University of Groningen Thermotropic liquid crystals from engineered polypeptides Pesce, Diego IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below. Document Version Publisher's PDF, also known as Version of record Publication date: 2015 Link to publication in University of Groningen/UMCG research database Citation for published version (APA): Pesce, D. (2015). Thermotropic liquid crystals from engineered polypeptides. [Groningen]: University of Groningen. Copyright Other than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons). Take-down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons the number of authors shown on this cover page is limited to 10 maximum. Download date: 09-07-2020
Transcript
University of Groningen
Thermotropic liquid crystals from engineered polypeptidesPesce, Diego
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2015
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Pesce, D. (2015). Thermotropic liquid crystals from engineered polypeptides. [Groningen]: University ofGroningen.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Abstract Liquid Crystals (LCs) can be found in various contexts ranging from DNA and cell membranes to sensor technology and soaps, providing a truly multidisciplinary environment for research at the interfaces of chemistry, biology, physics and engineering. Their physical behavior classifies them as a fourth state of matter, lying between the crystalline solid and disordered liquid states. The liquid crystalline mesopases transition is influenced by temperature and concentration, giving rise to different kinds of ordered structures. Particularly, LCs play an important role in biology because of their essential characteristic, the combination of order and mobility that is a basic requirement for self-organization and structure formation in living systems. Among biomacromolecules, many proteins and polypeptides can form liquid crystalline structures both in vivo and in vitro. A detailed systematic study of liquid crystalline behavior of proteins is required to better understand the nature of protein liquid crystals in biological systems and for implementing their utilization in medicine and material engineering. Here we reviewed examples from biological liquid crystalline materials with a focus on protein and polypeptides, both naturally occurring and synthetic. Furthermore, we highlighted their potential as versatile materials for applications in biotechnology and material science.
3
1.1 Liquid Crystalline mesophases In nature matter exists in three different states: solid, liquid and gas. In each of these states, atoms and molecules, the building blocks of matter, have a different degree and type of order with respect to their neighbors. Two kinds of order are distinguishable: i) positional order, where molecules are arranged in any sort of ordered lattice, and ii) orientational order, where molecules are mostly pointing in the same direction. In addition, the order can be either short-range (i.e. only between molecules close to each other) or long-range (i.e. extending to larger, sometimes macroscopic, dimensions). In orientational order the direction of preferred alignment is usually described by a unit vector, the director L, which is simply the symmetry axis of the orientational distribution (Figure 1). Depending on the kind of order the physical properties of matter are either uniform in all orientations and thus implying identical properties in all directions, or directionally dependent. These states of matter organization are isotropic and anisotropic, respectively1. Crystalline solids have highly ordered structures, whilst gases do not show any positional or orientational order and liquids possess only short-range but no long-range ordering. The borders between the different states are not always clear and there are a large number of other intermediate phases called mesophases. In particular, intermediate mesophases - which exhibit long-range orientational order - exist between solid and liquid states. Those mesophases are known as Liquid Crystals (LCs), since they reveal features belonging to both solid and fluid states. This fourth state of matter generally possesses orientational or weak positional order. Thus, it reveals several physical properties of crystalline state and, at the same time, has the ability to flow as a liquid. Because of the orientational order, most physical properties of LCs are anisoptropic. Additionally, there are new physical qualities, which do not appear in simple liquids such as: magnetic susceptibility, optical birefringence, elastic or frictional torques (rotational viscosity). All these properties are extremely sensitive to external perturbations because of the liquid-like fluidity of LCs. For example, the birefringence can be manipulated easily with the help of rather weak external magnetic, electric or optical fields, leading to magneto-optical, electro-optical and opto-optical effect. Therefore, the most successful application of LCs is in display technology, which takes advantage of the electro-optical effect. Comprehensively, LCs can be partitioned into two classes: thermotropic LCs
4
and lyotropic LCs. The phase transitions of thermotropic LCs depend on temperature, while those of lyotropic LCs depend on both temperature and concentration.
1.1.1 Thermotropic Liquid Crystals LCs are defined as thermotropic if the transitions between the phases are due to the action of heat alone. At high temperatures, the axes of the LC molecules randomly orient resulting in the isotropic phase as schematically represented in figure 1. One phase that forms with decreasing temperatures is the nematic phase (Figure 1a). The nematic phase is the least ordered of the mesophases, as the molecules possess only orientational but no long-range positional order. If a nematic LC is made of chiral molecules a cholesteric LC phase is also formed. The name cholesteric derives from the fact that the first molecules
FIGURE 1. THERMOTROPIC MESOPHASES. Schematic structures of various thermotropic LCs phases as function of the temperature. i) Cristalline, ii) smectic and iii) nematic are anisotropyic phases whereas iv) is the isotropic liquid state.
5
found to display these properties were those related to cholesterol. In cholesteric LC, the direction of preferred orientation forms a helical structure, with the helical axis being perpendicular to the director L. The most prominent characteristic of cholesteric LC is a set of equally spaced parallel lines (bright and dark lines), somewhat reminiscent of a fingerprint when observed by polarizing optical micrograph (POM) (Figure 4a). The distance between the alternating bright and dark lines is called the periodicity S, which is equal to half the pitch (p/2) of the torsion of cholesteric LC (Figure 1b). A second mesophase having positional ordering is known as the smectic phase (Figure 1a) and is due to layered molecular arrangement, characterized by additional degrees of positional order. There are different kinds of smectic phases, for example the smectic A, where the molecular orientation is perpendicular to the layers, and the smectic C, where the director is tilted (Figure 1b). Several other kinds of smectic phases can exist, depending on the extent and nature of positional order within the layers. Below a certain temperature, the crystalline state is eventually reached (Figure 1a). Such a temperature dependence of the material phases is reversible and a specific mesosphase can be equilibrated by temperature stabilization.
1.1.2 Lyotropic Liquid Crystals In lyotropic LCs, phase transition between mesophases may also be affected by the addition of appropriate solvents, such as surfactants, and their concentration. The molecules that form lyotropic LCs are amphipatic. The objects created by amphiphiles are usually spherical, as in the case of micelles, but may also be disc-like, rod-like or biaxial. These anisotropic self-assembled nano-structures can order themselves forming large-scale versions of all the thermotropic phases, in a similar way as thermotropic liquid crystals do. A generic progression of phases, going from low to high amphiphile concentration, is: i) discontinuous cubic phase (micellar cubic phase); ii) hexagonal phase (hexagonal columnar phase); iii) lamellar phase; iv) bicontinuous cubic phase; v) reverse hexagonal columnar phase; vi) and inverse cubic phase (inverse micellar phase) (Figure 2). A very simple model, which predicts lyotropic phase transitions, is the hard-rod model proposed by Lars Onsager1,2. This model predicts that a solution of rod-shaped objects, at sufficient concentration, will undergo a phase transition into a nematic phase.
