Page 1
University of Tennessee, Knoxville University of Tennessee, Knoxville
TRACE: Tennessee Research and Creative TRACE: Tennessee Research and Creative
Exchange Exchange
Masters Theses Graduate School
8-2015
Dietary Supplementation of Choline and Betaine in Heat-Stressed Dietary Supplementation of Choline and Betaine in Heat-Stressed
Broilers Broilers
Kouassi Remi Kpodo University of Tennessee - Knoxville, [email protected]
Follow this and additional works at: https://trace.tennessee.edu/utk_gradthes
Part of the Poultry or Avian Science Commons
Recommended Citation Recommended Citation Kpodo, Kouassi Remi, "Dietary Supplementation of Choline and Betaine in Heat-Stressed Broilers. " Master's Thesis, University of Tennessee, 2015. https://trace.tennessee.edu/utk_gradthes/3489
This Thesis is brought to you for free and open access by the Graduate School at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Masters Theses by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected] .
Page 2
To the Graduate Council:
I am submitting herewith a thesis written by Kouassi Remi Kpodo entitled "Dietary
Supplementation of Choline and Betaine in Heat-Stressed Broilers." I have examined the final
electronic copy of this thesis for form and content and recommend that it be accepted in partial
fulfillment of the requirements for the degree of Master of Science, with a major in Animal
Science.
Michael O. Smith, Major Professor
We have read this thesis and recommend its acceptance:
Brynn Voy, Agustin Rius, Arnold Saxton
Accepted for the Council:
Carolyn R. Hodges
Vice Provost and Dean of the Graduate School
(Original signatures are on file with official student records.)
Page 3
Dietary Supplementation of Choline and Betaine in Heat-Stressed Broilers
A Thesis Presented for the
Master of Science
Degree
The University of Tennessee, Knoxville
Kouassi Remi Kpodo
August 2015
Page 4
ii
Copyright © 2015 by Kouassi Remi Kpodo
All rights reserved.
Page 5
iii
Dedication
This thesis is dedicated to my family, my wife, brothers and sisters, and my friends whose
encouragement and moral support gave me the strength to pursue the Master’s Degree.
Page 6
iv
Acknowledgements
I would like to express my sincere gratitude to Dr. Michael Smith, my graduate mentor
for giving me the opportunity to pursue my Master’s Degree under his guidance, all his teaching
and transfer of knowledge and for his investment in helping me find a job after graduation. I
would like to thank Dr. Arnold Saxton, Dr. Brynn Voy and Dr. Agustin Rius for serving on my
graduate committee. Special thanks to Dr. Henry Kattesh for all his help with corticosterone
analysis, and Dr. Virginia Artegoitia for her help with choline and choline metabolites
determination.
Thanks to my fellow graduate students Rekek Negga, Xiang Liu and Tania Torchon for
their assistance during the live bird trial, and especially to Ronique Beckford for all his help and
constructive discussions about the experiment and the analysis of statistical outputs. Thanks also
to Eddie Jarboe and Linda Miller for all their help with taking blood samples as well as to
Barbara Gillespie and Sierra Lockwood for their help with corticosterone analysis, and JARTU
staffs for helping me raise the birds.
I would not finish without expressing my appreciation to Balchem Corporation and the
University of Tennessee AgResearch which partially provided the funding and materials for this
project. Finally, thanks to my family and friends whose support and encouragement gave me the
strength to finish the program.
Page 7
v
Abstract
A study was conducted to determine the effects of supplemental choline (CHO) and betaine
(BET) on broiler performance, carcass characteristics, corticosterone levels, immune organ
weights, intestinal morphology and choline metabolites under thermoneutral (TN: 23.9oC) and
heat stress (HS: 28-36oC) conditions. Eight hundred day-old chicks (400 per temperature
treatment) were assigned in groups of 10 to each of five dietary treatments: 1 (CON) the basal
diet; 2 (CHO500), basal diet plus 500 ppm methyl equivalents added CHO; 3 (CHO1000), basal
diet plus 1000 ppm methyl equivalents added CHO; 4 (BET500), basal diet plus 500 ppm methyl
equivalents added BET and 5 (BET1000), basal diet plus 1000 ppm methyl equivalents added
BET. Pen feed intake and body weights were recorded weekly. Foot pad dermatitis (FPD) was
assessed and litter samples were collected on day 42. On days 22 and 45, blood samples were
collected from eight birds per treatment to determine corticosterone and choline metabolites
concentrations. On day 52, birds were processed, spleen, bursa and thymus were weighed. Liver
and intestinal samples were collected for CHO and CHO metabolites determination and
histology (intestine). Breast color was measured and drip loss evaluated 4 and 7 days post
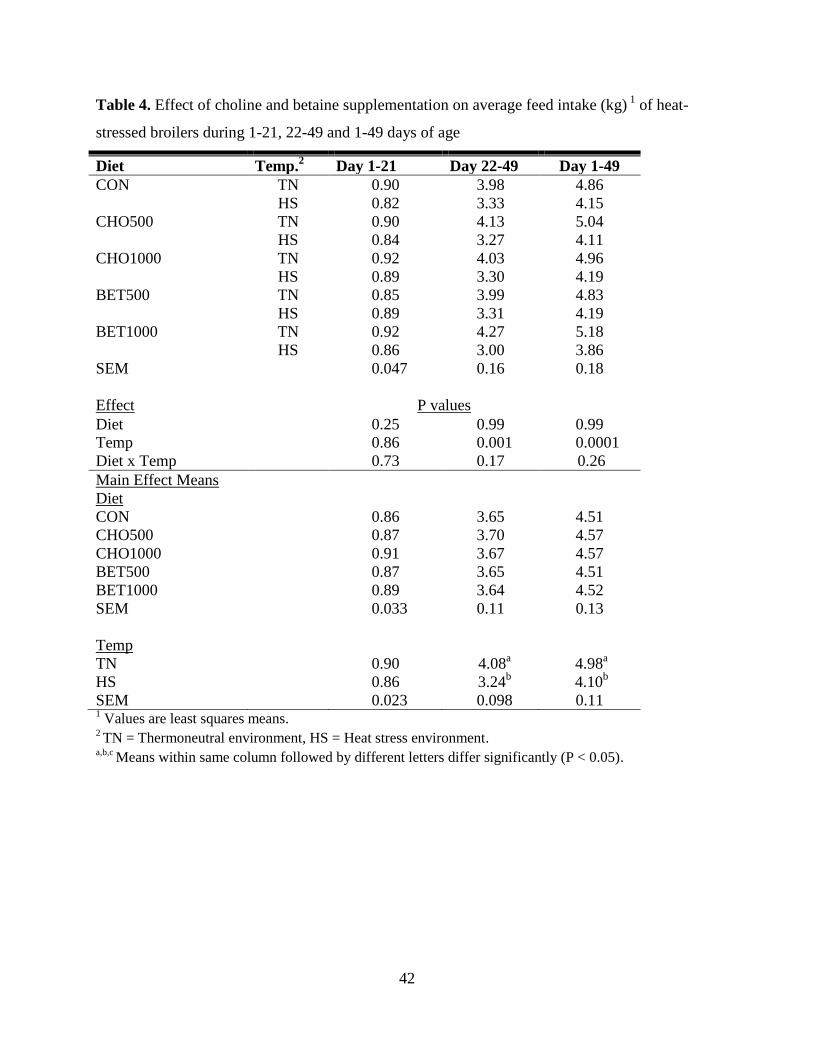
slaughter. Results showed that HS birds consumed 20.59% less feed, gained 23.35% less weight
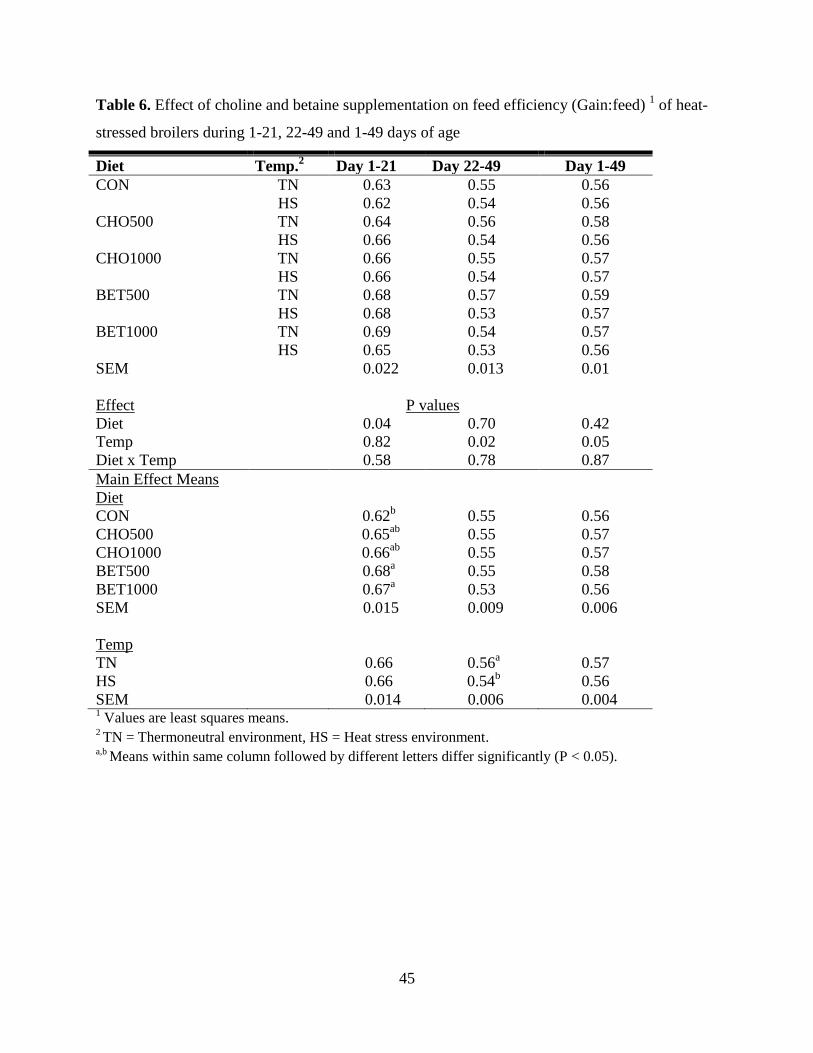
and had lower feed efficiency compared to TN birds (P < 0.05) during days 22-49. Foot pad
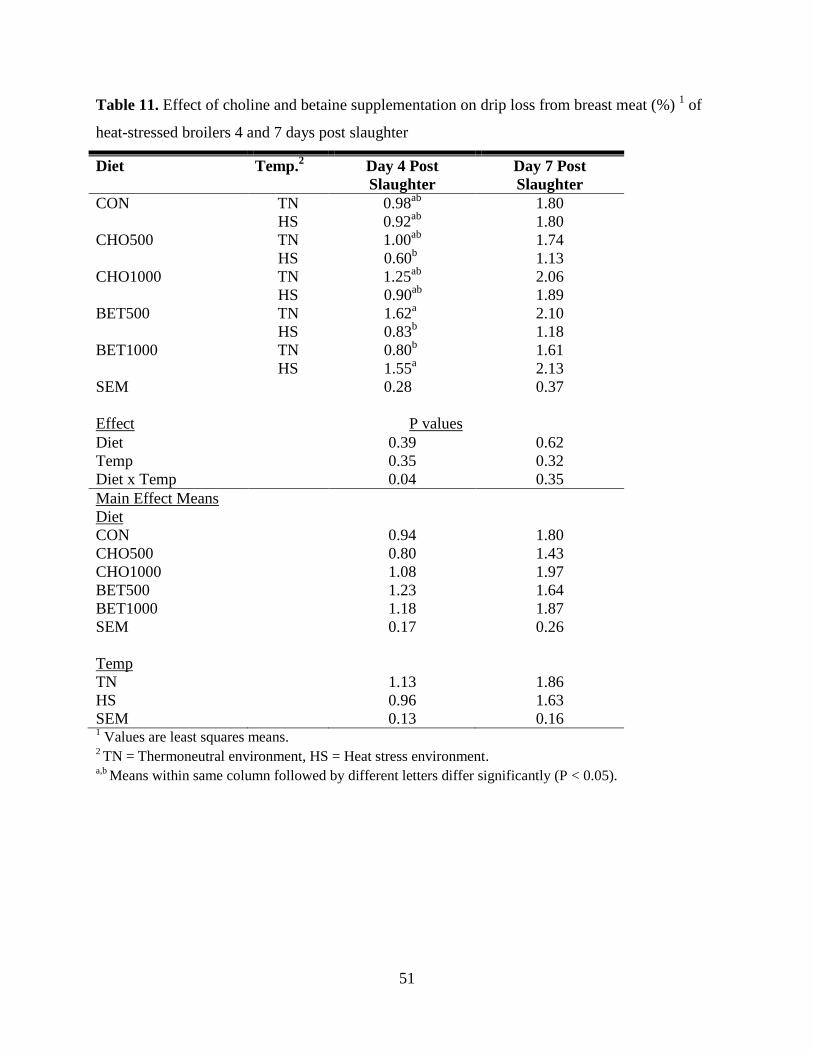
dermatitis was decreased (P = 0.003) by CHO500 and BET1000. Drip loss 4 days post slaughter
was reduced (P = 0.04) by CHO500 (0.60%) in HS and BET1000 (0.83%) in TN birds. On day
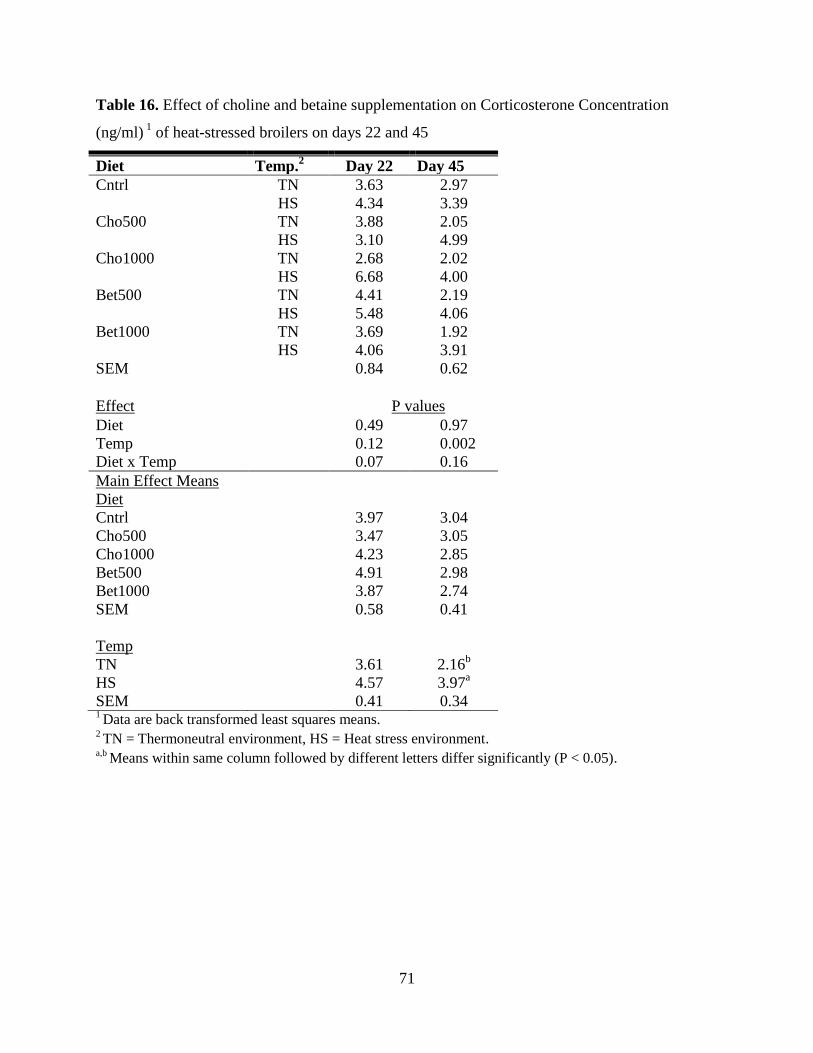
45, HS increased (P = 0.02) corticosterone levels (3.61 ng/ml HS vs 2.16 ng/ml TN). Thymus
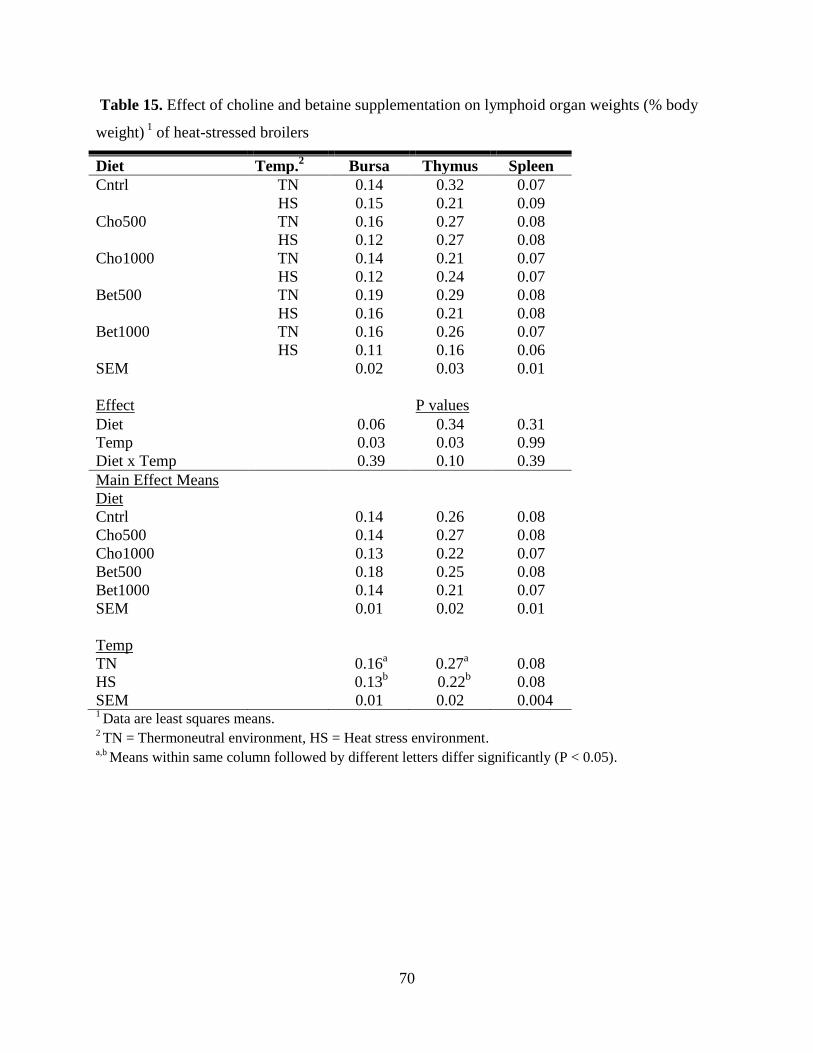
and bursa weights were reduced (P = 0.03) by HS. There was a diet X temperature interaction for
liver BET concentration (P = 0.03) whereas intestinal BET was increased only by diet (P = 0.02).
Liver CHO and glycerophosphocholine were increased by temperature (P = 0.01). Dietary
Page 8
vi
supplementation with CHO and BET did not negate the negative impacts of HS on broiler
performance and immune system. However, they reduced FDP as well as drip loss.
