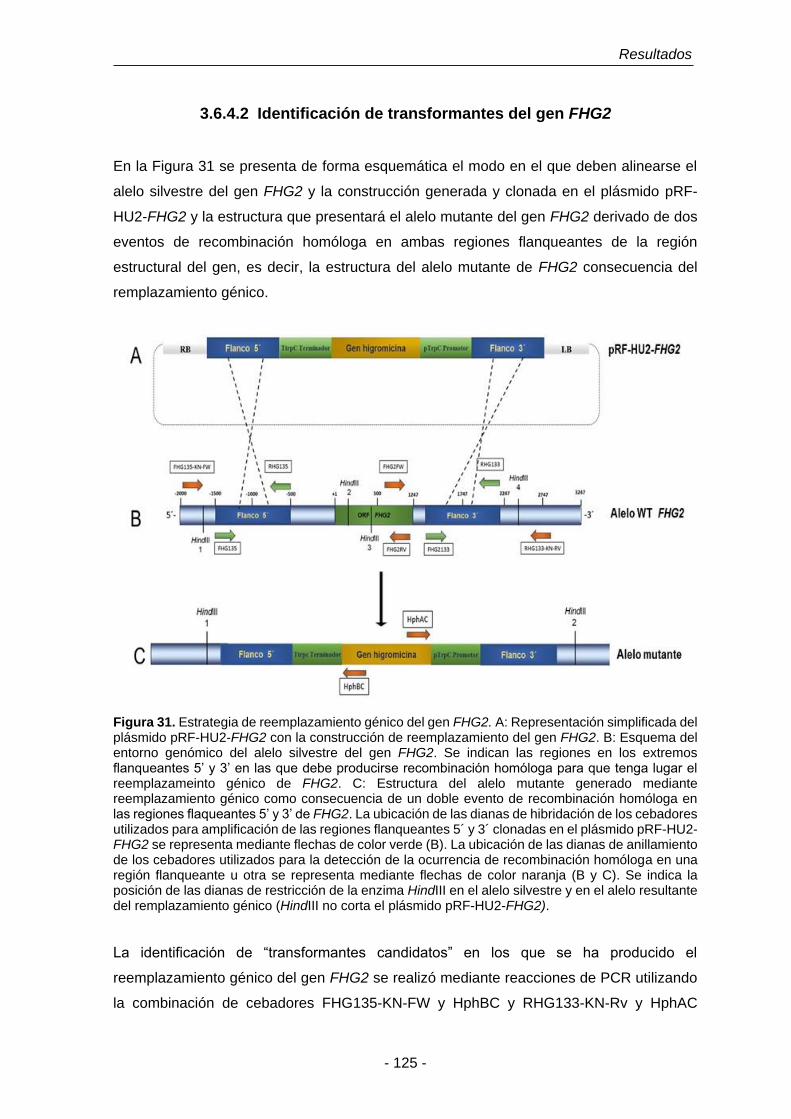
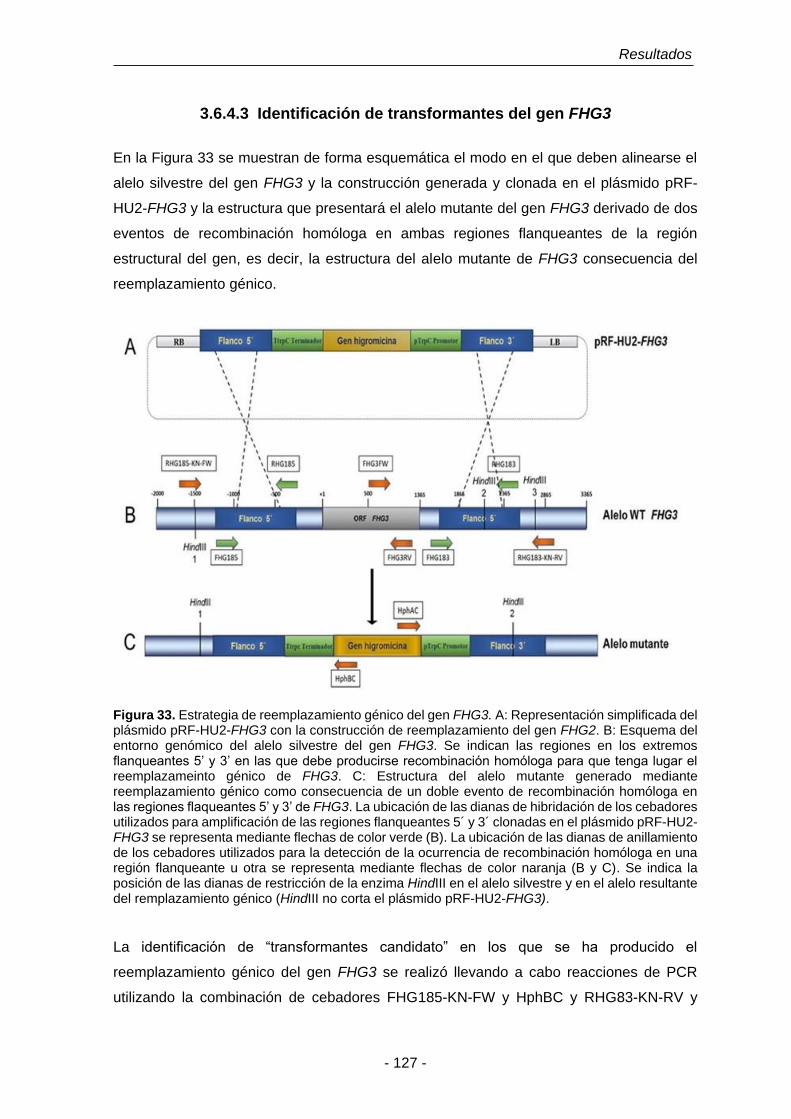
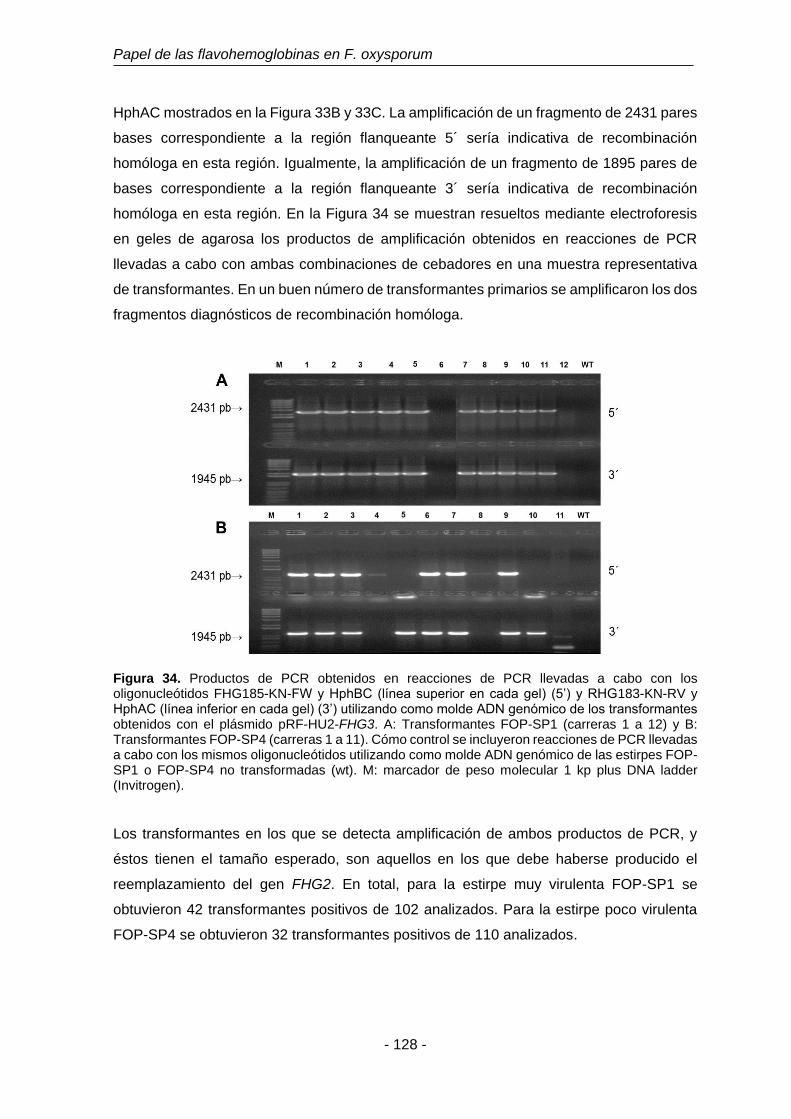
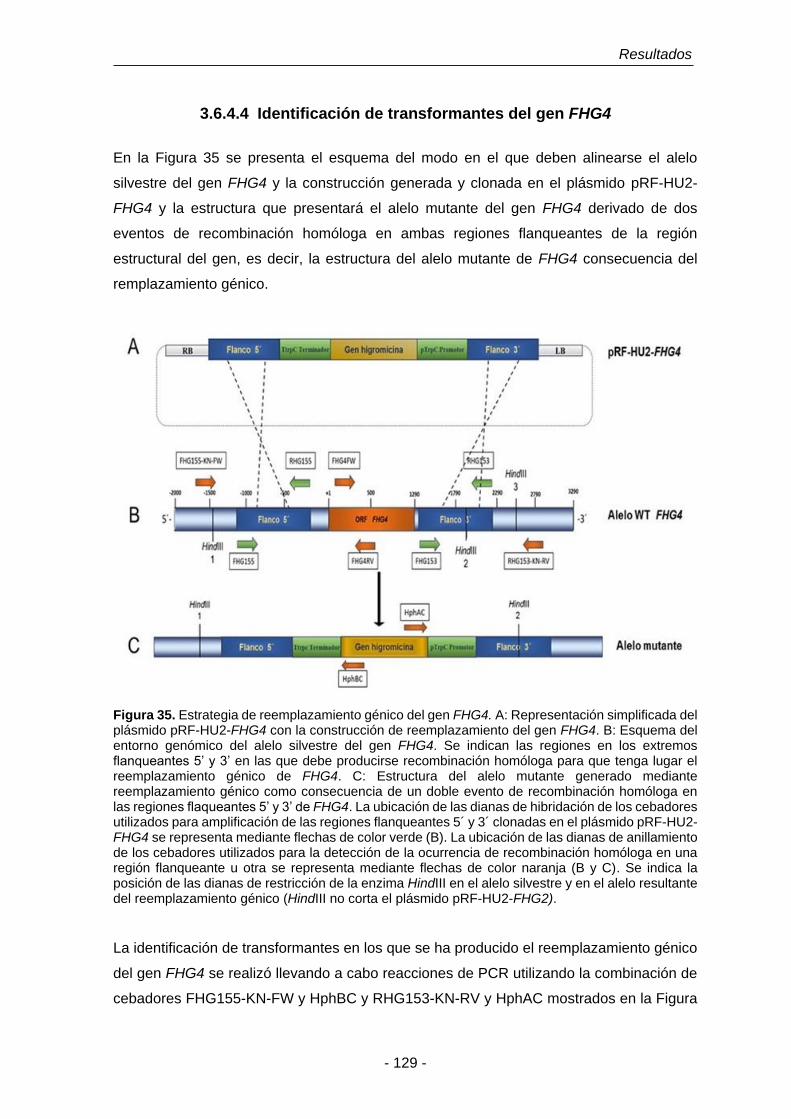
UNIVERSIDAD DE SALAMANCA DEPARTAMENTO DE MICROBIOLOGÍA Y GENÉTICA ÁREA DE GENÉTICA TESIS DOCTORAL Funciones fisiológicas de la detoxificación del óxido nítrico en Fusarium oxysporum f. sp. phaseoli: El papel de las flavohemoglobinas Eduardo Enrique Argotti Valencia Salamanca, 2016
Transcript
UNIVERSIDAD DE SALAMANCA
DEPARTAMENTO DE MICROBIOLOGÍA Y GENÉTICA
ÁREA DE GENÉTICA
TESIS DOCTORAL
Funciones fisiológicas de la detoxificación del óxido nítrico en
Fusarium oxysporum f. sp. phaseoli:
El papel de las flavohemoglobinas
Eduardo Enrique Argotti Valencia
Salamanca, 2016
UNIVERSIDAD DE SALAMANCA
DEPARTAMENTO DE MICROBIOLOGÍA Y GENÉTICA
ÁREA DE GENÉTICA
Funciones fisiológicas de la detoxificación del óxido nítrico (NO)
en Fusarium oxysporum f. sp. phaseoli:
El papel de las flavohemoglobinas
Tesis Doctoral
Programa de Doctorado: Agrobiotecnología
Órgano responsable del Programa de Doctorado:
Departamento de Microbiología y Genética
Eduardo Enrique Argotti Valencia
Salamanca, 2016
D. Luis Román Fernández Lago, Director del Departamento de Microbiología y Genética de la Facultad de Biología de la Universidad de Salamanca.
CERTIFICO: Que la presente Memoria titulada “Funciones fisiológicas de la detoxificación
del óxido nítrico en Fusarium oxysporum f. sp. phaseoli: El papel de las
flavohemoglobinas”, ha sido realizada en el Área de Genética del Departamento
de Microbiología y Genética y en el Instituto Hispano-Luso de Investigaciones
Agrarias de la Universidad de Salamanca por D. Eduardo Enrique Argotti
Valencia, bajo la dirección de los doctores Prof. Dr. Ernesto Pérez Benito y Prof.
Dr. José María Díaz Mínguez y cumple las condiciones exigidas para optar al grado
de Doctor por la Universidad de Salamanca.
Para que así conste, firmo el presente certificado en Salamanca a 12 de abril de
2016.
Fdo: D. Luis Román Fernández Lago
Dr. D. Ernesto Pérez Benito y Dr. D. José María Díaz Mínguez, Profesores Titulares del Área de Genética del Departamento de Microbiología y Genética de la Universidad de Salamanca
CERTIFICAMOS: Que la presente Memoria titulada “Funciones fisiológicas de la detoxificación
del óxido nítrico en Fusarium oxysporum f. sp. phaseoli: El papel de las
flavohemoglobinas”, ha sido realizada en el Área de Genética del Departamento
de Microbiología y Genética y en el Instituto Hispano-Luso de Investigaciones
Agrarias de la Universidad de Salamanca, bajo nuestra dirección, por D. Eduardo
Enrique Argotti Valencia, y cumple las condiciones exigidas para optar al grado
de Doctor por la Universidad de Salamanca.
Y para que así conste, extendemos el presente certificado en Salamanca a 12 de
abril de 2016.
Fdo: Dr. D. Ernesto Pérez Benito Fdo: Dr. D. José María Díaz Mínguez
Este trabajo se ha llevado a cabo en el Laboratorio 1 del Instituto Hispano-Luso de Investigaciones Agrarias (CIALE), Departamento de Microbiología y Genética, de la Universidad de Salamanca, bajo la dirección de los Profesores Dr. Ernesto Pérez Benito y Dr. José María Díaz Mínguez. Su ejecución ha sido posible gracias a la financiación de los proyectos AGL2012-39876-C02 y AGL2015-66131-C2-1-R (Ministerio de Economía y Competitividad). Durante el desarrollo de la Tesis, he disfrutado de una beca de la Secretaría Nacional de Educación Superior, Ciencia, Tecnología e Innovación del Ecuador
(SENESCYT).
AGRADECIMIENTOS
La presente Tesis Doctoral ha requerido un gran esfuerzo y una intensa dedicación sin la
cual no hubiese sido posible su elaboración y la cooperación de todas y cada una de las
personas e instituciones que a continuación citaré y muchas de las cuales han sido un
soporte muy fuerte en los momentos más duros y difíciles.
En primer lugar quiero agradecer al Gobierno del Ecuador, a la Secretaria de Educación
Superior, Ciencia, Tecnología e Innovación (SENESCYT), por concederme esta beca para
cursar estudios de Doctorado y hacer realidad este sueño que parecía inalcanzable.
A mis Tutores, Dr. Ernesto Pérez Benito y Dr. José María Díaz Mínguez, por haberme
aceptado en su grupo de investigación y haber confiado en mí, por brindarme su constante
apoyo y ayuda en los momentos que más he necesitado; sin su implicación, orientación y
supervisión en el día a día, así como en la redacción del presente manuscrito, esta
investigación no hubiese tenido lugar.
A todos los que son y han sido mis compañeros de laboratorio, con los que tantos días he
compartido: Walter Vargas, Vega Tello, Daniela Santander, Virginia Casado, Jonathan
Niño, José María Sanz, Raúl Martin y Wilson Acosta.
A mis suegros por su constante apoyo para que no desmaye durante estos cuatro años de
intenso trabajo y dedicación.
A mí madre por su constante apoyo a la distancia y siempre convencida de que tengo la
suficiente capacidad para llegar a donde yo quiera y vencer todos los obstáculos que se
presentan a diario.
A mí querida esposa, mi compañera, mi apoyo, mi fortaleza, cómplice de mis triunfos y que
jamás se ha opuesto a mis caprichos y objetivos que me he propuesto.
A mis hijos Kevin y Karen la razón de mi vida.
Eduardo Enrique Argotti Valencia
A mí Dios.
Por su infinito amor
ÍNDICES
Índice
I
ÍNDICE
ÍNDICES
3.5. ÍNDICE I
3.6. ÍNDICE DE FIGURAS IX
3.7. ÍNDICE DE TABLAS XV
3.8. SÍMBOLOS Y ABREVIATURAS XVII
RESUMEN XIX
1 INTRODUCCIÓN 1
1.1 Hongos fitopatógenos. 1
1.2 El género Fusarium. 1
1.2.1 Morfología. 2
1.2.2 Taxonomía del género Fusarium.
2
1.2.3 Especies anamórficas y teleomórficas del género Fusarium.
4
1.3. 1.1.1.1.1 El complejo de especies Fusarium oxysporum.
5
1.3.1 Descripción y características taxonómicas.
5
1.3.1.1 Morfología. 7
1.3.1.2 Ciclo de vida. 7
1.3.1.3 Capacidad infectiva: la fusariosis vascular
8
1.3.1.4. Genes esenciales en patogenicidad y virulencia de
F. oxysporum.
10
1.4 Genómica de F. oxysporum: genoma nuclear y genoma adaptativo,
expansiones génicas.
13
1.5 F. oxysporum f. sp. phaseoli.
14
1.5.1 Sintomatología y clasificación.
15
1.5.2 La importancia del cultivo de la judía.
15
1.6 El óxido nítrico.
17
1.6.1 Descripción
17
1.6.2 Funciones fisiológicas del NO.
18
1.6.2.1 Síntesis de NO en animales, plantas y hongos.
19
1.6.2.2 Óxido nítrico en la interacción planta-patógeno.
21
1.6.3 Fuentes de nitrógeno utilizadas por los hongos.
22
1.6.4 La denitrificación del nitrato en procariotas. 24
Papel de las flavohemoglobinas en F. oxysporum
II
1.6.5 Desnitrificación y producción de NO en F. oxysporum.
25
1.7 Las flavohemoglobinas.
26
1.7.1 Origen de las flavohemoglobinas.
27
1.7.2 Estructura de las flavohemoglobinas 27
1.7.3 Funciones de las flavohemoglobinas.
28
1.7.3.1 Respuesta de las flavohemoglobinas al estrés
nitrosativo y oxidativo.
28
1.7.3.2 Papel de las flavohemoglobinas en patogenicidad. 31
1.7.3.3 Papel de las flavohemoglobinas en la regulación del
desarrollo.
32
OBJETIVOS 35
2 MATERIALES Y MÉTODOS 37
2.1 Organismos. 37
2.1.1 Plantas. 37
2.1.2 Hongos. 37
2.1.3 Bacterias. 38
2.2 Medios de cultivo. 39
2.2.1 Plantas. 39
2.2.2 Hongos. 39
2.2.2.1 Cultivo en medio sólido de F. oxysporum. 39
2.2.2.2 Cultivo en medio líquido de F. oxysporum. 40
2.2.2.3 Obtención de cultivos monospóricos. 40
2.2.3 Bacterias 41
2.2.3.1 Cultivo de E. coli DHα5. 41
2.2.3.2 Cultivo de A. tumefaciens cepa AGL-1. 41
2.3 Ensayos de germinación. 42
2.3.1 Obtención de microconidios de F. oxysporum. 42
2.3.2 Cultivo estático de F. oxysporum. 43
2.4 Ensayos de crecimiento saprofítico. 43
2.4.1 Ensayos de sensibilidad a estrés nitrosativo. 43
2.5 Ensayos de infección in planta. 44
2.5.1 Ensayos de infección en invernadero. 44
2.5.2 Ensayos de infección en medio de cultivo hidropónico. 45
2.6 Extracción de ácidos nucleicos. 45
Índice
III
2.6.1 ADN genómico de F. oxysporum. 45
2.6.1.1 1.1.1.1.1.1 Método “TENSP”. 46
2.6.1.2 1.1.1.1.1.2 Método CTAB. 47
2.6.1.3 Extracción de ADN genómico mediante método
rápido.
47
2.6.2 Extracción de ADN plasmídico. 48
2.6.2.1 Minipreparación de ADN plasmídico mediante lisis
alcalina.
48
2.6.2.2 Minipreparación de ADN plasmídico con el kit
comercial NucleoSpin®Plasmid.
48
2.6.2.3 Preparación de grandes cantidades de ADN de
plásmidos de reducido número de copia.
49
2.6.3 Tratamientos enzimáticos del ADN. 50
2.6.3.1 Tratamiento con RNAsa A. 50
2.6.3.2 Digestión con enzimas de restricción. 51
2.6.3.3 Ligaciones. 51
2.6.3.4 Defosforilación con fosfatasa alcalina (CIEF). 51
2.6.4 Obtención de ARN total de F. oxysporum. 52
2.6.4.1 Método Trizol. 52
2.6.4.2 Obtención de ARN con el kit comercial SV Total
RNA Isolation System (Promega).
53
2.6.4.3 Tratamiento de ARN con DNAasa. 54
2.6.4.4 Cuantificación de ácidos nucleicos. 54
2.7 Electroforesis de ácidos nucleicos. 54
2.7.1 Electroforesis de ADN. 54
2.7.2 Electroforesis de ARN. 55
2.7.3 Purificación de fragmentos de ADN de geles de agarosa. 55
2.8 Vectores de clonación. 56
2.8.1 Vector de clonación pGEM-T Easy. 56
2.8.2 Vector de clonación pRF-HU2. 56
2.8.3 Vector de clonación pEA-FU. 58
2.8.4 Vector de silenciamiento pVIR-RNAi. 59
2.9 Transformaciones. 60
2.9.1 Preparación de células competentes de E. coli. 60
Papel de las flavohemoglobinas en F. oxysporum
IV
2.9.2 Digestión de los vectores de clonación PRF-HU2, pVIR-RNAi
y pEA-FU.
61
2.9.3 Ensayos de clonación con el sistema “USER Friendly Cloning”. 62
2.9.3.1 Diseño de oligonucleótidos con un adaptador de
uridina.
62
2.9.3.2 Amplificación de las regiones flanqueantes. 62
2.9.3.3 Reacción de ligación con USER Friendly Cloning. 63
2.9.4 Transformación de E. coli. 63
2.9.5 Transformación de A. tumefaciens. 63
2.9.5.1 Preparación de células competentes. 63
2.9.5.2 Electroporación de A. tumefaciens. 64
2.9.5.3 Transformación de F. oxysporum mediada por A.
tumefaciens (ATM).
64
2.9.5.4 Ensayo de cocultivo de A. tumefaciens con F.
oxysporum.
65
2.9.5.5 Identificación de transformantes. 65
2.10
.
Construcción de mutantes dobles. 66
2.10.1 Silenciamiento genético. 66
2.11
.
Complementación de genes. 66
2.12
.
Reacción en cadena de la polimerasa (PCR). 67
2.12.1 PCR estándar. 67
2.12.2 RT-PCR. 68
2.12.3 PCR en Tiempo Real. 68
2.12.4 Secuenciación. 69
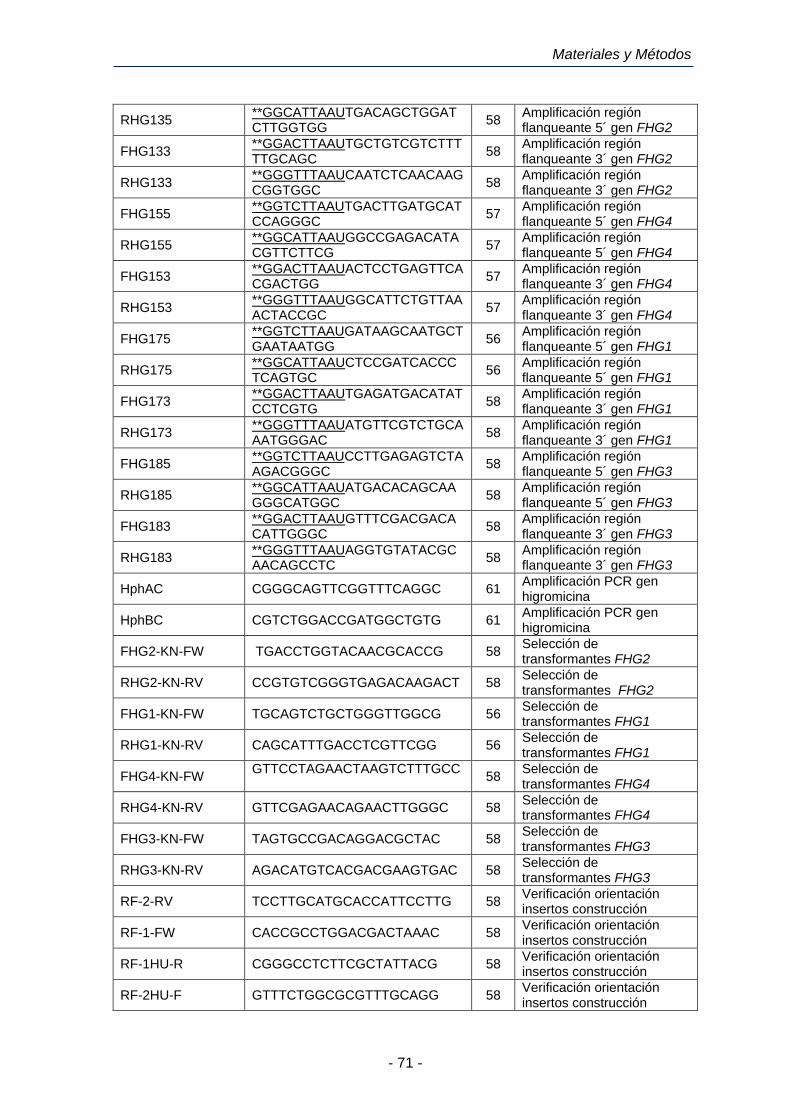
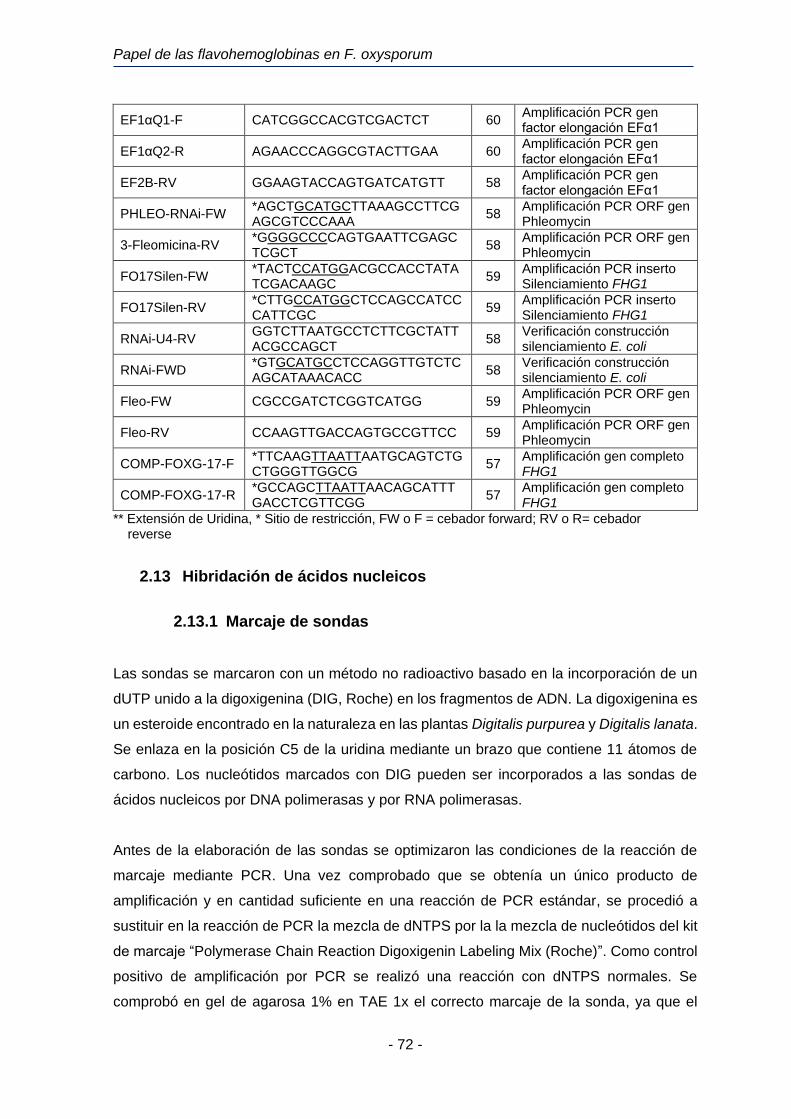
2.12.5 Oligonucleótidos utilizados en este trabajo. 70
2.13
.
Hibridación de ácidos nucleicos. 72
2.13.1 Marcaje de sondas. 72
2.13.2 Hibridación Southern blot. 73
2.13.3 Hibridación Northern blot. 75
2.14
.
Microscopía. 76
2.14.3 Microscopía óptica. 76
2.15
.
Análisis de datos. 76
2.15.1 Diseño de oligonucleótidos. 76
2.15.2 Diseño de plásmidos. 76
Índice
V
2.15.3 Alineamiento de secuencias. 76
2.15.4 Búsqueda y comparación de secuencias. 77
2.15.5 Diseño de árboles filogenéticos. 77
2.15.6 Búsqueda de secuencias de F. oxysporum. 77
2.15.7 Cuantificación de ADN y ARN. 77
2.15.8 Edición de texto y tablas. 77
2.15.9 Análisis estadístico. 78
3 RESULTADOS 79
3.1 Identificación de los genes codificadores de enzimas tipo
flavohemoglobina de F. oxysporum f. sp. phaseoli.
79
3.1.1 Análisis de las bases de datos de secuencias de genomas de
Fusarium.
79
3.1.2 Detección mediante PCR de genes codificadores de enzimas
de tipo flavohemoglobina de F. oxysporum f. sp. phaseoli.
80
3.1.3 Caracterización del entorno genómico de los genes
codificadores de flavohemoglobinas de F. oxysporum f. sp.
phaseoli.
83
3.2 Análisis estructural de los genes codificadores de flavohemoglobinas
de F. oxysporum f. sp. phaseoli.
85
3.2.1 Secuencia de los genes FHG de F. oxysporum f. sp. phaseoli. 86
3.2.1.1 Gen FHG1 de FOP-SP1. 86
3.2.1.2 Gen FHG2 de FOP-SP1. 87
3.2.1.3 Gen FHG3 de FOP-SP1. 88
3.2.1.4 Gen FHG4 de FOP-SP1. 89
3.2.2 Análisis de la región codificante de los genes FHG 90
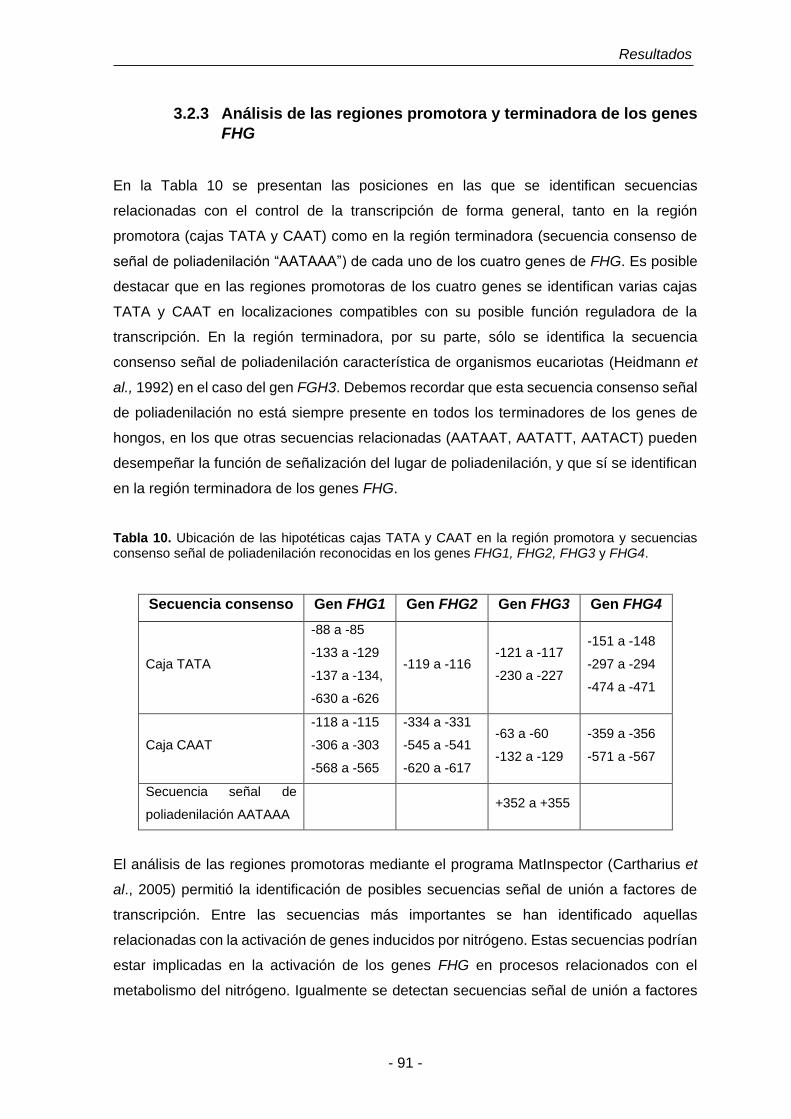
3.2.3 Análisis de las regiones promotora y terminadora de los genes
FHG.
91
3.3 Análisis de las proteínas deducidas codificadas por los genes FHG1,
FHG2, FHG3 y FHG4.
94
3.3.1 Dominios funcionales de las proteínas FHG. 94
3.3.2 Localización subcelular de las proteínas FHG. 99
3.4 Análisis filogenético de los genes codificadores de flavohemoglobinas. 101
3.5 Análisis de expresión de los genes FHG. 104
3.5.1 Condiciones para el análisis durante el crecimiento saprofítico
y en respuesta a NO.
104
Papel de las flavohemoglobinas en F. oxysporum
VI
3.5.2 Análisis de expresión de los genes FHG durante el
crecimiento saprofítico y en respuesta a NO.
107
3.5.2.1 3.8.2.1. Expresión de FHG1. 108
3.5.2.2 3.8.2.2. Expresión de FHG2. 109
3.5.2.3 3.8.2.3. Expresión de FHG3. 110
3.5.2.4 3.8.2.4. Expresión de FHG4. 112
3.5.3 Análisis de expresión de los genes FHG en plantas de judía
infectadas con estirpes de F. oxysporum f. sp. phaseoli.
112
3.5.3.1 Análisis de expresión de los genes FHG en plantas
de judía infectadas con FOP-SP1.
114
3.5.3.2 Análisis de expresión de los genes FHG en plantas
de judía infectadas con FOP-SP4.
115
3.6 3.5. Análisis funcional de los genes tipo flavohemoglobina. 117
3.6.1 Construcción de los vectores de transformación. 117
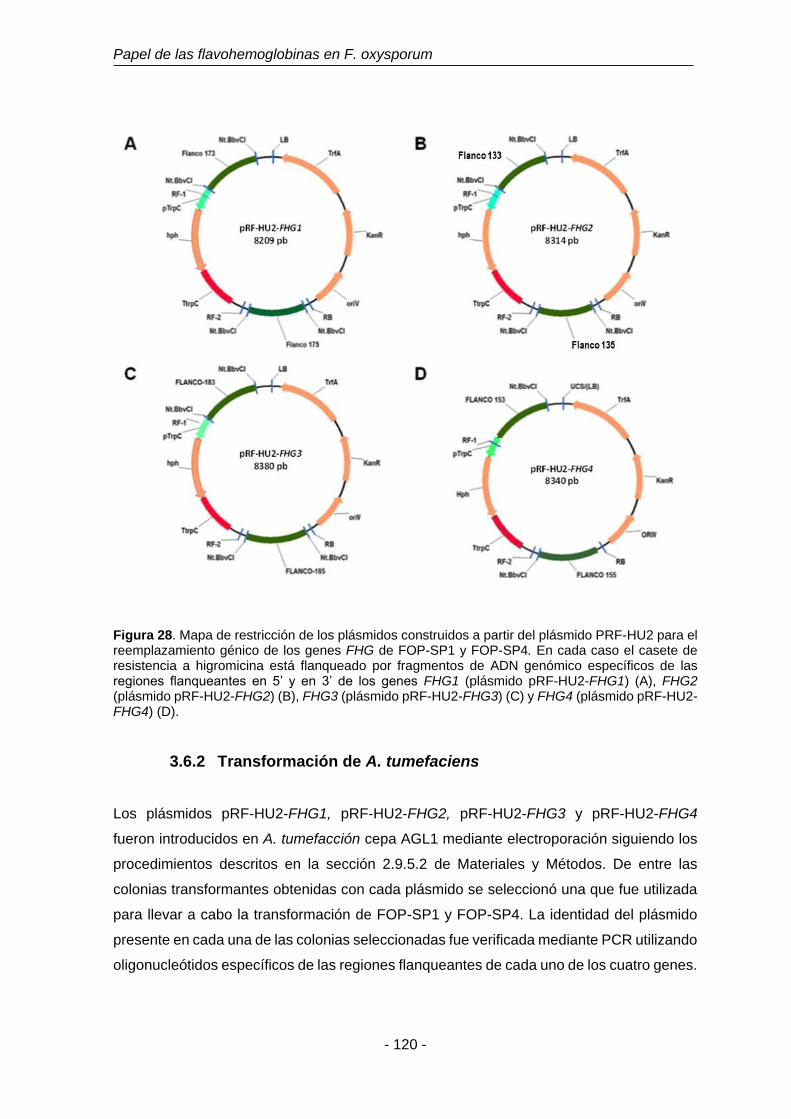
3.6.2 3.5.2. Transformación de A. tumefaciens. 120
3.6.3 3.5.3. Transformación de F. oxysporum de f. sp. phaseoli mediada
por A. tumefaciens.
121
3.6.4 3.5.4. Identificación de mutantes por reemplazamiento génico de F.
oxysporum f. sp. phaseoli.
121
3.6.4.1 3.5.4.1. Identificación de transformantes del gen FHG1. 123
3.6.4.2 3.5.4.2. Identificación de transformantes del gen FHG2. 125
3.6.4.3 3.5.4.3. Identificación de transformantes del gen FHG3. 127
3.6.4.4 3.5.4.4. Identificación de transformantes del gen FHG4. 129
3.7 Análisis mediante hibridación Southern de los transformantes de F.
oxysporum f. sp. phaseoli.
131
3.7.1 Análisis de los mutantes en el gen FHG1. 132
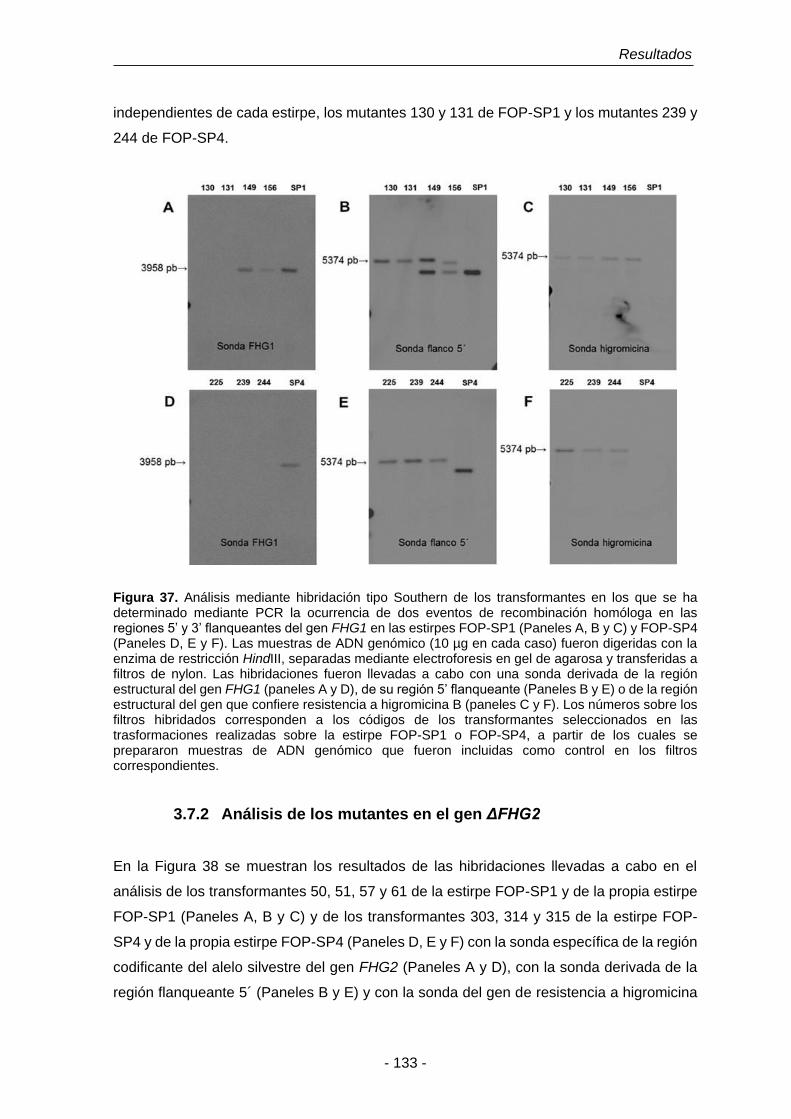
3.7.2 Análisis de los mutantes en el gen FHG2. 133
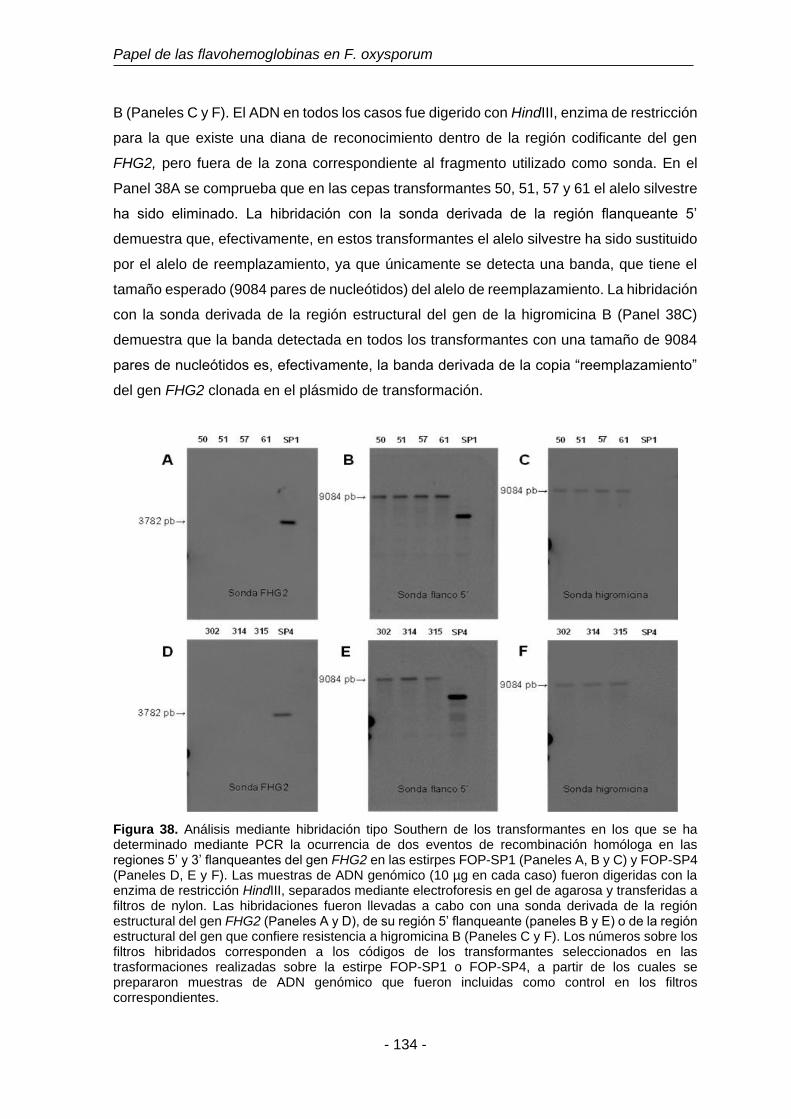
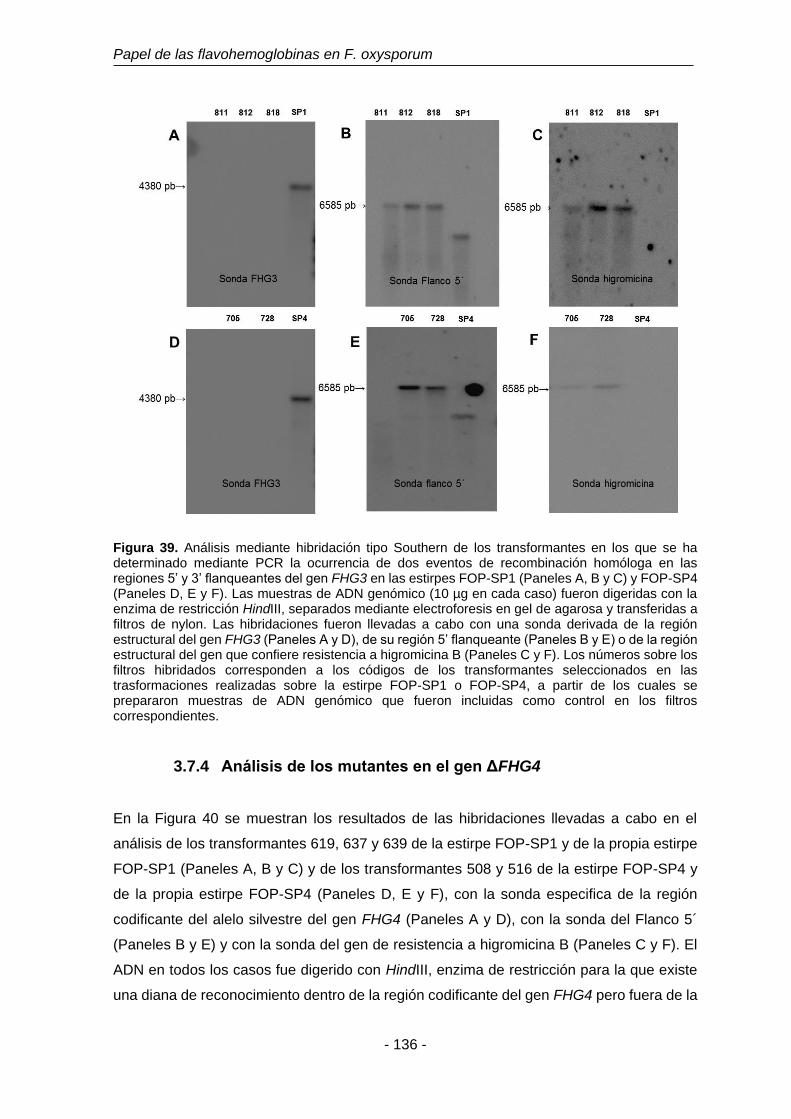
3.7.3 Análisis de los mutantes en el gen FHG3. 135
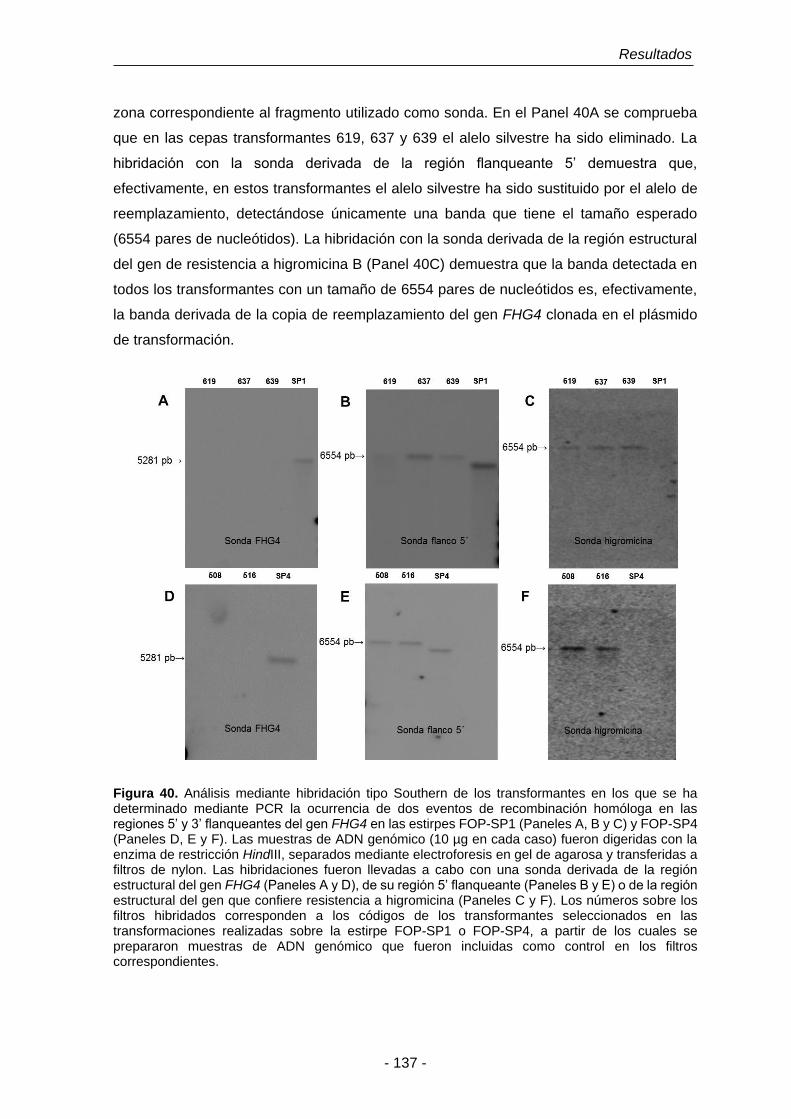
3.7.4 Análisis de los mutantes en el gen FHG4. 136
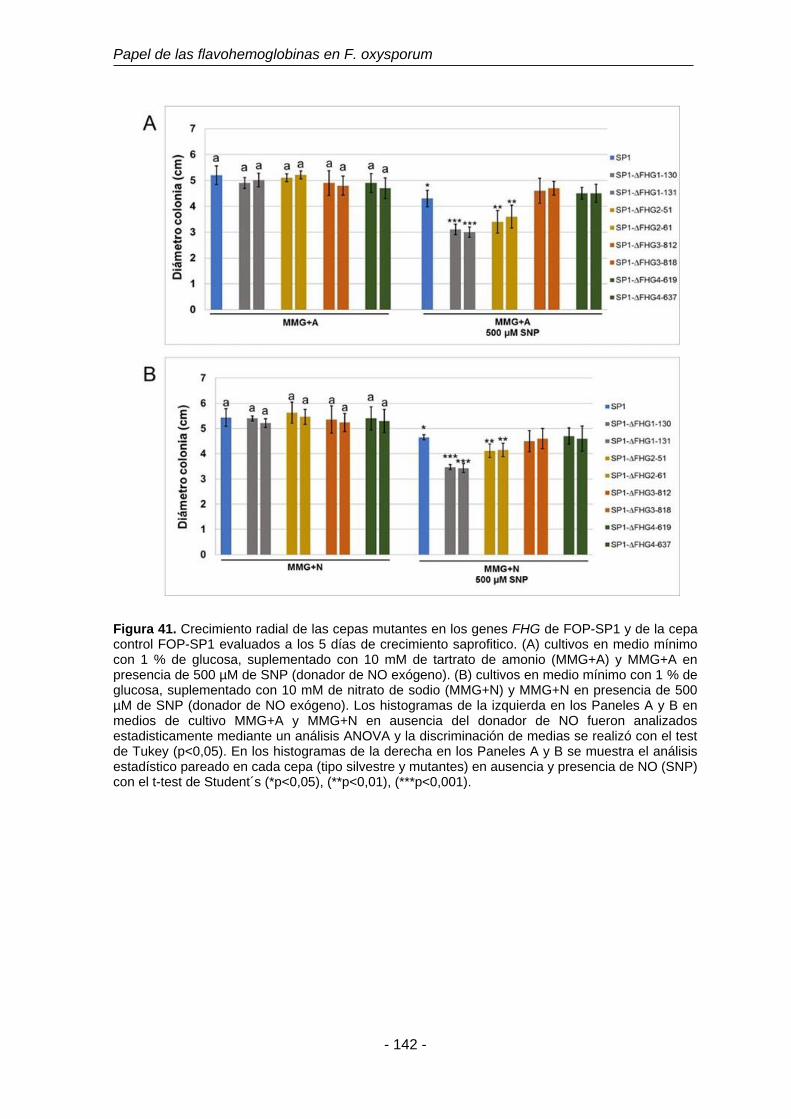
3.8 3.6. Caracterización fisiológica de los mutantes FHG. 138
3.8.1 Análisis de la capacidad de crecimiento saprofítico y de la
sensibilidad a estrés nitrosativo de los mutantes ∆FHG.
139
3.8.1.1 Análisis de mutantes ΔFHG de FOP-SP1. 140
3.8.1.2 Análisis de mutantes ΔFHG de FOP-SP4. 143
3.8.2 3.6.2. Análisis de germinación de microconidios de mutantes ΔFHG. 146
pRF-HU2-FHG1
Alelo WT FHG1
Alelo mutante
pRF-HU2-FHG1
Alelo WT FHG1
Índice
VII
3.8.2.1 3.6.2.1. Análisis de germinación de microconidios de
mutantes ΔFHG de FOP-SP1.
147
3.8.2.2 3.6.2.2. Análisis de germinación de microconidios de
mutantes ΔFHG de FOP-SP4.
150
3.8.3 3.6.3. Ensayos de infección in planta con los mutantes ΔFHG. 153
3.9 3.7. Ensayos de complementación de la mutación ΔFHG1. 155
3.9.1 Construcción del vector de complementación. 156
3.9.2 Transformación de A. tumefaciens. 156
3.9.3 3.7.2.1. Transformación de mutantes ∆FHG1 de FOP-SP1 y FOP-
SP4.
157
3.9.4 Identificación de transformantes que han incorporado la
construcción de complementación.
157
3.9.5 Análisis fenotípico de los mutantes portadores de la
construcción de complementación de la mutación ΔFHG1.
160
3.10
.
3.8. Obtención y caracterización de dobles mutantes ΔFHG2-FHG1RNAi. 163
3.10.1 Construcción del vector de silenciamiento. 164
3.10.2 Transformación de A. tumefaciens. 164
3.10.3 3.8.2.1. Transformación de mutantes ∆FHG2 con el vector de
silenciamiento pVIR-RNAi-FHG1.
165
3.8.2.2. 3.10.3.1 3.8.2.3. Identificación y selección de transformantes
ΔFHG2-FHG1RNAi.
166
3.8.2.4. 3.10.3.2 3.8.2.5. Análisis fenotípico de los dobles mutantes ΔFHG2-
FHG1RNAi.
169
3.8.2.6. 3.10.3.3 3.8.2.7. Ensayos de infección in planta con los dobles
mutantes ∆FHG2-FHG1RNAi.
174
4 DISCUSIÓN 177
4.1 Relaciones filogenéticas de los genes FHG de FOP. 177
4.2 Análisis de expresión y funcional de los genes de FHG. 179
4.3 3.9. Los genes FHG de FOP muestran patrones de expresión diferenciales. 180
4.4 Los genes FHG1 y FHG2 confieren resistencia a estrés nitrosativo. 185
4.5 3.9.2.1. El papel de FHG1 y FHG2 en la detoxificación de NO evidencia una
función del NO sobre germinación.
188
4.6 3.9.2.2. El papel del NO y de las flavohemoglobinas en diferenciación y
desarrollo.
190
4.7 El papel de los genes FHG durante la interacción con el huésped. 193
5 CONCLUSIONES 195
6 BIBLIOGRAFÍA 197
Papel de las flavohemoglobinas en F. oxysporum
VIII
Índice
IX
ÍNDICE DE FIGURAS
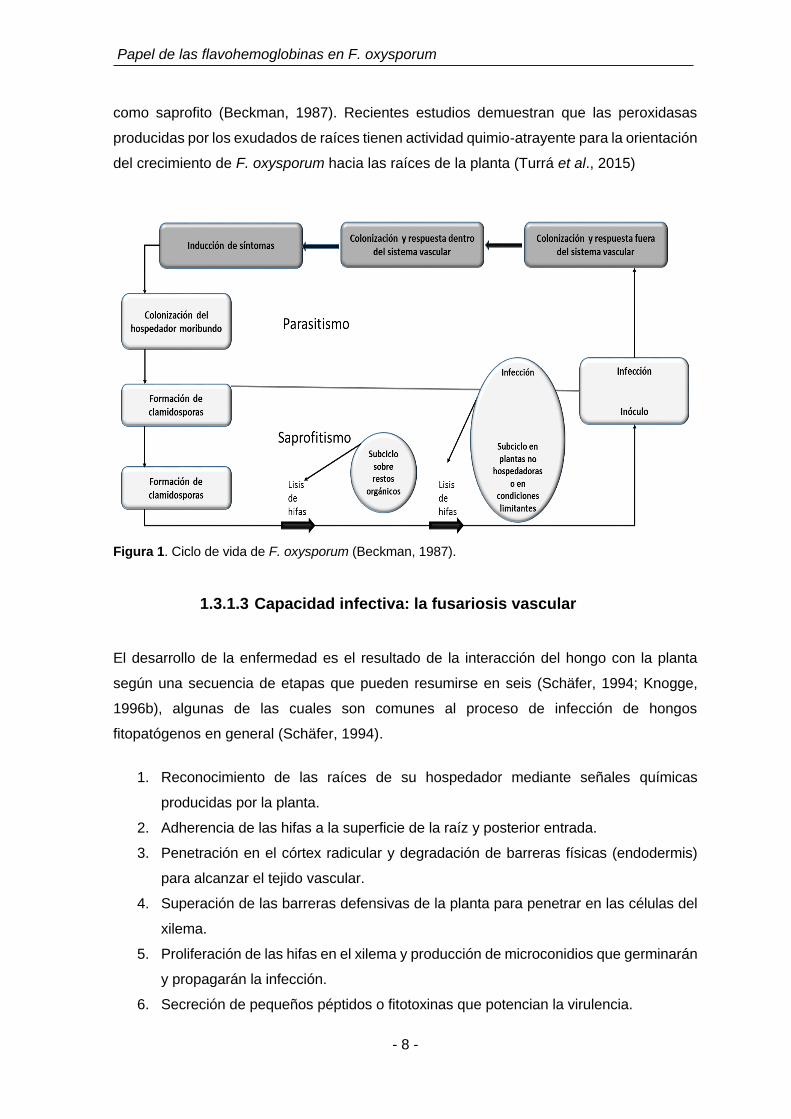
Figura 1. Ciclo de vida de F. oxysporum (Beckman, 1987).
8
Figura 2. Estructura tridimensional de la proteína flavohemoglobina de E.
coli.