6
1.2 Biological Liquid Crystals The main characteristics of biological materials are hierarchical structures, multi-functionality and self-assembly. The combination of order and mobility in living systems is a basic requirement for self-organization and structure formation in solution3,4, since solids do not easily respond to change and isotropic liquids have less capacity to include functionality. As a result, mesophases are present throughout nature5. All biological liquid crystals involve the formation of lyotropic phases as they are formed by aggregates of molecules in a solvent, mainly water. In the past decades, the study of lyotropic liquid crystals from biomolecules and bio molecular assemblies has attracted attention in diverse areas such as tissue engineering6, neurodegenerative diseases7, gene therapy8, food science9, and prebiotic chemistry 10. LC phases have been identified in many structural and functional biological materials, here we summarize examples ranging
FIGURE 2. LYOTROPIC MESOPHASES. Some lyotropic LC phases as a function of temperature and concentration of the amphiphilic molecules a) dissolved surfactant being in a disordered state, b) micellar, c) cubic, d) cylindrical and e) lamellar phases.
7
from phospholipid cell membranes, to some concentrated DNA and protein solutions in the secretion of spiders used to generate silk (Table 1). TABLE 1. BIOLOGICAL LIQUID CRYSTALS. Compilation of liquid crystals from biological molecules in vivo and in vitro.
BIOPOLYMER FUNCTION LC ORGANISATION OCCURRENCE Ref.
DNA
Chromosomal DNA Efficient packing Cholesteric In vivo 13,19
Phage DNA Efficient packing Columnar In vivo 12
Bacterial Plasmids Efficient packing Cholesteric In vivo / In vitro 20
Glycopolymers
Cellulose Structural Helicoidal / Cholesteric In vitro / In vivo 17
Starch Reservoir of energy Smectic In vitro 9
Chitin Structural Helicoidal / Cholesteric In vitro / In vivo 24,25
Lipids
Phospholipids Structural/Functional Layered lamellar In vivo / In vitro 88
Proteins
Collagen Structural Nematic / Cholesteric In vivo / In vitro 27,32
Spider Silk Structural / Functional Nematic In vivo / In vitro 28
Silkworm Silk Structural / Functional Nematic In vivo / In vitro 37
F-actin Structural / Functional Nematic / Smectic In vitro 30
Microtubules Structural / Functional Nematic In vitro 41
β-Lactoglobulin Functional Nematic In vitro 45
Lisozyme Functional Nematic In vitro 42
Insulin Functional Nematic In vitro 44
8
The liquid crystalline state is important for different features found in nature: i) efficient packing, as in concentrated DNA solutions11 in viruses12 and sperm13; ii) surface directed self assembly, as in surface supported growth in cellulose plywood14; iii) low viscosity, important in flow processing in silk spinning15; iv) sensor actuator ability as in outer hair cells16; v) mechanical strength, as in cellulose in plant cell walls, chitin in insect exocuticole and collagen in human bones17 (Table 1). The mesophase ordering in biological mesogens is usually chiral nematic, in particular cholesteric. Biological LCs and fibrous biological materials are also referred to as helicoidal plywoods18 because of their typical parallel fiber arrangement (Figure 3). In the next sections, a brief overview is given on naturally occurring liquid crystal phases according to molecular classes. The emphasis is laid on natural and synthetic polypeptides and proteins since they are a major subject of this thesis.
1.2.1 DNA LC formation of DNA is observed where efficient packing is required, examples are: the nuclei of cells such as dinoflagellates19, and sperm cells13, the heads of bacteriophages12, or plasmid DNA within bacteria20. In dinoflagellates, the twisted arrangement of chromosomes forms a cholesteric liquid crystal phase19. In bacteriophages, DNA is packed in a columnar fashion within concentric rings21. Plasmid DNA in E. coli cells forms dense
FIGURE 3. BIOLOGICAL PLYWOOD. (a–c) Schematics of the helicoidal plywood found in animals and plants. Oblique sections (c) give the characteristic arc pattern observed in extracellular fibrous composite biomaterials. (d and e) Side and normal view schematics of the helix axis, defined by the helix orientation h (dashed line in d), the pitch p0 and the rotation sense (counter-clockwise into the page in d) the layer distance between a 2π rotation of the fibres is the pitch p0, whose magnitude is in the micrometer range.18
9
clusters with long-range order, and in vitro it shows birefringent liquid crystal textures corresponding to a cholesteric phase at physiological concentrations20.
1.2.2 Glycopolymers/carbohydrates Glycopolymers are also known to form LCs. Cellulose is one of the most widely found biopolymers in nature and it is a component of the wall of many types of plant cell. Cellulose micro-fibrils within plant cell walls exhibit a helicoidal structure, which may be templated via a cholesteric liquid crystal phase17,22. The cholesteric phase has also been observed in vitro in concentrated solution of cellulose and derivatives of cellulose23. Starch has an important energy storage function in plants. It consists of the linear glucan amylose and the branched amylopectin. The amylopectin side chains form a smectic structure and the packing depends on the hydration level9. Chitin has a structural role in the exoskeleton of arthropods and is a long-chain polymer of N-acetylglucosamine. α-Chitin can be processed by acid hydrolysis to produce rod-like nanoparticles, which can form a chiral nematic phase24. Iridescent colouring of certain beetle wings results from helicoidal stacking of fibrous chitin layers, as for cellulose microfibrils25, and it is possible that LC phases may act as precursors for these structures.
1.2.3 Lipids and Membranes Lipids that constitute the cell membranes are perhaps the most important example of a biological LC. All biological membranes are liquid-crystalline structures and exhibit the typical layered lamellar structure26. Membranes exert structural and functional roles such as separation of different fluid compartments, selective transport and signal transduction. They consist of phospholipid bilayers, with simple and conjugate proteins immersed in it. Myelin, prominent in the transmission of electrical impulses by the nerve, is also a LC and it consists of concentric cylindrical phospholipid bilayers.
1.2.4 Polypeptides and proteins Polypeptides and proteins often self-assemble into fibrillar structures. In (aqueous) solution, at sufficiently high concentration, these fibrillar structures
10
can orient and pack into lyotropic mesophases. As mentioned above, the liquid crystalline state can play a pivotal role in the self-assembly process. Many of the living world’s structural materials are based on self-assembled protein fibers. Diverse examples include collagen27, silk28, tubulin29 and actin30. Some of these materials, like collagen and silk, have recently been described as a new class of matter: LC elastomers. LC elastomers can be defined as lightly cross-linked main-chain or side-chain liquid crystalline polymers showing elastomeric properties31. Interestingly, unlike conventional rubbers, at rest, LC elastomers are ordered rather than disordered.