Page 9
vii
Table of Contents
Chapter I: Introduction .................................................................................................................... 1
References ....................................................................................................................................... 5
Chapter II: Literature Review ......................................................................................................... 7
Heat Stress in Broilers ................................................................................................................. 8
Foot Pad Dermatitis ................................................................................................................... 10
Impact of Heat Stress on Intestinal Morphology ...................................................................... 12
Heat Stress and the Immune System ......................................................................................... 13
Choline ...................................................................................................................................... 14
Betaine ....................................................................................................................................... 16
Interrelation of Choline, Betaine and Methionine Metabolism................................................. 18
References ..................................................................................................................................... 21
Chapter III: Performance of heat-stressed broilers supplemented with dietary choline and betaine
....................................................................................................................................................... 29
Abstract ..................................................................................................................................... 30
Introduction ............................................................................................................................... 31
Materials and Methods .............................................................................................................. 33
Birds and Housing ................................................................................................................. 33
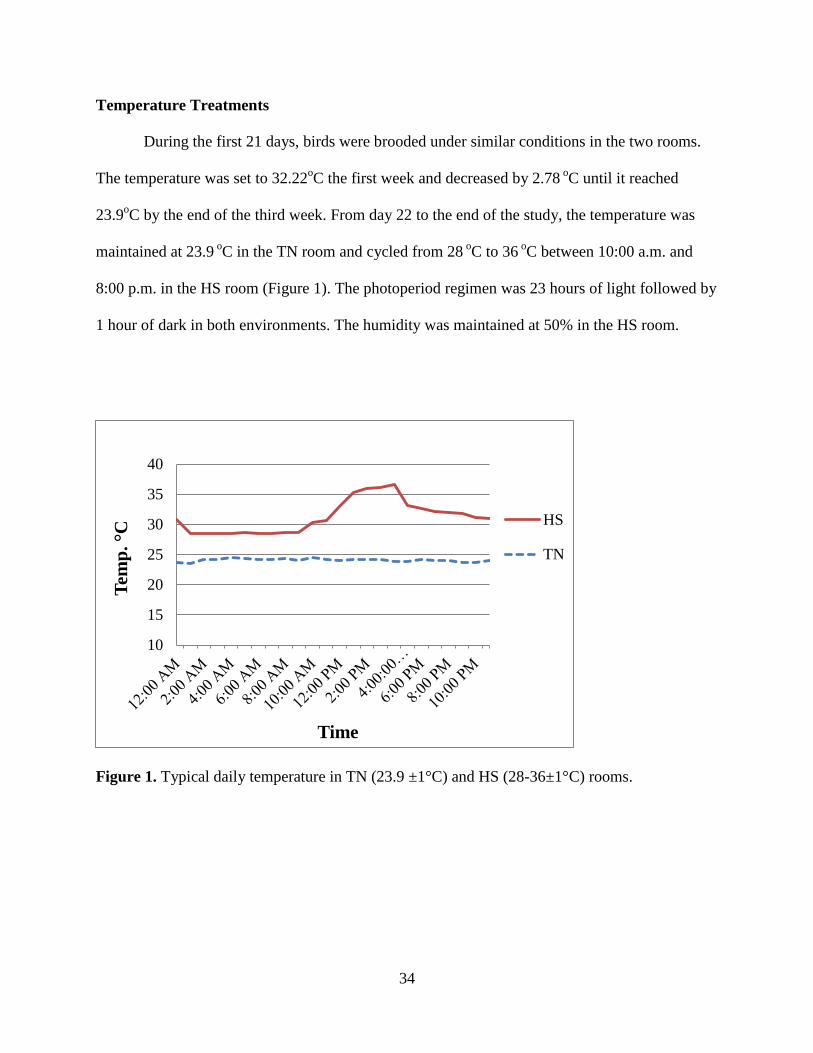
Temperature Treatments ........................................................................................................ 34
Dietary Treatments ................................................................................................................ 35
Growth Performance .............................................................................................................. 35
Litter Samples and Foot Pad Dermatitis ................................................................................ 35
Processing of Birds ................................................................................................................ 40
Drip Loss ............................................................................................................................... 40
Breast Meat Color .................................................................................................................. 40
Statistical Analysis ................................................................................................................ 41
Results ....................................................................................................................................... 41
Growth Performance .............................................................................................................. 41
Litter Moisture ....................................................................................................................... 43
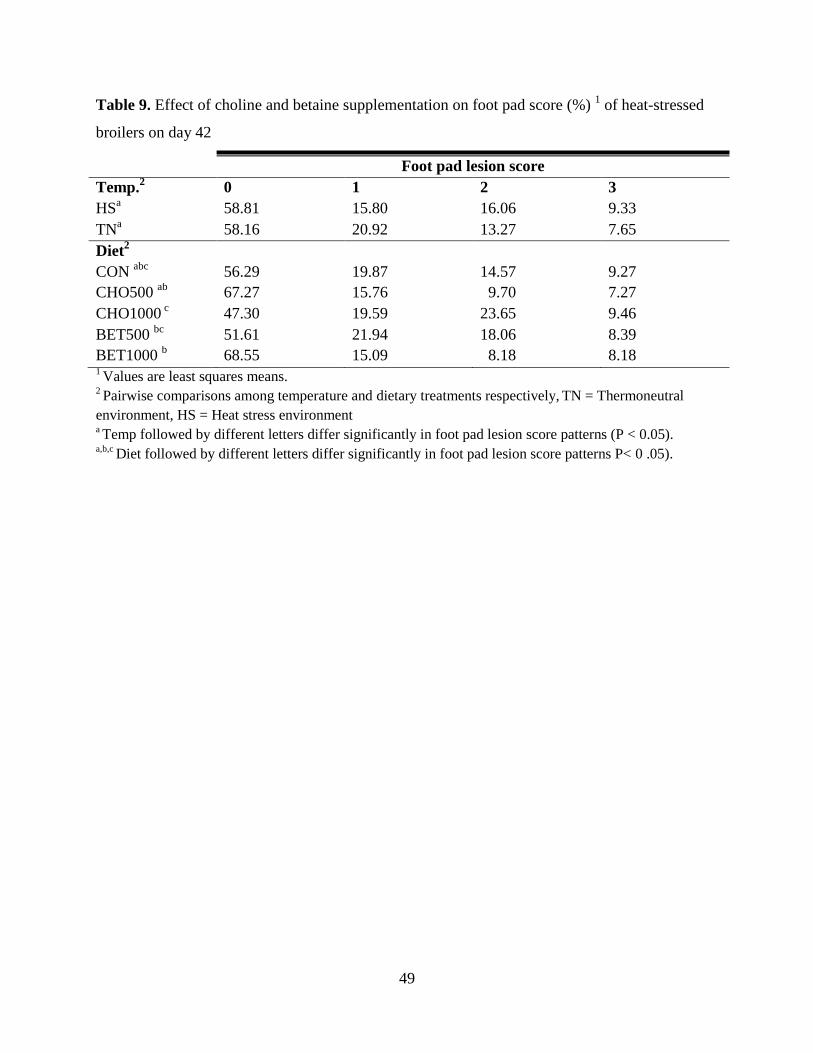
Foot Pad Dermatitis ............................................................................................................... 48
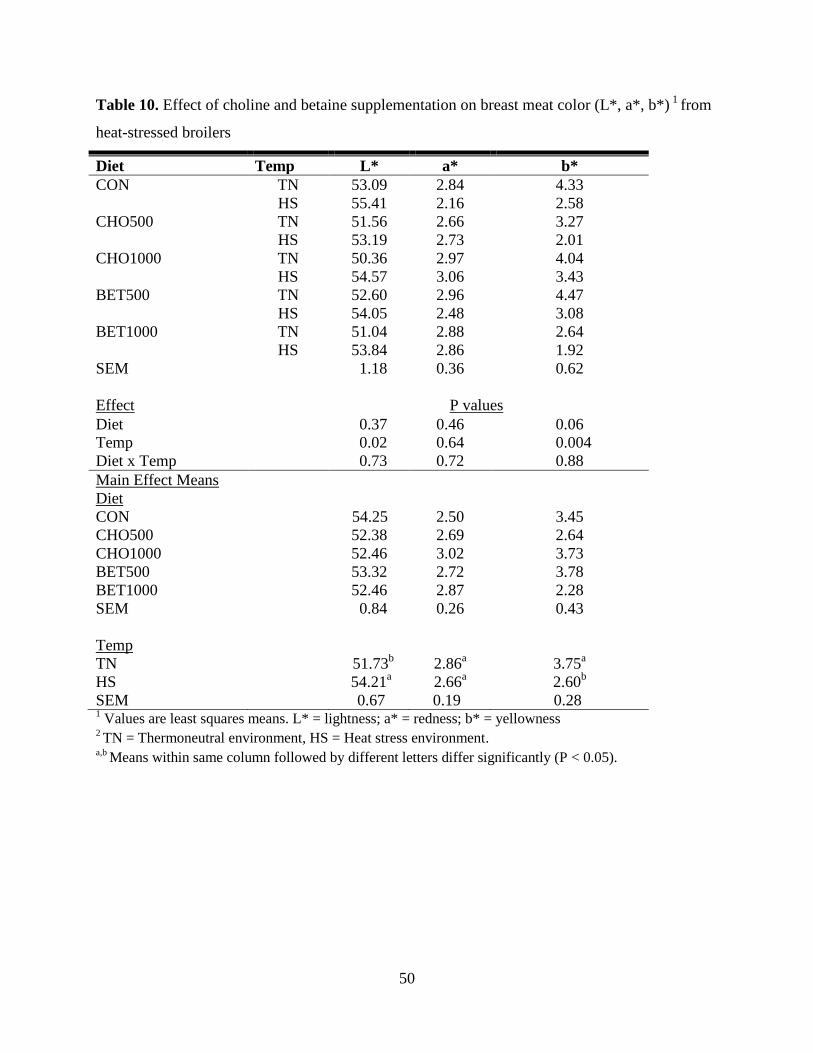
Breast Meat Color .................................................................................................................. 48
Page 10
viii
Breast Meat Drip Loss ........................................................................................................... 48
Discussion ................................................................................................................................. 52
References ..................................................................................................................................... 55
Chapter IV: Effect of dietary choline and betaine on corticosterone levels and immune organ
weights of heat-stressed broilers ................................................................................................... 58
Abstract ..................................................................................................................................... 59
Introduction ............................................................................................................................... 60
Materials and Methods .............................................................................................................. 62
Birds and Housing ................................................................................................................. 62
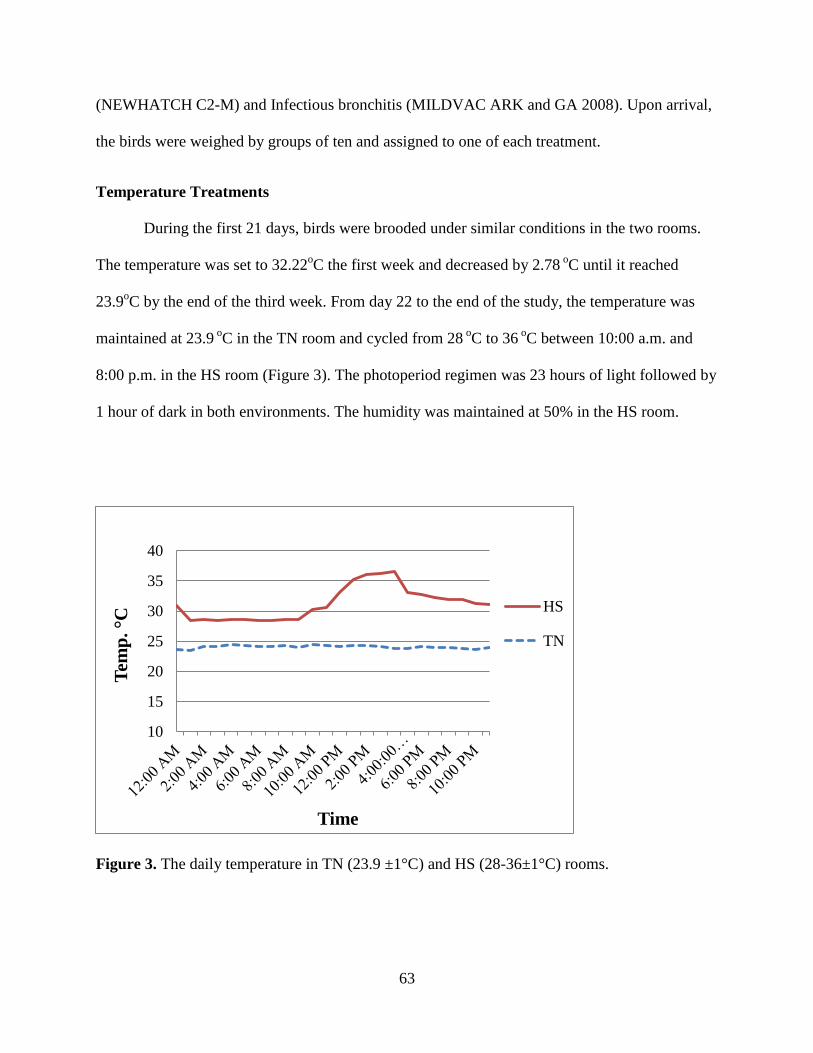
Temperature Treatments ........................................................................................................ 63
Dietary Treatments ................................................................................................................ 64
Blood Collection .................................................................................................................... 64
Processing of Birds ................................................................................................................ 64
Corticosterone Radioimmunoassay ....................................................................................... 68
Statistical Analysis ................................................................................................................ 68
Results ....................................................................................................................................... 69
Bursa of Fabricius, Spleen and Thymus Weights .................................................................. 69
Plasma Corticosterone ........................................................................................................... 69
Discussion ................................................................................................................................. 72
References ..................................................................................................................................... 74
Chapter V: Effect of dietary choline and betaine on choline metabolites and intestinal
morphology of heat-stressed broilers ............................................................................................ 76
Abstract ......................................................................................................................................... 77
Introduction ............................................................................................................................... 78
Materials and Methods .............................................................................................................. 81
Birds and Housing ................................................................................................................. 81
Temperature Treatments ........................................................................................................ 81
Dietary Treatments ................................................................................................................ 83
Blood Collection .................................................................................................................... 83
Processing of Birds ................................................................................................................ 83
Intestinal Histology................................................................................................................ 87
Page 11
ix
Choline Metabolites Analysis by HILC-MS/MS .................................................................. 87
Statistical Analysis ................................................................................................................ 88
Results ....................................................................................................................................... 88
Intestinal Histology................................................................................................................ 88
Intestinal Choline and Choline Metabolites .......................................................................... 88
Liver Choline and Choline Metabolites................................................................................. 91
Plasma Choline and Choline Metabolites .............................................................................. 93
Discussion ................................................................................................................................. 95
References ..................................................................................................................................... 98
Chapter VI: Summary and Conclusion ....................................................................................... 101
Vita .............................................................................................................................................. 103
Page 12
x
List of Tables
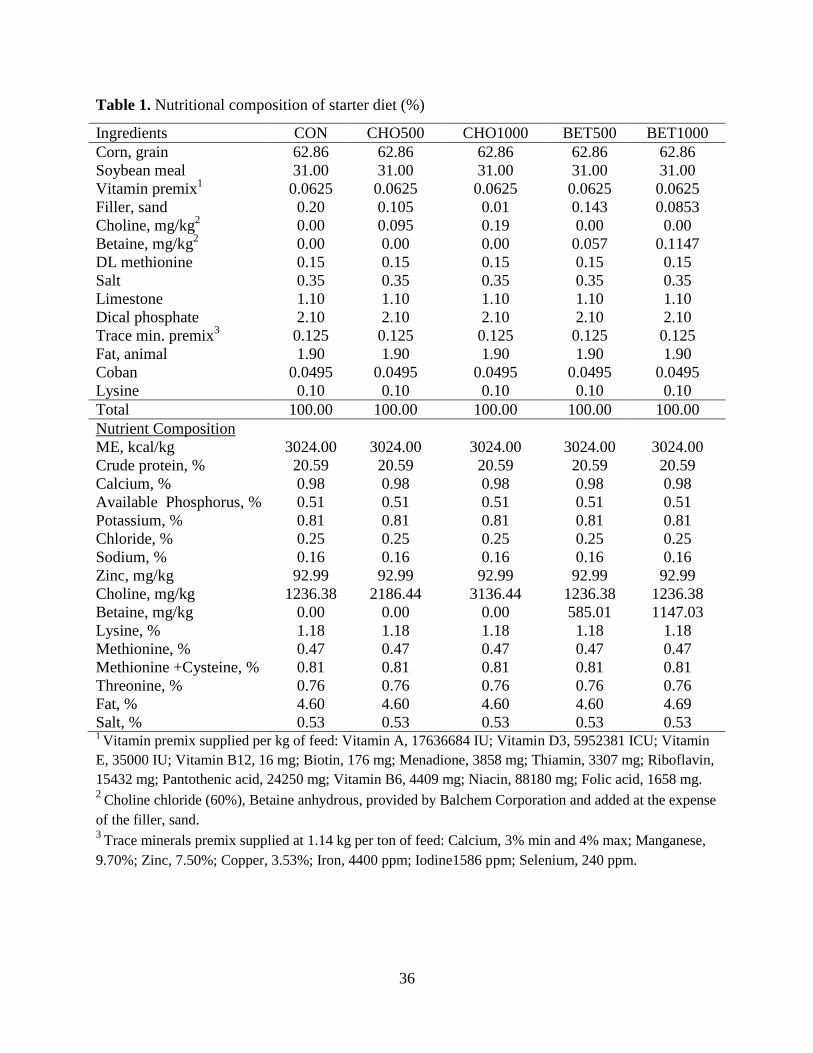
Table 1. Nutritional composition of starter diet (%) ..................................................................... 36
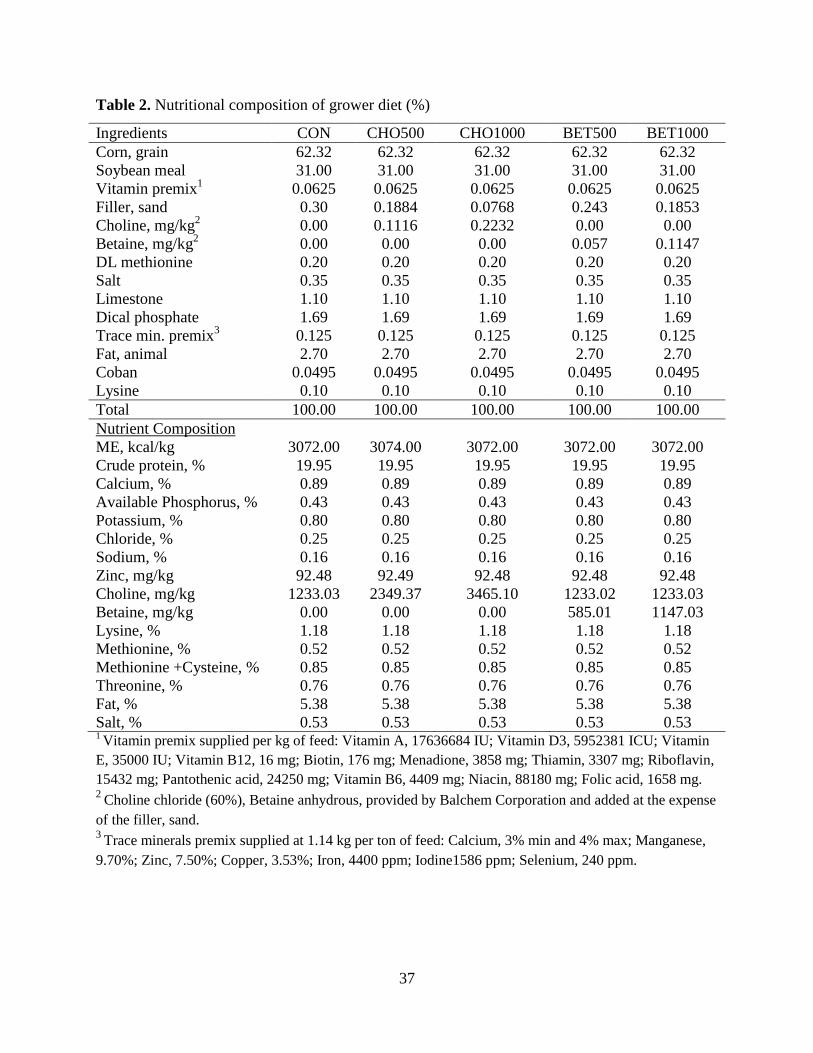
Table 2. Nutritional composition of grower diet (%) ................................................................... 37
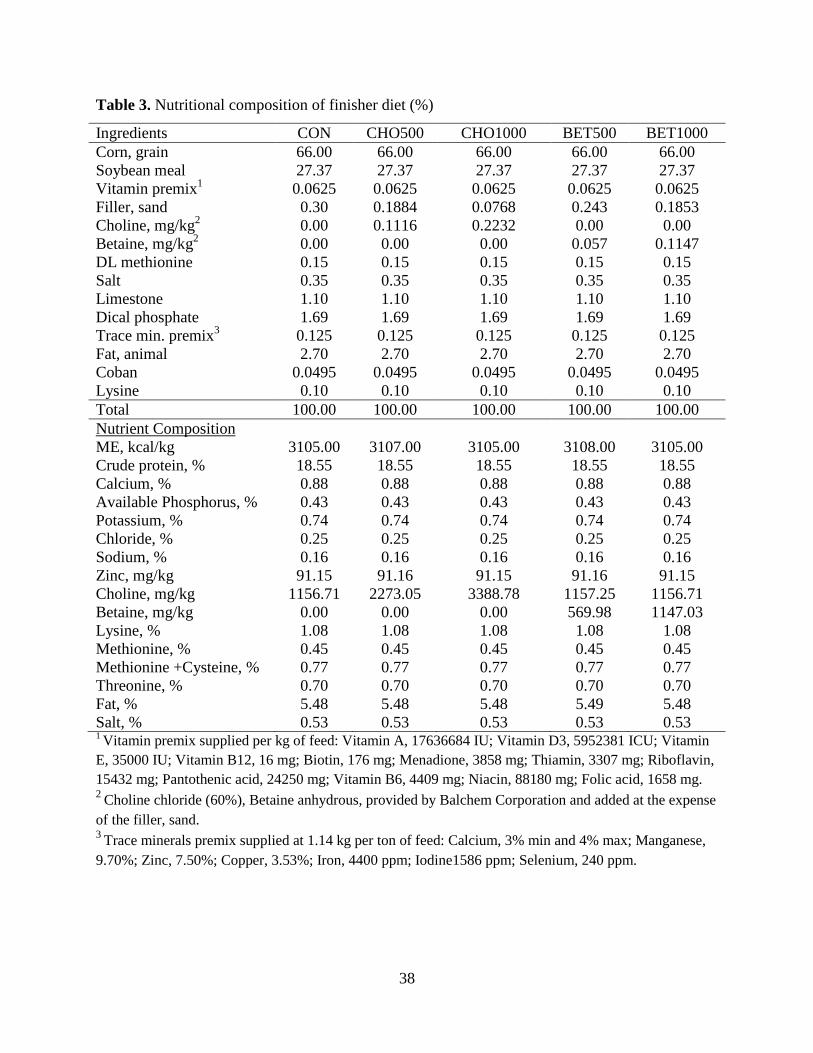
Table 3. Nutritional composition of finisher diet (%)................................................................... 38
Table 4. Effect of choline and betaine supplementation on average feed intake (kg) 1
of heat-
stressed broilers during 1-21, 22-49 and 1-49 days of age ........................................................... 42
Table 5. Effect of choline and betaine supplementation on average body weight gain (kg) 1
of
heat-stressed broilers during 1-21, 22-49 and 1-49 days of age ................................................... 44
Table 6. Effect of choline and betaine supplementation on feed efficiency (Gain:feed) 1 of heat-
