 Salix lasiandraSALIX055-08 07-JMS-1349 Salix lasiandraSALIX056-08 07-JMS-1350 Salix lasiandraSALIX067-08 07-JMS-0863 Salix lasiandraSALIX077-08 07-JMS-0899 Salix lasiandraSALIX080-08 07-JMS-0905 Salix lasiandraSALIX082-08 07-JMS-0923 Salix lasiandraSALIX088-08 07-JMS-0937 Salix lasiandraSALIX091-08 07-JMS-0943 Salix lasiandraSALIX106-08 07-JMS-0966 Salix lasiandraSALIX131-08 07-DPQC-0011 Salix lasiandraSALIX169-08 07-DPQC-0057 Salix lasiandraSALIX175-08 07-DPQC-natJerBeach Salix lasiandraSALIX178-08 07-JW-lasiand2 Salix lasiandraSALIX181-08 07-DPQC-MIBenBay Salix lasiandraSALIX185-08 07-DPQC-UBCFarm1 Salix lasiandraSALIX193-08 07-UBCBG-015012-0268-1974 Salix lasiandraSALIX251-08 07-BAB-198 Salix lasiandraSALIX262-08 07-BAB-231 Salix lasiandraSALIX296-08 07-CEG-25 Salix lasiandraSALIX351-08 07-GWA-14383 Salix lasiandraSALIX393-08 08-DPYC-02 Salix lasiandraSALIX406-08 07-CEG-17 Salix lasiandraSALIX423-08 08-DPQC-0075 Salix lasiandraSALIX425-08 08-DPQC-0077 Salix lasiandraSALIX426-08 08-DPQC-0078 Salix lasiandraSALIX434-08 08-DPQC-0086 Salix lasiandraSALIX438-08 08-DPQC-0090 Salix lasiandraSALIX445-08 08-DPQC-0096 Salix lasiandraSALIX447-08 08-DPQC-0100 Salix lasiandraSALIX449-08 08-DPQC-0102 Salix lasiandraSALIX476-08 08-DPQC-0131 Salix lasiandraSALIX477-08 08-DPQC-0132 Salix lasiandraSALIX108-08 07-JMS-0968 Salix sp.SALIX407-08 07-JMS-0887 Salix sp.SALIX070-08 07-JMS-0869 Salix scoulerianaSALIX071-08 07-JMS-0871 Salix scoulerianaSALIX076-08 07-JMS-0897 Salix scoulerianaSALIX087-08 07-JMS-0929 Salix scoulerianaSALIX107-08 07-JMS-0967 Salix scoulerianaSALIX179-08 07-DPQC-MI3 Salix scoulerianaSALIX180-08 07-DPQC-MI4 Salix scoulerianaSALIX409-08 07-JMS-0898B Salix scoulerianaSALIX394-08 08-DPYC-03 Salix sitchensisSALIX096-08 07-JMS-0954 Salix sp.SALIX039-08 07-JMS-1334 Salix arcticaSALIX156-08 07-DPQC-0040 Salix barrattianaSALIX258-08 07-BAB-224 Salix myrtillifoliaSALIX271-08 07-BAB-268 Salix pedicellarisSALIX283-08 07-BAB-293 Salix arcticaSALIX464-08 08-DPQC-0119 Salix glauca villosaSALIX480-08 08-DPQC-0135 Salix glauca villosaSALIX567-09 08-CAR-032 Salix reticulataSALIX298-08 07-CEG-26 Salix arizonicaSALIX511-08 08-CEG-13 Salix geyerianaSALIX299-08 07-CEG-28 Salix monticolaSALIX123-08 06-DPQC-S2 Salix sitchensisSALIX129-08 07-DPQC-0009 Salix bebbianaSALIX414-08 08-DPQC-0066 Salix bebbianaSALIX442-08 08-DPQC-0094 Salix bebbianaSALIX355-08 07-GWA-14387 Salix boothiiSALIX357-08 07-GWA-14390 Salix drummondianaSALIX359-08 07-GWA-14393 