Collagen plays a vital structural role in tissues such as tendon, skin, bone and cornea. It has a triple helix structure of peptide strands rich in glycine, in particular in sequences of type Gly-Pro-Y or Gly-X-Hyp (where X and Y are any amino acid other than glycine, proline or hydroxyproline). Many different types of collagen are known. Type I collagen monomers can form nematic, precholesteric and cholesteric phases in vitro, in dilute acid solution27. Their liquid crystallinity and orientation was first observed, on the basis of birefringence and magnetic field alignment in studies of collagen from rat-tail tendon under acidic conditions32 (Figure 4). Pro-collagen fibrils can also form nematic and precholesteric liquid crystal phases under physiological buffer conditions33 (Figure 4). It has been proposed that collagen fibrils belong to the
FIGURE 4. LIQUID CRYSTALLINITY IN COLLAGEN. a) Ordering of collagen molecules in concentrated media observed in polarized light microscopy. Collagen fingerprint patterns typical of a liquid crystalline cholesteric phase. bar =10 um. b) Structure of the collagen fibril. Thin section of rat tail tendon observed in transmission electron microscopy with uranyl acetate staining. bar = 100 nm.34
11
class of biological LC elastomers, since they are weakly cross-linked networks with elastic mechanical properties31. The arrangement of fibrils in vivo may also be LC-like, for instance the ordering of twisted fibrils of bone osteons resembles that of a cholesteric mesophase33. Moreover, liquid crystallinity is observed in collagen in solution under conditions close to the concentration in tissues (50–200 mg mL-1)34. Although direct evidences for liquid crystallinity of collagen in vivo are lacking, LC ordering has been proposed to play an important role in the determination of tissue form33. In this context, the ordering of collagen in bones, fish scales, cornea and in chitin fibers of arthropods has been suggested to rely on self-assembly within mesophases and being the underlying principle for formation of these extracellular matrices35. In another example, collagen (mainly type IV) extracted from bovine lens capsules gives an X-ray diffraction pattern that can be interpreted as nematic ordering of helical collagen chains36. Spider (Nephila clavipes) dragline silk has also been proposed to be a type of LC elastomer, due to its structural and mechanical properties31. Spider dragline silk protein comprises β-crystallites, enriched in the polyalanine repeats, within a matrix of the less ordered glycine-rich segments. The polyalanine segments form transverse lamellae within oriented nanofibrils, extruded from the duct31. These fibrils exhibit nematic ordering28,31. The efficiency of silk spinning depends on the viscosity of the protein solution dope, which is reduced by adopting well-aligned nematic ordering15. In contrast to fibers extruded from silk glands, mechanically drawn fibers show a banded texture, indicating a periodic variation in the orientation of the director37. Another type of silk, produced by silkworms (Bombyx mori), is consisting of β-sheets and is rich in GAGAGS repeats. Efforts are underway to understand the natural spinning process whereby an aqueous solution of silk fibroin is extruded under ambient conditions to produce aligned and water-insoluble fibrils. A nematic phase has been observed in aqueous solutions of silkworm silk fibroin37, and it was suggested to be an intermediate state, orienting the fibrils, in the in vivo processing route37,38. F-actin is the polymerized form of actin adopting helically twisted microfilaments. It has served as a model for a semiflexible biopolymer39. As an important motor protein, it is involved in force transduction in muscles and also plays a structural role in the cytoskeleton. Muscle cells are packed with oriented arrays of these microfilaments, which arrange themselves in a way reminiscent of nematic and smectic LC. Nematic-like ordering was observed
12
in concentrated solutions of actin30 and at the edge of evaporating droplets on glass40. Microtubules (neurofilaments) are the main component of the cytoskeleton in axons of vertebrates. They comprise superstructures of coiled-coil dimers of the central hydrophobic regions of the subunit proteins. The N- and C-terminal domains are unstructured and the latter adopt side arm structures around the filament body. The neurofilaments, which are the main components of neuronal cytoskeleton, can form nematic hydrogels under appropriate conditions of protein and salt concentration41. Besides in structural proteins, nematic phase formation was also observed for functional proteins such as hen lysozyme42, insulin43,44 and β-lactoglobulin45, all of which form amyloid fibrils under appropriate denaturing conditions (generally acidic pH, and/or high temperature) (Table 1). It is worth to mention that nematic order was also observed in supramolecular structures that do not form fibrils, as in the tobacco mosaic virus in colloidal solutions46,47 and the isolated gp8 coat protein of M13 bacteriophage48. Interestingly, various types of mucus are essentially liquid crystalline hydrogels, comprising an entangled network of chains with local rigidity due to glycosylated side chains. For example, the proteoglycan mucin is a polyelectrolyte with a bottlebrush structure characterized by highly charged carbohydrate side chains attached to a semiflexible protein backbone. Small-angle neutron scattering was used to show that mucin from pig stomach forms a nematic phase at high concentration49. Other proteoglycans such as those found in slug slime trails also form nematic phases50, which exhibit viscoelastic properties important to this mode of movement.
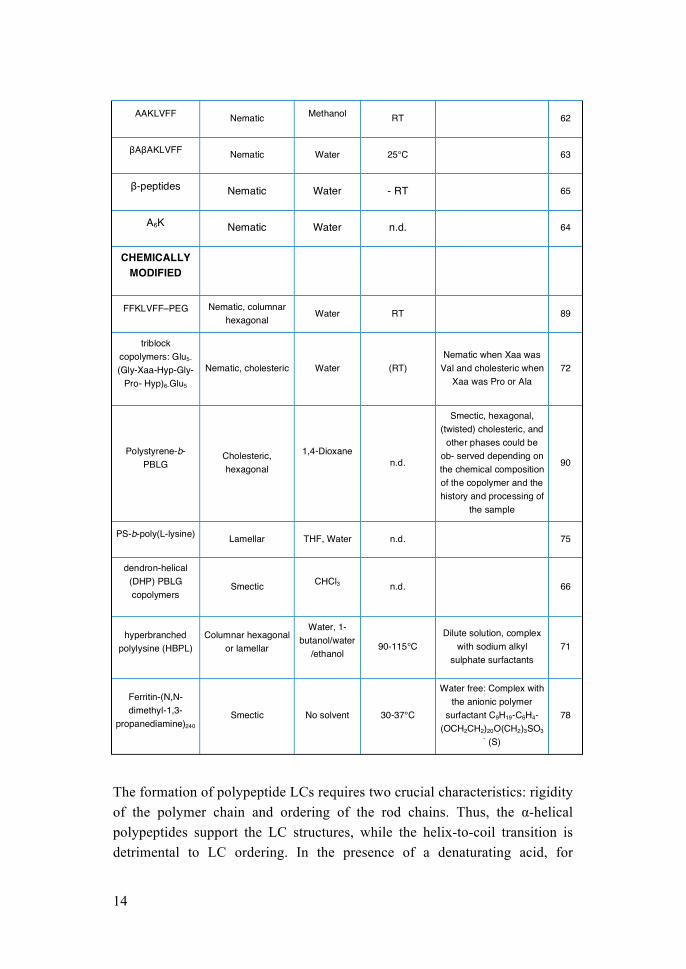
1.3 Liquid Crystals from synthetic polypeptides Several examples of LC phases have been obtained with synthetic and semi-synthetic peptides or polypeptides in vitro, in the presence of different kind of solvents (Table 2). Polypeptides generally adopt α-helix conformation when they are dissolved in organic solvents such as N,N-dimethylformamide, chloroform or benzene. At high concentration, the side-by-side packing of rigid polypeptide chains induces LC structures. Interestingly, the characteristic ordering of polypeptides was first observed from LC structures51.
13
TABLE 2. SYNTHETIC POLYPEPTIDE LIQUID CRYSTALS. Liquid crystals based on synthetic peptides and polypeptides reported in the literature.
POLYPEPTIDE LC
ORGANIZATION SOLVENT CONDITIONS NOTES Ref.
Poly(γ-benzyl L-glutamate) (PBLG) Cholesteric
Chloroform (CHCl3) n.d.