stressed broilers during 1-21, 22-49 and 1-49 days of age ........................................................... 45
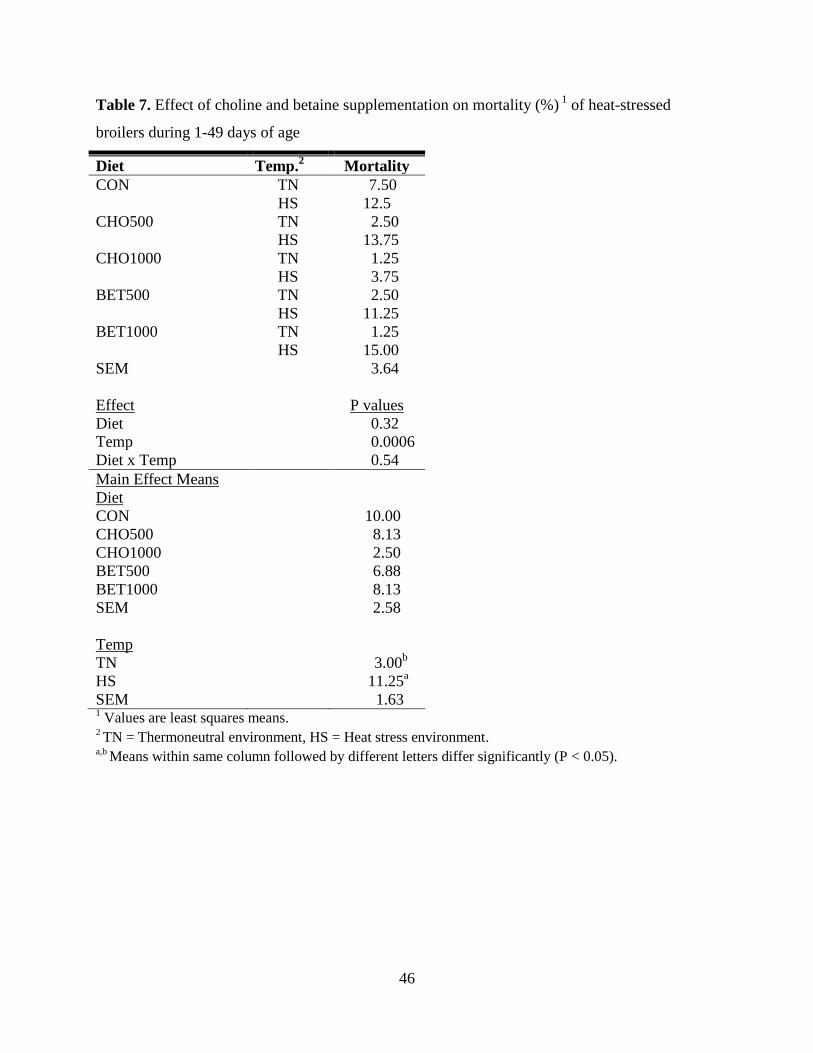
Table 7. Effect of choline and betaine supplementation on mortality (%) 1
of heat-stressed
broilers during 1-49 days of age ................................................................................................... 46
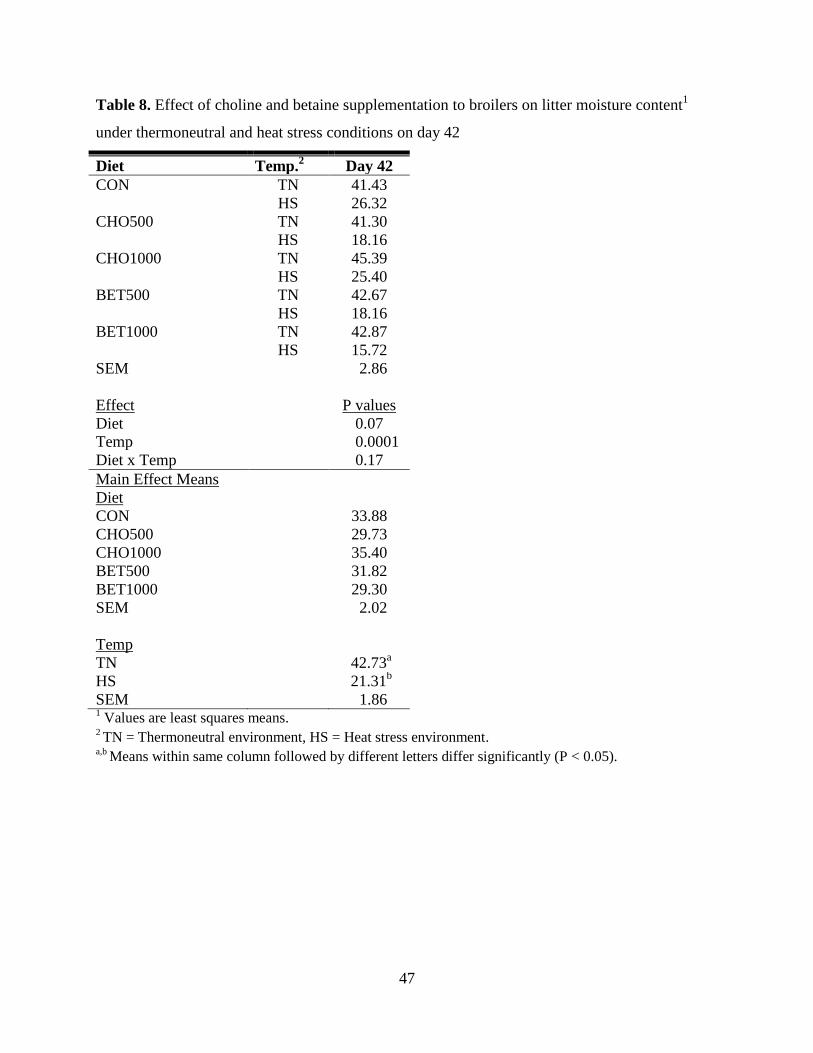
Table 8. Effect of choline and betaine supplementation to broilers on litter moisture content1
under thermoneutral and heat stress conditions on day 42 ........................................................... 47
Table 9. Effect of choline and betaine supplementation on foot pad score (%) 1 of heat-stressed
broilers on day 42.......................................................................................................................... 49
Table 10. Effect of choline and betaine supplementation on breast meat color (L*, a*, b*) 1
from
heat-stressed broilers ..................................................................................................................... 50
Table 11. Effect of choline and betaine supplementation on drip loss from breast meat (%) 1 of
heat-stressed broilers 4 and 7 days post slaughter ........................................................................ 51
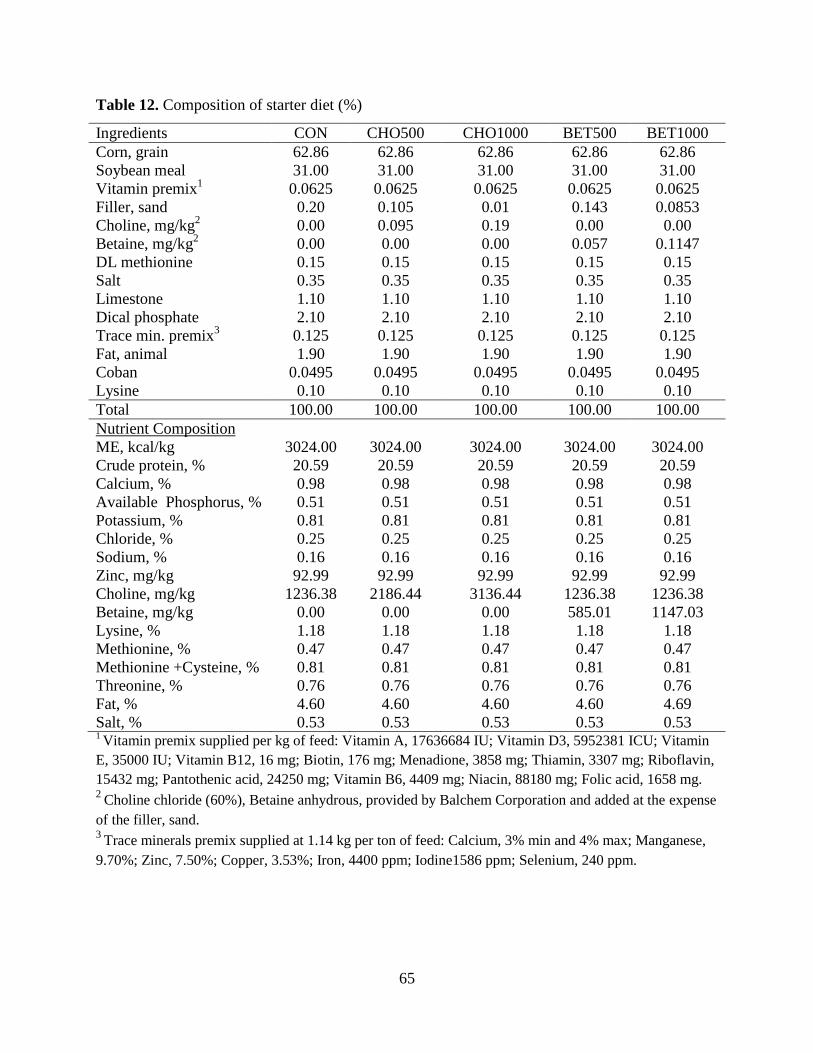
Table 12. Composition of starter diet (%) .................................................................................... 65
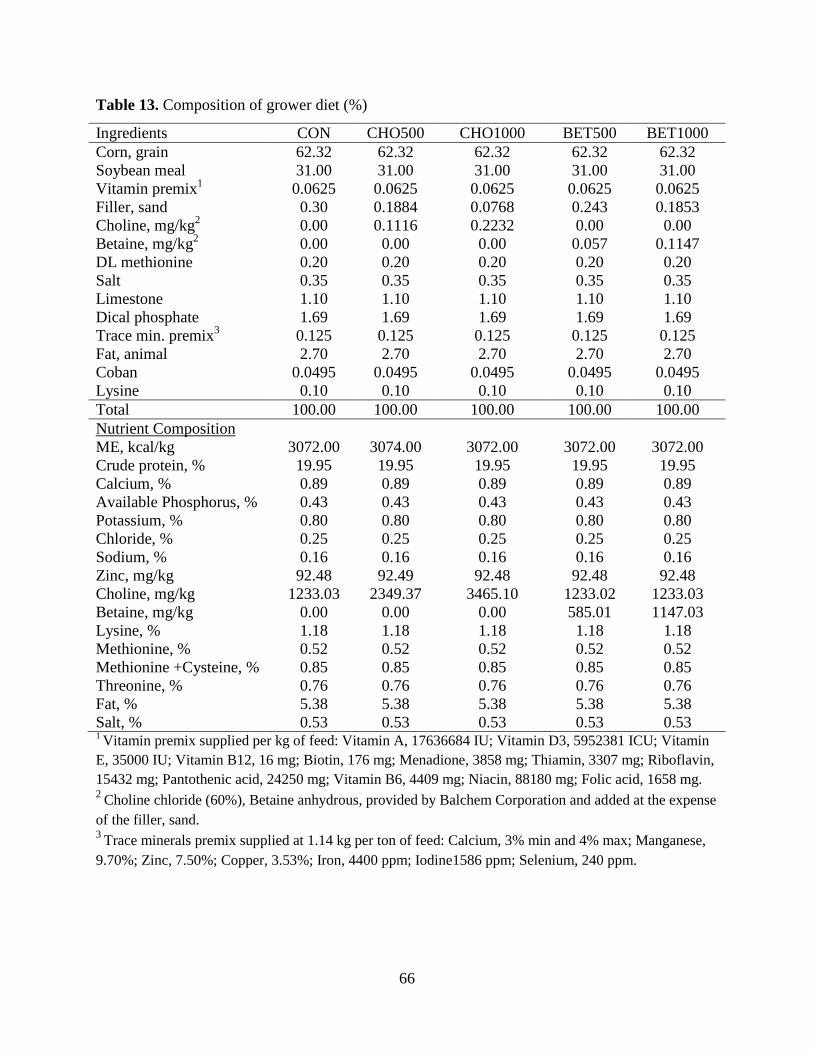
Table 13. Composition of grower diet (%) ................................................................................... 66
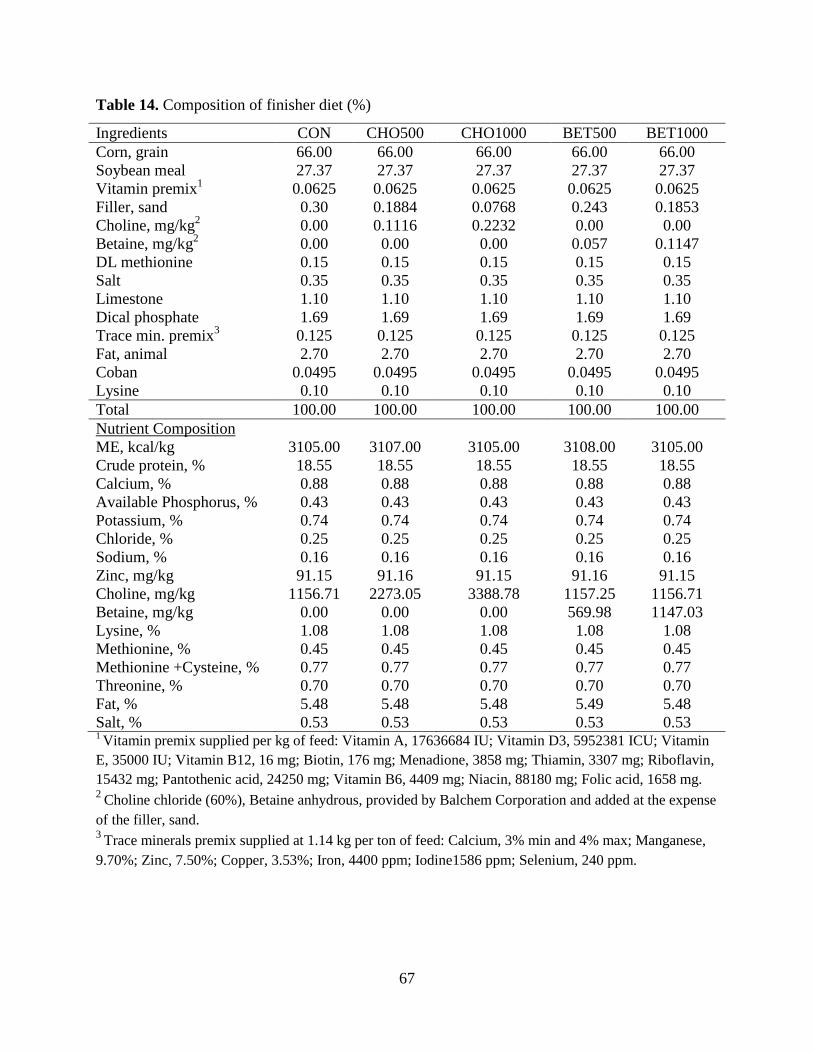
Table 14. Composition of finisher diet (%) .................................................................................. 67
Table 15. Effect of choline and betaine supplementation on lymphoid organ weights (% body
weight) 1
of heat-stressed broilers ................................................................................................. 70
Table 16. Effect of choline and betaine supplementation on Corticosterone Concentration (ng/ml)
1 of heat-stressed broilers on days 22 and 45 ................................................................................ 71
Table 17. Temp X Day interaction on corticosterone concentration (ng/ml) 1
of heat-stressed
broilers .......................................................................................................................................... 72
Table 18. Composition of experimental diet for starter period (%) .............................................. 84
Table 19. Composition of experimental diet for grower period (%) ............................................ 85
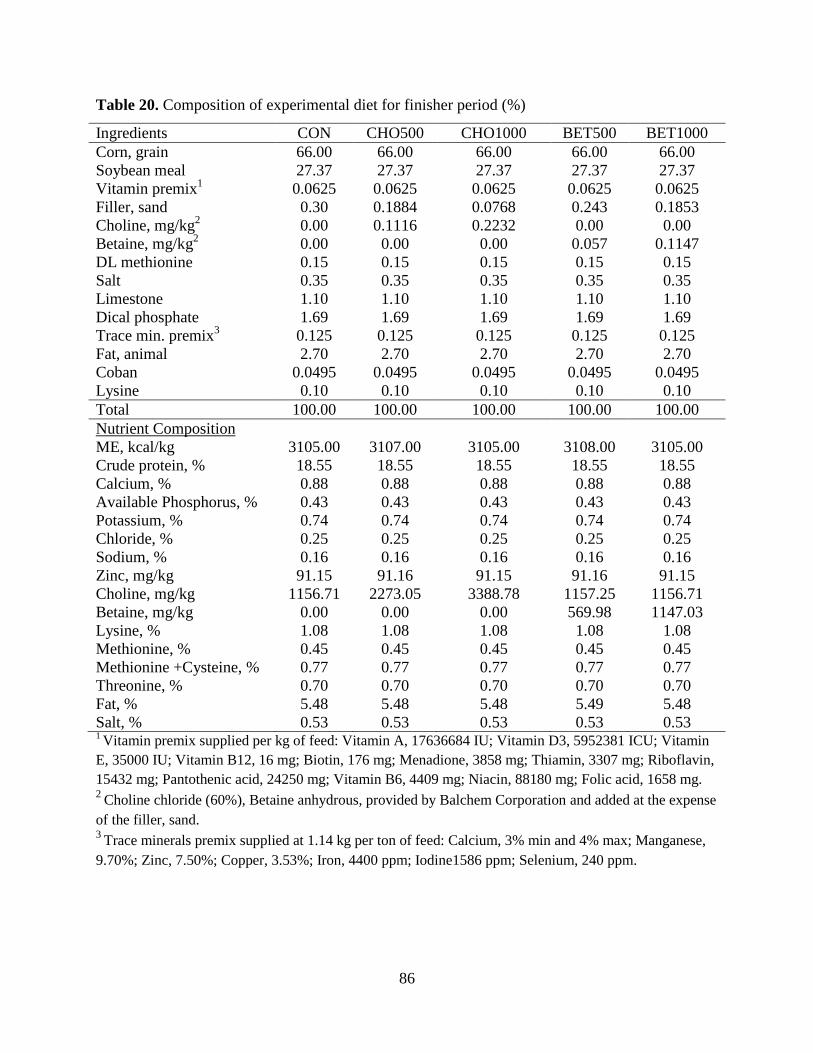
Table 20. Composition of experimental diet for finisher period (%) ........................................... 86
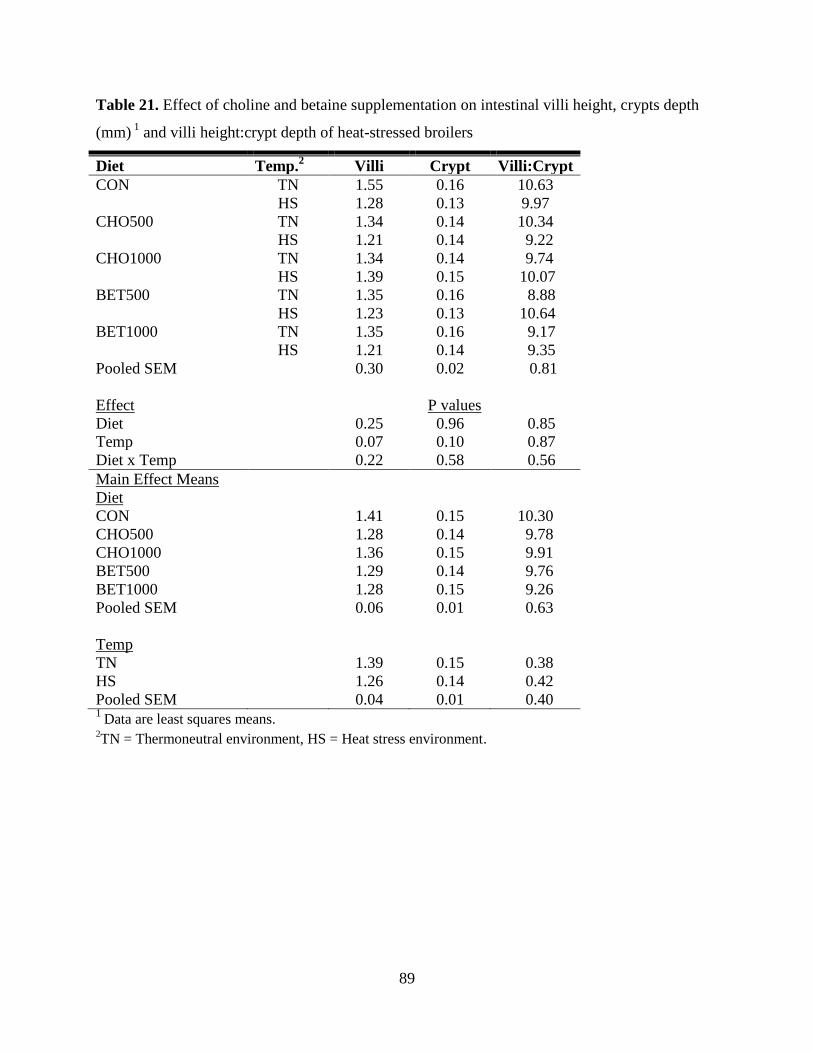
Table 21. Effect of choline and betaine supplementation on intestinal villi height, crypts depth
(mm) 1
and villi height:crypt depth of heat-stressed broilers ........................................................ 89
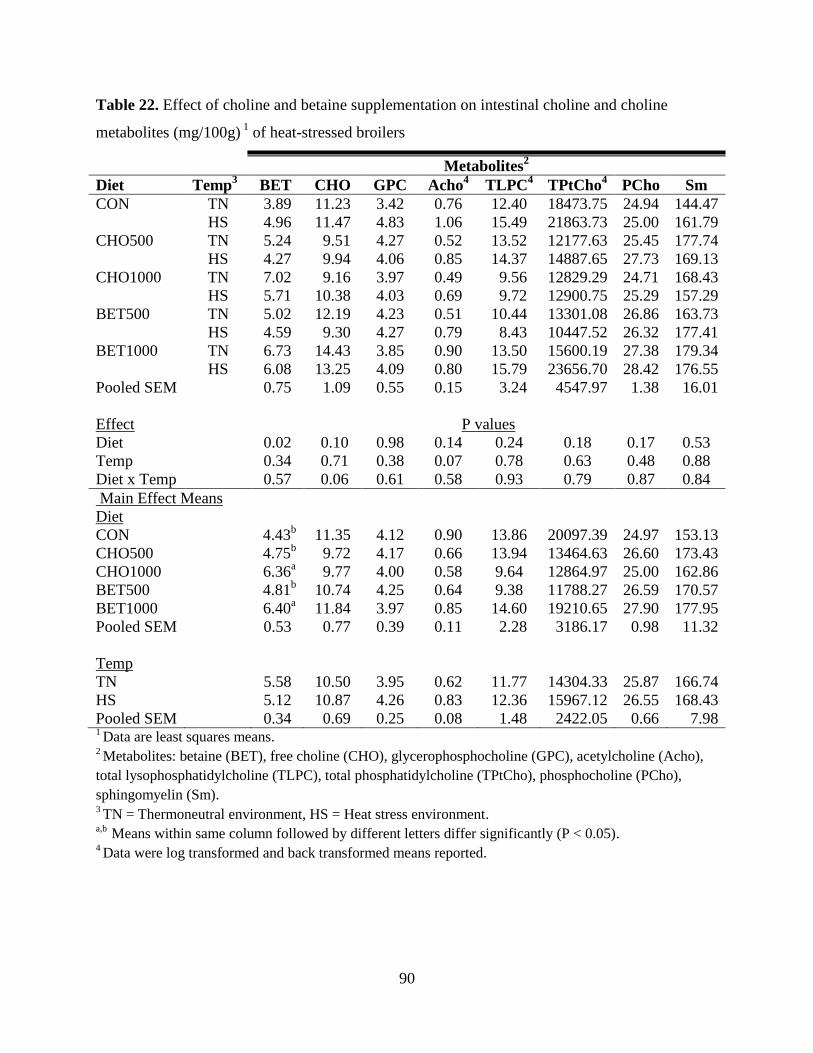
Table 22. Effect of choline and betaine supplementation on intestinal choline and choline
metabolites (mg/100g) 1
of heat-stressed broilers ......................................................................... 90
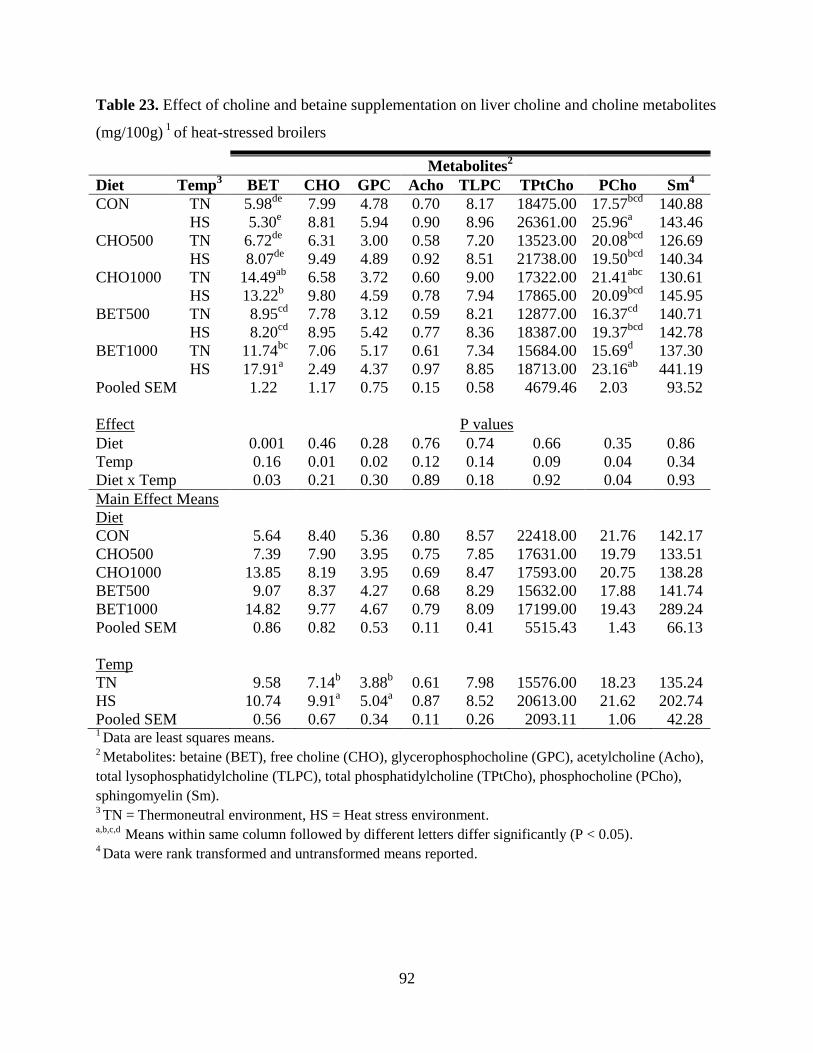
Table 23. Effect of choline and betaine supplementation on liver choline and choline metabolites
(mg/100g) 1
of heat-stressed broilers ............................................................................................. 92
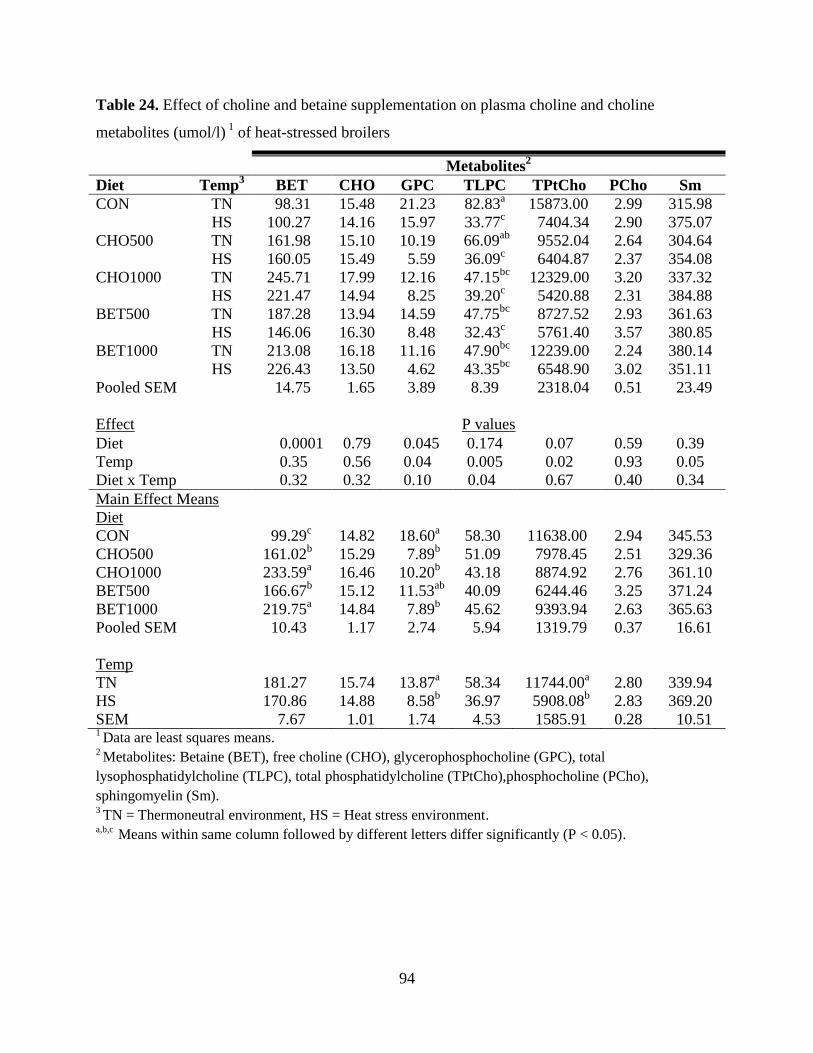
Table 24. Effect of choline and betaine supplementation on plasma choline and choline
metabolites (umol/l) 1
of heat-stressed broilers ............................................................................. 94
Page 13
xi
List of Figures
Figure 1. Typical daily temperature in TN (23.9 ±1°C) and HS (28-36±1°C) rooms. ................. 34
Figure 2. Scoring system for foot pad dermatitis (Mello, et al., 2011). ........................................ 39
Figure 3. The daily temperature in TN (23.9 ±1°C) and HS (28-36±1°C) rooms. ....................... 63
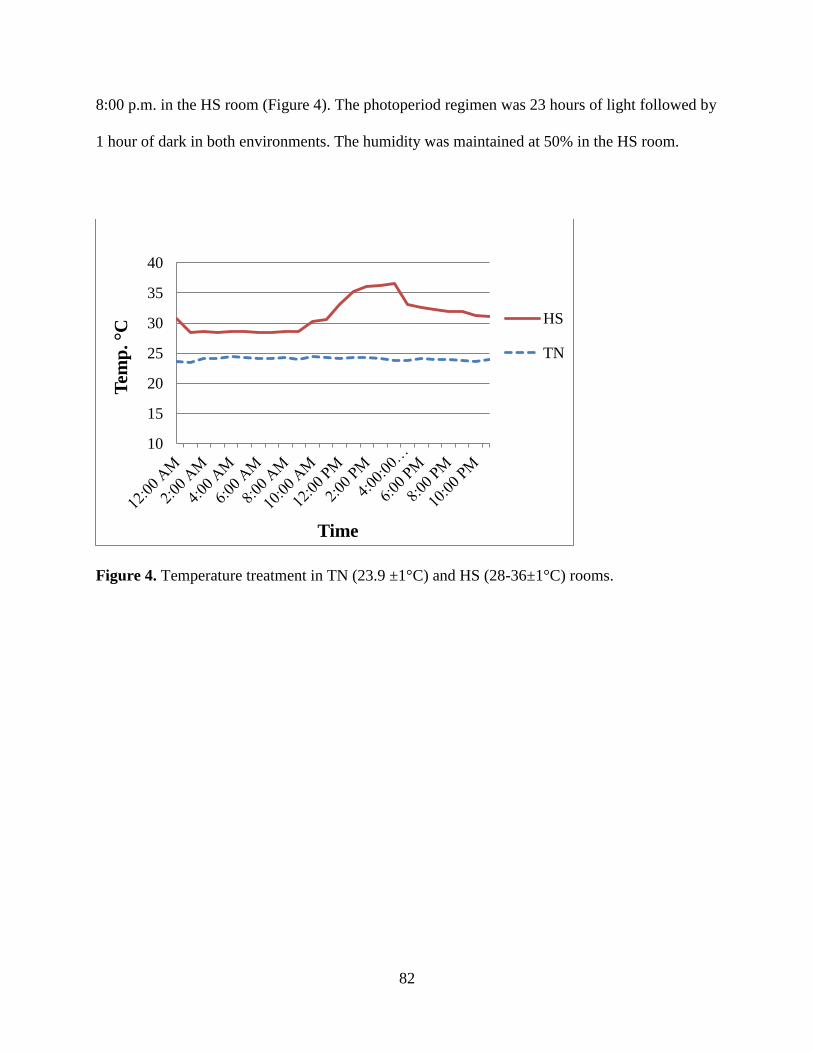
Figure 4. Temperature treatment in TN (23.9 ±1°C) and HS (28-36±1°C) rooms. ...................... 82
Page 14
1
Chapter I: Introduction
Page 15
2
The poultry industry has evolved from a dual purpose backyard operation to a vertically
integrated industry (USDA, 2002) with two distinct production systems, broiler (meat)
production and layer (egg) production. Both systems have benefited from various advances
allowing them to produce more efficiently through improved nutrition, genetics, environment
and management practices. Despite these achievements, the industry is still facing some
problems such as the effect of heat stress. Modern poultry production is highly intensive and
occurs in large enclosed houses with high stocking density and fast growing birds. Combination
of these factors leads to an increase of heat in chicken houses. Faster growing broilers produce
more heat (Cahaner and Leenstra, 1992; Yalcin, et al., 2001) and in addition, in the United
States, which is the highest producing country, the production of broilers is mostly concentrated
in the southeastern region (USDA, 2014) where extreme summer temperature combined with
high humidity creates the ideal condition for heat related stress (Konrad, 2013).
Heat stress occurs when the body produces or absorbs more heat than it can dissipate
which can result in negative consequences. Under heat stress, chickens eliminate the extra heat
by increasing their respiratory rate. This increased respiration may result in acid/base imbalance
or alkalosis (Borges, et al., 2004; Toyomizu, et al., 2005). Prolonged heat stress reduces visceral
blood supply to the intestine and causes damages to epithelial cells in the gut there by affecting
feed digestion and nutrients absorption (Cronje, 2007). Prolonged heat stress may also disrupt
intestinal barrier increasing the likelihood of pathogenic bacteria and endotoxin entry, which can
then result in excessive inflammation, decreasing production performance, and possibly death
(Quinteiro-Filho, et al., 2012). Heat stress prior to slaughter increases hemorrhage incidence
thereby decreasing meat quality post processing (Sandercock, et al., 2001). It also disrupts
intestinal integrity (Lambert, 2009) thus rendering it less resistant to breakage during mechanical
Page 16
3
processing. According to Rosenquist, et al. (2006), the highest rate of chicken carcass
contamination by campylobacter occurs during evisceration and is likely to cause food borne
illness or disease outbreak. Heat stress can be very costly to the poultry industry. In 2003, it was
estimated to cost the poultry industry approximately 128 million dollars annually (St-Pierre, et
al., 2003).
Over the years, producers have been trying to find various ways to alleviate the negative
impacts of heat stress on broilers. Management practices such as electrolyte supplementation
have been used to alleviate the negative impacts of heat stress. It has been shown that continuous
supply of electrolytes (K+ and Cl
-) improves weight gain (Smith and Teeter, 1992) and overall
performance. However, electrolytes are limited osmotic effectors and their excessive
accumulation in tissues may inhibit protein synthesis (Petronini, et al., 1992). Because of this
negative impact, an alternative to electrolytes is being sought, one such alternative being organic
osmolytes, such as choline and betaine. Choline and betaine are two important compounds that
benefit the poultry industry by improving production. Several studies have shown that not only
do they support growth but also alleviate the negative impacts of heat stress on chicken
physiology and production performances by attenuating cells dehydration (Attia, et al., 2009;
Farooqi, et al., 2005).
Choline plays various metabolic functions throughout the body including building and
maintenance of cell structure primarily as phosphatidylcholine, fat mobilization in the liver and
formation of acetylcholine (Combs Jr, 1991; McDowell, 1989). It is a component of molecules
important in the fetal development (Jones III, et al., 1999), in inflammation (Wang, et al., 2005)
and in blood clotting (Wells, 1964). Choline constitutes a source of osmolytes through its
metabolites, betaine and glycerophosphocholine, for regulating cell volume (Nakanishi and
Page 17
4
Burg, 1989). Glycerophosphocholine, mainly formed from phosphatidylcholine, is the
predominant osmolyte in the kidney that protects renal medullary cells from hypertonicity of the
interstitial fluid (Nakanishi and Burg, 1989). Betaine, an important methyl group donor to the
formation of methionine, has been shown to improve lipid metabolism and production
performance of broilers (Zhan, et al., 2006). In intestinal cells subjected to osmotic disorder and
dehydration, betaine is taken up and may have a stabilizing function (Kettunen, et al., 2001). It
also stimulates intestinal epithelial cell proliferation, feed digestibility and absorption of nutrients
(Augustine and Danforth, 1999).
Choline is already being included in broiler feed at required levels. Because of their
osmotic properties, it is possible that choline and betaine could be included at higher levels.
Various researchers have investigated this possibility, however further research is warranted to
understand the integrated role of these two compounds on chicken physiology and growth
performances especially in heat stress environment. Therefore, the objectives of this experiment
were to evaluate the osmoprotectant benefits of choline and betaine on growth performance and
on the intestine of heat-stressed broilers.
Page 18
5
References
Attia, Y., R. Hassan, and E. Qota. 2009. Recovery from adverse effects of heat stress on slow-
growing chicks in the tropics 1: Effect of ascorbic acid and different levels of betaine. Trop.
Anim. Health Prod. 41:807-818.
Augustine, P., and H. Danforth. 1999. Influence of betaine and salinomycin on intestinal
absorption of methionine and glucose and on the ultrastructure of intestinal cells and parasite
developmental stages in chicks infected with Eimeria acervulina. Avian Dis.:89-97.
Borges, S. A., A. V. F. da Silva, A. Majorka, D. M. Hooge, and K. R. Cummings. 2004.
Physiological responses of broiler chickens to heat stress and dietary electrolyte balance
(sodium plus potassium minus chloride, milliequivalents per kilogram). Poult. Sci. 83:1551-
1558.
Cahaner, A., and F. Leenstra. 1992. Effects of high temperature on growth and efficiency of
male and female broilers from lines selected for high weight gain, favorable feed conversion,