28
Figura 3. Mapa circular del Plásmido pGEM-T easy.
56
Figura 4. Mapa circular del Plásmido PRF-HU2 y estrategia de clonación.
57
Figura 5. Mapa de construcción del plásmido pEA-FU utilizado para
complementación.
58
Figura 6. Mapa circular del plásmido pVIR-RNAi utilizado para
silenciamiento genético.
60
Figura 7. Electroforesis en gel de agarosa de los productos de amplificación
por PCR de secuencias derivadas de la región codificante de los
genes codificadores de proteínas de tipo flavohemoglobina
utilizando ADN genómico de F. oxysporun f. sp. phaseoli.
82
Figura 8. Análisis tipo Southern de los genes codificadores de enzimas
flavohemoglobina en las diferentes estirpes de F. oxysporum f. sp.
phaseoli.
84
Figura 9. Secuencia de nucleótidos del gen FHG1 y de la secuencia de
aminoácidos de la proteína deducida.
86
Figura 10. Secuencia de nucleótidos del gen FHG2 y de la secuencia de
aminoácidos de la proteína deducida.
87
Figura 11 Secuencia de nucleótidos del gen FHG3 y de la secuencia de
aminoácidos de la proteína deducida.
88
Figura 12. Secuencia de nucleótidos del gen FHG4 y de la secuencia de
aminoácidos de la proteína deducida.
89
Figura 13. Alineamiento de secuencias de las proteínas tipo
flavohemoglobina de FOP-SP1.
95
Figura 14. Dominios funcionales característicos de las proteínas de tipo
flavohemoglobina, y sus posiciones relativas, identificados en las
proteínas deducidas codificadas por los genes tipo
flavohemoglobina de FOP-SP1.
96
Papel de las flavohemoglobinas en F. oxysporum
X
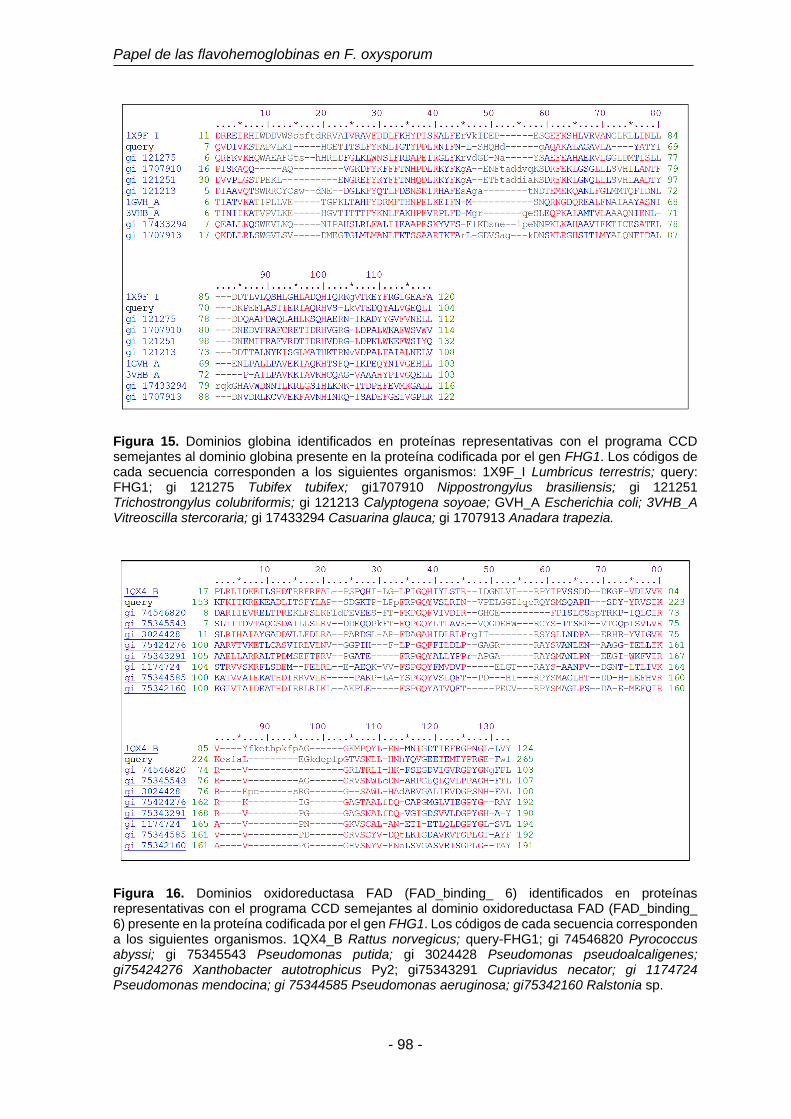
Figura 15. Dominios globina identificados en proteínas representativas con el
programa CCD semejantes al dominio globina presente en la
proteína codificada por el gen FHG1.
98
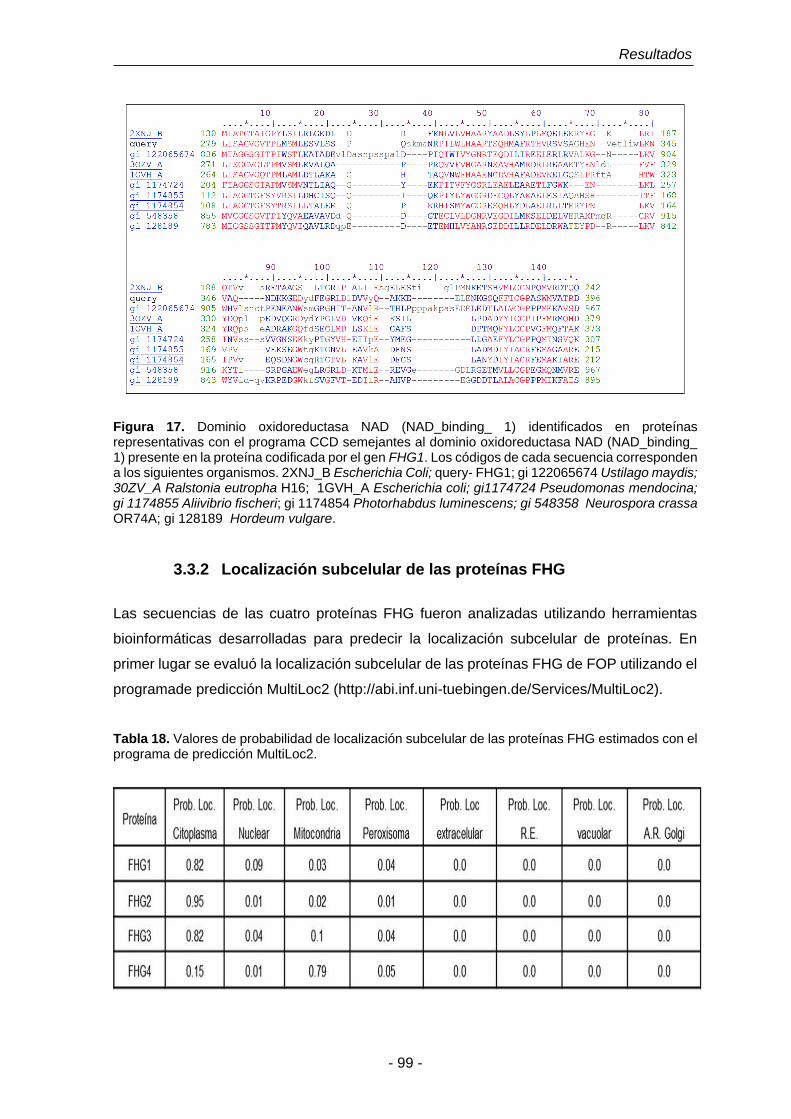
Figura 16. Dominios oxidoreductasa FAD (FAD_binding_ 6) identificados en
proteínas representativas con el programa CCD semejantes al
dominio oxidoreductasa FAD (FAD_binding_ 6) presente en la
proteína codificada por el gen FHG1.
98
Figura 17. Dominio oxidoreductasa NAD (NAD_binding_ 1) identificados en
proteínas representativas con el programa CCD semejantes al
dominio oxidoreductasa NAD (NAD_binding_ 1) presente en la
proteína codificada por el gen FHG1.
99
Figura 18. Árbol filogénetico de las proteínas de tipo flavohemoglobina.
102
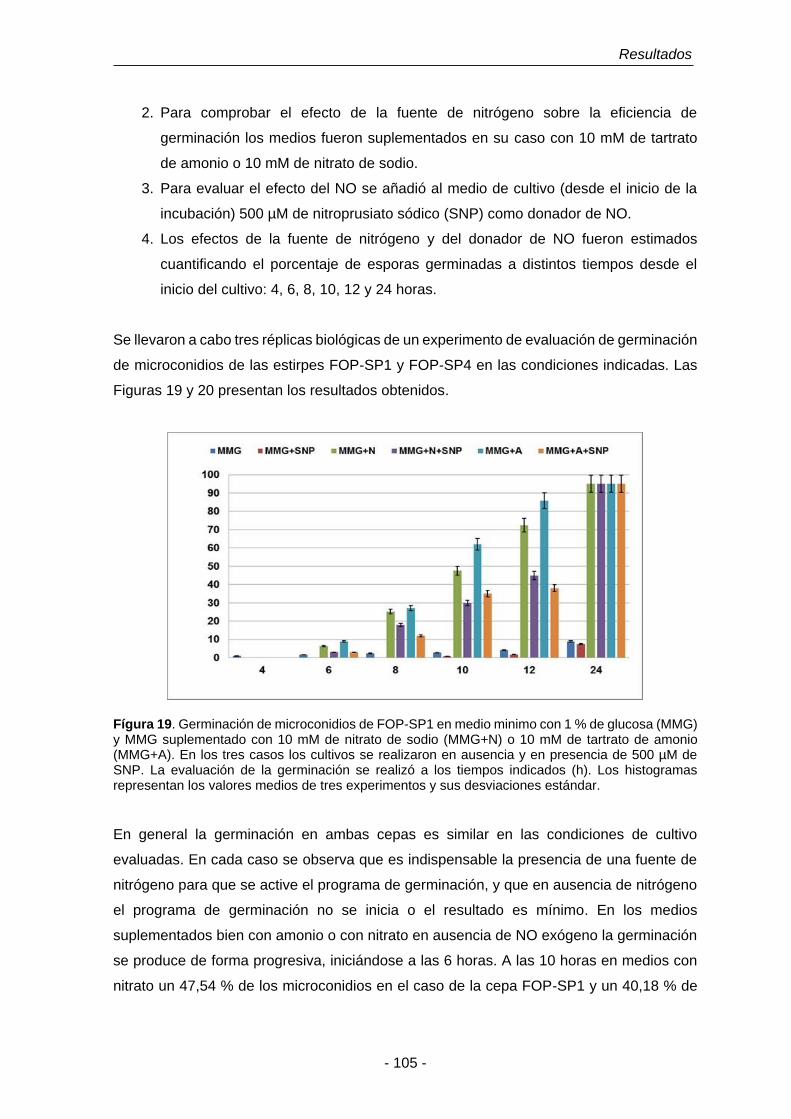
Figura 19. Germinación de microconidios de FOP-SP1 en medio mínimo con
1% de glucosa (MMG) y MMG suplementado con 10 mM de
tartrato de amonio (MMG+A) o 10 mM de nitrato de sodio
(MMG+N).
105
Figura 20. Germinación de microconidios de FOP-SP4 en medio mínimo
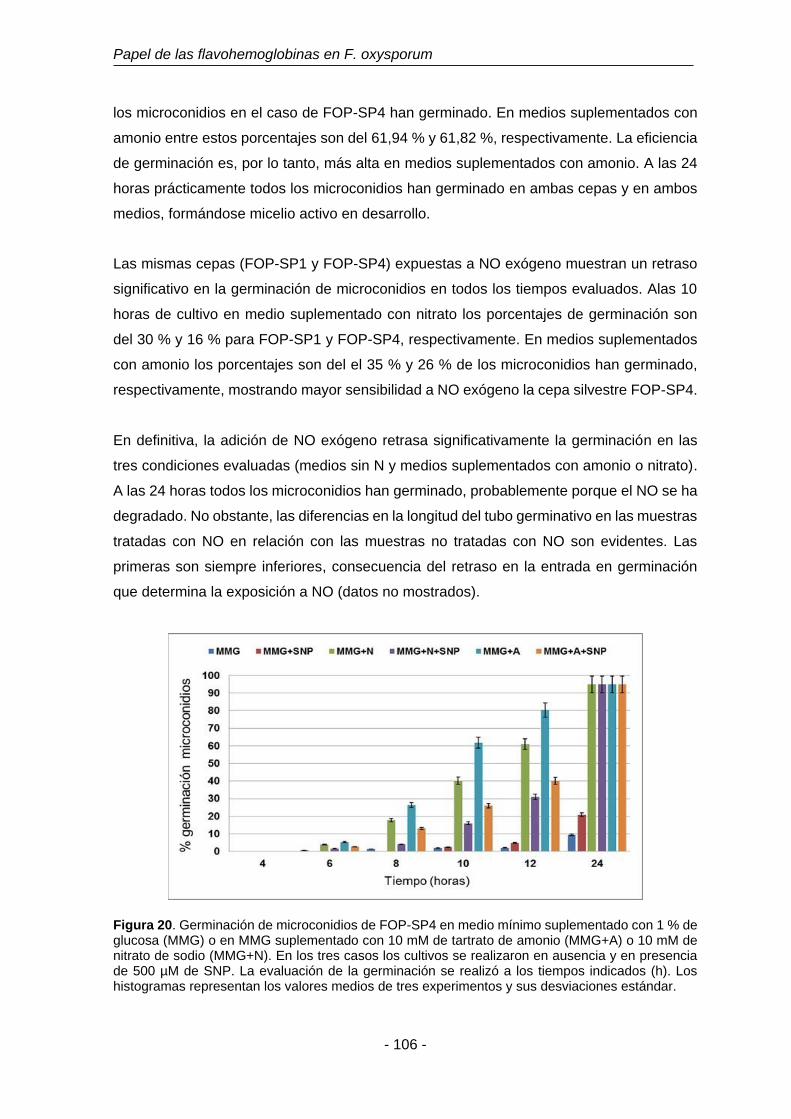
suplementado con 1% de glucosa (MMG) o en MMG
suplementado con 10 mM de tartrato de amonio (MMG+A) o 10
mM de nitrato de sodio (MMG+N).
106
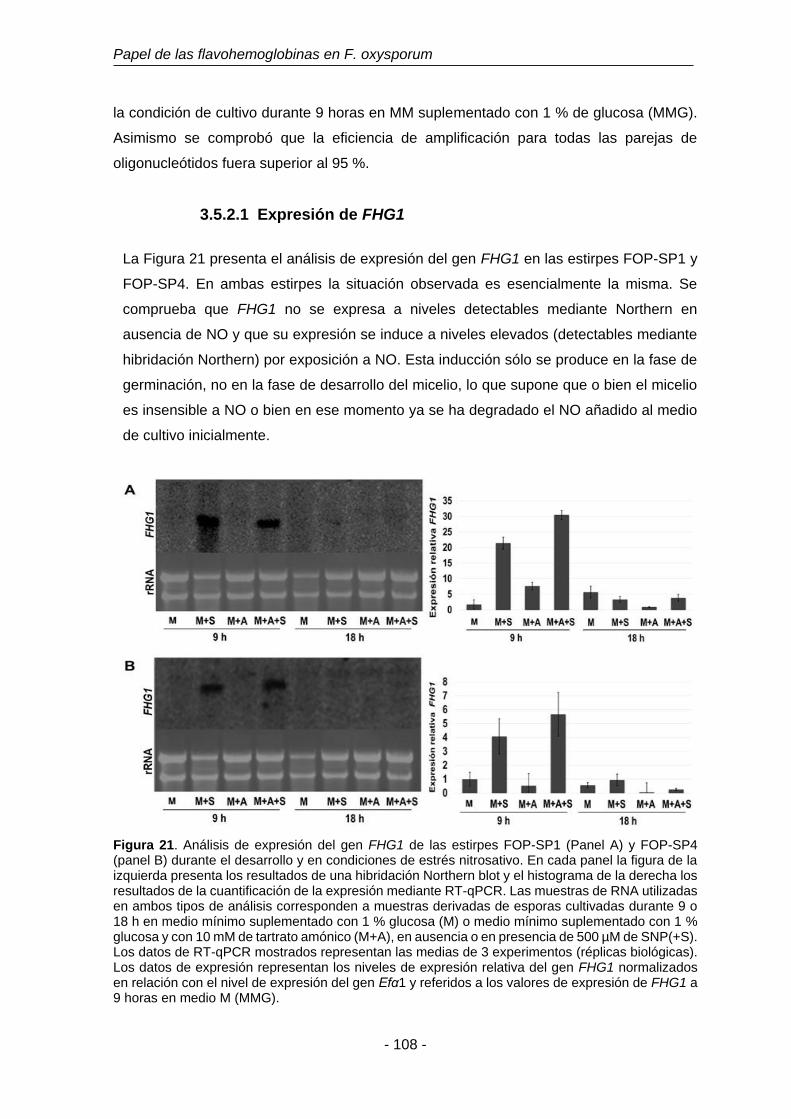
Figura 21. Análisis de expresión del gen FHG1 de las estirpes FOP-SP1 y
FOP-SP4.
108
Figura 22. Análisis de expresión del gen FHG2 de las estirpes FOP-SP1 y
FOP-SP4.
110
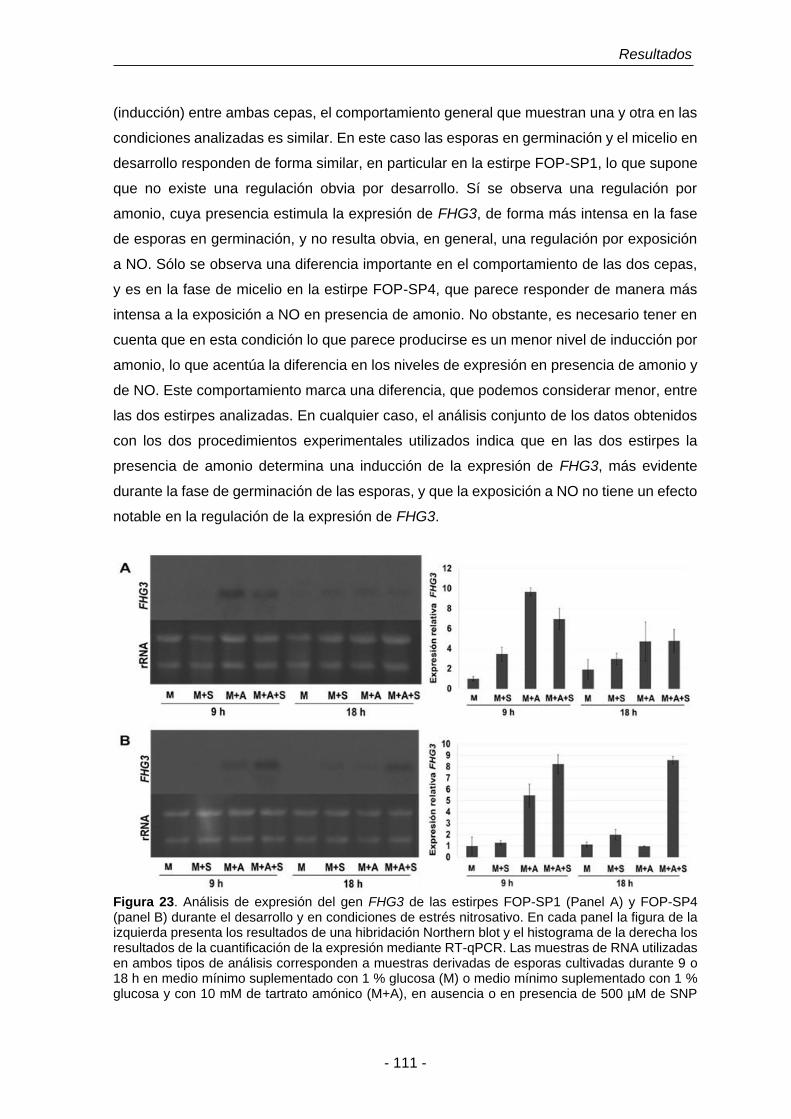
Figura 23. Análisis de expresión del gen FHG3 de las estirpes FOP-SP1 y
FOP-SP4.
111
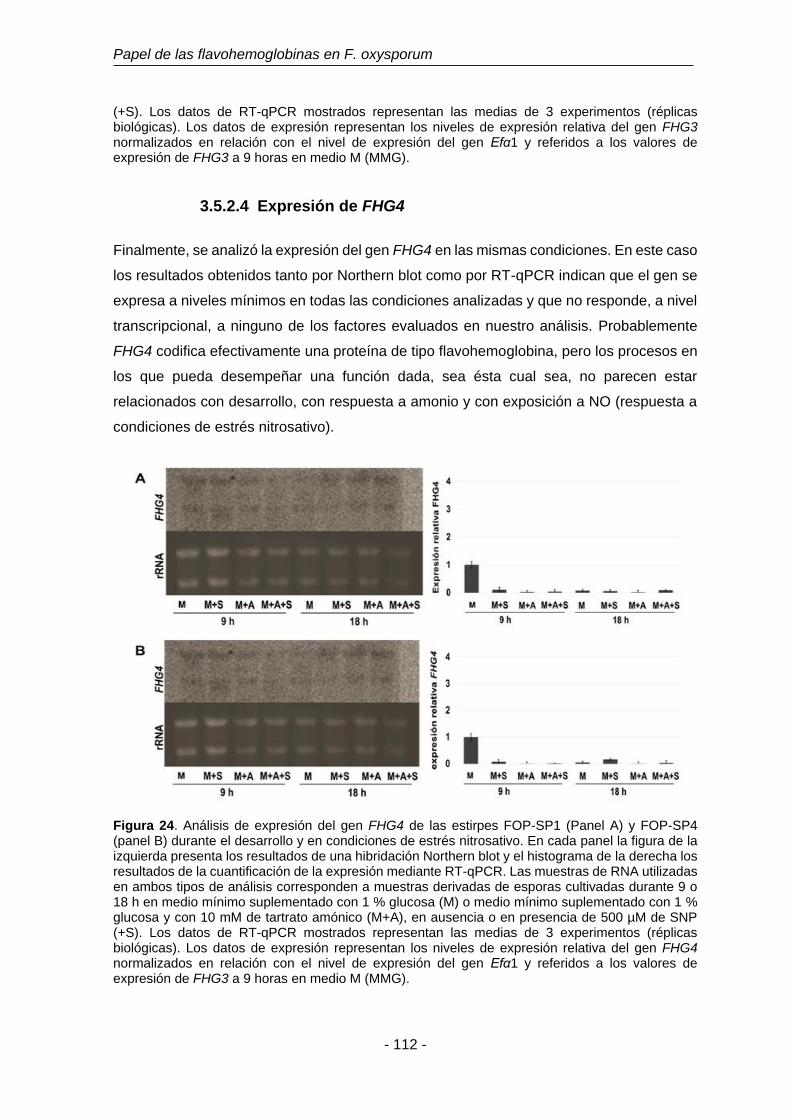
Figura 24. Análisis de expresión del gen FHG4 de las estirpes FOP-SP1 y
FOP-SP4.
112
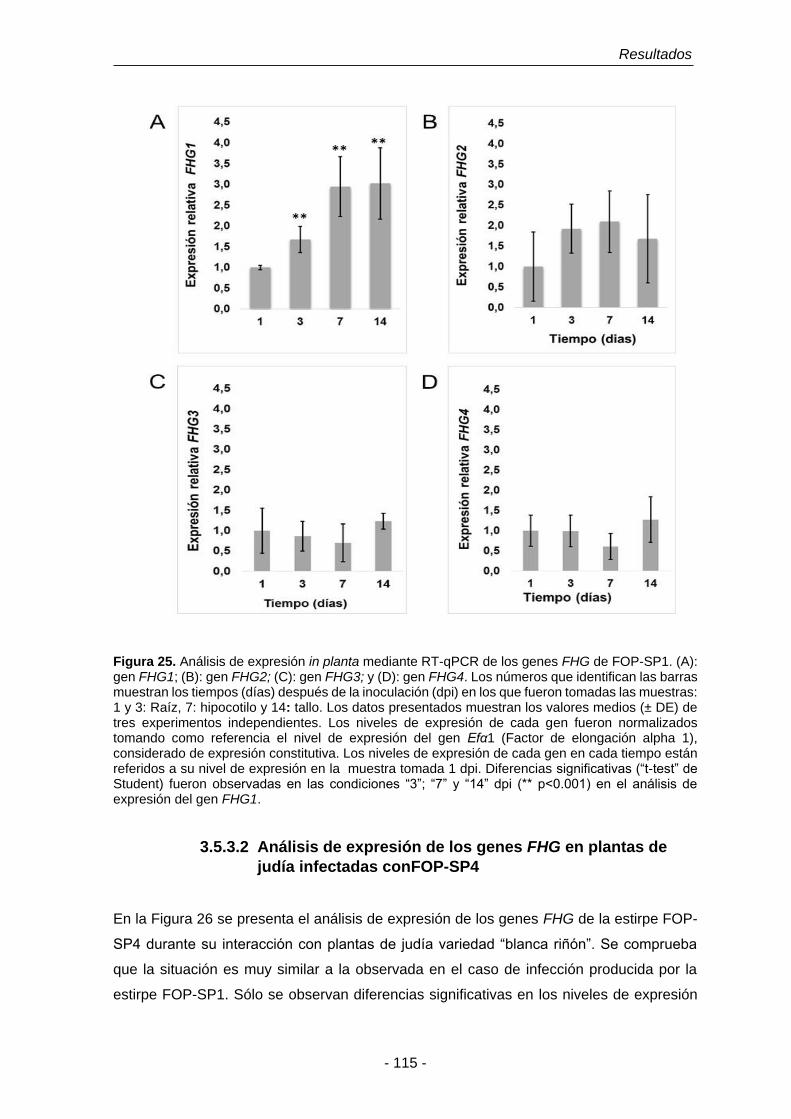
Figura 25. Análisis de expresión in planta mediante RT-qPCR de los genes
FHG de FOP-SP1.
115
Figura 26. Análisis de expresión in planta mediante RT-qPCR de los genes
FHG de FOP-SP4.
116
Figura 27. Productos de PCR obtenidos a partir de ADN genómico de FOP-
SP1 de las regiones flanqueantes 5´ y 3´ de los genes FHG.
119
Índice
XI
Figura 28. Mapa de restricción de los plásmidos construidos a partir del
plásmido PRF-HU2 para el reemplazamiento génico de los genes
FHG de FOP-SP1 y FOP-SP4.
120
Figura 29. Estrategia de reemplazamiento génico del gen FHG1 de FOP-SP1
y FOP-SP4.
123
Figura 30. Productos de PCR obtenidos en reacciones de PCR llevadas a
cabo con los oligonucleótidos FHG1-KN-FW y HphBC (5’) y RHG1-
KN-RV y HphAC (3’) utilizando como molde ADN genómico de los
transformantes obtenidos con el plásmido pRF-HU2-FHG1.
124
Figura 31. Estrategia de reemplazamiento génico del gen FHG2 de FOP-SP1
y FOP-SP4.
125
Figura 32. Productos de PCR obtenidos en reacciones de PCR llevadas a
cabo con los oligonucleótidos FHG2-KN-FW y HphBC (5’) y RHG2-
KN-RV y HphAC (3’) utilizando como molde ADN genómico de los
transformantes obtenidos con el plásmido pRF-HU2-FHG2.
126
Figura 33. Estrategia de reemplazamiento génico del gen FHG3 de FOP-SP1
y FOP-SP4.
127
Figura 34. Productos de PCR obtenidos en reacciones de PCR llevadas a
cabo con los oligonucleótidos FHG3-KN-FW y HphBC (5’) y RHG3-
KN-RV y HphAC (3’) utilizando como molde ADN genómico de los
transformantes obtenidos con el plásmido pRF-HU2-FHG3.
128
Figura 35. Estrategia de reemplazamiento génico del gen FHG4 de FOP-SP1
y FOP-SP4.
129
Figura 36. Productos de PCR obtenidos en reacciones de PCR llevadas a
cabo con los oligonucleótidos FHG4-KN-FW y (5’) y RHG4-KN-RV
y HphAC (3’) utilizando como molde ADN genómico de los
transformantes obtenidos con el plásmido pRF-HU2-FHG4.
130
Figura 37. Análisis mediante hibridación tipo Southern de los transformantes
en los que se ha determinado mediante PCR la ocurrencia de dos
eventos de recombinación homóloga en las regiones 5’ y 3’
flanqueantes del gen FHG1 en las estirpes FOP-SP1 y FOP-SP4.
133
Figura 38. Análisis mediante hibridación tipo Southern de los transformantes
en los que se ha determinado mediante PCR la ocurrencia de dos
eventos de recombinación homóloga en las regiones 5’ y 3’
flanqueantes del gen FHG2 en las estirpes FOP-SP1 y FOP-SP4.
134
Papel de las flavohemoglobinas en F. oxysporum
XII
Figura 39. Análisis mediante hibridación tipo Southern de los transformantes
en los que se ha determinado mediante PCR la ocurrencia de dos
eventos de recombinación homóloga en las regiones 5’ y 3’
flanqueantes del gen FHG3 en las estirpes FOP-SP1 y FOP-SP4.
136
Figura 40. Análisis mediante hibridación tipo Southern de los transformantes
en los que se ha determinado mediante PCR la ocurrencia de dos
eventos de recombinación homóloga en las regiones 5’ y 3’
flanqueantes del gen FHG4 en las estirpes FOP-SP1 y FOP-SP4.
137
Figura 41. Crecimiento radial de las cepas mutantes de los genes FHG de
FOP-SP1 y de la misma cepa control FOP-SP1 evaluados a los 5
días de crecimiento saprofitico.
142
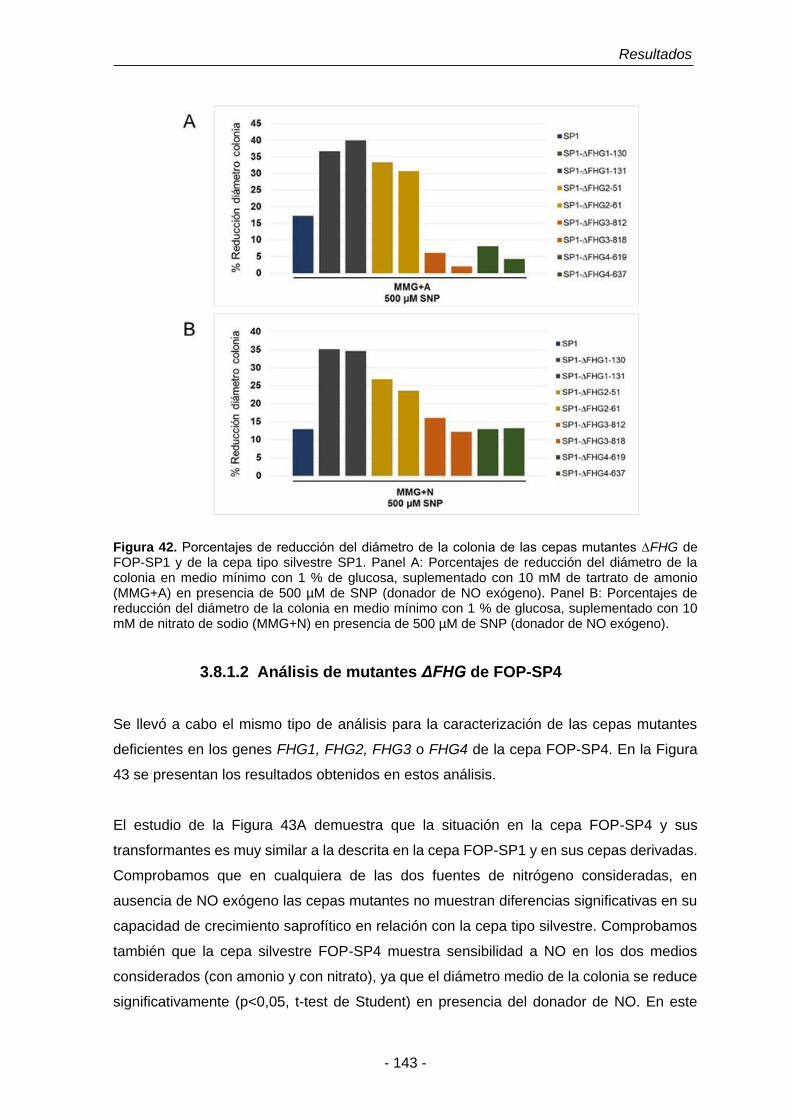
Figura 42. Porcentajes de reducción del diámetro de la colonia de las cepas
mutantes ∆FHG de FOP-SP1.
143
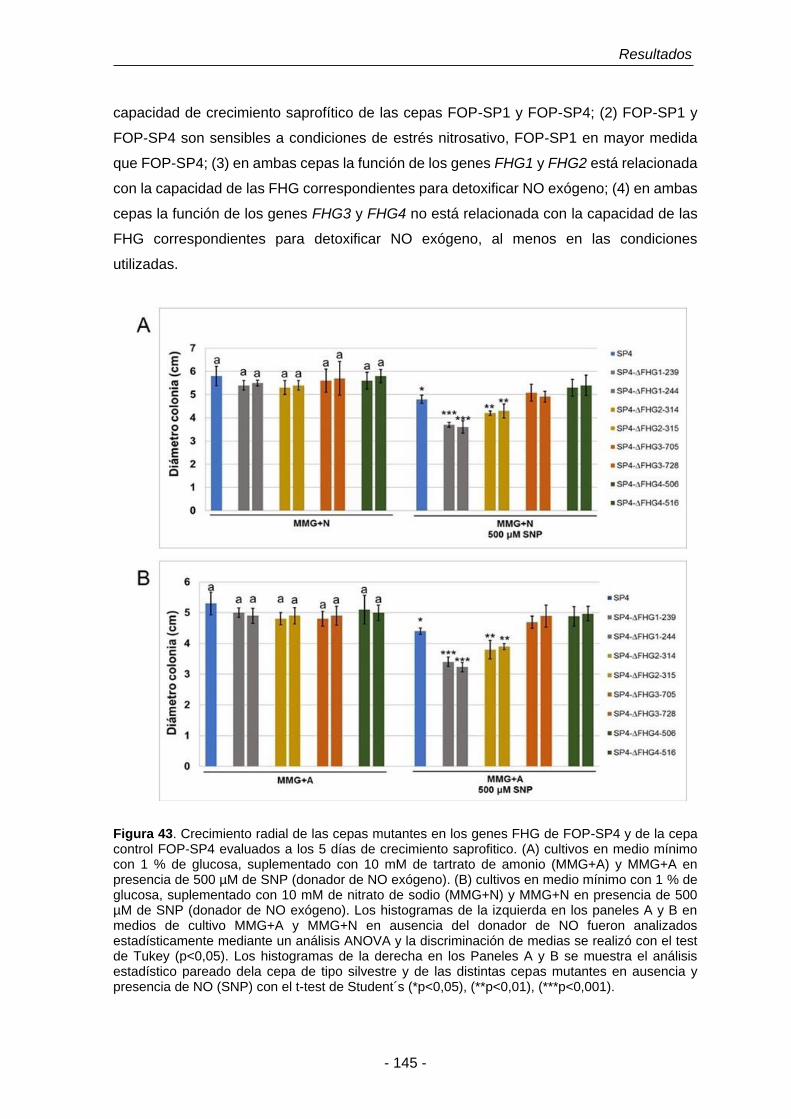
Figura 43. Crecimiento radial de las cepas mutantes de los genes FHG de
FOP-SP4 y de la cepa control FOP-SP4 evaluados a los 5 días de
crecimiento saprofitico.
145
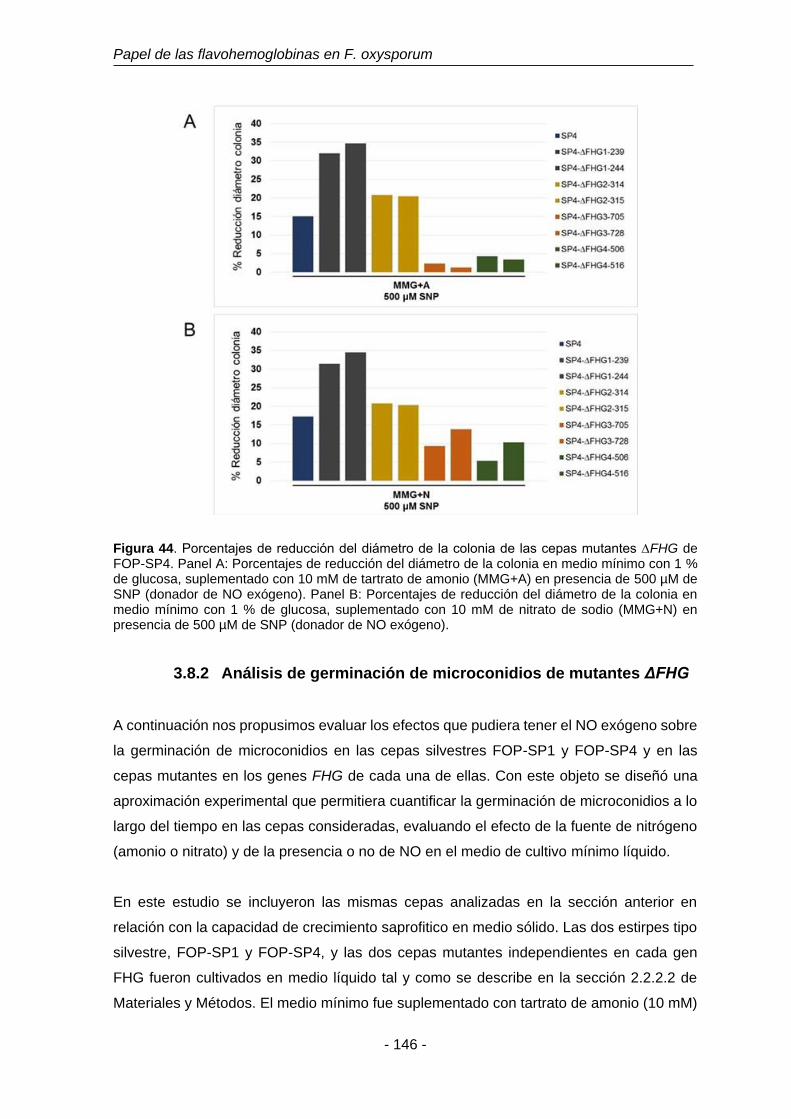
Figura 44. Porcentajes de reducción del diámetro de la colonia de las cepas
mutantes ∆FHG de FOP-SP4.
146
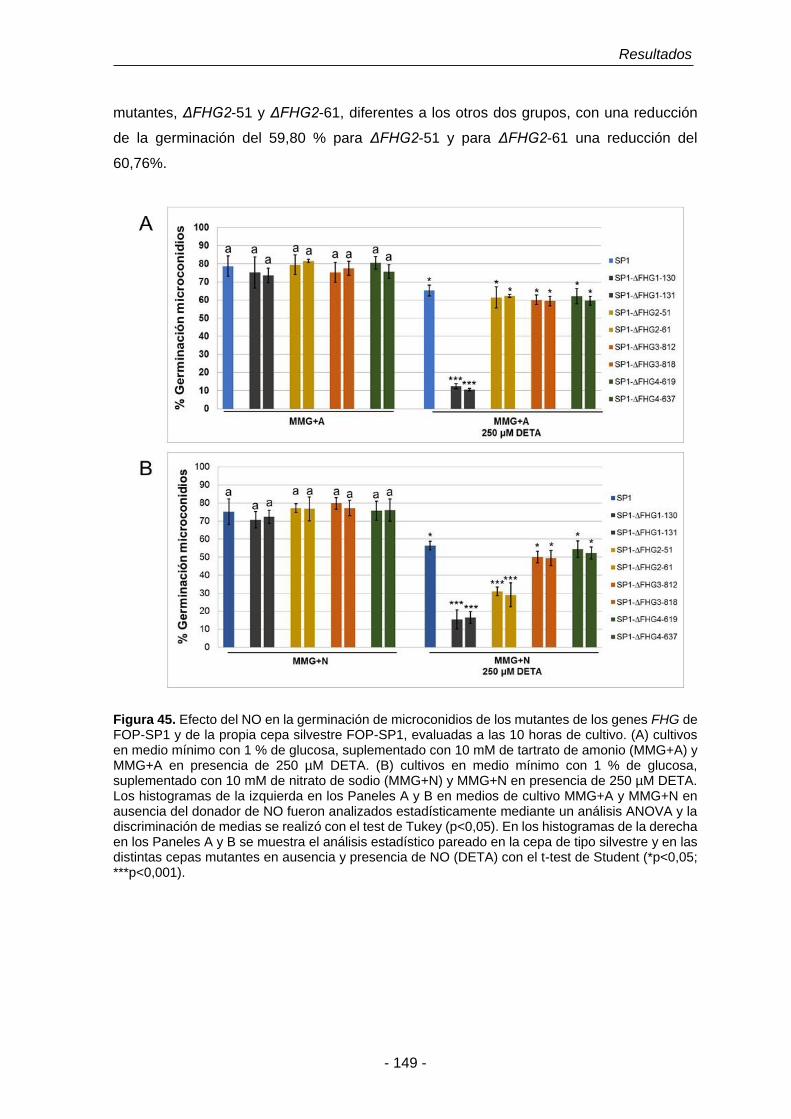
Figura 45. Efecto de NO en la germinación de microconidios de los mutantes
de los genes de FHG de SP1 y de la propia cepa SP1, evaluadas
a las 10 horas de cultivo.
149
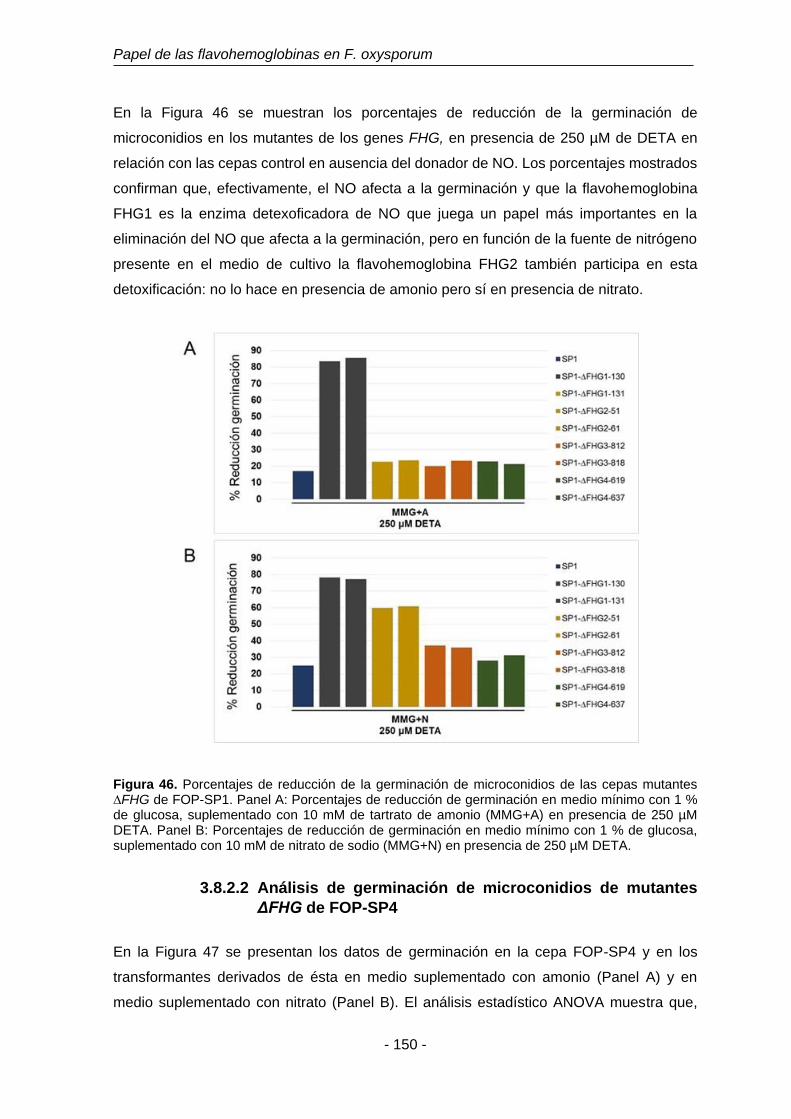
Figura 46. Porcentajes de reducción de la germinación de microconidios de
las cepas mutantes ∆FHG de FOP-SP1.
150
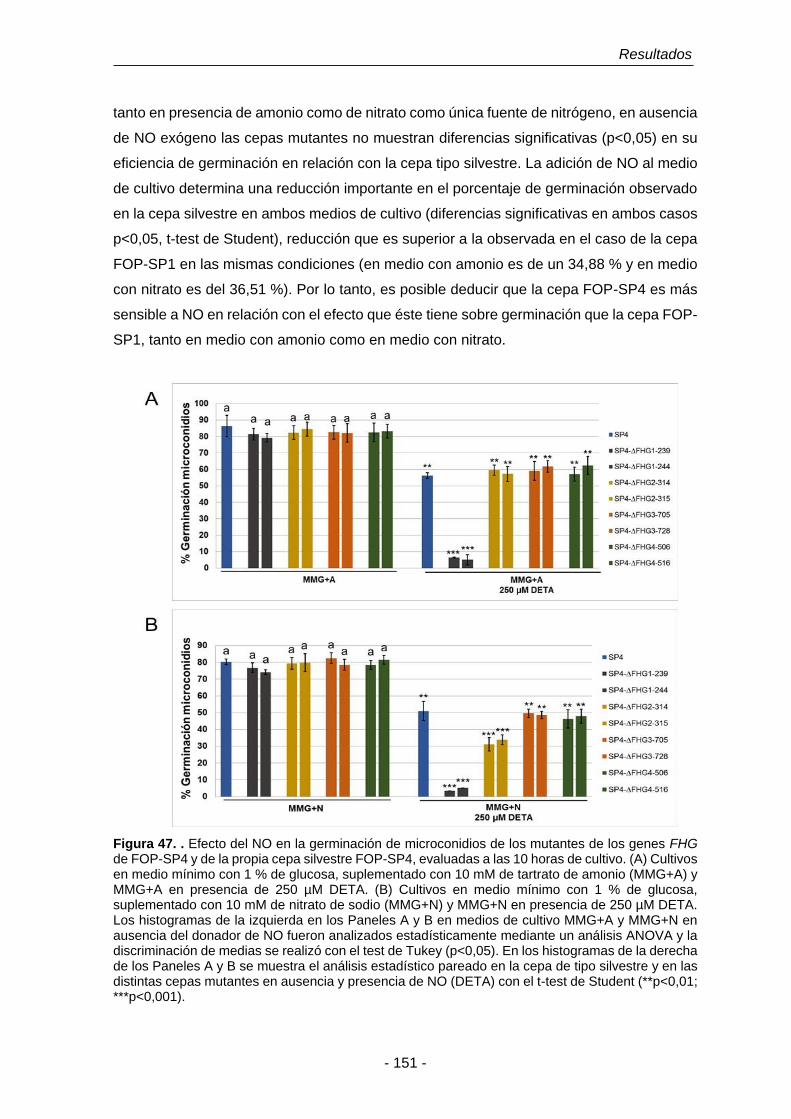
Figura 47. Efecto del NO en la germinación de microconidios de los mutantes
de los genes de FHG de SP4 y de la propia cepa SP4, evaluadas
a las 10 horas de cultivo.
151
Figura 48. Porcentajes de reducción de la germinación de microconidios de
las cepas mutantes ∆FHG de FOP-SP4.
152
Figura 49. Evaluación de la agresividad de los mutantes ∆FHG de FOP-SP1
y FOP-SP4.
155
Figura 50. Mapa de restricción del plásmido pEA-FU-FHG1 utilizado para la
complementación de la mutación ΔFHG1.
157
Índice
XIII
Figura 51. Identificación de los transformantes de las cepas SP1-ΔFHG1-131
y SP4-ΔFHG1-244 que han incorporado el gen FHG1 mediante
transformación con el plásmido pEA-FU-FHG1.
159
Figura 52. Evaluación de la sensibilidad a estrés nitrosativo de los mutantes
complementados SP1-ΔFHG1-131-C1 y SP4-ΔFHG1-244-C1
cultivados en MMG+A y MMG+N en ausencia y en presencia del
donador de NO SNP (500 µM).
161
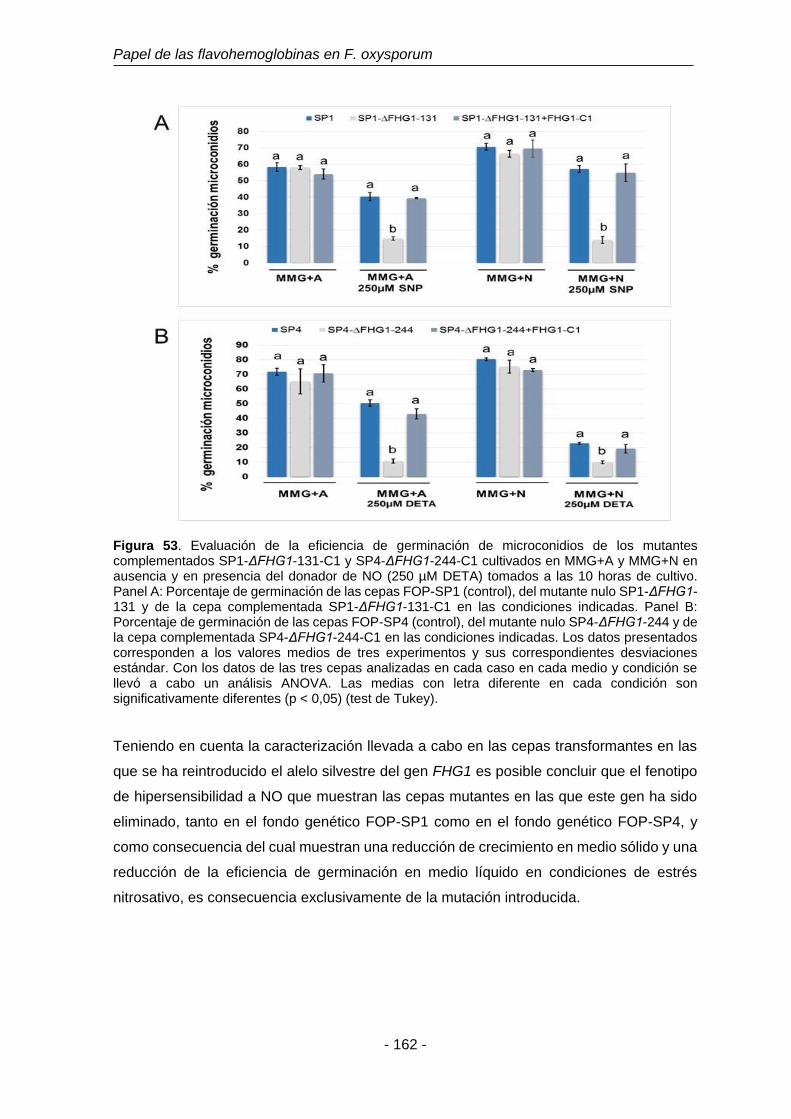
Figura 53. Evaluación de la eficiencia de germinación de microconidios de los
mutantes complementados SP1-ΔFHG1-131-C1 y SP4-ΔFHG1-
244-C1 cultivados en MMG+A y MMG+N en ausencia y en
presencia del donador de NO DETA (250 µM) tomados a las 10
horas de cultivo.
162
Figura 54. Producto de PCR amplificado a partir de ADN genómico de FOP-
SP1 con los cebadores Sil-FHG1-FW y Sil-FHG1-RV.
164
Figura 55. Mapa de restricción del plásmido pVIR-RNAi-FHG1 utilizado para
silenciamiento del gen FHG1.
165
Figura 56. Expresión relativa del gen FHG1 en los transformantes
seleccionados obtenidos con el plásmido pVIR-RNAi-FHG1 sobre
fondo genético de los mutantes SP1-∆FHG2-51 y SP4-∆FHG2-
314.
167
Figura 57. Caracterización molecular de los transformantes SP1-ΔFHG2-51 y
SP4-ΔFHG2-314 seleccionados en los experimentos de
transformación llevados a cabo con el plásmido de silenciamiento
pVIR-RNAi-FHG1.
169
Figura 58. Evaluación del crecimiento saprofitico de mutantes ΔFHG2 en los
que se ha silenciado el gen FHG1.
174
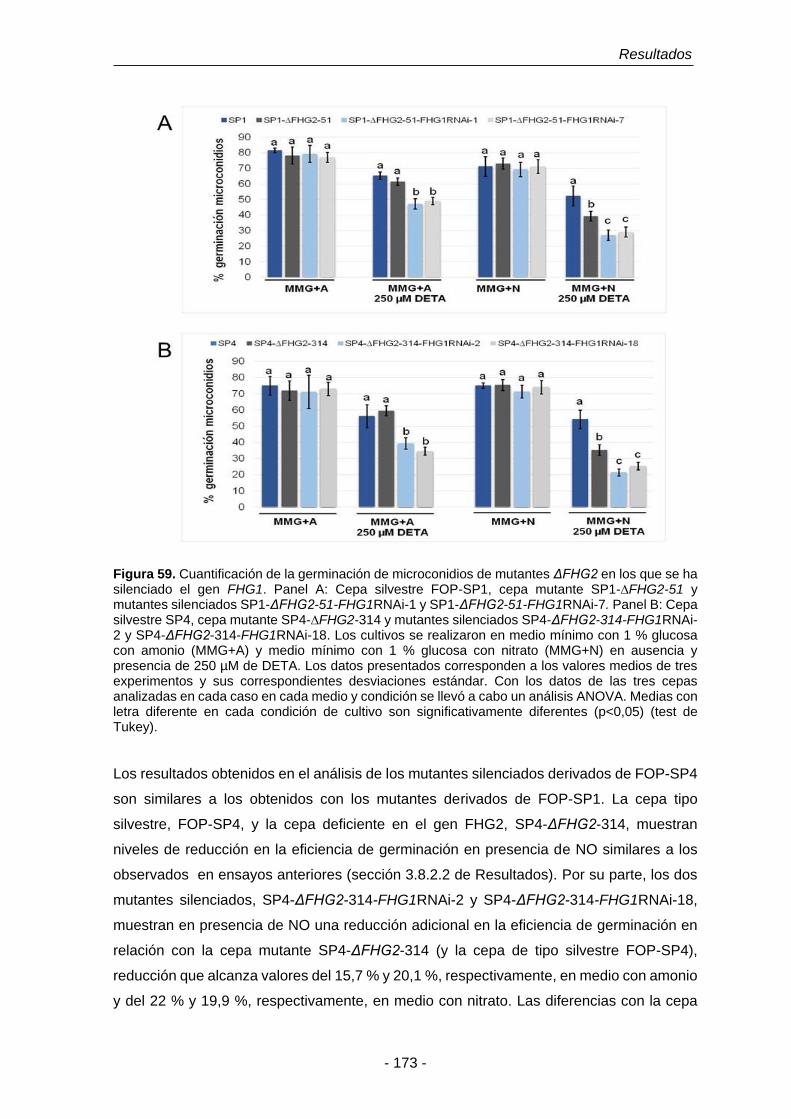
Figura 59. Cuantificación de la germinación de microconidios de mutantes
ΔFHG2 en los que se ha silenciado el gen FHG1.
173
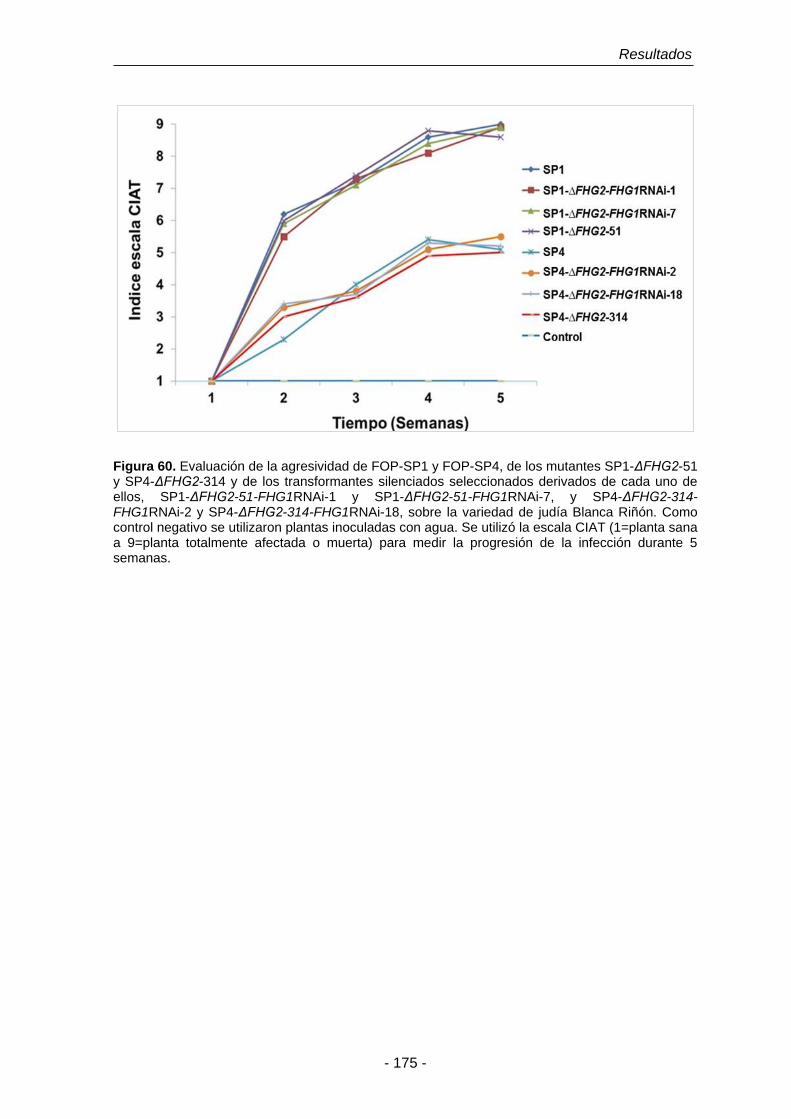
Figura 60. Ensayos de infección sobre la variedad de judía Blanca Riñón con
estirpes de F. oxysporum f. sp. phaseoli.
175
Papel de las flavohemoglobinas en F. oxysporum
XIV
Índice
XV
ÍNDICE DE TABLAS
Tabla 1. Grupo representativo de formas especiales del complejo de
especies de F. oxysporum.