54
PBLG right/left-handed
cholesteric, nematic m-cresol
30–60°C, >60°C
30–60°C right-handed cholesteric; 60°C
nematic; >60°C left-handed cholesteric
57
PBLG right/left-handed
cholesteric
Dioxane-dichloroethan
e 10-70°C
from right-handed to left-handed with increasing
volume fraction of dichloroethane
56
poly(β-p-chlorobenzyl L-
aspartate) (PClBLA)
right/left-handed cholesteric
Trichloroethylene (TCE) 25-100°C
left cholesteric (90°C), nematic (97°C), right cholesteric (102°C)
opposite to change in screw sense of the
polypeptide backbone
58
poly(β-phenethyl L-aspartate (PPLA)
right/left-handed cholesteric, nematic
Trichloroethylene/dichloroa
cetic acid 10-93°C
The screw sense of the polypeptide backbone
changed from left-handed (T<25°C) to
right-handed (25°C<T<89°C) and then to left-handed (T>89°C)
(L–R–L) with corresponding and opposite change in
cholesteric LCs (R-L-R) separate by nematic
phases
59
PBLG (recombinant DNA) Smectic
CHCl3/trifluoroacetic acid
(TFA) RT
Smectic order only when CHCl3 (97%) and TFA
(3%) 46
Ac-QQRFQWQFEQQ-
NH2 Nematic Water 20°C
60
NH2-FF-COOH nanowires Nematic CS2 RT
61
14
The formation of polypeptide LCs requires two crucial characteristics: rigidity of the polymer chain and ordering of the rod chains. Thus, the α-helical polypeptides support the LC structures, while the helix-to-coil transition is detrimental to LC ordering. In the presence of a denaturating acid, for
AAKLVFF Nematic Methanol RT
62
βAβAKLVFF Nematic Water 25°C
63
β-peptides Nematic Water - RT 65
A6K Nematic Water n.d. 64
CHEMICALLY MODIFIED
FFKLVFF–PEG Nematic, columnar hexagonal
Water RT
89
triblock copolymers: Glu5-
(Gly-Xaa-Hyp-Gly-Pro- Hyp)6-Glu5
Nematic, cholesteric Water (RT) Nematic when Xaa was
Val and cholesteric when Xaa was Pro or Ala
72
Polystyrene-b-PBLG
Cholesteric, hexagonal
1,4-Dioxane
n.d.
Smectic, hexagonal, (twisted) cholesteric, and
other phases could be ob- served depending on the chemical composition of the copolymer and the history and processing of
the sample
90
PS-b-poly(L-lysine) Lamellar THF, Water n.d.
75
dendron-helical (DHP) PBLG copolymers
Smectic CHCl3 n.d.
66
hyperbranched polylysine (HBPL)
Columnar hexagonal or lamellar
Water, 1-butanol/water
/ethanol 90-115°C
Dilute solution, complex with sodium alkyl
sulphate surfactants 71
Ferritin-(N,N-dimethyl-1,3-
propanediamine)240 Smectic No solvent 30-37°C
Water free: Complex with the anionic polymer
surfactant C9H19-C6H4-(OCH2CH2)20O(CH2)3SO3
- (S)
78
15
example, polypeptides tend to adopt a random coil state because the acid interferes with the hydrogen bonds involved in the helical architecture52. In that case an anisotropic–isotropic transition (known as reentrant transition) occurs at low temperature since the random coil is unable to induce LC ordering53. LCs of polypeptides were first described in the 1950s by Elliott and Ambrose, who observed a birefringent phase in a solution of poly(γ-benzyl L-glutamate)/ chloroform (PBLG/CHCl3) mixtures54. PBLG later became a model system for studying LCs. Polypeptides are, in general, able to form LCs with nematic, cholesteric and smectic phases (Figure 5).
1.3.1 Nematic and cholesteric phases In the nematic phase (Figure 5b), polypeptide chains have no positional order and self-align to have long-range directional order with their long axes roughly parallel. The nematic cholesteric phase (Figure 5c) is the most common LC structure observed in polypeptides. In the cholesteric phase, the periodicity S and the chirality are found to be dependent on several parameters such as the temperature, polymer concentration, solvent nature and molecular weight55-57. For example, PBLG in the solvent m-cresol, forms right-handed cholesteric LCs in the temperature range of 30–60 °C and the
FIGURE 5. POLYPEPTIDE LIQUID CRYSTALS. Classification of polypeptide LCs: a) smectic, b) nematic and c) cholesteric phases. S indicates the periodicity of cholesterol phase
16
periodicity S increases with increasing temperature57. At 60 °C, the cholesteric character disappears and nematic LCs are formed. Above this temperature, left-handed cholesteric structures appear and the periodicity S decreases with increasing temperature. Regarding the dependence on the nature of the solvent, results from the PBLG LCs formed in dioxane-dichloroethane mixed solvent showed that the chirality of the LC transformed from right-handed to left-handed with increasing volume fraction of dichloroethane56. For such behavior, the dielectric constant of the solvents is believed to be one of the crucial influencing factors. Generally, a solvent with lower dielectric constant supports a right-handed cholesteric structure. The chirality of polypeptide backbones was also found to have an influence on the handedness of LC structures. For example, Abe and collaborators found that the chirality of the LCs is opposite to the screw sense of the polypeptide backbone in the LC structure of poly(β-p-chlorobenzyl L-aspartate)/trichloroethylene (PClBLA/TCE) system58. At room temperature, PClBLA shows a right-handed α-helical conformation, while the chirality of the LC is left-handed. Between 80 and 100 °C the polypeptide α-helix changes its conformation from right to left-handed. Accordingly, the LC structures change from left-handed to right-handed cholesteric, separated by a nematic phase. In another work on poly(β-phenethyl L-aspartate)/trichloroethylene/dichloroacetic acid (PPLA/TCE/DCA) systems, the same authors found a more complex LC behavior59. With increasing temperature of the solution, the screw sense of the polypeptide backbone changed from left-handed to right-handed and then to left-handed (L–R–L). Simultaneously with the helix–helix transition, the handedness of LCs gradually changes from right to left and then to right (R–L–R) (Figure 6), each phase separated by a nematic phase. The inversion of the cholesteric sense is triggered by the reversal of the screw sense of the α-helical backbone. Simple nematic ordering has also been studied for de novo designed short peptides. For example, the peptide Ac-QQRFQWQFEQQ-NH2, designed to form β-tapes, has been shown to form nematic fluid and nematic gel phases60. Nematic liquid crystalline ordering of the aromatic dipeptide NH2-FF-COOH nanowires in the solvent CS2 has recently been discussed61. The stable dispersion of the peptide nanowires exhibited a LC phase for a broad concentration range allowing the rapid alignment of nanowires under an external electric field. Other nanotube and fibril forming peptides, such as AAKLVFF62, βAβAKLVFF (βA denotes the β2-alanine amino acid)63 and the model
17
amphiphilic peptide A6K64, form nematic phases in different solvents. Hartgerink and co-workers reported the nematic phase formation of self-assembling synthetic β-peptides65.