and high or low fat content. Poult. Sci. 71:1237-1250.
Combs Jr, G. F. 1991. The vitamins: fundamental aspects in nutrition and health. Am. J. Clin.
Nutr. 53:755-763.
Cronje, P. Year. Gut health, osmoregulation and resilience to heat stress in poultry. Proc. Proc.
19th Aust. Poult. Sci. Symp., Sydney, New South Wales, Australia.
Farooqi, H., M. Khan, M. Khan, M. Rabbani, K. Pervez, and J. Khan. 2005. Evaluation of
betaine and vitamin C in alleviation of heat stress in broilers. Int. J. Agric. Biol 5:744-746.
Jones III, J., W. H Meck, C. L. Williams, W. A. Wilson, and H. S. Swartzwelder. 1999. Choline
availability to the developing rat fetus alters adult hippocampal long-term potentiation. Dev.
Brain Res. 118:159-167.
Kettunen, H., S. Peuranen, K. Tiihonen, and M. Saarinen. 2001. Intestinal uptake of betaine in
vitro and the distribution of methyl groups from betaine, choline, and methionine in the body
of broiler chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 128:269-278.
Konrad, C. E., Fuhrmann, M. 2013. Climate of the Southeast USA: past, present and future.
Pages 8-36 in Climate of the Southeast Unites States. Ingram k., K. Dow, L. Carter, J.
Anderson, eds. Washington DC: Island Press.
Lambert, G. P. 2009. Stress-induced gastrointestinal barrier dysfunction and its inflammatory
effects. J. Anim. Sci. 87:E101-E108. doi 10.2527/jas.2008-1339
McDowell, L. R. 1989. Choline. Pages 565-596 in Vitamins in Animal and Human Nutrition.
2nd ed.
Page 19
6
Nakanishi, T., and M. B. Burg. 1989. Osmoregulation of glycerophosphorylcholine content of
mammalian renal cells. Am. J. Physiol. 257:C795-801.
Petronini, P. G., E. De Angelis, P. Borghetti, A. Borghetti, and K. P. Wheeler. 1992. Modulation
by betaine of cellular responses to osmotic stress. Biochem. J 282:69-73.
Quinteiro-Filho, W., A. Gomes, M. Pinheiro, A. Ribeiro, V. Ferraz-de-Paula, C. Astolfi-Ferreira,
A. Ferreira, and J. Palermo-Neto. 2012. Heat stress impairs performance and induces
intestinal inflammation in broiler chickens infected with Salmonella Enteritidis. Avian Pathol.
41:421-427.
Rosenquist, H., H. M. Sommer, N. L. Nielsen, and B. B. Christensen. 2006. The effect of
slaughter operations on the contamination of chicken carcasses with thermotolerant< i>
Campylobacter</i>. Int. J. Food Microbiol. 108:226-232.
Sandercock, D. A., R. R. Hunter, G. R. Nute, M. A. Mitchell, and P. M. Hocking. 2001. Acute
heat stress-induced alterations in blood acid-base status and skeletal muscle membrane
integrity in broiler chickens at two ages: Implications for meat quality. Poult. Sci. 80:418-425.
Smith, M. O., and R. G. Teeter. 1992. Effects of potassium chloride supplementation on growth
of heat-distressed broilers. J. Appl. Poult. Res. 1:321-324.
St-Pierre, N., B. Cobanov, and G. Schnitkey. 2003. Economic losses from heat stress by US
livestock industries. J. Dairy Sci. 86:E52-E77.
Toyomizu, M., M. Tokuda, A. Mujahid, and Y. Akiba. 2005. Progressive alteration to core
temperature, respiration and blood acid-base balance in broiler chickens exposed to acute heat
stress. J. Poult. Sci. 42:110-118.
USDA. 2002. U.S. Broiler Industry Structure. Pages 5 National Agricultural Statistics Service,
Washigton DC.
USDA. 2014. Poultry - Production and Value 2013 Summary. Pages 14.
Wang, Y., D.-M. Su, R.-H. Wang, Y. Liu, and H. Wang. 2005. Antinociceptive effects of choline
against acute and inflammatory pain. Neuroscience 132:49-56.
Wells, I. C. 1964. A blood clotting defect in choline deficient rats. Biochim. Biophys. Acta. Gen.
Subj. 86:339-345.
Yalcin, S., S. Özkan, L. Türkmut, and P. Siegel. 2001. Responses to heat stress in commercial
and local broiler stocks. 1. Performance traits. Br. Poult. Sci. 42:149-152.
Zhan, X., J. Li, Z. Xu, and R. Zhao. 2006. Effects of methionine and betaine supplementation on
growth performance, carcase composition and metabolism of lipids in male broilers. Br. Poult.
Sci. 47:576-580.
Page 20
7
Chapter II: Literature Review
Page 21
8
Heat Stress in Broilers
Broiler chickens experience heat stress when their body produces or absorbs more heat
than it can dissipate. Chicken brooding temperature is approximately 35oC during the first week
of age, however, the comfort zone for chickens declines to approximately 24oC by four weeks
(Teeter and Belay, 1996). When the environmental temperature exceeds the comfort limit,
chickens suffer heat related stress resulting in physiological and metabolic changes (Borges, et
al., 2007). Chickens are homeotherms and have developed adaptation mechanisms to maintain
their core body temperature in the normal range of 40-41oC. When the environmental
temperature exceeds their comfort zone, chickens reduce their feed intake in order to decrease
the heat production associated with feed consumption and metabolism, as well as increase non-
evaporative heat loss followed by evaporative cooling (Teeter and Belay, 1996).
Non-evaporative cooling constitutes the first response of chickens to heat stress (Teeter
and Belay, 1996). Birds increase their surface area and blood supply to the periphery to dissipate
heat through radiation, convection and conduction. As the environmental temperature rises closer
to the bird’s body temperature, non-evaporative cooling is no longer efficient and chickens rely
more on evaporative cooling (Teeter and Belay, 1996) which occurs through evaporation of
water from the respiratory system. Chickens also eliminate extra heat through hyperventilation
(Borges, et al., 2007; Bottje and Harrison, 1985; Donkoh, 1989; Teeter and Belay, 1996).
However, increased respiratory rate results in exhalation of too much carbon dioxide. This
reduces carbon dioxide partial pressure in blood while bicarbonate remains relatively constant
causing an elevation of blood pH thus resulting in acid-base imbalance known as respiratory
alkalosis (Borges, et al., 2007; Smith and Teeter, 1987b). Through renal compensation, more
bicarbonate is excreted and less hydrogen ions (H+) secreted to correct the acid-base imbalance
Page 22
9
(Borges, et al., 2004). Evidence suggests that in mammals (Rector Jr, et al., 1962), the decrease
in H+ secretion results in excretion of more potassium ions (K
+) which has been shown to alter
electrolytes balance. Smith and Teeter (1987b) reported 633% increase of K+ excretion in
chickens under heat stress (35oC) compared to thermoneutral (24
oC) conditions.
Several management practices have been developed and used to improve bird’s comfort
and survival thus mitigating the negative impacts of heat stress on broilers. These include house
insulation, ventilation, feed withdrawal (Mahmoud and Yaseen, 2005; Soutyrine, et al., 1998),
fasting before the onset of heat stress and acclimation of high temperature in younger birds (De
Basilio, et al., 2001; Yalcin, et al., 2001). Other techniques consist of dietary modification to
increase the energy content from fat in order to reduce heat increment under heat stress
conditions (Dale and Fuller, 1979). These approaches do not directly address the acid-base
imbalance and its subsequent electrolyte imbalance. However, several studies have shown that
electrolyte supplementation in feed or water improves performance (Ahmad and Sarwar, 2005;
Smith and Teeter, 1992), survival (Smith and Teeter, 1987a) and reduces alkalosis (Teeter and
Smith, 1986) in heat-stressed broilers. Smith and Teeter (1986) observed that, the addition of
adequate amount of ammonium chloride (NH4Cl) 0.3% or less reduced blood pH and increased
live weight gain. The supplementation of K+ as potassium chloride (KCl) increased weight gain
but had no effect on the blood pH (Teeter and Smith, 1986). The acid-base imbalance can also be
corrected by carbonating the drinking water or supplementing it with calcium chloride (CaCl2)
(Bottje and Harrison, 1985). While electrolytes have been shown to improve performance and
welfare of heat-stressed chickens, their excess may lead to adverse effects. High levels of NH4Cl
(Teeter and Smith, 1986), sodium bicarbonate (NaHCO3) and CaCl2 (Bottje and Harrison, 1985)
Page 23
10
used for correcting heat stress induced alkalosis can result in acidosis and reduced body weight
gain in heat-stressed broilers.
Sodium (Na+), chloride (Cl
-) and K
+ are electrolytes that participate in the maintenance of
body ionic and water balance. In a study utilizing chick embryos, Petronini, et al. (1992) reported
that the excess accumulation of Na+ and Cl
- interfere with cells’ proliferation and protein
synthesis as opposed to organic osmolytes such as betaine. Excess electrolytes in broiler feed or
drinking water can lead to increased excreta moisture and litter wetness (Francesch and Brufau,
2004; Smith, et al., 2000) which constitute the main source of foot pad dermatitis (Greene, et al.,
1985; Mayne, et al., 2007).
Foot Pad Dermatitis
Foot pad dermatitis (FPD), also known as pododermatitis or contact dermatitis, is
characterized by inflammation and lesion on the plantar surface of the feet and toes (Ekstrand, et
al., 1997; Greene, et al., 1985; Shepherd and Fairchild, 2010). In chickens, FPD appears similar
to that in turkey with few differences. The metatarsal, digital pad and interpad spaces are
ulcerated in chickens while only the pad is affected in turkey (Martland, 1985). In the early stage
of FPD, there is discoloration of the skin (Ekstrand, et al., 1997) that progresses in severe cases
into scab covered ulcers, intraepithelial inflammation, dermal capillary thrombosis and extensive
granulation tissue (Martland, 1985). Incidence of FPD may be related to high dietary
concentration of Na+, K
+ and phosphorous (P) which increase water intake and excreta moisture
in chickens and turkeys (Abd El‐Wahab, et al., 2013; Smith, et al., 2000). According to Berg and
Algers (2004), FPD could also be a result of high protein content or be related to dietary factors
such as indigestible fats leading to sticky droppings. McIlroy, et al. (1987) stated that litter
Page 24
11
quality, affected by stocking density and drinking water design could increase FPD. Poor
ventilation has been shown to increase litter moisture content (Martrenchar, et al., 2002) and
therefore FPD incidence. However, other factors could also contribute to FPD.
Foot pad dermatitis has been linked to genetics. According to Kjaer, et al. (2006) it may
be possible to decrease the incidence of FPD by genetic selection without negatively affecting
body weight. Another factor reportedly involved in FPD is biotin. However, there has been
mixed findings on its importance in the prevention of FPD. Harms and Simpson (1977) reported
that biotin supplementation decreased the severity of FPD when given to poults fed a diet
deficient in biotin and raised on wet litter as opposed to poults maintained on wet litter without
biotin supplementation. Conversely, Cengiz, et al. (2012) reported that supplemental biotin does
not minimize or prevent the development of FPD in broiler chickens.