Salix drummondianaSALIX144-08 07-DPQC-0028 Salix arcticaSALIX273-08 07-BAB-270 Salix pedicellarisSALIX266-08 07-BAB-246 Salix barclayiSALIX230-08 07-BAB-031 Salix richardsoniiSALIX371-08 07-GWA-14427 Salix lasiolepisSALIX372-08 07-GWA-14428 Salix lasiolepisSALIX220-08 07-BAB-013 Salix planifoliaSALIX222-08 07-BAB-015 Salix scoulerianaSALIX226-08 07-BAB-023 Salix alaxensisSALIX239-08 07-BAB-088 Salix alaxensisSALIX250-08 07-BAB-197 Salix alaxensisSALIX261-08 07-BAB-230 Salix alaxensisSALIX263-08 07-BAB-232 Salix alaxensisSALIX264-08 07-BAB-237 Salix alaxensisSALIX384-08 75-UBC-155629 Salix alaxensisSALIX385-08 78-UBC-162474 Salix alaxensisSALIX534-08 08-GWA-14451 Salix alaxensis x drummondianaSALIX060-08 07-JMS-1354 Salix arbusculoidesSALIX229-08 07-BAB-030 Salix arbusculoidesSALIX390-08 73-UBC-149089 Salix arbusculoidesSALIX392-08 69-UBC-149365 Salix arbusculoidesSALIX037-08 07-JMS-1330 Salix arcticaSALIX038-08 07-JMS-1333 Salix arcticaSALIX049-08 07-JMS-1343 Salix arcticaSALIX140-08 07-DPQC-0020 Salix arcticaSALIX142-08 07-DPQC-0025 Salix arcticaSALIX150-08 07-DPQC-0034 Salix arcticaSALIX154-08 07-DPQC-0038 Salix arcticaSALIX155-08 07-DPQC-0039 Salix arcticaSALIX167-08 07-DPQC-0055 Salix arcticaSALIX235-08 07-BAB-047B Salix arcticaSALIX380-08 61-UBC-119656 Salix arcticaSALIX383-08 73-UBC-152112 Salix arcticaSALIX247-08 07-BAB-186 Salix arctica x polarisSALIX297-08 07-CEG-27 Salix arizonicaSALIX510-08 08-CEG-12 Salix arizonicaSALIX517-08 08-GWA-14434 Salix athabascensisSALIX040-08 07-JMS-1335 Salix barclayiSALIX041-08 07-JMS-1336 Salix barclayiSALIX047-08 07-JMS-1341 Salix barclayiSALIX048-08 07-JMS-1342 Salix barclayiSALIX050-08 07-JMS-1344B Salix barclayiSALIX053-08 07-JMS-1347 Salix barclayiSALIX151-08 07-DPQC-0035 Salix barclayiSALIX152-08 07-DPQC-0036 Salix barclayiSALIX153-08 07-DPQC-0037 Salix barclayiSALIX162-08 07-DPQC-0050 Salix barclayiSALIX163-08 07-DPQC-0051 Salix barclayiSALIX164-08 07-DPQC-0052 Salix barclayiSALIX171-08 07-DPQC-0059 Salix barclayiSALIX194-08 07-UBCBG-037155-1003-2004 Salix barclayiSALIX199-08 07-UBCBG-native3 Salix barclayiSALIX227-08 07-BAB-025 Salix barclayiSALIX238-08 07-BAB-082 Salix barclayiSALIX260-08 07-BAB-227 Salix barclayiSALIX274-08 07-BAB-272 Salix barclayiSALIX279-08 07-BAB-287 Salix barclayiSALIX287-08 07-BAB-313 Salix barclayiSALIX404-08 07-BAB-024 Salix barclayiSALIX443-08 08-DPQC-0095A Salix barclayiSALIX444-08 08-DPQC-0095B Salix barclayiSALIX454-08 08-DPQC-0107 Salix barclayiSALIX455-08 