1.3.2 Smectic phases In the smectic phase (Figure 5a), as mentioned before, polypeptide chains are positionally ordered along one direction, forming well-defined layers that can slide over one another. Smectic ordering is common for low molecular weight LC compounds, is mainly found for polypeptides with identical degrees of polymerization and it is rarely observed in synthetic polypeptides due to the polydisperse nature of the materials. To overcome this problem, Tirrell’s group reported for the first time the formation of smectic LC structures from monodisperse polypeptides derived from PBLG, which was synthesized via
FIGURE 6. PPLA BACKBONE AND LC CHIRALITY. Schematic representation of PPLA conformation-induced transition of LC handedness.91
18
recombinant DNA technology46,47 (Figure 7). The LCs of modified PBLG were obtained in mixtures of CHCl3 (97%, v/v) and trifluoroacetic acid (TFA; 3%, v/v). The solution, observed under the polarized optical microscope (Figure 7a), was iridescent with a fan-like texture indicative for smectic order. The densitometer scan of the SAXD pattern of the film (Figure 7b, curve a) showed a well-defined spacing. In contrast, the suggested smectic-like ordering was not present in a solid film of polydisperse PBLG with comparable molecular weight (it yielded no small-angle reflections in a similar diffraction experiment) (Figure 7b, curve b). In figure 7c a illustration for the smectic LC structures is shown. The addition of TFA inhibits aggregation of PBLG in concentrated solutions while the low concentration does not destroy the rigidity of polymer chains. Another strategy is to cap the polypeptide ends with a compact and bulky group. During self-assembly in concentrated solutions, such modified polypeptides can self-assemble into a smectic LC phase66. For example, Winnik and colaborators found that dendron-helical (DHP) PBLG copolymers could form smectic phases in CHCl3
66. The dendritic block prevents the DHP copolymers from aligning into a nematic phase.
LC phase formation was also observed for some peptide based pharmaceutical active compounds - in particular, luteinizing-hormone releasing hormone (LHRH) agonists leuprolide67, nafarelin68 and detirelix68,69, all of which form β-sheet fibrils that form birefringent lyotropic gels in water, and in some cases
FIGURE 7. PBLG LIQUID CRYSTAL. a) POM image of PBLG in CHCl3/TFA. b) Densitometer scans of the SAXD patterns of films prepared from solutions of monodisperse PBLG (curve a) and polydisperse PBLG (curve b). c) Smectic structure of PBLG, showing the origin of the 12.5 A and 114.5 A reflections. As can be seen, the helical rods arrange in layers of thickness 114.5 A , and the distance between two PBLG rods is 12.5 A , which corresponds to the diameter of PBLG rods.91
19
in organic solvents. For the latter two peptides, the liquid crystal phase in aqueous solution was identified as nematic68. Certain compounds, such as the cyclic peptide cyclosporine (natural occurring peptide), may also form thermotropic liquid crystals70. The liquid crystal phase formation is actually a problem observed in formulation that leads to unwanted turbidity, precipitation and gelation69. Hyperbranched polylysine (HBPL) complexed with various anionic, sodium alkyl sulphate surfactants, were observed to form short-range liquid crystal-like order, columnar hexagonal packing or lamellar assembly, depending on the surfactant alkyl chain length71. Synthetic ‘‘triblock’’ copolymers containing Glu5 end blocks and a central collagen-like midblock, (Gly-Xaa-Hyp-Gly-Pro- Hyp)6, where Xaa was one of the amino acids Ala, Pro, Ser or Val, were also shown to exhibit liquid-crystal like ordering72. Nematic ordering was observed when Xaa was Val, and cholesteric-like ordering was obtained with Xaa being either Pro or Ala.
1.4 Liquid Crystals from polymer-peptide conjugate Besides pristine synthetic polypeptides, polystyrene-b-PBLG diblock copolymer may form cholesteric or hexagonal phases in organic solvents73. Swelling of melt structures for the same class of diblock also leads to lamellar phase formation in organic solvents74. In polystyrene-b-poly(L-lysine) diblock copolymer studied by the same authors, deswelling of lamellar structures was noted75. Diblocks of PBLG attached to short peptide blocks (e.g. leucine or leucine/ valine oligomers) may form cholesteric phases, although structures were only observed in cast (dried) films76. Hamley and collaborators have reported on nematic and hexagonal columnar phase formation by the peptide–PEG conjugate FFKLVFF–PEG in aqueous solution77. The peptide is based on a fragment of the amyloid β-peptide, KLVFF, extended at the N-terminus by two phenylalanine residues. Besides peptides, also proteins were transformed into liquid crystals by chemical modification. Recently, Mann and collaborators reported the first example of a solvent-free liquid protein. The protein melt, based on ferritin covalently coupled to N,N-dimethyl-1,3-propanediamine (DMPA) (240 groups per molecule) (Figure 8), exhibits thermotropic liquid-crystalline phase behavior with smectic micro-structure, when complexed with the anionic polymer surfactant (C9H19-C6H4-(OCH2CH2)20O(CH2)3SO3
- (S))78.
20
1.5 Applications and outlook Besides their importance for display technology, several new applications for LCs are emerging in recent years, in particular for LCs from biomacromolecules. Some reports have been published describing new liquid-crystal materials with notable biomedical and biological implications. For example, templates based on nematic and precholesteric (so-called ‘crimped’) structures of collagen were used to control the growth and morphology of fibroblasts79. It was suggested that the high concentration of collagen in the matrices could improve biodegradation compared to dermal equivalents79. Moreover, type I collagen showed to be a versatile template for the structuration of silica, allowing the replication of the chiral nematic organisation in a biologically inspired collagen-silica hybrid material80. Devices and configurations based on liquid-crystal materials are being developed for spectroscopy, imaging and microscopy, leading to new techniques for optically probing biological systems81. A series of
FIGURE 8. SOLVENT-FREE FERRITIN LIQUIDS AND LIQUID CRYSTALS. Proposed structure of the [C-Fn][S] melt at 32 °C showing a lamella liquid-crystalline phase viewed side-on to the layer stacking direction (black arrow; C-Fn (Cationized Ferritin) red, S (anionic polymer surfactant C9H19-C6H4-(OCH2CH2)20O(CH2)3SO3
-) green), with a liquid-like in-plane arrangement comprising short-range local ordering interactions between adjacent polymer chains.78
21
investigations have reported biosensors, which use the interface of thermotropic liquid crystals as a detection system82. With advancing technology, the preparation of peptide-modified interfaces has received a great deal of attention due to the potential utility of these interfaces for monitoring enzymatic activities, controlling cellular behaviors and manipulating peptide-protein interactions83,84. Lundgren and colleagues, for example, developed a liquid crystal pixel array for a high number of signal discrimination in array biosensors85. Regarding advanced materials processing, as mentioned before, regeneration of silk fibers via a wet spinning process from solutions containing silkworm cocoons is of intense interest in order to understand the natural processing route86,87, with the view that this might be used to process new genetically engineered silk proteins. The success in constructing highly ordered smectic phases in solution by using modified polydisperse polypeptide homopolymers, rather than monodisperse samples, is of significant value for the preparation of well-defined supramolecular structures. Highly ordered ultrathin PBLG films with controlled layer spacing and thickness can be prepared from smectic PBLG LCs. When the rods are oriented perpendicular to the substrate, they can be used as nonlinear optical and piezoelectric materials47. When the rods in the films are organized parallel to the substrate, the mono- or multilayer films are useful in creating patterned arrays for use in sensor technology. In addition, the LCs can be further functionalized through tethering bulky groups or polymer chains. In conclusion, the rapid growing synergism between materials science and molecular biology has led to new sources of engineering materials and to new applications for bioengineered proteins. Polypeptides can be produced that either i) copy the principal consensus amino acid repeats that characterize a natural protein; ii) contain the principal repeats from more than one protein, for example hybrid molecules of silk and elastin; iii) contain functional proteins in fusion; or, iv) even contain amino acids not found in nature. Because subsequent hierarchical assembly steps can be conducted in vitro with external control over the temperature, pressure and chemical environment, there is the opportunity for obtaining products with properties not found in nature. This holds especially true for obtaining novel peptide-based LCs with different mesophase for a vast range of potential applications.