The occurrence of FPD has significant welfare and economic implications for the broiler
industry. In severe cases, FPD lesions cause pain and walking difficulties in broilers (Berg and
Algers, 2004) with lameness (Greene, et al., 1985) thus reducing their access to feeders.
Martland (1985) suggested that turkeys experiencing severe FPD may develop pain-induced
inappetance which depresses growth rate and body weight causing economic losses for the
producer. Foot pad dermatitis causes birds to sit on their breast for long periods of time resulting
in the development of hock and breast lesions (Martland, 1985) which degrades the meat quality
(McIlroy, et al., 1987). With the recent development of an export market of chicken paw to
China and Hong Kong, in addition to welfare issue, FPD has become an important economic
component in the development of the broiler industry (USDA, 2013). Good litter management is
the key to preventing FPD (Hoffmann, et al., 2013). Litter wetness has been shown to be
increased by dietary concentration of Na+ and K
+, due to high moisture content of the excreta
Page 25
12
(Smith, et al., 2000). Therefore, the use of these electrolytes to minimize the negative impact of
heat stress on broilers must be done with respect to the dietary electrolytes balance (Ahmad and
Sarwar, 2005).
Impact of Heat Stress on Intestinal Morphology
The gastrointestinal tract serves in the digestion and absorption of nutrients and as a
barrier between the luminal content and the systemic circulation. The maintenance of the
physical integrity of the intestine is primordial for its functions. The intestinal barrier is
composed of apical membrane of cells of the epithelium and the tight junction between them
(Sun, et al., 1998). Several stress factors including osmotic stress (Mongin, et al., 1976) and heat
stress (Lambert, et al., 2002) affect gut integrity. In birds, the contents of the small intestine are
always hypertonic to the plasma osmolality of about 320 mOsmol (Klasing, et al., 2002; Mongin,
et al., 1976) exerting substantial osmotic pressure on the cells lining the gut. In heat-stressed
animals such as rats (Lambert, et al., 2002) , pigs (Pearce, et al., 2012) and chickens (Quinteiro-
Filho, et al., 2010), the tight junctions between epithelial cells are disrupted increasing the
intestinal permeability to large and normally restricted molecules from the intestinal lumen to the
blood. During heat stress, blood is diverted to the periphery to dissipate the extra heat, reducing
blood flow to the viscera. Prolonged reduced blood flow causes tissue hypoxia, generation of
reactive oxygen species and damage to enterocyte membrane (Hall, et al., 1999; Lambert, 2004)
resulting in increased intestinal permeability. The intestinal lumen contains commensal bacteria
and a number of other ingested pathogens. Endotoxins from bacteria present in the intestine
usually enter the internal environment at a basal level in normal conditions. However, with
compromised intestinal integrity, the level of endotoxin entry increases resulting in greater
Page 26
13
release of proinflammatory cytokines by the immune system causing further inflammation of the
intestinal epithelium (Lambert, 2009). Heat stress has also been proven to increase the absorption
of some nutrients. Pearce, et al. (2012) showed that there is an increase in glucose absorption in
heat-stressed pigs. Galactose and methionine absorption in the jejunum of chickens are increased
in response to heat stress (Mitchell and Carlisle, 1992).
Heat Stress and the Immune System
According to Selye (1976), stress is a non-specific response of the body to any stimulus
or stressor. Stress results in the activation of the hypothalamus-pituitary-adrenal axis (HPA) and
the secretion of glucocorticoid hormones: cortisol and corticosterone. Cortisol is the main
glucocorticoid in fish and most mammals, whereas corticosterone is the primary glucocorticoid
in birds, reptiles, amphibians and rodents (Cockrem, 2013). Glucocorticoids’ function in the
regulation of inflammatory response is well established. They inhibit proinflammatory cytokines
such as interleukin (IL)-12, tumor necrosis factor (TNF)-α and Interferon (IFN)-γ, and stimulate
the production of anti-inflammatory cytokines IL-10, IL-4 and transforming growth factor
(TGF)-β to control excessive immune response (Elenkov and Chrousos, 2002). An increase in
proinflammatory cytokines during excessive inflammation activates the HPA axis resulting in
increased production of glucocorticoids to reduce inflammation (Chrousos, 1995). Heat stress in
chickens has been shown to increase plasma corticosterone concentration (Beuving and Vonder,
1978; Quinteiro-Filho, et al., 2010) and a short term increase may be beneficial. In a study
conducted by Nasir, et al. (1999), adrenalectomised chickens were injected with corticosterone to
mimic an increase of corticosterone during stress. Results showed that elevated plasma
corticosterone stimulated higher feed intake, delayed food passage time and increased calcium,
Page 27
14
phosphorous and glucose absorption. Garriga, et al. (2006) also reported an increase in apical
glucose transport during heat stress. Enhanced gluconeogenesis following short term
administration of corticosterone in chickens was reported by Lin, et al. (2004) and Shini, et al.
(2008a). However, a production of glucocorticoids at a suboptimal level has been shown to
impair the immune system by decreasing inflammatory response (Chrousos, 1995) or by
reducing spleen and bursa weights (Shini, et al., 2008b).
Choline
Choline, the common name for 2-hydroxy-N,N,N-trimethylethanammonium (Combs Jr,
1991), is a water soluble vitamin-like compound found in the form of phosphatidylcholine, or
sphingomyelin in most foods (Combs Jr, 1991; McDowell, 1989) with the highest concentrations
in wheat germ, dried soybean, egg yolk, pork, and glandular meat such as beef and chicken liver
(Zeisel, et al., 2003). Depending on its functions in the body, choline is metabolized to different
compounds, including phosphatidylcholine essential for cell membrane integrity (Plagemann,
1968) and transmembrane signaling (Zeisel, 1993), acetylcholine involved in neurotransmission
(Cohen and Wurtman, 1976; Wecker, 1989), and betaine which donates methyl groups for the
methylation of homocysteine to methionine or functions as organic osmolyte (Zeisel, et al.,
1989). Choline also plays an important role in fat metabolism. Its deficiency is associated with
fatty liver (Cooke, et al., 2007; Vendemiale, et al., 2001) and low body fat (Daily III, et al.,
1998) in all animal species. Phosphatidylcholine is required for the synthesis of very low density
lipoprotein (VLDL) in the liver (Yao and Vance, 1988) or further metabolized to
glycerophosphocholine predominantly in renal medullary cells where it protects against hyper
salinity of the interstitial fluid (Beck, et al., 1992).
Page 28
15
Free choline is taken up via a carrier mediated pathway or by passive diffusion at low and
high concentration respectively (Herzberg and Lerner, 1973; Kamath, et al., 2003) and enter the
portal circulation. Choline is transported by three different mechanisms: high-affinity Na+-
dependent transporter that supplies choline for the synthesis of acetylcholine (Meyer Jr, et al.,
1982), low-affinity Na+-independent transporter that supplies choline for phosphatidylcholine
and other phospholipids synthesis in cell membranes, and very low-affinity Na+-independent
residual diffusion (Van Rossum and Boyd, 1998). Phosphatidylcholine, another form of choline
present in foods, is hydrolyzed in the intestine to lysophosphatidylcholine before absorption. In
the enterocytes, lysophosphatidylcholine is re-esterified to phosphatidylcholine, packed into
chylomicrons and sent to the lymphatic circulation (Le Kim and Betzing, 1976).
The signs of choline deficiency vary across animal species but the most common ones
include fatty liver, poor growth, hemorrhagic tissue and hypertension (McDowell, 1989). In
young chickens and turkeys, perosis (slipped tendon) is the primary sign of choline deficiency.
All animal species are able to synthesize sufficient amount of choline when enough methyl
groups, methionine or betaine are supplied in the diet (McDowell, 1989) except young chickens
until about 8 weeks of age (Cook, et al., 1984; Molitoris and Baker, 1976). Choline is therefore
required in poultry nutrition and has always been included in their diet to support growth and
prevent perosis. Fast growing broilers reach market weight at a minimum of 6 weeks and are
unable to synthesize adequate amount of choline required 1300 mg/kg for 0-3 weeks of age,
1000 mg/kg for 3-6 weeks of age and 750 mg/kg for 6-9 weeks of age (NRC., 1994).
Because of its multiple functions in the body and requirements for chickens, several
studies have investigated the effects of supplementation of choline in broiler diet for improving
performance. However these results have been conflicting and could be due to dietary
Page 29
16
methionine contents. Rafeeq, et al. (2011) and Pillai, et al. (2006) reported that choline increases
weight gain in broiler chickens fed a methionine deficient diet. A 300 mg/kg choline
supplementation to a diet containing adequate methionine improved growth performance (Dilger,
et al., 2007; Tillman and Pesti, 1986; Tsiagbe, et al., 1987) while other studies reported no
significant effect of choline on weight gain (Baker, et al., 1983; Waldroup and Fritts, 2005).
Supplemental choline was also shown to improve breast yield (Waldroup and Fritts, 2005) and
feed conversion ratio (Tillman and Pesti, 1986; Waldroup and Fritts, 2005).
Betaine
Betaine, a naturally occurring trimethylglycine (methylated amino acid), is found
predominantly in sugar beet, from which it was isolated for the first time, wheat bran, wheat and
spinach (Zeisel, et al., 2003). Chemically, betaine is a zwitterion quaternary ammonium
compound and a derivative of amino acid glycine (Craig, 2004). Its purified forms, anhydrous
betaine, betaine monophosphate and betaine chloride are used as feed additives (Eklund, et al.,
2005). Betaine can either be provided in the diet or synthesized by the mitochondria in the liver
and kidney from the oxidation of choline (Yamauchi, et al., 1992). As methylated amino acid,
betaine is mainly transported via betaine gamma amino butyric acid transporter (BGT-1) and
amino acid transport system A in most tissues (Petronini, et al., 2000; Yamauchi, et al., 1992). In
chickens, intestinal betaine is absorbed via Na+-dependent and Na
+-independent transport
systems. High concentration of betaine increases Na+-dependent transport resulting in higher
betaine uptake (Kettunen, et al., 2001b).
Betaine’s main functions are to serve as a methyl group donor for various methylation
reactions in the body or as osmolyte to protect cells subjected to osmotic pressure. As a methyl
Page 30
17
donor, betaine does not participate in most methylations but donates methyl groups to
homocysteine to form methionine (Vincent , et al., 1946). Its osmoprotectant property is also
established in plants (Storey and Jones, 1975; Yang, et al., 2003) and in bacteria (Sutherland, et
al., 1986). In bacteria, when subjected to mild hyperosmolar environment the primary
intracellular osmoprotectant is K+. However, with increased environmental salinity, a
corresponding internal K+ concentration is deleterious to enzymes function and betaine is
accumulated in preference to K+ to restore osmotic balance (Sutherland, et al., 1986). Virtanen,
et al. (1989), studying the adaptation of young Atlantic salmon, showed that betaine in an
effective osmolyte that helps maintain ionic and osmotic balance in sea water.
Due to its methyl donor and osmoprotectant properties, physiological and nutritional
effects of betaine have been investigated in livestock on performance and carcass quality
(Sakomura, et al., 2013; Waldroup and Fritts, 2005), intestinal infection states (Kettunen, et al.,
2001a; Klasing, et al., 2002) as well as stress conditions (Attia, et al., 2009). Betaine has been
shown to promote growth in pigs by enhancing growth hormone synthesis in finishing pigs
(Huang, et al., 2007). Pigs fed conjugated linoleic acid (1%) and betaine (0.5%) increased
leanness and decreased shoulder fat deposition (Rojas-Cano, et al., 2011). In broiler chickens,
betaine supplementation at various levels indicates some improvement on performance and
carcass quality. Improved breast yield was reported by McDevitt, et al. (2000) and Waldroup, et
al. (2006) when 0.5 g/kg betaine was supplemented. Waldroup, et al. (2006) also reported that
with 500 mg/kg or 1000 mg/kg betaine supplementation there was improved feed conversion
ratio at 35 and 42 days. Betaine supplementation at 0.072% and 0.144% decreased abdominal fat
deposition (Hassan, et al., 2005). Waldroup and Fritts (2005) saw an increase in dressing
percentage at 42 days of age in broiler chickens supplemented with 1000 mg/kg of betaine.
Page 31
18
Evidence strongly suggests that betaine acts as an osmolyte in intestinal cells. Betaine was
shown to improve diarrheal state and performance (Fetterer, et al., 2003) in coccidia-infected
chickens. Klasing, et al. (2002) showed that the supplementation of betaine at 0.5 g/kg and 1
g/kg improved osmolarity and restored villi length in the duodenum of infected chickens.
Supplemental betaine at 0.2% helps the duodenum maintain water balance in hypertonic
environment (Kettunen, et al., 2001a).
Interrelation of Choline, Betaine and Methionine Metabolism
Choline is considered a potential source of methyl groups along with methionine but is
not directly involved in methylation reactions unless oxidized into betaine (Borsook and
Dubnoff, 1947). Betaine donates its methyl group to homocysteine to form methionine, one of
the essential and limiting amino acids in chickens (Stucki and Harper, 1961). As reviewed by
Craig (2004), Eklund, et al. (2005) and Metzler-Zebeli, et al. (2009), methionine is first
converted to S-adenosylmethionine (SAM) by the enzyme methionine adenosyltransferase. S-
adenosylmethionine is the active methyl donor; it liberates methyl groups used in different
biosynthetic methylations, yielding S-adenosylhomocysteine. S-adenosylhomocysteine is
converted to homocysteine by the action of S-adenosylhomocysteine hydrolase. Homocysteine is
either irreversibly transformed to cysteine which can be utilized for protein synthesis or re-
methylated to methionine by other methyl group sources; betaine via betaine homocysteine-
methyltransferase (BHMT) or N5-methyl tetrahydrofolate via tetrahydrofolate-methyltransferase
(THMT).
Choline, betaine and methionine are interrelated and their ability to spare each other has
been extensively investigated in poultry nutrition with variable results. Various researchers have
Page 32
19
reported significant methionine sparing effect of choline and betaine. Baker, et al. (1983),
evaluated three choline (0, 217, and 434 mg/kg) and five methionine levels (0, 0.05, 0.10, 0.15
and 0.2%) in a 3x5 factorial design and found that supplementation of choline at 217 mg/kg with
0 and 0.05% methionine increased performance in poultry. By adding 0 or 0.05% supplemental
methionine to the basal diets containing 0.35% DL-methionine, the researchers made them
slightly deficient with regard to NRC requirements (0.5%). Supplementation of betaine and
choline to a diet containing 0.37% methionine (Pesti, et al., 1979) and betaine to 76% methionine
adequate diet (Zhan, et al., 2006) improved weight gain and feed conversion ratio suggesting that
choline and betaine can spare methionine only in a small range of methionine deficiency. In a
study conducted by Dilger, et al. (2007), graded levels of choline (0, 150,300 and 1000 mg/kg),
betaine (260 and 600 mg/kg) and methionine at 1289 mg/kg was added to choline-free diet. They
found that supplementation of choline-free diet with betaine and methionine showed a minimal
choline sparing effect. However, the addition of betaine to a diet containing a minimal level of
choline resulted in a substantial increase in growth performances. They concluded that 50% of
dietary requirement of choline must be supplied as choline and the rest can be replaced by
betaine. While the 50% reduction might be questionable, it is noteworthy that because choline
has multiple functions in the body, betaine could only spare for its methyl donor property.
Studies have shown no methionine sparing effects of choline and betaine. Esteve-Garcia and
Mack (2000) conducted a study in which two levels of betaine (0 and 0.5 g/kg) and three levels
of methionine (0, 0.6 and 1.2 g/kg) were supplemented to a methionine deficient diet. They
found that unlike betaine, methionine supplementations improved weight gain and feed
efficiency confirming that betaine does not spare methionine. Similarly, Schutte, et al. (1997)
added two levels of betaine (0 and 0.04%) and three levels of methionine (0, 0.05 and 0.1%) to
Page 33
20
different combinations of methionine deficient diets and reported that betaine did not improve
growth contrary to methionine supplementations.
Page 34
21
References
Abd El‐Wahab, A., C. Visscher, A. Beineke, M. Beyerbach, and J. Kamphues. 2013. Effects of
high electrolyte contents in the diet and using floor heating on development and severity of
foot pad dermatitis in young turkeys. J. Anim. Physiol. Anim. Nutr. (Berl.) 97:39-47.
Ahmad, T., and M. Sarwar. 2005. Influence of varying sources of dietary electrolytes on the
performance of broilers reared in a high temperature environment. Anim. Feed Sci. Technol.
120:277-298.
Attia, Y., R. Hassan, and E. Qota. 2009. Recovery from adverse effects of heat stress on slow-
growing chicks in the tropics 1: Effect of ascorbic acid and different levels of betaine. Trop.
Anim. Health Prod. 41:807-818.
Baker, D., K. Halpin, G. Czarnecki, and C. Parsons. 1983. The choline-methionine
interrelationship for growth of the chick. Poult. Sci. 62:133-137.
Beck, F. X., M. Schmolke, W. G. Guder, A. Dorge, and K. Thurau. 1992. Osmolytes in renal
medulla during rapid changes in papillary tonicity. Am. J. Physiol. 262:F849-F856.
Berg, C., and B. Algers. Year. The effect of floor heating and feed protein level on the incidence
of foot pad dermatitis in turkeys poults. Proc. EAAP-55th Annual Meeting L.
Beuving, G., and G. Vonder. 1978. Effect of stressing factors on corticosterone levels in the
plasma of laying hens. Gen. Comp. Endocrinol. 35:153-159.
Borges, S., F. Da Silva, A. Maiorka, D. Hooge, and K. Cummings. 2004. Effects of diet and
cyclic daily heat stress on electrolyte, nitrogen and water intake, excretion and retention by
colostomized male broiler chickens. Int. J. Poult. Sci 3:313-321.
Borges, S. A., A. V. F. Da Silva, and A. Maiorka. 2007. Acid-base balance in broilers. Worlds
Poult. Sci. J. 63:73-81. doi Doi 10.1079/Wps2005128
Borsook, H., and J. W. Dubnoff. 1947. Methionine formation by transmethylation in vitro. J.
Biol. Chem. 169:247-258.
Bottje, W. G., and P. C. Harrison. 1985. The effect of tap water, carbonated water, sodium
bicarbonate, and calcium chloride on blood acid-base balance in cockerels subjected to heat
stress. Poult. Sci. 64:107-113.
Cengiz, Ö., J. Hess, and S. Bilgili. 2012. Dietary biotin supplementation does not alleviate the
development of footpad dermatitis in broiler chickens. J. Appl. Poult. Res. 21:764-769.
Chrousos, G. P. 1995. The Hypothalamic–Pituitary–Adrenal Axis and Immune-Mediated
Inflammation. N. Engl. J. Med. 332:1351-1363. doi doi:10.1056/NEJM199505183322008
Page 35
22
Cockrem, J. F. 2013. Individual variation in glucocorticoid stress responses in animals. Gen.
Comp. Endocrinol. 181:45-58.
Cohen, E. L., and R. J. Wurtman. 1976. Brain acetylcholine: control by dietary choline. Science
191:561-562.
Combs Jr, G. F. 1991. The vitamins: fundamental aspects in nutrition and health. Am. J. Clin.
Nutr. 53:755-763.
Cook, M., W. Springer, and J. Hebert. 1984. Enhanced incidence of leg abnormalities in reovirus
WVU 2937-infected chickens fed various dietary levels of selected vitamins. Avian Dis.:548-
561.
Cooke, R., N. S. Del Rio, D. Caraviello, S. Bertics, M. Ramos, and R. Grummer. 2007.
Supplemental choline for prevention and alleviation of fatty liver in dairy cattle. J. Dairy Sci.
90:2413-2418.
Craig, S. A. 2004. Betaine in human nutrition. Am. J. Clin. Nutr. 80:539-549.
Daily III, J. W., N. Hongu, R. L. Mynatt, and D. S. Sachan. 1998. Choline supplementation
increases tissue concentrations of carnitine and lowers body fat in guinea pigs. J. Nutr.
Biochem. 9:464-470.
Dale, N., and H. Fuller. 1979. Effects of Diet Composition on Feed Intake and Growth of Chicks
under Heat Stress I. Dietary Fat Levels. Poult. Sci. 58:1529-1534.
De Basilio, V., M. Vilarino, S. Yahav, and M. Picard. 2001. Early age thermal conditioning and
a dual feeding program for male broilers challenged by heat stress. Poult. Sci. 80:29-36.
Dilger, R. N., T. A. Garrow, and D. H. Baker. 2007. Betaine can partially spare choline in chicks
but only when added to diets containing a minimal level of choline. J. Nutr. 137:2224-2228.
Donkoh, A. 1989. Ambient temperature: a factor affecting performance and physiological
response of broiler chickens. Int. J. Biometeorol. 33:259-265.
Eklund, M., E. Bauer, J. Wamatu, and R. Mosenthin. 2005. Potential nutritional and
physiological functions of betaine in livestock. Nutr. Res. Rev. 18:31-48. doi
doi:10.1079/NRR200493
Ekstrand, C., B. Algers, and J. Svedberg. 1997. Rearing conditions and foot-pad dermatitis in
Swedish broiler chickens. Prev. Vet. Med. 31:167-174.
Elenkov, I. J., and G. P. Chrousos. 2002. Stress Hormones, Proinflammatory and
Antiinflammatory Cytokines, and Autoimmunity. Ann. N. Y. Acad. Sci. 966:290-303. doi
10.1111/j.1749-6632.2002.tb04229.x
Esteve-Garcia, E., and S. Mack. 2000. The effect of DL-methionine and betaine on growth
performance and carcass characteristics in broilers. Anim. Feed Sci. Technol. 87:85-93.
Page 36
23
Fetterer, R., P. Augustine, P. Allen, and R. Barfield. 2003. The effect of dietary betaine on
intestinal and plasma levels of betaine in uninfected and coccidia-infected broiler chicks.
Parasitol. Res. 90:343-348.
Francesch, M., and J. Brufau. 2004. Nutritional factors affecting excreta/litter moisture and
quality. Worlds Poult. Sci. J. 60:64-75. doi 10.1079/wps20035
Garriga, C., R. R. Hunter, C. Amat, J. M. Planas, M. A. Mitchell, and M. Moretó. 2006. Heat
stress increases apical glucose transport in the chicken jejunum. Am. J. Physiol. Regul. Intgr.
Comp. Physiol. 290:R195-R201.
Greene, J. A., R. M. McCracken, and R. T. Evans. 1985. A contact dermatitis of broilers -clinical
and pathological findings. Avian Pathol. 14:23-38. doi 10.1080/03079458508436205
Hall, D. M., K. R. Baumgardner, T. D. Oberley, and C. V. Gisolfi. 1999. Splanchnic tissues
undergo hypoxic stress during whole body hyperthermia. Am. J. Physiol. Gastrointest. Liver.
Physiol. 276:G1195-G1203.
Harms, R. H., and C. F. Simpson. 1977. Influence of wet litter and supplemental biotin on foot
pad dermatitis in turkey poults. Poult. Sci. 56:2009-2012.
Hassan, R., Y. Attia, and E. El-Ganzory. 2005. Growth, carcass quality and serum constituents of
slow growing chicks as affected by betaine addition to diets containing 1. Different levels of
choline. Int. J. Poult. Sci. 4:840-850.