08-DPQC-0108 Salix barclayiSALIX458-08 08-DPQC-0112 Salix barclayiSALIX139-08 07-DPQC-0019 Salix barrattianaSALIX173-08 07-DPQC-0061 Salix barrattianaSALIX246-08 07-BAB-168 Salix barrattianaSALIX381-08 43-UBC-65549 Salix barrattianaSALIX382-08 73-UBC-149084 Salix barrattianaSALIX522-08 08-GWA-14437A Salix barrattianaSALIX004-08 07-JMS-1056 Salix bebbianaSALIX005-08 07-JMS-1059 Salix bebbianaSALIX008-08 07-JMS-1094 Salix bebbianaSALIX066-08 07-JMS-1365 Salix bebbianaSALIX113-08 07-EM-0315 Salix bebbianaSALIX115-08 07-EM-0472 Salix bebbianaSALIX118-08 07-EM-0583 Salix bebbianaSALIX121-08 07-EM-0628 Salix bebbianaSALIX134-08 07-DPQC-0014 Salix bebbianaSALIX198-08 07-UBCBG-037023-1003-2003 Salix bebbianaSALIX200-08 07-UBCBG-34916-5555-99 Salix bebbianaSALIX221-08 07-BAB-014 Salix bebbianaSALIX248-08 07-BAB-193 Salix bebbianaSALIX253-08 07-BAB-200 Salix bebbianaSALIX275-08 07-BAB-273 Salix bebbianaSALIX348-08 07-GWA-14380 Salix bebbianaSALIX417-08 08-DPQC-0069 Salix bebbianaSALIX419-08 08-DPQC-0071 Salix bebbianaSALIX428-08 08-DPQC-0080 Salix bebbianaSALIX432-08 08-DPQC-0084 Salix bebbianaSALIX463-08 08-DPQC-0118 Salix bebbianaSALIX472-08 08-DPQC-0127 Salix bebbianaSALIX513-08 08-CEG-15 Salix bebbianaSALIX514-08 08-CEG-16 Salix bebbianaSALIX065-08 07-JMS-1364 Salix boothiiSALIX293-08 07-CEG-19 Salix boothiiSALIX360-08 07-GWA-14395 Salix boothiiSALIX515-08 08-CEG-17 Salix boothiiSALIX516-08 08-GWA-14433 Salix boothiiSALIX526-08 08-GWA-14443 Salix brachycarpaSALIX527-08 08-GWA-14444 Salix brachycarpaSALIX533-08 08-GWA-14450 Salix calicolaSALIX395-08 08-ISQC-CanIs Salix canariensisSALIX120-08 07-EM-0618 Salix candidaSALIX243-08 07-BAB-130 Salix candidaSALIX259-08 07-BAB-226 Salix candidaSALIX312-08 07-GWA-14337 Salix candidaSALIX313-08 07-GWA-14338 Salix candidaSALIX315-08 07-GWA-14340 Salix candidaSALIX485-08 08-DPQC-0141 Salix candidaSALIX569-09 09-DPQC-11 Salix candidaSALIX573-09 09-DPQC-26 Salix candidaSALIX143-08 07-DPQC-0027 Salix commutataSALIX161-08 07-DPQC-0049 Salix commutataSALIX529-08 08-GWA-14446 Salix commutataSALIX342-08 07-GWA-14371 Salix cordataSALIX400-08 07-DW-6000 Salix cordataSALIX402-08 07-DW-6002 Salix cordataSALIX054-08 07-JMS-1348 Salix drummondianaSALIX059-08 07-JMS-1353 Salix drummondianaSALIX064-08 07-JMS-1363 Salix drummondianaSALIX135-08 07-DPQC-0015 Salix drummondianaSALIX276-08 07-BAB-274 Salix drummondianaSALIX430-08 08-DPQC-0082 Salix drummondianaSALIX435-08 08-DPQC-0087 Salix drummondianaSALIX457-08 