22
1.6 Summary Phases of the matter in between the liquid isotropic and the solid anisotropic states are called Liquid Crystals and comprise different mesophases. These mesophases vary depending on the temperature (thermotropic LCs) and concentration of the mesogens (lyotropic LCs). LC is, thus, a “delicate phase of matter” and it is characteristic of many biological materials from DNA to membranes. The perfect balance between order and fluidity appear to be a necessary prerequisite for life and the self-assembly driven by the formation of lyotropic liquid crystal phases takes part in several biological processes. Several protein and polypeptide LC mesogens are present in nature and they take over important structural and functional, cellular and extracellular processes. Moreover, synthetic peptides and polypeptides that show liquid crystalline behavior have been produced thanks to advances of chemical synthesis and genetic engineering. There are many challenges arising for LCs based on proteins and polypeptides. They range from the design of naturally inspired new materials, with extreme control over the synthesis and introduction of functionalities, to a better understanding of fundamental processes in biology.
1.7 Motivation and thesis overview The overall goal of the work described in this thesis was to explore the physical properties of a novel anhydrous thermotropic liquid crystalline material based on protein polyelectrolytes. In Chapter 2 we describe the construction of a genetically engineered elastin-like polypeptide (ELP) carrying several negative charges. The protein electrolyte was monodisperse and had a precisely defined amino acid composition. We further explored the production of thermotropic liquid crystals in the absence of water. To perform this we used a generic method based on the formation of a complex with cationic surfactants containing flexible alkyl tails followed by dehydration. In the second chapter, the phase behavior of these polypeptide-surfactant complexes was investigated employing different techniques such as differential scanning calorimetry (DSC), polarized optical microscopy (POM), small-angle x-ray scattering (SAXS) and wide-angle x-ray scattering (WAXS). After thoroughly characterizing the liquid crystal properties of the pristine
23
complexes, we tested the possibility of incorporating a functional protein within the anhydrous thermotropic LCs, as described in Chapter 3. We first fused the ELP proteins to a Green Fluorescent Protein (GFP) and then we studied the formation of LC complexes. The ELP-GFP fusion proteins were still able to form LCs structures without hampering the fluorescence of GFP even in the anhydrous mesophase. Additionally, we analysed the effect of the length of the ELP segment and of the alkyl chain of the surfactant on the liquid-crystalline behavior. In Chapter 4 we investigate the elastic properties and rheological studies of the ELP-GFP surfactant complexes. They showed elastic behavior similar to native elastin. The elastic values were comparable to those of liquid crystal elastomers in which the mesogens usually need to be cross-linked by polymerization to achieve elastic behavior. Moreover, it was shown how the elasticity of the material can be controlled by varying the lengths of alkyl chains of the surfactants or the molecular weight of the ELP segment. In Chapter 5 we tested the properties of the pristine ELP-GFP material in the absence of surfactant. In particular, we investigated the cytotoxicity and the ability of the ELP-GFP to be internalized by mammalian cells. Thus, we generated positively charged variant of ELP-GFP fusion proteins and tested their cytotoxicity and uptake in mammalian cells. The ELP-GFP fusion proteins showed low cytotoxicity and six-fold enhanced cellular uptake compared to the GFP alone. Finally, the mechanism of cellular uptake was investigated.
24
1.8 Bibliography 1. Sengupta, A. Liquid Crystal Theory. 7–36 (2013). 2. Onsager, L. The effects of shape on the interaction of colloidal
particles. Ann. N. Y. Acad. Sci. 51, 627–659 (1949). 3. Hamley, I. W. Liquid crystal phase formation by biopolymers. Soft
Matter 6, 1863–1871 (2010). 4. Hamley, I. W. & Castelletto, V. Biological Soft Materials. Angew.
Chem. Int. Ed. 46, 4442–4455 (2007). 5. Rey, A. D., Herrera-Valencia, E. E. & Murugesan, Y. K. Structure and
dynamics of biological liquid crystals. Liq. Cryst. 41, 430–451 (2014). 6. Chung, W. J. et al. Biomimetic self-templating supramolecular
structures. Nature 478, 364–368 (2011). 7. Conway, K. A. et al. Acceleration of oligomerization, not fibrillization,
is a shared property of both alpha-synuclein mutations linked to early-onset Parkinson's disease: Implications for pathogenesis and therapy. Proc. Natl. Acad. Sci. 97, 571–576 (2000).
8. Koltover, I., Salditt, T., Rädler, J. O. & Safinya, C. R. An inverted hexagonal phase of cationic liposome-DNA complexes related to DNA release and delivery. Science 281, 78–81 (1998).
9. Waigh, T. A., Perry, P., Riekel, C., Gidley, M. J. & Donald, A. M. Chiral side-chain liquid-crystalline polymeric properties of starch. Macromolecules 31, 7980–7984 (1998).
10. Nakata, M. et al. End-to-End Stacking and Liquid Crystal Condensation of 6- to 20-Base Pair DNA Duplexes. Science 318, 1276–1279 (2007).
11. Livolant, F. & Bouligand, Y. Liquid crystalline phases given by helical biological polymers (DNA, PBLG and xanthan). Columnar textures. J. Phys. France 47, 1813–1827 (1986).
12. Earnshaw, W. C. & Casjens, S. R. DNA packaging by the double-stranded DNA bacteriophages. Cell 21, 319–331 (1980).
13. Livolant, F. Ordered phases of DNA in vivo and in vitro. Physica A 176, 117–137 (1991).
14. Giraud-Guille, M. M. Plywood structures in nature. Curr. Opin. Solid St. M. 3, 221–227 (1998).
15. Vollrath, F. & Knight, D. P. Liquid crystalline spinning of spider silk. Nature 410, 541–548 (2001).
25
16. Bouligand, Y. Liquid crystals and biological morphogenesis: Ancient and new questions. Comptes Rendus Chimie 11, 281–296 (2008).
17. Neville, A. C. A Pipe-Cleaner Molecular-Model for Morphogenesis of Helicoidal Plant-Cell Walls Based on Hemicellulose Complexity. J. Theor. Biol. 131, 243–254 (1988).
18. Rey, A. D. Liquid crystal models of biological materials and processes. Soft Matter 6, 3402 (2010).
19. Livolant, F. & Bouligand, Y. New observations on the twisted arrangement of dinoflagellate chromosomes. Chromosoma 68, 21–44 (1978).
20. Reich, Z., Wachtel, E. J. & Minsky, A. Liquid-crystalline mesophases of plasmid DNA in bacteria. Science 264, 1460–1463 (1994).
21. Cerritelli, M. Encapsidated Conformation of Bacteriophage T7 DNA. Cell 91, 271–280 (1997).
22. Roland, J. C., Vian, B. & Reis, D. Further Observations on Cell-Wall Morphogenesis and Polysaccharide Arrangement During Plant-Growth. Protoplasma 91, 125–141 (1977).