6
Tabla 2. Genes de patogenicidad y virulencia identificados en F. oxysporum
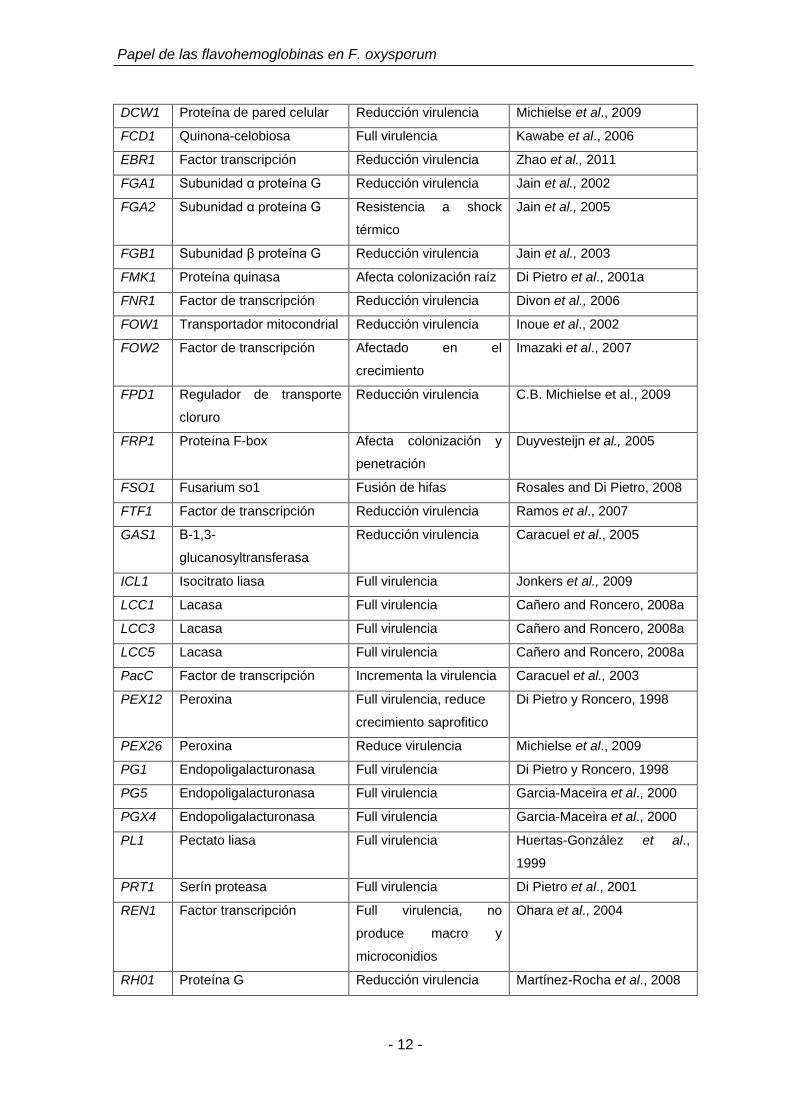
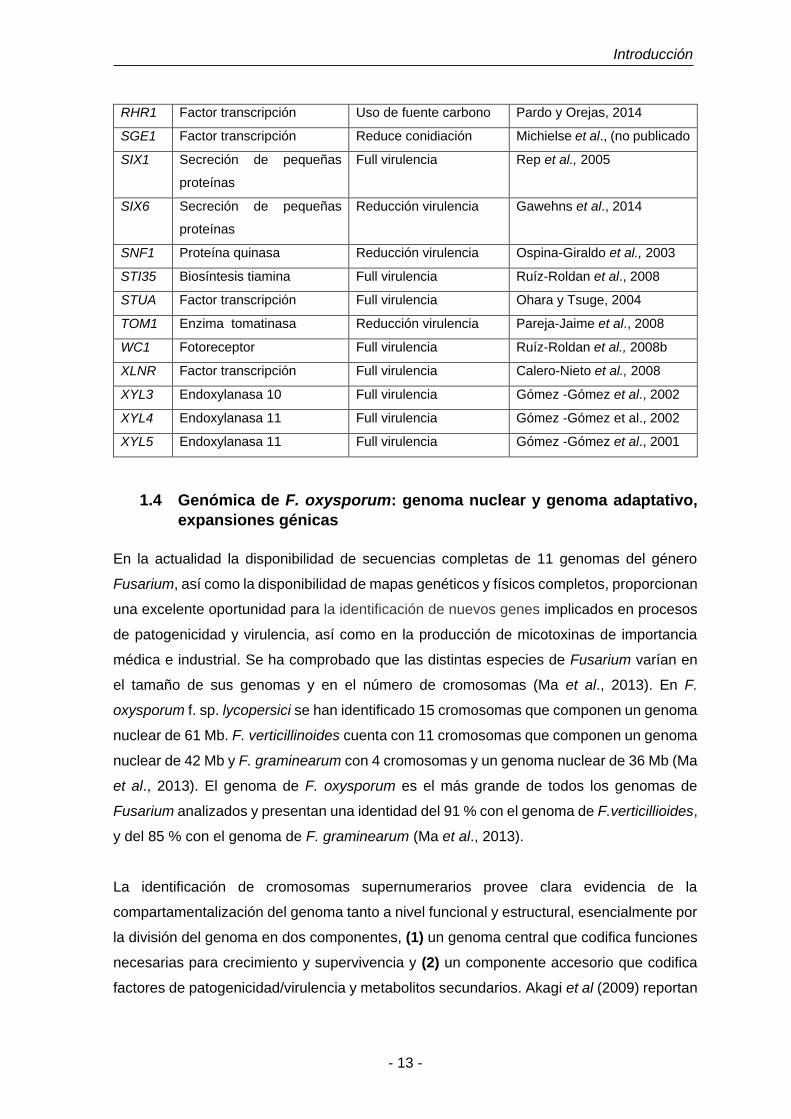
(tomado de Michielse and Rep, 2009).
11
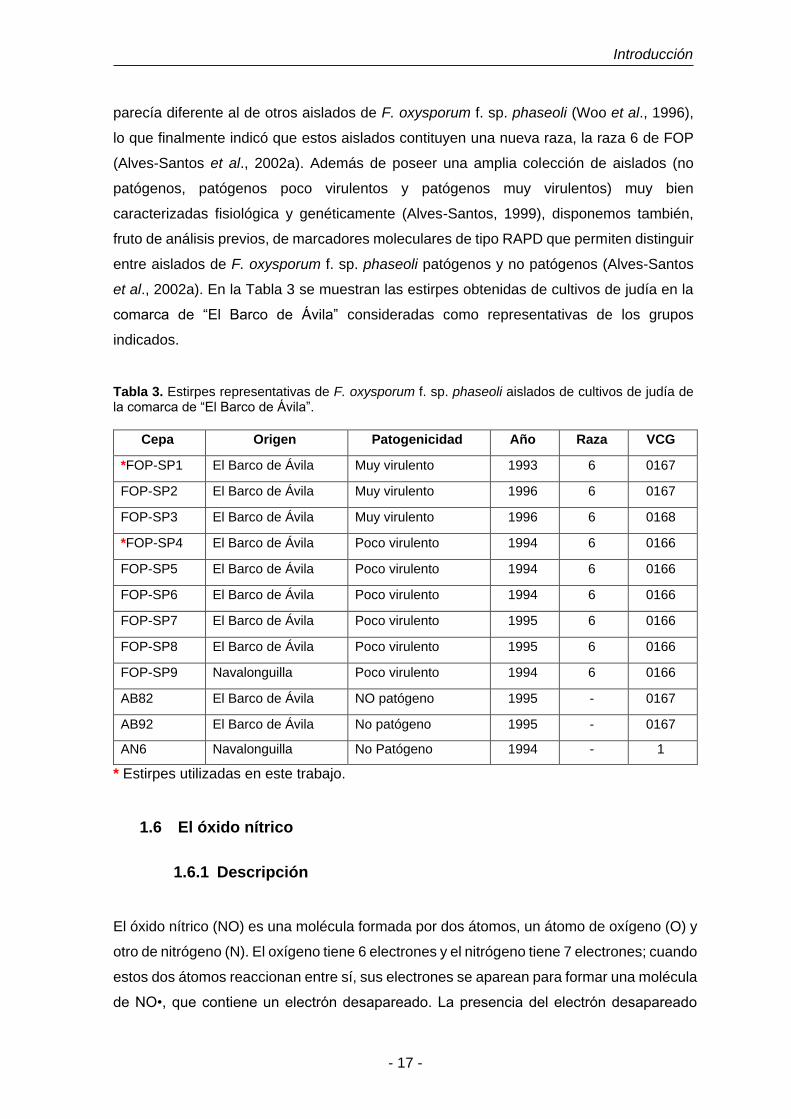
Tabla 3. Estirpes representativas de F. oxysporum f. sp. phaseoli aislados de
cultivos de judía de la comarca de “El Barco de Ávila”.
17
Tabla 4. Estirpes silvestres y transformantes de F. oxysporum utilizadas en
esta investigación.
37
Tabla 5. Medios de cultivo utilizados en la transformación de F. oxysporum
mediada por A. tumefaciens.
41
Tabla 6. Oligonucleótidos utilizados en este trabajo de investigación.
70
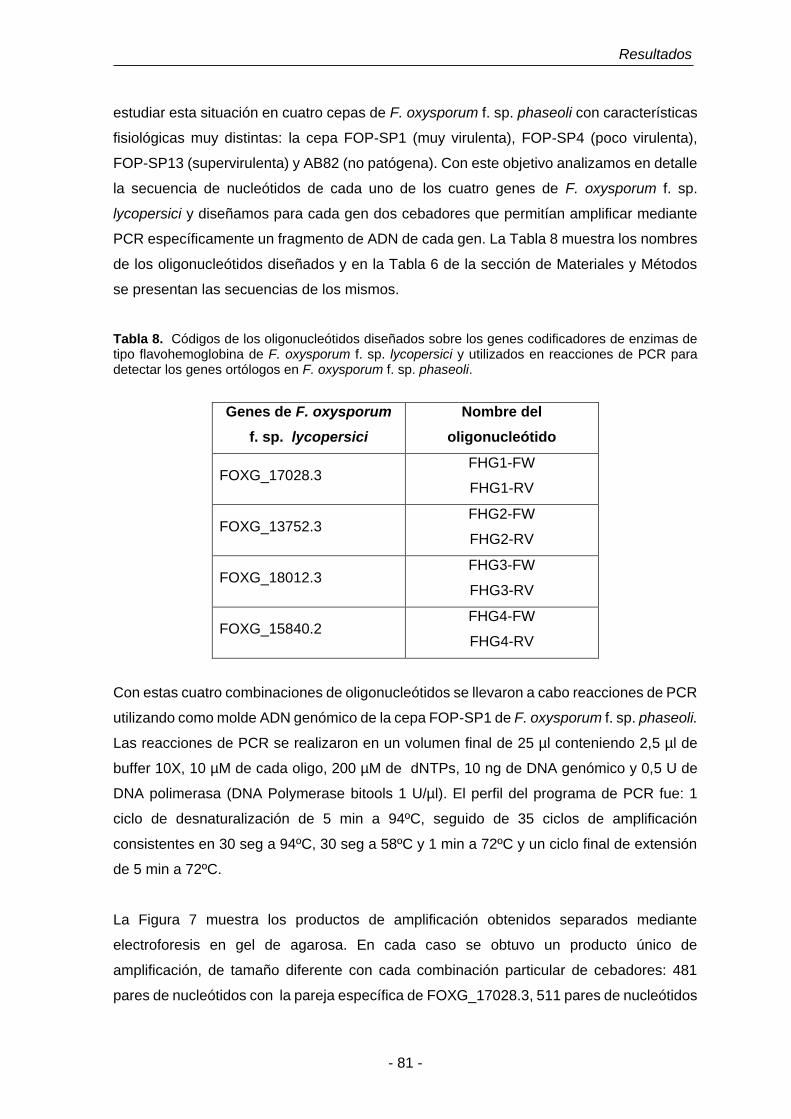
Tabla 7. Códigos de genes codificadores de proteínas tipo flavohemoglobina
del género Fusarium. Columna 1: F. oxysporum f. sp. lycopersici,
columna 2: F. verticillinoides; columna 3: F. graminearum.
80
Tabla 8. Códigos de los oligonucleótidos diseñados sobre los genes
codificadores de enzimas de tipo flavohemoglobina de F. oxysporum
f. sp. lycopersici y utilizados en reacciones de PCR para detectar los
genes ortólogos en F. oxysporum f. sp. phaseoli.
81
Tabla 9. Porcentajes de identidad entre las secuencias de nucleótidos de las
regiones codificantes de los genes FHG1, FHG2, FHG3 y FHG4.
90
Tabla 10. Ubicación de las hipotéticas cajas TATA y CAAT en la región
promotora y secuencias consenso señal de poliadenilación
reconocidos en los genes FHG1, FHG2, FHG3 y FHG4.
91
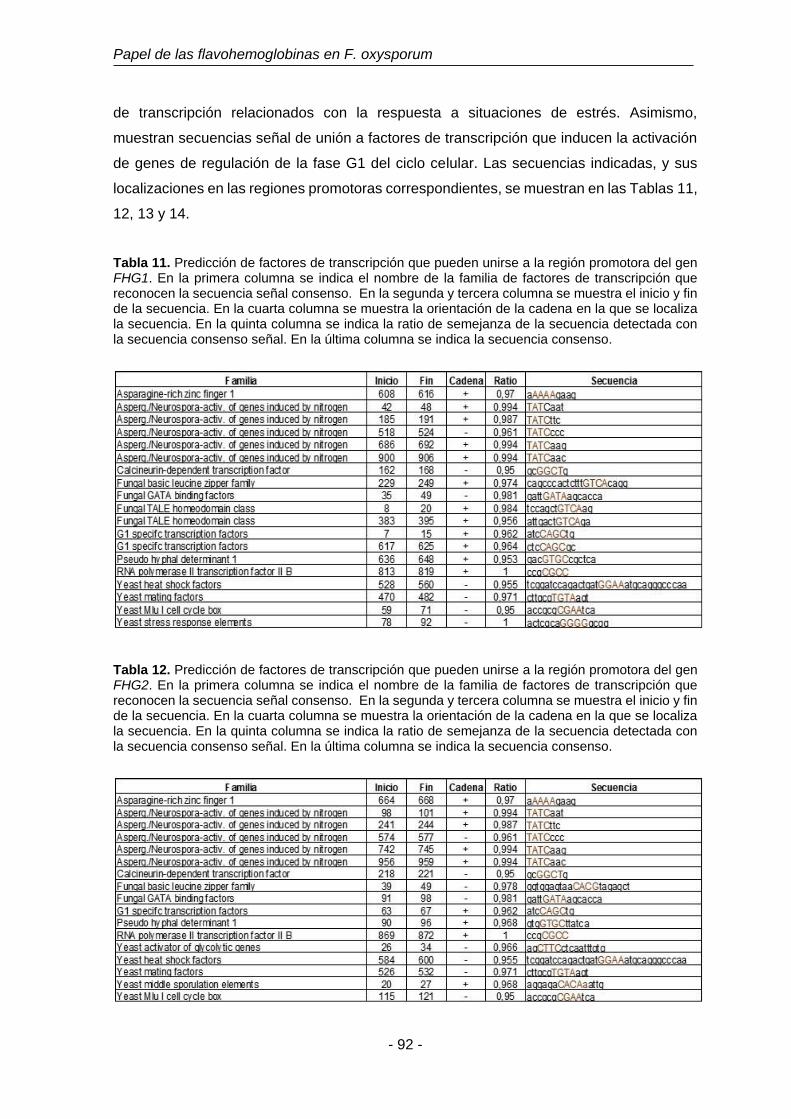
Tabla 11. Predicción de factores de transcripción que pueden unirse a la
región promotora del gen FHG1.
92
Tabla 12. Predicción de factores de transcripción que pueden unirse a la
región promotora del gen FHG2.
92
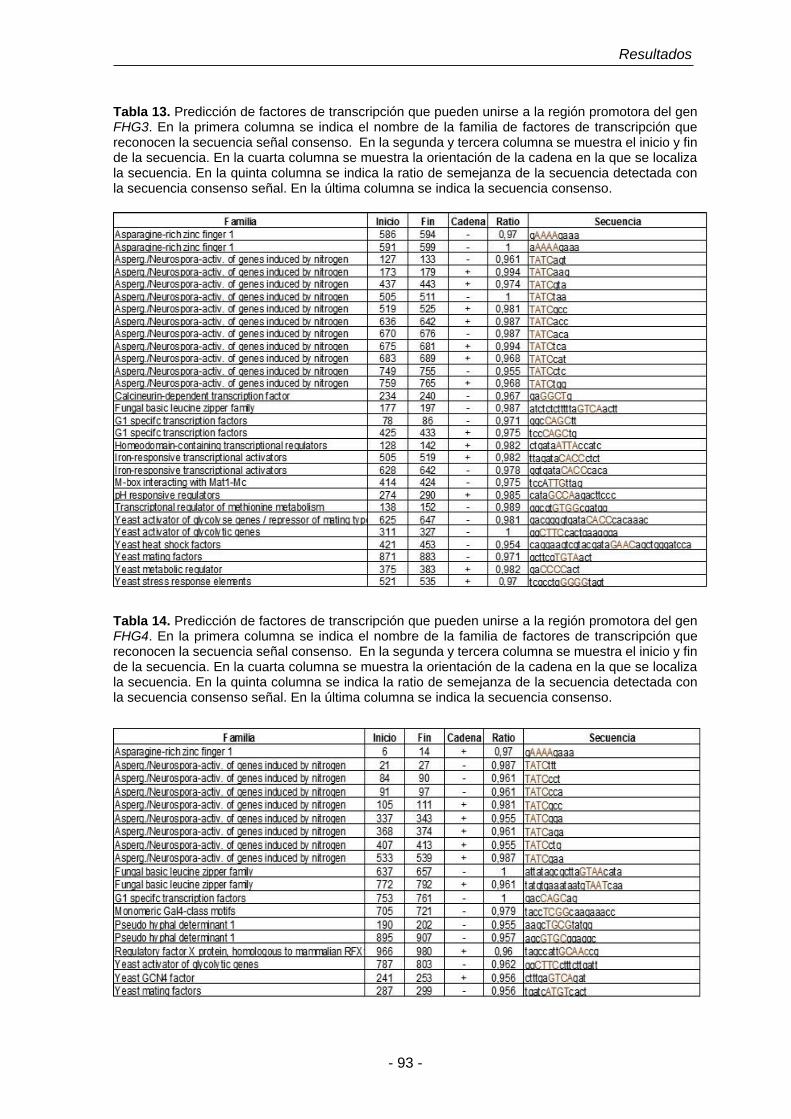
Tabla 13. Predicción de factores de transcripción que pueden unirse a la
región promotora del gen FHG3.
93
Papel de las flavohemoglobinas en F. oxysporum
XVI
Tabla 14. Predicción de factores de transcripción que pueden unirse a la
región promotora del gen FHG4.
93
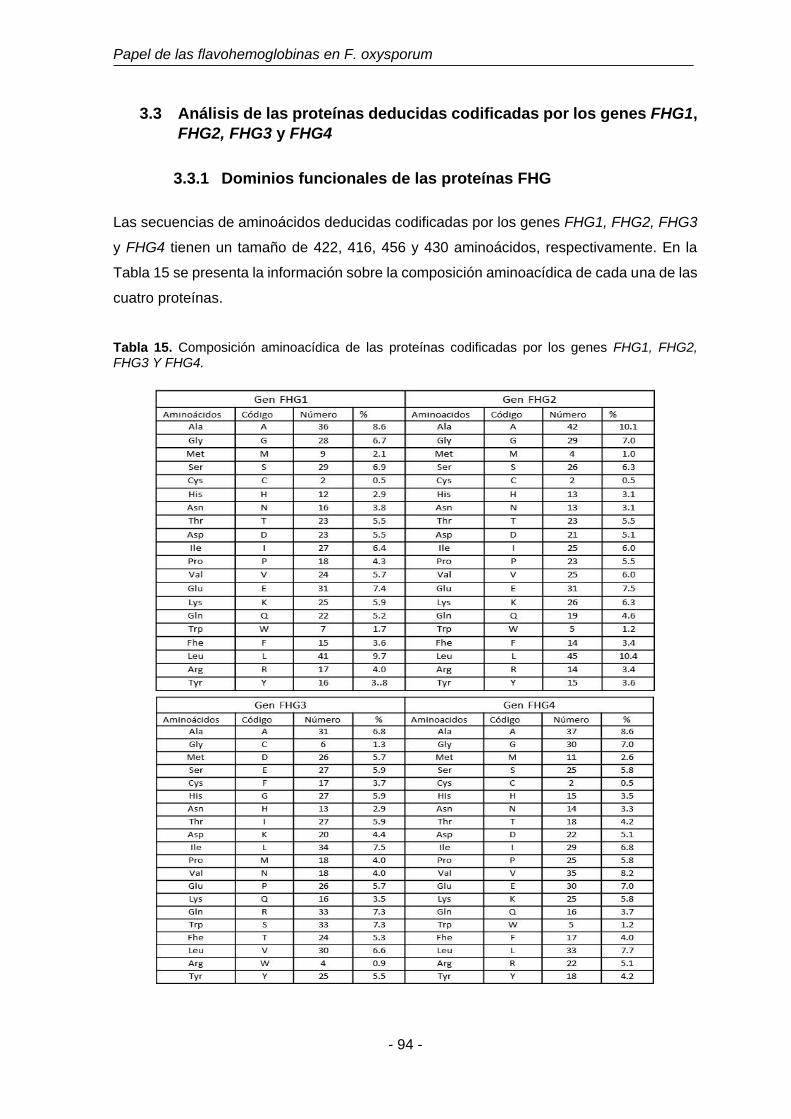
Tabla 15. Composición aminoacídica de las proteínas codificadas por los
genes FHG1, FHG2, FHG3 y FHG4.
94
Tabla 16. Porcentaje de semejanza de las secuencias aminoacídicas
deducidas de las proteínas codificadas por los genes FHG1, FHG2,
FHG3 y FHG4 de FOP-SP1.
95
Tabla 17. Dominios funcionales presentes en las proteínas FHG1, FHG2,
FHG3 y FHG4.
97
Tabla 18. Valores de probabilidad de localización subcelular de las proteínas
FHG estimados con el programa de predicción MultiLoc2.
99
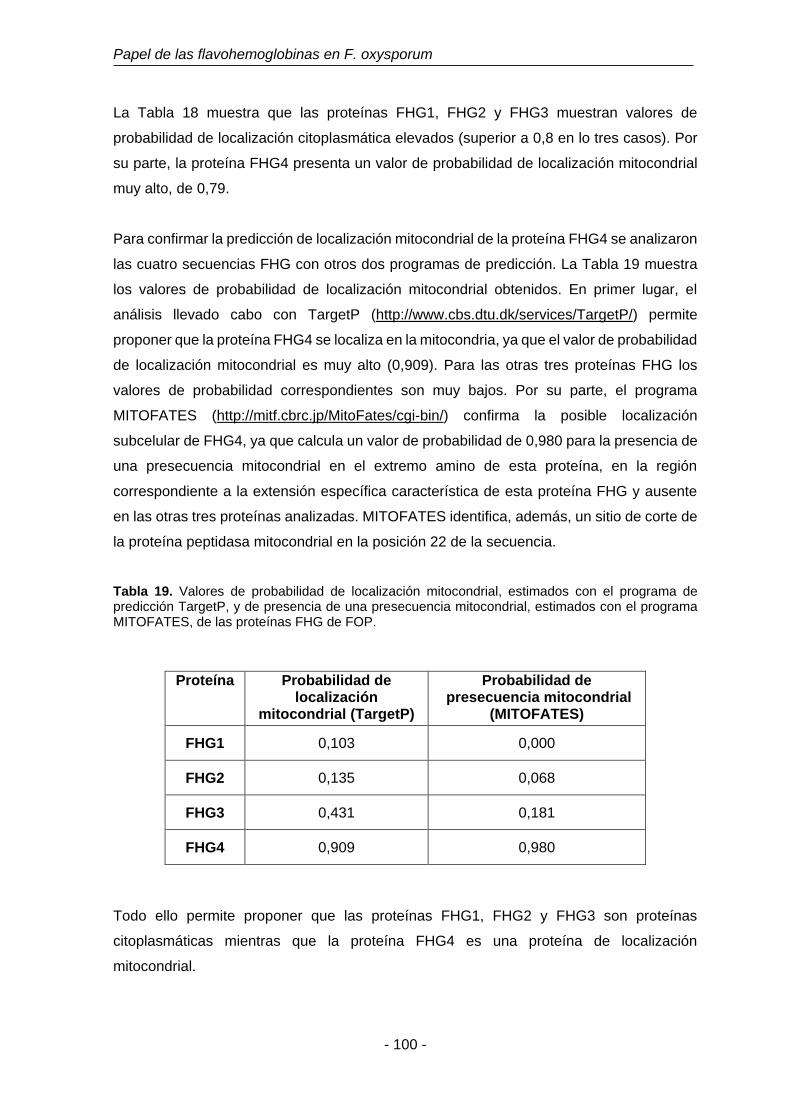
Tabla 19 Valores de probabilidad de localización mitocondrial, estimada con
el programa de predicción TargetP, y de presencia de una
presecuencia mitocondrial, estimada con el programa MITOFATES,
de las proteínas FHG de FOP.
100
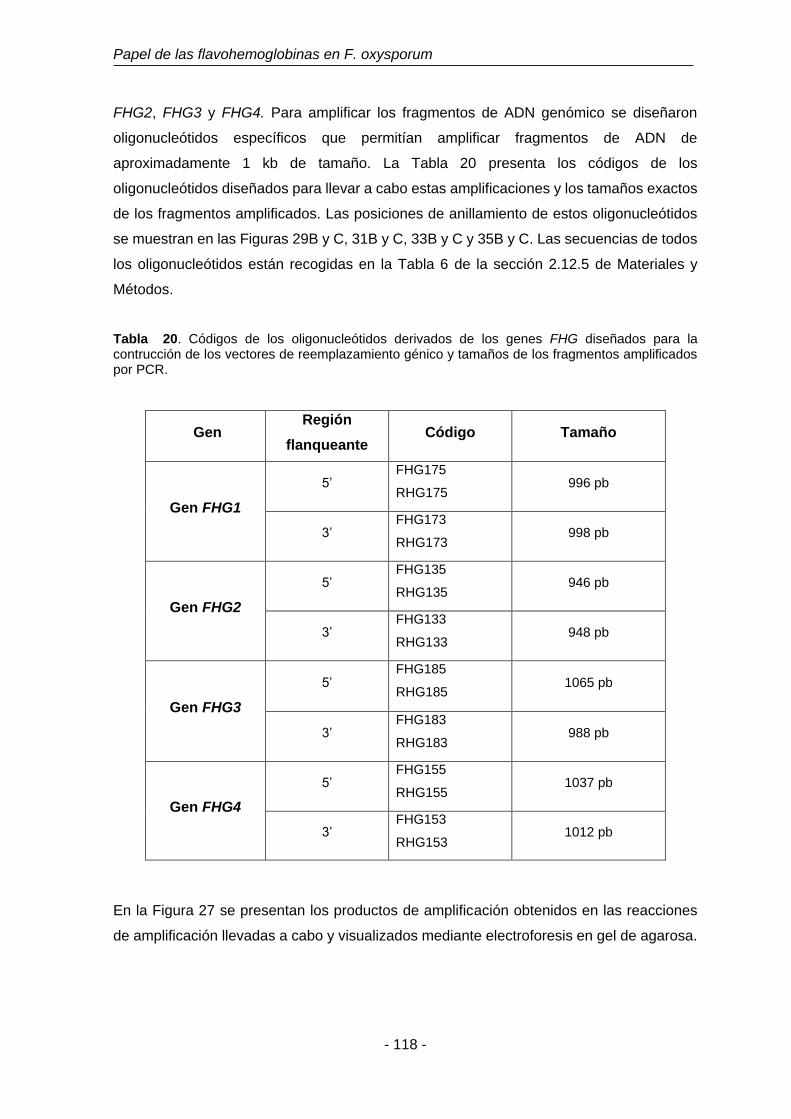
Tabla 20. Códigos de los oligonucleótidos derivados de los genes FHG
diseñados para la construcción de los vectores de reemplazamiento
génico y tamaños de los fragmentos amplificados por PCR.
118
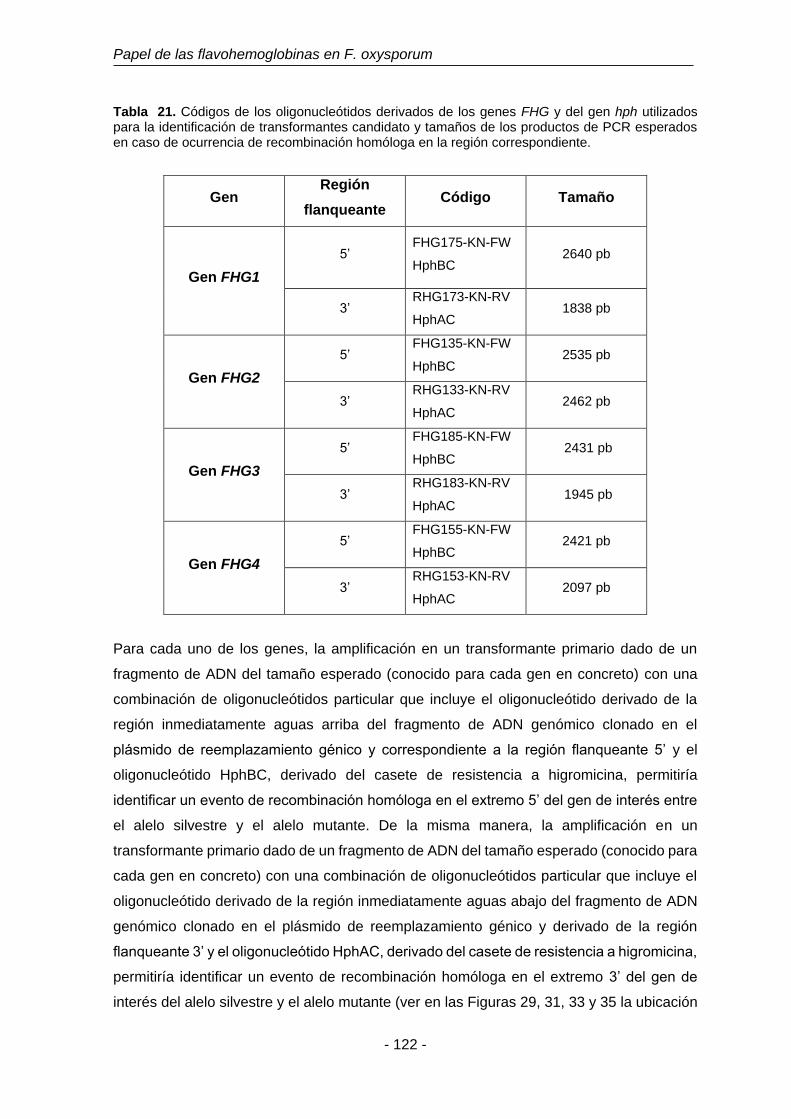
Tabla 21. Códigos de los oligonucleótidos derivados de los genes FHG y del
gen hph utilizados para la identificación de transformantes candidato
y tamaños de los productos de PCR esperados en caso ocurrencia
de recombinación homóloga en la región correspondiente.
122
Tabla 22. Estirpes silvestres y cepas mutantes ΔFHG generadas en este
trabajo y seleccionadas para su análisis fisiológico.
139
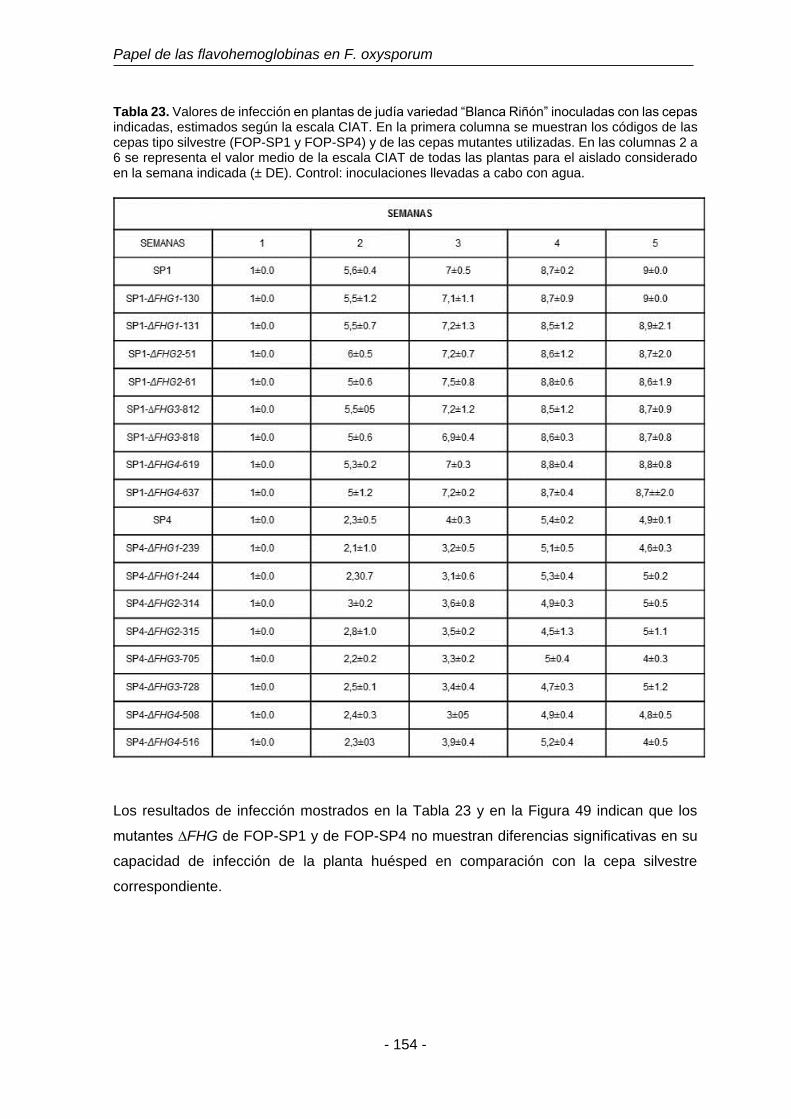
Tabla 23. Valores de infección en plantas de judía variedad “Blanca Riñón”
con las cepas mutantes ΔFHG de FOP estimados con la escala
CIAT.
154
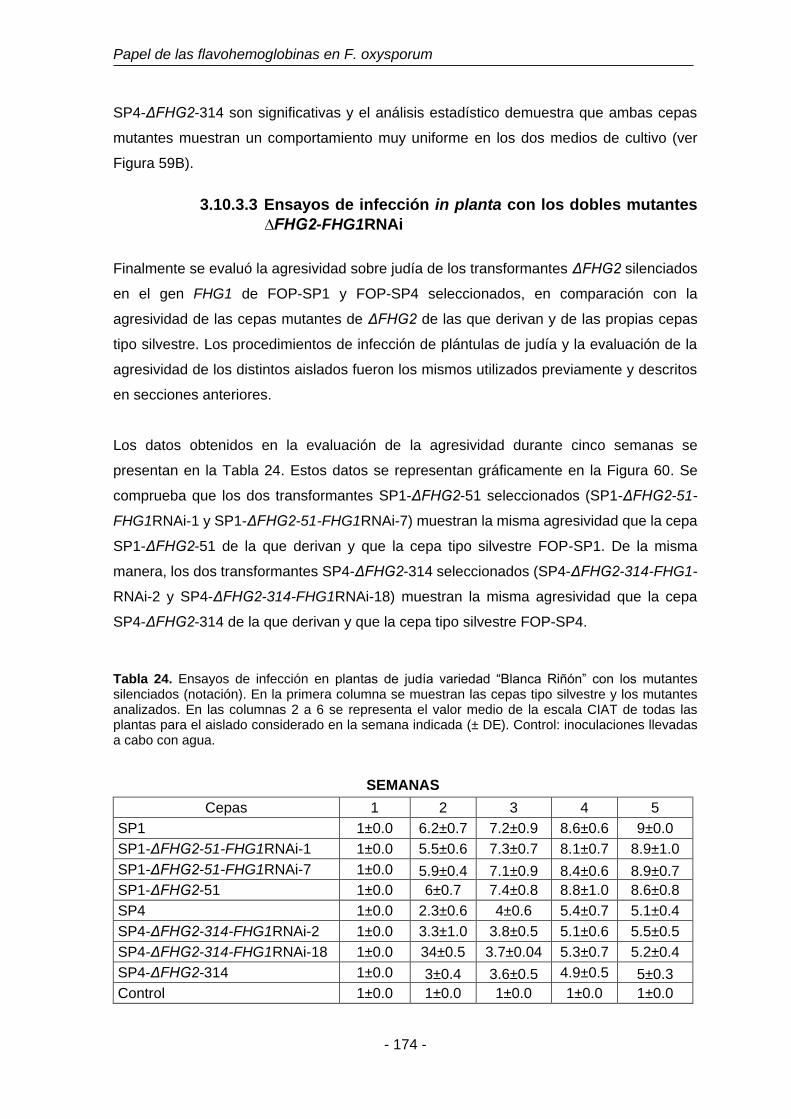
Tabla 24. Ensayos de infección en plantas de judía variedad “Blanca Riñón”
con los mutantes dobles. Evaluada con la escala CIAT.
y en el subdominio de unión a NAD el glutamato (Glu) 388 está muy conservado (Gardner
et al., 2005). La Figura 2 muestra la estructura tridimensional de la hmp de E. coli derivada
de datos experimentales obtenidos mediante rayos X (Ilari et al., 2002).
Papel de las flavohemoglobinas en F. oxysporum
- 28 -
Figura 2. Estructura tridimensional de la flavohemoglobina de E. coli. El dominio globina se muestra en color rojo. El dominio de oxidoreducción se representa en color turquesa (Ermler et al., 1995).
1.7.3 Funciones de las flavohemoglobinas
Las proteínas flavohemoglobinas (FHG´s) se encuentran ampliamente distribuidas en
microorganismos procariotas y eucariotas. En bacterias las proteínas FHG´s están
relacionadas con la eliminación del oxido nítrico (NO). En hongos el papel funcional de las
flavohemoglobinas aún es una cuestión controvertida (Cassanova et al., 2005; Smagghe
et al., 2008). La mayoría de las investigaciones apuntan a que las funciones de las FHG´s
en hongos están relacionadas con el metabolismo del NO, estrés nitrosativo y oxidativo,
señalización celular, germinación, conidiación y desarrollo sexual (Cassanova et al., 2005;
Smagghe et al., 2008; Marcos et al., 2016).
1.7.3.1 Respuesta de las flavohemoglobinas al estrés nitrosativo
y oxidativo
El primer gen codificador de una enzima FHG fue identificado en la bacteria E. coli (gen
hmp) (Vasudevan et al., 1991). La caracterización molecular y funcional de la enzima fue
planteada una decada más tarde, describiéndose como una dioxigenasa de NO
dependiente de NADPH, FAD y O2, cuya función es proteger a la bacteria del estrés
Introducción
- 29 -
nitrosativo mediante la conversión del NO en nitrato (NO3) (Gardner et al., 1998; Hausladen
et al., 1998). En condiciones aeróbicas el gen hmp es inducido por NO (Gardner et al.,
1998; Hausladen et al., 1998; Membrillo-Hernandez et al., 1998). En condiciones
anaeróbicas la flavohemoglobina de E. coli consume NO generando N2O con la
participación de NADH (Durner et al., 1999). Los mutantes de la FHG de E. coli no sólo
acumulan mayor cantidad de NO, sino que son incapaces de producir NO detectable,
limitación que se correlaciona con una reducción de la actividad de la enzima nitrito
reductasa responsable de la producción de NO (Corker y Poole, 2003).
En la bacteria Salmonella typhimurium se ha demostrado que los mutantes deficientes de
la flavohemoglobina (hmp) son capaces de producir NO tanto aeróbicamente como
anaeróbicamente, lo que implica que esta flavohemoglobina interviene en el control de la
concentración de NO (Gilberthorpe y Poole, 2008).
En la bacteria Bacillus subtilis el gen hmp es inducido bajo condiciones aeróbicas y
anaeróbicas en respuesta a tratamientos con NO (Moore et al., 2004). Sin embargo, este
mismo estudio muestra que la respuesta al estrés del NO gas y del donador SNP son
diferentes. A pesar de que ambos tratamientos inducen el regulón σβ, el NO gas lo hace
mediante la vía energética y el SNP mediante la vía de estrés ambiental. La diferencia de
la respuesta trascripcional dependiendo de la fuente de NO ha sido también observada en
S. cerevisiae (Sarver y DeRisi, 2005). Este fenómeno es debido probablemente a la
cantidad de NO liberada por el donador y el tiempo de exposición a NO que tiene cada
donador determinado por su vida media (Fitzhugh and Keefer, 2000).
La flavohemoglobina de S. cerevisiae (yhb1) protege a la levadura del estrés nitrosativo
producido por el NO en condiciones aeróbicas y anaeróbicas. También se ha demostrado
que yhb1 facilita la denitrosilación de proteínas. Se ha comprobado que los mutantes son
sensibles a estrés nitrosativo y que muestran un incremento en la concentración de las
proteínas S-nitrosiladas al ser comparados con las cepas silvestres (Liu et al., 2000). La
flavohemoglobina yhb1 se localiza en dos compartimentos celulares distintos: el citosol y
la matriz mitocondrial (Cassanova et al., 2005). La localización mitocondrial de yhb1 tiene
implicaciones en el control de la cadena respiratoria, ya que el NO que degrada es capaz
de inhibir la citocromo C oxidasa y la respiración (Boveris et al., 2000; Szibor et al., 2001)
y de reaccionar con el superóxido para formar peroxinitrito (Cadenas et al., 2000). De esta
forma, en ausencia de la flavohemoglobina los niveles de NO se incrementan (Liu et al.,
2000), se inhibe la citocromo C oxidasa, se incrementa la producción de superóxido y,
Papel de las flavohemoglobinas en F. oxysporum
- 30 -
consecuentemente, los niveles de peroxinitrito. Así, la levadura puede reducir los niveles
de superóxido no sólo mediante la superóxido dismutasa sino reduciendo su formación vía
cadena respiratoria al eliminar NO mediante la flavohemoglobina. De este modo, yhb1 es
esencial para la supervivencia de la célula bajo condiciones de estrés oxidativo y nitrosativo
(Liu et al., 2000). La distribución de la flavohemoglobina entre el citosol y la matriz
mitocondrial se regula diferencialmente dependiendo de las condiciones de anoxia y del
tipo de genoma mitocondrial, permitiendo de manera indirecta la eliminación del estrés
oxidativo (Cassanova et al., 2005). Las proteínas localizadas en uno y otro compartimiento
subcelular son codificadas por el mismo gen, ya que la mutación de yhb1 elimina ambas
proteínas y la reintroducción del gen restaura la aparición de la proteína en el citosol y en
la matriz mitocondrial (Cassanova et al., 2005). La flavohemoglobina de S. cerevisiae
(yhb1) muestra expresión constitutiva que no se ve incrementada con tratamientos con el
donador de NO DETA/NO en cultivos logarítmicos o estacionarios bajo condiciones
aeróbicas o anaeróbicas (Liu et al., 2000). Por el contrario, la respuesta de S. cerevisiae al
tratamiento con DPTA/NONOate incrementa la expresión de yhb1 unas 10 veces (Sarver
y DeRisi, 2005). Este comportamiento puede ser debido a las diferencias en los donadores
de NO, medios de cultivo, cepas de levadura, sensibilidad del método de medida o
condiciones de cultivo utilizadas. La expresión de yhb1 tampoco es significativamente
afectada por químicos que imponen estrés oxidativo como el H2O2, paraquat y menadiona
(Buisson and Labbe-Bois, 1998). Sin embargo, la expresión de yhb1 es menor en cultivos
anaeróbicos de la levadura que en aeróbicos (Liu et al., 2000).
C. albicans posee tres genes que codifican proteínas de tipo flavohemoglobina, pero
sólamente Cayhb1 es inducido por NO y es el responsable de la detoxificación del NO
producido por los macrófagos. Los mutantes delecionados del gen Cayhb1 son
hipersensibles a NO (DETA) (Ullman et al., 2004). En A. oryzae y A. nidulans se ha
reportado la presencia de dos genes tipo FHG. La flavohemoglobina FHb1 de A. oryzae, al
igual que la flavohemoglobina fhbA de A. nidulans, están presentes en el citosol y muestran
altos niveles de expresión en medios suplementados con NO3, siendo mayor la expresión
en medios con un pH ligeramente ácido. Ambos genes (FHb1 y fhbA) codifican las
flavohemogloginas responsables de proteger al hongo del estrés nitrosativo mediante la
conversión del NO a NO3 (Schinko et al., 2010). Las flavohemoglobinas FHb2 y fhbB de A.
oryzae y de A. nidulans, respectivamente, han sido localizadas en la matriz mitocondrial y
su función aún resulta una cuestión controvertida. Ambas muestran una secuencia señal
mitocondrial y probablemente están implicadas en la detoxificación de NO en la mitocondria
(Zhou et al., 2009; Schinko et al., 2013). Schinko et al. (2010) y Marcos et al. (2016)
Introducción
- 31 -
reportaron que la expresión del gen fhbA de A. nidulans es inducido por el gen NirA y no
por AreA. Sin embargo, en el promotor del gen fhbA se localizan cuatro secuencias señal
de unión para AreA. Los autores demuestran que AreA regula la expresión de fhbA durante
la fase de conidiación y no durante la fase de desarrollo (Schinko et al., 2010; Marcos et
al., 2016). También se ha demostrado que fhbA es inducido en presencia de elevadas
concentraciones de NO2 y pH ligeramente ácido (Schinko et al., 2010). En A. fumigatus la
flavohemoglobina fhbA protege al hongo del estrés nitrosativo producido por el NO,
mostrando altos niveles de expresión sobre nitrato y nitrito. Los mutantes delecionados del
gen fhbA muestran un incremento en la sensibilidad a DETA-NO comparado con la cepa
de tipo silvestre, indicando que la flavohemoglobina fhbA juega un rol importante en la
defensa contra las especies reactivas de nitrógeno (Lapp et al., 2014).
En B. cinerea la flavohemoglobina (BCFHG1) es la proteína responsable de la
detoxificación de NO, protegiendo al hongo del estrés nitrosativo. Los mutantes con pérdida
de función de BCFHG1 son sensibles a NO (Turrión-Goméz and Benito, 2011). Bcfhg1 se
expresa en todas las fases de desarrollo en ausencia de NO exógeno, mostrando un
máximo de expresión durante la ventana termporal de germinación (entre las 4 y las 6 h de
culvito en medio mínimo líquido), y la exposición a NO exógeno determina un aumento de
sus niveles de expresión (Turrión-Gómez et al., 2010).
1.7.3.2 Papel de las flavohemoglobinas en patogenicidad
Las flavohemoglobinas juegan un papel importante en los organismos patógenos,
protegiéndoles del estrés nitrosativo creado por el hospedero como mecanismo de defensa
(Arasimowicz‐Jelonek and Floryszak‐Wieczorek, 2014). Así se ha comprobado en la
bacteria fitopatógena Erwinia chrysanthemi, sistema en el que la flavohemoglobina
codificada por el gen HmpX es inducida durante la infección y es esencial para el desarrollo
de la bacteria (Boccara et al., 2005), ya que inactiva el NO y previene la inhibición de la
respiración celular que éste causa. Pero además, la flavohemoglobina del patógeno puede
condicionar la respuesta del huésped. Así, en plantas de Saintpaulia ionantha inoculadas
con mutantes deficientes en el gen HmpX de E. chrysantemi se acumulan altos niveles de
NO y se observa activación del programa de muerte celular (Boccara et al., 2005). Las
plantas transgénicas de Arabidopsis que expresan el gen hmp de E. coli muestran una
reducción en los niveles de NO acumulado in planta y una atenuación del choque de NO
producido por el patógeno (Zeier et al., 2004). Tomando todos estos datos en su conjunto
Papel de las flavohemoglobinas en F. oxysporum
- 32 -
podemos deducir que las flavohemoglobinas son unos potentes detoxificadores del NO que
reducen la Respuesta Hipersensible (HR) en plantas y el estrés nitrosativo en el patógeno.
La flavohemoglobina (yhb1) de C. neoformans es esencial para la virulencia del hongo en
ratones al degradar el NO producido por la iNOS, ya que la virulencia de los mutantes
deficientes en el gen YHB1 disminuye en animales de tipo silvestre y se reestablece en
animales iNOS-/- (de Jesus-Berrios et al., 2003). Es interesante destacar que en el mismo
trabajo se indica que el estudio y caracterización de dobles mutantes de la
flavohemoglobina y de la superóxido dismutasa, responsable esta última de la protección
de C. neoformans al estrés oxidativo, demuestran un efecto sinérgico de ambos sistemas
en la reducción de la virulencia sobre ratones.
La reducción de vilurencia de los mutantes de flavohemoglobina se ha descrito en C.
albicans (Hromatka et al., 2005; Ullmann et al., 2004). Como hemos indicado
anteriormente, en el genoma de C. albicans se han identificado tres genes FHG, pero sólo
uno, Cayhb1, es inducido por NO y responsable de la degradación de NO. La eliminación
de este gen determina una reducción de la virulencia (Ullmann et al., 2004). Sin embargo,
los mutantes del gen Cayhb1 de C. albicans no recuperan la virulencia en mutantes de
ratón carentes de NOS2, lo que supone que en el modelo de investigación utilizado la
producción de NO por parte del huésped no es un determinante fundamental de la
virulencia del patógeno y que la reducción de virulencia observada en los mutantes
analizados debe ser consecuencia de algún otro proceso de naturaleza aún por determinar
(Hromatka et al., 2005).
En la bacteria patógena humana S. enterica su flavohemoglobina es necesaria para la
infección de ratones, restaurándose la virulencia en mutantes de ratones carentes de la
iNOS (Bang et al., 2006).
1.7.3.3 Papel de las flavohemoglobinas en la regulación del
desarrollo
Varios estudios realizados con proteínas tipo flavohemoglobina de microorganismos
procariotas y eucariotas demuestran que estas proteínas además de estar implicadas en
la protección frente al estrés nitrosativo producido por el hospedador infectado, también
están implicadas en funciones de desarrollo y crecimiento. Investigaciones realizadas por
Membrillo-Hernández et al. (2009) mostraron que el gen hmp de E. coli regula el
Introducción
- 33 -
crecimiento de la bacteria en la fase estacionaria, ya que los mutantes deficientes en hmp
muestran un ligero retraso en el crecimiento (Membrillo-Hernández et al., 1999). En C.
albicans se ha mostrado que el producto génico de CaYHB1 regula el desarrollo de la
levadura, puesto que los mutantes deficientes en este gen muestran un desarrollo
hiperfilamentoso (Hromatka et al., 2005). El producto génico del gen fHb1 de A. oryzae
regula el desarrollo de hifas, ya que los mutantes deficientes en FHb1 presentan
alteraciones morfológicas en el micelio (Zhou et al., 2009). En A. nidulans la
flavohemoglobina FhbA regula la conidiación y el desarrollo de hifas. El gen fhbA es
fuertemente expresado en la fase inicial del desarrollo y su expresión decrece luego
gradualmente, mientras que fhbB se expresa de forma moderada en la fase inicial del
desarrollo y su función parece estar más relacionada con la regulación de la transición de
la fase de crecimiento vegetativo a conidiación. Los mutantes deficientes en fhbA y fhbB
muestran un crecimiento superficial y anormal del micelio (Marcos et al., 2016). También
se ha reportado que el gen fhbA regula la formación de cuerpos de fructificación (Baidya et
al., 2011).
En C. coccodes utilizando aproximaciones farmacológicas se ha demostrado que el NO
juega un papel importante durante la germinación, ya que el exceso de NO retrasa la
germinación y el uso de secuestradores de NO acelera la germinación, lo que supone que
el NO participa en los procesos fisiológicos de germinación y desarrollo (Wang and Higgins,
2005). En C. minitans la adición de NO exógeno inhibe la formación de conidios (Gong et
al., 2007). El papel del NO en la formación y desarrollo de estructuras reproductivas y
esporas también se ha evidenciado en N. crassa (Niennemann and Maier, 1996), P.
blakesleeanus (Maier et al., 2001) y Flammulina velutipes (Song et al., 2000). La ameba
Dictyostelium discoideum posee dos genes que codifican proteínas tipo flavohemoglobina
(DdFHa y DdFHb) cuya función está relacionada con la protección a estrés nitrosativo,
metabolismo de la glucosa, crecimiento y desarrollo celular. Los mutantes deficientes de
DdFHa y DdFHb muestran un marcado incremento en el tamaño de la célula, reducida
viabilidad bajo altas concentraciones de glucosa y sensibilidad a estrés nitrosativo. La
expresión tanto de DdFHa como de DdFHb es inducida bajo condiciones sumergidas,
incrementándose sus niveles de expresión de 3 a 4 veces en relación con los niveles
observados en las células utilizadas como control (Lijima et al., 2000).
Papel de las flavohemoglobinas en F. oxysporum
- 34 -
OBJETIVOS
Objetivos
- 35 -
OBJETIVOS
Las proteínas de tipo flavohemoglobina están ampliamente distribuidas en
microorganismos procariotas y eucariotas. Las evidencias experimentales acumuladas
demuestran que éstas constituyen uno de los principales sistemas enzimáticos de
detoxificación de NO en estos grupos de organismos. Mediante esta investigación
pretendemos determinar cuáles son las funciones fisiológicas en las que participan las
proteínas de tipo flavohemoglobina en el hongo fitopatógeno F. oxysporum f. sp. phaseoli.
Son objetivos del presente trabajo los siguientes:
1. Identificación de genes codificadores de enzimas de tipo flavohemoglobina, genes
FHG, en las estirpes SP1, muy virulenta, y SP4, poco virulenta, del F. oxysporum f.
sp. phaseoli.
2. Determinación de los patrones de expresión de los genes FHG durante el
crecimiento saprofítico en las estirpes SP1 y SP4 del patógeno en respuesta a NO
exógeno y durante su interacción con la planta hospedera.
3. Obtención de mutantes deficientes en los genes FHG en ambas estirpes de F.
oxysporum f. sp. phaseoli.
4. Caracterización funcional y fisiológica de los mutantes deficientes en los genes FHG
de ambas estirpes de F. oxysporum f. sp. phaseoli durante el crecimiento saprofitico
y durante la interacción con la planta hospedera.
Papel de las flavohemoglobinas en F. oxysporum
- 36 -
MATERIALES Y MÉTODOS
Materiales y Métodos
- 37 -
2 MATERIALES Y MÉTODOS
2.1 Organismos
2.1.1 Plantas
Para los ensayos de infección se utilizaron plantas de judía (P. vulgaris L.) de la variedad
“blanca riñón”.
2.1.2 Hongos
En la Tabla 4 se muestran las estirpes de F. oxysporum utilizadas en esta investigación,
las mismas que fueron colectadas por nuestro laboratorio de plantas de judía (P. vulgaris)
infectadas en la zona de El Barco de Ávila (Alves-Santos et al., 1999), y conservadas a -
80ºC.
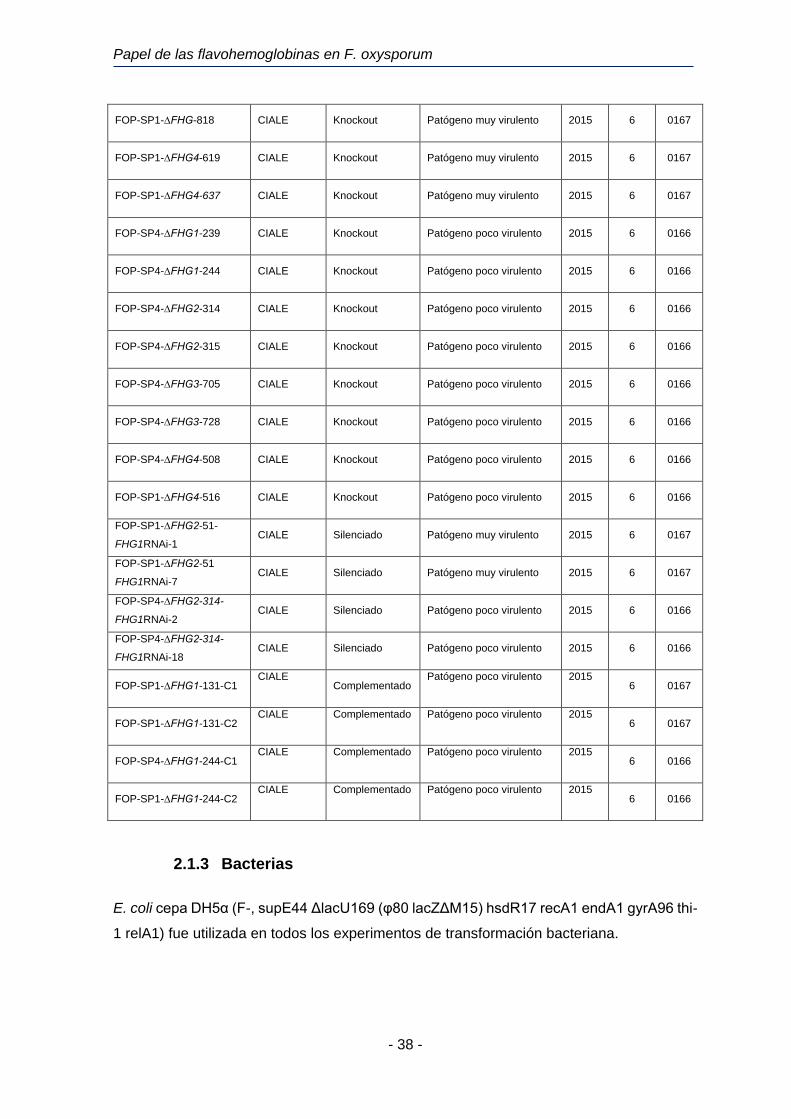
Tabla 4. Estirpes silvestres y transformantes de F. oxysporum utilizadas en esta investigación
Aislado Origen Genotipo Patogenicidad Año Raza VCG
FOP-SP1 La Carrera
(CIALE) Silvestre
Patógeno muy virulento 1993 6 0167
FOP-SP4 La Carrera
(CIALE) Silvestre
Patógeno poco virulento 1994 6 0166
FOP-SP13 Villafranca
(CIALE) Silvestre
Patógeno súper virulento 6 0169
AB82 El Barco de
Ávila
(CIALE)
Silvestre No patógeno 1995 -- 0167
4287 (F. oxysporum f. sp.
lycopersici)
J. Tello
(INIA) Silvestre Patógeno muy virulento 2 0030
FOP-SP1-∆FHG1-130 CIALE Knockout Patógeno muy virulento 2015 6 0167
FOP-SP1-∆FHG1-131 CIALE Knockout Patógeno muy virulento 2015 6 0167
FOP-SP1-∆FHG2-51 CIALE Knockout Patógeno muy virulento 2015 6 0167
FOP-SP1-∆FHG2-61 CIALE Knockout Patógeno muy virulento 2015 6 0167
FOP-SP1-∆FHG3-812 CIALE Knockout Patógeno muy virulento 2015 6 0167
Papel de las flavohemoglobinas en F. oxysporum
- 38 -
FOP-SP1-∆FHG-818 CIALE Knockout Patógeno muy virulento 2015 6 0167
FOP-SP1-∆FHG4-619 CIALE Knockout Patógeno muy virulento 2015 6 0167
FOP-SP1-∆FHG4-637 CIALE Knockout Patógeno muy virulento 2015 6 0167
FOP-SP4-∆FHG1-239 CIALE Knockout Patógeno poco virulento 2015 6 0166
FOP-SP4-∆FHG1-244 CIALE Knockout Patógeno poco virulento 2015 6 0166
FOP-SP4-∆FHG2-314 CIALE Knockout Patógeno poco virulento 2015 6 0166
FOP-SP4-∆FHG2-315 CIALE Knockout Patógeno poco virulento 2015 6 0166
FOP-SP4-∆FHG3-705 CIALE Knockout Patógeno poco virulento 2015 6 0166
FOP-SP4-∆FHG3-728 CIALE Knockout Patógeno poco virulento 2015 6 0166
FOP-SP4-∆FHG4-508 CIALE Knockout Patógeno poco virulento 2015 6 0166
FOP-SP1-∆FHG4-516 CIALE Knockout Patógeno poco virulento 2015 6 0166
FOP-SP1-∆FHG2-51-
FHG1RNAi-1 CIALE Silenciado Patógeno muy virulento 2015 6 0167
FOP-SP1-∆FHG2-51
FHG1RNAi-7 CIALE Silenciado Patógeno muy virulento 2015 6 0167
FOP-SP4-∆FHG2-314-
FHG1RNAi-2 CIALE Silenciado Patógeno poco virulento 2015 6 0166
FOP-SP4-∆FHG2-314-
FHG1RNAi-18 CIALE Silenciado Patógeno poco virulento 2015 6 0166
FOP-SP1-∆FHG1-131-C1 CIALE
Complementado Patógeno poco virulento 2015
6 0167
FOP-SP1-∆FHG1-131-C2 CIALE Complementado Patógeno poco virulento 2015
6 0167
FOP-SP4-∆FHG1-244-C1 CIALE Complementado Patógeno poco virulento 2015
6 0166
FOP-SP1-∆FHG1-244-C2 CIALE Complementado Patógeno poco virulento 2015
reacción de defosforilación se realizó en un volumen final de 10 µl, con 1 µg de ADN
digerido, 1 µl de tampón 10X y 2 µl de fosfatasa alcalina CIP (Roche) diluida hasta
0,01U/pmol (0,5 M Tris-HCl pH 9,0; 10 mM MgCl2; 1 mM ZnCl2 y 10 mM espermicida) y agua
MiliQ. Se incubó a 37ºC durante 30 minutos. La desactivación de la enzima se realizó
mediante incubación a 65ºC durante 15 minutos. Los fragmentos de ADN tratados fueron
purificados utilizando el kit comercial NucleoSpin® Gel and PR clean-up (Macherey-Nagel).