Herzberg, G. R., and J. Lerner. 1973. Intestinal absorption of choline in the chick. Biochim.
Biophys. Acta, Biomembr. 307:234-242.
Hoffmann, G., C. Ammon, L. Volkamer, C. Suerie, and D. Radko. 2013. Sensor-based
monitoring of the prevalence and severity of foot pad dermatitis in broiler chickens. Br. Poult.
Sci. 54:553-561. doi 10.1080/00071668.2013.830174
Huang, Q. C., Z. R. Xu, X. Y. Han, and W. F. Li. 2007. Effect of betaine on growth hormone
pulsatile secretion and serum metabolites in finishing pigs. J. Anim. Physiol. Anim. Nutr.
(Berl.) 91:85-90.
Kamath, A. V., I. M. Darling, and M. E. Morris. 2003. Choline uptake in human intestinal Caco-
2 cells is carrier-mediated. J. Nutr. 133:2607-2611.
Kettunen, H., S. Peuranen, and K. Tiihonen. 2001a. Betaine aids in the osmoregulation of
duodenal epithelium of broiler chicks, and affects the movement of water across the small
intestinal epithelium in vitro. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 129:595-603.
Kettunen, H., S. Peuranen, K. Tiihonen, and M. Saarinen. 2001b. Intestinal uptake of betaine in
vitro and the distribution of methyl groups from betaine, choline, and methionine in the body
of broiler chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 128:269-278.
Page 37
24
Kjaer, J. B., G. Su, B. L. Nielsen, and P. Sørensen. 2006. Foot pad dermatitis and hock burn in
broiler chickens and degree of inheritance. Poult. Sci. 85:1342-1348.
Klasing, K., K. Adler, J. Remus, and C. Calvert. 2002. Dietary betaine increases intraepithelial
lymphocytes in the duodenum of coccidia-infected chicks and increases functional properties
of phagocytes. J. Nutr. 132:2274-2282.
Lambert, G. P. 2004. Role of gastrointestinal permeability in exertional heatstroke. Exerc. Sport
Sci. Rev. 32:185-190.
Lambert, G. P. 2009. Stress-induced gastrointestinal barrier dysfunction and its inflammatory
effects. J. Anim. Sci. 87:E101-E108. doi 10.2527/jas.2008-1339
Lambert, G. P., C. V. Gisolfi, D. J. Berg, P. L. Moseley, L. W. Oberley, and K. C. Kregel. 2002.
Selected contribution: Hyperthermia-induced intestinal permeability and the role of oxidative
and nitrosative stress. J. Appl. Physiol. 92:1750-1761. doi 10.1152/japplphysiol.00787.2001
Le Kim, D., and H. Betzing. 1976. Intestinal absorption of polyunsaturated phosphatidylcholine
in the rat. Hoppe-Seyler´ s Z. Physiol. Chem. 357:1321-1332.
Lin, H., E. Decuypere, and J. Buyse. 2004. Oxidative stress induced by corticosterone
administration in broiler chickens (Gallus gallus domesticus): 1. Chronic exposure. Comp.
Biochem. Physiol. B Mol. Integr. Physiol. 139:737-744.
Mahmoud, K., and A. Yaseen. 2005. Effect of feed withdrawal and heat acclimatization on stress
responses of male broiler and layer-type chickens (Gallus gallus domesticus). Asian-Aust. J.
Anim. Sci. 18:1445.
Martland, M. 1985. Ulcerative dermatitis dm broiler chickens: The effects of wet litter. Avian
Pathol. 14:353-364.
Martrenchar, A., E. Boilletot, D. Huonnic, and F. Pol. 2002. Risk factors for foot-pad dermatitis
in chicken and turkey broilers in France. Prev. Vet. Med. 52:213-226.
Mayne, R., R. Else, and P. Hocking. 2007. High litter moisture alone is sufficient to cause
footpad dermatitis in growing turkeys. Br. Poult. Sci. 48:538-545.
McDevitt, R., S. Mack, and I. Wallis. 2000. Can betaine partially replace or enhance the effect of
methionine by improving broiler growth and carcase characteristics? Br. Poult. Sci. 41:473-
480.
McDowell, L. R. 1989. Choline. Pages 565-596 in Vitamins in Animal and Human Nutrition.
2nd ed.
McIlroy, S., E. Goodall, and C. McMurray. 1987. A contact dermatitis of broilers‐epidemiological findings. Avian Pathol. 16:93-105.
Page 38
25
Metzler-Zebeli, B. U., M. Eklund, and R. Mosenthin. 2009. Impact of osmoregulatory and
methyl donor functions of betaine on intestinal health and performance in poultry. Worlds
Poult. Sci. J. 65:419-442. doi 10.1017/s0043933909000300
Meyer Jr, E., D. Engel, and J. Cooper. 1982. Acetylation and phosphorylation of choline
following high or low affinity uptake by rat cortical synaptosomes. Neurochem. Res. 7:749-
759.
Mitchell, M., and A. Carlisle. 1992. The effects of chronic exposure to elevated environmental
temperature on intestinal morphology and nutrient absorption in the domestic fowl (Gallus
domesticus). Comp. Biochem. Physiol. Part A Physiol. 101:137-142.
Molitoris, B. A., and D. H. Baker. 1976. The choline requirement of broiler chicks during the
seventh week of life. Poult. Sci. 55:220-224.
Mongin, P., M. Larbier, N. Carbobaptista, D. Licois, and P. Coudert. 1976. Comparison of
osmotic pressures along digestive-tract of domestic-fowl and rabbit. Br. Poult. Sci. 17:379-
382. doi 10.1080/00071667608416291
Nasir, A., R. Moudgal, and N. Singh. 1999. Involvement of corticosterone in food intake, food
passage time and in vivo uptake of nutrients in the chicken (Gallus domesticus). Br. Poult.
Sci. 40:517-522.
NRC. 1994. Nutrient Requirements of Poultry. 9th rev ed. Natl. Acad. Press, Washington, DC.
Pearce, S., V. Mani, R. Boddicker, J. Johnson, T. Weber, J. Ross, L. Baumgard, and N. Gabler.
2012. Heat stress reduces barrier function and alters intestinal metabolism in growing pigs. J.
Anim. Sci. 90:257-259.
Pesti, G., A. Harper, and M. Sunde. 1979. Sulfur amino acid and methyl donor status of corn-soy
diets fed to starting broiler chicks and turkey poults. Poult. Sci. 58:1541-1547.
Petronini, P.-G., R. R. Alfieri, M. N. Losio, A. E. Caccamo, A. Cavazzoni, M. A. Bonelli, A. F.
Borghetti, and K. P. Wheeler. 2000. Induction of BGT-1 and amino acid system A transport
activities in endothelial cells exposed to hyperosmolarity. Am. J. Physiol. Regul. Intgr. Comp.
Physiol. 279:R1580-R1589.
Petronini, P. G., E. De Angelis, P. Borghetti, A. Borghetti, and K. P. Wheeler. 1992. Modulation
by betaine of cellular responses to osmotic stress. Biochem. J 282:69-73.
Pillai, P., A. Fanatico, M. Blair, and J. Emmert. 2006. Homocysteine remethylation in broilers
fed surfeit choline or betaine and varying levels and sources of methionine from eight to
twenty-two days of age. Poult. Sci. 85:1729-1736.
Plagemann, P. G. 1968. Choline metabolism and membrane formation in rat hepatoma cells
grown in suspension culture: I. Incorporation of choline into phosphatidylcholine of
mitochondria and other membranous structures and effect of metabolic inhibitors. Arch.
Biochem. Biophys. 128:70-87.
Page 39
26
Quinteiro-Filho, W., A. Ribeiro, V. Ferraz-de-Paula, M. Pinheiro, M. Sakai, L. Sá, A. Ferreira,
and J. Palermo-Neto. 2010. Heat stress impairs performance parameters, induces intestinal
injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 89:1905-1914.
Rafeeq, M., T. Pasha, N. Rashid, B. Hilal, and I. Shahzad. 2011. Effect of supplementation of
methionine, betaine and choline on the performance of broiler chicken in early life fed
methionine deficient ration. . J. Anim. Plant. Sci. 21:778-780.
Rector Jr, F. C., H. Buttram, and D. W. Seldin. 1962. An analysis of the mechanism of the
inhibitory influence of K+ on renal H+ secretion. J. Clin. Invest. 41:611.
Rojas-Cano, M., L. Lara, M. Lachica, J. Aguilera, and I. Fernandez-Figares. 2011. Influence of
betaine and conjugated linoleic acid on development of carcass cuts of Iberian pigs growing
from 20 to 50kg body weight. Meat. Sci. 88:525-530.
Sakomura, N. K., N. A. A. Barbosa, F. A. Longo, E. P. da Silva, M. A. Bonato, and J. B. K.
Fernandes. 2013. Effect of dietary betaine supplementation on the performance, carcass yield,
and intestinal morphometrics of broilers submitted to heat stress. Braz. J. Poultry Sci. 15:105-
111.
Schutte, J., J. De Jong, W. Smink, and M. Pack. 1997. Replacement value of betaine for DL-
methionine in male broiler chicks. Poult. Sci. 76:321-325.
Selye, H. 1976. Forty years of stress research: principal remaining problems and misconceptions.
Can. Med. Assoc. J. 115:53.
Shepherd, E., and B. Fairchild. 2010. Footpad dermatitis in poultry. Poult. Sci. 89:2043-2051.
Shini, S., P. Kaiser, A. Shini, and W. L. Bryden. 2008a. Biological response of chickens (Gallus
gallus domesticus) induced by corticosterone and a bacterial endotoxin. Comp. Biochem.
Physiol. B Mol. Integr. Physiol. 149:324-333.
Shini, S., P. Kaiser, A. Shini, and W. L. Bryden. 2008b. Differential alterations in ultrastructural
morphology of chicken heterophils and lymphocytes induced by corticosterone and
lipopolysaccharide. Vet. Immunol. Immunopathol. 122:83-93.
Smith, A., S. Rose, R. Wells, and V. Pirgozliev. 2000. Effect of excess dietary sodium,
potassium, calcium and phosphorus on excreta moisture of laying hens. Br. Poult. Sci. 41:598-
607.
Smith, M. O., and R. G. Teeter. 1987a. Effect of ammonium chloride and potassium chloride on
survival of broiler chicks during acute heat stress. Nutr. Res. 7:677-681.
Smith, M. O., and R. G. Teeter. 1987b. Potassium Balance of the 5 to 8-Week-Old Broiler
Exposed to Constant Heat or Cycling High Temperature Stress and the Effects of
Supplemental Potassium Chloride on Body Weight Gain and Feed Efficiency. Poult. Sci.
66:487-492. doi 10.3382/ps.0660487
Page 40
27
Smith, M. O., and R. G. Teeter. 1992. Effects of potassium chloride supplementation on growth
of heat-distressed broilers. J. Appl. Poult. Res. 1:321-324.
Soutyrine, A., M. Smith, and B. Sivanadian. 1998. Feed withdrawal, potassium chloride, and
carbonated water effects on broiler thermotolerance. J. Appl. Poult. Res. 7:138-143.
Storey, R., and R. W. Jones. 1975. Betaine and choline levels in plants and their relationship to
NaCl stress. Plant Sci. Lett. 4:161-168.
Stucki, W., and A. Harper. 1961. Importance of dispensable amino acids for normal growth of
chicks. J. Nutr. 74:377-383.
Sun, Z., X. Wang, X. Deng, Å. Lasson, R. Wallen, E. Hallberg, and R. Andersson. 1998. The
influence of intestinal ischemia and reperfusion on bidirectional intestinal barrier
permeability, cellular membrane integrity, proteinase inhibitors, and cell death in rats. Shock
10:203-212.
Sutherland, L., J. Cairney, M. Elmore, I. Booth, and C. Higgins. 1986. Osmotic regulation of
transcription: induction of the proU betaine transport gene is dependent on accumulation of
intracellular potassium. J. Bacteriol. 168:805-814.
Teeter, R., and M. Smith. 1986. High chronic ambient temperature stress effects on broiler acid-
base balance and their response to supplemental ammonium chloride, potassium chloride, and
potassium carbonate. Poult. Sci. 65:1777-1781.
Teeter, R. G., and T. Belay. 1996. Broiler management during acute heat stress. Anim. Feed Sci.
Technol. 58:127-142.
Tillman, P., and G. Pesti. 1986. The response of male broiler chicks to a corn-soy diet
supplemented with L-methionine, L-cystine, choline, sulfate, and vitamin B12. Poult. Sci.
65:1741-1748.
Tsiagbe, V., M. Cook, A. Harper, and M. Sunde. 1987. Enhanced immune responses in broiler
chicks fed methionine-supplemented diets. Poult. Sci. 66:1147-1154.
USDA. 2013. International Egg & Poultry Review. Pages 2 U.S Department of Agriculture.
Van Rossum, G., and C. Boyd. 1998. Sodium-dependent and-independent choline uptake by type
II epithelial cells from rat lung. J. Membr. Biol. 162:147-156.
Vendemiale, G., I. Grattagliano, P. Caraceni, G. Caraccio, M. Domenicali, M. Dall'Agata, F.
Trevisani, F. Guerrieri, M. Bernardi, and E. Altomare. 2001. Mitochondrial oxidative injury
and energy metabolism alteration in rat fatty liver: effect of the nutritional status. Hepatology
33:808-815.
Vincent , B., S. S. Vigneaud, J. P. Chandler, and M. Cohn. 1946. A further investigation of the
role of betaine in transmethylation reactions in vivo. The Journal of Biological Chemestry
165:639-648.
Page 41
28
Virtanen, E., M. Junnila, and A. Soivio. 1989. Effects of food containing betaine/amino acid
additive on the osmotic adaptation of young Atlantic salmon, Salmo salar L. Aquaculture
83:109-122.
Waldroup, P., and C. Fritts. 2005. Evaluation of separate and combined effects of choline and
betaine in diets for male broilers. Int. J. Poult. Sci 4:442-448.
Waldroup, P. W., M. A. Motl, F. Yan, and C. A. Fritts. 2006. Effects of betaine and choline on
response to methionine supplementation to broiler diets formulated to industry standards. J.
Appl. Poult. Res. 15:58-71.
Wecker, L. 1989. Dietary choline: a limiting factor for the synthesis of acetylcholine by the
brain. Adv. Neurol. 51:139-145.
Yalcin, S., S. Özkan, L. Türkmut, and P. Siegel. 2001. Responses to heat stress in commercial
and local broiler stocks. 1. Performance traits. Br. Poult. Sci. 42:149-152.
Yamauchi, A., S. Uchida, H. Kwon, A. Preston, R. B. Robey, A. Garcia-Perez, M. Burg, and J.
Handler. 1992. Cloning of a Na (+)-and Cl (-)-dependent betaine transporter that is regulated
by hypertonicity. J. Biol. Chem. 267:649-652.
Yang, W.-J., P. J. Rich, J. D. Axtell, K. V. Wood, C. C. Bonham, G. Ejeta, M. V. Mickelbart,
and D. Rhodes. 2003. Genotypic variation for glycinebetaine in sorghum. Crop Sci. 43:162-
169.
Yao, Z., and D. E. Vance. 1988. The active synthesis of phosphatidylcholine is required for very
low density lipoprotein secretion from rat hepatocytes. J. Biol. Chem. 263:2998-3004.
Zeisel, S. 1993. Choline phospholipids: signal transduction and carcinogenesis. FASEB J. 7:551-
557.
Zeisel, S. H., M.-H. Mar, J. C. Howe, and J. M. Holden. 2003. Concentrations of choline-
containing compounds and betaine in common foods. J. Nutr. 133:1302-1307.
Zeisel, S. H., T. Zola, K.-A. daCosta, and E. A. Pomfret. 1989. Effect of choline deficiency on S-
adenosylmethionine and methionine concentrations in rat liver. Biochem. J 259:725-729.
Zhan, X., J. Li, Z. Xu, and R. Zhao. 2006. Effects of methionine and betaine supplementation on
growth performance, carcase composition and metabolism of lipids in male broilers. Br. Poult.
Sci. 47:576-580.
Page 42
29
Chapter III: Performance of heat-stressed broilers supplemented with dietary choline and
betaine
Page 43
30
Abstract
A study was conducted to determine the effects of supplemental choline (CHO) and betaine
(BET) on broiler performance and carcass characteristics under different temperature conditions,
thermoneutral (TN: 23.9oC) and heat stress (HS: 28-36
oC). Eight hundred day-old mixed sex
Cobb500 (female) x Hubbard (M99 male) chicks were weighed, wing tagged and equally divided
into the two environments. Eight replicates of 10 chicks were randomly assigned to each of five
dietary treatments: treatment 1 (CON) basal diet; treatment 2 (CHO500) basal diet plus 500 ppm
methyl equivalents added CHO; treatment 3 (CHO1000) basal diet plus 1000 ppm methyl
equivalents added CHO; treatment 4 (BET500) basal diet plus 500 ppm methyl equivalents
added BET and treatment 5 (BET1000) basal diet plus 1000 ppm methyl equivalents added BET.
The corn-soy bean basal diet was formulated to meet NRC requirements for broilers. Feed intake
and body weight were recorded weekly by pen. On day 24, foot pads were assessed for health
and litter samples collected from each pen. At slaughter on day 52, breast meat was collected for
drip loss evaluation at 4 and 7 days post slaughter. Breast meat lightness (L*), redness (a*) and
yellowness (b*) were measured. Data were analyzed using mixed model analysis of variance
(SAS 9.3, Cary, NC). Results showed that HS birds consumed 20.59% less feed and gained
23.34% less weight than TN birds (P < 0.05) during 22-49 days of age. The overall feed intake
and weight gain during days 1-49 were similarly reduced in HS birds. During 1-21 days of age,
birds consuming BET500 (0.68) and BET1000 (0.67) had higher feed efficiency compared to
those consuming the CON (0.62). Heat stress decreased feed efficiency during days 22-49 (P =
0.02). HS birds (0.54) had lower feed efficiency when compared to TN birds (0.56). There was a
diet X temperature interaction (P = 0.04) on drip loss 4 days post slaughter. The lowest drip loss
occurred with CHO500 (0.60%) in HS and BET1000 (0.83%) in TN birds. Breast meat color of
Page 44
31
HS birds was significantly (P = 0.02) lighter (54.21) while that of TN birds was significantly (P
= 0.004) more yellow (3.75). Temperature did not affect pododermatitis (P = 0.22) however,
there was an effect of diet (P = 0.003) with CHO500 and BET1000 showing the lowest
occurrence. In this study, breast meat drip loss was influenced by dietary CHO and BET while
meat color was affected by rearing temperature. Dietary CHO and BET supplementation did not
improve weight loss induced by HS conditions.
Introduction
Heat stress is one of the major problems facing the poultry industry (St-Pierre, et al.,
2003). During high environmental temperature and high relative humidity, chickens adapt by
reducing feed intake to decrease endogenous heat production and hyperventilating or panting to
eliminate extra heat. However, the results are detrimental for physiology with slow growth rate,
low body weight gain (Cooper and Washburn, 1998; Donkoh, 1989; Quinteiro-Filho, et al.,
2010) and respiratory alkalosis (Borges, et al., 2004; Bottje and Harrison, 1985). Several
management practices have been developed and used to improve birds’ comfort, survival and to
mitigate the negative impact of heat stress on broilers. These include housing insulation,
ventilation, evaporative cooling systems, feed withdrawal (Mahmoud and Yaseen, 2005;
Soutyrine, et al., 1998), fasting before the onset of heat stress and acclimation of high
temperature in younger age (De Basilio, et al., 2001; Yalcin, et al., 2001). These practices don’t
directly address the acid-base imbalance (respiratory alkalosis) even though they improve
broilers performances.
Respiratory alkalosis caused by excessive loss of carbon dioxide due to hyperventilation
(Borges, et al., 2007; Teeter and Smith, 1986) subsequently results in electrolyte imbalance in
Page 45
32
chickens primarily due to potassium excretion. Smith et al (1987) reported a 633% increase of
K+ excretion via excreta during heat stress (35
oC) compared to thermoneutral (24
oC)
environment. Supplementing heat-stressed broilers with essential electrolytes could be beneficial
in correcting acid-base imbalance thereby improving broilers comfort and performance.
Supplementation of water with potassium chloride (KCl) was shown to increase body weight
gain (Ahmad, et al., 2008; Smith and Teeter, 1987) of heat-stressed broilers. Dietary electrolytes
balance for maximum production was estimated to be 240 mEq/kg (Borges, et al., 2003).
However, close attention must be paid to the use of dietary electrolytes to alleviate the negative
impacts of heat stress on broilers. Addition of the electrolytes Na+, Cl
-, and K
+ in drinking water
was shown to cause an increase in water consumption that resulted in litter wetness, the primary
cause of foot pad dermatitis (Berg and Algers, 2004). Additionally, excess accumulation of Na+
and Cl- may inhibit cells proliferation and protein synthesis as opposed to organic osmolytes
(Petronini, et al., 1992).
Organic osmolytes such as choline and betaine could be alternatives to electrolytes.
Currently, betaine is not required in a typical broiler diet, however choline is usually added at
different rates depending on age: 1300 mg/kg for 0-3 weeks of age, 1000 mg/kg for 3-6 weeks of
age and 750 mg/kg for 6-9 weeks of age (NRC., 1994). Betaine can be synthesized from choline
and has been shown to affect the movement of water across small intestine of broiler chicks
(Kettunen, et al., 2001). The supplementation of choline and betaine in broiler diet may be
beneficial in improving osmotic pressure and decreasing excreta moisture in heat-stressed
chickens and could improve their performance and foot health. Even though the effect of
supplemental choline and betaine on broiler performance has been the objective of several
studies, very little has been done concerning their osmoprotectant benefit if any, under heat stress
Page 46
33
conditions. Therefore, this study was designed to evaluate the impact of dietary supplementation
of choline and betaine on the performance and carcass characteristics of broilers under HS
conditions.
Materials and Methods
Birds and Housing
The procedures were reviewed and approved by the University of Tennessee Institutional
Animal Care and Use Committee (IACUC), and the research was conducted at the Johnson
Animal Research and Teaching Unit (JARTU). Two adjacent rooms, thermoneutral (TN) and
heat stress (HS) were used. Each room was divided into 40 pens equipped with individual gravity
feeders, water nipple line and bedding made of shredded paper. The rooms were equipped with
HOBO®1
data loggers to record daily temperature and humidity. In addition, a humidifier was
placed in the heat stress room to ensure that humidity remained above 50%. Eight hundred day-
old mixed sex Cobb500 (female) x Hubbard (M99 male) chicks were obtained from Pilgrim’s
Pride hatchery in Cohutta, Georgia. They were vaccinated at the hatchery against Marek’s and
Gumboro diseases (VAXXITEK HVT-IBD); Escherichia coli, Salmonella typhimuriun and
Pseudomonas aeruginosa (GARASOL); Coccidiosis (COCCIVAC-B), Newcastle
(NEWHATCH C2-M) and Infectious bronchitis (MILDVAC ARK and GA 2008). Upon arrival,
the birds were weighed by groups of ten and assigned to one of each treatment.
1 Onset Computer Corporation
470 MacArthur Blvd. Bourne, MA 02532
Page 47
34
Temperature Treatments
During the first 21 days, birds were brooded under similar conditions in the two rooms.