23. Gilbert, R.-D. & Patton, P. A. Liquid crystal formation in cellulose and cellulose derivatives. Prog. Polym. Sci. 9, 115–131 (1983).
24. Belamie, E., Davidson, P. & Giraud-Guille, M. M. Structure and chirality of the nematic phase in α-chitin suspensions. J. Phys. Chem. B 108, 14991–15000 (2004).
25. Jewell, S. A., Vukusic, P. & Roberts, N. W. Circularly polarized colour reflection from helicoidal structures in the beetle Plusiotis boucardi. New J. Phys. 9, 99–99 (2007).
26. Goodby, B. J. Liquid crystals and life. Liq. Cryst. 24, 25–38 (1998). 27. Giraud-Guille, M. M. Liquid Crystallinity in Condensed Type-I
Collagen Solutions - a Clue to the Packing of Collagen in Extracellular Matrices. J. Mol. Biol. 224, 861–873 (1992).
28. Knight, D. P. & Vollrath, F. Liquid crystals and flow elongation in a spider's silk production line. P. Roy. Soc. B-Biol. Sci. 266, 519–523 (1999).
29. Janmey, P. A. Mechanical properties of cytoskeletal polymers. Curr. Opin. Cell Biol. 3, 4–11 (1991).
30. Coppin, C. M. & Leavis, P. C. Quantitation of liquid-crystalline ordering in F-actin solutions. Biophys. J. 63, 794–807 (1992).
31. Knight, D. P. & Vollrath, F. Biological liquid crystal elastomers. Phil. Tr. Roy. Soc. B 357, 155–163 (2002).
26
32. Murthy, N. S. Liquid Crystallinity in Collagen Solutions and Magnetic Orientation of Collagen Fibrils. Biopolymers 23, 1261–1267 (1984).
33. Martin, R. et al. Liquid crystalline ordering of procollagen as a determinant of three-dimensional extracellular matrix architecture. J. Mol. Biol. 301, 11–17 (2000).
34. Giraud-Guille, M.-M., Mosser, G. & Belamie, E. Liquid crystallinity in collagen systems in vitro and in vivo. Curr. Opin. Colloid In. 13, 303–313 (2008).
35. Belamie, E., Mosser, G., Gobeaux, F. & Giraud-Guille, M.-M. Possible transient liquid crystal phase during the laying out of connective tissues: α-chitin and collagen as models. J. Phys-Condens. Mat. 18, 115-129 (2006).
36. Barnard, K. & Gathercole, L. J. Short and long range order in basement membrane type IV collagen revealed by enzymic and chemical extraction. Int. J. Biol. Macromol. 13, 359–365 (1991).
37. Kerkam, K., Viney, C., Kaplan, D. & Lombardi, S. Liquid Crystallinity of Natural Silk Secretions. Nature 349, 596–598 (1991).
38. Viney, C. & Bell, F. I. Inspiration versus duplication with biomolecular fibrous materials: learning nature‘s lessons without copying nature’s limitations. Curr. Opin. Solid St. M. 8, 165–171 (2004).
39. Käs, J. et al. F-actin, a model polymer for semiflexible chains in dilute, semidilute, and liquid crystalline solutions. Biophys. J. 70, 609–625 (1996).
40. Vonna, L., Limozin, L., Roth, A. & Sackmann, E. Single-filament dynamics and long-range ordering of semiflexible biopolymers under flow and confinement. Langmuir 21, 9635–9643 (2005).
41. Jones, J. B. & Safinya, C. R. Interplay between liquid crystalline and isotropic gels in self-assembled neurofilament networks. Biophys. J. 95, 823–835 (2008).
42. Corrigan, A. M., Müller, C. & Krebs, M. R. H. The Formation of Nematic Liquid Crystal Phases by Hen Lysozyme Amyloid Fibrils. J. Am. Chem. Soc. 128, 14740–14741 (2006).
43. Krebs, M. R. H., Bromley, E. H. C., Rogers, S. S. & Donald, A. M. The Mechanism of Amyloid Spherulite Formation by Bovine Insulin. Biophys. J. 88, 2013–2021 (2005).
44. Krebs, M. R. H. et al. The formation of spherulites by amyloid fibrils of bovine insulin. Proc. Natl. Acad. Sci. 101, 14420–14424 (2004).
27
45. Sagis, L. M. C., Veerman, C. & van der Linden, E. Mesoscopic Properties of Semiflexible Amyloid Fibrils. Langmuir 20, 924–927 (2004).
46. Yu, S., Soto, C. M. & Tirrell, D. A. Nanometer-scale smectic ordering of genetically engineered rodlike polymers: Synthesis and characterization of monodisperse derivatives of poly(gamma-benzyl alpha,L-glutamate). J. Am. Chem. Soc. 122, 6552–6559 (2000).
47. Tirrell, D. A. et al. Smectic ordering in solutions and films of a rod-like polymer owing to monodispersity of chain length : Article : Nature. Nature 389, 167–170 (1997).
48. Fiester, S. E., Jákli, A. & Woolverton, C. J. Liquid crystal properties of a self-assembling viral coat protein. Liq. Cryst. 38, 1153–1157 (2011).
49. Waigh, T. A. et al. Entanglement Coupling in Porcine Stomach Mucin. Langmuir 18, 7188–7195 (2002).
50. Viney, C., Huber, A. E. & Verdugo, P. Liquid-Crystalline Order in Mucus. Macromolecules 26, 852–855 (1993).
51. Robinson, C. Liquid-Crystalline Structures in Solutions of a Polypeptide. T. Faraday Soc. 52, 571–592 (1956).
52. Lin, J. P. Re-entrant isotropic transition of polypeptide liquid crystal. Polymer 38, 4837–4841 (1997).
53. Lin, J. P. Reentrant isotropic transition of polypeptide liquid crystal: effect of steric and orientation-dependent interactions. Polymer 39, 5495–5500 (1998).
54. Elliott, A. & Ambrose, E. J. Evidence of Chain Folding in Polypeptides and Proteins. Discuss. Faraday Soc. 246–251 (1950).
55. Watanabe, J. & Nagase, T. Thermotropic Polypeptides .5. Temperature-Dependence of Cholesteric Pitches Exhibiting a Cholesteric Sense Inversion. Macromolecules 21, 171–175 (1988).
56. Toriumi, H., Minakuchi, S., Uematsu, Y. & Uematsu, I. Helical Twisting Power of Poly(Gamma-Benzyl L-Glutamate) Liquid-Crystals in Mixed-Solvents. Polym. J. 12, 431–437 (1980).
57. Toriumi, H., Kusumi, Y., Uematsu, I. & Uematsu, Y. Thermally Induced Inversion of the Cholesteric Sense in Lyotropic Polypeptide Liquid-Crystals. Polym. J. 11, 863–869 (1979).
58. Abe, A., Hiraga, K., Imada, Y., Hiejima, T. & Furuya, H. Screw‐sense inversion characteristic of α‐helical poly (β‐p‐chlorobenzyl L‐aspartate) and comparison with other related polyaspartates. Pept. Sci. 80, 249–257 (2005).
28
59. Abe, A., Furuya, H. & Okamoto, S. Spatial configurations, transformation, and reorganization of mesophase structures of polyaspartates - A highly intelligent molecular system. Biopolymers 43, 405–412 (1997).