08-DPQC-0111 Salix drummondianaSALIX466-08 08-DPQC-0121 Salix drummondianaSALIX470-08 08-DPQC-0125 Salix drummondianaSALIX478-08 08-DPQC-0133 Salix drummondianaSALIX520-08 08-GWA-14437 1 Salix drummondianaSALIX521-08 08-GWA-14437 2 Salix drummondianaSALIX528-08 08-GWA-14445 Salix drummondianaSALIX530-08 08-GWA-14447 Salix drummondianaSALIX574-09 07-GWA-14436B Salix drummondianaSALIX375-08 07-GWA-14431 Salix eastwoodiaeSALIX191-08 07-UBCBG-013854-0013-1976 Salix elaeagnosSALIX219-08 07-CAN-532085 Salix eriocephalaSALIX334-08 07-GWA-14363 Salix eriocephalaSALIX309-08 07-GWA-14334 Salix famelicaSALIX310-08 07-GWA-14335 Salix famelicaSALIX524-08 08-GWA-14439 Salix farriaeSALIX570-09 09-DPQC-13 Salix farriaeSALIX109-08 07-JMS-0974 Salix geyerianaSALIX168-08 07-DPQC-0056 Salix glaucaSALIX174-08 07-DPQC-0062 Salix glaucaSALIX224-08 07-BAB-017 Salix glaucaSALIX252-08 07-BAB-199 Salix glaucaSALIX286-08 07-BAB-299 Salix glaucaSALIX362-08 07-GWA-14397 Salix glaucaSALIX374-08 07-GWA-14430 Salix glaucaSALIX388-08 46-UBC-61323 Salix glaucaSALIX389-08 47-UBC-60831 Salix glaucaSALIX441-08 08-DPQC-0093 Salix glaucaSALIX471-08 08-DPQC-0126 Salix glaucaSALIX486-08 08-DPQC-0142 Salix glaucaSALIX487-08 08-DPQC-0143 Salix glaucaSALIX459-08 08-DPQC-0113 Salix glauca villosaSALIX460-08 08-DPQC-0114 Salix glauca villosaSALIX465-08 08-DPQC-0120 Salix glauca villosaSALIX531-08 08-GWA-14448 Salix glauca villosaSALIX386-08 56-UBC-58897 Salix glauca x brachycarpaSALIX024-08 07-JMS-1297 Salix hookerianaSALIX069-08 07-JMS-0865 Salix hookerianaSALIX074-08 07-JMS-0885 Salix hookerianaSALIX078-08 07-JMS-0900 Salix hookerianaSALIX098-08 07-JMS-0956 Salix hookerianaSALIX100-08 07-JMS-0958 Salix hookerianaSALIX102-08 07-JMS-0960 Salix hookerianaSALIX103-08 07-JMS-0961 Salix hookerianaSALIX189-08 07-DPQC-UBCFarm5 Salix hookerianaSALIX192-08 07-UBCBG-037096-1003-2004 Salix hookerianaSALIX341-08 07-GWA-14370 Salix hookerianaSALIX399-08 08-DPsitchSB Salix hookerianaSALIX290-08 07-CEG-15 Salix irrorataSALIX291-08 07-CEG-16 Salix irrorataSALIX364-08 07-GWA-14411 Salix jepsoniiSALIX365-08 07-GWA-14413 Salix jepsoniiSALIX370-08 07-GWA-14426 Salix jepsoniiSALIX292-08 07-CEG-18 Salix lasiolepisSALIX353-08 07-GWA-14385 Salix lemmoniiSALIX354-08 07-GWA-14386 Salix lemmoniiSALIX288-08 07-CEG-13 Salix ligulifoliaSALIX289-08 07-CEG-14 Salix ligulifoliaSALIX500-08 08-CEG-03A Salix ligulifoliaSALIX501-08 08-CEG-03B Salix ligulifoliaSALIX396-08 08-DPSlind Salix lindleyanaSALIX350-08 