The temperature was set to 32.22oC the first week and decreased by 2.78
oC until it reached
23.9oC by the end of the third week. From day 22 to the end of the study, the temperature was
maintained at 23.9 oC in the TN room and cycled from 28
oC to 36
oC between 10:00 a.m. and
8:00 p.m. in the HS room (Figure 1). The photoperiod regimen was 23 hours of light followed by
1 hour of dark in both environments. The humidity was maintained at 50% in the HS room.
Figure 1. Typical daily temperature in TN (23.9 ±1°C) and HS (28-36±1°C) rooms.
10
15
20
25
30
35
40
Tem
p. °C
Time
HS
TN
Page 48
35
Dietary Treatments
Five dietary treatments were tested. The basal formulation with choline or betaine
supplementation was used to make the following dietary treatments:
Control (CON), basal formulation
Low choline (CHO500), basal diet plus 500 ppm methyl equivalents added choline
High choline (CHO1000), basal diet plus 1000 ppm methyl equivalents added choline
Low betaine (BET500), basal diet plus 500 ppm methyl equivalents added betaine
High betaine (BET1000), basal diet plus 1000 ppm methyl equivalents added betaine
The basal diet was formulated to meet broilers energy, protein, minerals and vitamins
requirements (NRC., 1994) except for methionine which was 20% lower. The birds were fed
starter diet (Table 1) from days 1-21, grower diet (Table 2) from days 22-35 and finisher diet
(Table 3) from days 36-49.
Growth Performance
The feed was weighed and added every three days to ensure continuous access. Once per
week, the remaining feed was weighed to determine feed intake. Similarly, the birds were
weighed per pen to determine body weight gain. When a bird died, the carcass weight was
recorded and accounted for in the calculation of weekly average weight gain and feed intake.
Litter Samples and Foot Pad Dermatitis
The litter made of shredded paper was not changed during the entire experiment. On day
42, litter samples were collected at approximately the same area between the nipple lines and the
feeders and stored at -20oC for later determination of moisture content. On day 42, all the birds
were scored for foot pad dermatitis. The pad of both feet were observed, compared to necrotic
patterns and ranked using the following scores: 0-no lesions, 1-small lesion, 2-moderate lesion in
Page 49
36
Table 1. Nutritional composition of starter diet (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 62.86 62.86 62.86 62.86 62.86
Soybean meal 31.00 31.00 31.00 31.00 31.00
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.20 0.105 0.01 0.143 0.0853
Choline, mg/kg2 0.00 0.095 0.19 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.15 0.15 0.15 0.15 0.15
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 2.10 2.10 2.10 2.10 2.10
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 1.90 1.90 1.90 1.90 1.90
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3024.00 3024.00 3024.00 3024.00 3024.00
Crude protein, % 20.59 20.59 20.59 20.59 20.59
Calcium, % 0.98 0.98 0.98 0.98 0.98
Available Phosphorus, % 0.51 0.51 0.51 0.51 0.51
Potassium, % 0.81 0.81 0.81 0.81 0.81
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 92.99 92.99 92.99 92.99 92.99
Choline, mg/kg 1236.38 2186.44 3136.44 1236.38 1236.38
Betaine, mg/kg 0.00 0.00 0.00 585.01 1147.03
Lysine, % 1.18 1.18 1.18 1.18 1.18
Methionine, % 0.47 0.47 0.47 0.47 0.47
Methionine +Cysteine, % 0.81 0.81 0.81 0.81 0.81
Threonine, % 0.76 0.76 0.76 0.76 0.76
Fat, % 4.60 4.60 4.60 4.60 4.69
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 50
37
Table 2. Nutritional composition of grower diet (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 62.32 62.32 62.32 62.32 62.32
Soybean meal 31.00 31.00 31.00 31.00 31.00
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.30 0.1884 0.0768 0.243 0.1853
Choline, mg/kg2 0.00 0.1116 0.2232 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.20 0.20 0.20 0.20 0.20
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 1.69 1.69 1.69 1.69 1.69
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 2.70 2.70 2.70 2.70 2.70
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3072.00 3074.00 3072.00 3072.00 3072.00
Crude protein, % 19.95 19.95 19.95 19.95 19.95
Calcium, % 0.89 0.89 0.89 0.89 0.89
Available Phosphorus, % 0.43 0.43 0.43 0.43 0.43
Potassium, % 0.80 0.80 0.80 0.80 0.80
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 92.48 92.49 92.48 92.48 92.48
Choline, mg/kg 1233.03 2349.37 3465.10 1233.02 1233.03
Betaine, mg/kg 0.00 0.00 0.00 585.01 1147.03
Lysine, % 1.18 1.18 1.18 1.18 1.18
Methionine, % 0.52 0.52 0.52 0.52 0.52
Methionine +Cysteine, % 0.85 0.85 0.85 0.85 0.85
Threonine, % 0.76 0.76 0.76 0.76 0.76
Fat, % 5.38 5.38 5.38 5.38 5.38
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 51
38
Table 3. Nutritional composition of finisher diet (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 66.00 66.00 66.00 66.00 66.00
Soybean meal 27.37 27.37 27.37 27.37 27.37
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.30 0.1884 0.0768 0.243 0.1853
Choline, mg/kg2 0.00 0.1116 0.2232 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.15 0.15 0.15 0.15 0.15
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 1.69 1.69 1.69 1.69 1.69
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 2.70 2.70 2.70 2.70 2.70
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3105.00 3107.00 3105.00 3108.00 3105.00
Crude protein, % 18.55 18.55 18.55 18.55 18.55
Calcium, % 0.88 0.88 0.88 0.88 0.88
Available Phosphorus, % 0.43 0.43 0.43 0.43 0.43
Potassium, % 0.74 0.74 0.74 0.74 0.74
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 91.15 91.16 91.15 91.16 91.15
Choline, mg/kg 1156.71 2273.05 3388.78 1157.25 1156.71
Betaine, mg/kg 0.00 0.00 0.00 569.98 1147.03
Lysine, % 1.08 1.08 1.08 1.08 1.08
Methionine, % 0.45 0.45 0.45 0.45 0.45
Methionine +Cysteine, % 0.77 0.77 0.77 0.77 0.77
Threonine, % 0.70 0.70 0.70 0.70 0.70
Fat, % 5.48 5.48 5.48 5.49 5.48
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 52
39
one or both feet, and 3-extensive lesion in one or both feet (Figure 2). The scoring was done by
the same person for consistency. To determine litter moisture, samples were thawed at room
temperature, weighed and dried in the oven for 48 hours at 16oC. After 48 hours, the samples
were weighed and moisture content determined as difference between wet and dried litter weight
and expressed as a percent of wet litter.
Figure 2. Scoring system for foot pad dermatitis (Mello, et al., 2011).
Page 53
40
Processing of Birds
Prior to harvesting on day 52, birds were deprived of feed for 12 hours to allow for the
emptying of the digestive track. Eighty birds (eight birds randomly selected from each treatment
per room), were weighed, hung on shackles, stunned with an electric knife, and then killed by
severing the jugular vein. They were allowed to bleed for 3 minutes then scalded in hot water
(59.44 oC) for 40 seconds, and their feathers removed using a de-feathering machine. Birds were
then eviscerated and carefully dissected. The carcasses were weighed and the breast meat
separated. The breasts were cut into half, weighed, individually bagged and chilled on ice for
later drip loss determination.
Drip Loss
The left half of each breast was hung in the refrigerator at 4oC for drip loss determination.
The breasts were weighed at day four and again at day seven post slaughter. Drip loss constitutes
water loss during storage and was determined by the difference in weights between two periods
of storage time. For four days post slaughter, drip loss was determined by taking the difference in
breast weights between days four and one and is expressed as a percent of day one breast weight.
Drip loss seven days post slaughter is determined using the same principle and expressed as a
percent of day one breast weight.
Breast Meat Color
Color of breast meat was determined using photo colorimeter (Konica Minolta)2 and
reported using L*a*b* color space (CIELAB) from CIE (International Commission on
Illumination). L* represents lightness and the breast meat is considered light when L* > 50.0 and
2 Konica Minolta, Inc. JP Tower,
272 Marunouchi Chiyoda-ku, Tokyo 100-7015, Japan
Page 54
41
dark when L* < 45.0. Redness (a*) and yellowness (b*) represent the chromaticity coordinates.
Terms a* and b* indicate color direction: the more positive a* the higher the redness and the
more negative a* the greener the color, the more positive b* the more yellow and the more
negative b* the more blue the color (McGuire, 1992). Each color value is given by the average of
three measurements on an individual breast sample.
Statistical Analysis
The experimental design was a completely randomized design split plot with temperature
in the whole plots and diet in the subplots. Data were analyzed using mixed model analysis of
variance procedure SAS 9.3 (SAS Institute, Cary, NC, USA) and least squares means compared
using protected LSD at 5% level of significance. Data on foot pad dermatitis were analyzed
using contingency table.
Results
Growth Performance
Average feed consumption for days 1-21, 22-49 and 1-49 are presented in Table 4. There
was no diet or temperature effect nor was there an interaction on feed intake during days 1-21 (P
> 0.20). During days 22-49 there was no diet X temperature interaction on feed intake (P = 0.17).
However, temperature significantly affected feed intake (P = 0.001) with thermoneutral bird
(4.08 kg) consuming in average 20.59% more than heat stress bird (3.24 kg). The feed
consumption during days 1-49 was similarly affected only by temperature (P = 0.0001). The
thermoneutral bird average consumption (4.98 kg) was 17.67% higher than that of the heat-
stressed bird (4.10 kg).
Page 55
42
Table 4. Effect of choline and betaine supplementation on average feed intake (kg) 1
of heat-
stressed broilers during 1-21, 22-49 and 1-49 days of age
Diet Temp.2
Day 1-21 Day 22-49 Day 1-49
CON TN 0.90
3.98 4.86
HS 0.82 3.33 4.15
CHO500 TN 0.90 4.13 5.04
HS 0.84 3.27 4.11
CHO1000 TN 0.92 4.03 4.96
HS 0.89 3.30 4.19
BET500 TN 0.85 3.99 4.83
HS 0.89 3.31 4.19
BET1000 TN 0.92 4.27 5.18
HS 0.86 3.00 3.86
SEM 0.047
0.16 0.18
Effect P values
Diet 0.25 0.99 0.99
Temp 0.86 0.001 0.0001
Diet x Temp 0.73 0.17 0.26
Main Effect Means
Diet
CON 0.86 3.65
4.51
CHO500 0.87 3.70 4.57
CHO1000 0.91 3.67 4.57
BET500 0.87 3.65 4.51
BET1000 0.89 3.64 4.52
SEM 0.033 0.11 0.13
Temp
TN 0.90 4.08a
4.98a
HS 0.86 3.24b
4.10b
SEM 0.023 0.098 0.11 1 Values are least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b,c Means within same column followed by different letters differ significantly (P < 0.05).
Page 56
43
Body weight gains are reported in Table 5. There was no diet or temperature effect nor
was there their interaction on body weight gain during the starter period (P > 0.50). During days
22-49, only temperature significantly affected body weight gain (P = 0.001). The average body
weight gain of heat-stressed bird (1.74 kg) was 23.35% lower compared to thermoneutral bird
(2.27 kg). During days 1-49, body weight gain was similarly affected by heat stress (P = 0.001)
with heat-stressed birds (2.86 kg) gaining 19.23% less weight that the thermoneutral bird (2.31
kg).
There was no diet X temperature interaction on feed efficiency (Table 6). During days 1-
21, BET500 (0.68) and BET1000 (0.67) increased (P = 0.04) feed efficiency compared to the
CON (0.62). During days 22-49 only temperature increased feed efficiency (P = 0.02). Heat-
stressed birds had lower feed efficiency (0.54) compared to TN birds (0.56). Overall during days
1-49, temperature tended to decrease feed efficiency (P = 0.05).
The mortality rate (Table 7) was not affected by diet or diet X temperature interaction.
However, mortality in the heat stress treatment (11.25%) was significantly higher than that of the
thermoneutral environment (3%) (P = 0.0006).
Litter Moisture
Litter moisture content expressed as a percent of wet litter weight is reported in Table 8.
There was no diet X temperature interaction on litter moisture (P = 0.17). Litter moisture was
significantly lower (P = 0.0001) in the heat stress (21.31%) room compared to thermoneutral
room (42.73%). CHO500 (29.73%) and BET1000 (29.73%) tended to lower litter moisture (P =
0.07).
Page 57
44
Table 5. Effect of choline and betaine supplementation on average body weight gain (kg) 1
of
heat-stressed broilers during 1-21, 22-49 and 1-49 days of age
Diet Temp.2
Day 1-21 Day 22-49 Day 1-49
CON TN 0.57 2.19 2.75
HS 0.52 1.80 2.32
CHO500 TN 0.58 2.33 2.91
HS 0.55 1.76 2.31
CHO1000 TN 0.61 2.24 2.85
HS 0.60 1.78 2.38
BET500 TN 0.57 2.27 2.84
HS 0.61 1.77 2.38
BET1000 TN 0.64 2.30 2.94
HS 0.57 1.59 2.15
SEM 0.042 0.091 0.11
Effect P values
Diet 0.46 0.82 0.87
Temp 0.53 0.001 0.001
Diet x Temp 0.65 0.35 0.36
Main Effect Means
Diet
CON 0.54 1.99 2.54
CHO500 0.57 2.04 2.61
CHO1000 0.60 2.01 2.61
BET500 0.59 2.02 2.61
BET1000 0.60 1.94 2.54
SEM 0.029 0.065 0.077
Temp
TN 0.59 2.27a
2.86a
HS 0.57 1.74b
2.31b
SEM 0.024 0.052 0.065 1 Values are least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Page 58
45
Table 6. Effect of choline and betaine supplementation on feed efficiency (Gain:feed) 1 of heat-
stressed broilers during 1-21, 22-49 and 1-49 days of age
Diet Temp.2
Day 1-21 Day 22-49 Day 1-49
CON TN 0.63 0.55 0.56
HS 0.62 0.54 0.56
CHO500 TN 0.64 0.56 0.58
HS 0.66 0.54 0.56
CHO1000 TN 0.66 0.55 0.57
HS 0.66 0.54 0.57
BET500 TN 0.68 0.57 0.59
HS 0.68 0.53 0.57
BET1000 TN 0.69 0.54 0.57
HS 0.65 0.53 0.56
SEM 0.022 0.013 0.01
Effect P values
Diet 0.04 0.70 0.42
Temp 0.82 0.02 0.05
Diet x Temp 0.58 0.78 0.87
Main Effect Means
Diet
CON 0.62b
0.55 0.56
CHO500 0.65ab
0.55 0.57
CHO1000 0.66ab
0.55 0.57
BET500 0.68a
0.55 0.58
BET1000 0.67a
0.53 0.56
SEM 0.015 0.009 0.006
Temp
TN 0.66 0.56a 0.57
HS 0.66 0.54b 0.56
SEM 0.014 0.006 0.004 1 Values are least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Page 59
46
Table 7. Effect of choline and betaine supplementation on mortality (%) 1
of heat-stressed
broilers during 1-49 days of age
Diet Temp.2
Mortality
CON TN 7.50
HS 12.5
CHO500 TN 2.50
HS 13.75
CHO1000 TN 1.25
HS 3.75
BET500 TN 2.50
HS 11.25
BET1000 TN 1.25
HS 15.00
SEM 3.64
Effect P values
Diet 0.32
Temp 0.0006
Diet x Temp 0.54
Main Effect Means
Diet
CON 10.00
CHO500 8.13
CHO1000 2.50
BET500 6.88
BET1000 8.13
SEM 2.58
Temp
TN 3.00b
HS 11.25a
SEM 1.63 1 Values are least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Page 60
47
Table 8. Effect of choline and betaine supplementation to broilers on litter moisture content1
under thermoneutral and heat stress conditions on day 42
Diet Temp.2
Day 42
CON TN 41.43
HS 26.32
CHO500 TN 41.30
HS 18.16
CHO1000 TN 45.39
HS 25.40
BET500 TN 42.67
HS 18.16
BET1000 TN 42.87
HS 15.72
SEM 2.86
Effect P values
Diet 0.07
Temp 0.0001
Diet x Temp 0.17
Main Effect Means
Diet
CON 33.88
CHO500 29.73
CHO1000 35.40
BET500 31.82
BET1000 29.30
SEM 2.02
Temp
TN 42.73a
HS 21.31b
SEM 1.86 1 Values are least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Page 61
48
Foot Pad Dermatitis
Foot pad lesion score is presented in Table 9. There was no diet X temperature interaction
on foot pad lesion score (P > 0.05). Scores did not differ among temperature treatments (P =
0.22) but were affected by diets (P = 0.003). The pairwise comparisons showed that the pattern
of foot pad lesion scores for the CON did not differ from that of the rest of the diets. However,
BET1000 and CHO500 showed similar pattern with the highest score (score 0; 68.55% and
67.27 % respectively).
Breast Meat Color
The mean L*, a* and b* values for breast meat are presented Table 10. There was no diet
or diet X temperature interaction on lightness and yellowness. Lightness (54.51) was
significantly higher (P = 0.02) for HS birds compared to TN (51.73). Conversely, yellowness
was significantly higher (P= 0.004) for TN (3.75) compared to HS (2.60) birds.
Breast Meat Drip Loss
Drip loss assessed on days four and seven post slaughter is presented in Table 11. There
was diet X temperature interaction for day four (P = 0.04). The lowest drip loss occurred for
BET1000 (0.80%) and CHO500 (0.60%) in thermoneutral and heat stress environments
respectively. For seven days post slaughter, drip loss was not significantly affected by diet,
temperature or by their interaction (P > 0.30).
Page 62
49
Table 9. Effect of choline and betaine supplementation on foot pad score (%) 1 of heat-stressed
broilers on day 42
Foot pad lesion score
Temp.2
0 1 2 3
HSa
58.81 15.80 16.06 9.33
TNa
58.16 20.92 13.27 7.65
Diet2
CON abc
56.29 19.87 14.57 9.27
CHO500 ab
67.27 15.76 9.70 7.27
CHO1000 c 47.30 19.59 23.65 9.46
BET500 bc
51.61 21.94 18.06 8.39
BET1000 b
68.55 15.09 8.18 8.18 1 Values are least squares means.
2 Pairwise comparisons among temperature and dietary treatments respectively,
TN = Thermoneutral
environment, HS = Heat stress environment a Temp followed by different letters differ significantly in foot pad lesion score patterns (P < 0.05).
a,b,c Diet followed by different letters differ significantly in foot pad lesion score patterns P< 0 .05).
Page 63
50
Table 10. Effect of choline and betaine supplementation on breast meat color (L*, a*, b*) 1
from
heat-stressed broilers
Diet Temp L* a* b*
CON TN 53.09 2.84 4.33
HS 55.41 2.16 2.58
CHO500 TN 51.56 2.66 3.27
HS 53.19 2.73 2.01
CHO1000 TN 50.36 2.97 4.04
HS 54.57 3.06 3.43
BET500 TN 52.60 2.96 4.47
HS 54.05 2.48 3.08
BET1000 TN 51.04 2.88 2.64
HS 53.84 2.86 1.92
SEM 1.18 0.36 0.62
Effect P values
Diet 0.37 0.46 0.06
Temp 0.02 0.64 0.004
Diet x Temp 0.73 0.72 0.88
Main Effect Means
Diet
CON 54.25 2.50 3.45
CHO500 52.38 2.69 2.64
CHO1000 52.46 3.02 3.73
BET500 53.32 2.72 3.78
BET1000 52.46 2.87 2.28
SEM 0.84 0.26 0.43
Temp
TN 51.73b
2.86a
3.75a
HS 54.21a
2.66a
2.60b
SEM 0.67 0.19 0.28 1 Values are least squares means. L* = lightness; a* = redness; b* = yellowness
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Page 64
51
Table 11. Effect of choline and betaine supplementation on drip loss from breast meat (%) 1 of
heat-stressed broilers 4 and 7 days post slaughter
Diet Temp.2
Day 4 Post
Slaughter
Day 7 Post
Slaughter
CON TN 0.98ab
1.80
HS 0.92ab
1.80
CHO500 TN 1.00ab
1.74
HS 0.60b
1.13
CHO1000 TN 1.25ab
2.06
HS 0.90ab
1.89
BET500 TN 1.62a
2.10
HS 0.83b
1.18
BET1000 TN 0.80b
1.61
HS 1.55a
2.13
SEM 0.28
0.37
Effect P values
Diet 0.39 0.62
Temp 0.35 0.32
Diet x Temp 0.04 0.35
Main Effect Means
Diet
CON 0.94 1.80
CHO500 0.80 1.43
CHO1000 1.08 1.97
BET500 1.23 1.64
BET1000 1.18 1.87
SEM 0.17 0.26
Temp
TN 1.13 1.86
HS 0.96 1.63
SEM 0.13 0.16 1 Values are least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Page 65
52
Discussion
This study was conducted to evaluate the impact of dietary supplementation with choline
and betaine on heat-stressed broilers. The performance parameters evaluated were weight gain,
feed consumption, feed efficiency and mortality. Numerous studies have reported the effect of
high environmental temperature on the performance of broilers. Heat stress has been shown to
negatively impact broiler chicken performance (Dale and Fuller, 1980; Smith and Teeter, 1987).
In the current study, heat stress decreased feed intake and weight gain during days 22-49. The
reduction in feed consumption could be due to an effort by the bird to reduce heat generated
from feed consumption, digestion and metabolism. Feed efficiency was affected by diet during
the starter phase. BET500 and BET1000 showed higher feed efficiency compared to the CON.
During 22-49 days of age, feed efficiency was decreased only by heat stress. These results are
consistent with those reported by Bartlett and Smith (2003) who found that broilers under cyclic
and chronic heat stress (23.9oC to 37
oC) had lower feed intake compared to thermoneutral birds
(23.9oC).
In this study dietary supplementation of choline and betaine to provide 500 ppm and 1000
ppm methyl equivalents to a diet low in methionine did not improve weight gain, feed intake nor
feed efficiency during the grower and finisher periods. Waldroup, et al. (2006) evaluated choline
and betaine sparing effect of methionine by adding the 500 mg or 1000 mg of choline or betaine
to the diet and reported an improvement in feed intake in birds reared in thermoneutral
environment. Esteve-Garcia and Mack (2000) fed methionine diet supplemented with 0.5 g/kg
betaine and found no positive effect on broiler performance. Similarly Rafeeq, et al. (2011)
found that addition of 700 mg/kg betaine and choline to a methionine deficient diet did not
improve performance. According to Baker, et al. (1983), the beneficial effect of choline
Page 66
53
regarding its methionine sparing effect is possible only in a diet already containing a level of
choline and deficient in methionine. Although methionine level was deliberately decreased
compared to NRC requirements, choline sparing effect of methionine and its translation into
weight gain was not apparent in this study.
This study showed that mortality rate was increased by heat stress conditions. The
introduction of the temperature treatment resulted in 11.25% mortality compared to 3.00% in
thermoneutral environment. De Basilio, et al. (2001) obtained a higher mortality rate of 38%
with thermal challenged chickens. This could be due to the difference between the temperatures
used in both studies. In the current study the maximum temperature in the heat stress room was
36oC while 38
oC was used by De Basilio, et al. (2001). Supplementing diets with choline and
betaine did not reduced mortality rate. This is consistent with the results reported by Waldroup
and Fritts (2005) who found no effect of choline and betaine on broilers mortality.