60. Aggeli, A. et al. Hierarchical self-assembly of chiral rod-like molecules as a model for peptide beta-sheet tapes, ribbons, fibrils, and fibers. Proc. Natl. Acad. Sci. 98, 11857–11862 (2001).
61. Han, T. H. et al. Liquid Crystalline Peptide Nanowires. Adv. Mater. 19, 3924–3927 (2007).
62. Krysmann, M. J. et al. Self-assembly of Peptide nanotubes in an organic solvent. Langmuir 24, 8158–8162 (2008).
63. Castelletto, V., Hamley, I. W., Hule, R. A. & Pochan, D. Helical‐Ribbon Formation by a β‐Amino Acid Modified Amyloid β‐Peptide Fragment. Angew. Chem. Int. Ed. Engl. 48, 2317–2320 (2009).
64. Bucak, S., Cenker, C., Nasir, I., Olsson, U. & Zackrisson, M. Peptide Nanotube Nematic Phase. Langmuir 25, 4262–4265 (2009).
65. Pomerantz, W. C. et al. Nanofibers and Lyotropic Liquid Crystals from a Class of Self‐Assembling β‐Peptides. Angew. Chem. 120, 1261–1264 (2008).
66. Kim, K. T., Park, C., Kim, C., Winnik, M. A. & Manners, I. Self-assembly of dendron-helical polypeptide copolymers: organogels and lyotropic liquid crystals. Chem. Commun. 13, 1372–1374 (2006).
67. Tan, M. M., Corley, C. A. & Stevenson, C. L. Effect of gelation on the chemical stability and conformation of leuprolide. Pharm. Res. 15, 1442–1448 (1998).
68. Powell, M. F., Sanders, L. M., Rogerson, A. & Si, V. Parenteral Peptide Formulations: Chemical and Physical Properties of Native Luteinizing Hormone-Releasing Hormone (LHRH) and Hydrophobic Analogues in Aqueous Solution. Pharm. Res. 08, 1258–1263 (1991).
69. Powell, M. F., Fleitman, J., Sanders, L. M. & Si, V. C. Peptide Liquid Crystals: Inverse Correlation of Kinetic Formation and Thermodynamic Stability in Aqueous Solution. Pharm. Res. 11, 1352–1354 (1994).
70. Lechuga Ballesteros, D., Abdul Fattah, A., Stevenson, C. L. & Bennett, D. B. Properties and stability of a liquid crystal form of cyclosporine—the first reported naturally occurring peptide that exists as a thermotropic liquid crystal. J. Pharm. Sci. 92, 1821–1831 (2003).
29
71. Canilho, N., Scholl, M., Klok, H.-A. & Mezzenga, R. Thermotropic Ionic Liquid Crystals via Self-Assembly of Cationic Hyperbranched Polypeptides and Anionic Surfactants. Macromolecules 40, 8374–8383 (2007).
72. Martin, R., Waldmann, L. & Kaplan, D. L. Supramolecular assembly of collagen triblock peptides. Biopolymers 70, 435–444 (2003).
73. Schlaad, H. Solution properties of polypeptide-based copolymers. 53–73 (2006).
74. Douy, A. & Gallot, B. Block copolymers with a polyvinyl and a polypeptide block: factors governing the folding of the polypeptide chains. Polymer 23, 1039–1044 (1982).
75. Billot, J.-P., Douy, A. & Gallot, B. Synthesis and structural study of block copolymers with a hydrophobic polyvinyl block and a hydrophilic polypeptide block: Copolymers polystyrene/poly(L-lysine) and polybutadiene/poly(L-lysine) - Billot - 2003 - Die Makromolekulare Chemie - Wiley Online Library. Makromol. Chem. 177, 1889–1893 (1976).
76. Minich, E. A., Nowak, A. P., Deming, T. J. & Pochan, D. J. Rod–rod and rod–coil self-assembly and phase behavior of polypeptide diblock copolymers. Polymer 45, 1951–1957 (2004).
77. Hamley, I. W., Krysmann, M. J., Castelletto, V. & Noirez, L. Multiple Lyotropic Polymorphism of a Poly(ethylene glycol)-Peptide Conjugate in Aqueous Solution. Adv. Mater. 20, 4394–4397 (2008).
78. Perriman, A. W., Cölfen, H., Hughes, R. W., Barrie, C. L. & Mann, S. Solvent-free protein liquids and liquid crystals. Angew. Chem. Int. Ed. Engl. 48, 6242–6246 (2009).
79. Besseau, L., Coulomb, B., Lebreton-Decoster, C. & Giraud-Guille, M. M. Production of ordered collagen matrices for three-dimensional cell culture. Biomaterials 23, 27–36 (2002).
80. Eglin, D., Mosser, G., Giraud-Guille, M.-M., Livage, J. & Coradin, T. Type I collagen, a versatile liquid crystal biological template for silica structuration from nano- to microscopic scales. Soft Matter 1, 129–131 (2005).
81. Woltman, S. J., Jay, G. D. & Crawford, G. P. Liquid-crystal materials find a new order in biomedical applications. Nat. Mater. 6, 929–938 (2007).
82. Lowe, A. M. & Abbott, N. L. Liquid Crystalline Materials for Biological Applications. Chem. Mater. 24, 746–758 (2012).
30
83. Park, J.-S. & Abbott, N. L. Ordering transitions in thermotropic liquid crystals induced by the interfacial assembly and enzymatic processing of oligopeptide amphiphiles. Adv. Mater. 20, 1185–1190 (2008).
84. Park, J.-S. et al. Formation of Oligopeptide-Based Polymeric Membranes at Interfaces between Aqueous Phases and Thermotropic Liquid Crystals. Chem. Mater. 18, 6147–6151 (2006).
85. Lundgren, J. S., Watkins, A. N., Racz, D. & Ligler, F. S. A liquid crystal pixel array for signal discrimination in array biosensors. Biosens. Bioelectron. 15, 417–421 (2000).
86. Ha, S.-W., Tonelli, A. E. & Hudson, S. M. Structural Studies of BombyxmoriSilk Fibroin during Regeneration from Solutions and Wet Fiber Spinning. Biomacromolecules 6, 1722–1731 (2005).
87. Liivak, O., Blye, A., Shah, N. & Jelinski, L. W. A microfabricated wet-spinning apparatus to spin fibers of silk proteins. Structure-property correlations. Macromolecules 31, 2947–2951 (1998).
88. Luzzati, V. & Husson, F. The structure of the liquid-crystalline phasis of lipid-water systems. J. Cell Biol. 12, 207–219 (1962).
89. Hamley, I. W. et al. Nematic and Columnar Ordering of a PEG-Peptide Conjugate in Aqueous Solution. Chem. Eur. J. 14, 11369–11375 (2008).
90. Gallot, B. Comb-like and block liquid crystalline polymers for biological applications. Prog. Polym. Sci. 21, 1035–1088 (1996).
91. Cai, C., Lin, J., Zhuang, Z. & Zhu, W. Ordering of Polypeptides in Liquid Crystals, Gels and Micelles. Controlled Polymerization and Polymeric Structures: Flow Microreactor Polymerization, Micelles Kinetics, Polypeptide Ordering, Light Emitting Nanostructures 259, 159–199 (2013).