07-GWA-14382 Salix luteaSALIX352-08 07-GWA-14384 Salix luteaSALIX479-08 08-DPQC-0134 Salix maccallianaSALIX483-08 08-DPQC-0139 Salix maccallianaSALIX484-08 08-DPQC-0140 Salix maccallianaSALIX488-08 08-DPQC-0144 Salix maccallianaSALIX489-08 08-DPQC-0145 Salix maccallianaSALIX535-08 08-GWA-14452 Salix maccallianaSALIX195-08 07-UBCBG-023877-0368-1984 Salix magnificaSALIX300-08 07-CEG-29 Salix monticolaSALIX512-08 08-CEG-14 Salix monticolaSALIX403-08 07-DW-6003 Salix myricoidesSALIX223-08 07-BAB-016 Salix myrtillifoliaSALIX277-08 07-BAB-276 Salix myrtillifoliaSALIX377-08 07-WAGNER-MYR1 Salix myrtillifoliaSALIX196-08 07-UBCBG-26352-117-87 Salix nakamuranaSALIX236-08 07-BAB-051 Salix niphocladaSALIX268-08 07-BAB-250 Salix niphocladaSALIX138-08 07-DPQC-0018 Salix nivalisSALIX149-08 07-DPQC-0033 Salix nivalisSALIX157-08 07-DPQC-0041 Salix nivalisSALIX158-08 07-DPQC-0042 Salix nivalisSALIX523-08 08-GWA-14438 Salix nivalisSALIX036-08 07-JMS-1320 Salix pentandraSALIX058-08 07-JMS-1352 Salix planifoliaSALIX249-08 07-BAB-194 Salix planifoliaSALIX366-08 07-GWA-14416 Salix planifoliaSALIX367-08 07-GWA-14417 Salix planifoliaSALIX422-08 08-DPQC-0074 Salix planifoliaSALIX461-08 08-DPQC-0115 Salix planifoliaSALIX285-08 07-BAB-298 Salix polarisSALIX148-08 07-DPQC-0032 Salix prolixaSALIX421-08 08-DPQC-0073 Salix prolixaSALIX427-08 08-DPQC-0079 Salix prolixaSALIX429-08 08-DPQC-0081 Salix prolixaSALIX439-08 08-DPQC-0091 Salix prolixaSALIX450-08 08-DPQC-0103 Salix prolixaSALIX452-08 08-DPQC-0105 Salix prolixaSALIX469-08 08-DPQC-0124 Salix prolixaSALIX475-08 08-DPQC-0130 Salix prolixaSALIX491-08 08-DPQC-0148 Salix prolixaSALIX492-08 08-DPQC-0149 Salix prolixaSALIX117-08 07-EM-0579 Salix pseudomonticolaSALIX267-08 07-BAB-247 Salix pseudomonticolaSALIX314-08 07-GWA-14339 Salix pseudomonticolaSALIX316-08 07-GWA-14342 Salix pseudomonticolaSALIX468-08 08-DPQC-0123 Salix pseudomonticolaSALIX474-08 08-DPQC-0129 Salix pseudomonticolaSALIX482-08 08-DPQC-0137 Salix pseudomonticolaSALIX116-08 07-EM-0475 Salix pseudomyrsinitesSALIX245-08 07-BAB-167 Salix pseudomyrsinitesSALIX278-08 07-BAB-286 Salix pseudomyrsinitesSALIX378-08 07-WAGNER-PS1 Salix pseudomyrsinitesSALIX379-08 07-WAGNER-PS2 Salix pseudomyrsinitesSALIX405-08 07-BAB-344 Salix pseudomyrsinitesSALIX225-08 07-BAB-018 Salix pulchraSALIX281-08 07-BAB-291 Salix pulchraSALIX347-08 07-GWA-14379 Salix pyrifoliaSALIX418-08 08-DPQC-0070 Salix pyrifoliaSALIX536-09 08-CAR-001 Salix reticulataSALIX537-09 08-CAR-002 