2.6.4 Obtención de ARN total de F. oxysporum
Para la obtención de ARN total de F. oxysporum se utilizaron los siguientes métodos: el
método Trizol (Invitrogen) y el método basado en la utilización del kit SV Total RNA Isolation
System (Promega).
2.6.4.1 Método Trizol
El Trizol es una solución mono-básica de fenol e isocianato de guanidina que durante la
homogeneización de la muestra mantiene la integridad del ARN al mismo tiempo que altera
la estabilidad de las células y rompe los componentes celulares. El volumen de la muestra
no debe exceder del 10 % del volumen del trizol. La adición de cloroformo seguida de
centrifugación separa la muestra en dos fases, una de ellas acuosa y la otra orgánica. El
ARN permanece exclusivamente en la fase acuosa y puede ser recuperado por precipitación
con alcohol isopropílico.
Las muestras de micelio o tejido vegetal congelado a -80ºC fueron trituradas en un mortero
con nitrógeno líquido. El micelio o tejido vegetal pulverizado fue transferido a un tubo
eppendorf de 1,5 ml, al que se añadió 1 ml de Trizol® Reagent (Ambion RNA life
technologies), mezclando cuidadosamente con una pipeta para homogeneizar la muestra.
Ésta fue incubada a temperatura ambiente durante 10 minutos para permitir la disociación
completa de las proteínas. A continuación la muestra fue centrifugada durante 10 minutos a
12.000 rpm y 4ºC y el sobrenadante fue transferido a un nuevo tubo que fue incubado
durante 5 minutos en hielo. Se añadieron 200 µl de cloroformo por cada ml de Trizol utilizado
y se agitó la muestra manualmente o en un vórtex durante 15 segundos, siendo incubada
posteriormente durante 5 minutos en hielo. A continuación se centrifugó durante 15 minutos
a 12.000 rpm y 4ºC. La muestra se separa ahora en dos fases, una fase roja que
Materiales y Métodos
- 53 -
corresponde al trizol-cloroformo y una fase incolora que corresponde a la fase acuosa. El
ARN permanece exclusivamente en la fase incolora.
La fase acuosa fue transferida a un nuevo tubo y se añadió 1 volumen de cloroformo/alcohol
isoamílico (24:1). El tubo se agitó en el vortex, fue centrifugado durante 10 minutos a 12.000
rpm y 4ºC y la fase acuosa fue transferida a un nuevo tubo eppendorf. Dependiendo de la
pureza y calidad del ARN que deseamos obtener el tratamiento con 1 volumen
cloroformo/alcohol isoamílico (24:1) se puede repetir 1 o 2 veces. Se añadieron al
sobrenadante 250 µl de isopropanol y 250 µl de citrato de sodio (citrato de sodio 0,8 M y
NaCl 1,2 M) y se incubó la mezcla durante 10 minutos o toda la noche. Pasado este tiempo
se centrifugó durante 15 minutos a 12.000 rpm y 4ºC. Se eliminó la fase acuosa y se añadió
1 ml de etanol 70 %. La suspensión fue centrifugada durante 5 minutos a 7.500 rpm y 4ºC
y el sedimento resultante se resuspendió en 25 a 50 µl de agua MiliQ tratada con DEPC 0,1
%. El ARN así preparado se conservó a -80ºC hasta su utilización.
2.6.4.2 Obtención de ARN con el kit comercial SV Total RNA
Isolation System (Promega)
El Kit comercial SV Total RNA Isolation System (Promega) permite obtener ARN de buena
calidad y pureza de forma rápida y sencilla. Se realizó la extracción de ARN de micelio o
tejido de planta siguiendo las recomendaciones del fabricante.
El micelio o tejido de planta conservados a -80ºC fueron triturados en un mortero con
nitrógeno líquido. El polvo resultante fue transferido a un tubo eppendorf de 1,5 ml con 175
µl de buffer de lisis de ARN (RLA+BME) y la muestra fue homogeneizada con ayuda de una
pipeta o por inversión. Se añadieron 350 µl de buffer de dilución de ARN (RDA, blue),
mezclando la muestra por inversión de 3 a 4 veces e incubando 3 minutos a 70ºC. Se
centrifugó 10 minutos a 13.000 rpm. La fase transparente fue transferida a un tubo
eppendorf con 200 µl de etanol 95 % (etanol absoluto 99 % + agua DEPC 0,1 %), se mezcló
la muestra con una pipeta para homogeneizar la muestra, se transfirió la suspensión a una
columna y ésta fue centrifugada durante 1 minuto a 13.000 rpm. Se eliminó el lisado y se
añadieron 600 µl de tampón de lavado de ARN (RWA+etanol 95 %). La columna se
centrifugó durante 1 minuto a 13.000 rpm, se eliminó el sobrenadante y se añadieron 50 µl
de DNAsa (40 µl de Yellow core buffer; 5 µl de MnCl2 0,09 M; 5 µl de DNAsa I), y se incubó
15 minutos a temperatura ambiente. La inactivación de la DNAsa se realizó añadiendo 200
Papel de las flavohemoglobinas en F. oxysporum
- 54 -
µl de solución DNAsa stop (DSA + etanol 95 %) y centrifugando durante 1 minuto a 13.000
rpm. El filtro con el ARN fue lavado 2 veces con 600 µl y 250 µl de tampón de lavado (RWA
+ etanol 95 %) y centrifugado durante 2 minutos a 13.000 rpm. El sedimento de ARN fue
recuperado en 50 µl de agua libre de RNAsas.
2.6.4.3 Tratamiento de ARN con DNAasa
El ARN fue tratado con DNAsa utilizando el kit DNA-free™ Kit TURBO™ DNase Treatment
and Removal Reagents (Ambion Life Technologies, USA). Las muestras fueron tratadas
con 1 µl de DNAsa a 37ºC durante 30 minutos. Transcurrido este tiempo se añadieron 5 µl
del inactivador de la DNAsa y las muestras se incubaron durante 2 minutos a temperatura
ambiente. Posteriormente fueron centifugadas durante 1 minuto a 13.000 rpm y el
sobrenadante fue transferido a un nuevo tubo eppendorf. El ARN se conservó a -80ºC.
2.6.4.4 Cuantificación de ácidos nucleicos
Para estimar la pureza del ADN se utilizó un espectrofotómetro NanoDrop N-1000. Los
valores de absorbancia de 1,8 a 2,0 a 260 nm y 280 nm, respectivamente, son indicativos
de ADN puro. Los valores inferiores indican la presencia de proteínas o de otros
contaminantes. Una segunda valoración de la pureza de ácidos nucleicos es la estimación
de la relación Absorbancia 260/ Absorbancia 230. Los valores de 2,0 a 2,2 indican pureza.
Una relación inferior supone la presencia de contaminantes como carbohidratos o fenol.
La integridad del ADN también se puede evaluar mediante electroforesis en gel de agarosa.
Si el ADN está integro, se debe observar una banda estrecha de alto peso molecular. Si
está fragmentado se observará una banda más o menos ancha y más o menos difusa, en
función del grado de degradación, pudiendo aparecer a lo largo de todo el carril de
electroforesis. El ADN fragmentado dificulta la amplificación de productos de PCR de alto
peso molecular, afectando la reproducibilidad de las técnicas.
2.7 Electroforesis de ácidos nucleícos
2.7.1 Electroforesis de ADN
Las electroforesis de ADN se realizaron en geles de agarosa de 0,6 al 2 % preparados en
tampón TAE 1X (40 mM Tris-acetato y 1 mM EDTA). Se utilizaron cubetas horizontales de
Materiales y Métodos
- 55 -
15 o 30 cm de largo, con tampón de electroforesis TAE 1X. El ADN se mezcló en proporción
6:1 con el tampón de carga de electroforesis (30 % glicerol; 0,25 % azul de bromofenol y
0,25 % xylene cyanol, Sigma). Se cargaron las muestras en el gel de agarosa incluyendo
en cada gel un pocillo para el marcador de peso molecular (1 kb Plus DNA ladder, Fisher).
Los geles se sometieron a campos eléctricos de 80 a 100 voltios dependiendo del rango de
tamaños a resolver y de la cantidad de ADN. Después de 45 a 60 minutos los geles se
tiñeron con bromuro de etidio en una concentración de 0,1 a 0,5 mg/ml en TAE 1X, durante
20 minutos. El ADN se visualizó por medio de luz ultravioleta y se captaron las imágenes
por medio del sistema Kodak de grabación digital de imágenes.
2.7.2 Electroforesis de ARN
Las electroforesis de ARN se realizaron en condiciones desnaturalizantes, siguiendo el
procedimiento descrito en Sambrook y colaboradores (1989) y las indicaciones mostradas
en el manual de marcaje de sondas DIG (Roche Molecular Biochemicals). Los geles de ARN
proporcionan la matriz y las condiciones desnaturalizantes para una correcta separación y
detección del ARN. Se prepararon geles de agarosa al 1 % en tampón MOPS 1X (20 mM
MOPS, 5 mM acetato sódico, 1 mM EDTA pH 7) con 2 % de formaldehído y 0,5 µg/ml de
bromuro de etidio.
Las muestras de ARN fueron ajustadas a una concentración de 1 µg/µl y se mezclaron en
2 volúmenes de tampón de carga para Northern Blot. Para 500 µl de tampón se añadieron
250 µl de formamida deionizada, 83 µl de formaldehído, 50 µl glicerol 100 %, 10 µl de azul
de bromofenol y 57 µl de agua DEPC 0,1 %. Las muestras se incubaron durante 10 minutos
a 65ºC para desnaturalizar el ARN, se incubaron en hielo y se depositaron en el gel. El
tampón de electroforesis utilizado fue MOPS 1x. Las muestras migraron a 40 voltios durante
4 horas en una campana de extracción. Debido a la poca conductibilidad eléctrica del
tampón MOPS, cada 15 minutos se removió el tampón del polo positivo al negativo. La
integridad del ARN se comprobó en el transluminador de luz ultravioleta.
2.7.3 Purificación de fragmentos de ADN de geles de agarosa
Las muestras de ADN a ser recuperadas fueron preparadas en gel de agarosa del 0,7 % en
TAE 1X (la concentración puede variar en función del tamaño del fragmento que se desea
purificar). Las condiciones de la electroforesis se realizaron garantizando la máxima
resolución y separación del fragmento a ser recuperado. Una vez que el gel había sido
Papel de las flavohemoglobinas en F. oxysporum
- 56 -
teñido con bromuro de etidio se procedió a recuperar el fragmento deseado cortando la
sección del gel de agarosa correspondiente, que fue depositada en un tubo eppendorf de
1,5 ml. El fragmento recuperado fue purificado con el kit comercial NucleoSpin® Gel and PR
clean-up siguiendo las recomendaciones del fabricante.
2.8 Vectores de clonacion
2.8.1 Vector de clonación pGEM-T Easy
El vector de clonación pGEM-T Easy (Promega) linearizado se utilizó para la clonación de
fragmentos amplificados por PCR siguiendo las especificaciones de la casa comercial.
pGEM-T Easy se caracteriza por tener una desoxitimidina 3` terminal en ambos extremos,
característica que mejora la eficiencia de clonación de productos de PCR debido a que las
polimerasas termoestables frecuentemente añaden un residuo de desoxiadenosina de
forma no dependiente de molde en los extremos 3` de los fragmentos amplificados. Posee
el gen de resistencia a ampicilina como marcador seleccionable (Figura 3). Además tiene
una región codificante del péptido α de la enzima β-galactosidasa (lacZ) cuya inactivación
insercional permite identificar por color los clones recombinantes. El tamaño del vector es
de 3015 pb.
Figura 3. Mapa circular del plásmido pGEM-T easy.
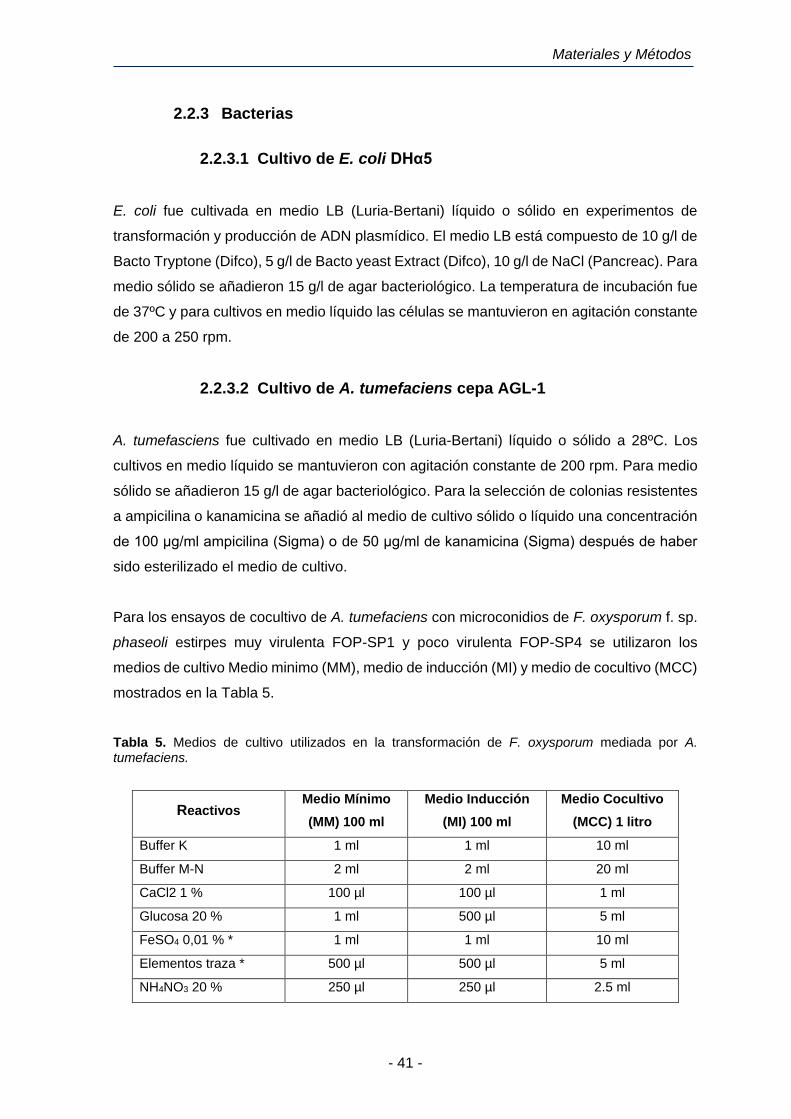
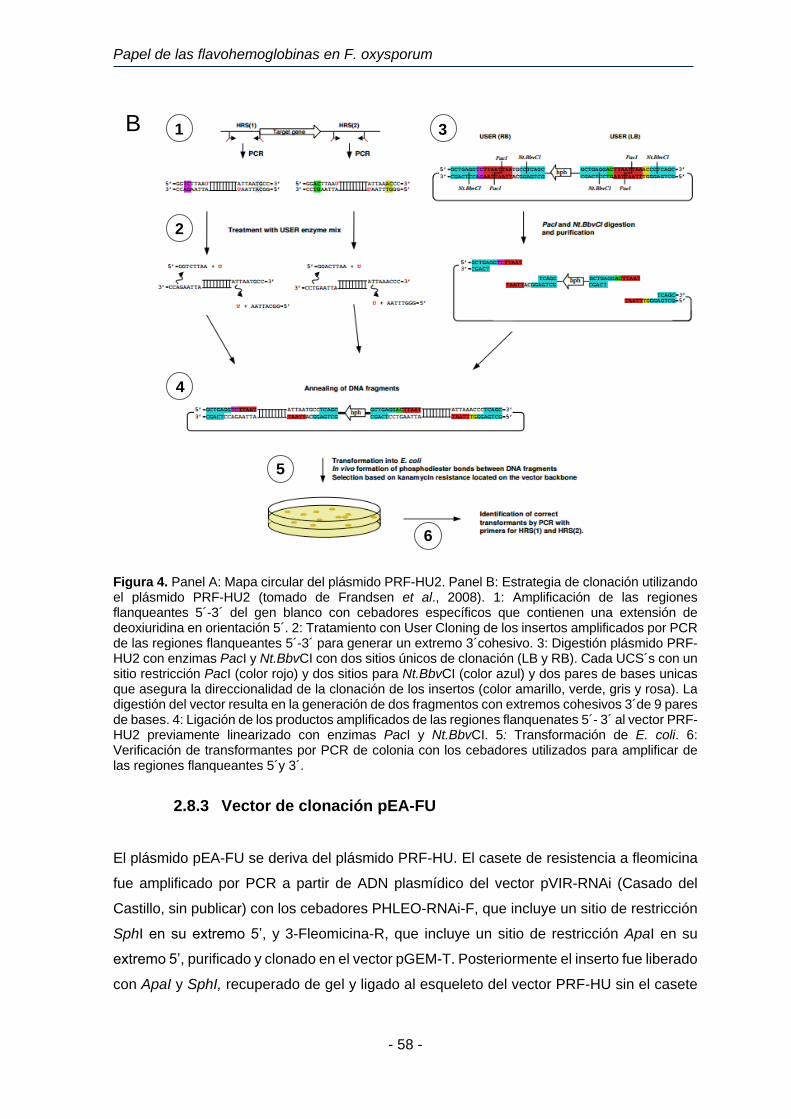
2.8.2 Vector de clonación pRF-HU2
El vector de clonación PRF-HU2 es un vector binario de 6.323 pb utilizado para la clonación
de dos fragmentos de ADN en un solo paso (Frandsen et al., 2008) (Figura 4, Panel I).
Materiales y Métodos
- 57 -
Posee un gen de resistencia a kanamicina (KanR) como marcador seleccionable en E. coli;
un gen de resistencia a higromicina B (hph) como marcador seleccionable en hongos y un
gen de iniciación de la replicación del plásmido (TrfA). El vector PRF-HU2 posee dos sitios
únicos de clonación (UCS´s), uno de ellos situado cerca del borde izquierdo (LB) y el otro
situado cerca del borde derecho (RB). Cada sitio único de clonación presenta una diana
para la enzima de restricción PacI, flanqueada por dianas para la enzima de restricción
Nt.BbvCI (Biolabs, Inglaterra). Los bordes LB y RB constituyen los límites del T-DNA (ADN
transferido). El T-DNA es un fragmento de ADN derivado del plásmido Ti de A. tumefaciens
que es transferido desde la bacteria hacia la célula vegetal huésped.
La digestión de PRF-HU2 con PacI y Nt.Bbv.CI divide el plásmido en 2 fragmentos lineales
de ADN, cada uno con 2 extremos cohesivos de 9 pares de bases, de las cuales 7 son
idénticas en todos los extremos y dos son características de cada extremo (Figura 4B-2).
Estas dos bases características de cada extremo proporcionan la direccionalidad de la
reacción de clonación, asegurando que los fragmentos de ADN a clonar que previamente
han sido generados mediante amplificación por PCR y digestión con la mezcla de enzimas
USER se liguen al vector y al fragmento con el gen de resistencia a higromicina en la
ordenación correcta, tal y como se indica en la Figura 4.
A
Papel de las flavohemoglobinas en F. oxysporum
- 58 -
Figura 4. Panel A: Mapa circular del plásmido PRF-HU2. Panel B: Estrategia de clonación utilizando el plásmido PRF-HU2 (tomado de Frandsen et al., 2008). 1: Amplificación de las regiones flanqueantes 5´-3´ del gen blanco con cebadores específicos que contienen una extensión de deoxiuridina en orientación 5´. 2: Tratamiento con User Cloning de los insertos amplificados por PCR de las regiones flanqueantes 5´-3´ para generar un extremo 3´cohesivo. 3: Digestión plásmido PRF-HU2 con enzimas PacI y Nt.BbvCI con dos sitios únicos de clonación (LB y RB). Cada UCS´s con un sitio restricción PacI (color rojo) y dos sitios para Nt.BbvCI (color azul) y dos pares de bases unicas que asegura la direccionalidad de la clonación de los insertos (color amarillo, verde, gris y rosa). La digestión del vector resulta en la generación de dos fragmentos con extremos cohesivos 3´de 9 pares de bases. 4: Ligación de los productos amplificados de las regiones flanquenates 5´- 3´ al vector PRF-HU2 previamente linearizado con enzimas PacI y Nt.BbvCI. 5: Transformación de E. coli. 6: Verificación de transformantes por PCR de colonia con los cebadores utilizados para amplificar de las regiones flanqueantes 5´y 3´.
2.8.3 Vector de clonación pEA-FU
El plásmido pEA-FU se deriva del plásmido PRF-HU. El casete de resistencia a fleomicina
fue amplificado por PCR a partir de ADN plasmídico del vector pVIR-RNAi (Casado del
Castillo, sin publicar) con los cebadores PHLEO-RNAi-F, que incluye un sitio de restricción
SphI en su extremo 5’, y 3-Fleomicina-R, que incluye un sitio de restricción ApaI en su
extremo 5’, purificado y clonado en el vector pGEM-T. Posteriormente el inserto fue liberado
con ApaI y SphI, recuperado de gel y ligado al esqueleto del vector PRF-HU sin el casete
B 1 3
2
4
5
6
Materiales y Métodos
- 59 -
de resistencia a Higromicina B (el casete de resistencia a higromicina fue retirado por
digestión con las enzimas de restricción ApaI y SphI), obteniéndose el vector pEA-FU. El
vector pEA-FU posee un sitio único de clonación (UCS) con sitios de restricción únicos para
las enzimas PacI y Nt.Bbv.CI. Tiene un gen de resistencia a kanamicina (KanR) como
marcador seleccionable en E. coli y un gen de iniciación de la replicación del plásmido (TrfA).
El casete de resistencia a fleomicina esta formado por el promotor del gen codificador de la
enzima glyceraldehydephosphate deshydrogenase (Pgpd) de A.awamori; el gen (ble) como
marcador seleccionable en hongos y el terminador transcripcional del gen codificador de la
enzima cytochrome oxidase (Tcyc1) de S. cerevisiae. El vector pEA-FU linearizado posee
extremos cohesivos de 9 pb, lo que proporciona la direccionalidad de la clonación en la
reacción con sistema USER Friendly Cloning y asegurando el correcto clonaje del fragmento
de ADN durante la reacción de clonación. El vector tiene un tamaño de 5.459 pb (Figura 5).
Figura 5. Mapa de construcción del plásmido pEA-FU. Panel A se muestra el plásmido PRF-HU con el casete de resistencia a higromicina que fue eliminado por digestión enzimática con las enzimas de restricción ApaI y SphI. Panel B se muestra el plásmido pVIR-RNAi del cual se amplificó el casete de resistencia a fleomicina con los cebadores PHLEO-RNAi-F que posee un sitio de restricción SphI en el extremo 5´y Fleomicina-R que posee un sitio de restricción ApaI en el extremos 5´. Panel C se muestra el plásmido pEA-FU que fue utilizado para ensayos de complementación.
Papel de las flavohemoglobinas en F. oxysporum
- 60 -
2.8.4 Vector de silenciamiento pVIR-RNAi
El vector de silenciamiento pVIR-RNAi (Casado del Castillo, sin publicar) posee un gen de
resistencia a kanamicina (KanR) como marcador seleccionable en E. coli; un gen de
iniciación de la replicación del plásmido (TrfA). El casete de resistencia a fleomicina esta
formado por el promotor del gen codificador de la enzima glyceraldehydephosphate
deshydrogenase (Pgpd) de A. awamori; el gen (ble) como marcador seleccionable en
hongos y el terminador transcripcional del gen codificador de la enzima cytochrome oxidase
(Tcyc1) de S. cerevisiae. Este vector presenta un casete de silenciamiento formado por el
promotor del gen codificador de la enzima glyceraldehydephosphate deshydrogenase
(Pgpd) de A. awamori y el promotor del gen que codifica la enzima isopenicillin N-synthetase
(PpcbC) de P. chrysogenum, orientados en sentidos opuestos. Entre ambos promotores se
ubica un sitio único de clonación NcoI, en el cual se inserta el fragmento de ADN del gen
diana a ser silenciado. El vector pVIR-RNAi tiene un tamaño de 7.254 pb (Figura 6).
Figura 6. Mapa circular plásmido pVIR-RNAi utilizado para silenciamiento genético.
2.9 Transformaciones
2.9.1 Preparación de células competentes de E. coli
Para la preparación de células competentes de E. coli se siguió el siguiente procedimiento.
Una alícuota de E. coli almacenada a -80ºC se sembró en una placa petri con medio LB que
fue incubada toda la noche a 37ºC. Al siguiente día se inoculó una colonia única en 5 ml de
medio de cultivo líquido SOB y el cultivo se incubó toda la noche a 37ºC y 200 rpm. Se tomó
Materiales y Métodos
- 61 -
1 ml de preinóculo que se añadió a 250 ml de medio de cultivo SOB (20 g/l de Triptona, 5
g/l de Extracto de levadura, 0,5 g/l de NaCl, 2,3 ml de glucosa 40 %; 10 ml de KCl 250 mM,
ajustado el pH a 7,0 y esterilizado en autoclave. A este medio posteriormente se le añade 5
ml de MgCl2 2M) extendiendo la incubación 10 a 12 horas a 37ºC y 200 rpm (hasta alcanzar
una DO600nm = 0,6). A partir de este momento se trabajó todo el tiempo en hielo (el rotor y la
centrífuga deben estar refrigerados). Los 250 ml de cultivo bacteriano fueron transferidos a
4 tubos para la centrífuga (Beckman Avanti®J-25I, USA) y fueron incubados en hielo
durante 10 minutos antes de proceder a su centrifugación a 3.000 rpm durante 10 minutos
y 4°C en un ROTOR JLA-16-250.
Se eliminó el medio de cultivo y las células sedimentadas fueron resuspendidas en 80 ml
de buffer TB frio (PIPES 10 mM, CaCl2 15 mM, KCl 250 mM, ajustar el pH a 6,7 con KOH
1M, y añadir finalmente 55 mM de MnCl2 4H2O). La suspensión fue homogeneizada
agitando manualmente (no se debe usar el vórtex), incubada en hielo durante 10 minutos y
centrifugada a 3.000 rpm y 4ºC. Se eliminaron los restos de tampón y las células fueron
transferidas a 20 ml de buffer TB frio, homogeneizando la suspensión de células de nuevo
agitando manualmente. A continuación se añadieron 1,4 ml de DMSO 7%. Las células
fueron resuspendidas mediante agitación manual, incubadas en hielo durante 10 minutos y
a continuación repartidas en alícuotas de 10 y 100 µl en tubos eppendorf que fueron
congelados en nitrógeno líquido y posteriormente almacenados a -80°C.
2.9.2 Digestión de los vectores de clonación PRF-HU2, pVIR-RNAi y
pEA-FU
Aproximadamente 10 µg de ADN del vector de clonación PRF-HU2 fueron digeridos con 70
U (7 µl) de la enzima PacI en un volumen total de 200 µl a 37ºC durante toda la noche. Al
día siguiente se añadieron 20 U (2 µl) de la enzima PacI y 40 U (4 µl) de la enzima Nt.Bbv.CI
y la suspensión se incubó durante 3 horas. La digestión completa del vector PRF-HU2 fue
verificada mediante electroforesis en gel de agarosa. Una vez comprobada la digestión total
del plásmido, el ADN digerido fue purificado sobre columna con el kit comercial
NucleoSpin®Gel and PR clean-up, siguiendo las recomendaciones del fabricante. La
concentración final del vector digerido fue aproximadamente de 100 ng/µl en un volumen
final de 100 µl. El vector pVIR-RNAi utilizado para la obtención de transformantes
silenciados fue linearizado con la enzima NcoI. El plásmido pEA-FU, utilizado para la
complementación de los mutantes deficientes en el gen FHG1, fue digerido con las enzimas
PacI y Nt.Bbv.CI como se ha indicado en el caso del plásmido pRF-HU2.
Papel de las flavohemoglobinas en F. oxysporum
- 62 -
2.9.3 Ensayos de clonación con el sistema “USER Friendly Cloning”
2.9.3.1 Diseño de oligonucleótidos con un adaptador de uridina
Los oligonucleótidos para la construcción de mutantes deficientes en los genes que
codifican las enzimas tipo flavohemoglobina de F. oxysporum f. sp. phaseoli fueron
diseñados en base al genoma anotado de F. oxysporum f. sp. lycopersici depositado en la
base de datos del Broad Institute de la Universidad de Harvard (U.S.A.)
(www.broadinstitute.org/annotation/genome/fusarium_group). A partir de las secuencias de
las regiones flanqueantes 5´ y 3’ de cada gen se diseñaron oligonucleótidos específicos a
los que se añadió en cada caso un adaptador de 9 pb en el extremo 5´ (Primer 1:
GGTCTTAAU; primer 2: GGCATTAAU; primer 3: GGACTTAAU; primer 4: GGGTTTAAU).
Cada adaptador incluye un nucleótido de Deoxiuridina para permitir la direccionalidad de la
clonación con el sistema USER Cloning (Biolabs, Inglaterra). La síntesis de los
oligonucleotidos fue realizada por la empresa Integrated DNA Technologies. Las secuencias
de los oligonucleótidos diseñados para cada uno de los cuatro genes codificadores de
enzimas de tipo flavohemoglobina de F. oxysproum se presentan en la Tabla 6.
2.9.3.2 Amplificación de las regiones flanqueantes
Los fragmentos correspondientes de las regiones flaqueantes 5’ y 3’ de cada uno de los
genes FHG necesarios para la creación de los vectores de remplazamiento génico utilizando
el sistema USER Friendly Cloning fueron amplificados por PCR con la enzima pfuTurbo Cx
Hoststart ADN polimerasa (Agilent Technologies), que reconoce nucleótidos de
deoxiuridina. La ampificación de las regiones flanqueantes 5´-3´se realizaron con el
siguiente progama de PCR: un ciclo inicial de 5 minutos a 95 ºC, seguido de 35 ciclos de
95ºC (desnaturalización) 30 segundos y 35 ciclos de 55ºC a 58 ºC dependiendo del cebador
(Tabla 6) y una la extensión a 72ºC durante 1 a 1,5 minutos. Los productos de PCR fueron
visualizados en gel de agarosa 1 % en TAE 1X, verificando que tuvieran el tamaño adecuado
y la concentración suficiente para las reacciones de clonación. Los productos de PCR fueron
purificados con el kit comercial NucleoSpin® Gel and PR clean-up, siguiendo las
nuestro análisis permitió identificar la presencia de cuatro genes “de tipo
flavohemoglobina”. En la versión 2015 se identifican únicamente 3 genes. Detectadas
estas inconsistencias, que atribuimos a variaciones debidas a los distintos programas
utilizados para la anotación automática de los genomas y su actualización, nuestro grupo
de investigación decidió continuar trabajando con los cuatro genes identificados en la
versión del año 2012, todos ellos caracterizados por codificar proteínas que presentan los
dominios funcionales característicos específicos de proteínas de tipo flavohemoglobina.
Los códigos de los cuatro genes identificados en el genoma de F. oxysporum f. sp.
lycopersici son: FOXG_17028.3, FOXG_13752.3, FOXG_18012.3 y FOXG_15840.2 (Tabla
7). Con los mismos criterios de búsqueda se analizó el genoma de las cepas secuenciadas
de F. verticilloides (7600) y F. graminearum (PH-1). En estos casos se identificaron 3 y 2
“genes codificadores de enzimas de tipo flavohemoglobina”, respectivamente. Los códigos
identificativos de todos ellos se incluyen en la Tabla 7. En la misma, los genes identificados
en las distintas cepas se presentan ordenados en función de la similitud que presentan
entre ellos (es decir, en la misma fila de la tabla se presentan los genes de las distintas
cepas que pueden ser considerados como ortólogos).
Tabla 7. Códigos de genes codificadores de proteínas tipo flavohemoglobina del género Fusarium. Columna 1: F. oxysporum f. sp. lycopersici, columna 2: F. verticillinoides; columna 3: F. graminearum.
estudiar esta situación en cuatro cepas de F. oxysporum f. sp. phaseoli con características
fisiológicas muy distintas: la cepa FOP-SP1 (muy virulenta), FOP-SP4 (poco virulenta),
FOP-SP13 (supervirulenta) y AB82 (no patógena). Con este objetivo analizamos en detalle
la secuencia de nucleótidos de cada uno de los cuatro genes de F. oxysporum f. sp.
lycopersici y diseñamos para cada gen dos cebadores que permitían amplificar mediante
PCR específicamente un fragmento de ADN de cada gen. La Tabla 8 muestra los nombres
de los oligonucleótidos diseñados y en la Tabla 6 de la sección de Materiales y Métodos
se presentan las secuencias de los mismos.
Tabla 8. Códigos de los oligonucleótidos diseñados sobre los genes codificadores de enzimas de tipo flavohemoglobina de F. oxysporum f. sp. lycopersici y utilizados en reacciones de PCR para detectar los genes ortólogos en F. oxysporum f. sp. phaseoli.
Genes de F. oxysporum
f. sp. lycopersici
Nombre del
oligonucleótido
FOXG_17028.3 FHG1-FW
FHG1-RV
FOXG_13752.3 FHG2-FW
FHG2-RV
FOXG_18012.3 FHG3-FW
FHG3-RV
FOXG_15840.2 FHG4-FW
FHG4-RV
Con estas cuatro combinaciones de oligonucleótidos se llevaron a cabo reacciones de PCR
utilizando como molde ADN genómico de la cepa FOP-SP1 de F. oxysporum f. sp. phaseoli.
Las reacciones de PCR se realizaron en un volumen final de 25 µl conteniendo 2,5 µl de
buffer 10X, 10 µM de cada oligo, 200 µM de dNTPs, 10 ng de DNA genómico y 0,5 U de
DNA polimerasa (DNA Polymerase bitools 1 U/µl). El perfil del programa de PCR fue: 1
ciclo de desnaturalización de 5 min a 94ºC, seguido de 35 ciclos de amplificación
consistentes en 30 seg a 94ºC, 30 seg a 58ºC y 1 min a 72ºC y un ciclo final de extensión
de 5 min a 72ºC.
La Figura 7 muestra los productos de amplificación obtenidos separados mediante
electroforesis en gel de agarosa. En cada caso se obtuvo un producto único de
amplificación, de tamaño diferente con cada combinación particular de cebadores: 481
pares de nucleótidos con la pareja específica de FOXG_17028.3, 511 pares de nucleótidos
con la pareja específica de FOXG_13752.3, 661 pares de nucleótidos con la pareja
específica de FOXG_18012.3 y 591 pares de nucleótidos con la pareja específica de
FOXG_15840.2.
Figura 7. Electroforesis en gel de agarosa de los productos de amplificación por PCR de secuencias derivadas de la región codificante de los genes codificadores de proteínas de tipo flavohemoglobina utilizando ADN genómico de F. oxysporun f. sp. phaseoli (FOP-SP1) y las combinaciones de oligonucleótidos específicas de (A): gen FOXG_17028.3, (B): gen FOXG_13752.3, (C): gen FOXG_18012.3 y (D): gen FOXG_15840.2. En el carril M: marcador de peso molecular 1 kp plus DNA ladder (Invitrogen).
El hecho de que con cada combinación de cebadores se generara un único producto de
amplificación, producto de amplificación diferente y específico de cada combinación de
cebadores, junto con el hecho de que éstos hayan sido diseñados teniendo en cuenta la
secuencia de cuatro genes diferentes codificadores de enzimas de tipo flavohemoglobina
de F. oxysporum f. sp. lycopersici, nos permite proponer que ésta es probablemente la
situación también en F. oxysporum f. sp. phaseoli cepa FOP-SP1. Asumimos así
inicialmente la existencia de cuatro genes “FHG” en el genoma de FOP-SP1, codificadores
de enzimas tipo flavohemoglobina, que fueron denominados de la siguiente manera:
3.1.3 Caracterización del entorno genómico de los genes
codificadores de flavohemoglobinas de F. oxysporum f. sp.
phaseoli
Para caracterizar el entorno genómico de cada uno de los cuatro posibles genes
codificadores de enzimas de tipo flavohemoglobina en las cuatro cepas de F. oxysporum f.
sp. phaseoli objeto de estudio en este trabajo (FOP-SP1, FOP-SP4, FOP-SP13 y AB82)
en comparación con F. oxysporym f. sp. lycopersici y para determinar si éstos están
presentes en copia única, se llevó a cabo un análisis de hibridación tipo Southern. Para
ello se utilizaron como sondas los fragmentos de ADN genómico amplificados mediante
PCR con las cuatro combinaciones de cebadores descritas en la sección 3.1.2 de
Resultados (ver Figura 7) (marcadas también mediante PCR) que fueron probadas sobre
filtros de nylon a los que habían sido transferidos los fragmentos resultantes de las
digestiones de ADN genómico de las cuatro cepas de F. oxysporum f. sp. phaseoli y de la
estirpe 4287 de F. oxysporum f. sp. lycopersici (FOL) con las enzimas de restricción HindIII
y BamHI.
La Figura 8 recoge los resultados obtenidos en las hibridaciones llevadas a cabo con cada
una de las cuatro sondas. De su estudio se derivan diversas consideraciones de interés.
En primer lugar, en cada caso el patrón de hibridación obtenido es específico de cada
sonda, lo que permite proponer que cada sonda detecta una región particular del genoma
de las estirpes analizadas y, por lo tanto, que existen cuatro genes codificadores de
enzimas de tipo flavohemoglobina en el genoma de las cuatro estirpes de F. oxysporum f.
sp. phaseoli analizadas, así como en la estirpe 4287 de F. oxysporum f. sp. lycopersici.
En segundo lugar, con ninguna de las cuatro sondas se detecta hibridación cruzada en
condiciones de alta astringencia, lo que supone que los cuatro genes, aun teniendo cierto
grado de similitud, son lo suficientemente diferentes en su secuencia de nucleótidos como
para no ser detectados de forma cruzada en condiciones de hibridación restrictivas.
En tercer lugar, en las digestiones con la enzima HindIII se detecta en todos los casos una
única banda de hibridación (lo que supone que la enzima en ningún caso corta en la
secuencia interna de la sonda). Es posible deducir, entonces, que son genes de copia única
y que no existen secuencias muy relacionadas en el genoma de las estirpes analizadas.
Papel de las flavohemoglobinas en F. oxysporum
- 84 -
Y finalmente, los patrones de hibridación son esencialmente idénticos entre las cinco cepas
analizadas en relación con las dos digestiones consideradas en estas hibridaciones. Sólo
se detecta un polimorfismo (RFLP) entre los patrones de hibridación de las cuatro cepas
de F. oxysporum f. sp. phaseoli y el de la cepa de F. oxysporum f. sp. lycopersici en la
hibridación llevada a cabo con la sonda derivada del gen FHG1, el correspondiente a la
banda BamHI de 12.300 nucleótidos característica de esta última, de un tamaño
ligeramente superior a la banda detectada en la cuatro estirpes de F. oxysporum f. sp
phaseoli. Es interesante destacar que entre las cuatro estirpes de F. oxysporum f. sp.
phaseoli también se detecta un polimorfismo, tanto en la digestión con la enzima HindIII
como en la digestión con la enzima BamHI, en la hibridación llevada a cabo con la sonda
derivada del gen FHG4, y que marca una diferencia entre la estirpe AB82, la única estirpe
no patogénica, y todas las demás, incluida la cepa 4287 de FOL (Figura 8).
Figura 8. Análisis tipo Southern de los genes codificadores de enzimas flavohemoglobina en las diferentes estirpes de F. oxysporum f. sp. phaseoli. En la parte izquierda de cada panel se muestran las hibridaciones correspondientes a digestiones de ADN genómico realizadas con la enzima HindIII, y en la parte derecha las digestiones de ADN genómico realizadas con la enzima BamHI. A: hibridación con la sonda derivada del gen FHG1; B: hibridación con la sonda derivada del gen FHG2; C: hibridación con la sonda derivada del gen FHG3; D: hibridación con la sonda derivada del gen FHG4. Los números del 1 al 5 corresponden a las muestras de ADN genómico de las estirpes FOP-SP1 (1), FOP-SP4 (2), FOP-SP13 (3), AB82 (4) y F. oxysporum f. sp. lycopersici (4287) (5).
Resultados
- 85 -
En resumen, los resultados presentados en la Figura 8 permiten asegurar que el genoma
de los cuatro estirpes de F. oxysporum f. sp. phaseoli analizadas, FOP-SP1, FOP-SP4,
FOP-SP13 y AB82, incluye cuatro genes de copia única codificadores de enzimas de tipo
flavohemoglobina. La situación es, por lo tanto, la misma que la descrita en la estirpe de F.
oxysporum f. sp. lycopersici utilizada como referencia en este trabajo y cuya consideración
nos ha permitido identificar de forma sencilla los genes ortólogos en F. oxysporum f. sp
phaseoli antes de disponer del genoma de esta forma especial.
3.2 Análisis estructural de los genes codificadores de
flavohemoglobinas de F. oxysporum f. sp. phaseoli
En el curso de este trabajo y una vez identificados mediante PCR e hibridación Southern
los cuatro genes codificadores de enzimas de tipo flavohemoglobina en F. oxysporum f. sp
phaseoli fue posible disponer de la secuencia completa del genoma de las estirpes FOP-
SP1, FOP-SP4, FOP-SP13 y AB82. Estas secuencias fueron obtenidas en el curso de un
proyecto de investigación sobre genómica de hongos fitopatógenos que desarrolla nuestro
grupo de investigación (AGL2012-39876-C02-01). Un análisis detallado de la secuencia
del genoma de la estirpe FOP-SP1 permitió confirmar la presencia en el mismo de cuatro
genes codificadores de enzimas de tipo flavohemoglobina, los genes que hemos
denominado previamente FHG1, FHG2, FHG3 y FHG4. La secuencia de nucleótidos de la
región codificante y de las regiones flanqueantes en dirección 5’ y en dirección 3’ de cada
gen fueron rescatadas, revisadas detalladamente y anotadas. Se presenta a continuación
la secuencia de cada uno de ellos.
Papel de las flavohemoglobinas en F. oxysporum
- 86 -
3.2.1 Secuencia de los genes FHG de F. oxysporum f. sp. phaseoli
3.2.1.1 Gen FHG1 de FOP-SP1
La región codificante del gen FHG1 consta de 1266 nucleótidos y carece de intrones. Su
traducción da lugar a una secuencia de proteína deducida de 422 aminoácidos (Figura 9).
Figura 9. Secuencia de nucleótidos del gen FHG1 y secuencia de aminoácidos de la proteína deducida. La secuencia de la región codificante (1266 nt) está representada en negro y numerada a partir del codón de iniciación ATG. Los codones de iniciación y terminación de la traducción se muestran subrayados en color rojo. La secuencia de aminoácidos de la proteína deducida se muestra en color rojo debajo de la secuencia de nucleótidos. Se incluyen 756 nucleótidos de la región 5’ flanqueante y 555 nucleótidos de la región 3’ flanqueante. Las posibles cajas CAAT y TATA identificadas en la región promotora se presentan marcadas en color rojo y subrayado.
Resultados
- 87 -
3.2.1.2 Gen FHG2 de FOP-SP1
La región codificante del gen FHG2 consta de 1247 nucleótidos y carece de intrones. Su
traducción da lugar a una secuencia de proteína deducida de 416 aminoácidos (Figura 10).
Figura 10. Secuencia de nucleótidos del gen FHG2 y secuencia de aminoácidos de la proteína deducida. La secuencia de la región codificante (1247 nt) está representada en negro y numerada a partir del codón de iniciación ATG. Los codones de iniciación y terminación de la traducción se muestran subrayados en color rojo. La secuencia de aminoácidos de la proteína deducida codificada se muestra en color rojo debajo de la secuencia de nucleótidos. Se incluyen 723 nucleótidos de la región 5’ flanqueante y 846 nucleótidos de la región 3’ flanqueante. Las posibles cajas CAAT y TATA identificadas en la región promotora se presentan marcadas en color rojo y subrayado.
Papel de las flavohemoglobinas en F. oxysporum
- 88 -
3.2.1.3 Gen FHG3 de FOP-SP1
La región codificante del gen FHG3 consta de 1365 nucleótidos y carece de intrones. Su
traducción da lugar a una secuencia de proteína deducida de 456 aminoácidos (Figura 11).
Figura 11. Secuencia de nucleótidos del gen FHG3 y secuencia de aminoácidos de la proteína deducida. La secuencia de la región codificante (1365 nt) está representada en negro y numerada a partir del codón de iniciación ATG. Los codones de iniciación y terminación de la traducción se muestran subrayados en color rojo. La secuencia de aminoácidos de la proteína deducida se muestra en color rojo debajo de la secuencia de nucleótidos. Se incluyen 807nucleótidos de la región 5’ flanqueante y 609 nucleótidos de la región 3’ flanqueante. Las posibles cajas CAAT y TATA identificadas en la región promotora se presentan marcadas en color rojo y subrayado. La posible secuencia de poliadenilación AATAAA se muestra en color morado y subrayada.
Resultados
- 89 -
3.2.1.4 Gen FHG4 de FOP-SP1
La región codificante del gen FHG4 consta de 1290 nucleótidos y carece de intrones. Su
traducción da lugar a una secuencia de proteína deducida de 430 aminoácidos (Figura 12).
Figura 12. Secuencia de nucleótidos del gen FHG4 y secuencia de aminoácidos de la proteína deducida. La secuencia de la región codificante (1290 nt) está representada en negro y numerada a partir del codón de iniciación ATG. Los codones de iniciación y terminación de la traducción se muestran subrayados en color rojo. La secuencia de aminoácidos de la proteína deducida codificada se muestra en color rojo debajo de la secuencia de nucleótidos. Se incluyen 751 nucleótidos de la región 5’ flanqueante y 669 nucleótidos de la región 3’ flanqueante. Las posibles cajas CAAT y TATA identificadas en la región promotora se presentan marcadas en color rojo y subrayado.
3.2.2 Análisis de la región codificante de los genes FHG
Estructuralmente los genes FHG de F. oxysporum f. sp. phaseoli de FOP-SP1 son muy
similares: todos ellos carecen de intrones y las correspondientes regiones estructurales
tienen tamaños muy parecidos entre sí (1266, 1247, 1365 y 1290 nucleótidos para los
genes FHG1, FHG2, FHG3 Y FHG4, respectivamente) y similares a los tamaños de la
regiones codificantes de otros genes codificadores de enzimas de tipo flavohemoglobina
de hongos (Gardner et al., 2005).
Cuando se comparan las secuencias de nucleótidos de los genes de FHG de FOP-SP1
con los genes ortólogos de FOL se comprueba que en todos los casos existe un nivel de
semejanza muy elevado, como cabe esperar tratándose de estirpes del mismo complejo
de especies, si bien correspondientes a formas especiales distintas, aunque variable según
el gen. Se detectan 17 sustituciones nucleotídicas cuando se comparan el gen FHG1 y el
gen FOXG_17028.3, 4 cuando se comparan el gen FHG2 y el gen FOXG_13752.3, 12
cuando se comparan el gen FHG3 y el gen FOXG_18012.3 y 20 cuando se comparan el
gen FHG4 y el gen FOXG_15840.2.
El alineamiento de las secuencias de nucleótidos de los cuatro genes de FHG de FOP-SP1
y FOP-SP4 indica que el grado de identidad entre ellos es superior al 30 % y que los genes
FHG1, FHG2 y FHG3 están más relacionados entre sí que con el gen FHG4 (ver Tabla 9).
Tabla 9. Porcentajes de identidad entre las secuencias de nucleótidos de las regiones codificantes de los genes FHG1, FHG2, FHG3 Y FHG4.
FGH1 FHG2 FHG3 FHG4
FHG1 100 %
FHG2 48 % 100 %
FHG3 35 % 47 % 100 %
FHG4 33 % 35 % 33 % 100 %
Resultados
- 91 -
3.2.3 Análisis de las regiones promotora y terminadora de los genes
FHG
En la Tabla 10 se presentan las posiciones en las que se identifican secuencias
relacionadas con el control de la transcripción de forma general, tanto en la región
promotora (cajas TATA y CAAT) como en la región terminadora (secuencia consenso de
señal de poliadenilación “AATAAA”) de cada uno de los cuatro genes de FHG. Es posible
destacar que en las regiones promotoras de los cuatro genes se identifican varias cajas
TATA y CAAT en localizaciones compatibles con su posible función reguladora de la
transcripción. En la región terminadora, por su parte, sólo se identifica la secuencia
consenso señal de poliadenilación característica de organismos eucariotas (Heidmann et
al., 1992) en el caso del gen FGH3. Debemos recordar que esta secuencia consenso señal
de poliadenilación no está siempre presente en todos los terminadores de los genes de
hongos, en los que otras secuencias relacionadas (AATAAT, AATATT, AATACT) pueden
desempeñar la función de señalización del lugar de poliadenilación, y que sí se identifican
en la región terminadora de los genes FHG.
Tabla 10. Ubicación de las hipotéticas cajas TATA y CAAT en la región promotora y secuencias consenso señal de poliadenilación reconocidas en los genes FHG1, FHG2, FHG3 y FHG4.
Secuencia consenso Gen FHG1 Gen FHG2 Gen FHG3 Gen FHG4
Caja TATA
-88 a -85
-133 a -129
-137 a -134,
-630 a -626
-119 a -116 -121 a -117
-230 a -227
-151 a -148
-297 a -294
-474 a -471
Caja CAAT
-118 a -115
-306 a -303
-568 a -565
-334 a -331
-545 a -541
-620 a -617
-63 a -60
-132 a -129
-359 a -356
-571 a -567
Secuencia señal de
poliadenilación AATAAA +352 a +355
El análisis de las regiones promotoras mediante el programa MatInspector (Cartharius et
al., 2005) permitió la identificación de posibles secuencias señal de unión a factores de
transcripción. Entre las secuencias más importantes se han identificado aquellas
relacionadas con la activación de genes inducidos por nitrógeno. Estas secuencias podrían
estar implicadas en la activación de los genes FHG en procesos relacionados con el
metabolismo del nitrógeno. Igualmente se detectan secuencias señal de unión a factores
Papel de las flavohemoglobinas en F. oxysporum
- 92 -
de transcripción relacionados con la respuesta a situaciones de estrés. Asimismo,
muestran secuencias señal de unión a factores de transcripción que inducen la activación
de genes de regulación de la fase G1 del ciclo celular. Las secuencias indicadas, y sus
localizaciones en las regiones promotoras correspondientes, se muestran en las Tablas 11,
12, 13 y 14.
Tabla 11. Predicción de factores de transcripción que pueden unirse a la región promotora del gen FHG1. En la primera columna se indica el nombre de la familia de factores de transcripción que reconocen la secuencia señal consenso. En la segunda y tercera columna se muestra el inicio y fin de la secuencia. En la cuarta columna se muestra la orientación de la cadena en la que se localiza la secuencia. En la quinta columna se indica la ratio de semejanza de la secuencia detectada con la secuencia consenso señal. En la última columna se indica la secuencia consenso.
Tabla 12. Predicción de factores de transcripción que pueden unirse a la región promotora del gen FHG2. En la primera columna se indica el nombre de la familia de factores de transcripción que reconocen la secuencia señal consenso. En la segunda y tercera columna se muestra el inicio y fin de la secuencia. En la cuarta columna se muestra la orientación de la cadena en la que se localiza la secuencia. En la quinta columna se indica la ratio de semejanza de la secuencia detectada con la secuencia consenso señal. En la última columna se indica la secuencia consenso.
Resultados
- 93 -
Tabla 13. Predicción de factores de transcripción que pueden unirse a la región promotora del gen FHG3. En la primera columna se indica el nombre de la familia de factores de transcripción que reconocen la secuencia señal consenso. En la segunda y tercera columna se muestra el inicio y fin de la secuencia. En la cuarta columna se muestra la orientación de la cadena en la que se localiza la secuencia. En la quinta columna se indica la ratio de semejanza de la secuencia detectada con la secuencia consenso señal. En la última columna se indica la secuencia consenso.
Tabla 14. Predicción de factores de transcripción que pueden unirse a la región promotora del gen FHG4. En la primera columna se indica el nombre de la familia de factores de transcripción que reconocen la secuencia señal consenso. En la segunda y tercera columna se muestra el inicio y fin de la secuencia. En la cuarta columna se muestra la orientación de la cadena en la que se localiza la secuencia. En la quinta columna se indica la ratio de semejanza de la secuencia detectada con la secuencia consenso señal. En la última columna se indica la secuencia consenso.
Papel de las flavohemoglobinas en F. oxysporum
- 94 -
3.3 Análisis de las proteínas deducidas codificadas por los genes FHG1,
FHG2, FHG3 y FHG4
3.3.1 Dominios funcionales de las proteínas FHG
Las secuencias de aminoácidos deducidas codificadas por los genes FHG1, FHG2, FHG3
y FHG4 tienen un tamaño de 422, 416, 456 y 430 aminoácidos, respectivamente. En la
Tabla 15 se presenta la información sobre la composición aminoacídica de cada una de las
cuatro proteínas.
Tabla 15. Composición aminoacídica de las proteínas codificadas por los genes FHG1, FHG2, FHG3 Y FHG4.
Resultados
- 95 -
En la Figura 13, se muestra el alineamiento de las secuencias de las proteínas codificadas
por los genes FHG1, FHG2, FHG3 y FHG4, las cuales muestran porcentajes de identidad
al 40 %. Las proteínas FHG1 y FHG2 muestran el mayor porcentaje de semejanza con el
53,41 %, en tanto que el menor corresponde a FHG1 y FHG3, con el 40,73 % (ver Tabla
13).
Tabla 16. Porcentaje de semejanza de las secuencias aminoacídicas deducidas de las proteínas codificadas por los genes FHG1, FHG2, FHG3 y FHG4 de FOP-SP1.
FGH1 FHG2 FHG3 FHG4
FHG1 100 %
FHG2 53,41 % 100 %
FHG3 40,73 % 47,30 % 100 %
FHG4 43,11 % 41,37 % 40,20 % 100 %
Figura 13. Alineamiento de secuencias de las proteínas tipo flavohemoglobina de FOP-SP1. El color rojo representa el 90 % de consenso y el azul más del 50 % de consenso en las secuencias de aminoácidos. Los códigos de las secuencias utilizadas corresponden a los genes codificadores de enzimas tipo flavohemoglobina de F. oxysporum f. sp. phaseoli: FHG1, FHG2, FHG3 y FHG4.
Papel de las flavohemoglobinas en F. oxysporum
- 96 -
El tamaño de las cuatro proteínas deducidas es similar. La más pequeña es la proteína
FHG2, de 416 aminoácidoss, y la de mayor tamaño es la proteína FHG3, de 456
aminoácidos. El estudio de los alineamientos y del grado de conservación de aminoácidos
demuestra que la estructura general de las proteínas es similar en los cuatro casos (Figura
13). La proteína FHG3 tiene una extensión exclusiva de 37 aminoácidos en el extremo C-
carboxilo de la proteína. Por su parte, la proteína FHG4 presenta una extensión particular
en su extremo N-amino de 22 aminoácidos. Como veremos más adelante, esta extensión
marca una diferencia fundamental en relación con las otras tres secuencias FHG.
El análisis de las secuencias de aminoácidos deducidas permite identificar en las cuatro
proteínas tres dominios funcionales bien definidos, localizados en posiciones muy
similares, y con tamaños muy parecidos: un dominio globina (globin) en el extremo amino
de la proteína, un dominio oxidoreductasa de unión a FAD (FAD_binding_6) en la región
central y un dominio oxidoreductasa de unión a NAD (NAD_binding_1) en el extremo
carboxilo. La Figura 14 muestra una representación simplificada de la organización de cada
una de las cuatro proteínas analizadas y de la localización de los dominios funcionales
conservados que se han identificado en cada caso.
Figura 14. Dominios funcionales característicos de las proteínas de tipo flavohemoglobina, y sus posiciones relativas, identificados en las proteínas deducidas codificadas por los genes (A): FHG1, (B): FHG2, (C): FHG3 y (D): FHG4 de FOP-SP1.
Resultados
- 97 -
En la Tabla 17 se resume la información relacionada con la posición de cada uno de los
tres dominios funcionales identificados en la proteína correspondiente y el valor E que el
análisis bioinformático asigna en cada caso.
Tabla 17. Dominios funcionales presentes en las proteínas FHG1, FHG2, FHG3 y FHG4.
Gen Inicio Fin E-Value Dominios
FHG1
7 104 8,7051e-07 Globin
153 263 0,000221694 FAD_binding_6
279 396 4,98703e-10 NAD_binding_1
FHG2
7 104 1,42061e-08 Globin
153 269 2,65117e-05 FAD_binding_6
283 385 1,53381e-09 NAD_binding_1
FHG3
10 101 0,00250719 Globin
153 270 2,73308e-07 FAD_binding_6
282 387 1,3732e-08 NAD_binding_1
FHG4
32 129 2,45399e-10 Globin
182 288 8,79661e-10 FAD_binding_6
298 407 8,17601e-20 NAD_binding_1
El programa CCD del NCBI (National Center for Biotechnology Information) (Marchler-
Bauer et al., 2015) permite visualizar en forma de alineamientos locales las semejanzas
entre los dominios funcionales recogidos en la base de datos y los posibles dominios
funcionales presentes en las secuencias de aminoácidos objeto de análisis en cada caso.
Las Figuras 15, 16 y 17 muestran los alineamientos obtenidos en el análisis llevado a cabo
con la proteína FHG1. Los resultados obtenidos en los análisis de FHG2, FHG3 y FHG4
Figura 15. Dominios globina identificados en proteínas representativas con el programa CCD semejantes al dominio globina presente en la proteína codificada por el gen FHG1. Los códigos de cada secuencia corresponden a los siguientes organismos: 1X9F_I Lumbricus terrestris; query: FHG1; gi 121275 Tubifex tubifex; gi1707910 Nippostrongylus brasiliensis; gi 121251 Trichostrongylus colubriformis; gi 121213 Calyptogena soyoae; GVH_A Escherichia coli; 3VHB_A Vitreoscilla stercoraria; gi 17433294 Casuarina glauca; gi 1707913 Anadara trapezia.
Figura 16. Dominios oxidoreductasa FAD (FAD_binding_ 6) identificados en proteínas representativas con el programa CCD semejantes al dominio oxidoreductasa FAD (FAD_binding_ 6) presente en la proteína codificada por el gen FHG1. Los códigos de cada secuencia corresponden a los siguientes organismos. 1QX4_B Rattus norvegicus; query-FHG1; gi 74546820 Pyrococcus abyssi; gi 75345543 Pseudomonas putida; gi 3024428 Pseudomonas pseudoalcaligenes; gi75424276 Xanthobacter autotrophicus Py2; gi75343291 Cupriavidus necator; gi 1174724 Pseudomonas mendocina; gi 75344585 Pseudomonas aeruginosa; gi75342160 Ralstonia sp.