Salix reticulataSALIX538-09 08-CAR-003 Salix reticulataSALIX539-09 08-CAR-004 Salix reticulataSALIX540-09 08-CAR-005 Salix reticulataSALIX541-09 08-CAR-006 Salix reticulataSALIX542-09 08-CAR-007 Salix reticulataSALIX044-08 07-JMS-1338 Salix reticulataSALIX045-08 07-JMS-1339 Salix reticulataSALIX046-08 07-JMS-1340 Salix reticulataSALIX233-08 07-BAB-039 Salix reticulataSALIX244-08 07-BAB-165 Salix reticulataSALIX543-09 08-CAR-008 Salix reticulataSALIX544-09 08-CAR-009 Salix reticulataSALIX545-09 08-CAR-010 Salix reticulataSALIX546-09 08-CAR-011 Salix reticulataSALIX547-09 08-CAR-012 Salix reticulataSALIX550-09 08-CAR-015 Salix reticulataSALIX551-09 08-CAR-016 Salix reticulataSALIX552-09 08-CAR-017 Salix reticulataSALIX553-09 08-CAR-018 Salix reticulataSALIX555-09 08-CAR-020 Salix reticulataSALIX556-09 08-CAR-021 Salix reticulataSALIX559-09 08-CAR-024 Salix reticulataSALIX560-09 08-CAR-025 Salix reticulataSALIX561-09 08-CAR-026 Salix reticulataSALIX564-09 08-CAR-029 Salix reticulataSALIX565-09 08-CAR-030 Salix reticulataSALIX566-09 08-CAR-031 Salix reticulataSALIX568-09 08-CAR-033 Salix reticulataSALIX228-08 07-BAB-029 Salix richardsoniiSALIX255-08 07-BAB-209 Salix richardsoniiSALIX284-08 07-BAB-294 Salix richardsoniiSALIX242-08 07-BAB-108 Salix rotundifoliaSALIX001-08 07-JMS-1037 Salix scoulerianaSALIX002-08 07-JMS-1038 Salix scoulerianaSALIX003-08 07-JMS-1050 Salix scoulerianaSALIX006-08 07-JMS-1081 Salix scoulerianaSALIX014-08 07-JMS-1141 Salix scoulerianaSALIX015-08 07-JMS-1142 Salix scoulerianaSALIX017-08 07-JMS-1211 Salix scoulerianaSALIX025-08 07-JMS-1298 Salix scoulerianaSALIX026-08 07-JMS-1299 Salix scoulerianaSALIX033-08 07-JMS-1316 Salix scoulerianaSALIX034-08 07-JMS-1317 Salix scoulerianaSALIX052-08 07-JMS-1346 Salix scoulerianaSALIX062-08 07-JMS-1361 Salix scoulerianaSALIX085-08 07-JMS-0926 Salix scoulerianaSALIX086-08 07-JMS-0927 Salix scoulerianaSALIX128-08 07-DPQC-0008 Salix scoulerianaSALIX130-08 07-DPQC-0010 Salix scoulerianaSALIX133-08 07-DPQC-0013 Salix scoulerianaSALIX145-08 07-DPQC-0029 Salix scoulerianaSALIX146-08 07-DPQC-0030 Salix scoulerianaSALIX147-08 07-DPQC-0031 Salix scoulerianaSALIX257-08 07-BAB-211 Salix scoulerianaSALIX280-08 07-BAB-288 Salix scoulerianaSALIX358-08 07-GWA-14392 Salix scoulerianaSALIX502-08 08-CEG-04 Salix scoulerianaSALIX503-08 08-CEG-05 Salix scoulerianaSALIX009-08 07-JMS-1119 Salix sitchensisSALIX010-08 07-JMS-1123 Salix sitchensisSALIX012-08 07-JMS-1132 Salix sitchensisSALIX018-08 07-JMS-1212 Salix sitchensisSALIX021-08 07-JMS-1259 Salix sitchensisSALIX022-08 07-JMS-1260 Salix sitchensisSALIX023-08 07-JMS-1281 Salix sitchensisSALIX029-08 07-JMS-1310 Salix sitchensisSALIX030-08 07-JMS-1313 Salix sitchensisSALIX032-08 07-JMS-1315 Salix sitchensisSALIX035-08 07-JMS-1319 Salix sitchensisSALIX051-08 07-JMS-1345 Salix sitchensisSALIX068-08 07-JMS-0864 Salix sitchensisSALIX072-08 07-JMS-0878 Salix sitchensisSALIX073-08 07-JMS-0884 Salix sitchensisSALIX075-08 07-JMS-0891 Salix sitchensisSALIX079-08 07-JMS-0901 Salix sitchensisSALIX083-08 07-JMS-0924 Salix sitchensisSALIX084-08 07-JMS-0925 Salix sitchensisSALIX089-08 07-JMS-0941 Salix sitchensisSALIX092-08 07-JMS-0948 Salix sitchensisSALIX093-08 07-JMS-0949 Salix sitchensisSALIX094-08 07-JMS-0952 Salix sitchensisSALIX095-08 07-JMS-0953 Salix sitchensisSALIX097-08 07-JMS-0955 Salix sitchensisSALIX099-08 07-JMS-0957 Salix sitchensisSALIX101-08 07-JMS-0959 Salix sitchensisSALIX104-08 07-JMS-0962 Salix sitchensisSALIX105-08 07-JMS-0965 Salix sitchensisSALIX110-08 07-JMS-0977 Salix sitchensisSALIX124-08 07-DPQC-0001A Salix sitchensisSALIX132-08 07-DPQC-0012 Salix sitchensisSALIX136-08 07-DPQC-0016 Salix sitchensisSALIX166-08 07-DPQC-0054 Salix sitchensisSALIX172-08 07-DPQC-0060 Salix sitchensisSALIX182-08 07-DPQC-MINavChan Salix sitchensisSALIX186-08 07-DPQC-UBCFarm2 Salix sitchensisSALIX202-08 07-DPQC-VanIs1 Salix sitchensisSALIX203-08 07-DPQC-VanIs2 Salix sitchensisSALIX206-08 07-DPQC-WhistRd2 Salix sitchensisSALIX209-08 07-DPQC-WhistRd5 Salix sitchensisSALIX210-08 07-DPQC-WhistRd6 Salix sitchensisSALIX408-08 07-JMS-0895 Salix sitchensisSALIX437-08 08-DPQC-0089 Salix sitchensisSALIX448-08 08-DPQC-0101 Salix sitchensisSALIX451-08 08-DPQC-0104 Salix sitchensisSALIX456-08 08-DPQC-0110 Salix sitchensisSALIX473-08 08-DPQC-0128 Salix sitchensisSALIX205-08 07-DPQC-WhistRd1 Salix sp.SALIX207-08 07-DPQC-WhistRd3 Salix sp.SALIX208-08 07-DPQC-WhistRd4 Salix sp.SALIX413-08 08-DPQC-0065 Salix sp.SALIX420-08 08-DPQC-0072 Salix sp.SALIX571-09 09-DPQC-17 Salix sp.SALIX572-09 09-DPQC-25 Salix sp.SALIX184-08 07-PMP-20885 Salix uvaursiSALIX518-08 08-GWA-14435 Salix vestitaSALIX519-08 08-GWA-14436 Salix vestitaSALIX204-08 07-DPQC-vimEur ope Salix viminalisSALIX020-08 07-JMS-1254 Salix x rubens SALIX183-08 07-PMP-20866 Salix exiguaSALIX112-08 07-JMS-0981 Salix interiorSALIX254-08 07-BAB-201 Salix interiorSALIX265-08 07-BAB-242 Salix interiorSALIX317-08 07-GWA-14343 Salix interiorSALIX331-08 