Figura 17. Dominio oxidoreductasa NAD (NAD_binding_ 1) identificados en proteínas representativas con el programa CCD semejantes al dominio oxidoreductasa NAD (NAD_binding_ 1) presente en la proteína codificada por el gen FHG1. Los códigos de cada secuencia corresponden a los siguientes organismos. 2XNJ_B Escherichia Coli; query- FHG1; gi 122065674 Ustilago maydis; 30ZV_A Ralstonia eutropha H16; 1GVH_A Escherichia coli; gi1174724 Pseudomonas mendocina; gi 1174855 Aliivibrio fischeri; gi 1174854 Photorhabdus luminescens; gi 548358 Neurospora crassa OR74A; gi 128189 Hordeum vulgare.
3.3.2 Localización subcelular de las proteínas FHG
Las secuencias de las cuatro proteínas FHG fueron analizadas utilizando herramientas
bioinformáticas desarrolladas para predecir la localización subcelular de proteínas. En
primer lugar se evaluó la localización subcelular de las proteínas FHG de FOP utilizando el
La Tabla 18 muestra que las proteínas FHG1, FHG2 y FHG3 muestran valores de
probabilidad de localización citoplasmática elevados (superior a 0,8 en lo tres casos). Por
su parte, la proteína FHG4 presenta un valor de probabilidad de localización mitocondrial
muy alto, de 0,79.
Para confirmar la predicción de localización mitocondrial de la proteína FHG4 se analizaron
las cuatro secuencias FHG con otros dos programas de predicción. La Tabla 19 muestra
los valores de probabilidad de localización mitocondrial obtenidos. En primer lugar, el
análisis llevado cabo con TargetP (http://www.cbs.dtu.dk/services/TargetP/) permite
proponer que la proteína FHG4 se localiza en la mitocondria, ya que el valor de probabilidad
de localización mitocondrial es muy alto (0,909). Para las otras tres proteínas FHG los
valores de probabilidad correspondientes son muy bajos. Por su parte, el programa
MITOFATES (http://mitf.cbrc.jp/MitoFates/cgi-bin/) confirma la posible localización
subcelular de FHG4, ya que calcula un valor de probabilidad de 0,980 para la presencia de
una presecuencia mitocondrial en el extremo amino de esta proteína, en la región
correspondiente a la extensión específica característica de esta proteína FHG y ausente
en las otras tres proteínas analizadas. MITOFATES identifica, además, un sitio de corte de
la proteína peptidasa mitocondrial en la posición 22 de la secuencia.
Tabla 19. Valores de probabilidad de localización mitocondrial, estimados con el programa de predicción TargetP, y de presencia de una presecuencia mitocondrial, estimados con el programa MITOFATES, de las proteínas FHG de FOP.
Proteína Probabilidad de localización
mitocondrial (TargetP)
Probabilidad de presecuencia mitocondrial
(MITOFATES)
FHG1 0,103 0,000
FHG2 0,135 0,068
FHG3 0,431 0,181
FHG4 0,909 0,980
Todo ello permite proponer que las proteínas FHG1, FHG2 y FHG3 son proteínas
citoplasmáticas mientras que la proteína FHG4 es una proteína de localización
Figura 18. Árbol filogénetico de las proteinas de tipo flavohemoglobina.
Los números en los nodos indican el valor de boostrap que proporciona la fiabilidad de la intersección. La longitud de la ramas indica la distancia filogénetica. Números de acceso de las secuencias de proteinas utilizadas: AAP41066 Cryptococcus neoformans 1, CAP74387 Botrytis cinerea 1, XP_001588433 Sclerotinia sclerotiorum 1, XP_664773 Aspergillus nidulans 1,
Una vez identificada la presencia de los cuatro genes codificadores de enzimas de tipo
flavohemoglobina, FHG1, FHG2, FHG3 y FHG4, en las estirpes de F. oxysporum f. sp.
phaseoli, nos propusimos determinar la naturaleza de las funciones fisiológicas en las que
la detoxificación del NO pueda jugar un papel relevante en este hongo. Consideramos sus
posibles funciones tanto durante el crecimiento saprofítico del hongo como durante la
interacción del mismo con su huésped. Como una primera fuente de información sobre la
función de cada uno de los cuatro genes FHG decidimos llevar a cabo un análisis de
expresión detallado de los mismos.
3.5.1 Condiciones para el análisis durante el crecimiento saprofítico
y en respuesta a NO
Las enzimas de tipo flavohemoglobina constituyen uno de los mecanismos de
detoxificación de NO más importantes, si no el principal, en los microorganismos, y la
expresión de los genes codificadores de flavohemoglobinas en distintos sistemas es
inducida por exposición a NO exógeno. Se ha comprobado, por otra parte, que el NO juega
un papel relevante en procesos de diferenciación y desarrollo, en particular en la
germinación de las esporas en hongos. Por estas razones decidimos evaluar y cuantificar
inicialmente la expresión de los genes FHG durante el desarrollo y la germinación en F.
oxysporum f. sp. phaseoli durante el ciclo de vida saprofítico. Para lo cual es necesario
determinar previamente las condiciones experimentales más apropiadas para llevar a cabo
esta cuantificación.
Decidimos estudiar dos estirpes diferentes de F. oxysporum f. sp. phaseoli: una estirpe muy
virulenta, la estirpe FOP-SP1, y una estirpe poco virulenta, la estirpe FOP-SP4, y en ambos
casos se evaluaría la eficiencia y la cinética de la germinación de los microconidios en
medio de cultivo líquido. Después de tener en cuenta distintas variables y consideraciones
y de evaluar algunos experimentos preliminares decidimos utilizar de forma rutinaria las
siguientes condiciones experimentales:
1. Como medio de cultivo base se utilizó el medio de cultivo mínimo líquido
suplementado con glucosa 1 % como fuente de carbono (MMG) (ver sección
2.2.2.2 de Materiales y Métodos).
Resultados
- 105 -
2. Para comprobar el efecto de la fuente de nitrógeno sobre la eficiencia de
germinación los medios fueron suplementados en su caso con 10 mM de tartrato
de amonio o 10 mM de nitrato de sodio.
3. Para evaluar el efecto del NO se añadió al medio de cultivo (desde el inicio de la
incubación) 500 µM de nitroprusiato sódico (SNP) como donador de NO.
4. Los efectos de la fuente de nitrógeno y del donador de NO fueron estimados
cuantificando el porcentaje de esporas germinadas a distintos tiempos desde el
inicio del cultivo: 4, 6, 8, 10, 12 y 24 horas.
Se llevaron a cabo tres réplicas biológicas de un experimento de evaluación de germinación
de microconidios de las estirpes FOP-SP1 y FOP-SP4 en las condiciones indicadas. Las
Figuras 19 y 20 presentan los resultados obtenidos.
Fígura 19. Germinación de microconidios de FOP-SP1 en medio minimo con 1 % de glucosa (MMG) y MMG suplementado con 10 mM de nitrato de sodio (MMG+N) o 10 mM de tartrato de amonio (MMG+A). En los tres casos los cultivos se realizaron en ausencia y en presencia de 500 µM de SNP. La evaluación de la germinación se realizó a los tiempos indicados (h). Los histogramas representan los valores medios de tres experimentos y sus desviaciones estándar.
En general la germinación en ambas cepas es similar en las condiciones de cultivo
evaluadas. En cada caso se observa que es indispensable la presencia de una fuente de
nitrógeno para que se active el programa de germinación, y que en ausencia de nitrógeno
el programa de germinación no se inicia o el resultado es mínimo. En los medios
suplementados bien con amonio o con nitrato en ausencia de NO exógeno la germinación
se produce de forma progresiva, iniciándose a las 6 horas. A las 10 horas en medios con
nitrato un 47,54 % de los microconidios en el caso de la cepa FOP-SP1 y un 40,18 % de
Papel de las flavohemoglobinas en F. oxysporum
- 106 -
los microconidios en el caso de FOP-SP4 han germinado. En medios suplementados con
amonio entre estos porcentajes son del 61,94 % y 61,82 %, respectivamente. La eficiencia
de germinación es, por lo tanto, más alta en medios suplementados con amonio. A las 24
horas prácticamente todos los microconidios han germinado en ambas cepas y en ambos
medios, formándose micelio activo en desarrollo.
Las mismas cepas (FOP-SP1 y FOP-SP4) expuestas a NO exógeno muestran un retraso
significativo en la germinación de microconidios en todos los tiempos evaluados. Alas 10
horas de cultivo en medio suplementado con nitrato los porcentajes de germinación son
del 30 % y 16 % para FOP-SP1 y FOP-SP4, respectivamente. En medios suplementados
con amonio los porcentajes son del el 35 % y 26 % de los microconidios han germinado,
respectivamente, mostrando mayor sensibilidad a NO exógeno la cepa silvestre FOP-SP4.
En definitiva, la adición de NO exógeno retrasa significativamente la germinación en las
tres condiciones evaluadas (medios sin N y medios suplementados con amonio o nitrato).
A las 24 horas todos los microconidios han germinado, probablemente porque el NO se ha
degradado. No obstante, las diferencias en la longitud del tubo germinativo en las muestras
tratadas con NO en relación con las muestras no tratadas con NO son evidentes. Las
primeras son siempre inferiores, consecuencia del retraso en la entrada en germinación
que determina la exposición a NO (datos no mostrados).
Figura 20. Germinación de microconidios de FOP-SP4 en medio mínimo suplementado con 1 % de glucosa (MMG) o en MMG suplementado con 10 mM de tartrato de amonio (MMG+A) o 10 mM de nitrato de sodio (MMG+N). En los tres casos los cultivos se realizaron en ausencia y en presencia de 500 µM de SNP. La evaluación de la germinación se realizó a los tiempos indicados (h). Los histogramas representan los valores medios de tres experimentos y sus desviaciones estándar.
Resultados
- 107 -
3.5.2 Análisis de expresión de los genes FHG durante el crecimiento
saprofítico y en respuesta a NO
Teniendo en cuenta estas observaciones decidimos llevar a cabo el análisis de expresión
de los genes FHG en las estirpes FOP-SP1 y FOP-SP4 en dos momentos de desarrollo
diferentes, durante la germinación de las esporas (elegimos el tiempo 9 h por corresponder
a un momento en que el programa de germinación ha comenzado y la germinación es muy
evidente en ausencia de NO) y las 18 h, momento en el cual las esporas han germinado y
el micelio está ramificándose y creciendo activamente. Corresponden a dos estadios de
desarrollo claramente diferentes sobre los que se evaluarán los niveles de expresión de los
genes FHG en respuesta a los factores considerados. Como fuente de Nitrógeno (N) se
seleccionó el tartrato de amonio porque la eficiencia de germinación en su presencia es
superior en todos los tiempos considerados (Ver Figuras 19 y 20).
Los cultivos se realizaron en matraces con 50 ml de medio mínimo con 1 % glucosa (MMG),
suplementado con 10 mM de tartrato de amonio con y sin exposición a NO (SNP) exógeno,
inoculados con 1 x 106 microconidios/ml de las estirpes de F. oxysporum f. sp. phaseoli
(FOP-SP1) y (FOP-SP4) e incubados a 25ºC con agitación constante de 120 rpm. Las
muestras se recolectaron a las 9 y 18 horas en nitrógeno líquido y se conservaron a -80ºC
hasta su utilización.
A partir de estas muestras se obtuvieron las preparaciones de ARN total para el análisis de
expresión de los genes FHG1, FHG2, FHG3 Y FHG4, tanto mediante hibridación Northern
como mediante PCR a tiempo real (RT-qPCR). Para el análisis mediante Northern se
prepararon filtros de nylon a los que se transfirieron 10 µg de ARN total de las muestras de
interés (ver Figuras 21, 22, 23 y 24). Como control de carga se utilizó la intensidad de las
bandas del ARN ribosómico 18S y 28S en geles de agarosa teñidos con bromuro de etidio
antes de ser transferidos a las membranas. Las hibridaciones fueron llevadas a cabo en
todos los casos en condiciones restrictivas. Se utilizaron como sondas de hibridación
amplicones específicos de cada gen, obtenidos mediante PCR con las parejas de
oligonucleótidos mostrados en la Tabla 6 y marcados con digoxigenina. Para el análisis
mediante RT-qPCR se diseñaron oligonucleótidos a partir de las secuencias de las
regiones codificantes de cada uno de los cuatro genes FHG y que permitían amplificar
fragmentos específicos de cada gen (ver Tabla 6). Los datos de expresión relativa fueron
normalizados en relación con la expresión del gen de expresión constitutiva Efα1 (Factor
de elongación alfa 1) y fueron referidos en cada caso al valor de expresión de cada gen en
Papel de las flavohemoglobinas en F. oxysporum
- 108 -
la condición de cultivo durante 9 horas en MM suplementado con 1 % de glucosa (MMG).
Asimismo se comprobó que la eficiencia de amplificación para todas las parejas de
oligonucleótidos fuera superior al 95 %.
3.5.2.1 Expresión de FHG1
La Figura 21 presenta el análisis de expresión del gen FHG1 en las estirpes FOP-SP1 y
FOP-SP4. En ambas estirpes la situación observada es esencialmente la misma. Se
comprueba que FHG1 no se expresa a niveles detectables mediante Northern en
ausencia de NO y que su expresión se induce a niveles elevados (detectables mediante
hibridación Northern) por exposición a NO. Esta inducción sólo se produce en la fase de
germinación, no en la fase de desarrollo del micelio, lo que supone que o bien el micelio
es insensible a NO o bien en ese momento ya se ha degradado el NO añadido al medio
de cultivo inicialmente.
Figura 21. Análisis de expresión del gen FHG1 de las estirpes FOP-SP1 (Panel A) y FOP-SP4 (panel B) durante el desarrollo y en condiciones de estrés nitrosativo. En cada panel la figura de la izquierda presenta los resultados de una hibridación Northern blot y el histograma de la derecha los resultados de la cuantificación de la expresión mediante RT-qPCR. Las muestras de RNA utilizadas en ambos tipos de análisis corresponden a muestras derivadas de esporas cultivadas durante 9 o 18 h en medio mínimo suplementado con 1 % glucosa (M) o medio mínimo suplementado con 1 % glucosa y con 10 mM de tartrato amónico (M+A), en ausencia o en presencia de 500 µM de SNP(+S). Los datos de RT-qPCR mostrados representan las medias de 3 experimentos (réplicas biológicas). Los datos de expresión representan los niveles de expresión relativa del gen FHG1 normalizados en relación con el nivel de expresión del gen Efα1 y referidos a los valores de expresión de FHG1 a 9 horas en medio M (MMG).
Resultados
- 109 -
El análisis de expresión mediante qPCR confirma el patrón detectado mediante hibridación
Northern, permitiendo esta técnica realizar una cuantificación de los niveles de expresión
del gen en relación con una condición de referencia (los valores representados muestran
los valores medios de expresión en tres réplicas biológicas independientes). Se comprueba
que el patrón de expresión de FHG1 es muy parecido en ambas estirpes y que en ambos
casos sólo hay un aumento importante de expresión durante la fase de germinación y en
respuesta a NO exógeno. El nivel de inducción es mayor en la estirpe FOP-SP4
(aproximadamente 20X en MMG en presencia de NO y 30X en MMG+A en presencia de
NO) que en la estirpe FOP-SP1 (6X y 7X, respectivamente), pero la respuesta en ambas
estirpes es esencialmente la misma.
3.5.2.2 Expresión de FHG2
El análisis del patrón de expresión del gen FHG2 en las condiciones indicadas se presenta
en la Figura 22. También en este caso el perfil de expresión es esencialmente el mismo en
ambas estirpes. El análisis comparado entre ellas mediante hibridación Northern blot
resulta menos informativo en primera aproximación porque la intensidad de las bandas de
hibridación detectadas en el caso de la estirpe FOP-SP1 es, en conjunto, menor, pero
ciertamente el perfil de expresión en ambas estirpes es similar.
Tomando como sistema de referencia la estirpe FOP-SP4, se comprueba que la expresión
de FHG2 está regulada durante el desarrollo. Ésta es máxima en micelio cultivado durante
18 h en medio mínimo con glucosa y en ausencia de donador de NO, y resulta reprimida
en presencia de amonio, condición, debemos recordar, que estimula la germinación. Es
interesante destacar que en la fase de micelio, cuando éste es cultivado en medio mínimo
con glucosa la exposición a NO reprime la expresión de FHG2 (o anula la inducción
observada en ausencia de NO). En presencia de amonio, condición en la que la expresión
de FHG2 es reprimida, la exposición a NO sí tiene un efecto inductor de la expresión.
Durante la fase de germinación de las esporas el perfil de expresión es diferente.
Considerando los datos de cuantificación mediante RT-qPCR se comprueba que en
ausencia de NO exógeno la adición de amonio al medio de cultivo también reprime la
expresión de FHG2, aunque la magnitud de la represión es menor que en la fase de micelio
en desarrollo. Sin embargo, tanto en presencia como en ausencia de amonio la adición de
NO exógeno induce la expresión de FHG2.
Papel de las flavohemoglobinas en F. oxysporum
- 110 -
Es resumen, en el caso de FHG2 se detecta una regulación de la expresión durante el
desarrollo (probablemente consecuencia de una sensibilidad diferente al NO en las dos
fases consideradas, esporas en germinación y micelio) sobre la cual se superpone una
regulación por fuente de nitrógeno. Ésta determina que en la fase de micelio la adición de
amonio reprima la expresión de FHG2 y que también lo haga la adición de NO en ausencia
de amonio que determina que la adición de amonio reprima la expresión de FHG2 y que la
exposición a NO en presencia de amonio aumente la expresión de FHG2. Sólo en el caso
de la fase de desarrollo de micelio y en ausencia de amonio la exposición a NO no
determina un aumento de la expresión de FHG2.
Figura 22. Análisis de expresión del gen FHG2 de las estirpes FOP-SP1 (Panel A) y FOP-SP4 (panel B) durante el desarrollo y en condiciones de estrés nitrosativo. En cada panel la figura de la izquierda presenta los resultados de una hibridación Northern blot y el histograma de la derecha los resultados de la cuantificación de la expresión mediante RT-qPCR. Las muestras de RNA utilizadas en ambos tipos de análisis corresponden a muestras derivadas de esporas cultivadas durante 9 o 18 h en medio mínimo suplementado con 1 % glucosa (M) o medio mínimo suplementado con 1 % glucosa y con 10 mM de tartrato amónico (M+A), en ausencia o en presencia de 500 µM de SNP (+S). Los datos de RT-qPCR mostrados representan las medias de 3 experimentos (réplicas biológicas). Los datos de expresión representan los niveles de expresión relativa del gen FHG2 normalizados en relación con el nivel de expresión del gen Efα1 y referidos a los valores de expresión de FHG2 a 9 horas en medio M (MMG).
3.5.2.3 Expresión de FHG3
Los patrones de expresión de FHG3 en las estirpes FOP-SP1 y FOP-SP4 se presentan en
la Figura 23. Los patrones de expresión en ambas estirpes son similares. Aunque es
posible considerar algunas diferencias en la magnitud de los valores de expresión
Resultados
- 111 -
(inducción) entre ambas cepas, el comportamiento general que muestran una y otra en las
condiciones analizadas es similar. En este caso las esporas en germinación y el micelio en
desarrollo responden de forma similar, en particular en la estirpe FOP-SP1, lo que supone
que no existe una regulación obvia por desarrollo. Sí se observa una regulación por
amonio, cuya presencia estimula la expresión de FHG3, de forma más intensa en la fase
de esporas en germinación, y no resulta obvia, en general, una regulación por exposición
a NO. Sólo se observa una diferencia importante en el comportamiento de las dos cepas,
y es en la fase de micelio en la estirpe FOP-SP4, que parece responder de manera más
intensa a la exposición a NO en presencia de amonio. No obstante, es necesario tener en
cuenta que en esta condición lo que parece producirse es un menor nivel de inducción por
amonio, lo que acentúa la diferencia en los niveles de expresión en presencia de amonio y
de NO. Este comportamiento marca una diferencia, que podemos considerar menor, entre
las dos estirpes analizadas. En cualquier caso, el análisis conjunto de los datos obtenidos
con los dos procedimientos experimentales utilizados indica que en las dos estirpes la
presencia de amonio determina una inducción de la expresión de FHG3, más evidente
durante la fase de germinación de las esporas, y que la exposición a NO no tiene un efecto
notable en la regulación de la expresión de FHG3.
Figura 23. Análisis de expresión del gen FHG3 de las estirpes FOP-SP1 (Panel A) y FOP-SP4 (panel B) durante el desarrollo y en condiciones de estrés nitrosativo. En cada panel la figura de la izquierda presenta los resultados de una hibridación Northern blot y el histograma de la derecha los resultados de la cuantificación de la expresión mediante RT-qPCR. Las muestras de RNA utilizadas en ambos tipos de análisis corresponden a muestras derivadas de esporas cultivadas durante 9 o 18 h en medio mínimo suplementado con 1 % glucosa (M) o medio mínimo suplementado con 1 % glucosa y con 10 mM de tartrato amónico (M+A), en ausencia o en presencia de 500 µM de SNP
Papel de las flavohemoglobinas en F. oxysporum
- 112 -
(+S). Los datos de RT-qPCR mostrados representan las medias de 3 experimentos (réplicas biológicas). Los datos de expresión representan los niveles de expresión relativa del gen FHG3 normalizados en relación con el nivel de expresión del gen Efα1 y referidos a los valores de expresión de FHG3 a 9 horas en medio M (MMG).
3.5.2.4 Expresión de FHG4
Finalmente, se analizó la expresión del gen FHG4 en las mismas condiciones. En este caso
los resultados obtenidos tanto por Northern blot como por RT-qPCR indican que el gen se
expresa a niveles mínimos en todas las condiciones analizadas y que no responde, a nivel
transcripcional, a ninguno de los factores evaluados en nuestro análisis. Probablemente
FHG4 codifica efectivamente una proteína de tipo flavohemoglobina, pero los procesos en
los que pueda desempeñar una función dada, sea ésta cual sea, no parecen estar
relacionados con desarrollo, con respuesta a amonio y con exposición a NO (respuesta a
condiciones de estrés nitrosativo).
Figura 24. Análisis de expresión del gen FHG4 de las estirpes FOP-SP1 (Panel A) y FOP-SP4 (panel B) durante el desarrollo y en condiciones de estrés nitrosativo. En cada panel la figura de la izquierda presenta los resultados de una hibridación Northern blot y el histograma de la derecha los resultados de la cuantificación de la expresión mediante RT-qPCR. Las muestras de RNA utilizadas en ambos tipos de análisis corresponden a muestras derivadas de esporas cultivadas durante 9 o 18 h en medio mínimo suplementado con 1 % glucosa (M) o medio mínimo suplementado con 1 % glucosa y con 10 mM de tartrato amónico (M+A), en ausencia o en presencia de 500 µM de SNP (+S). Los datos de RT-qPCR mostrados representan las medias de 3 experimentos (réplicas biológicas). Los datos de expresión representan los niveles de expresión relativa del gen FHG4 normalizados en relación con el nivel de expresión del gen Efα1 y referidos a los valores de expresión de FHG3 a 9 horas en medio M (MMG).
Resultados
- 113 -
3.5.3 Análisis de expresión de los genes FHG en plantas de judía
infectadas con estirpes de F. oxysporum f. sp. phaseoli
Los patrones de expresión de los cuatro genes FHG en relación con los factores analizados
son diferentes, lo que permite deducir que las proteínas codificadas pueden participar en
procesos distintos. En el contexto de nuestro trabajo estamos interesados de forma
particular en la posible participación de las enzimas de tipo flavohemoglobina en desarrollo
y en patogenicidad. Una vez analizados los patrones de expresión de los cuatro genes
FHG durante el crecimiento saprofítico decidimos abordar a continuación el estudio de los
niveles de expresión de los genes FHG durante la interacción del patógeno con su
huésped.
Decidimos realizar ensayos de infección sobre plantas de judía con las estirpes silvestres
de F. oxysporum FOP-SP1 y FOP-SP4 utilizando un sistema de cultivo hidropónico con el
medio de cultivo de plantas Plant growth medium PGM (descrito en la sección 2.2.1. de
Materiales y Métodos) por considerarse un sistema de cultivo limpio y libre de materia
orgánica y organismos parásitos y patógenos. Además mediante este tipo de cultivo los
experimentos son independientes de fenómenos meteorológicos como luz, temperatura y
humedad relativa, obteniéndose además muestras biológicas libres de contaminación. Otra
de las ventajas es la facilidad para realizar un seguimiento continuo del estado de
desarrollo de la planta y del progreso de la enfermedad. En ningún caso el progreso de la
enfermedad es igual al observado en experimentos de infección realizados en macetas con
medio de cultivo sólido. Ésta progresa siempre más lentamente en cultivos hidropónicos.
No obstante, la distinción entre plantas inoculadas con la estirpe muy virulenta y con la
estirpe poco virulenta, tanto en el número de hojas cloróticas como en el grado de necrosis
en el sistema vascular, puede hacerse de manera muy similar en ambos condiciones de
cultivo de las plantas.
Las raíces de plántulas de 10 días de edad se lavaron con agua destilada estéril y con
tijeras estériles se cortaron los ápices radiculares. A continuación éstos fueron sumergidos
en una solución de esporas 1 x 106 esporas/ml durante 5 minutos. Seguidamente se
retiraron y se traspasaron a tubos falcon de 50 ml con 30 ml de medio de cultivo líquido
(PGM), tubos que fueron cubiertos con papel aluminio y sellados con parafilm para evitar
la transpiración y con el objeto de mantener constante el volumen de medio de cultivo. Los
cultivos hidropónicos fueron incubados a 25ºC en un fitotrón a 60-80 % de humedad relativa
con un fotoperiodo de 16 horas luz y 8 horas oscuridad. La evaluación se realizó tomando
Papel de las flavohemoglobinas en F. oxysporum
- 114 -
muestras de raíz a los 1, 2 y 3 días después de la inoculación, muestras de hipocotilo a los
5 y 7 días después de la inoculación y muestras de tallo a los 14 y 21 días después de la
inoculación. Las muestras de plantas inoculadas con F. oxysporum f. sp. phaseoli fueron
colectadas y conservadas a -80ºC. El ARN total se extrajo siguiendo el método descrito en
la sección 2.6.4.2 de Materiales y Métodos. La calidad del ARN se evaluó por electroforesis
en gel de agarosa en TAE 1X y por espectrofotometría en el NanoDrop N-1000.
En base a los resultados obtenidos en el análisis de expresión de los genes de FHG durante
el crecimiento saprofítico mediante análisis tipo Northern y RT-qPCR se decidió evaluar la
expresión de estos genes en la interacción con el huésped mediante RT-qPCR. La síntesis
de cDNA y el procedimiento de preparación de las reacciones de RT-qPCR se realizaron
siguiendo los procedimientos descritos en la sección 2.12.2 y 2.12.3 de Materiales y
Métodos. Los oligonucleótidos utilizados en reacciones de RT-qPCR son los que se
utilizaron para los análisis de expresión durante el crecimiento saprofitico descritos en las
secciones 3.5.1 y 3.5.2 de Resultados.
3.5.3.1 Análisis de expresión de los genes FHG en plantas de
judía infectadas con FOP-SP1
En la Figura 25 se presentan los patrones de expresión de los genes FHG de la estirpe
FOP-SP1 durante la interacción con plantas de judía variedad “blanca riñón”. En el caso
de FHG1 se observa un aumento en su nivel de expresión en raíz a los 3 dpi, duplicándose
éste en relación con la situación de refencia considerada, 1 dpi. En hipocotilo a los 7 dpi el
nivel de expresión detectado ha aumentado aún más (se triplica en relación con la situación
de referencia) y parece mantenerse constante en fases posteriores del proceso de
infección, concretamente a los 14 dpi en tallo (Panel A). En el caso de FHG2 el análisis
llevado a cabo sugiere que también parece aumentar, aunque en menor medida, su nivel
de expresión desde el día 3 dpi. Sin embargo, el análisis estadístico demuestra que las
diferencias observadas no son significativas (Panel B). La cuantificación de los niveles de
expresión de los genes FHG3 (Panel C) y FHG4 (Panel D) indica que ésta es muy uniforme
a lo largo del proceso de infección, no observándose variaciones notables en los tiempos
considerados en este estudio para ninguno de los dos genes.
Resultados
- 115 -
Figura 25. Análisis de expresión in planta mediante RT-qPCR de los genes FHG de FOP-SP1. (A): gen FHG1; (B): gen FHG2; (C): gen FHG3; y (D): gen FHG4. Los números que identifican las barras muestran los tiempos (días) después de la inoculación (dpi) en los que fueron tomadas las muestras: 1 y 3: Raíz, 7: hipocotilo y 14: tallo. Los datos presentados muestran los valores medios (± DE) de tres experimentos independientes. Los niveles de expresión de cada gen fueron normalizados tomando como referencia el nivel de expresión del gen Efα1 (Factor de elongación alpha 1), considerado de expresión constitutiva. Los niveles de expresión de cada gen en cada tiempo están referidos a su nivel de expresión en la muestra tomada 1 dpi. Diferencias significativas (“t-test” de Student) fueron observadas en las condiciones “3”; “7” y “14” dpi (** p<0.001) en el análisis de expresión del gen FHG1.
3.5.3.2 Análisis de expresión de los genes FHG en plantas de
judía infectadas conFOP-SP4
En la Figura 26 se presenta el análisis de expresión de los genes FHG de la estirpe FOP-
SP4 durante su interacción con plantas de judía variedad “blanca riñón”. Se comprueba
que la situación es muy similar a la observada en el caso de infección producida por la
estirpe FOP-SP1. Sólo se observan diferencias significativas en los niveles de expresión
Papel de las flavohemoglobinas en F. oxysporum
- 116 -
en relación con la condición de referencia (1 dpi) en el caso del gen FHG1, cuya expresión
aumenta en raíz a los 3 dpi y lo hace aún más en hipocotilo a los 7 dpi (Panel A). Este
aumento no parece mantenerse a lo largo de tiempo, marcando una diferencia menor en
comparación con lo que sucede en la interacción con FOP-SP1, ya que decae a los 14 dpi.
En este sistema los niveles de expresión de los genes FHG2, FHG3 y FHG4 no muestran
variaciones a lo largo del tiempo durante la interacció10n (Paneles B, C y D).
Figura 26. Análisis de expresión in planta mediante qPCR de los genes FHG de FOP-SP4. (A): gen FHG1; (B): gen FHG2; (C): gen FHG3; y (D): gen FHG4. Los números que identifican las barras muestran los tiempos (días) después de la inoculación (dpi) en los que fueron tomadas las muestras: 1 y 3: Raíz; 7: hipocotilo; y 14: tallo. Los datos presentados muestran los valores medios (± SD) de tres experimentos independientes. Los niveles de expresión de cada gen fueron normalizados tomando como referencia el nivel de expresión del gen Efα1 (Factor de elongación alpha 1), considerado de expresión constitutiva. Los niveles de expresión de cada gen en cada tiempo están referidos a su nivel de expresión en la muestra tomada 1 dpi. Diferencias significativas (“t-test” de Student) fueron observadas en las condiciones “3” y “14” dpi (* p<0,05) y “7” dpi (** p<0.001) en el análisis de expresión del gen FHG1.
Resultados
- 117 -
3.6 Análisis funcional de los genes FHG
Los análisis de expresión durante el crecimiento saprofítico en medio de cultivo indican que
los cuatro genes codificadores de enzimas de tipo flavohemoglobina de F. oxysporum f. sp
phaseoli muestran patrones diferentes lo que permite proponer que las proteínas
codificadas puedan participar en procesos fisiológicos distintos. Con el objeto de obtener
información sobre los procesos en los que cada una de las cuatro proteínas FHG pudiera
participar decidimos llevar a cabo la caracterización funcional de los genes codificadores
de las mismas mediante la obtención y caracterización de mutantes deficientes de cada
uno de ellos. Para generar mutantes deficientes en cada uno de los genes se diseñó una
estrategia basada en el reemplazamiento génico del alelo silvestre correspondiente por un
alelo mutante en el que la región estructural completa del gen, o una parte del mismo, ha
sido sustituida por una fusión génica que permite la expresión del gen de resistencia a
higromicina B bajo el control de secuencias reguladoras funcionales en hongos
filamentosos. Esta fusión génica queda flanqueada en el plásmido de reemplazamiento
génico por secuencias de ADN genómico específicas del gen de interés, en cada caso
derivadas de las regiones inmediatamente adyacentes a los extremos 5’ y 3’ de la región
codificante.
El esquema de trabajo en todos los casos fue el mismo. Para cada gen, a partir de ADN
genómico de FOP-SP1 se amplificó un fragmento de ADN de aproximadamente 1 kb de
tamaño de la región flanqueante 5’ y un segundo fragmento de ADN del mismo tamaño de
la región flanqueante 3’. Estos fragmentos se amplificaron con oligonucleótidos específicos
diseñados a tal efecto. Para cada gen se construyó un plásmido binario clonando los
fragmentos flanqueantes 5’ y 3’ específicos a ambos lados del casete de expresión de la
higromicina. Con cada uno de estos plásmidos se transformó A. tumefaciens por
electroporación y con las cepas transformantes seleccionadas en cada caso se llevó a cabo
una transformación de FOP-SP1 y de FOP-SP4. Posteriormente, entre los transformantes
obtenidos se identificaron mediante PCR y mediante hibridación Southern aquéllos en los
que el alelo silvestre del gen considerado había sido reemplazado por el alelo mutante
construído en cada caso.
3.6.1 Construcción de los vectores de transformación
Para la construcción de los vectores de transformación previamente se amplificaron
mediante PCR fragmentos de ADN de las regiones flanqueantes 5´y 3´ de los genes FHG1,
Papel de las flavohemoglobinas en F. oxysporum
- 118 -
FHG2, FHG3 y FHG4. Para amplificar los fragmentos de ADN genómico se diseñaron
oligonucleótidos específicos que permitían amplificar fragmentos de ADN de
aproximadamente 1 kb de tamaño. La Tabla 20 presenta los códigos de los
oligonucleótidos diseñados para llevar a cabo estas amplificaciones y los tamaños exactos
de los fragmentos amplificados. Las posiciones de anillamiento de estos oligonucleótidos
se muestran en las Figuras 29B y C, 31B y C, 33B y C y 35B y C. Las secuencias de todos
los oligonucleótidos están recogidas en la Tabla 6 de la sección 2.12.5 de Materiales y
Métodos.
Tabla 20. Códigos de los oligonucleótidos derivados de los genes FHG diseñados para la contrucción de los vectores de reemplazamiento génico y tamaños de los fragmentos amplificados por PCR.
Gen Región
flanqueante Código Tamaño
Gen FHG1
5’ FHG175
RHG175 996 pb
3’ FHG173
RHG173 998 pb
Gen FHG2
5’ FHG135
RHG135 946 pb
3’ FHG133
RHG133 948 pb
Gen FHG3
5’ FHG185
RHG185 1065 pb
3’ FHG183
RHG183 988 pb
Gen FHG4
5’ FHG155
RHG155 1037 pb
3’ FHG153
RHG153 1012 pb
En la Figura 27 se presentan los productos de amplificación obtenidos en las reacciones
de amplificación llevadas a cabo y visualizados mediante electroforesis en gel de agarosa.
Resultados
- 119 -
Figura 27. Productos de PCR obtenidos a partir de ADN genómico de FOP-SP1 de las regiones flanqueantes 5´ (carriles 1) y 3´ (carriles 2) de los genes FHG con los oligonucleótidos específicos en cada caso. (A): FHG1, (B): FHG2, (C): FHG3 y (D): FHG4. Las combinaciones de cebadores utilizadas en cada amplificación son las indicadas en la Tabla 20. Los productos de PCR fueron resueltos mediante electroforesis en geles de agarosa al 1 %. M: marcador de peso molecular 1 kp plus DNA ladder (invitrogen).
Los productos de PCR de cada uno de los genes fueron purificados y ligados en el vector
pRF-HU2 previamente digerido con las enzimas de restricción PacI y NT.BbvCI utilizando
el sistema User Friendly Cloning. Con las reacciones de ligación se llevaron a cabo
transformaciones de células de E. coli cepa DH5α mediante choque térmico. Para
identificar colonias portadoras del plásmido de interés se llevó a cabo un escrutinio
mediante PCR directa sobre colonia de una muestra representativa de las colonias
obtenidas utilizando en cada caso los oligonucleótidos utilizados para amplificar los
fragmentos de ADN de las regiones flanqueantes correspondientes. Se seleccionó
finalmente una colonia positiva portadora del plásmido deseado en cada caso. Los
plásmisdos generados fueron denominados pRF-HU2-FHG1, pRF-HU2-FHG2, pRF-HU2-
FHG3 y pRF-HU2-FHG4. La Figura 28 muestra los mapas de restricción de estos
plásmidos.
Papel de las flavohemoglobinas en F. oxysporum
- 120 -
Figura 28. Mapa de restricción de los plásmidos construidos a partir del plásmido PRF-HU2 para el reemplazamiento génico de los genes FHG de FOP-SP1 y FOP-SP4. En cada caso el casete de resistencia a higromicina está flanqueado por fragmentos de ADN genómico específicos de las regiones flanqueantes en 5’ y en 3’ de los genes FHG1 (plásmido pRF-HU2-FHG1) (A), FHG2 (plásmido pRF-HU2-FHG2) (B), FHG3 (plásmido pRF-HU2-FHG3) (C) y FHG4 (plásmido pRF-HU2-FHG4) (D).
3.6.2 Transformación de A. tumefaciens
Los plásmidos pRF-HU2-FHG1, pRF-HU2-FHG2, pRF-HU2-FHG3 y pRF-HU2-FHG4
fueron introducidos en A. tumefacción cepa AGL1 mediante electroporación siguiendo los
procedimientos descritos en la sección 2.9.5.2 de Materiales y Métodos. De entre las
colonias transformantes obtenidas con cada plásmido se seleccionó una que fue utilizada
para llevar a cabo la transformación de FOP-SP1 y FOP-SP4. La identidad del plásmido
presente en cada una de las colonias seleccionadas fue verificada mediante PCR utilizando
oligonucleótidos específicos de las regiones flanqueantes de cada uno de los cuatro genes.
Resultados
- 121 -
3.6.3 Transformación de F. oxysporum f. sp. phaseoli mediada por A.
tumefaciens
La transformación de F. oxysporum mediada por A. tumefaciens se realizó siguiendo la
metodología mostrada en los apartados 2.9.5.3 y 2.9.5.4 de Materiales y Métodos. Se
transformó tanto la estirpe FOP-SP1 como la estirpe FOP-SP4. Los transformantes
primarios que fueron apareciendo en las placas de medio selectivo en presencia de
higromicina B fueron transferidos individualmente a placas de PDA con 75 µg/ml de
higromicina e incubados durante 5 días a 25ºC. En la estirpe muy virulenta FOP-SP1 se
seleccionaron 96 transformantes para el gen FHG1, 72 transformantes para el gen FHG2,
102 transformantes para el gen FHG3 y 85 transformantes para el gen FGH4. En la estirpe
poco virulenta FOP-SP4 se seleccionaron 92 transformantes para el gen FHG1, 86
transformantes para el gen FHG2, 110 transformantes para el gen FHG3 y 112
transformantes para el gen FHG4.
3.6.4 Identificación de mutantes por reemplazamiento génico de F.
oxysporum f. sp. phaseoli
Para el análisis de los transformantes primarios y la identificación de mutantes por
reemplazamiento génico se partió de micelio tomado de placas PDA cultivadas durante 5
días. El micelio se transfirió a tubos eppendorf de 1,5 ml con Tris-HCl 10X. Este material
fue utilizado para la extracción de ADN genómico haciendo uso del método de extracción
rápido descrito en la sección 2.6.1.2 de Materiales y Métodos. El ADN genómico de cada
transformante fue utilizado para identificar mediante PCR de forma rápida transformantes
en los cuales se pudieran haber producido eventos de recombinación homóloga tanto en
el extremo flanqueante 5’ como en el extremo flanqueante 3’ entre el alelo silvestre del gen
y el alelo mutante en cada caso clonado en el plásmido de transformación correspondiente.
Para la discriminación de transformantes “candidato”, en cada uno de los de los cuatro
genes analizados se diseñaron oligonucleótidos derivados de secuencias localizadas en
las regiones flanqueantes, bien 5’ bien 3’, pero inmediatamente aguas arriba o aguas abajo
de los fragmentos de ADN clonados en el plásmido de reemplazamiento génico
correspondiente. Los códigos de los oligonucleótidos diseñados se presentan en la Tabla
21 y las posiciones que ocupan las secuencias diana de hibridación de los mismos se
muestran en las Figuras 29B y C, 31B y C, 33B y C y 35B y C. Sus secuencias se presentan
en la Tabla 6 de la sección 2.12.5 de Materiales y Métodos.
Papel de las flavohemoglobinas en F. oxysporum
- 122 -
Tabla 21. Códigos de los oligonucleótidos derivados de los genes FHG y del gen hph utilizados para la identificación de transformantes candidato y tamaños de los productos de PCR esperados en caso de ocurrencia de recombinación homóloga en la región correspondiente.
Gen Región
flanqueante Código Tamaño
Gen FHG1
5’ FHG175-KN-FW
HphBC 2640 pb
3’ RHG173-KN-RV
HphAC 1838 pb
Gen FHG2
5’ FHG135-KN-FW
HphBC 2535 pb
3’ RHG133-KN-RV
HphAC 2462 pb
Gen FHG3
5’ FHG185-KN-FW
HphBC 2431 pb
3’ RHG183-KN-RV
HphAC 1945 pb
Gen FHG4
5’ FHG155-KN-FW
HphBC 2421 pb
3’ RHG153-KN-RV
HphAC 2097 pb
Para cada uno de los genes, la amplificación en un transformante primario dado de un
fragmento de ADN del tamaño esperado (conocido para cada gen en concreto) con una
combinación de oligonucleótidos particular que incluye el oligonucleótido derivado de la
región inmediatamente aguas arriba del fragmento de ADN genómico clonado en el
plásmido de reemplazamiento génico y correspondiente a la región flanqueante 5’ y el
oligonucleótido HphBC, derivado del casete de resistencia a higromicina, permitiría
identificar un evento de recombinación homóloga en el extremo 5’ del gen de interés entre
el alelo silvestre y el alelo mutante. De la misma manera, la amplificación en un
transformante primario dado de un fragmento de ADN del tamaño esperado (conocido para
cada gen en concreto) con una combinación de oligonucleótidos particular que incluye el
oligonucleótido derivado de la región inmediatamente aguas abajo del fragmento de ADN
genómico clonado en el plásmido de reemplazamiento génico y derivado de la región
flanqueante 3’ y el oligonucleótido HphAC, derivado del casete de resistencia a higromicina,
permitiría identificar un evento de recombinación homóloga en el extremo 3’ del gen de
interés del alelo silvestre y el alelo mutante (ver en las Figuras 29, 31, 33 y 35 la ubicación
Resultados
- 123 -
de los cebadores representados en color naranja). La amplificación simultánea de ambos
productos de amplificación permitiría identificar transformantes primarios derivados de un
evento de reemplazamiento génico que serían seleccionados para análisis posteriores.
3.6.4.1 Identificación de transformantes del gen FHG1
En la Figura 29 se presenta de forma esquemática el modo en el que deben alinearse el
alelo silvestre del gen FHG1 y la construcción generada y clonada en el plásmido pRF-
HU2-FHG1 y la estructura que presentará el alelo mutante del gen FHG1 derivado de dos
eventos de recombinación homóloga en ambas regiones flanqueantes de la región
estructural del gen, es decir, la estructura del alelo mutante de FHG1 consecuencia del
remplazamiento génico.
Figura 29. Estrategia de reemplazamiento génico del gen FHG1. A: Representación simplificada del plásmido pRF-HU2-FHG1 con la construcción de reemplazamiento del gen FHG1. B: Esquema del entorno genómico del alelo silvestre del gen FHG1. Se indican (cajas de color azul intenso) las regiones en los extremos flanqueantes 5’ y 3’ en las que debe producirse recombinación homóloga para que tenga lugar el reemplazameinto génico de FHG1. C: Estructura del alelo mutante generado mediante reemplazamiento génico como consecuencia de un doble evento de recombinación homóloga en las regiones flaqueantes 5’ y 3’ de FHG1. La ubicación de las dianas de hibridación de los cebadores utilizados para amplificación de las regiones flanqueantes 5´ y 3´ clonadas en el plásmido pRF-HU2-FHG1 se representa mediante flechas de color verde (B). La ubicación de las diana de anillamiento de los cebadores utilizados para la detección de la ocurrencia de recombinación homóloga en una región flanqueante u otra se representa mediante flechas de color naranja (B y C). Se indica la posición de las dianas de restricción de la enzima HindIII en el alelo silvestre y en el alelo resultante del remplazamiento génico (HindIII no corta el plásmido pRF-HU2-FHG1).
Papel de las flavohemoglobinas en F. oxysporum
- 124 -
La identificación de “transformantes candidatos” en los que se ha producido el
reemplazamiento génico del gen FHG1 se realizó mediante reacciones de PCR utilizando
la combinación de cebadores FHG175-KN-FW y HphBC y RHG173-KN-RV y HphAC
mostrados en la Figura 29B y 29C. En el primer caso, la amplificación de un fragmento de
2640 pares bases correspondiente a la región flanqueante 5´ sería indicativa de
recombinación homóloga en esta región. En el segundo caso, la amplificación de un
fragmento de 1838 pares de bases correspondiente a la región flanqueante 3´ sería
indicativa de recombinación homóloga en esta región. En la Figura 30, se muestran
resueltos mediante electroforesis en geles de agarosa los productos de amplificación
obtenidos en reacciones de PCR llevadas a cabo con ambas combinaciones de cebadores
en una muestra representativa de transformantes. En un buen número de transformantes
primarios se amplificaron los dos fragmentos diagnósticos de recombinación homóloga.
Estos son transformantes en los que debe haberse producido el reemplazamiento del gen
FHG1. En total, para la estirpe muy virulenta FOP-SP1 se obtuvieron 16 transformantes
positivos candidatos de 96 analizados. Para la estirpe poco virulenta FOP-SP4 se
obtuvieron 22 transformantes positivos de 92 analizados. En la Figura 30 se muestran los
productos de PCR obtenidos a partir de ADN genómico de las cepas mutantes del gen
FHG1 de FOP-SP1 y FOP-SP4.
Figura 30. Productos de PCR obtenidos en reacciones de PCR llevadas a cabo con los oligonucleótidos FHG175-KN-FW y HphBC (línea superior en cada gel) (5’) y RHG173-KN-RV y HphAC (línea inferior en cada gel) (3’) utilizando como molde ADN genómico de los transformantes obtenidos con el plásmido pRF-HU2-FHG1. A: Transformantes FOP-SP1 (carreras 1 a 18) y B: Transformantes FOP-SP4 (carreras 1 a 18). Cómo control se incluyeron reacciones de PCR llevadas a cabo con los mismos oligonucleótidos utilizando como molde ADN genómico de la estirpes FOP-SP1 o FOP-SP4 no transformadas. M: marcador de peso molecular 1 kp plus DNA ladder (Invitrogen).
Resultados
- 125 -
3.6.4.2 Identificación de transformantes del gen FHG2 En la Figura 31 se presenta de forma esquemática el modo en el que deben alinearse el
alelo silvestre del gen FHG2 y la construcción generada y clonada en el plásmido pRF-
HU2-FHG2 y la estructura que presentará el alelo mutante del gen FHG2 derivado de dos
eventos de recombinación homóloga en ambas regiones flanqueantes de la región
estructural del gen, es decir, la estructura del alelo mutante de FHG2 consecuencia del
remplazamiento génico.
Figura 31. Estrategia de reemplazamiento génico del gen FHG2. A: Representación simplificada del plásmido pRF-HU2-FHG2 con la construcción de reemplazamiento del gen FHG2. B: Esquema del entorno genómico del alelo silvestre del gen FHG2. Se indican las regiones en los extremos flanqueantes 5’ y 3’ en las que debe producirse recombinación homóloga para que tenga lugar el reemplazameinto génico de FHG2. C: Estructura del alelo mutante generado mediante reemplazamiento génico como consecuencia de un doble evento de recombinación homóloga en las regiones flaqueantes 5’ y 3’ de FHG2. La ubicación de las dianas de hibridación de los cebadores utilizados para amplificación de las regiones flanqueantes 5´ y 3´ clonadas en el plásmido pRF-HU2-FHG2 se representa mediante flechas de color verde (B). La ubicación de las dianas de anillamiento de los cebadores utilizados para la detección de la ocurrencia de recombinación homóloga en una región flanqueante u otra se representa mediante flechas de color naranja (B y C). Se indica la posición de las dianas de restricción de la enzima HindIII en el alelo silvestre y en el alelo resultante del remplazamiento génico (HindIII no corta el plásmido pRF-HU2-FHG2).
La identificación de “transformantes candidatos” en los que se ha producido el
reemplazamiento génico del gen FHG2 se realizó mediante reacciones de PCR utilizando
la combinación de cebadores FHG135-KN-FW y HphBC y RHG133-KN-Rv y HphAC
pRF-HU2-FHG1
Alelo WT FHG1
Alelo mutante
Papel de las flavohemoglobinas en F. oxysporum
- 126 -
mostrados en la Figura 31B y 31C. En el primer caso, la amplificación de un fragmento de
2537 pares bases correspondiente a la región flanqueante 5´ sería indicativa de
recombinación homólogo en esta región. En el segundo caso, la amplificación de un
fragmento de 2462 pares de bases correspondiente a la región flanqueante 3´ sería
indicativa de recombinación homóloga en esta región.
En la Figura 32 se muestran resueltos mediante electroforesis en geles de agarosa los
productos de amplificación obtenidos en reacciones de PCR llevadas a cabo con ambas
combinaciones de cebadores en una muestra representativa de transformantes. También
en este caso en un buen número de transformantes primarios se amplificaron los dos
fragmentos diagnósticos de recombinación homóloga.
Figura 32. Productos de PCR obtenidos en reacciones de PCR llevadas a cabo con los oligonucleótidos FHG135-KN-FW y HphBC (línea superior en cada gel) (5’) y RHG133-KN-RV y HphAC (línea inferior en cada gel) (3’) utilizando como molde ADN genómico de los transformantes obtenidos con el plásmido pRF-HU2-FHG2. A: Transformantes FOP-SP1 (carreras 1 a 18) y B: Transformantes FOP-SP4 (carreras 1 a 18). Cómo control se incluyeron reacciones de PCR llevadas a cabo con los mismos oligonucleótidos utilizando como molde ADN genómico de las estirpes FOP-SP1 o FOP-SP4 no transformadas (wt). M: marcador de peso molecular 1 kp plus DNA ladder (Invitrogen).
Estos son transformantes en los que debe haberse producido el reemplazamiento del gen
FHG2. Los fragmentos amplificados mostraron el tamaño esperado y posiblemente ocurrió
un doble evento de recombinación homóloga. En total, para la estirpe muy virulenta FOP-
SP1 se obtuvieron 48 transformantes positivos candidatos de 72 analizados. Para la estirpe
poco virulenta FOP-SP4 se obtuvieron 42 transformantes positivos de 88 analizados.
Resultados
- 127 -
3.6.4.3 Identificación de transformantes del gen FHG3
En la Figura 33 se muestran de forma esquemática el modo en el que deben alinearse el
alelo silvestre del gen FHG3 y la construcción generada y clonada en el plásmido pRF-
HU2-FHG3 y la estructura que presentará el alelo mutante del gen FHG3 derivado de dos
eventos de recombinación homóloga en ambas regiones flanqueantes de la región
estructural del gen, es decir, la estructura del alelo mutante de FHG3 consecuencia del
reemplazamiento génico.
Figura 33. Estrategia de reemplazamiento génico del gen FHG3. A: Representación simplificada del plásmido pRF-HU2-FHG3 con la construcción de reemplazamiento del gen FHG2. B: Esquema del entorno genómico del alelo silvestre del gen FHG3. Se indican las regiones en los extremos flanqueantes 5’ y 3’ en las que debe producirse recombinación homóloga para que tenga lugar el reemplazameinto génico de FHG3. C: Estructura del alelo mutante generado mediante reemplazamiento génico como consecuencia de un doble evento de recombinación homóloga en las regiones flaqueantes 5’ y 3’ de FHG3. La ubicación de las dianas de hibridación de los cebadores utilizados para amplificación de las regiones flanqueantes 5´ y 3´ clonadas en el plásmido pRF-HU2-FHG3 se representa mediante flechas de color verde (B). La ubicación de las dianas de anillamiento de los cebadores utilizados para la detección de la ocurrencia de recombinación homóloga en una región flanqueante u otra se representa mediante flechas de color naranja (B y C). Se indica la posición de las dianas de restricción de la enzima HindIII en el alelo silvestre y en el alelo resultante del remplazamiento génico (HindIII no corta el plásmido pRF-HU2-FHG3).
La identificación de “transformantes candidato” en los que se ha producido el
reemplazamiento génico del gen FHG3 se realizó llevando a cabo reacciones de PCR
utilizando la combinación de cebadores FHG185-KN-FW y HphBC y RHG83-KN-RV y
Papel de las flavohemoglobinas en F. oxysporum
- 128 -
HphAC mostrados en la Figura 33B y 33C. La amplificación de un fragmento de 2431 pares
bases correspondiente a la región flanqueante 5´ sería indicativa de recombinación
homóloga en esta región. Igualmente, la amplificación de un fragmento de 1895 pares de
bases correspondiente a la región flanqueante 3´ sería indicativa de recombinación
homóloga en esta región. En la Figura 34 se muestran resueltos mediante electroforesis
en geles de agarosa los productos de amplificación obtenidos en reacciones de PCR
llevadas a cabo con ambas combinaciones de cebadores en una muestra representativa
de transformantes. En un buen número de transformantes primarios se amplificaron los dos
fragmentos diagnósticos de recombinación homóloga.
Figura 34. Productos de PCR obtenidos en reacciones de PCR llevadas a cabo con los oligonucleótidos FHG185-KN-FW y HphBC (línea superior en cada gel) (5’) y RHG183-KN-RV y HphAC (línea inferior en cada gel) (3’) utilizando como molde ADN genómico de los transformantes obtenidos con el plásmido pRF-HU2-FHG3. A: Transformantes FOP-SP1 (carreras 1 a 12) y B: Transformantes FOP-SP4 (carreras 1 a 11). Cómo control se incluyeron reacciones de PCR llevadas a cabo con los mismos oligonucleótidos utilizando como molde ADN genómico de las estirpes FOP-SP1 o FOP-SP4 no transformadas (wt). M: marcador de peso molecular 1 kp plus DNA ladder (Invitrogen).
Los transformantes en los que se detecta amplificación de ambos productos de PCR, y
éstos tienen el tamaño esperado, son aquellos en los que debe haberse producido el
reemplazamiento del gen FHG2. En total, para la estirpe muy virulenta FOP-SP1 se
obtuvieron 42 transformantes positivos de 102 analizados. Para la estirpe poco virulenta
FOP-SP4 se obtuvieron 32 transformantes positivos de 110 analizados.
Resultados
- 129 -
3.6.4.4 Identificación de transformantes del gen FHG4
En la Figura 35 se presenta el esquema del modo en el que deben alinearse el alelo
silvestre del gen FHG4 y la construcción generada y clonada en el plásmido pRF-HU2-
FHG4 y la estructura que presentará el alelo mutante del gen FHG4 derivado de dos
eventos de recombinación homóloga en ambas regiones flanqueantes de la región
estructural del gen, es decir, la estructura del alelo mutante de FHG4 consecuencia del
remplazamiento génico.
Figura 35. Estrategia de reemplazamiento génico del gen FHG4. A: Representación simplificada del plásmido pRF-HU2-FHG4 con la construcción de reemplazamiento del gen FHG4. B: Esquema del entorno genómico del alelo silvestre del gen FHG4. Se indican las regiones en los extremos flanqueantes 5’ y 3’ en las que debe producirse recombinación homóloga para que tenga lugar el reemplazamiento génico de FHG4. C: Estructura del alelo mutante generado mediante reemplazamiento génico como consecuencia de un doble evento de recombinación homóloga en las regiones flaqueantes 5’ y 3’ de FHG4. La ubicación de las dianas de hibridación de los cebadores utilizados para amplificación de las regiones flanqueantes 5´ y 3´ clonadas en el plásmido pRF-HU2-FHG4 se representa mediante flechas de color verde (B). La ubicación de las dianas de anillamiento de los cebadores utilizados para la detección de la ocurrencia de recombinación homóloga en una región flanqueante u otra se representa mediante flechas de color naranja (B y C). Se indica la posición de las dianas de restricción de la enzima HindIII en el alelo silvestre y en el alelo resultante del reemplazamiento génico (HindIII no corta el plásmido pRF-HU2-FHG2).
La identificación de transformantes en los que se ha producido el reemplazamiento génico
del gen FHG4 se realizó llevando a cabo reacciones de PCR utilizando la combinación de
cebadores FHG155-KN-FW y HphBC y RHG153-KN-RV y HphAC mostrados en la Figura
Papel de las flavohemoglobinas en F. oxysporum
- 130 -
35B y 35C. La amplificación de un fragmento de 2421 pares bases correspondiente a la
región flanqueante 5´ sería indicativa de recombinación homóloga en esta región. Por otro
lado, la amplificación de un fragmento de 2097 pares de bases correspondiente a la región
flanqueante 3´ sería indicativa de recombinación homóloga en esta región. Como se
observa en la Figura 36, en la que se muestran resueltos mediante electroforesis en geles
de agarosa los productos de amplificación obtenidos en reacciones de PCR llevadas a
cabo con ambas combinaciones de cebadores en una muestra representativa de
transformantes, en un buen número de transformantes primarios amplificaron los dos
fragmentos indicadores de recombinación homóloga. Estos transformantes son aquellos
en los que debe haberse producido el reemplazamiento del gen FHG4. En los mismos se
detectó la amplificación de los fragmentos derivados de una y otra regíón flanqueante y
ambos presentaban el tamaño esperado. En total, para la estirpe muy virulenta FOP-SP1
se obtuvieron 7 transformantes positivos candidatos de 110 analizados. Para la estirpe
poco virulenta FOP-SP4 se obtuvieron 5 transformantes positivos de 112 analizados.
Figura 36. Productos de PCR obtenidos en reacciones de PCR llevadas a cabo con los oligonucleótidos FHG155-KN-FW y HphBC (línea superior en cada gel) (5’) y RHG153-KN-RV y HphAC (línea inferior en cada gel) (3’) utilizando como molde ADN genómico de los transformantes obtenidos con el plásmido pRF-HU2-FHG4. A: Transformantes FOP-SP1 (carreras 1 a 18) y B: Transformantes FOP-SP4 (carreras 1 a 18). Cómo control se incluyeron reacciones de PCR llevadas a cabo con los mismos oligonucleótidos utilizando como molde ADN genómico de las estirpes FOP-SP1 o FOP-SP4 no transformadas (wt). M: marcador de peso molecular 1 kp plus DNA ladder (Invitrogen).
Resultados
- 131 -
3.7 Análisis mediante hibridación Southern de los transformantes de F.
oxysporum f. sp. phaseoli
Los transformantes en los que fue posible identificar mediante PCR la ocurrencia de dos
eventos de doble recombinación homóloga en las regiones flanqueantes 5´ y 3’ bien del
gen FHG1, del gen FHG2, del gen FHG3 o del gen FHG4 fueron analizados mediante
hibridación de tipo Southern. Con el objeto de eliminar posibles núcleos no transformados
y de garantizar la homocariosis del alelo “reemplazamiento” en los transformantes
seleccionados, a partir de cada uno de ellos se obtuvieron cultivos monospóricos
seleccionando esporas individuales en medio PDA suplementado con 75 µg/ml de
higromicina B (en la mayor parte de los casos a partir de cada cepa en forma de cultivo
monospórico se obtuvo un cultivo monospórico de “segunda generación” y de éste un
cultivo monospórico de tercera generación, dados los problemas de heterocariosis que en
algunos casos hemos detectado).