07-GWA-14360 Salix interiorSALIX336-08 07-GWA-14365 Salix interiorSALIX337-08 07-GWA-14366 Salix interiorSALIX338-08 07-GWA-14367 Salix interiorSALIX525-08 08-GWA-14440 Salix interiorSALIX212-08 07-CAN-483931 Salix interior x eriocephalaSALIX217-08 07-CAN-532065 Salix interior x eriocephalaSALIX216-08 07-CAN-532063 Salix interior x petiolarisSALIX398-08 08-DPweepSB Salix sepulcralisSALIX081-08 07-JMS-0912 Salix x sepulcralisSALIX176-08 07-DPQC-weepJerBeach Salix x sepulcralisSALIX201-08 07-DMP-UniGuelph Salix albaSALIX016-08 07-JMS-1206 Salix x rubensSALIX327-08 07-GWA-14353 Salix serissimaSALIX328-08 07-GWA-14354 Salix serissimaSALIX397-08 08-DPsmallSB Salix sepulcralisSALIX027-08 07-JMS-1303 Salix x rubensSALIX019-08 07-JMS-1213 Salix x sepulcralisSALIX320-08 07-GWA-14346 Salix amygdaloidesSALIX332-08 07-GWA-14361 Salix amygdaloidesSALIX506-08 08-CEG-08 Salix amygdaloides x goodingiiSALIX507-08 08-CEG-09 Salix amygdaloides x goodingiiSALIX340-08 07-GWA-14369 Salix candidaSALIX343-08 07-GWA-14372 Salix cordataSALIX344-08 07-GWA-14375 Salix cordataSALIX114-08 07-EM-0359 Salix discolorSALIX319-08 07-GWA-14345 Salix discolorSALIX321-08 07-GWA-14347 Salix discolorSALIX329-08 07-GWA-14355 Salix discolorSALIX339-08 07-GWA-14368 Salix discolorSALIX431-08 08-DPQC-0083 Salix discolorSALIX345-08 07-GWA-14376 Salix eriocephalaSALIX346-08 07-GWA-14377 Salix eriocephalaSALIX127-08 07-DPQC-0003 Salix exiguaSALIX349-08 07-GWA-14381 Salix exiguaSALIX373-08 07-GWA-14429 Salix exiguaSALIX490-08 08-DPQC-0147 Salix exiguaSALIX499-08 08-CEG-02 Salix exiguaSALIX504-08 08-CEG-06 Salix exiguaSALIX508-08 08-CEG-10 Salix gooddingiiSALIX509-08 08-CEG-11 Salix gooddingiiSALIX330-08 07-GWA-14356 Salix humilisSALIX335-08 07-GWA-14364 Salix humilisSALIX214-08 07-CAN-483950 Salix interior x petiolarisSALIX294-08 07-CEG-23 Salix laevigataSALIX295-08 07-CEG-24 Salix laevigataSALIX324-08 07-GWA-14350 Salix lucidaSALIX326-08 07-GWA-14352 Salix lucidaSALIX333-08 07-GWA-14362 Salix nigraSALIX213-08 07-CAN-483942 Salix petiolarisSALIX318-08 07-GWA-14344 Salix petiolarisSALIX322-08 07-GWA-14348 Salix petiolarisSALIX325-08 07-GWA-14351 Salix petiolarisSALIX433-08 08-DPQC-0085 Salix planifoliaSALIX410-08 07-PMP-21007 Salix sp.SALIX411-08 07-PMP-21162 Salix sp.SALIX412-08 07-PMP-21311 Salix sp.0.05 changes
G I II III IV V VII - 3 8 7 7 9II - 11 10 10 12III - 1 1 3IV - 0 2V - 2VI -
matK 28 PIC
I
II
I
VI IIIIV+V
IVIV
0
© 2014 John Wiley & Sons Ltd
20 D. M. PERCY ET AL.