Finalmente los transformantes seleccionados y purificados fueron cultivados en medio
líquido PDB con 50 µg/ml de higromicina y a partir del micelio recuperado se extrajo ADN
genómico que fue utilizado en el análisis de hibridación Southern. En todos los casos, es
decir, para la caracterización de los transformantes seleccionados para cada uno de los
cuatro genes considerados, el análisis siguió la misma estrategia. En primer lugar se
determinaría la presencia o ausencia en cada transformante de la banda de hibridación
característica del alelo silvestre mediante hibridación con una sonda derivada de la región
estructural del gen analizado. En segundo lugar se detectarían bien la banda
correspondiente al alelo silvestre, bien la banda correspondiente al alelo remplazamiento,
mediante hibridación con una sonda derivada de la región 5’ flanqueante del gen. Y
finalmente, se confirmaría la identidad de la banda remplazamiento mediante hibridación
con una sonda derivada de la región estructural del gen que determina la resistencia a
higromicina. Esta última hibridación permitiría, además, detectar la ocurrencia de
integraciones adicionales del casete de resistencia a higromicina B en el genoma de los
transformantes. Las sondas derivadas de la región codificante y de la región flanqueante
5’ de cada gen en estudio y la sonda del gen de resistencia a higromicina B fueron
marcadas mediante PCR con los cebadores mostrados en la Tabla 6 (ver Figuras 37, 38,
39 y 40 para visualizar los fragmentos de amplificación esperados en cada caso).
Papel de las flavohemoglobinas en F. oxysporum
- 132 -
3.7.1 Análisis de los mutantes en el gen ΔFHG1
En la Figura 37 se muestran los resultados de las hibridaciones de tipo Southern llevadas
a cabo en el análisis de los transformantes 130, 131, 149 y 156 de la estirpe FOP-SP1 y
de la propia estirpe FOP-SP1 (Paneles A, B y C), así como de los transformantes 226, 239
y 244 de la estirpe FOP-SP4 y de la propia estirpe FOP-SP4 (Paneles D, E y F), con la
sonda específica de la región codificante del alelo silvestre del gen FHG1 (paneles A y D),
con la sonda del Flanco 5´ del mismo (Paneles B y E) y con la sonda del gen de resistencia
a higromicina (Paneles C y F). El ADN en todos los casos fue digerido con HindIII, enzima
de restricción para la que existe una diana de reconocimiento dentro de la región
codificante de FHG1 pero fuera de la zona correspondiente al fragmento utilizado como
sonda. En el Panel 37A se comprueba que en las cepas transformantes 149 y 156 de FOP-
SP1 se detecta la banda de hibridación correspondiente al alelo silvestre de FHG1, de 3958
pares de nucleótidos, banda que también se detecta en la cepa FOP-SP1. En las cepas
130 y 131 no se detecta esta banda, lo cual indica que en estos dos transformantes el alelo
silvestre ha sido eliminado.
La hibridación con la sonda derivada de la región flanqueante 5’ demuestra que,
efectivamente, en los transformantes 130 y 131 el alelo silvestre ha sido sustituido por el
alelo de reemplazamiento, ya que únicamente se detecta una banda de hibridación, que
tiene el tamaño esperado (5374 pares de nucleótidos) del alelo “reemplazamiento”. En los
transformantes 149 y 156 se detectan ambas bandas con la sonda derivada de la región 5’
flanqueante, la correspondiente al alelo silvestre y la correspondiente al alelo
reemplazamiento, observación compatible con una integración en tándem, consecuencia
de un único evento de recombinación homóloga bien en la región flanqueante 5’, bien en
la región flanqueante 3’, o con la ocurrencia de heterocariosis. La hibridación con la sonda
derivada de la región estructural del gen de resistencia a higromicina B (Panel 37C)
demuestra que la banda detectada en todos los transformantes con una tamaño de 5374
pares de nucleótidos es, efectivamente, la banda derivada de la copia de reemplazamiento
de FHG1 clonada en el plásmido de transformación. Por lo tanto, sólo los transformantes
130 y 131 de la estirpe FOP-SP1 son auténticos mutantes por reemplazamiento génico del
gen FHG1. El mismo tipo de análisis llevado a cabo con los transformantes 225, 239 y 244
de la estirpe FOP-SP4 demuestra que en este caso el alelo silvestre FHG1 ha sido
reemplazado por el alelo mutante en todos los transformantes identificados mediante PCR
y seleccionados. Para todos los análisis posteriores se seleccionaron dos mutantes
Resultados
- 133 -
independientes de cada estirpe, los mutantes 130 y 131 de FOP-SP1 y los mutantes 239 y
244 de FOP-SP4.
Figura 37. Análisis mediante hibridación tipo Southern de los transformantes en los que se ha determinado mediante PCR la ocurrencia de dos eventos de recombinación homóloga en las regiones 5’ y 3’ flanqueantes del gen FHG1 en las estirpes FOP-SP1 (Paneles A, B y C) y FOP-SP4 (Paneles D, E y F). Las muestras de ADN genómico (10 µg en cada caso) fueron digeridas con la enzima de restricción HindIII, separadas mediante electroforesis en gel de agarosa y transferidas a filtros de nylon. Las hibridaciones fueron llevadas a cabo con una sonda derivada de la región estructural del gen FHG1 (paneles A y D), de su región 5’ flanqueante (Paneles B y E) o de la región estructural del gen que confiere resistencia a higromicina B (paneles C y F). Los números sobre los filtros hibridados corresponden a los códigos de los transformantes seleccionados en las trasformaciones realizadas sobre la estirpe FOP-SP1 o FOP-SP4, a partir de los cuales se prepararon muestras de ADN genómico que fueron incluidas como control en los filtros correspondientes.
3.7.2 Análisis de los mutantes en el gen ΔFHG2
En la Figura 38 se muestran los resultados de las hibridaciones llevadas a cabo en el
análisis de los transformantes 50, 51, 57 y 61 de la estirpe FOP-SP1 y de la propia estirpe
FOP-SP1 (Paneles A, B y C) y de los transformantes 303, 314 y 315 de la estirpe FOP-
SP4 y de la propia estirpe FOP-SP4 (Paneles D, E y F) con la sonda específica de la región
codificante del alelo silvestre del gen FHG2 (Paneles A y D), con la sonda derivada de la
región flanqueante 5´ (Paneles B y E) y con la sonda del gen de resistencia a higromicina
Papel de las flavohemoglobinas en F. oxysporum
- 134 -
B (Paneles C y F). El ADN en todos los casos fue digerido con HindIII, enzima de restricción
para la que existe una diana de reconocimiento dentro de la región codificante del gen
FHG2, pero fuera de la zona correspondiente al fragmento utilizado como sonda. En el
Panel 38A se comprueba que en las cepas transformantes 50, 51, 57 y 61 el alelo silvestre
ha sido eliminado. La hibridación con la sonda derivada de la región flanqueante 5’
demuestra que, efectivamente, en estos transformantes el alelo silvestre ha sido sustituido
por el alelo de reemplazamiento, ya que únicamente se detecta una banda, que tiene el
tamaño esperado (9084 pares de nucleótidos) del alelo de reemplazamiento. La hibridación
con la sonda derivada de la región estructural del gen de la higromicina B (Panel 38C)
demuestra que la banda detectada en todos los transformantes con una tamaño de 9084
pares de nucleótidos es, efectivamente, la banda derivada de la copia “reemplazamiento”
del gen FHG2 clonada en el plásmido de transformación.
Figura 38. Análisis mediante hibridación tipo Southern de los transformantes en los que se ha determinado mediante PCR la ocurrencia de dos eventos de recombinación homóloga en las regiones 5’ y 3’ flanqueantes del gen FHG2 en las estirpes FOP-SP1 (Paneles A, B y C) y FOP-SP4 (Paneles D, E y F). Las muestras de ADN genómico (10 µg en cada caso) fueron digeridas con la enzima de restricción HindIII, separados mediante electroforesis en gel de agarosa y transferidas a filtros de nylon. Las hibridaciones fueron llevadas a cabo con una sonda derivada de la región estructural del gen FHG2 (Paneles A y D), de su región 5’ flanqueante (paneles B y E) o de la región estructural del gen que confiere resistencia a higromicina B (Paneles C y F). Los números sobre los filtros hibridados corresponden a los códigos de los transformantes seleccionados en las trasformaciones realizadas sobre la estirpe FOP-SP1 o FOP-SP4, a partir de los cuales se prepararon muestras de ADN genómico que fueron incluidas como control en los filtros correspondientes.
Resultados
- 135 -
Por lo tanto, los transformantes 50, 51, 57 y 61 de la estirpe FOP-SP1 son auténticos
mutantes por remplazamiento génico de FHG2. El mismo tipo de análisis llevado a cabo
con los transformantes 302, 314 y 315 de la estirpe FOP-SP4 demuestra que en este caso
el alelo silvestre FHG2 ha sido reemplazado por el alelo mutante en todos los
transformantes identificados mediante PCR y seleccionados. Para todos los análisis
posteriores se seleccionaron dos mutantes independientes de cada estirpe, los mutantes
51 y 61 de FOP-SP1 y los mutantes 314 y 315 de FOP-SP4.
3.7.3 Análisis de los mutantes en el gen ΔFHG3
En la Figura 39 se muestran los resultados de las hibridaciones realizadas con los
transformantes 811, 812 y 818 de la estirpe FOP-SP1 y de la propia estirpe FOP-SP1
(Paneles A, B y C) y de los transformantes 705 y 728 de la estirpe FOP-SP4 y de la propia
estirpe FOP-SP4 (Paneles D, E y F) con la sonda especifica de la región codificante del
alelo silvestre del gen FHG3 (Paneles A y D), con la sonda derivada de la región
flanqueante 5´ (Paneles B y E) y con la sonda del gen de resistencia a higromicina B
(Paneles C y F). El ADN en todos los casos fue digerido con HindIII, enzima de restricción
para la que existe una diana de reconocimiento dentro de la región codificante del gen
FHG3 pero fuera de la zona correspondiente al fragmento utilizado como sonda. En el
Panel 39A se comprueba que en las cepas transformantes 811, 812 y 818 el alelo silvestre
ha sido eliminado. La hibridación con la sonda derivada de la región flanqueante 5’
demuestra que, efectivamente, en estos transformantes el alelo silvestre ha sido sustituido
por el alelo remplazamiento, ya que únicamente se detecta una banda, que tiene el tamaño
esperado (6585 pares de nucleótidos). La hibridación con la sonda derivada de la región
estructural del gen de resistencia a higromicina B (Panel 39C) demuestra que la banda
detectada en todos los transformantes tiene un tamaño de 6585 pares de nucleótidos y es,
efectivamente, la banda derivada de la copia de reemplazamiento del gen FHG3 clonada
en el plásmido de transformación. Por lo tanto, los transformantes 811, 812 y 818 de la
estirpe FOP-SP1 son auténticos mutantes por remplazamiento génico del gen FHG3. El
mismo tipo de análisis llevado a cabo con los transformantes 705 y 728 de la estirpe FOP-
SP4 demuestra que en este caso el alelo silvestre FHG3 ha sido reemplazado por el alelo
mutante en los dos transformantes identificados mediante PCR y seleccionados. Para
todos los análisis posteriores se seleccionaron dos mutantes independientes de cada
estirpe, los mutantes 812 y 818 de FOP-SP1 y los mutantes 705 y 728 de FOP-SP4.
Papel de las flavohemoglobinas en F. oxysporum
- 136 -
Figura 39. Análisis mediante hibridación tipo Southern de los transformantes en los que se ha determinado mediante PCR la ocurrencia de dos eventos de recombinación homóloga en las regiones 5’ y 3’ flanqueantes del gen FHG3 en las estirpes FOP-SP1 (Paneles A, B y C) y FOP-SP4 (Paneles D, E y F). Las muestras de ADN genómico (10 µg en cada caso) fueron digeridas con la enzima de restricción HindIII, separados mediante electroforesis en gel de agarosa y transferidas a filtros de nylon. Las hibridaciones fueron llevadas a cabo con una sonda derivada de la región estructural del gen FHG3 (Paneles A y D), de su región 5’ flanqueante (Paneles B y E) o de la región estructural del gen que confiere resistencia a higromicina B (Paneles C y F). Los números sobre los filtros hibridados corresponden a los códigos de los transformantes seleccionados en las trasformaciones realizadas sobre la estirpe FOP-SP1 o FOP-SP4, a partir de los cuales se prepararon muestras de ADN genómico que fueron incluidas como control en los filtros correspondientes.
3.7.4 Análisis de los mutantes en el gen ΔFHG4
En la Figura 40 se muestran los resultados de las hibridaciones llevadas a cabo en el
análisis de los transformantes 619, 637 y 639 de la estirpe FOP-SP1 y de la propia estirpe
FOP-SP1 (Paneles A, B y C) y de los transformantes 508 y 516 de la estirpe FOP-SP4 y
de la propia estirpe FOP-SP4 (Paneles D, E y F), con la sonda especifica de la región
codificante del alelo silvestre del gen FHG4 (Paneles A y D), con la sonda del Flanco 5´
(Paneles B y E) y con la sonda del gen de resistencia a higromicina B (Paneles C y F). El
ADN en todos los casos fue digerido con HindIII, enzima de restricción para la que existe
una diana de reconocimiento dentro de la región codificante del gen FHG4 pero fuera de la
Resultados
- 137 -
zona correspondiente al fragmento utilizado como sonda. En el Panel 40A se comprueba
que en las cepas transformantes 619, 637 y 639 el alelo silvestre ha sido eliminado. La
hibridación con la sonda derivada de la región flanqueante 5’ demuestra que,
efectivamente, en estos transformantes el alelo silvestre ha sido sustituido por el alelo de
reemplazamiento, detectándose únicamente una banda que tiene el tamaño esperado
(6554 pares de nucleótidos). La hibridación con la sonda derivada de la región estructural
del gen de resistencia a higromicina B (Panel 40C) demuestra que la banda detectada en
todos los transformantes con un tamaño de 6554 pares de nucleótidos es, efectivamente,
la banda derivada de la copia de reemplazamiento del gen FHG4 clonada en el plásmido
de transformación.
Figura 40. Análisis mediante hibridación tipo Southern de los transformantes en los que se ha determinado mediante PCR la ocurrencia de dos eventos de recombinación homóloga en las regiones 5’ y 3’ flanqueantes del gen FHG4 en las estirpes FOP-SP1 (Paneles A, B y C) y FOP-SP4 (Paneles D, E y F). Las muestras de ADN genómico (10 µg en cada caso) fueron digeridas con la enzima de restricción HindIII, separados mediante electroforesis en gel de agarosa y transferidas a filtros de nylon. Las hibridaciones fueron llevadas a cabo con una sonda derivada de la región estructural del gen FHG4 (Paneles A y D), de su región 5’ flanqueante (Paneles B y E) o de la región estructural del gen que confiere resistencia a higromicina (Paneles C y F). Los números sobre los filtros hibridados corresponden a los códigos de los transformantes seleccionados en las transformaciones realizadas sobre la estirpe FOP-SP1 o FOP-SP4, a partir de los cuales se prepararon muestras de ADN genómico que fueron incluidas como control en los filtros correspondientes.
Papel de las flavohemoglobinas en F. oxysporum
- 138 -
Por lo tanto, los transformantes 619, 637 y 639 de la estirpe FOP-SP1 son auténticos
mutantes por reemplazamiento génico del gen FHG4. El mismo tipo de análisis llevado a
cabo con los transformantes 508 y 516 de la estirpe FOP-SP4 demuestra que en este caso
el alelo silvestre FHG4 ha sido reemplazado por el alelo mutante en los dos transformantes
identificados mediante PCR y seleccionados. Para todos los análisis posteriores se
seleccionaron dos mutantes independientes de cada estirpe, los mutantes 619 y 637 de
FOP-SP1 y los mutantes 508 y 516 de FOP-SP4.
3.8 Caracterización fisiológica de los mutantes ∆FHG
Los cuatro genes FHG analizados en este trabajo codifican proteínas que presentan los
dominios funcionales característicos de proteínas tipo flavohemoglobina. Por lo tanto, es
posible asumir que estas proteínas están implicadas en la modulación de los niveles de
NO a los que puede resultar expuesto F. oxysporum f. sp. phaseoli (FOP). Los cuatro genes
muestran patrones de expresión diferentes en respuesta a distintos factores o estímulos,
tales como el desarrollo, la fuente de nitrógeno o la exposición a NO exógeno. Estas
observaciones sugieren que cada una de estas proteínas puede participar en el mismo
proceso fisiológico, respondiendo de manera diferente a un mismo estímulo, o en procesos
fisiológicos distintos, quizás en un caso y otro como consecuencia de cierta especialización
funcional. Para tratar de identificar los procesos fisiológicos en los que la detoxificación de
NO mediada por cada una de estas enzimas pudiera jugar un papel relevante decidimos
llevar a cabo una caracterización funcional de los mutantes deficientes en cada uno de los
cuatro genes FHG. Los aspectos sobre los que decidimos trabajar inicialmente evaluando
la ocurrencia de posibles alteraciones en comparación con las estirpes silvestres fueron:
(1) la capacidad de crecimiento saprofítico, (2) la sensibilidad frente a condiciones de estrés
nitrosativo, (3) la germinación de los microconidios y (4) la agresividad sobre judía.
Para la caracterización fisiológica de las cepas mutantes en cada gen FHG en comparación
con la correspondiente estirpe silvestre se seleccionaron en cada caso dos cepas mutantes
independientes, como hemos indicado anteriormente. El objeto de incluir dos cepas
mutantes independientes de cada gen en cada estirpe es comprobar que, en el caso de
detectar una variación dada en relación con la cepa silvestre en alguna de las cepas
mutantes, ésta es observada también en la segunda cepa mutante alterada en el mismo
gen. En una primera aproximación, la consistencia de las observaciones realizadas en
ambas cepas mutantes para un gen dado permite adscribir el fenotipo observado a la
mutación introducida. La Tabla 22 presenta las cepas mutantes seleccionadas en nuestro
Resultados
- 139 -
análisis de posibles alteraciones fisiológicas, indicando la cepa tipo silvestre de la que
derivan, el gen que ha sido eliminado en cada caso y el código que le ha sido asignado.
En todos los análisis llevados a cabo y que se describen a continuación las dos cepas
mutantes fueron comparadas con la correspondiente cepa silvestre en experimentos que
fueron repetidos al menos en tres ocasiones.
Tabla 22. Estirpes silvestres y cepas mutantes ΔFHG generadas en este trabajo y seleccionadas para su análisis fisiológico.
WT ∆FHG1 ∆FHG2 ∆FHG3 ∆FHG4
FOP-SP1 ∆FHG1-130 ∆FHG2-51 ∆FHG3-812 ∆FHG4-619
∆FHG1-131 ∆FHG2-61 ∆FHG3-818 ∆FHG4-637
FOP-SP4 ∆FHG1-239 ∆FHG2-314 ∆FHG3-705 ∆FHG4-508
∆FHG1-244 ∆FHG2-315 ∆FHG3-728 ∆FHG4-516
3.8.1 Análisis de la capacidad de crecimiento saprofítico y de la
sensibilidad a estrés nitrosativo de los mutantes ∆FHG
En primer lugar se evaluó la capacidad de crecimiento saprofítico de las cepas mutantes
en comparación con su correspondiente cepa tipo silvestre en ausencia y en presencia de
un donador de NO. La valoración del crecimiento saprofítico en ausencia de donador de
NO y, por lo tanto, la presencia de NO exógeno, nos proporcionará, en primer lugar,
información sobre posibles alteraciones nutricionales o de desarrollo que pudieran
derivarse de la mutación analizada en cada caso y, en segundo lugar, un estado de
referencia para evaluar un posible aumento en la sensibilidad frente a condiciones de
estrés nitrosativo generadas por la adición de NO exógeno y derivada de la eliminación de
una enzima flavohemoglobina dada.
Para el análisis de crecimiento saprofitico y sensibilidad frente a estrés nitrosativo se
utilizaron placas de medio de cultivo sólido. El medio de cultivo basal elegido fue medio
mínimo suplementado con 1 % glucosa (MMG). Como fuente de nitrógeno se utilizó bien
amonio, bien nitrato (ver en la sección 2.2.2.1 de Materiales y Métodos). Las cepas
seleccionadas fueron precultivadas en los medios antes mencionados en cada caso y a
partir del borde de crecimiento de la colonia se tomaron cilindros de agar con micelio en
crecimiento activo de 5 mm de diámetro que fueron transferidos al centro de las placas con
Papel de las flavohemoglobinas en F. oxysporum
- 140 -
medio de cultivo fresco. En un caso y otro, es decir, en MMG+A o MMG+N, las distintas
cepas analizadas fueron cultivadas en ausencia y en presencia del donador de NO (SNP
500 µM). En cada experimento, para cada cepa (mutante y tipo silvestre) se sembraron
tres placas de Petri y el crecimiento fue evaluado estimando el diámetro medio de la colonia
en crecimiento a los 2, 3, 4 y 5 días. El ensayo de evaluación de crecimiento en cada
condición fue repetido al menos tres veces y los datos registrados fueron analizados
estadísticamente mediante análisis de varianza (ANOVA) y la discriminación de medias
con el test de Tukey (p<0,05). Las diferencias estadísticas entre cepas de tipo silvestre y
las distintas cepas mutantes ∆FHG en ausencia y presencia de NO exógeno se analizaron
mediante un análisis pareado con t-test de Student´s (p<0,05).
3.8.1.1 Análisis de mutantes ΔFHG de FOP-SP1
La Figura 41 muestra los resultados obtenidos en el análisis de crecimiento saprofitico de
los mutantes ∆FHG seleccionados en comparación con la cepa silvestre FOP-SP1. En el
Panel 41A los datos presentados son los derivados del cultivo de las distintas cepas en
medio suplementado con amonio como fuente de nitrógeno. El Panel 41B presenta los
datos obtenidos sobre medio suplementado con nitrato como fuente de nitrógeno. El
estudio de estos resultados ofrece varios elementos de información. En primer lugar, en
ambos medios de cultivo, MMG+A y MMG+N, y en ausencia de donador de NO, el
comportamiento de todas las cepas mutantes consideradas es similar al de la cepa
silvestre. El análisis estadístico (ANOVA) demuestra que no existen diferencias
significativas (p<0,05) en ningún caso entre los valores medios de diámetro de la colonia
obtenidos en el caso de las cepas mutantes consideradas y la cepa FOP-SP1. Por lo tanto,
la capacidad de crecimiento saprofítico de las cepas mutantes analizadas no se ve alterada
de forma significativa como consecuencia de las mutaciones en los genes FHG1, FHG2,
FHG3 o FHG4.
En segundo lugar, se comprueba que la cepa silvestre FOP-P1 es sensible a las
condiciones de estrés nitrosativo que impone la adición de NO (SNP 500 µM). Tanto en
medio con amonio como en medio con nitrato se detectan diferencias significativas (p<0,05,
t-test de Student) en el diámetro medio de colonia de la cepa FOP-SP1 en ausencia y en
presencia de NO. En medio con amonio la adición del donador de NO determina una
reducción del diámetro de la colonia de un 17,3 % y en medio con nitrato una reducción del
12,96 %. Esta reducción de crecimiento ofrece una herramienta de cuantificación de la
sensibilidad de FOP-SP1 al NO.
Resultados
- 141 -
En tercer lugar, las dos cepas mutantes deficientes en el gen ΔFHG1 (130 y 131) y las dos
cepas mutantes deficientes en el gen ΔFHG2 (51 y 61) muestran un fenotipo de
hipersensibilidad a las condiciones de estrés nitrosativo utilizadas en este trabajo. Tanto
en medio con amonio como en medio con nitrato las cepas mutantes indicadas muestran
diferencias significativas significativas (p<0,05, t-test de Student) en el diámetro de la
colonia en ausencia y en presencia de NO, presentando una reducción de crecimiento en
presencia de NO superior a la observada en la cepas tipo silvestre correspondiente. Se
comprueba que la reducción en el diámetro de la colonia observado en presencia de SNP
en medio suplementado con amonio es muy superior a la que muestra la propia cepa tipo
silvestre FOP-SP1, con una reducción del 36,73 % en el caso de la cepa ΔFHG1-130, un
40 % en el caso de la cepa ΔFHG1-131, un 33,33 % en el caso de la cepa ΔFHG2-51 y un
30,76 % en el caso de la cepa ΔFHG2-61. Las diferencias en el diámetro de la colonia entre
estas cuatro cepas no son significativas, aunque de forma consistente las dos cepas
deficientes en el gen FHG1 muestra un mayor grado de sensibilidad al donador de NO que
las dos cepas deficientes en el gen FHG2. En medio suplementado con nitrato también se
observa hipersensibilidad a estrés nitrosativo en estas cuatro cepas mutantes, si bien en
un grado ligeramente inferior (la reducción en el diámetro de la colonia observado en este
caso es de 33,33 % y 30,76 %, para los mutantes ΔFHG1-130 y ΔFHG1-131,
respectivamente, y de 26,79 % y 23,64 %, para los mutantes ΔFHG2-51 y ΔFHG2-61,
respectivamente).
Finalmente, las dos cepas mutantes deficientes en el gen ΔFHG3 (812 y 818) y las dos
cepas mutantes deficientes en el gen ΔFHG4 (619 y 637) no muestran hipersensibilidad a
condiciones de estrés nitrosativo en medio suplementado con amonio, ya que en estas
cuatro cepas el diámetro medio de la colonia en presencia de donador de óxido nítrico (NO)
no presenta diferencias significativas con el diámetro de la colonia en ausencia de NO
(Figura 41A). En presencia de nitrato como fuente de nitrógeno las dos cepas mutantes
deficientes en el gen ΔFHG3 y las dos cepas mutantes deficientes en el gen ΔFHG4
tampoco muestran sensibilidad a NO en comparación con las cepas cultivadas en ausencia
de NO (Figura 41B). En la Figura 42 se muestran los porcentajes de reducción del diámetro
de colonia en presencia de NO (500 µM de SNP) en los mutantes ΔFHG y en la cepa
control SP1 considerando los datos presentados en las Figuras 41A y 41B.
Papel de las flavohemoglobinas en F. oxysporum
- 142 -
Figura 41. Crecimiento radial de las cepas mutantes en los genes FHG de FOP-SP1 y de la cepa control FOP-SP1 evaluados a los 5 días de crecimiento saprofitico. (A) cultivos en medio mínimo con 1 % de glucosa, suplementado con 10 mM de tartrato de amonio (MMG+A) y MMG+A en presencia de 500 µM de SNP (donador de NO exógeno). (B) cultivos en medio mínimo con 1 % de glucosa, suplementado con 10 mM de nitrato de sodio (MMG+N) y MMG+N en presencia de 500 µM de SNP (donador de NO exógeno). Los histogramas de la izquierda en los Paneles A y B en medios de cultivo MMG+A y MMG+N en ausencia del donador de NO fueron analizados estadisticamente mediante un análisis ANOVA y la discriminación de medias se realizó con el test de Tukey (p<0,05). En los histogramas de la derecha en los Paneles A y B se muestra el análisis estadístico pareado en cada cepa (tipo silvestre y mutantes) en ausencia y presencia de NO (SNP) con el t-test de Student´s (*p<0,05), (**p<0,01), (***p<0,001).
Resultados
- 143 -
Figura 42. Porcentajes de reducción del diámetro de la colonia de las cepas mutantes ∆FHG de FOP-SP1 y de la cepa tipo silvestre SP1. Panel A: Porcentajes de reducción del diámetro de la colonia en medio mínimo con 1 % de glucosa, suplementado con 10 mM de tartrato de amonio (MMG+A) en presencia de 500 µM de SNP (donador de NO exógeno). Panel B: Porcentajes de reducción del diámetro de la colonia en medio mínimo con 1 % de glucosa, suplementado con 10 mM de nitrato de sodio (MMG+N) en presencia de 500 µM de SNP (donador de NO exógeno).
3.8.1.2 Análisis de mutantes ΔFHG de FOP-SP4
Se llevó a cabo el mismo tipo de análisis para la caracterización de las cepas mutantes
deficientes en los genes FHG1, FHG2, FHG3 o FHG4 de la cepa FOP-SP4. En la Figura
43 se presentan los resultados obtenidos en estos análisis.
El estudio de la Figura 43A demuestra que la situación en la cepa FOP-SP4 y sus
transformantes es muy similar a la descrita en la cepa FOP-SP1 y en sus cepas derivadas.
Comprobamos que en cualquiera de las dos fuentes de nitrógeno consideradas, en
ausencia de NO exógeno las cepas mutantes no muestran diferencias significativas en su
capacidad de crecimiento saprofítico en relación con la cepa tipo silvestre. Comprobamos
también que la cepa silvestre FOP-SP4 muestra sensibilidad a NO en los dos medios
considerados (con amonio y con nitrato), ya que el diámetro medio de la colonia se reduce
significativamente (p<0,05, t-test de Student) en presencia del donador de NO. En este
Papel de las flavohemoglobinas en F. oxysporum
- 144 -
caso la reducción de crecimiento observada en medio con amonio, de un 15,09 %, es de
menor magnitud que en el caso de la cepa FOP-SP1 (17,3 %). En medio con nitrato la
reducción de crecimiento determinada en la cepa FOP-SP4, 17,2 %, es superior a la
observada para la cepa FOP-SP1 (12,96 %). Por lo tanto, es posible concluir que ambas
cepas tipo silvestre son sensibles a NO y en similar medida, mostrando diferencias
menores en función de la fuente de nitrógeno en presencia de la cual una y otra son
cultivadas.
Comprobamos, además, que las cepas mutantes deficientes en el gen FHG1 (239 y 244)
y las cepas mutantes alteradas en el gen FHG2 (314 y 315) muestran hipersensibilidad
frente a condiciones de estrés nitrosativo. Tanto en amonio como en nitrato como fuente
de nitrógeno estas cepas muestran diferencias significativas en el diámetro de la colonia
(p<0,05, t-test de Student) en presencia de NO en relación con su comportamiento en
ausencia de NO. En comparación con las cepas equivalentes en el fondo genético de FOP-
SP1, se observa que el grado de reducción de crecimiento en las cepas mutantes ΔFHG1
y ΔFHG2 es ligeramente menor que en aquéllas. Así, en presencia de amonio la reducción
de diámetro medio de la colonia en condiciones de estrés nitrosativo es del 32,65 % para
la cepa ΔFHG1-239, y 34,69 % para la cepa ΔFHG1-244, 20,83 % para la cepa ΔFHG2-
314 y 20,41 % para la cepa ΔFHG2-315. En presencia de nitrato las reducciones
observadas son 31,48 % para ΔFHG1-239 y 34,54 % para ΔFHG1-244. Para las cepas
ΔFHG2-314 y ΔFHG2-315 las reducciones observadas son del 20,75 % y 19,31 %,
respectivamente (ver Figura 43A y 43B).
Finalmente, el análisis estadístico demuestra que no hay diferencias significativas (p<0,05,
t-test de Student) en la reducción del diámetro medio de la colonia entre los mutantes de
los genes ∆FHG3 y ∆FHG4 en la comparación llevada a cabo en presencia y en ausencia
del donador de NO.
En la Figura 44 se muestran los porcentajes medios de reducción del diámetro de colonia
en presencia de NO (500 µM de SNP) en los mutantes ΔFHG de SP4 y en la cepa control
SP4 considerando los datos presentados en las Figuras 43A y 43B.
En resumen, del análisis de la capacidad de crecimiento saprofitico y la evaluación de la
sensibilidad a condiciones de estrés nitrosativo de las cepas tipo silvestre FOP-SP1 y FOP-
SP4 y de los mutantes FHG de una y otra se derivan las siguientes conclusiones: (1) Las
mutaciones analizadas no afectan a funciones metabólicas básicas relacionadas con la
Resultados
- 145 -
capacidad de crecimiento saprofítico de las cepas FOP-SP1 y FOP-SP4; (2) FOP-SP1 y
FOP-SP4 son sensibles a condiciones de estrés nitrosativo, FOP-SP1 en mayor medida
que FOP-SP4; (3) en ambas cepas la función de los genes FHG1 y FHG2 está relacionada
con la capacidad de las FHG correspondientes para detoxificar NO exógeno; (4) en ambas
cepas la función de los genes FHG3 y FHG4 no está relacionada con la capacidad de las
FHG correspondientes para detoxificar NO exógeno, al menos en las condiciones
utilizadas.
Figura 43. Crecimiento radial de las cepas mutantes en los genes FHG de FOP-SP4 y de la cepa control FOP-SP4 evaluados a los 5 días de crecimiento saprofitico. (A) cultivos en medio mínimo con 1 % de glucosa, suplementado con 10 mM de tartrato de amonio (MMG+A) y MMG+A en presencia de 500 µM de SNP (donador de NO exógeno). (B) cultivos en medio mínimo con 1 % de glucosa, suplementado con 10 mM de nitrato de sodio (MMG+N) y MMG+N en presencia de 500 µM de SNP (donador de NO exógeno). Los histogramas de la izquierda en los paneles A y B en medios de cultivo MMG+A y MMG+N en ausencia del donador de NO fueron analizados estadísticamente mediante un análisis ANOVA y la discriminación de medias se realizó con el test de Tukey (p<0,05). Los histogramas de la derecha en los Paneles A y B se muestra el análisis estadístico pareado dela cepa de tipo silvestre y de las distintas cepas mutantes en ausencia y presencia de NO (SNP) con el t-test de Student´s (*p<0,05), (**p<0,01), (***p<0,001).
Papel de las flavohemoglobinas en F. oxysporum
- 146 -
Figura 44. Porcentajes de reducción del diámetro de la colonia de las cepas mutantes ∆FHG de FOP-SP4. Panel A: Porcentajes de reducción del diámetro de la colonia en medio mínimo con 1 % de glucosa, suplementado con 10 mM de tartrato de amonio (MMG+A) en presencia de 500 µM de SNP (donador de NO exógeno). Panel B: Porcentajes de reducción del diámetro de la colonia en medio mínimo con 1 % de glucosa, suplementado con 10 mM de nitrato de sodio (MMG+N) en presencia de 500 µM de SNP (donador de NO exógeno).
3.8.2 Análisis de germinación de microconidios de mutantes ΔFHG
A continuación nos propusimos evaluar los efectos que pudiera tener el NO exógeno sobre
la germinación de microconidios en las cepas silvestres FOP-SP1 y FOP-SP4 y en las
cepas mutantes en los genes FHG de cada una de ellas. Con este objeto se diseñó una
aproximación experimental que permitiera cuantificar la germinación de microconidios a lo
largo del tiempo en las cepas consideradas, evaluando el efecto de la fuente de nitrógeno
(amonio o nitrato) y de la presencia o no de NO en el medio de cultivo mínimo líquido.
En este estudio se incluyeron las mismas cepas analizadas en la sección anterior en
relación con la capacidad de crecimiento saprofitico en medio sólido. Las dos estirpes tipo
silvestre, FOP-SP1 y FOP-SP4, y las dos cepas mutantes independientes en cada gen
FHG fueron cultivados en medio líquido tal y como se describe en la sección 2.2.2.2 de
Materiales y Métodos. El medio mínimo fue suplementado con tartrato de amonio (10 mM)
Resultados
- 147 -
o con nitrato de sodio (10 mM) en presencia y en ausencia de donador de NO. En estos
experimentos se utilizó como donador de oxido nítrico DETA (250 µM) en lugar de
nitroprusiato sódico (SNP). DETA-NO dentro del grupo de los donadores de NO es el de
mejor calidad y pureza, libera NO de forma lenta pero continua durante más tiempo, a
diferencia del donador de NO nitroprusiato sódico (SNP) que libera elevadas cantidades
de NO y durante periodos cortos de tiempo, y no provoca efectos secundarios dañinos para
la célula, en medio acuoso produce ion cianuro y posteriormente tiocianato que son
compuestos altamente tóxicos para la supervivencia de la célula (Moncada et al., 1991;
Perazzolli et al., 2004). La germinación de los microconidios se evaluó a las 4, 6, 8, 10 y
12 horas. En cada momento considerado se llevaron a cabo los recuentos considerando
como microconidios germinados aquéllos que presentaron un tubo germinativo con una
longitud igual o superior al tamaño del microconidio no germinado. Se realizaron tres
réplicas con diferentes muestras biológicas tanto de las cepas mutantes como de las cepas
silvestres FOP-SP1 y FOP-SP4. Los datos generados fueron analizados estadísticamente
mediante análisis de varianza (ANOVA) y la discriminación de medias se realizó con el test
de Tukey (p<0,05). Las diferencias estadisticas en las cepas tipo silvestre y en las distintas
cepas mutantes en ausencia y en presencia de NO exógeno se realizó llevando a cabo un
análisis pareado con el t-test de Student´s (p<0,05).
3.8.2.1 Análisis de germinación de microconidios de mutantes
ΔFHG de FOP-SP1
La Figura 45 muestra los resultados obtenidos en el análisis de la germinación de
microconidios de los mutantes SP1-∆FHG seleccionados en comparación con la cepa
silvestre FOP-SP1 a las 10 horas de cultivo. En el Panel A se presentan los datos de la
germinación en medios de cultivo con amonio como única fuente de nitrógeno en ausencia
y presencia de NO (DETA 250 µM). En el Panel B se presentan los datos derivados del
mismo análisis llevado a cabo en medios de cultivo suplementados con nitrato como fuente
de nitrógeno. En ambos medios de cultivo, MMG+A y MMG+N, en ausencia del donador
de NO (DETA) el comportamiento de todas las cepas mutantes consideradas es similar al
de la cepa silvestre y el análisis estadístico demuestra que no existen diferencias
significativas (p<0,05) en la germinación de microconidios entre las cepas mutantes
∆FHG1, ∆FHG2, ∆FHG3 o ∆FHG4 y la cepa FOP-SP1. Por lo tanto, la eficiencia de la
germinación de las cepas mutantes analizadas no se ve alterada de forma significativa
como consecuencia de las mutaciones inducidas en estos genes en ausencia de NO
exógeno.
Papel de las flavohemoglobinas en F. oxysporum
- 148 -
En la cepa silvestre FOP-SP1 la eficiencia de la germinación de microconidios es afectada
significativamente (p<0,05, T-test de Student) por la presencia de NO (DETA 250 µM) tanto
en medio con amonio como en medio con nitrato, en comparación con los cultivos en
ausencia del donador de NO. En medio con amonio la adición del donador de NO determina
un retraso en la germinación del 19,40 % y en medio con nitrato un retraso del 25,01 %.
En cuanto al comportamiento de los mutantes deficientes en el gen ΔFHG1 (130 y 131) se
observa que en presencia de NO estos mutantes muestran diferencia significativas
(p<0,05, T-test de Student) en medio con amonio en relación con la situación “ausencia de
NO exógeno”, con una reducción en la eficiencia de germinación muy superior a la que
muestra la cepa tipo silvestre FOP-SP1 (83,59 % de reducción en el caso de la cepa
ΔFHG1-130 y un 85,67 % en el caso de la cepa ΔFHG1-131 (Panel 45A). En este medio
los mutantes ΔFHG2, ΔFHG3 y ΔFHG4, también muestran un retraso en la germinación
en relación con la situación observada en ausencia de NO, pero este retraso tiene la misma
magnitud que el observado en la cepa silvestre (el análisis estadístico agrupa estas seis
cepas mutantes con la cepa silvestre). Esto supone que el NO afecta a la germinación en
FOP-SP1, determinando un retraso de la misma, y que probablemente la flavohemoglobina
codificada por FHG1 juega un papel fundamental en la eliminación del NO que retrasa la
germinación. Las flavohemoglobinas codificadas por los genes FHG2, FHG3 y FHG4 no
tienen un papel importante en la eliminación del NO que afecta a la germinación.
En medio con nitrato la situación que se observa cuando se añade el donador de NO es
similar a la descrita en medio con amonio, pero con algunas diferencias importantes. En
este caso en general el efecto de la adición de NO es más evidente (la reducción en el
porcentaje de germinación en las distinas cepas analizadas es mayor). Las cepas que
manifiestan una mayor reducción, como en el caso anterior, son las cepas mutantes
deficientes en el gen FHG1 y las que muestran una menor reducción son las cepas
deficientes en el gen FHG3 y las cepas deficientes en el gen FHG4 (que se comportan
como la cepa silvestre FOP-SP1). Las cepas deficientes en el gen FHG2 muestran un
comportamiento particular en este medio de cultivo, mostrando diferencias significativas en
la condición “presencia de NO” en relación con la condición “ausencia de NO”, con una
reducción intermedia en el porcentaje de germinación con valores intermedios entre las
correspondientes cepas ΔFHG1 y las correspondientes a los de las las cepas ΔFHG3 y
ΔFHG4 (que son iguales al de la cepa silvestre). De hecho, el análisis estadísitico detecta
la existencia de diferencias significativas (p<0,05, T-test de Student) entre medias en el
conjunto de cepas analizadas y genera un grupo particular que incluye a estas dos cepas
Resultados
- 149 -
mutantes, ΔFHG2-51 y ΔFHG2-61, diferentes a los otros dos grupos, con una reducción
de la germinación del 59,80 % para ΔFHG2-51 y para ΔFHG2-61 una reducción del
60,76%.
Figura 45. Efecto del NO en la germinación de microconidios de los mutantes de los genes FHG de FOP-SP1 y de la propia cepa silvestre FOP-SP1, evaluadas a las 10 horas de cultivo. (A) cultivos en medio mínimo con 1 % de glucosa, suplementado con 10 mM de tartrato de amonio (MMG+A) y MMG+A en presencia de 250 µM DETA. (B) cultivos en medio mínimo con 1 % de glucosa, suplementado con 10 mM de nitrato de sodio (MMG+N) y MMG+N en presencia de 250 µM DETA. Los histogramas de la izquierda en los Paneles A y B en medios de cultivo MMG+A y MMG+N en ausencia del donador de NO fueron analizados estadísticamente mediante un análisis ANOVA y la discriminación de medias se realizó con el test de Tukey (p<0,05). En los histogramas de la derecha en los Paneles A y B se muestra el análisis estadístico pareado en la cepa de tipo silvestre y en las distintas cepas mutantes en ausencia y presencia de NO (DETA) con el t-test de Student (*p<0,05; ***p<0,001).
Papel de las flavohemoglobinas en F. oxysporum
- 150 -
En la Figura 46 se muestran los porcentajes de reducción de la germinación de
microconidios en los mutantes de los genes FHG, en presencia de 250 µM de DETA en
relación con las cepas control en ausencia del donador de NO. Los porcentajes mostrados
confirman que, efectivamente, el NO afecta a la germinación y que la flavohemoglobina
FHG1 es la enzima detexoficadora de NO que juega un papel más importantes en la
eliminación del NO que afecta a la germinación, pero en función de la fuente de nitrógeno
presente en el medio de cultivo la flavohemoglobina FHG2 también participa en esta
detoxificación: no lo hace en presencia de amonio pero sí en presencia de nitrato.
Figura 46. Porcentajes de reducción de la germinación de microconidios de las cepas mutantes ∆FHG de FOP-SP1. Panel A: Porcentajes de reducción de germinación en medio mínimo con 1 % de glucosa, suplementado con 10 mM de tartrato de amonio (MMG+A) en presencia de 250 µM DETA. Panel B: Porcentajes de reducción de germinación en medio mínimo con 1 % de glucosa, suplementado con 10 mM de nitrato de sodio (MMG+N) en presencia de 250 µM DETA.
3.8.2.2 Análisis de germinación de microconidios de mutantes
ΔFHG de FOP-SP4
En la Figura 47 se presentan los datos de germinación en la cepa FOP-SP4 y en los
transformantes derivados de ésta en medio suplementado con amonio (Panel A) y en
medio suplementado con nitrato (Panel B). El análisis estadístico ANOVA muestra que,
Resultados
- 151 -
tanto en presencia de amonio como de nitrato como única fuente de nitrógeno, en ausencia
de NO exógeno las cepas mutantes no muestran diferencias significativas (p<0,05) en su
eficiencia de germinación en relación con la cepa tipo silvestre. La adición de NO al medio
de cultivo determina una reducción importante en el porcentaje de germinación observado
en la cepa silvestre en ambos medios de cultivo (diferencias significativas en ambos casos
p<0,05, t-test de Student), reducción que es superior a la observada en el caso de la cepa
FOP-SP1 en las mismas condiciones (en medio con amonio es de un 34,88 % y en medio
con nitrato es del 36,51 %). Por lo tanto, es posible deducir que la cepa FOP-SP4 es más
sensible a NO en relación con el efecto que éste tiene sobre germinación que la cepa FOP-
SP1, tanto en medio con amonio como en medio con nitrato.
Figura 47. . Efecto del NO en la germinación de microconidios de los mutantes de los genes FHG de FOP-SP4 y de la propia cepa silvestre FOP-SP4, evaluadas a las 10 horas de cultivo. (A) Cultivos en medio mínimo con 1 % de glucosa, suplementado con 10 mM de tartrato de amonio (MMG+A) y MMG+A en presencia de 250 µM DETA. (B) Cultivos en medio mínimo con 1 % de glucosa, suplementado con 10 mM de nitrato de sodio (MMG+N) y MMG+N en presencia de 250 µM DETA. Los histogramas de la izquierda en los Paneles A y B en medios de cultivo MMG+A y MMG+N en ausencia del donador de NO fueron analizados estadísticamente mediante un análisis ANOVA y la discriminación de medias se realizó con el test de Tukey (p<0,05). En los histogramas de la derecha de los Paneles A y B se muestra el análisis estadístico pareado en la cepa de tipo silvestre y en las distintas cepas mutantes en ausencia y presencia de NO (DETA) con el t-test de Student (**p<0,01; ***p<0,001).
Papel de las flavohemoglobinas en F. oxysporum
- 152 -
Establecida esta diferencia de sensibilidad de la cepa FOP-SP4 en comparación con la
cepa FOP-SP1, el análisis global de los datos generados en la situación “presencia de NO”
demuestra que el comportamiento de las correspondientes cepas mutantes deficientes en
cada uno de los cuatro genes FHG en la cepa FOP-SP4 es el mismo que el observado en
el caso de los mutantes en la cepa FOP-SP1. Así, en medio con amonio es la
flavohemoglobina codificada por FHG1 la enzima que probablemente es la responsable de
la detoxificación del NO que afecta a la germinación. En medio con nitrato, además de
FHG1, la flavohemoglobina codificada por el gen FHG2 parece tener un papel relevante en
la detoxificación de NO.
En la Figura 48, se presenta un resumen de los porcentajes de reducción de la germinación
de microconidios de las distintas cepas mutantes en los genes FHG derivados de la cepa
FOP-SP4 de la que se derivan los respectivos mutantes, en medios de cultivo con amonio
y nitrato en ausencia y en presencia de 250 µM DETA y mostrados en las Figuras 47A y
47B.
Figura 48. Porcentajes de reducción de la germinación de microconidios de las cepas mutantes ∆FHG de FOP-SP4. Panel A: Porcentajes de reducción de germinación en medio mínimo con 1 % de glucosa, suplementado con 10 mM de tartrato de amonio (MMG+A) en presencia de 250 µM DETA. Panel B: Porcentajes de reducción de germinación en medio mínimo con 1 % de glucosa, suplementado con 10 mM de nitrato de sodio (MMG+N) en presencia de 250 µM DETA.
Resultados
- 153 -
3.8.3 Ensayos de infección in planta con los mutantes ΔFHG
Los trabajos descritos en las secciones anteriores indican que las FHG codificadas por al
menos dos de los genes analizados, FHG1 y FHG2, desempeñan una función relevante en
la detoxificación de NO exógeno y en la resistencia a condiciones de estrés nitrosativo.
Además, el análisis de la eficiencia y dinámica de germinación de las cepas mutantes
correspondientes demuestra que el NO tiene un efecto notable sobre la germinación de los
microconidios, y que este efecto parece estar modulado por las enzimas de tipo
flavohemoglobina codificadas por estos dos genes. Teniendo en cuenta que el NO es
producido por la planta durante la interacción con distintos tipos de patógenos y que su
función como molécula señalizadora y ejecutora que coordina y orquesta la respuesta
defensiva de la planta es esencial (Arasimowicz‐Jelonek & Floryszak‐Wieczorek, 2014),
decidimos evaluar la agresividad de los mutantes ΔFHG de FOP-SP1 y FOP-SP4 durante
las interacciones que una y otra cepa establecen con el huésped. Para ello diseñamos una
serie de ensayos de infección de plantas de judía variedad “blanca riñón” que fueron
llevados a cabo siguiendo el procedimiento descrito en el apartado 2.5.1 de Materiales y
Métodos.
En cada ensayo de infección se inocularon 10 plantas de judía con cada estirpe silvestre
considerada, FOP-SP1 y FOP-SP4, y 10 plantas con cada una de las cepas mutantes
seleccionadas (las mismas cepas mutantes analizadas en las secciones anteriores (Tabla
23). Los síntomas y el progreso de la enfermedad fueron evaluados en las plantas
inoculadas con las cepas mutantes y comparadas con los síntomas y el progreso de la
enfermedad en las plantas inoculadas con las estirpes silvestres de las que derivan los
mutantes. El ensayo con cada estirpe fue repetido 3 veces en plantas infectadas con
diferentes lotes de microconidios. El progreso de la enfermedad fue evaluado
semanalmente utilizando la escala CIAT, en la cual las plantas que muestran un índice de
1 a 3 son consideradas como resistentes al patógeno mientras que las plantas con un
índice en la escala de 3,1 a 9 son consideradas como susceptibles al patógeno.
Papel de las flavohemoglobinas en F. oxysporum
- 154 -
Tabla 23. Valores de infección en plantas de judía variedad “Blanca Riñón” inoculadas con las cepas indicadas, estimados según la escala CIAT. En la primera columna se muestran los códigos de las cepas tipo silvestre (FOP-SP1 y FOP-SP4) y de las cepas mutantes utilizadas. En las columnas 2 a 6 se representa el valor medio de la escala CIAT de todas las plantas para el aislado considerado en la semana indicada (± DE). Control: inoculaciones llevadas a cabo con agua.
Los resultados de infección mostrados en la Tabla 23 y en la Figura 49 indican que los
mutantes ∆FHG de FOP-SP1 y de FOP-SP4 no muestran diferencias significativas en su
capacidad de infección de la planta huésped en comparación con la cepa silvestre
correspondiente.
Resultados
- 155 -
Figura 49. Evaluación de la agresividad de los mutantes ∆FHG de FOP-SP1 y FOP-SP4 (ver leyenda) sobre la variedad de judía Blanca Riñón. Como control negativo se utilizaron plantas no infectadas. Se utilizó la escala CIAT (1=planta sana a 9=planta totalmente afectada o muerta) para medir la severidad de los síntomas.
3.9 Ensayos de complementación de la mutación ΔFHG1
Los análisis de expresión llevados a cabo demuestran que FHG1 es el gen codificador de
FHGs que muestra un mayor nivel de inducción en su expresión en respuesta a la
exposición a NO exógeno. Por otra parte, los análisis de sensibilidad a condiciones de
estrés nitrosativo indican que la enzima codificada por este gen es la que juega un papel
más destacado en la resistencia a estrés nitrosativo del patógeno. Finalmente, es en los
mutantes ΔFHG1 en los que el efecto de inhibición de la germinación que observamos es
más evidente. La enzima codificada por FHG1 parece ser, por lo tanto, la enzima más
importante desde el punto de vista funcional en la resistencia a estrés nitrosativo. Para
confirmar que las alteraciones fenotípicas observadas son consecuencia, efectivamente,
de la mutación ΔFHG1, decidimos llevar a cabo un experimento de complementación de la
mutación reintroduciendo la copia silvestre del gen FHG1 en los mutantes SP1-ΔFHG1-
131 y SP4-ΔFHG1-244.
Para ello diseñamos en primer lugar dos oligonucleótidos que permiten amplificar la copia
genómica del gen FHG1. Las secuencias complementarias de estos oligonucleótidos se
Papel de las flavohemoglobinas en F. oxysporum
- 156 -
encuentran en las regiones flanqueantes del gen FHG1, aproximadamente a 1 kb aguas
arriba del codón de iniciación del gen el primero de ellos, y a 1 kb aguas abajo del codón
de terminación del gen el segundo de ellos.
3.9.1 Construcción del vector de complementación
Los oligonucleótidos COMP-FOXG-17-F y COMP-FOXG-17-R (ver Tabla 6) poseen en su
extremo 5’ un sitio de restricción para las enzimas PacI el primero de ellos y NT.Bbv.CI el
segundo. Con estos oligonucleótidos se amplificó mediante PCR un fragmento de ADN
genómico de FOP-SP1 de 3.6 kb que incluye la región estructural del gen FHG1 y 1 kb de
la región 5’ flanqueante y 1 kb de la región flanqueante 3´. El fragmento fue purificado y
clonado en el vector pGEM-T easy. Posteriormente, el inserto clonado fue liberado
mediante digestión con las enzimas PacI y NT.Bbv.CI, recuperado de gel y ligado al vector
pEA-FU (Figura 5) (previamente linearizado con las enzimas PacI y NT.Bbv.CI) utilizando
el sistema User Friendly Cloning. Con el producto de la ligación se transformaron células
competentes de E. coli cepa DHα5. Entre los transformantes obtenidos se identificó un clon
portador del plásmido buscado, que fue denominado vector de transformación pEA-FU-
FHG1, con un tamaño de 5537 pares de bases (ver Figura 50A y Figura 50B).
3.9.2 Transformación de A. tumefaciens
El plásmido pEA-FU-FHG1 (Figura 50A) fue introducido en A. tumefacción cepa AGL1
mediante electroporación siguiendo los procedimientos descritos en la sección 2.9.5.2 de
Materiales y Métodos. De entre las colonias transformantes obtenidas con este plásmido
se seleccionó una que fue utilizada para llevar a cabo la transformación de los mutantes
∆FHG1-131 de FOP-SP1 y ∆FHG1-244 de FOP-SP4. La identidad del plásmido presente
en la colonia seleccionada fue verificada mediante PCR utilizando oligonucleótidos
específicos de las regiones flanqueantes y de la región codificante del gen FHG1.
Resultados
- 157 -
Figura 50. Panel A: Mapa de restricción del plásmido pEA-FU-FHG1 utilizado para la complementación de la mutación ΔFHG1. El fragmento de ADN que incluye la región codificante del gen FHG1, su región promotora y su región terminadora, fue amplificado con los cebadores COMP-FOXG-17-F y COMP-FOXG-17-R. El plásmido incluye el gen ble de resistencia a fleomicina para la selección de transformantes en F. oxysporum. Panel B: Representación detallada de la construcción generada para la complementación de la mutación ΔFHG1 en la que se indica la ubicación de los oligonucleótidos utilizados para la discriminación de transformantes que han incorporado la construcción para la complementación de la mutación ΔFHG1.
3.9.3 Transformación de mutantes ∆FHG1 de FOP-SP1 y FOP-SP4
La transformación de F. oxysporum mediada por A. tumefaciens se realizó siguiendo la
metodología mostrada en los apartados 2.9.5.3 y 2.9.5.4 de Materiales y Métodos. Se
transformó el mutante ∆FHG1-131 de la estirpe FOP-SP1 y el mutante ∆FHG1-244 de la
estirpe FOP-SP4. Los transformantes primarios que fueron apareciendo en las placas de
medio selectivo en presencia de fleomicina fueron transferidos individualmente a placas de
PDA con 400 a 600 µg/ml de fleomicina e incubados durante 5 días a 25ºC. De la estirpe
∆FHG1-131 se seleccionaron 10 transformantes resistentes a fleomicina y de la estirpe
∆FHG1-244 se seleccionaron 8 transformantes.
3.9.4 Identificación de transformantes que han incorporado la
construcción de complementación
La discriminación de transformantes que habían incorporado la copia silvestre del gen
FHG1 se realizó mediante PCR con los cebadores FHG1-FW y FHG1-RV, que permiten
Papel de las flavohemoglobinas en F. oxysporum
- 158 -
amplificar un fragmento de 481 pares de bases derivado de la región codificante del mismo.
La Figura 51A, muestra los productos de amplificación obtenidos utilizando como ADN
molde en las reacciones de PCR ADN genómico de dos transformantes obtenidos en la
estirpe SP1-ΔFHG1-131, los identificados con los códigos SP1-ΔFHG1-131-C1 y SP1-
ΔFHG1-131-C2. En las carreras correspondientes se visualiza el fragmento de ADN del
tamaño esperado (481 nt), el del fragmento amplificado en la reacción preparada utilizando
como ADN molde ADN genómico de la estirpe silvestre FOP-SP1 y ausente en la reacción
de PCR preparada utilizando como ADN molde ADN genómico de la estirpe mutante SP1-
ΔFHG1-131. En la Figura 51B se presenta el análisis correspondiente llevado a cabo sobre
dos transformantes candidato obtenidos en la estirpe SP4-ΔFHG1-244, los identificados
con los códigos SP4-ΔFHG1-244-C1 y SP4-ΔFHG1-244-C2. La amplificación del
fragmento esperado es indicativa de la reintroducción del alelo silvestre del gen FHG1 en
ambas estirpes mutantes.
Las cuatro cepas seleccionadas como candidatas fueron sometidas a un análisis tipo
Southern. Éstas fueron cultivadas en medio de cultivo PDB y a partir del micelio recogido
se extrajo ADN genómico que fue utilizado en el análisis de tipo Southern. Las muestras
de ADN genómico fueron digeridas con la enzima HindIII, que no presenta sitios de
restricción dentro de la construcción de transformación, y como sonda de hibridación se
utilizó un fragmento de ADN derivado de la región 5’ flanqueante del gen FHG1, amplificado
y marcado mediante PCR utilizando los oligonucleótidos FHG175 y RHG175, que permite
detectar tanto el alelo silvestre del gen FHG1 como el alelo mutante derivado del
reemplazamiento del alelo silvestre del gen FHG1.
La Figura 51 muestra los patrones de hibridación obtenidos en el análisis de los
transformantes candidatos obtenidos en la cepa SP1-ΔFHG1-131 (Panel B) y de los
transformantes candidatos obtenidos en la cepa SP4-ΔFHG1-244 (Panel D). Se
comprueba que en ambos casos, es decir, tanto en las cepas derivadas del mutante SP1-
ΔFHG1-131 como en las cepas derivadas del mutante SP4-ΔFHG1-244, es posible
detectar la banda de hibridación que representa el alelo mutante (consecuencia del
reemplazamiento génico), la de mayor tamaño, y una segunda banda de menor tamaño
que representa el alelo silvestre introducido mediante transformación clonado en el
plásmido pEA-FU-FHG1, y que está ausente en las cepas mutantes SP1-ΔFHG1-131 y
SP4-ΔFHG1-244. En la cepa SP1-ΔFHG1-131-C2 comprobamos que la banda
correspondiente al alelo silvestre detectada en la cepa SP1-ΔFHG1-131-C1 no está
presente, lo que supone que esta cepa constituye un falso positivo identificado en el
Resultados
- 159 -
análisis mediante PCR, pero que no puede ser considerado como una cepa en la que se
ha reintroducido el alelo silvestre FHG1.
Llevado a cabo este análisis fueron seleccionados para su análisis funcional posterior un
representante de cada pareja de transformantes analizados, concretamente el
transformante SP1-ΔFHG1-131-C1 de FOP-SP1 y el transformante SP4-ΔFHG1-244-C1
de FOP-SP4.
Figura 51. Identificación de los transformantes de las cepas SP1-ΔFHG1-131 y SP4-ΔFHG1-244 que han incorporado el gen FHG1 mediante transformación con el plásmido pEA-FU-FHG1. Panel A: Productos de PCR amplificados con los cebadores FHG1-FW y FHG1-RV utilizando ADN genómico de los transformantes seleccionados, SP1-ΔFHG1-131-C1 y SP1-ΔFHG1-131-C2 (carriles 1 y 2), de la cepa mutante SP1-ΔFHG1-131 no transformada (carril 3) y de la estirpe silvestre FOP-SP1 (Carril 4). Carril M: marcador de peso molecular 1 kp plus DNA ladder (Invitrogen) Panel B: Análisis Southern de los transformantes seleccionados SP1-ΔFHG1-131-C1 y SP1-ΔFHG1-131-C2 (carriles 1 y 2), de la cepa mutante SP1 -ΔFHG1-131 (carril 3) y cepa silvestre FOP-SP1 (carriles 4). Panel C: Productos de PCR amplificados con los cebadores FHG1-FW y FHG1-RV utilizando ADN genómico de transformantes seleccionados, SP4-ΔFHG1-244-C1 y SP4-ΔFHG1-244-C2 (carriles 1 y 2), de la cepa mutante SP4-ΔFHG1-244 no transformada (carril 3) y de la estirpe silvestre SP4 (Carril 4). Carril M: marcador de peso molecular 1 kp plus DNA ladder (invitrogen). Panel D: Análisis Southern de los transformantes seleccionados, SP4-ΔFHG1-244-C1 y SP4-ΔFHG1-244-C2 (carriles 1 y 2), de la cepa mutante SP4-ΔFHG1-131 (carril 3) y de la cepa silvestre FOP-SP4 (carril 4). En las hibridaciones cuyos resultados se presentan en los paneles B y D, el ADN genómico fue digerido con la enzima HindIII y la sonda utilizada fue una sonda derivada del extremo 5’ flanqueante del gen FHG1.
Papel de las flavohemoglobinas en F. oxysporum
- 160 -
3.9.5 Análisis fenotípico de los mutantes portadores de la
construcción de complementación de la mutación ΔFHG1
El fenotipo de los transformantes seleccionados en los que se había reintroducido el alelo
silvestre del gen FHG1 fue comparado con el de las cepas silvestres correspondientes y
con el de sus cepas mutantes ΔFHG1, en relación con los dos aspectos fisiológicos
considerados inicialmente, es decir, la sensibilidad a condiciones de estrés nitrosativo y la
eficiencia de germinación. Este análisis se llevó a cabo en las mismas condiciones
experimentales y aplicando los mismos procedimientos utilizados entonces. Así, para la
evaluación de la sensibilidad a estrés nitrosativo las cepas analizadas fueron cultivadas en
placas de Petri con medios MMG+A y MMG+N sólidos, en presencia y en ausencia del
donador de NO (SNP 500 µM). Para cada cepa y condición de cultivo se prepararon 3
placas y el crecimiento de cada colonia fue evaluado a los 4 y 5 días de cultivo. Para la
evaluación de la eficiencia de germinación las suspensiones de microconidios de las cepas
analizadas fueron cultivadas en medios MMG+A y MMG+N líquidos en presencia y en
ausencia del donador de NO (250 µM DETA) y los recuentos se efectuaron a las 10 horas
de cultivo. El experimento fue repetido tres veces.
En la figura 52 se presentan los resultados obtenidos en la evaluación de la sensibilidad a
estrés nitrosativo de las cepas SP1-ΔFHG1-131-C1 (Panel A) y SP4-ΔFHG1-244-C1
(Panel B) en comparación con sus respectivas cepas de referencia. Los datos de diámetro
de la colonia fueron tomados a los 5 días de crecimiento saprofitico en las condiciones
indicadas. En su análisis se comprueba que las dos cepas mutantes, SP1-ΔFHG1-131 y
SP4-ΔFHG1-244, y en ambos medios de cultivo (en presencia de amonio y en presencia
de nitrato como fuente de nitrógeno), manifiestan el fenotipo de hipersensibilidad a NO
observado previamente, y que este fenotipo es revertido una vez reintroducido el alelo
silvestre del gen FHG1, ya que el diámetro de colonia de las cepas SP1-ΔFHG1-131-C1 y
SP4-ΔFHG1-244-C1 es igual al de las colonias de las cepas FOP-SP1 (panel A) y FOP-
SP4 (Panel B), respectivamente. Es decir, la reintroducción del alelo silvestre permite
restaurar el fenotipo silvestre de resistencia a condiciones de estrés nitrosativo. Se puede
deducir, por lo tanto, que la reducción del crecimiento en respuesta a NO exógeno
mostrada por los mutantes SP1-∆FHG1 y SP4-∆FHG1 es consecuencia de la mutación
inducida en el gen FHG1.
Resultados
- 161 -
Figura 52. Evaluación de la sensibilidad a estrés nitrosativo de los mutantes complementados SP1-ΔFHG1-131-C1 y SP4-ΔFHG1-244-C1 cultivados en MMG+A y MMG+N en ausencia y en presencia del donador de NO (500 µM SNP). Panel A: Diámetro de la colonia de las cepas: FOP-SP1 (control), mutante nulo SP1-ΔFHG1-131 y cepa complementada SP1-ΔFHG1-131-C1. Panel B: Diámetro de la colonia de las cepas FOP-SP4 (control), mutante nulo SP4-ΔFHG1-244 y cepa complementada SP4-ΔFHG1-244-C1 en las condiciones indicadas. Los datos presentados corresponden a los valores medios de tres experimentos y sus correspondientes desviaciones estándar. Con los datos de las tres cepas analizadas en cada caso en cada medio y condición se llevó a cabo un análisis ANOVA. Medias con letra diferente en cada combinación son significativamente diferentes (p< 0,05) (test de Tukey).
En la figura 53 se presentan los resultados obtenidos en el análisis de la germinación de
microconidios de las cepas SP1-ΔFHG1-131-C1 (Panel A) y SP4-ΔFHG1-244-C1 (Panel
B) en comparación con sus respectivas cepas de referencia. Estos resultados confirman
también las observaciones realizadas inicialmente, indicando que la exposición a NO
determina una reducción de la germinación de los dos cepas silvestres utilizadas en
nuestro trabajo, FOP-SP1 y FOP-SP4, tanto en presencia de amonio como en presencia
de nitrato. La reducción es más evidente en el caso de los mutantes SP1-ΔFHG1-131 y
SP4-ΔFHG1-244. Los transformantes en los que se ha reintroducido el alelo silvestre del
gen FHG1, SP1-ΔFHG1-131-C1 y SP4-ΔFHG1-244-C1, recuperan los niveles de
germinación característicos de la correspondiente cepa silvestre en cada medio de cultivo.
Papel de las flavohemoglobinas en F. oxysporum
- 162 -
Figura 53. Evaluación de la eficiencia de germinación de microconidios de los mutantes complementados SP1-ΔFHG1-131-C1 y SP4-ΔFHG1-244-C1 cultivados en MMG+A y MMG+N en ausencia y en presencia del donador de NO (250 µM DETA) tomados a las 10 horas de cultivo. Panel A: Porcentaje de germinación de las cepas FOP-SP1 (control), del mutante nulo SP1-ΔFHG1-131 y de la cepa complementada SP1-ΔFHG1-131-C1 en las condiciones indicadas. Panel B: Porcentaje de germinación de las cepas FOP-SP4 (control), del mutante nulo SP4-ΔFHG1-244 y de la cepa complementada SP4-ΔFHG1-244-C1 en las condiciones indicadas. Los datos presentados corresponden a los valores medios de tres experimentos y sus correspondientes desviaciones estándar. Con los datos de las tres cepas analizadas en cada caso en cada medio y condición se llevó a cabo un análisis ANOVA. Las medias con letra diferente en cada condición son significativamente diferentes (p < 0,05) (test de Tukey).
Teniendo en cuenta la caracterización llevada a cabo en las cepas transformantes en las
que se ha reintroducido el alelo silvestre del gen FHG1 es posible concluir que el fenotipo
de hipersensibilidad a NO que muestran las cepas mutantes en las que este gen ha sido
eliminado, tanto en el fondo genético FOP-SP1 como en el fondo genético FOP-SP4, y
como consecuencia del cual muestran una reducción de crecimiento en medio sólido y una
reducción de la eficiencia de germinación en medio líquido en condiciones de estrés
nitrosativo, es consecuencia exclusivamente de la mutación introducida.
Resultados
- 163 -
3.10 Obtención y caracterización de dobles mutantes ΔFHG2-FHG1RNAi.
El nivel de inducción de la expresión de FHG1 en respuesta a NO exógeno durante la fase
de germinación de las esporas, y el fenotipo de los mutantes ΔFHG1 en las cepas FOP-
SP1 y FOP-SP4, hipersensibles a condiciones de estrés nitrosativo, indican que este gen
codifica la enzima flavohemoglobina que desempeña un papel más relevante en relación
con la resistencia a condiciones de estrés nitrosativo de FOP. El gen FHG2 también
responde a nivel transcripcional a la exposición a NO exógeno, aunque de manera mucho
más limitada. Además, los mutantes ΔFHG2 muestran un cierto aumento en su sensibilidad
a condiciones de estrés nitrosativo, si bien también en este caso de forma mucho más
limitada que los mutantes ΔFHG1. Estas observaciones nos llevaron a plantear la
conveniencia de obtener mutantes dobles alterados simultáneamente en ambos genes,
FHG1 y FHG2, y analizar su fenotipo para determinar en qué medida ambas enzimas
contribuyen a la resistencia a estrés nitrosativo. El patrón de expresión de FHG2 indica que
este gen responde a otros estímulos relacionados con la fuente de carbono, la fuente de
nitrógeno y el desarrollo, además de la exposición a NO exógeno. Es posible asumir que
la enzima flavohemoglobina codificada por FHG2 pueda estar implicada en más de una
función fisiológica.
La primera estrategia experimental consistió en generar, mediante reemplazamiento
génico, mutantes alterados en FHG2 en un fondo genético ΔFHG1 (tanto en la cepa
silvestre FOP-SP1 como en la cepa FOP-SP4). Después de varios intentos de
transformación con resultados negativos (no se obtuvieron transformantes) decidimos
aplicar una estrategia alternativa, basada en silenciamiento génico. En la misma se
consideró inducir el silenciamiento del gen FHG1 sobre el fondo genético de los mutantes
∆FHG2 de FOP-SP1 y FOP-SP4. Para ello, se amplificó una secuencia de la región
codificante del gen FHG1 y se ligó a un vector de silenciamiento que posee dos promotores
en direcciones opuestas cuya actividad genera un ARN bicatenario que es reconocido por el
complejo de silenciamiento Dicer. La aplicación de esta estrategia supuso desarrollar las
siguientes fases y procedimientos: (1) diseño de cebadores para la amplificación de un
fragmento de ADN de la región codificante del gen FHG1 para su clonación en un vector
de silenciamiento; (2) construcción de un vector de silenciamiento en un plásmido portador
de un marcador de resistencia a fleomicina, dado que los mutantes deficientes en el gen
FHG2 ya poseen un gen de resistencia a higromicina B; (3) transformación de E. coli y A.
tumefaciens con los plásmidos generados; (4) transformación de mutantes deficientes en
FHG2 de FOP-SP1 y FOP-SP4 con el vector de silenciamiento; (5) identificación de
Papel de las flavohemoglobinas en F. oxysporum
- 164 -
transformantes en los que se ha silenciado el gen FHG1; y (6) análisis fisiológico de
mutantes silenciados de FOP-SP1 y FOP-SP4.
3.10.1 Construcción del vector de silenciamiento
Con los cebadores Sil-FHG1-FW Y Sil-FHG1-RV (Tabla 6) (diseñados con un sitio de
restricción NcoI en el extremos 5’ para facilitar su clonación) se amplificó un fragmento de
281 pares de nucleótidos de la región codificante del gen FHG1 (ver Figura 54). Este
fragmento fue purificado y clonado en el vector pGEM-T easy. Una vez liberado del
plásmido derivado mediante la digestión con la enzima NcoI el inserto fue ligado al vector
pVIR-RNAi (Figura 5) (previamente linearizado con la enzima NcoI). Con la mezcla de
ligación se transformó E. coli cepa DHα5. Entre los transformantes obtenidos se identificó
una colonia portadora del plásmido deseado, que recibió el nombre de pVIR-RNAi-FHG1,
cuyo tamaño es de 7535 pares de bases (ver Figura 55A).
Figura 54. Producto de PCR amplificado a partir de ADN genómico de FOP-SP1 con los cebadores Sil-FHG1-FW y Sil-FHG1-RV visualizado en gel de agarosa 2 % (carril 1). Carril (M): Marcador de peso molecular 1 Kb Plus DNA ladder (Invitrogen).
3.10.2 Transformación de A. tumefaciens
El plásmido pVIR-RNAi-FHG1 (Figura 55A) fue introducido en A. tumefaciens cepa AGL1
mediante electroporación siguiendo los procedimientos descritos en la sección 2.9.5.2 de
Materiales y Métodos. De entre las colonias transformantes obtenidas se seleccionó una
que fue utilizada para llevar a cabo la transformación de los mutantes SP1-∆FHG2-51 de
FOP-SP1 y SP4-∆FHG2-314 de FOP-SP4. La identidad del plásmido presente en la colonia
Resultados
- 165 -
seleccionada fue verificada mediante PCR utilizando oligonucleótidos específicos de los
promotores del casete de silenciamiento y de la región codificante del gen de resistencia a
fleomicina (Figura 55B).
Figura 55. Panel A: Mapa de restricción del plásmido pVIR-RNAi-FHG1 utilizado para el silenciamiento del gen FHG1. Panel B: Representación detallada de la región clonada entre los extremos RB y LB del plásmido pVIR-RNAi-FHG1. Se indica la ubicación de los sitios de unión de los cebadores utilizados para la discriminación de transformantes que han incorporado el plásmido de la construcción de silenciamiento.
3.10.3 Transformación de mutantes ∆FHG2 con el vector de
silenciamiento pVIR-RNAi-FHG1
La transformación de F. oxysporum mediada por A. tumefaciens se realizó siguiendo la
metodología mostrada en los apartados 2.9.5.3 y 2.9.5.4 de Materiales y Métodos. Se
transformaron los mutantes deficientes del gen FHG2, tanto en la cepa FOP-SP1,
identificado como mutante SP1-∆FHG2-51, como en la cepa FOP-SP4, identificado como
mutante SP4-∆FHG2-314. Los transformantes primarios que fueron apareciendo en las
placas de medio selectivo en presencia de fleomicina fueron transferidos individualmente
a placas de PDA con 400 a 600 µg/ml de fleomicina e incubados durante 5 días a 25ºC. Se
seleccionaron 10 transformantes para cada estirpe (SP1 y SP4).
Papel de las flavohemoglobinas en F. oxysporum
- 166 -
3.10.3.1 Identificación y selección de transformantes ΔFHG2-
FHG1RNAi
Los transformantes seleccionados fueron cultivados en medio de cultivo PBD durante 3
días a 25ºC y 120 rpm. El micelio producido fue recuperado por filtración y utilizado para la
extracción de ARN total con el protocolo descrito en el apartado 2.6.4.2 de Materiales y
Métodos. La cantidad y calidad del ARN fueron verificadas por espectrofotometría y
electroforesis en gel de agarosa 1 % en MOPS 1X.
La síntesis de cDNA a partir de ARN total y el procedimiento para la preparación de las
muestras para las reacciones de RT-qPCR se describe en los apartados 2.12.2 y 2.12.3 de
Materiales y Métodos. Los niveles de expresión del gen FHG1 en las muestras preparadas
fueron normalizados con los niveles de expresión constitutiva del gen codificador del factor
de elongación 1α (EF1α) y referidos a los valores de expresión del gen FHG1 de las cepas
silvestres correspondientes (la cepa FOP-SP1 o la cepa FOP-SP4).
En la Figura 56 se presentan los resultados de los análisis de cuantificación de los niveles
de expresión del gen FHG1 en los transformantes seleccionados a partir de las cepas
mutantes SP1-ΔFHG2-51 (Panel A) y SP4-ΔFHG2-314 (Panel B) transformadas con el
vector de silenciamiento pVIR-RNAi-FHG1. En cada panel cada barra representa el nivel
de expresión relativa del gen FHG1 en la cepa indicada en relación con el nivel de
expresión en la cepa silvestre. La expresión se cuantificó en esporas germinadas tras 9 h
de incubación en medio MMG+A+SNP, condiciones en las cuales se había comprobado
previamente que se produce el mayor incremento de expresión de FHG1 (ver en la sección
3.5.2.1 Resultados). Se comprueba que los distintos transformantes silenciados muestran
niveles de expresión variables del gen FHG1. Para los siguientes análisis se seleccionaron
los transformantes que mostraron los niveles más bajos de expresión, esto es SP1-ΔFHG2-
FHG1RNAi-17, SP4-ΔFHG2-314-FHG1RNAi-18 y SP4-ΔFHG2-314-FHG1RNAi-22.
Figura 56. . Expresión relativa del gen FHG1 en los transformantes seleccionados obtenidos con el plásmido pVIR-RNAi-FHG1 sobre el fondo genético de los mutantes SP1-∆FHG2-51 y SP4-∆FHG2-314. Las muestras de RNA utilizadas en este análisis se obtuvieron de esporas cultivadas durante 9 h en medio mínimo con 1 % glucosa, suplementado con 10 mM de tartrato amónico (MMG+A), en presencia de 500 µM de SNP. Panel A: Cepa silvestre FOP-SP1, cepa mutante SP1-∆FHG2-51 y mutantes silenciados SP1-∆FHG2-51-FHG1RNAi-1, 2, 3, 4, 7, 17, 19, 22, 27 y 30 (carreras 1 a 30). Panel B: Cepa silvestre FOP-SP4, cepa mutante SP4-∆FHG2-314 y mutantes silenciados SP4-∆FHG2-314-FHG1RNAi-1, 2, 3, 16, 17, 18, 20, 22, 31 y 32 (carreras 1 a 32). Los datos de expresión representan los niveles de expresión relativa del gen FHG1 normalizados en relación con el nivel de expresión del gen Efα1 y referidos en cada caso a los valores de expresión de las cepas silvestre
Para demostrar que en los transformantes en los que hemos detectado silenciamiento está
presente, efectivamente, la construcción de silenciamiento decidimos llevar a cabo una
caracterización de los mismos mediante PCR y mediante hibridación tipo Southern. Para
ello en primer lugar se extrajo ADN genómico de las cepas mutantes silenciadas, de las
cepas SP1-∆FHG2-51 y SP4-∆FHG2-314 y de las estirpes silvestres FOP-SP1 y FOP-SP4.
Para la caracterización mediante PCR las muestras de ADN genómico obtenidas fueron
utilizadas directamente. Para el análisis mediante hibridación Southern el ADN en cada
caso fue digerido con la enzima HindIII y separado por electroforesis en gel de agarosa 1
Papel de las flavohemoglobinas en F. oxysporum
- 168 -
% y transferido a filtros de nylon. Los filtros resultantes fueron hibridados con una sonda
derivada de la región de los promotores de silenciamiento incluidos en el plásmido de
transformación amplificada por PCR con los cebadores RNAi-U4-FW y Sil-FHG1-RV (ver
ubicación de los cebadores en la Figura 55B).
En la Figura 57 se presentan los resultados obtenidos en esta caracterización. En el Panel
A se muestran los productos de PCR amplificados a partir del ADN genómico de la cepa
mutante SP1-∆FHG2-51 y de los transformantes SP1-ΔFHG2-51-FHG1RNAi-1, 2, 3, 4 y 7
en reacciones de PCR llevadas a cabo con los cebadores RNAi-U4-FW y Sil-FHG1-RV
diseñados a partir de la secuencia del ADN del plásmido pVIR-RNAi-FHG1. El cebador Sil-
FHG1-RV hibrida específicamente con un extremo del fragmento de ADN con la secuencia
de silenciamiento y el cebador RNAi-U4-FW hibrida con la región correspondientes al
promotor 1 del casete de silenciamiento (ver Figura 57B). Con el molde adecuado esta
combinación de cebadores permite amplificar un fragmento de 768 pares de bases. Como
se comprueba en el Panel A de la Figura 57, en todos los transformantes seleccionados se
detecta la amplificación de un fragmento de ADN del tamaño esperado. En el Panel B de
la Figura 57 se presenta el análisis Southern blot de estos mismos transformantes. En este
análisis los filtros preparados fueron probados con una sonda marcada por PCR en una
reacción llevada a cabo utilizando los cebadores RNAi-U4-FW y Sil-FHG1-RV (descritos
anteriormente y utilizados en el análisis mediante PCR). Los resultados obtenidos muestran
integración del casete de silenciamiento en diferentes posiciones en el genoma de la cepa
receptora (sólo en el caso de los transformanes 3 y 4 el tamaño de la banda de hibridación
detectada parece ser el mismo). En la Figura 57, Paneles C y D se presentan los resultados
obtenidos aplicando las mismas aproximaciones experimentales (análisis mediante PCR
en el caso del Panel C y análisis mediante hibridación Southern en el caso del Panel D) de
los transformantes SP4-ΔFHG2-314-FHG1RNAi-2, 3, 16, 17, 18 y 22. En todos los
transformantes analizados se detecta amplificación de un producto de PCR del tamaño
esperado, confirmando la presencia del casete de silenciamiento, y las bandas de
hibridación detectadas con la sonda utilizada muestran tamaños diferentes, indicativos de
integraciones del casete de silenciamiento en posiciones diferentes del genoma de la cepa
receptora.
Bansándonos en los resultados mostrados decidimos seleccionar dos transformantes
silenciados en cada cepa mutante transformada para los posteriores análisis fisiológicos.
Estos fueron los transformantes SP1-∆FHG2-51-FHG1RNAi-1 y SP1-∆FHG2-51-
Resultados
- 169 -
FHG1RNAi-7 derivados de la cepa SP1-∆FHG2-51 y los transformantes SP4-∆FHG2-314-
FHG1RNAi-2 y SP4-∆FHG2-314-FHG1RNAi-18 derivados de la cepa SP4-∆FHG1-314.
Figura 57. . Caracterización molecular de los transformantes SP1-ΔFHG2-51 y SP4-ΔFHG2-314 seleccionados en los experimentos de transformación llevados a cabo con el plásmido de silenciamiento pVIR-RNAi-FHG1. Panel A: Productos de PCR amplificados a partir de ADN genómico de transformantes silenciados de SP1-ΔFHG2-51 con los cebadores RNAi-U4-FW y Sil-FHG1-RV. Carril M: Marcador de peso molecular 1kp plus DNA ladder (invitrogen). Carriles (1, 2, 3, 4 y 7) producto de amplificación de ADN en los mutantes silenciados SP1-∆FHG2-51-FHG1RNAi-1, 2, 3, 4 y 7. Panel B: Análisis mediante hibridación Southern de transformantes silenciados de SP1-ΔFHG2-51 hibridado con la sonda derivada del promotor e inserto del casete de silenciamiento. Carril 51: ADN de la cepa SP1-∆FHG2-51. Carriles (1, 2, 3, 4 y 7) ADN de los mutantes silenciados SP1-∆FHG2-51-FHG1RNAi-1, 2, 3, 4 y 7. Panel C: Producto de PCR amplificado a partir de ADN genómico de transformantes silenciados de SP4-ΔFHG2-314 con los cebadores RNAi-U4-FW y Sil-FHG1-RV. Carril M: Marcador de peso molecular 1kp plus DNA ladder (invitrogen). Carriles (2, 3, 16, 17, 18 22) producto de amplificación de ADN en los mutantes silenciados SP4-∆FHG2-314-FHG1RNAi-2, 3, 16, 17, 18 22. Panel D: Análisis mediante hibridación Southern de transformantes silenciados de FOP-SP1 hibridado con la sonda derivada del promotor e inserto del casete de silenciamiento. Carril 314: ADN de la cepa SP4-∆FHG2-314. Carriles (2, 3, 16, 17, 18y 22) ADN de los mutantes silenciados SP4-∆FHG2-314-FHG1RNAi-2, 3, 16, 17, 18 22.
3.10.3.2 Análisis fenotípico de los dobles mutantes ΔFHG2-
FHG1RNAi
Se evaluó la capacidad de crecimiento saprofítico y la sensibilidad a estrés nitrosativo de
los transformantes silenciados seleccionados en comparación con los mutantes ∆FHG2 de
los que se derivan, SP1-∆FHG2-51 y SP1-∆FHG2-314, y sus correspondientes cepas tipo
silvestre, FOP-SP1 y FOP-SP4, en ausencia y en presencia de un donador de NO. Para
Papel de las flavohemoglobinas en F. oxysporum
- 170 -
ello todas las cepas objeto de estudio fueron cultivadas en placas Petri con medio mínimo
con 1 % de glucosa suplementado con amonio o nitrato, en presencia y ausencia del
donador de NO (500 µM de SNP) en las mismas condiciones que las descritas en la sección
3.8.1 de Resultados.
En la Figura 58 se muestran los resultados del análisis de resistencia/sensibilidad a
condiciones de estrés nitrosativo de las cepas mutantes SP1-∆FHG2-51-FHG1RNAi-1 y
SP1-∆FHG2-51-FHG1RNAi-7 de FOP-SP1 y SP4-∆FHG2-314-FHG1RNAi-2 y SP4-
∆FHG2-314-FHG1RNAi-18 de FOP-SP4 en comparación con las cepas mutantes
deficientes en el gen FHG2 de las que derivan (SP1-∆FHG2-51 y SP4-∆FHG2-314) y de
las propias cepas tipo silvestre FOP-SP1 y FOP-SP4. En el Panel A se presenta el análisis
de las distintas cepas derivadas de FOP-SP1 en medios suplementados con amonio y
nitrato como fuente de nitrógeno y en el Panel B el análisis correspondiente llevado a cabo
con las cepas derivadas de FOP-SP4.
El análisis estadístico demuestra que los mutantes silenciados de FOP-SP1 en las dos
fuentes de nitrógeno consideradas y en ausencia de NO exógeno no muestran diferencias
significativas en el diámetro de la colonia en relación con la cepa tipo silvestre y con la cepa
deficiente del gen ΔFHG2. La cepa silvestre FOP-SP1 y la cepa deficiente del gen FHG2,
SP1-ΔFHG2-51, muestran sensibilidad a NO en ambos medios (con amonio y con nitrato),
en unos niveles similares a los observados en los ensayos llevados a cabo anteriormente
(sección 3.8.1.1 de Resultados). Efectivamente, FOP-SP1 muestra sensibilidad a NO y es
más sensible aún la cepa mutante SP1-ΔFHG2-51. El análisis del comportamiento de los
mutantes silenciados permite comprobar que la cepa SP1-ΔFHG2-51-FHG1RNAi-1
muestra un aumento adicional de su sensibilidad a NO, detectándose diferencias
significativas en relación con la cepa mutante SP1-ΔFHG2-51 en el diámetro medio de la
colonia en presencia de NO (p<0,05), con una reducción del 20,5 % en medio con amonio
y del 18,42 % en medio con nitrato en relación con aquél (ver Figura 58A). El segundo
mutante silenciado seleccionado mostró un aumento de sensibilidad a NO mínimo en
relación con la cepa mutante SP1-ΔFHG2-51, no detectándose diferencias significativas
en el análisis estadístico.
El comportamiento de los mutantes silenciados de FOP-SP4 es similar al observado en los
mutantes de FOP-SP1. La cepa de tipo silvestre y la cepa mutante en el gen FHG2, SP4-
ΔFHG2-314, muestran niveles de sensibilidad a NO similares a los mostrados en ensayos
anteriores (sección 3.8.1.2 de Resultados). En cuanto al comportamiento de los mutantes
Resultados
- 171 -
silenciados en el gen FHG1 analizados, se comprueba que en los dos medios
considerados, tanto en presencia de amonio como de nitrato como fuente de nitrógeno,
ambas cepas, SP4-ΔFHG2-314-FHG1RNAi-2 y SP4-ΔFHG2-314-FHG1RNAi-18,
muestran un aumento adicional de sensibilidad a NO en comparación con la cepa SP4-
ΔFHG2-314, y que ambas cepas tienen un comportamiento muy similar que reconoce el
análisis estadístico. La reducción del diámetro de la colonia en relación con la cepa mutante
SP4-ΔFHG2-314 es del 21,62 % y del 18,9 % para las cepas SP4-ΔFHG2-314-FHG1RNAi-
2 y SP4-ΔFHG2-314-FHG1RNAi-18, respectivamente, en medio con amonio y del 15,79 %
y 18,42 % en medio con nitrato (ver Figura 58B).
Figura 58. Evaluación del crecimiento saprofitico de mutantes ΔFHG2 en los que se ha silenciado el gen FHG1. Panel A: Cepa silvestre FOP-SP1, cepa mutante SP1-∆FHG2-51 y mutantes silenciados SP1-ΔFHG2-51-FHG1RNAi-1 y SP1-ΔFHG2-51-FHG1RNAi-7. Panel B: Cepa silvestre SP4, cepa mutante SP4-∆FHG2-314 y mutantes silenciados SP4-ΔFHG2-314-FHG1RNAi-2 y FOP-SP4-ΔFHG2-314-FHG1RNAi-18. Los cultivos se realizaron en medio mínimo con 1 % glucosa con amonio (MMG+A) y medio mínimo con 1 % glucosa con nitrato (MMG+N) en ausencia y presencia de 500 µM de SNP. Los datos presentados corresponden a los valores medios de tres experimentos y sus correspondientes desviaciones estándar. Con los datos de las tres cepas analizadas en cada caso en cada medio y condición se llevó a cabo un análisis ANOVA. Las medias con letra diferente en cada condición de cultivo son significativamente diferentes (p<0,05) (test de Tukey).
Papel de las flavohemoglobinas en F. oxysporum
- 172 -
En resumen, en los mutantes silenciados de FOP-SP1, solamente la cepa silenciada SP1-
ΔFHG2-51-FHG1RNAi-1 muestra una reducción adicional del diámetro de la colonia
estadísticamente significativa en presencia de NO, tanto en medio suplementado con
amonio como en medio suplementado con nitrato como fuente de nitrógeno, en relación
con la cepa utilizada como referencia, el mutante SP1-ΔFHG2-51. En los mutantes
silenciados de FOP-SP4, tanto la cepa SP4-ΔFHG2-314-FHG1RNAi-2 como la cepa SP4-
ΔFHG2-314-FHG1RNAi-18 muestran sensibilidad adicional a NO (SNP) en ambos medios
de cultivo en relación con la cepa de referencia, en este caso el mutante SP4- ΔFHG2-314.
Para evaluar la eficiencia de germinación de microconidios en los mutantes silenciados se
utilizaron las mismas cepas utilizadas en los ensayos de evaluación de crecimiento
saprofitico y de sensibilidad a estrés nitrosativo. Como fuente de NO se utilizó DETA 250
µM. El propósito de estos ensayos fue evaluar los efectos fisiológicos que pudiera tener el
NO exógeno sobre la germinación de microconidios en los mutantes silenciados en
comparación con las cepas deficientes en el gen ∆FHG2 en las cepas SP1 y SP4 (cepas
SP1-∆FHG2-51 y SP4-∆FHG2-314) de las cuales se derivan cada uno de los mutantes
silenciados y con sus respectivas cepas silvestres FOP-SP1 y FOP-SP4, de las mismas
que ya se conoce su comportamiento en ausencia y presencia de NO (DETA). Los medios
de cultivo, el tiempo de evaluación y el análisis estadístico fueron los mismos utilizados en
ensayos anteriores y descritos en la sección 3.8.2 de Resultados.
Los resultados obtenidos se presentan en la Figura 59. Los mutantes silenciados de FOP-
SP1 no muestran diferencias significativas en la eficiencia de germinación en ausencia de
NO en relación con su cepa tipo silvestre FOP-SP1 y con la cepa deficiente del gen ΔFHG2
correspondiente, SP1-ΔFHG2-51. Por otro lado, estas dos cepas muestran sensibilidad a
NO en los dos medios considerados (con amonio y con nitrato) y unos niveles de reducción
en la eficiencia de germinación en presencia de NO similares a los observados en ensayos
anteriores (sección 3.8.2.1 de Resultados). En cuanto a los mutantes silenciados, SP1-
ΔFHG2-51-FHG1RNAi-1 y SP1-ΔFHG2-51-FHG1RNAi-7, se comprueba que ambas cepas
muestran una reducción adicional en la eficiencia de germinación en presencia de NO
(DETA 250 µM) al ser comparadas con la cepa mutante SP1-ΔFHG2-51 (y con la propia
cepa de tipo silvestre FOP-SP1), reducción en el eficiencia de germinación de un 19,8 % y
17,1 %, respectivamente, en medio con amonio y del 24 % y 22 %, respectivamente, en
medio con nitrato (ver Figura 59A).
Resultados
- 173 -
Figura 59. Cuantificación de la germinación de microconidios de mutantes ΔFHG2 en los que se ha silenciado el gen FHG1. Panel A: Cepa silvestre FOP-SP1, cepa mutante SP1-∆FHG2-51 y mutantes silenciados SP1-ΔFHG2-51-FHG1RNAi-1 y SP1-ΔFHG2-51-FHG1RNAi-7. Panel B: Cepa silvestre SP4, cepa mutante SP4-∆FHG2-314 y mutantes silenciados SP4-ΔFHG2-314-FHG1RNAi-2 y SP4-ΔFHG2-314-FHG1RNAi-18. Los cultivos se realizaron en medio mínimo con 1 % glucosa con amonio (MMG+A) y medio mínimo con 1 % glucosa con nitrato (MMG+N) en ausencia y presencia de 250 µM de DETA. Los datos presentados corresponden a los valores medios de tres experimentos y sus correspondientes desviaciones estándar. Con los datos de las tres cepas analizadas en cada caso en cada medio y condición se llevó a cabo un análisis ANOVA. Medias con letra diferente en cada condición de cultivo son significativamente diferentes (p<0,05) (test de Tukey).
Los resultados obtenidos en el análisis de los mutantes silenciados derivados de FOP-SP4
son similares a los obtenidos con los mutantes derivados de FOP-SP1. La cepa tipo
silvestre, FOP-SP4, y la cepa deficiente en el gen FHG2, SP4-ΔFHG2-314, muestran
niveles de reducción en la eficiencia de germinación en presencia de NO similares a los
observados en ensayos anteriores (sección 3.8.2.2 de Resultados). Por su parte, los dos
mutantes silenciados, SP4-ΔFHG2-314-FHG1RNAi-2 y SP4-ΔFHG2-314-FHG1RNAi-18,
muestran en presencia de NO una reducción adicional en la eficiencia de germinación en
relación con la cepa mutante SP4-ΔFHG2-314 (y la cepa de tipo silvestre FOP-SP4),
reducción que alcanza valores del 15,7 % y 20,1 %, respectivamente, en medio con amonio
y del 22 % y 19,9 %, respectivamente, en medio con nitrato. Las diferencias con la cepa
Papel de las flavohemoglobinas en F. oxysporum
- 174 -
SP4-ΔFHG2-314 son significativas y el análisis estadístico demuestra que ambas cepas
mutantes muestran un comportamiento muy uniforme en los dos medios de cultivo (ver
Figura 59B).
3.10.3.3 Ensayos de infección in planta con los dobles mutantes
∆FHG2-FHG1RNAi
Finalmente se evaluó la agresividad sobre judía de los transformantes ΔFHG2 silenciados
en el gen FHG1 de FOP-SP1 y FOP-SP4 seleccionados, en comparación con la
agresividad de las cepas mutantes de ΔFHG2 de las que derivan y de las propias cepas
tipo silvestre. Los procedimientos de infección de plántulas de judía y la evaluación de la
agresividad de los distintos aislados fueron los mismos utilizados previamente y descritos
en secciones anteriores.
Los datos obtenidos en la evaluación de la agresividad durante cinco semanas se
presentan en la Tabla 24. Estos datos se representan gráficamente en la Figura 60. Se
comprueba que los dos transformantes SP1-ΔFHG2-51 seleccionados (SP1-ΔFHG2-51-
FHG1RNAi-1 y SP1-ΔFHG2-51-FHG1RNAi-7) muestran la misma agresividad que la cepa
SP1-ΔFHG2-51 de la que derivan y que la cepa tipo silvestre FOP-SP1. De la misma
manera, los dos transformantes SP4-ΔFHG2-314 seleccionados (SP4-ΔFHG2-314-FHG1-
RNAi-2 y SP4-ΔFHG2-314-FHG1RNAi-18) muestran la misma agresividad que la cepa
SP4-ΔFHG2-314 de la que derivan y que la cepa tipo silvestre FOP-SP4.
Tabla 24. Ensayos de infección en plantas de judía variedad “Blanca Riñón” con los mutantes silenciados (notación). En la primera columna se muestran las cepas tipo silvestre y los mutantes analizados. En las columnas 2 a 6 se representa el valor medio de la escala CIAT de todas las plantas para el aislado considerado en la semana indicada (± DE). Control: inoculaciones llevadas a cabo con agua.
Figura 60. Evaluación de la agresividad de FOP-SP1 y FOP-SP4, de los mutantes SP1-ΔFHG2-51 y SP4-ΔFHG2-314 y de los transformantes silenciados seleccionados derivados de cada uno de ellos, SP1-ΔFHG2-51-FHG1RNAi-1 y SP1-ΔFHG2-51-FHG1RNAi-7, y SP4-ΔFHG2-314-FHG1RNAi-2 y SP4-ΔFHG2-314-FHG1RNAi-18, sobre la variedad de judía Blanca Riñón. Como control negativo se utilizaron plantas inoculadas con agua. Se utilizó la escala CIAT (1=planta sana a 9=planta totalmente afectada o muerta) para medir la progresión de la infección durante 5 semanas.
Papel de las flavohemoglobinas en F. oxysporum
- 176 -
DISCUSIÓN
Discusión
- 177 -
4 DISCUSIÓN
4.1 Relaciones filogenéticas de los genes FHG de FOP
Las proteínas de tipo flavohemoglobina derivan de globinas como consecuencia de
procesos que tuvieron lugar antes de la emergencia de los archea, los eucariotas y las
bacterias (Vinogradov et al., 2005). Los ancestros de las actuales flavohemoglobinas
probablemente desaparecieron en los archea, permaneciendo en bacterias y en eucariotas
(Vinogradov et al., 2005). Es interesante destacar el hecho de que en estos últimos las
flavohemoglobinas sólo se encuentran en distintos grupos de microorganismos eucariotas,
en particular de hongos.
La reconstrucción filogenética generada en este trabajo, teniendo en cuenta las secuencias
de flavohemoglobinas representativas de distintos grupos taxonómicos, muestra en líneas
generales las relaciones observadas previamente por otros autores que han trabajado en
este grupo de proteínas en los organismos en los que estamos interesados (Turrión-Gómez
et al., 2010; te Biesebeke et al., 2010). Durante los últimos años el número de secuencias
relacionadas a disposición del investigador ha aumentado constantemente, permitiendo
llevar a cabo análisis cada vez más precisos y robustos. En nuestro estudio identificamos
en primer lugar un clado particular, el clado 4, en el que se agrupan la mayor parte de las
flavohemoglobinas de origen bacteriano. Más próximas a éstas que a las
flavohemoglobinas de otros hongos se encuentran las enzimas correspondientes descritas
en distintas especies de levaduras. La mayor parte de las secuencias de
flavohemoglobinas de hongos se integran en un agrupamiento específico de este grupo de
organismos (que se conforma con un sólido soporte estadístico). En esta agrupación, que
incluye los clados 1 y 2 mostrados en la Figura 18, se localizan las proteínas
flavohemoglobinas FHG1, FHG2 y FHG3 de FOP. Es necesario llamar la atención sobre el
hecho de que FHG4 no se incluye en esta agrupación. La proteína FHG4 se localiza en un
clado diferente, el clado 3, ubicado entre la agrupación de los clados 1 y 2 y el clado 4, y
de constitución muy particular, ya que incluye flavohemoglobinas presentes únicamente en
algunas especies del género Aspergillus y que están relacionadas con flavohemoglobinas
presentes en especies bacterianas muy concretas: Chromobacterium, Bortedella,
Burkholderia y Sinorhizobium. Las tres primeras, FHG1, FHG2 y FHG3, por un lado, y esta
Papel de las flavohemoglobinas en F. oxysporum
- 178 -
última, FHG4, por otro, representan probablemente tipos de flavohemoglobinas de
características diferentes y presentan relaciones evolutivas particulares y diferenciales.
El número de copias de los genes FHG en hongos varía de unas especies a otras. En
ascomicetes, se ha descrito una copia en especies como S. cerevisiae (Zhu et al., 1992) y
B. cinerea (Turrión-Gómez et al., 2010), dos en P. anserina, N. crassa y F. graminearum,
tres en C. albicans (Ullman et al., 2004) y en F. verticilloides y cuatro en F. oxysporum (este
trabajo). Y observamos que entre los basidiomicetes la representación de genes FHG es
más reducida: no han sido identificados en el genoma de especies representativas, como
Ustilago maydis y Coprinus cinereus y Laccaria bicolor, y sólo se ha identificado una copia
en el genoma de la especie C. neoformans (de Jesús-Berrios et al., 2003). En aquellas
especies en las que están presentes varias copias de los genes FHG, éstas no se agrupan
en una misma rama del cladograma. En estos casos, además, una copia dada presenta
generalmente más homología con copias de otras especies que con otras copias presentes
en la misma especie. Teniendo en cuenta estas consideraciones, el estudio de nuestro
cladograma y de las relaciones filogenéticas que en él se presentan, en particular en
relación con los clados 1 y 2, se pueden proponer varias hipótesis para explicar la evolución
de los genes FHG en los fusaria: (1) Sucesivas duplicaciones y divergencia a partir de un
gen ancestral, tal y como se ha propuesto para otros genes de F. oxysporum (Niño-
Sánchez et al., 2016); (2) adquisición por transferencia horizontal, hipótesis que parece la
más razonable para explicar la aparición de regiones específicas de F. oxysporum (Lineage
Specific, LS), las cuales provendrían de otras especies del género Fusarium. Es muy
posible que ambas explicaciones sean plausibles. Proponemos, que los genes FHG de F.
oxysporum, F. verticillioides y F. graminearum que aparecen próximos en el clado 2,
procederían de un ancestro común a las tres especies, si bien uno de los tres genes se
habría perdido en F. graminearum. Por el contrario, el gen de F. oxysporum que aparece
en el clado 3 (FHG4), próximo a genes de FHG de Aspergillus y Proteobacterias, podría
haberse originado por transferencia horizontal (explicado en el siguiente párrafo).
El análisis de los genes de FHGs en especies del género Aspergillus ofrece resultados
particulares y llamativos. En las distintas especies analizadas se han descrito dos copias
de genes FHG. Aquellas de las especies A. niger y A. clavatus, junto con una copia de las
especies A. fumigatus, A. kawachii, A. oryzae y A. terreus, se agrupan en el clado 1 junto
con la única copia del gen FHG descrita en B. cinerea y en S. sclerotiorum, y no aparece
ninguna copia en el clado 2. La segunda copia descrita en las especies A. terreus, A.
fumigatus, A. nidulans y A. oryzae aparece en el clado 3, que incluye como describíamos
anteriormente, genes correspondientes a ciertos grupos bacterianos. Llama
Discusión
- 179 -
poderosamente la atención la composición de este clado, que es el clado en el que se
localiza la cuarta copia de un gen FHG de FOP. Resulta tentador, a la vista de la
composición de este clado, proponer que hayan tenido lugar procesos de transferencia
horizontal de genes entre especies de bacterias y alguna de estas especies de hongos,
probablemente en tiempos muy tempranos, que permitan explicar la relación existente
entre las secuencias correspondientes.
Como reflexionan diversos autores (Levasseur et al., 2007; Te Bieseke et al., 2010) y como
recogen los libros de texto, los modelos de genética de poblaciones predicen que no es
posible mantener copias completamente redundantes de genes dados en el genoma
durante largos periodos de tiempo, ya que se irán acumulando mutaciones deletéreas y
tenderán a ser eliminados. Y a la inversa, la divergencia funcional de copias duplicadas
favorecerá el mantenimiento de las mismas. Acorde con este razonamiento, nuestra
hipótesis de partida para el presente trabajo fue desentrañar la posible especialización
funcional de los genes codificadores de proteínas FHG en F. oxysporum.
4.2 Análisis de expresión y funcional de los genes FHG
El hecho de encontrar varios genes codificadores de una determinada proteína en el
genoma de una especie dada y de que éstos sean mantenidos en el proceso evolutivo
permite considerar que las distintas copias sean funcionales. En el caso de FOP se han
identificado cuatro copias de los genes FHG. El objetivo fundamental de nuestro trabajo ha
sido determinar en qué procesos fisiológicos participan las enzimas codificadas por estos
genes en las estirpes FOP-SP1 y FOP-SP4. Estas dos estirpes de F. oxysporum son
patógenas de la judía común (por lo tanto forman parte de la forma especial phaseoli), y
son representativas de estirpes muy virulentas y poco virulentas de esta forma especial
(Alves-Santos et al, 2002b; de Vega-Bartol, 2011). En el curso de este trabjo se han
generado evidencias experimentales de tres tipos: las derivadas del análisis bioinformático
de los genes y de las proteínas codificadas, las derivadas del análisis de expresión de los
genes FHG en respuesta a distintos factores o estímulos, y las que se derivan de la
caracterización fenotípica de los mutantes en los genes FHG.
Es necesario en este momento hacer una consideración particular. Para llevar a cabo el
análisis de expresión y el análisis funcional descrito en primer lugar fue necesario
establecer las condiciones óptimas de cultivo con el fin de conocer el comportamiento de
las estirpes FOP-SP1 y FOP-SP4 en la germinación y desarrollo. Dado que inicialmente
Papel de las flavohemoglobinas en F. oxysporum
- 180 -
era importante conocer el efecto sobre la sensibilidad a NO y sobre desarrollo, decidimos
utilizar como medio de cultivo de referencia el medio en el que la germinación es más
eficiente. Por esta razón se llevó a cabo el análisis de expresión en medios suplementados
con amonio como fuente de nitrógeno. El amonio y/o la glutamina son las fuentes de
nitrógeno preferidas por los hongos y en su presencia es reprimida la expresión de genes
implicados en la asimilación del nitrato o nitrito (Marzluf, 1997; Berger et al., 2008). Por lo
tanto, nuestro análisis de expresión ha tenido en cuenta la condición de ausencia de fuente
de nitrógeno y la condición de presencia de amonio, pero no ha valorado el efecto de la
presencia de nitrato. Esta condición sí ha sido tenida en cuenta en el análisis fisiológico de
los mutantes.
4.3 Los genes FHG de FOP muestran patrones de expresión
diferenciales
La función principal de las proteínas de tipo FHG, ampliamente distribuidas en
microorganismos procariotas y eucariotas, es la detoxificación de NO la cual llevan a cabo
gracias a su actividad dioxigenasa dependiente de NADPH, FAD y O2. Esta actividad
dioxigenasa de la flavohemoglobina fue inicialmente descrita en la enzima hmp de E. coli
(Gardner et al., 1998; Hausladen et al., 1998) y posteriormente en las flavohemoglobinas
de Alcaligenes eutrophus (Ermler et al., 1995) y S. cerevisiae (Zhao et al., 1996). La
capacidad detoxificadora de la flavohemoglobina ha sido descrita desde entonces en
numerosas especies de bacterias (Crawford and Golberg, 1998b; Cramm et al., 1994;
Favey et al., 1995; LaCelle et al., 1996) y de hongos (Hromatka et al., 2005; Zhu et al.,
2006; Zhou et al., 2010; Turrión-Goméz et al. 2010; Lapp et al. 2014). En microorganismos
la exposición a NO determina un aumento de la expresión del gen o de los genes
codificadores de enzimas flavohemoglobinas, activando así un mecanismo inducible de
resistencia frente a condiciones de estrés nitrosativo (Crawford and Goldberg, 1998a;
Ullmann et al., 2004; Poole et al., 1996; Moore et al., 2004; Sarver and DeRisi, 2005;
Hromatka et al., 2005; Zhou et al., 2010; Turrión-Goméz et al. 2010; Lapp et al., 2014).
Los microorganismos eucariotas poseen frecuentemente varios genes codificadores de
enzimas flavohemoglobina en sus genomas. Dentro del grupo de los hongos encontramos
ejemplos de las distintas situaciones que cabe considerar. B. cinerea posee sólo uno,
Bcghg1 (Turrión-Gómez et al., 2010). Distintas especies del género Aspergillus poseen dos
genes FHG. En el caso de A. oryzae, Fhb1 codifica una flavohemoglobina de localización
citoplasmática implicada en la detoxificación de NO en el citoplasma y Fhb2 codifica una
Efectivamente, la aproximación farmacológica basada en la adición de NO exógeno ha sido
considerada en varios hongos a la hora de investigar la naturaleza de los procesos en los
Discusión
- 191 -
que el NO tiene un efecto. Los resultados obtenidos sugieren que el NO, también en este
grupo de organismos, regula procesos de diferenciación y/o desarrollo, aunque
dependiendo de los sistemas, en ocasiones parece ejercer una regulación en sentidos
opuestos. Así, se ha demostrado que en N. crassa la adición de NO induce la producción
de esporas asexuales (Ninnemann and Maier, 1996). En P. blakesleeanus regula el
desarrollo del esporangióforo (Maier et al., 2001). En Coniothyrium minitans induce la
conidiación (Gong et al., 2007). En A. nidulans, en uno de los estudios más completos al
respecto, se ha comprobado recientemente que la exposición a niveles elevados de NO
reduce la germinación de las esporas asexuales e induce el desarrollo de las estructuras
características de la fase de reproducción sexual, los cleistotecios (Marcos et al., 2016). En
F. oxysporum nosotros observamos una reducción en la eficiencia de la germinación de las
esporas, lo que supone que F. oxysporum y A. nidulans responden de manera similar a NO
exógeno. Dado que F. oxysporum no presenta reproducción sexual conocida, la
estimulación o no de la producción de estructuras sexuales no ha podido ser evaluada.
Es interesante destacar que la participación del NO en procesos de desarrollo y
morfogénesis se ha descrito en distintos sistemas, en particular en plantas (Beligni y
Lamattina, 2000; Pagnussat et al., 2002, 2003; He et al., 2004; Lombardo et al., 2006; Lee
et al., 2008; Fernández-Marcos et al., 2011; Giudice et al., 2011; Kwon et al., 2012). En el
contexto de nuestro trabajo resulta especialmente interesante el trabajo de Wang et al.
(2012) analizando procesos de desarrollo en la especie vegetal Camelia sinensis.
Basándose en los resultados obtenidos con un donador de NO estos autores sugieren que
el NO retrasa tanto la germinación del grano de polen como el desarrollo del tubo polínico
en C. sinensis. Esto supone que observamos efectos parecidos del NO en el desarrollo de
estructuras similares en especies pertenecientes a grupos taxonómicos muy diferentes,
plantas y hongos. Este paralelismo probablemente indique un grado de conservación de
los componentes implicados en la formación de dichas estructuras.
La aproximación farmacológica considerada en los estudios que hemos descrito utiliza
donadores de NO. En la mayor parte de los sistemas investigados la exposición a NO
exógeno determina la inducción de la expresión del gen, o de los genes, FHG, como parte
de un sistema de defensa inducible frente a estrés nitrosativo. Pero también en la mayor
parte de los sistemas vivos se ha demostrado la producción de NO endógeno, en particular
en especies de hongos en los que se ha demostrado esta producción tanto mediante la vía
oxidativa como mediante la vía reductiva (revisado en Roszer, 2012). El microorganismo
debe hacer frente también, indudablemente, al NO producido de forma endógena, haciendo
Papel de las flavohemoglobinas en F. oxysporum
- 192 -
uso de mecanismos de detoxificación en los que participan las enzimas FHG. El control de
los niveles de NO, resultantes de la producción endógena y de la detoxificación por enzimas
FHG, permite considerar este control como un mecanismo importante en la regulación de
procesos de diferenciación y desarrollo en este grupo de organismos. En este sentido
apuntan las observaciones llevadas a cabo en B. cinerea y en A. nidulans. En el primero
se ha demostrado la producción de NO por parte del hongo, aunque no ha sido posible
determinar el mecanismo mediante el cual éste es generado (Turrión-Gómez and Benito,
2011). En este sistema la expresión de Bcfhg1 está regulada durante el desarrollo,
mostrando un máximo de expresión coincidente con la germinación en ausencia de NO
exógeno (Turrión-Gómez et al., 2010). A. nidulans también produce NO, en este caso
mediante un sistema dependiente al menos parcialmente de la actividad NR, y esta
producción está regulada durante el desarrollo, aumentando en las fases tempranas de la
transición de crecimiento vegetativo a conidiación (Marcos et al., 2016). Es interesante
destacar que en A. nidulans niaD y fhbA muestran una inducción transitoria de la expresión
durante la inducción de la conidiación. Por su parte, fhbB muestra un patrón de expresión
diferente caracterizado por una expresión moderada en la fase de inicio de la conidiación
y una fuerte inducción en fases más tardías (Marcos et al., 2016). Todo ello sugiere que en
este grupo de organismos, representados por B. cinerea y A. nidulans, el NO endógeno
puede condicionar procesos tales como la conidiación o la germinación de las esporas y
que éstos son regulados mediante la modulación de los niveles de expresión de los genes
codificadores de enzimas FHG que detoxifican el NO. En los sistemas que poseen varios
genes FHG esto puede implicar una regulación diferencial de la expresión de unos y otros,
y la asunción de papeles o funciones específicas.
F. oxysporum posee, como se demuestra en este trabajo, cuatro genes codificadores de
FHG, los cuales muestran patrones de expresión diferentes. En el curso de este trabajo
hemos obtenido, utilizando procedimientos basados en microscopía de fluorescencia, unas
primeras evidencias de que F. oxysporum también produce NO (resultados no mostrados).
Todas estas observaciones contribuyen a dibujar el comportamiento de un hongo muy
“versátil” desde un punto de vista metabólico, que puede crecer tanto en amonio como en
nitrato en condiciones de aerobiosis, que también puede desarrollarse en condiciones de
denitrificación y crecer en anaerobiosis, y que puede resultar expuesto a posibles fuentes
internas y externas de NO. La disponibilidad de distintas enzimas FHG puede conferir
ventajas adaptativas en condiciones ambientales y de acceso a nutrientes muy diversas.
Discusión
- 193 -
4.7 El papel de los genes FHG durante la interacción con el huésped
El análisis de expresión de los genes FHG de F. oxysporum in planta demuestra que los
niveles de expresión de los genes FHG2, FHG3 y FHG4 son muy estables a lo largo del
proceso de infección en los tiempos considerados en nuestro estudio, desde 1 dpi hasta
14 dpi. Sólo en el caso del gen FHG1 se detectan variaciones significativas en el nivel de
expresión, que aumenta a lo largo del tiempo (del progreso de la infección), triplicando su
nivel de expresión a 7 dpi. La situación es muy similar en ambas cepas. Dado que éstas
difieren significativamente en la virulencia frente a plantas de judía, no parece que la
expresión de los genes FHG esté correlacionada con la viruencia exhibida durante la
colonización del huésped.
Como hemos comprobado en este trabajo, FHG1 responde a nivel transcripcional de forma
específica a la exposición a NO durante el cultivo en medios sintéticos. Teniendo en cuenta
esta observación es posible proponer que durante la interacción tanto de la cepa FOP-SP1
como de la cepa FOP-SP4 de F. oxysporum con su huésped, el patógeno debe resultar
expuesto a NO. Aunque no puede descartarse que el patógeno pueda producir por sí
mismo NO a niveles superiores en respuesta a algún tipo de estímulo de la planta durante
la interacción con ésta, parece razonable asumir que es la propia planta la que produce
NO como consecuencia de la activación de los mecanismos de defensa, en el curso de los
cuales se producen y liberan cantidades importantes de AOS y RNS (Delledone et al., 1998;
Delledone et al., 2001) a los que el hongo resultará expuesto. En esta situación, y dada la
toxicidad del NO sobre los microorganismos, la eliminación de componentes importantes
en la detoxificación de NO bien podría determinar una reducción de la capacidad de
desarrollo del patógeno in planta con la consiguiente reducción de virulencia. Esta situación
ha sido descrita en algunas interacciones planta-patógeno (Favey et al., 1995; Boccara et
al., 2005) y también en algunas interacciones en las que participan microorganismos
patógenos de la especie humana (Crawford and Goldberg, 1998; de Jesús-Barrios et al.,
2003; Ullmann et al., 2004). En F. oxysporum, el trabajo descrito en esta memoria
demuestra que la eliminación de FHG1, al igual que la de los otros tres genes FHG, no
determina reducción de la capacidad de infección de ninguna de las dos estirpes
analizadas. Tampoco se observa reducción de virulencia en los mutantes ΔFHG2 en los
que se ha silenciado el gen FHG1. Es posible concluir, por lo tanto, que los genes FHG no
son factores de patogenicidad en FOP. Esta es una situación descrita recientemente en
distintos sistemas en los que a priori se suponía un papel importante de las enzimas de
tipo flavohemoglobina como factores de patogenicidad. Por ejemplo, en B. cinerea los
Papel de las flavohemoglobinas en F. oxysporum
- 194 -
mutantes deficientes en el gen Bcfhg1, único gen codificador de una enzima
flavohemoglobina en este hongo necrótrofo, son hipersensibles a NO pero no muestran
una reducción en su patogenicidad ni virulencia (Turrión-Gómez et al., 2010). En A.
fumigatus los mutantes deficientes en los genes codificadores de enzimas
flavohemoglobina muestran la misma agresividad en sistemas modelo de infección
(ratones) que la cepa tipo silvestre (Lapp et al., 2014). Estas observaciones, que provienen
de sistemas muy diferentes, proporcionan evidencias que contradicen, al menos en parte,
la visión más clásica de la interacción, según la cual el microorganismo sufre una situación
de estrés nitrosativo, además de estrés oxidativo, durante su interacción con el huésped.
En el caso de FOP, el hecho de que los mutantes ΔFHG infecten a su huésped de igual
manera que las cepas tipo silvestre supone que, o bien los niveles de NO producidos
durante la interacción no son lo suficientemente elevados como para determinar una
limitación en su crecimiento in planta, o bien que F. oxysporum dispone de otros sistemas
de detoxificación de NO.
CONCLUSIONES
Conclusiones
- 195 -
5 CONCLUSIONES
- Las estirpes FOP-SP1 y FOP-SP4 de F. oxysporum f. sp. phaseoli poseen cuatro genes
codificadores de enzimas de tipo flavohemoglobina, FHG1, FHG2, FHG3 y FHG4. Son
cuatro genes de copia única y carecen de intrones. Las proteínas codificadas presentan la
organización en dominios funcionales característica de las enzimas de tipo
flavohemoglobina. El análisis bioinformático indica que FHG1, FHG2 y FHG3 son proteínas
de localización citoplasmática mientras que FHG4 es una proteína de localización
mitocondrial.
- El patrón de expresión de los genes FHG en relación con los factores “estado de
desarrollo”, “fuente de nitrógeno” y “exposición a NO”, es específico de cada gen y similar
en ambas cepas.
- FHG1, y en menor medida FHG2, incrementan sus niveles de expresión durante la
germinación de las esporas en respuesta a la exposición a NO. La expresión de FHG2 está
regulada, además, por la fuente de nitrógeno y el estado de desarrollo. La regulación de la
expresión de FHG3 es compleja, pero no resulta obvia, en general, una regulación por
exposición a NO. FHG4 no responde a nivel transcripcional a ninguno de los factores
evaluados en nuestro análisis.
- El nivel de expresión de FHG1 aumenta a medida que progresa la infección, tanto en la
interacción de plantas de judía FOP-SP1 como en la interacción con la estirpe FOP-SP4.
El nivel de expresión de los genes FHG2, FHG3 y FHG4 no varía a lo largo del proceso de
infección.
- Los mutantes simples deficientes en cada uno de los genes FHG de las estirpes FOP-SP1
y FOP-SP4 no muestran ninguna alteración evidente que afecte a su capacidad de
crecimiento en medios de cultivo artificiales o a su desarrollo.
- Las flavohemoglobinas FHG1 y FHG2, pero no las flavohemoglobinas FHG3 y FHG4, son
esenciales para proporcionar los niveles de resistencia frente a condiciones de estrés
nitrosativo mostrados por las estirpes silvestres FOP-SP1 y FOP-SP4. .
Papel de las flavohemoglobinas en F. oxysporum
- 196 -
- La exposición a NO en FOP-SP1 y en FOP-SP4 determina un retraso en la germinación
de los microconidios. La caracterización funcional de los mutantes ΔFHG demuestra que
FHG1 es la flavohemoglobina que desempeña una función principal en la eliminación del
NO que afecta a la germinaciónde las esporas.
- Las flavohemoglobinas codificadas por los genes analizados en este trabajo no constituyen
factores de virulencia en F. oxysporum.
BIBLIOGRAFÍA
Bibliografía
- 197 -
6 BIBLIOGRAFÍA
Adams, J. M., & Cory, S. 2002. Apoptosomes: engines for caspase activation. Current Opinion in Cell Biology 14(6):715-720.
Agrios, G.N. 1997. Plant pathology, 4th edn. San Diego, CA, USA: Academic Press Ltd. Akagi, Y., Akamatsu, H., Otani, H., & Kodama, M. 2009. Horizontal chromosome transfer,
a mechanism for the evolution and differentiation of a plant-pathogenic fungus. Eukaryotic Cell 8(11):1732-1738.
Alderton, W. K., Cooper, C. H., & Knowles, R. 2001. Nitric oxide synthases: structure,
function and inhibition. Biochemistry Journal 357:593-615. Aloj, B., Maziano, F., Zoina, A. & Noviello, C. 1983. La tracheofusariosi del fagiolo in Italia.
Informatore Fitopatologico 11: 63-66. Aloj, B., Marziano, F., Zoina, A. and Noviello, C. 1987. Osservazioni su una nuova razza
fisiologica de Fusarium oxysporum f.sp. phaseoli. "Annali" della FAcoltà di Scienze Agrarie dell´Università di Napoli in Portici. Serie IV, vol. XXI, pp. 51-55.
Alves-Santos, F. M., Benito, E. P., Eslava, A. P., & Díaz-Mínguez, J. 1999. Genetic Diversity
of Fusarium oxysporum Strains from Common Bean Fields in Spain. Applied and Environmental Microbiology 65(8):3335-3340.
Alves‐Santos, F. M., Cordeiro‐Rodrigues, L., Sayagués, J. M., Martín‐Domínguez, R.,
García‐Benavides, P., Crespo, M. C., & Eslava, A. P. 2002a. Pathogenicity and race characterization of Fusarium oxysporum f. sp. phaseoli isolates from Spain and Greece. Plant Pathology 51(5):605-611.
Alves-Santos, F. M., Ramos, B., García-Sánchez, M. A., Eslava, A. P., & Díaz-Mínguez, J.
M. 2002b. A DNA-based procedure for in planta detection of Fusarium oxysporum f. sp. phaseoli. Phytopathology 92(3):237-244.
Andrianopoulos, A., Kourambas, S., Sharp, J. A., Davis, M. A., & Hynes, M. J. 1998.
Characterization of the Aspergillus nidulans nmrA gene involved in nitrogen metabolite repression. Journal of Bacteriology 180(7):1973-1977.
Appel, D. J., & Gordon, T. R. 1995. Intraspecific variation within populations of Fusarium
oxysporum based on RFLP analysis of the intergenic spacer region of the rDNA. Experimental Mycology 19(2):120-128.
Arasimowicz‐Jelonek, M., & Floryszak‐Wieczorek, J. 2014. Nitric oxide: an effective weapon
of the plant or the pathogen. Molecular Plant Pathology 15(4):406-416. Baayen, R. P., O'Donnell, K., Bonants, P. J., Cigelnik, E., Kroon, L. P., Roebroeck, E. J., &
Waalwijk, C. 2000. Gene genealogies and AFLP analyses in the Fusarium oxysporum complex identify monophyletic and nonmonophyletic formae speciales causing wilt and rot disease. Phytopathology 90(8):891-900.
Papel de las flavohemoglobinas en F. oxysporum
- 198 -
Baidya, S., Cary, J. W., Grayburn, W. S., & Calvo, A. M. 2011. Role of nitric oxide and flavohemoglobin homolog genes in Aspergillus nidulans sexual development and mycotoxin production. Applied and Environmental Microbiology 77(15): 5524-5528.
Bang, I. S., Liu, L., Vazquez-Torres, A., Crouch, M. L., Stamler, J. S., & Fang, F. C. 2006.
Maintenance of nitric oxide and redox homeostasis by the Salmonella flavohemoglobin hmp gene. Journal of Biological Chemistry 281(38): 28039-28047.
Beckman, C.H. 1987. The nature of wilt diseases of plants. American Phytopathological
Society. EEUU. Belenghi, B., Romero-Puertas, M. C., Vercammen, D., Brackenier, A., Inzé, D., Delledonne,
M., & Van Breusegem, F. 2007. Metacaspase activity of Arabidopsis thaliana is regulated by S-nitrosylation of a critical cysteine residue. Journal of Biological Chemistry 282(2): 1352-1358.
Beligni, M. V. & Lamattina, L. 2000. Nitric oxide stimulates seed germination and de-
etiolation, and inhibits hypocotyl elongation, three light-inducible responses in plants. Planta 210(2): 215-21.
Benlloch, M, Cañizo JD. 1926. La enfermedad de las alubias en barco de ávila. Boletín de
la Estación de Patología Vegetal 1: 2-7.
Berger, H., Basheer, A., Böck, S., Reyes‐Dominguez, Y., Dalik, T., Altmann, F., & Strauss, J. 2008. Dissecting individual steps of nitrogen transcription factor cooperation in the Aspergillus nidulans nitrate cluster. Molecular Microbiology 69(6): 1385-1398.
Bernreiter, A., Ramón, A., Fernández-Martínez, J., Berger, H., Araújo-Bazan, L., Espeso,
E. A., & Strauss, J. 2007. Nuclear export of the transcription factor NirA is a regulatory checkpoint for nitrate induction in Aspergillus nidulans. Molecular and Cellular Biology 27(3): 791-802.
Besson-Bard, A., Pugin, A., & Wendehenne, D. 2008. New insights into nitric oxide signaling
in plants. Annual Review Plant Biology 59: 21-39. Bethke, P. C., Gubler, F., Jacobsen, J. V., & Jones, R. L. 2004. Dormancy of Arabidopsis
seeds and barley grains can be broken by nitric oxide. Plant Journal 219(5): 847-855.
Boccara, M., Mills, C.E., Zeier, J., Anzi, C., Lamb, C., Poole, R.K., & Delledonne, M. 2005.
Flavohaemoglobin HmpX from Erwinia chrysanthemi confers nitrosative stress tolerance and affects the plant hypersensitive reaction by intercepting nitric oxide produced by the host. The Plant Journal 43:226-37.
Bonfoco, E., Krainc, D., Ankarcrona, M., Nicotera, P., & Lipton, S.A. 1995. Apoptosis and
necrosis - 2 distinct events induced, respectively, by mild and intense insults with N-methyl-D-aspartate or nitric oxide superoxide in cortical cell cultures. Proceedings of the National Academy of Science of the. U. S. A. 92:7162-7166.
Booth, C., Moss, M. O., & Smith, J. E. 1984. The Fusarium problem: historical, economic
and taxonomic aspects. In The Applied Mycology of Fusarium. Symposium of the British Mycological Society held at Queen Mary College, London, September 1982. (pp. 1-13). Cambridge University Press.
Bibliografía
- 199 -
Boveris, A., Costa, L. E., Poderoso, J. J., Carreras, M. C., & Cadenas, E. 2000. Regulation of mitochondrial respiration by oxygen and nitric oxide. Annals of the New York Academy of Sciences 899(1): 121-135.
Bright, J., Desikan, R., Hancock, J. T., Weir, I. S., & Neill, S. J. 2006. ABA‐induced NO
generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. The Plant Journal 45(1): 113-122.
Buisson, N., & Labbe-Bois, R. 1998. Flavohemoglobin Expression and Function in
Saccharomyces cerevisiae NO relationship with respiration and complex response to oxidative stress. Journal of Biological Chemistry 273(16): 9527-9533.
Burnett, J. H. 1984. Aspects of Fusarium genetics. The Applied Mycology of Fusarium In
pp. 39-69.Held at Queen Mary Collage of London. Cadenas, E., & Davies, K. J. 2000. Mitochondrial free radical generation, oxidative stress,
and aging. Free Radical Biology and Medicine 29(3): 222-230. Cañero, D. C., & Roncero, M. I. G. 2008. Functional analyses of laccase genes from
Fusarium oxysporum. Phytopathology 98(5): 509-518. Cartharius, K., Frech, K., Grote, K., Klocke, B., Haltmeier, M., Klingenhoff, A., & Werner, T.
2005. MatInspector and beyond: promoter analysis based on transcription factor binding sites. Bioinformatics 21(13): 2933-2942.
Cassanova, N., O'Brien, K. M., Stahl, B. T., McClure, T., & Poyton, R. O. 2005. Yeast
Flavohemoglobin, a Nitric Oxide Oxidoreductase, Is Located in Both the Cytosol and the Mitochondrial Matrix effects of respiration, anoxia, and the mitochondrial genome on its intracellular level and distribution. Journal of Biological Chemistry 280(9): 7645-7653.
Cevahir, G., Aytamka, E., & Erol, Ç. 2007. The role of nitric oxide in plants. Biotechnology
& Biotechnological Equipment 21(1): 13-17. Coppin, E., Debuchy, R., Arnaise, S., & Picard, M. 1997. Mating types and sexual
development in filamentous ascomycetes. Microbiology and Molecular Biology Reviews 61(4): 411-428.
Corker, H., & Poole, R. K. 2003. Nitric oxide formation by Escherichia coli dependence on
nitrite reductase, the no-sensing regulator Fnr, and flavohemoglobin Hmp. Journal of Biological Chemistry 278(34): 31584-31592.
Corpas, F. J., Barroso, J. B., & del Rıo, L. A. 2001. Peroxisomes as a source of reactive
oxygen species and nitric oxide signal molecules in plant cells. Trends in Plant Science 6(4): 145-150.
Courtois, C., Besson, A., Dahan, J., Bourque, S., Dobrowolska, G., Pugin, A., &
Wendehenne, D. 2008. Nitric oxide signalling in plants: interplays with Ca2+ and protein kinases. Journal of Experimental Botany 59(2): 155-163.
Cove, D. J. 1979. Genetic studies of nitrate assimilation in Aspergillus nidulans. Biological
Reviews 54(3): 291-327.
Papel de las flavohemoglobinas en F. oxysporum
- 200 -
Chichkova, N.V., Kim, S.H., Titova, E.S., Kalkum, M., Morozov, V.S., Rubtsov, Y.P., Kalinina, N.O., Taliansky, M.E., & Vartapetian, A.B. 2004. A plant caspase-like protease activated during the hypersensitive response. Plant Cell 16: 157-171.
Cramm, R., Siddiqui, R. A., & Friedrich, B. 1994. Primary sequence and evidence for a
physiological function of the flavohemoprotein of Alcaligenes eutrophus. Journal of Biological Chemistry 269(10): 7349-7354.
Crawford, M. J., & Goldberg, D. E. 1998b. Role for the Salmonella flavohemoglobin in
protection from nitric oxide. Journal of Biological Chemistry 273(20): 12543-12547. Crawford, M. J., & Goldberg, D. E. 1998a. Regulation of the Salmonella typhimurium
Flavohemoglobin Gene a new pathway for bacterial gene expression in response to nitric oxide. Journal of Biological Chemistry 273(51): 34028-34032.
Cueto, M., Hernández-Perera, O., Martín, R., Bentura, M. L., Rodrigo, J., Lamas, S., &
Golvano, M. P. 1996. Presence of nitric oxide synthase activity in roots and nodules of Lupinus albus. FEBS Letters 398(2): 159-164.
Czymmek, K. J., Fogg, M., Powell, D. H., Sweigard, J., Park, S. Y., & Kang, S. 2007. In vivo
time-lapse documentation using confocal and multi-photon microscopy reveals the mechanisms of invasion into the Arabidopsis root vascular system by Fusarium oxysporum. Fungal Genetics and Biology 44(10): 1011-1023.
Daboussi, M. J., & Capy, P. 2003. Transposable elements in filamentous fungi. Annual
Reviews in Microbiology 57(1): 275-299. de Jesús-Berrıos, M., Liu, L., Nussbaum, J. C., Cox, G. M., Stamler, J. S., & Heitman, J.
2003. Enzymes that counteract nitrosative stress promote fungal virulence. Current Biology 13(22): 1963-1968.
de Vega-Bartol, J. J., Tello, V., Niño, J., Casado, V., & Díaz-Mínguez, J. M. 2013.
Quantitative PCR Analysis of Double-Stranded RNA-Mediated Gene Silencing in Fungi. In Laboratory Protocols in Fungal Biology (pp. 279-287). Springer New York
de Vega-Bartol, J. J., Martín-Dominguez, R., Ramos, B., García-Sánchez, M. A., & Díaz-
Mínguez, J. M. 2011. New virulence groups in Fusarium oxysporum f. sp. phaseoli: the expression of the gene coding for the transcription factor ftf1 correlates with virulence. Phytopathology 101(4): 470-479.
Deighton, F. C., Stevenson, J. A., & Cummins, G. B. 1962. Formae speciales and the
code. International Association for Plant Taxonomy 11(3): 70-71. Delledonne, M., Xia, Y., Dixon, R. A., Lamb, C. 1998. Nitric oxide functions as a signal in
plant disease resistance. Nature 394(6693): 585-8. Delledonne, M., Zeier, J., Marocco, A., Lamb, C. 2001. Signal interactions between nitric
oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proceedings of the National Academy of Sciences of the U. S. A. 98(23): 13454-9.
Bibliografía
- 201 -
Di Pietro, A. D., Madrid, M. P., Caracuel, Z., Delgado‐Jarana, J., & Roncero, M. I. G. 2003. Fusarium oxysporum: exploring the molecular arsenal of a vascular wilt fungus. Molecular Plant Pathology 4(5): 315-325.
Di Pietro, A., García‐Maceira, F. I., Meglecz, E., & Roncero, M. I. G. 2001. A MAP kinase
of the vascular wilt fungus Fusarium oxysporum is essential for root penetration and pathogenesis. Molecular Microbiology 39(5): 1140-1152.
Díaz-Mínguez, J. M., Alves-Santos, F. M., Benito, E. P., & Eslava, A. P. 1996. Fusarium wilt
of common bean in the Castilla y Leon region of Spain. Plant Disease 80(5): 600. Dunn-Coleman, N. S., Tomsett, A. B., & Garrett, R. H. 1981. The regulation of nitrate
assimilation in Neurospora crassa: biochemical analysis of the nmr-1 mutants. Molecular and General Genetics MGG 182(2): 234-239.
Durner, J., & Klessig, D. F. 1999. Nitric oxide as a signal in plants. Current Opinion in Plant
Biology 2(5): 369-374. Ermler, U., Siddiqui, R. A., Cramm, R., & Friedrich, B. 1995. Crystal structure of the
flavohemoglobin from Alcaligenes eutrophus at 1.75 Aº resolution. The EMBO Journal 14(24): 6067.
Favey, S., Labesse, G., Vouille, V., & Boccara, M. 1995. Flavohaemoglobin HmpX: a new
pathogenicity determinant in Erwinia chrysanthemi strain 3937. Microbiology 141(4): 863-871.
Feechan, A., Kwon, E., Yun, B. W., Wang, Y., Pallas, J. A., & Loake, G. J. 2005. A central
role for S-nitrosothiols in plant disease resistance. Proceedings of the National Academy of Sciences of the U. S. A. 102(22): 8054-8059.
Fernández-Marcos, M., Sanz, L., Lewis, D. R., Muday, G. K., & Lorenzo, O. 2011. Nitric
oxide causes root apical meristem defects and growth inhibition while reducing PIN-FORMED 1 (PIN1)-dependent acropetal auxin transport. Proceedings of the National Academy of Sciences of the U. S. A. 108(45): 18506-18511.
Fitzhugh, A. L., & Keefer, L. K. 2000. Diazeniumdiolates: Pro-and antioxidant applications
of the “NONOates. Free Radical Biology and Medicine 28(10): 1463-1469. Frandsen, R. J., Andersson, J. A., Kristensen, M. B., & Giese, H. 2008. Efficient four
fragment cloning for the construction of vectors for targeted gene replacement in filamentous fungi. BMC Molecular Biology 9(1): 1.
Freschi, L., Rodrigues, M. A., Domingues, D. S., Purgatto, E., Van Sluys, M. A., Magalhaes,
J. R., & Mercier, H. 2010. Nitric oxide mediates the hormonal control of Crassulacean acid metabolism expression in young pineapple plants. Plant Physiology 152(4): 1971-1985.
García-Martínez, J., Ádám, A. L., & Avalos, J. 2012. Adenylyl cyclase plays a regulatory
role in development, stress resistance and secondary metabolism in Fusarium fujikuroi. PloS One 7(1): e28849.
Papel de las flavohemoglobinas en F. oxysporum
- 202 -
Gardner, P.R., Gardner, A.M., Martin, L.A., & Salzman, A.L. 1998. Nitric oxide dioxygenase: an enzymic function for flavohemoglobin. Proceedings of the National Academy of Sciences of the U. S. A. 95:10378-83.
Gardner, P. R. 2005. Nitric oxide dioxygenase function and mechanism of flavohemoglobin,
hemoglobin, myoglobin and their associated reductases. Journal of Inorganic Biochemistry 99(1): 247-266.
Geiser, D. M., Jiménez-Gasco, M. M., Kang, S., Makalowska, I., Veeraraghavan, N., Ward,
T. J., Zhang, N., Kuldau, A. & O´Donell, K. 2004. Fusarium-ID v. 1.0: A DNA sequence database for identifying Fusarium. European Journal of Plant Pathology 110: 473-479.
Gepts, P., & Bliss, F. A. 1988. Dissemination pathways of common bean (Phaseolus
vulgaris, Fabaceae) deduced from phaseolin electrophoretic variability. II. Europe and Africa. Economic Botany 42(1): 86-104.
Gerlach, W. & Niremberg, H. 1982. The genus Fusarium, a pictorical atlas. Mitteilungen aus
der Bioloischen Bundesansalt für Land- und Forstwirschaft. 406. Gilberthorpe, N. J., & Poole, R. K. 2008. Nitric Oxide Homeostasis in Salmonella
typhimurium roles of respiratory nitrate reductase and flavohemoglobin. Journal of Biological Chemistry 283(17): 11146-11154.
Godard, P., Urrestarazu, A., Vissers, S., Kontos, K., Bontempi, G., van Helden, J., & André,
B. 2007. Effect of 21 different nitrogen sources on global gene expression in the yeast Saccharomyces cerevisiae. Molecular and Cellular Biology 27(8): 3065-3086.
Gong, X., Fu, Y., Jiang, D., Li, G., Yi, X., & Peng, Y. 2007. L-arginine is essential for
conidiation in the filamentous fungus Coniothyrium minitans. Fungal Genetics and Biology 44(12): 1368-1379.
Gordon, T. R., & Martyn, R. D. 1997. The evolutionary biology of Fusarium
oxysporum. Annual Review of Phytopathology 35(1): 111-128. Glass, N. L., Jacobson, D. J., & Shiu, P. K. 2000. The genetics of hyphal fusion and
vegetative incompatibility in filamentous ascomycete fungi. Annual review of genetics 34(1): 165-186.
Guadet, J., Julien, J., Lafay, J. F., & Brygoo, Y. 1989. Phylogeny of some Fusarium species,
as determined by large-subunit rRNA sequence comparison. Molecular Biology and Evolution 6(3): 227-242.
Hagedorn, D. J. 1991. Compendium of bean diseases. Hall, R. (Ed). APS Press, pp. 20. Han, Y., Liu, X., Benny, U., Kistler, H. C., & VanEtten, H. D. 2001. Genes determining
pathogenicity to pea is clustered on a supernumerary chromosome in the fungal plant pathogen Nectria haematococca. The Plant Journal 25(3): 305-314.
Hausladen, A., Gow, A. J., & Stamler, J. S. 1998. Nitrosative stress: metabolic pathway
involving the flavohemoglobin. Proceedings of the National Academy of Sciences of the U.S.A. 95(24): 14100-14105.
Bibliografía
- 203 -
He, Y., Tang, R. H., Hao, Y., Stevens, R. D., Cook, C. W., Ahn, S. M., Jing, L., Yang, Z., Chen, L., Guo, F., Fiorani, F., Jackson, R. B., Crawford, N. M., & Pei, Z. M. 2004. Nitric oxide represses the Arabidopsis floral transition. Science 305(5692): 1968-71.
Heidmann, S., Obermaier, B., Vogel, K., & Domdey, H. 1992. Identification of pre-mRNA
polyadenylation sites in Saccharomyces cerevisiae. Journal of Biological Chemistry 12: 4215-4229.
Hernández-Urzúa, E., Mills, C. E., White, G. P., Contreras-Zentella, M. L., Escamilla, E.,
Vasudevan, S. G., & Poole, R. K. 2003. Flavohemoglobin Hmp, but not its individual domains, confers protection from respiratory inhibition by nitric oxide in Escherichia coli. Journal of Biological Chemistry 278(37): 34975-34982.
Hess, D. T., Matsumoto, A., Kim, S. O., Marshall, H. E., & Stamler, J. S. 2005. Protein S-
Hromatka, B.S., Noble, S.M., y Johnson, A.D. 2005. Transcriptional response of Candida
albicans to nitric oxide and the role of the YHB1 gene in nitrosative stress and virulence. Journal of Biological Chemistry 16: 4814-4826.
Houterman, P. M., Ma, L., Van Ooijen, G., De Vroomen, M. J., Cornelissen, B. J., Takken,
F. L., & Rep, M. 2009. The effector protein Avr2 of the xylem‐colonizing fungus
Fusarium oxysporum activates the tomato resistance protein I‐2 intracellularly. The Plant Journal 58(6): 970-978.
Ignarro, L. J. 1990. Biosynthesis and metabolism of endothelium-derived nitric
oxide. Annual Review of Pharmacology and Toxicology 30(1): 535-560. Napoli, C., & Ignarro, L. J. 2009. Nitric oxide and pathogenic mechanisms involved in the
development of vascular diseases. Archives of Pharmacal Research 32(8): 1103-1108.
Iijima, M., Shimizu, H., Tanaka, Y., & Urushihara, H. 2000. Identification and
characterization of two flavohemoglobin genes in Dictyostelium discoideum. Cell Structure and Function 25(1): 47-55.
Ilari, A., Bonamore, A., Farina, A., Johnson, K. A., & Boffi, A. 2002. The X-ray Structure of
Ferric Escherichia coli Flavohemoglobin Reveals an Unexpected Geometry of the Distal Heme Pocket. Journal of Biological Chemistry 277(26): 23725-23732.
Imazaki, I., Kurahashi, M., Iida, Y., & Tsuge, T. 2007. Fow2, a Zn (II) 2Cys6‐type
transcription regulator, controls plant infection of the vascular wilt fungus Fusarium oxysporum. Molecular Microbiology 63(3): 737-753.
Jasid, S., Simontacchi, M., Bartoli, C. G., & Puntarulo, S. 2006. Chloroplasts as a nitric
oxide cellular source. Effect of reactive nitrogen species on chloroplastic lipids and proteins. Plant Physiology 142(3): 1246-1255.
Jonkers, W., Xayamongkhon, H., Haas, M., Olivain, C., Does, H. C., Broz, K., & Kistler, H.
C. 2014. EBR1 genomic expansion and its role in virulence of Fusarium species. Environmental Microbiology 16(7): 1982-2003.
Papel de las flavohemoglobinas en F. oxysporum
- 204 -
Kaiser, W. M., Gupta, K. J., & Planchet, E. 2006. Higher plant mitochondria as a source for NO. In Nitric Oxide in Plant Growth, Development and Stress Physiology (pp. 1-14). Springer Berlin Heidelberg.
Kanadia, R. N., Kuo, W. N., Mcnabb, M., & Botchway, A. 1998. Constitutive nitric oxide
synthase in Saccharomyces cerevisiae. IUBMB Life 45(6): 1081-1087. Kazan, K., Gardiner, D. M., & Manners, J. M. 2012. On the trail of a cereal killer: recent
advances in Fusarium graminearum pathogenomics and host resistance. Molecular Plant Pathology 13(4): 399-413.
Kendrick, J. B. & Snyder, W. C. 1942. Fusarium yellows of beans. Phytopathology 32: 1010-
1014. Kerényi, Z., Moretti, A., Waalwijk, C., Oláh, B., & Hornok, L. 2004. Mating type sequences
in asexually reproducing Fusarium species. Applied and Environmental Microbiology 70(8): 4419-4423.
Keszthelyi, A., Jeney, A., Kerényi, Z., Mendes, O., Waalwijk, C., & Hornok, L. 2007. Tagging
target genes of the MAT1-2-1 transcription factor in Fusarium verticillioides (Gibberella fujikuroi MP-A). Antonie Van Leeuwenhoek, 91(4): 373-391.
Kistler, H. C. 2001. Evolution of host specificity in Fusarium oxysporum. In: Summerell BA,
Leslie JF, Backhouse WL, Burgess LW, editors. Fusarium: Paul E Nelson Memorial Symposium. St. Paul: APS Press. pp. 70–82.
Kistler, H. C., Benny, U., Boehm, E. W. A., & Katan, T. 1995. Genetic duplication in
Fusarium oxysporum. Current Genetics 28(2): 173-176. Knogge, W. 1996. Molecular basis of specificity in host/fungus interactions. European
Journal of Plant Pathology 102(9): 807-816. Kobayashi, M., & Shoun, H. 1995. The copper-containing dissimilatory nitrite reductase
involved in the denitrifying system of the fungus Fusarium oxysporum. Journal of Biological Chemistry 270(8): 4146-4151.
Kudla, B., Caddick, M. X., Langdon, T., Martinez-Rossi, N. M., Bennett, C. F., Sibley, S., &
Arst Jr, H. N. 1990. The regulatory gene areA mediating nitrogen metabolite repression in Aspergillus nidulans. Mutations affecting specificity of gene activation alter a loop residue of a putative zinc finger. The EMBO Journal 9(5): 1355.
Kuo, P. C., & Schroeder, R. A. 1995. The emerging multifaceted roles of nitric oxide. Annals
of Surgery 221(3): 220. Kwon, E., Feechan, A., Yun, B. W., Hwang, B. H., Pallas, J. A., Kang, J. G., & Loake, G. J.
2012. AtGSNOR1 function is required for multiple developmental programs in Arabidopsis. Plant Journal 236(3): 887-900.
LaCelle, M., Kumano, M., Kurita, K., Yamane, K., Zuber, P., & Nakano, M. M. 1996. Oxygen-
controlled regulation of the flavohemoglobin gene in Bacillus subtilis. Journal of Bacteriology 178(13): 3803-3808.
Laday, M., & Szecsi, A. 2002. Identification of Fusarium species by isozyme analysis. Acta
Microbiologica et Immunologica Hungarica 49(2-3): 321-330.
Bibliografía
- 205 -
Lapp, K., Vödisch, M., Kroll, K., Strassburger, M., Kniemeyer, O., Heinekamp, T., & Brakhage, A. A. 2014. Characterization of the Aspergillus fumigatus detoxification systems for reactive nitrogen intermediates and their impact on virulence. Frontiers in Microbiology 5:469.
Lazo, G.R., Stein, P.A. & Ludwig, R.A. 1991. A DNA transformation-competent Arabidopsis
genomic library in Agrobacterium. Biotechnology 9: 963-7. Lee, J., Myong, K., Kim, J. E., Kim, H. K., Yun, S. H., & Lee, Y. W. 2012. FgVelB globally
regulates sexual reproduction, mycotoxin production and pathogenicity in the cereal pathogen Fusarium graminearum. Microbiology 158(7): 1723-1733.
Lee, U., Wie, C., Fernandez, B. O., Feelisch, M., & Vierling, E. 2008. Modulation of
nitrosative stress by S-nitrosoglutathione reductase is critical for thermotolerance and plant growth in Arabidopsis. Plant Cell 20(3): 786-802.
Leslie, J. F. 1993. Fungal vegetative compatibility. Annual Review of Phytopathology
31:127-150. Leslie, J. F., & Summerell, B. A. 2006. The Fusarium laboratory manual (Vol. 2, No. 10).
Ames, IA, USA: Blackwell Pub. Levasseur, A., Orlando, L., Bailly, X., Milinkovitch, M. C., Danchin, E. G., & Pontarotti, P.
2007. Conceptual bases for quantifying the role of the environment on gene evolution: the participation of positive selection and neutral evolution. Biological Reviews 82(4): 551-572.
Li, B., Fu, Y., Jiang, D., Xie, J., Cheng, J., Li, G., & Yi, X. 2010. Cyclic GMP as a second
messenger in the nitric oxide-mediated conidiation of the mycoparasite Coniothyrium minitans. Applied and Environmental Microbiology 76(9): 2830-2836.
Lievens, B., Brouwer, M., Vanachter, A. C., Levesque, C. A., Cammue, B. P. & Thomma,
B. P. 2003. Design and development of a DNA array for rapid detection and identification of multiple tomato vascular wilt pathogens. FEMS Microbiology Letters 223: 113-122.
Liu, J., & Zuber, P. 2000. The ClpX protein of Bacillus subtilis indirectly influences RNA
Lombardo, Graziano, M., Polacco, J. C., & Lamattina, L. 2006. Nitric oxide functions as a
positive regulator of root hair development. Plant Signal Behavior 1(1): 28-33. López-Berges, M. S., Rispail, N., Prados-Rosales, R. C., & Di Pietro, A. 2010. A nitrogen
response pathway regulates virulence functions in Fusarium oxysporum via the protein kinase TOR and the bZIP protein MeaB. The Plant Cell 22(7): 2459-2475.
Ma, L. J., Geiser, D. M., Proctor, R. H., Rooney, A. P., O'Donnell, K., Trail, F. & Kazan, K. 2013. Fusarium pathogenomics. Annual Review of Microbiology 67: 399-416.
Ma, L. J., Van Der Does, H. C., Borkovich, K. A., Coleman, J. J., Daboussi, M. J., Di Pietro,
A., & Houterman, P. M. 2010. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 464(7287): 367-373.
Papel de las flavohemoglobinas en F. oxysporum
- 206 -
MacMicking, J. D., North, R. J., La Course, R., Mudgett, J. S., Shah, S. K., & Nathan, C. F. 1997. Identification of nitric oxide synthase as a protective locus against tuberculosis. Proceedings of the National Academy of Sciences of the U.S.A. 94(10): 5243-5248.
Magasanik, B. 1992. Regulation of Nitrogen Utilization. Cold Spring Harbor Monograph
Archive 21: 283-317. Magasanik, B. 2005. The transduction of the nitrogen regulation signal in Saccharomyces
cerevisiae. Proceedings of the National Academy of Sciences of the U.S.A. 102(46): 16537-16538.
Magasanik, B., & Kaiser, C. A. 2002. Nitrogen regulation in Saccharomyces cerevisiae.
Gene 290(1-2): 1-18. Maier, J., Hecker, R., Rockel, P., & Ninnemann, H. 2001. Role of nitric oxide synthase in
the light-induced development of sporangiophores in Phycomyces blakesleeanus. Plant Physiology 126: 1323-1330.
Mannick, J. B., & Schonhoff, C. M. 2002. Nitrosylation: the next phosphorylation. Archives
of Biochemistry and Biophysics 408(1): 1-6. Marchler-Bauer, A. 2015, CDD: NCBI conserved domain database. Nucleic Acids Research
43:222-6. Martín-Urdíroz, M., Roncero, M. I. G., González-Reyes, J. A., & Ruiz-Roldán, C. 2008.
ChsVb, a class VII chitin synthase involved in septation, is critical for pathogenicity in Fusarium oxysporum. Eukaryotic cell 7(1): 112-121.
Marcos, A. T., Ramos, M. S., Marcos, J. F., Carmona, L., Strauss, J., & Cánovas, D. 2016.
Nitric oxide synthesis by nitrate reductase is regulate during development in Aspergillus.Mole cular Microbiology 99(1): 15-33.
Martin, S. J., & Green, D. R. 1995. Protease activation during apoptosis: death by a
thousand cuts. Cell 82(3): 349-352. Marzluf, G. A. 1997. Genetic regulation of nitrogen metabolism in the fungi. Microbiology
and Molecular Biology Reviews 61(1): 17-32. Membrillo-Hernández, J., Coopamah, M. D., Anjum, M. F., Stevanin, T. M., Kelly, A.,
Hughes, M. N., & Poole, R. K. 1999. The flavohemoglobin of Escherichia coli confers resistance to a nitrosating agent, a “nitric oxide releaser,” and paraquat and is essential for transcriptional responses to oxidative stress. Journal of Biological Chemistry 274(2): 748-754.
Meyer, C., Lea, U. S., Provan, F., Kaiser, W. M., & Lillo, C. 2005. Is nitrate reductase a
major player in the plant NO (nitric oxide) game. Photosynthesis Research 83(2): 181-189.
Michaelis, T. E. 1991. Compendium of Bean Diseases. APS Press. Michielse, C. B., & Rep, M. 2009. Pathogen profile update: Fusarium oxysporum. Molecular
Plant Pathology 10(3): 311-324.
Bibliografía
- 207 -
Minehart, P. L., & Magasanik, B. O. R. I. S. 1991. Sequence and expression of GLN3, a positive nitrogen regulatory gene of Saccharomyces cerevisiae encoding a protein with a putative zinc finger DNA-binding domain. Molecular and Cellular Biology 11(12): 6216-6228.
Mirete, S., Patino, B., Vázquez, C., Jiménez, M., Hinojo, M. J., Soldevilla, C., & González-
Jaén, M. T. 2003. Fumonisin production by Gibberella fujikuroi strains from Pinus species. International Journal of Food Microbiology 89(2): 213-221.
Moncada, S., Rees, D. D., Schulz, R., & Palmer, R. M. 1991. Development and mechanism
of a specific supersensitivity to nitrovasodilators after inhibition of vascular nitric oxide synthesis in vivo. Proceedings of the National Academy of Sciences of the U.S.A 88(6): 2166-2170.
Moore, C.M., Nakano, M.M., Wang, T., Ye, R.W., & Helmann, J.D. 2004. Response of
Bacillus subtilis to nitric oxide and the nitrosating agent sodium nitroprusside. Journal Bacteriology 186: 4655-4664.
Booth, C., Moss, M. O., & Smith, J. E. 1984. The Fusarium problem: historical, economic
and taxonomic aspects. In The applied mycology of Fusarium. Symposium of the British Mycological Society held at Queen Mary College, London, September 1982. (pp. 1-13). Cambridge University Press.
Mullins, E. D., Chen, X., Romaine, P., Raina, R., Geiser, D. M., & Kang, S. 2001.
Agrobacterium-mediated transformation of Fusarium oxysporum: an efficient tool for insertional mutagenesis and gene transfer. Phytopathology 91(2): 173-180.
Mur, L. A., Kenton, P., Lloyd, A. J., Ougham, H., & Prats, E. 2008. The hypersensitive
response; the centenary is upon us but how much do we know? Journal of Experimental Botany 59(3): 501-520.
Nakanishi, Y., Zhou, S., Kim, S. W., Fushinobu, S., Maruyama, J. I., Kitamoto, K., & Shoun,
H. 2010. A eukaryotic copper-containing nitrite reductase derived from a NirK homolog gene of Aspergillus oryzae. Bioscience, Biotechnology and Biochemistry 74(5): 984-991.
Namiki, F., Matsunaga, M., Okuda, M., Inoue, I., Nishi, K., Fujita, Y., & Tsuge, T. 2001.
Mutation of an arginine biosynthesis gene causes reduced pathogenicity in Fusarium oxysporum f. sp. melonis. Molecular Plant-Microbe Interactions 14(4): 580-584.
Napoli, C., & Ignarro, L. J. 2009. Nitric oxide and pathogenic mechanisms involved in the
development of vascular diseases. Archives of Pharmacal Research 32(8): 1103-1108.
Narendja, F., Goller, S. P., Wolschek, M., & Strauss, J. 2002. Nitrate and the GATA factor AreA are necessary for in vivo binding of NirA, the pathway‐specific transcriptional activator of Aspergillus nidulans. Molecular Microbiology 44(2): 573-583.
Neill, S. J., Desikan, R., & Hancock, J. T. 2003. Nitric oxide signalling in plants. New
Phytologist 159(1): 11-35. Nelson, P. E. 1981. Life cylcle ande epidemiology of Fusarium oxysporum. In vascular wilt,
Nelson, P. E., Desjardins, A. E., & Plattner, R. D. 1996. Fumonisins, mycotoxins produced by Fusarium species: biology, chemistry, and significance. Annual review of phytopathology 31(1): 233-252.
Nelson, P. E., Dignani, M. C., & Anaissie, E. J. 1994. Taxonomy, biology, and clinical
aspects of Fusarium species. Clinical Microbiology Reviews 7(4): 479-504. Nelson, P. E., Toussoun, T. A., & Marasas, W. F. O. 1983. Fusarium species: an illustrated
manual for identification, Pennsylvania State University Press. Ninnemann, H. & Maier, J. 1996. "Indications for the occurrence of nitric oxide synthases in
fungi and plants and the involvement in photoconidiation of Neurospora crassa." Photochemmistry and Photobiology 64(2): 393-8.
Niño‐Sánchez, J., Casado‐del Castillo, V., Tello, V., de Vega‐Bartol, J. J., Ramos, B.,
Sukno, S. A., & Díaz Mínguez, J. M. 2016. The FTF gene family regulates virulence and expression of SIX effectors in Fusarium oxysporum. Molecular plant pathology.
Niño-Sánchez, J., Tello, V., Casado-del Castillo, V., Thon, M. R., Benito, E. P., & Díaz-
Mínguez, J. M. 2015. Gene expression patterns and dynamics of the colonization of common bean (Phaseolus vulgaris L.) by highly virulent and weakly virulent strains of Fusarium oxysporum. Frontiers in Microbiology 6: 234
O´Donnell, K. 2000. Molecular phylogeny of the Nectria haematococca-Fusarium solani
species complex. Mycologia 92: 919-938. O´Donnell, K., Kistler, H. C., Cigelnik, E. & Ploetz, R. C. 1998. Multiple evolutionary origins
of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proceedings of the National Academy of Sciences U.S.A. 95:2044-2049.
O'Donnell, K., Sarver, B. A., Brandt, M., Chang, D. C., Noble-Wang, J., Park, B. J., & Geiser,
D. M. 2007. Phylogenetic diversity and microsphere array-based genotyping of human pathogenic fusaria, including isolates from the multistate contact lens-associated US keratitis outbreaks of 2005 and 2006.Journal of Clinical Microbiology 45(7): 2235-2248.
O'Donnell, K., Sutton, D. A., Rinaldi, M. G., Magnon, K. C., Cox, P. A., Revankar, S. G., &
Padhye, A. 2004. Genetic diversity of human pathogenic members of the Fusarium oxysporum complex inferred from multilocus DNA sequence data and amplified fragment length polymorphism analyses: evidence for the recent dispersion of a geographically widespread clonal lineage and nosocomial origin. Journal of Clinical Microbiology 42(11): 5109-5120.
Olivain, C., & Alabouvette, C. 1999. Process of tomato root colonization by a pathogenic
strain of Fusarium oxysporum f. sp. lycopersici in comparison with a non‐pathogenic strain. New Phytologist 141(3): 497-510.
Ortoneda M., Guarro, J., Madrid, M. P., Caracuel, Z., Roncero, M. I. G., Mayayo, E., & Di
Pietro, A. 2004. Fusarium oxysporum as a multihost model for the genetic dissection of fungal virulence in plants and mammals. Infection and Immunity 72(3): 1760-1766.
Bibliografía
- 209 -
Pagnussat, G. C., Simontacchi, M., Puntarulo, S., & Lamattina, L. 2002. Nitric oxide is required for root organogenesis. Molecular Plant Pathology 129(3): 954-6.
Pareja-Jaime, Y., Roncero, M. I. G., & Ruiz-Roldán, M. C. 2008. Tomatinase from Fusarium
oxysporum f. sp. lycopersici is required for full virulence on tomato plants. Molecular Plant-Microbe Interactions 21(6): 728-736.
Pareja‐Jaime, Yolanda., Martín‐Urdíroz, Magdalena., Roncero, Mig., González‐Reyes, J.
A., & Roldán, M. D. C. R. 2010. Chitin synthase‐deficient mutant of Fusarium
oxysporum elicits tomato plant defence response and protects against wild‐type infection. Molecular Plant Pathology 11(4): 479-493.
Pastor-Corrales, M. A., & Abawi, G. S. 1987. Reactions of selected bean germ plasms to
infection by Fusarium oxysporum f. sp. phaseoli. Plant Disease 71(11): 990-993. Perazzolli, M., Dominici, P., Romero-Puertas, M. C., Zago, E., Zeier, J., Sonoda, M. &
Delledonne, M. 2004. Arabidopsis nonsymbiotic hemoglobin AHb1 modulates nitric oxide bioactivity. The Plant Cell 16(10): 2785-2794.
Pérez-Nadales, E., & Di Pietro, A. 2011. The membrane mucin Msb2 regulates invasive
growth and plant infection in Fusarium oxysporum. The Plant Cell 23(3): 1171-1185. Ploetz, R. C., Trec, Ifas and Correll, J. C. 1988. Vegetative compatibility among races of
Fusarium oxysporum f.sp. cubense. Plant Disease 72(4): 325-328. Poole, R. K., & Hughes, M. N. 2000. New functions for the ancient globin family: bacterial
responses to nitric oxide and nitrosative stress. Molecular Microbiology 36(4): 775-783.
Poole, R. K., Ioannidis, N., & Orii, Y. 1994. Reactions of the Escherichia coli
flavohaemoglobin (Hmp) with oxygen and reduced nicotinamide adenine dinucleotide: evidence for oxygen switching of flavin oxidoreduction and a mechanism for oxygen sensing. Proceedings of the Royal Society of London B: Biological Sciences 255(1344): 251-258.
Poole, R.K., Anjum, M.F., Membrillo-Hernandez, J., Kim, S.O., Hughes, M.N., & Stewart, V.
1996. Nitric oxide, nitrite, and Fnr regulation of hmp (flavohemoglobin) gene expression in Escherichia coli K-12. Journal Bacteriology 178: 5487-92.
Prats, E., Carver, T. L., & Mur, L. A. 2008. Pathogen-derived nitric oxide influences
formation of the appressorium infection structure in the phytopathogenic fungus Blumeria graminis. Research in Microbiology 159(6): 476-480.
Price D. 1984. Fusarium and plant pathology: the reservoir of infection. pp. 71-93 In: Moss
MO, Smith JE. (eds.) The applied mycology of Fusarium. Cambridge University Press, Cambridge.
Price, M. N., Dehal, P. S., & Arkin, A. P. 2009. FastTree: computing large minimum evolution
trees with profiles instead of a distance matrix. Molecular Biology and Evolution 26(7): 1641-1650.
Puhalla, J. E. 1985. Classification of strains of Fusarium oxysporum based on vegetative
compatibility. Canadian Journal of Botany 63: 179-183.
Papel de las flavohemoglobinas en F. oxysporum
- 210 -
Ramos, B., Alves-Santos, F. M., García-Sánchez, M. A., Martín-Rodrigues, N., Eslava, A. P., & Díaz-Mínguez, J. M. 2007. The gene coding for a new transcription factor (ftf1) of Fusarium oxysporum is only expressed during infection of common bean. Fungal Genetics and Biology 44(9): 864-876.
Ribeiro, R. D. L., & Hagedorn, D. J. 1979. Screening for resistance to and pathogenic
specialization of Fusarium oxysporum f. sp. phaseoli, the causal agent of bean yellows. Phytopathology 69(3): 272-276.
subcellular localization of nitric oxide synthase in maize cells. FEBS Letters 445:283-286.
Rispail, N., & Di Pietro, A. 2009. Fusarium oxysporum Ste12 controls invasive growth and
virulence downstream of the Fmk1 MAPK cascade. Molecular Plant-microbe Interactions 22(7): 830-839.
Rockel, P., Strube, F., Rockel, A., Wildt, J., & Kaiser, W. M. 2002. Regulation of nitric oxide
(NO) production by plant nitrate reductase in vivo and in vitro. Journal of Experimental Botany 53(366): 103-110.
Rodriguez-Galvez, E., & Mendgen, K. 1995. Cell wall synthesis in cotton roots after infection
with Fusarium oxysporum. Planta 197(3): 535-545. Romero-Puertas, M. C., Laxa, M., Mattè, A., Zaninotto, F., Finkemeier, I., Jones, A. M., &
Delledonne, M. 2007. S-nitrosylation of peroxiredoxin II E promotes peroxynitrite-mediated tyrosine nitration. The Plant Cell 19(12): 4120-4130.
Roszer, T. 2012. The biology of subcellular nitric oxide. London. Heidelberg, Springer. Salgado, M. O., & Schwartz, H. F. 1993. Physiologycal specialization and effects of
inoculum concentration of Fusarium oxysporum f.sp. phaseoli on common beans. Plant Disease 77(5): 492-496.
Samalova, M., Johnson, J., Illes, M., Kelly, S., Fricker, M., & Gurr, S. 2013. Nitric oxide
generated by the rice blast fungus Magnaporthe oryzae drives plant infection. New Phytologist 197(1): 207-222.
Samolski, I., de Luis, A., Vizcaíno, J. A., Monte, E., & Suárez, M. B. 2009. Gene expression
analysis of the biocontrol fungus Trichoderma harzianum in the presence of tomato plants, chitin, or glucose using a high-density oligonucleotide microarray. BMC Microbiology 9(1): 217.
Samuels, G. J., & Blackwell, M. 2001. Pyrenomycetes fungi with perithecia. In Systematics
and Evolution (pp. 221-255). Springer Berlin Heidelberg. Sambrook, J., Fritsch, E. F., & Maniatis, T. 1989. Molecular cloning (Vol. 2, pp. 14-9). New
Yor. Cold Spring Harbor Laboratory Press. Santander, D. 2014. Estudio del metabolismo del óxido nítrico (NO) en Botrytis cinerea:
mecanismos de producción y efectos fisiológicos. Tesis Doctoral. Stamler, J. S., Simon, D. I., Osborne, J. A., Mullins, M. E., Jaraki, O., Michel, T., & Loscalzo,
J. 1992. S-nitrosylation of proteins with nitric oxide: synthesis and characterization
Bibliografía
- 211 -
of biologically active compounds. Proceedings of the National Academy of Sciences of the U.S.A 89(1): 444-448.
Sarver, A., & DeRisi, J. 2005. Fzf1p regulates an inducible response to nitrosative stress in
Saccharomyces cerevisiae. Molecular Biology of the Cell 16(10): 4781-4791. Simontacchi, M., Jasid, S., & Puntarulo, S. 2004. Nitric oxide generation during early
germination of sorghum seeds. Plant Science 167(4): 839-847. Southern, E. M. 1975. Detection of specific sequences among DNA fragments separated
by gel electrophoresis. Journal of Molecular Biology 98(3): 503-517. Schäfer, W. 1994. Molecular mechanisms of fungal pathogenicity to plants. Annual Review
of Phytopathology 32: 461-477. Schäfer, K., Di Pietro, A., Gow, N. A., & MacCallum, D. 2014. Murine model for Fusarium
oxysporum invasive fusariosis reveals organ-specific structures for dissemination and long-term persistence. PloS one 9(2): e89920.
Stajich, J. E., Harris, T., Brunk, B. P., Brestelli, J., Fischer, S., Harb, O. S., & Stoeckert, C.
J. 2012. FungiDB: an integrated functional genomics database for fungi. Nucleic Acids Research 40(D1): D675-D681.
Knogge, W. (1996). Fungal infection of plants. The Plant Cell 8(10): 1711. Schinko, T., Berger, H., Lee, W., Gallmetzer, A., Pirker, K., Pachlinger, R. & Strauss, J.
2010. Transcriptome analysis of nitrate assimilation in Aspergillus nidulans reveals connections to nitric oxide metabolism. Molecular Microbiology 78(3): 720-738.
Schinko, T., Gallmetzer, A., Amillis, S., & Strauss, J. 2013. Pseudo-constitutivity of nitrate-
responsive genes in nitrate reductase mutants. Fungal Genetics and Biology 54: 34-41.
Seifert, K. A., & Gams, W. 2001. The taxonomy of anamorphic fungi. In Systematics and
Evolution (pp. 307-347). Springer Berlin Heidelberg. Seifert, K. A., & Levesque, C. A. 2004. Phylogeny and molecular diagnosis of mycotoxigenic
fungi. In Molecular Diversity and PCR-detection of Toxigenic Fusarium Species and Ochratoxigenic Fungi (pp. 449-471). Springer Netherlands.
Shoun, H., Kim, D. H., Uchiyama, H., & Sugiyama, J. 1992. Denitrification by fungi. FEMS
Microbiology Letters 94(3): 277-281. Smagghe, B. J., Trent III, J. T., & Hargrove, M. S. 2008. NO dioxygenase activity in
hemoglobins is ubiquitous in vitro, but limited by reduction in vivo. PloS one 3(4): e2039.
Song, N. K., Jeong, C.S., & Choi, H.S. 2000. Identification of nitric oxide synthase in
Flammulina velutipes. Mycologia (92): 1027-1032. Spiro, S. 2007. Regulators of bacterial responses to nitric oxide. FEMS Microbiology
Reviews 31(2): 193-211.
Papel de las flavohemoglobinas en F. oxysporum
- 212 -
Stakman, E. C., & Jensen, L. 1915. Infection experiments with timothy rust. Journal of Agricultural Research 5(5): 211
Stamler, J. S., Simon, D. I., Osborne, J. A., Mullins, M. E., Jaraki, O., Michel, T., & Loscalzo,
J. 1992. S-nitrosylation of proteins with nitric oxide: synthesis and characterization of biologically active compounds. Proceedings of the National Academy of Sciences of the U.S.A 89(1): 444-448.
Stuehr, D. J. 1999. Mammalian nitric oxide synthases. Biochimica et Biophysica Acta
(BBA)-Bioenergetics 1411(2): 217-230. Suga, H., Ikeda, S., Taga, M., Kageyama, K. and Hyakumachi, M. 2002. Electrophoretic
karyotyping and gene mapping of seven formae speciales in Fusarium solani. Current Genetics 41: 254-260.
Szibor, M., Richter, C., & Ghafourifar, P. 2001. Redox control of mitochondrial functions.
Antioxidants and Redox Signaling 3(3): 515-523. Takaya, N. 2002. Dissimilatory nitrate reduction metabolisms and their control in fungi.
Journal of Bioscience and Bioengineering 94(6): 506-510. Takaya, N., Suzuki, S., Matsuo, M., & Shoun, H. 1997. Purification and characterization of
a flavohemoglobin from the denitrifying fungus Fusarium oxysporum. FEBS Letters 414(3): 545-548.
Tan, M. K. & Niessen, M. 2003. Analysis of rDNA ITS sequences to determine genetic
relationships among, and provide a basis for simplified diagnosis of Fusarium species causing crown rot and head blight of cereals. Molecular Genetics and Genomics 107: 811-821.
Te Biesebeke, R., Levasseur, A., Boussier, A., Record, E., Van den Hondel, C. A., & Punt,
P. J. 2010. Phylogeny of fungal hemoglobins and expression analysis of the Aspergillus oryzae flavohemoglobin gene fhbA during hyphal growth. Fungal Biology 114(2): 135-143.
Teunissen, H. A., Verkooijen, J., Cornelissen, B. J. & Haring, M. A. 2002. Genetic exchange
of avirulence determinants and extensive karyotype rearrangements in parasexual recombinants of Fusarium oxysporum. Molecular Genetics and Genomics. 268: 298-310.
Thompson, J. D., Higgins, D. G., & Gibson, T. J. 1994. CLUSTAL W: improving the
sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic acids Research 22(22): 4673-4680.
Turrà, D., & Di Pietro, A. 2015. Chemotropic sensing in fungus–plant interactions. Current
Opinion in Plant Biology 26: 135-140. Turrión-Goméz, J. L., & Benito, E. P. 2011. Flux of nitric oxide between the necrotrophic
pathogen Botrytis cinerea and the host plant. Molecular Plant Pathology 12(6): 606-616.
Turrión-Goméz, J. L., Eslava, A. P., & Benito, E. P. 2010. The flavohemoglobin BCFHG1 is
the main NO detoxification system and confers protection against nitrosative
Bibliografía
- 213 -
conditions but is not a virulence factor in the fungal necrotroph Botrytis cinerea. Fungal Genetics and Biology 47(5): 484-496.
Ullmann, B. D., Myers, H., Chiranand, W., Lazzell, A. L., Zhao, Q., Vega, L. A., Lopez-Ribot,
J. L., Gardner, P. R., & Gustin, M. C. 2004. Inducible defense mechanism against nitric oxide in Candida albicans. Eukaryot Cell 3(3): 715-23.
Urban, M., Daniels, S., Mott, E., & Hammond‐Kosack, K. 2002. Arabidopsis is susceptible
to the cereal ear blight fungal pathogens Fusarium graminearum and Fusarium culmorum. The Plant Journal 32(6): 961-973.
Valderrama, R., Corpas, F. J., Carreras, A., Fernández-Ocaña, A., Chaki, M., Luque, F., &
Barroso, J. B. 2007. Nitrosative stress in plants. FEBS Letters 581(3): 453-461. Vallance, P., & Collier, J. 1994. Biology and clinical relevance of nitric oxide. BMJ: British
Medical Journal 309(6952): 453. Van Schoonhoven, A., & Pastor-Corrales, M. A. 1987. CIAT. Standard evaluation of bean
germplasm. Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia. Vasudevan, S. G., Armarego, W. L., Shawl, D. C., Lilley, P. E., Dixon, N. E., & Poole, R. K.
1991. Isolation and nucleotide sequence of the hmp gene that encodes a haemoglobin-like protein in Escherichia coli K-12. Molecular and General Genetics MGG 226(12): 49-58.
Vieira, A. L., Linares, E., Augusto, O., & Gomes, S. L. 2009. Evidence of a Ca 2+-NO-cGMP
signaling pathway controlling zoospore biogenesis in the aquatic fungus Blastocladiella emersonii. Fungal Genetics and Biology 46(8): 575-584.
Vinogradov, S. N. 2008. Chapter Thirty-One-Tracing Globin Phylogeny Using PSIBLAST
Searches Based on Groups of Sequences. Methods in Enzymology 436: 571-583. Vinogradov, S. N., Hoogewijs, D., Bailly, X., Arredondo-Peter, R., Guertin, M., Gough, J., &
Vanfleteren, J. R. 2005. Three globin lineages belonging to two structural classes in genomes from the three kingdoms of life. Proceedings of the National Academy of Sciences of the U.S.A 102(32): 11385-11389.
Vinogradov, S. N., Hoogewijs, D., Bailly, X., Mizuguchi, K., Dewilde, S., Moens, L., &
Vanfleteren, J. R. 2007. A model of globin evolution. Gene 398(1): 132-142. Voigt, K., Schleier, S., & Brückner, B. 1995. Genetic variability in Gibberella fujikuroi and
some related species of the genus Fusarium based on random amplification of polymorphic DNA (RAPD). Current Genetics 27(6): 528-535.
Wang, J., & Higgins, V. J. 2005. Nitric oxide has a regulatory effect in the germination of
conidia of Colletotrichum coccodes. Fungal Genetics and Biology 42(4): 284-292. Wang, Y., Jiang, C. J., Li, Y. Y., Wei, C. L., & Deng, W. W. 2012. CsICE1 and CsCBF1: two
transcription factors involved in cold responses in Camellia sinensis. Plant cell Reports 31(1): 27-34.
Papel de las flavohemoglobinas en F. oxysporum
- 214 -
Wittenberg, J. B., Bolognesi, M., Wittenberg, B. A., & Guertin, M. 2002. Truncated hemoglobins: a new family of hemoglobins widely distributed in bacteria, unicellular eukaryotes, and plants. Journal of Biological Chemistry 277(2): 871-874.
Wiemann, P., Brown, D. W., Kleigrewe, K., Bok, J. W., Keller, N. P., Humpf, H. U., &
Tudzynski, B. 2010. FfVel1 and FfLae1, components of a velvet‐like complex in Fusarium fujikuroi, affect differentiation, secondary metabolism and virulence. Molecular Microbiology 77(4): 972-994.
Wong, K. H., Hynes, M. J., & Davis, M. A. 2008. Recent advances in nitrogen regulation: a
comparison between Saccharomyces cerevisiae and filamentous fungi. Eukaryotic Cell 7(6): 917-925.
Woo, S. L., Zoina, A., Del Sorbo, G., Lorito, M., Nanni, B., Scala, F. & Noviello, C. 1996.
Characterization of Fusarium oxysporum f.sp. phaseoli by pathogenic races, VCGs, RFLPs, and RAPD. Phytopathology 86(9): 966-973.
Comparison of epidemics of Botrytis fruit rot and powdery mildew of strawberry in large plastic tunnel and field production systems. Plant Disease 85: 901-909.
Yedidia, I., Benhamou, N., & Chet, I. 1999. Induction of defense responses in cucumber
plants (Cucumis sativus L.) by the biocontrol agent.Trichoderma harzianum. Applied and Environmental Microbiology 65(3): 1061-1070.
Yoder, W. T. and Christianson, L. M. 1998. Species-specific primers resolve members of
Fusarium section Fusarium. Fungal Genetic Biology 23: 68-80. Zafra, A., Rodríguez-García, M. I., & Alché, J. D. 2010. Cellular localization of ROS and NO
in olive reproductive tissues during flower development. BMC Plant Biology 10(1): 36.
Zeier, J., Delledonne, M., Mishina, T., Severi, E., Sonoda, M., & Lamb, C. 2004. Genetic
elucidation of nitric oxide signaling in incompatible plant-pathogen interactions. Plant Physiology 136(1): 2875-2886.
Zhao, X. J., Raitt, D., Burke, P. V., Clewell, A. S., Kwast, K. E., & Poyton, R. O. 1996.
Function and Expression of Flavohemoglobin in Saccharomyces cerevisiae evidence for a role in the oxidative stress response. Journal of Biological Chemistry 271(41): 25131-25138.
Zhou, S., Fushinobu, S., Kim, S. W., Nakanishi, Y., Wakagi, T., & Shoun, H. 2010.
Aspergillus oryzae flavohemoglobins promote oxidative damage by hydrogen peroxide. Biochemical and Biophysical Research Communications 394(3): 558-561.
Zhou, S., Fushinobu, S., Nakanishi, Y., Kim, S. W., Wakagi, T., & Shoun, H. 2009. Cloning
and characterization of two flavohemoglobins from Aspergillus oryzae. Biochemical and Biophysical Research Communications 381(1): 7-11.
Zhou, Z., Takaya, N., Nakamura, A., Yamaguchi, M., Takeo, K., & Shoun, H. 2002.
Ammonia fermentation, a novel anoxic metabolism of nitrate by fungi. Journal of Biological Chemistry 277(3): 1892-1896.
Bibliografía
- 215 -
Zhou, Z., Takaya, N., Sakairi, M. A. C., & Shoun, H. 2001. Oxygen requirement for denitrification by the fungus Fusarium oxysporum. Archives of Microbiology 175(1): 19-25.
Zhu, H., & Riggs, A. F. 1992. Yeast flavohemoglobin is an ancient protein related to globins
and a reductase family. Proceedings of the National Academy of Sciences of the U.S.A 89(11): 5015-5019.
Zhu, J., Jambhekar, A., Sarver, A., & DeRisi, J. 2006. A Bayesian network driven approach
to model the transcriptional response to nitric oxide in Saccharomyces cerevisiae. PloS One 1(1): e94.
Zumft, W. G. 1997. Cell biology and molecular basis of denitrification. Microbiology and