Litter moisture was reduced by 50% in the heat stress room. This is not surprising due to
the fact that in the heat stress environment, the temperature cycled between 28 and 36oC. This
allowed the moisture in the litter to evaporate and the bedding to remain drier. Litter wetness has
been positively linked to foot pad dermatitis (Greene, et al., 1985; Mayne, et al., 2007). Hence, it
is very important to maintain litter dryness. Various factors can increase litter wetness and
contribute to the development of foot pad dermatitis (Francesch and Brufau, 2004). These
include infectious diseases, dietary composition and increase water consumption due electrolytes
supplementation (Lichtorowicz, et al., 2012). Water consumption was not measured in this study.
In evaluating the impact of choline and betaine supplementation on litter wetness, results showed
that CHO500 and BET1000 tended to decrease litter moisture compared to the CON. The trend
was similar in food pad scores. Even though CHO500 and BET1000 did not statistically differ
Page 67
54
from the CON, they showed less incidence of foot pad dermatitis (highest score 0). Chicken feet
health is a welfare issue, and it has been shown that turkey poults with severe necrosis develop
pain induced inappetance resulting in decrease feed intake and weight gain (Martland, 1984).
Additionally, there is an increasing market of chickens’ paws in China and Japan and keeping the
highest 0 score is economically important for poultry producers.
Meat color and drip loss are important quality parameters that could influence
consumers’ selection. The results of this study showed that choline and betaine supplementation
did not affect breast meat color nor was there a diet X temperature interaction. However, heat
stress increased breast meat lightness and decreased yellowness. Similarly, in an experiment
conducted by Akşit, et al. (2006) in which broilers were exposed to cyclic temperature, breast
meat lightness was increased by heat stress. However, heat stress did not affect meat yellowness
in their study. There was a diet X temperature interaction for breast meat drip loss. This was
caused by BET1000 increasing drip loss by 0.75 (0.80 % TN to 1.55 % HS) while CHO500 and
BET500 decreased it by 0.4 (1.00% TN to 0.60 % HS) and 0.79 (1.62 % TN to 0.83 % HS)
respectively. This suggests that the supplementation of BET1000 under thermoneutral
environment and CHO500 under heat stress could reduce water loss during short term storage
(up to four days). The supplementation of BET1000 under heat stress conditions may have
caused betaine to effectively accumulate in cells and help retain water in heat-stressed birds. This
could explain the higher drip loss of breast meat from heat-stressed birds. In order to fully
appreciate the negative impact of heat stress on the meat quality, further research need to be done
including the determination of the meat pH, cook loss and shear force.
Page 68
55
References
Ahmad, T., T. Khalid, T. Mushtaq, M. Mirza, A. Nadeem, M. Babar, and G. Ahmad. 2008.
Effect of potassium chloride supplementation in drinking water on broiler performance under
heat stress conditions. Poult. Sci. 87:1276-1280.
Akşit, M., S. Yalcin, S. Özkan, K. Metin, and D. Özdemir. 2006. Effects of temperature during
rearing and crating on stress parameters and meat quality of broilers. Poult. Sci. 85:1867-
1874.
Baker, D., K. Halpin, G. Czarnecki, and C. Parsons. 1983. The choline-methionine
interrelationship for growth of the chick. Poult. Sci. 62:133-137.
Bartlett, J. R., and M. O. Smith. 2003. Effects of different levels of zinc on the performance and
immunocompetence of broilers under heat stress. Poult. Sci. 82:1580-1588.
Berg, C., and B. Algers. Year. The effect of floor heating and feed protein level on the incidence
of foot pad dermatitis in turkeys poults. Proc. EAAP-55th Annual Meeting L.
Borges, S., A. F. Da Silva, J. Ariki, D. Hooge, and K. Cummings. 2003. Dietary electrolyte
balance for broiler chickens under moderately high ambient temperatures and relative
humidities. Poult. Sci. 82:301-308.
Borges, S. A., A. V. F. Da Silva, and A. Maiorka. 2007. Acid-base balance in broilers. Worlds
Poult. Sci. J. 63:73-81. doi Doi 10.1079/Wps2005128
Borges, S. A., A. V. F. da Silva, A. Majorka, D. M. Hooge, and K. R. Cummings. 2004.
Physiological responses of broiler chickens to heat stress and dietary electrolyte balance
(sodium plus potassium minus chloride, milliequivalents per kilogram). Poult. Sci. 83:1551-
1558.
Bottje, W. G., and P. C. Harrison. 1985. The effect of tap water, carbonated water, sodium
bicarbonate, and calcium chloride on blood acid-base balance in cockerels subjected to heat
stress. Poult. Sci. 64:107-113.
Cooper, M. A., and K. W. Washburn. 1998. The relationships of body temperature to weight
gain, feed consumption, and feed utilization in broilers under heat stress. Poult. Sci. 77:237-
242. doi 10.1093/ps/77.2.237
Dale, N. M., and H. L. Fuller. 1980. Effect of Diet Composition on Feed Intake and Growth of
Chicks Under Heat Stress.: II. Constant vs. Cycling Temperatures. Poult. Sci. 59:1434-1441.
doi 10.3382/ps.0591434
De Basilio, V., M. Vilarino, S. Yahav, and M. Picard. 2001. Early age thermal conditioning and
a dual feeding program for male broilers challenged by heat stress. Poult. Sci. 80:29-36.
Page 69
56
Donkoh, A. 1989. Ambient temperature: a factor affecting performance and physiological
response of broiler chickens. Int. J. Biometeorol. 33:259-265.
Esteve-Garcia, E., and S. Mack. 2000. The effect of DL-methionine and betaine on growth
performance and carcass characteristics in broilers. Anim. Feed Sci. Technol. 87:85-93.
Francesch, M., and J. Brufau. 2004. Nutritional factors affecting excreta/litter moisture and
quality. Worlds Poult. Sci. J. 60:64-75. doi 10.1079/wps20035
Greene, J. A., R. M. McCracken, and R. T. Evans. 1985. A contact dermatitis of broilers -clinical
and pathological findings. Avian Pathol. 14:23-38. doi 10.1080/03079458508436205
Kettunen, H., S. Peuranen, and K. Tiihonen. 2001. Betaine aids in the osmoregulation of
duodenal epithelium of broiler chicks, and affects the movement of water across the small
intestinal epithelium in vitro. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 129:595-603.
Lichtorowicz, K., J. Jankowski, Z. Zdunczyk, and J. Juskiewicz. 2012. The effect of different
dietary sodium levels on blood electrolytes, growth performance and foot pad dermatitis
incidence in turkeys. J. Elem. 17.
Mahmoud, K., and A. Yaseen. 2005. Effect of feed withdrawal and heat acclimatization on stress
responses of male broiler and layer-type chickens (Gallus gallus domesticus). Asian-Aust. J.
Anim. Sci. 18:1445.
Martland, M. 1984. Wet litter as a cause of plantar pododermatitis, leading to foot ulceration and
lameness in fattening turkeys. Avian Pathol. 13:241-252.
Mayne, R., R. Else, and P. Hocking. 2007. High litter moisture alone is sufficient to cause
footpad dermatitis in growing turkeys. Br. Poult. Sci. 48:538-545.
McGuire, R. G. 1992. Reporting of objective color measurements. HortScience 27:1254-1255.
Mello, J., H. Borba, A. Giampietro-Ganeco, L. Vieira, M. Boiago, P. Souza, and M. Martins.
2011. Incidence of contact pododermatitis in broilers subjected to heat stress. Accessed. Aug.
2014. http://en.engormix.com/MA-poultry-industry/management/articles/incidence-contact-
pododermatitis-broilers-t1806/124-p0.htm.
NRC. 1994. Nutrient Requirements of Poultry. 9th rev ed. Natl. Acad. Press, Washington, DC.
Petronini, P. G., E. De Angelis, P. Borghetti, A. Borghetti, and K. P. Wheeler. 1992. Modulation
by betaine of cellular responses to osmotic stress. Biochem. J 282:69-73.
Quinteiro-Filho, W., A. Ribeiro, V. Ferraz-de-Paula, M. Pinheiro, M. Sakai, L. Sá, A. Ferreira,
and J. Palermo-Neto. 2010. Heat stress impairs performance parameters, induces intestinal
injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 89:1905-1914.
Page 70
57
Rafeeq, M., T. Pasha, N. Rashid, B. Hilal, and I. Shahzad. 2011. Effect of supplementation of
methionine, betaine and choline on the performance of broiler chicken in early life fed
methionine deficient ration. . J. Anim. Plant. Sci. 21:778-780.
Smith, M. O., and R. G. Teeter. 1987. Potassium Balance of the 5 to 8-Week-Old Broiler
Exposed to Constant Heat or Cycling High Temperature Stress and the Effects of
Supplemental Potassium Chloride on Body Weight Gain and Feed Efficiency. Poult. Sci.
66:487-492. doi 10.3382/ps.0660487
Soutyrine, A., M. Smith, and B. Sivanadian. 1998. Feed withdrawal, potassium chloride, and
carbonated water effects on broiler thermotolerance. J. Appl. Poult. Res. 7:138-143.
St-Pierre, N., B. Cobanov, and G. Schnitkey. 2003. Economic losses from heat stress by US
livestock industries. J. Dairy Sci. 86:E52-E77.
Teeter, R., and M. Smith. 1986. High chronic ambient temperature stress effects on broiler acid-
base balance and their response to supplemental ammonium chloride, potassium chloride, and
potassium carbonate. Poult. Sci. 65:1777-1781.
Waldroup, P., and C. Fritts. 2005. Evaluation of separate and combined effects of choline and
betaine in diets for male broilers. Int. J. Poult. Sci 4:442-448.
Waldroup, P. W., M. A. Motl, F. Yan, and C. A. Fritts. 2006. Effects of betaine and choline on
response to methionine supplementation to broiler diets formulated to industry standards. J.
Appl. Poult. Res. 15:58-71.
Yalcin, S., S. Özkan, L. Türkmut, and P. Siegel. 2001. Responses to heat stress in commercial
and local broiler stocks. 1. Performance traits. Br. Poult. Sci. 42:149-152.
Page 71
58
Chapter IV: Effect of dietary choline and betaine on corticosterone levels and immune
organ weights of heat-stressed broilers
Page 72
59
Abstract
A study was conducted to determine the effects of supplemental choline (CHO) and betaine
(BET) on broiler corticosterone concentration and immune organ weights under different
temperature conditions, thermoneutral (TN: 23.9oC) and heat stress (HS: 28-36
oC). Eight
hundred day-old mixed sex Cobb500 (female) x Hubbard (M99 male) chicks were weighed,
wing tagged and equally divided into the two environments. Eight replicates of 10 chicks were
randomly assigned to each of five dietary treatments: treatment 1 (CON) the basal diet; treatment
2 (CHO500) basal diet plus 500 ppm methyl equivalents added CHO; treatment 3 (CHO1000)
basal diet plus 1000 ppm methyl equivalents added CHO; treatment 4 (BET500) basal diet plus
500 ppm methyl equivalents added BET and treatment 5 (BET1000) basal diet plus 1000 ppm
methyl equivalents added BET. The corn-soy bean basal diet was formulated to meet NRC
requirements for broilers. The birds were brooded under standard conditions for three weeks
prior to heat stress imposition. Plasma corticosterone was determined before and three weeks
after imposition of heat stress. At the termination of the study, birds were processed and immune
organs (spleen, bursa of Fabricius and thymus) collected and weighed. Data were analyzed using
mixed model analysis of variance (SAS 9.3, Cary, NC). On day 22, there was no effect of diet or
temperature and no diet X temperature interaction on corticosterone levels. Day 45
corticosterone levels (HS, 3.97 ng/ml and TN, 2.16 ng/ml) were affected by temperature (P =
0.002) while there was no diet effect or diet X temperature effect. There was day X temperature
interaction (P = 0.03) for corticosterone levels. This was caused by corticosterone levels
decreasing less by 0.6 (4.57 ng/ml to 3.97 ng/ml) in HS birds while decreasing more by 1.45
(3.61 ng/ml to 2.16 ng/ml) in TN birds from days 22 to 45. There was no diet effect or diet X
temperature interaction on immune organ weights. However, chronic heat stress significantly (P
Page 73
60
= 0.03) reduced both thymus and bursa weights. Thymus (r = - 0.33, P = 0.002) and bursa (r = -
0.36, P = 0.0008) are negatively and moderately related to corticosterone levels. Betaine and
choline supplementation at 500 ppm and 1000 ppm did not protect broilers from heat stress and
did not affect immune organ weights.
Introduction
Exposure of chickens to high environmental temperature causes them to become heat
stressed. Birds respond to being stressed by decreasing feed intake which results in decreased
weight gain and feed efficiency (Cooper and Washburn, 1998; Sakomura, et al., 2013). Other
physiological changes also occur such as an increase in plasma corticosterone levels (Edens and
Siegel, 1976; Quinteiro-Filho, et al., 2012b) . Corticosterone is the main stress hormone in
chickens. The increase in corticosterone levels is the result of the activation of hypothalamus-
pituitary-adrenal (HPA) axis in avian species (Beuving and Vonder, 1978; Etches, 1976;
Quinteiro-Filho, et al., 2010) and is a response of the body to a stressor (Selye, 1976). The
stressor is detected by the cortex in the brain which activates HPA by sending a signal to the
hypothalamus. Corticotropin releasing hormone (CRH) from the hypothalamus causes the
pituitary gland to release adrenocorticotropic hormone (ACTH) which stimulates the synthesis of
glucocorticoids by the adrenal cortex (Chrousos, 1995). Glucocorticoids release is also
stimulated in response to inflammation. An increase in proinflammatory cytokines during
excessive inflammation also activates the HPA resulting in increased production of
glucocorticoids which reduces the inflammation (Chrousos, 1995).
Corticosterone serves as an indicator of stress in chickens. During acute heat stress,
plasma corticosterone increased after 70 minutes exposure but decreased rapidly to base line in
Page 74
61
birds heated for 120 min (Edens and Siegel, 1975). Several studies, in which either
corticosterone was fed or ACTH injected to simulate chicken response to high temperature, have
shown that an increase in corticosterone for a short period of time may be beneficial. In a study
conducted by Nasir, et al. (1999), corticosterone was injected to adrenalectomised chickens to
mimic an increase of corticosterone during stress. They found that elevated plasma
corticosterone stimulated higher feed intake, delayed food passage time and increased calcium,
phosphorous and glucose absorption. Similarly Garriga, et al. (2006) found an increase in apical
glucose transport during heat stress. Enhanced gluconeogenesis following short term
administration of corticosterone in chickens was reported by Lin, et al. (2004b) and Shini, et al.
(2008b).
Immune organs (bursa of Fabricius, spleen and thymus) are important components of the
immune system (McArthur, et al., 1971). Thymus and bursa are central lymphoid tissues; thymus
produces small lymphocytes and is responsible for the cell mediated immunity while bursa
produces large lymphocytes and is involved in humoral immunity (Kendall, 1980). Spleen, a
peripheral lymphoid tissue, is considered a final stage of lymphocyte differentiation for the
production of memory cells involved in secondary immune response (John, 1994). According to
(Chrousos, 1995), the production of glucocorticoids at a suboptimal level has detrimental effects
on the immune system. Shini, et al. (2008a) showed that increased levels of corticosterone
decreased spleen and bursa weights. Similarly, Post, et al. (2003) reported a reduction in weight
gain and spleen weight in chickens treated with high corticosterone to simulate heat stress
conditions. Quinteiro-Filho, et al. (2012a) showed that acute heat stress increased plasma
corticosterone but found no negative impact on immune organ (spleen, thymus and bursa)
weights. Few studies have evaluated the effect of chronic heat stress on corticosterone levels in
Page 75
62
broiler chickens and its effect on lymphoid organs. Quinteiro-Filho, et al. (2010) reported an
increase in corticosterone levels in chronic heat-stressed birds and a decrease in lymphoid organ
weights. Therefore the objective of the present study was to ascertain the effect of chronic heat
stress on chicken plasma corticosterone levels and immune organ weights and to determine if
dietary supplementation of choline and betaine could decrease the negative impacts of heat
stress.
Materials and Methods
Birds and Housing
The procedures were reviewed and approved by the University of Tennessee Institutional
Animal Care and Use Committee (IACUC), and the research was conducted at the Johnson
Animal Research and Teaching Unit (JARTU). Two adjacent rooms, thermoneutral (TN) and
heat stress (HS) were used. Each room was divided into 40 pens equipped with individual gravity
feeders, water nipple line and bedding made of shredded paper. The rooms were equipped with
HOBO®3
data loggers to record daily temperature and humidity. In addition, a humidifier was
placed in the heat stress room to ensure that humidity remained above 50%. Eight hundred day-
old mixed sex Cobb500 (female) x Hubbard (M99 male) chicks were obtained from Pilgrim’s
Pride hatchery in Cohutta, Georgia. They were vaccinated at the hatchery against Marek’s and
Gumboro diseases (VAXXITEK HVT-IBD); Escherichia coli, Salmonella typhimuriun and
Pseudomonas aeruginosa (GARASOL); Coccidiosis (COCCIVAC-B), Newcastle
3 Onset Computer Corporation
470 MacArthur Blvd. Bourne, MA 02532
Page 76
63
(NEWHATCH C2-M) and Infectious bronchitis (MILDVAC ARK and GA 2008). Upon arrival,
the birds were weighed by groups of ten and assigned to one of each treatment.
Temperature Treatments
During the first 21 days, birds were brooded under similar conditions in the two rooms.
The temperature was set to 32.22oC the first week and decreased by 2.78
oC until it reached
23.9oC by the end of the third week. From day 22 to the end of the study, the temperature was
maintained at 23.9 oC in the TN room and cycled from 28
oC to 36
oC between 10:00 a.m. and
8:00 p.m. in the HS room (Figure 3). The photoperiod regimen was 23 hours of light followed by
1 hour of dark in both environments. The humidity was maintained at 50% in the HS room.
Figure 3. The daily temperature in TN (23.9 ±1°C) and HS (28-36±1°C) rooms.
10
15
20
25
30
35
40
Tem
p. °C
Time
HS
TN
Page 77
64
Dietary Treatments
Five dietary treatments were tested. The basal formulation with choline or betaine
supplementation was used to make the following dietary treatments:
Control (CON), basal formulation
Low choline (CHO500), basal diet plus 500 ppm methyl equivalents added choline
High choline (CHO1000), basal diet plus 1000 ppm methyl equivalents added choline
Low betaine (BET500), basal diet plus 500 ppm methyl equivalents added betaine
High betaine (BET1000), basal diet plus 1000 ppm methyl equivalents added betaine
The basal diet was formulated to meet broilers energy, protein, minerals and vitamins
requirements (NRC., 1994) except for methionine which was 20% lower. The birds were fed
starter diet (Table 12) from days 1-21, grower diet from (Table 13) days 22-35 and finisher diet
(Table 14) from days 36-49.
Blood Collection
On day 49, one bird was randomly selected from each pen in each room and blood was
collected from their brachial vein into heparinized tubes and stored on ice. Blood was
centrifuged at 2000 rpm for 10 min, and plasma stored in micro-centrifuge tubes at -80oC for
later corticosterone analysis.
Processing of Birds
Prior to harvesting on day 52, birds were deprived of feed for 12 hours to allow for the
emptying of the digestive track. Eighty birds (eight birds randomly selected from each treatment
per room), were weighed, hung on shackles, stunned with an electric knife, and then killed by
severing the jugular vein. They were allowed to bleed for 3 minutes then scalded in hot water
(59.44oC) for 40 second, and their feathers removed using a de-feathering machine. Birds were
Page 78
65
Table 12. Composition of starter diet (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 62.86 62.86 62.86 62.86 62.86
Soybean meal 31.00 31.00 31.00 31.00 31.00
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.20 0.105 0.01 0.143 0.0853
Choline, mg/kg2 0.00 0.095 0.19 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.15 0.15 0.15 0.15 0.15
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 2.10 2.10 2.10 2.10 2.10
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 1.90 1.90 1.90 1.90 1.90
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3024.00 3024.00 3024.00 3024.00 3024.00
Crude protein, % 20.59 20.59 20.59 20.59 20.59
Calcium, % 0.98 0.98 0.98 0.98 0.98
Available Phosphorus, % 0.51 0.51 0.51 0.51 0.51
Potassium, % 0.81 0.81 0.81 0.81 0.81
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 92.99 92.99 92.99 92.99 92.99
Choline, mg/kg 1236.38 2186.44 3136.44 1236.38 1236.38
Betaine, mg/kg 0.00 0.00 0.00 585.01 1147.03
Lysine, % 1.18 1.18 1.18 1.18 1.18
Methionine, % 0.47 0.47 0.47 0.47 0.47
Methionine +Cysteine, % 0.81 0.81 0.81 0.81 0.81
Threonine, % 0.76 0.76 0.76 0.76 0.76
Fat, % 4.60 4.60 4.60 4.60 4.69
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 79
66
Table 13. Composition of grower diet (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 62.32 62.32 62.32 62.32 62.32
Soybean meal 31.00 31.00 31.00 31.00 31.00
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.30 0.1884 0.0768 0.243 0.1853
Choline, mg/kg2 0.00 0.1116 0.2232 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.20 0.20 0.20 0.20 0.20
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 1.69 1.69 1.69 1.69 1.69
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 2.70 2.70 2.70 2.70 2.70
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3072.00 3074.00 3072.00 3072.00 3072.00
Crude protein, % 19.95 19.95 19.95 19.95 19.95
Calcium, % 0.89 0.89 0.89 0.89 0.89
Available Phosphorus, % 0.43 0.43 0.43 0.43 0.43
Potassium, % 0.80 0.80 0.80 0.80 0.80
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 92.48 92.49 92.48 92.48 92.48
Choline, mg/kg 1233.03 2349.37 3465.10 1233.02 1233.03
Betaine, mg/kg 0.00 0.00 0.00 585.01 1147.03
Lysine, % 1.18 1.18 1.18 1.18 1.18
Methionine, % 0.52 0.52 0.52 0.52 0.52
Methionine +Cysteine, % 0.85 0.85 0.85 0.85 0.85
Threonine, % 0.76 0.76 0.76 0.76 0.76
Fat, % 5.38 5.38 5.38 5.38 5.38
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 80
67
Table 14. Composition of finisher diet (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 66.00 66.00 66.00 66.00 66.00
Soybean meal 27.37 27.37 27.37 27.37 27.37
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.30 0.1884 0.0768 0.243 0.1853
Choline, mg/kg2 0.00 0.1116 0.2232 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.15 0.15 0.15 0.15 0.15
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 1.69 1.69 1.69 1.69 1.69
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 2.70 2.70 2.70 2.70 2.70
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3105.00 3107.00 3105.00 3108.00 3105.00
Crude protein, % 18.55 18.55 18.55 18.55 18.55
Calcium, % 0.88 0.88 0.88 0.88 0.88
Available Phosphorus, % 0.43 0.43 0.43 0.43 0.43
Potassium, % 0.74 0.74 0.74 0.74 0.74
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 91.15 91.16 91.15 91.16 91.15
Choline, mg/kg 1156.71 2273.05 3388.78 1157.25 1156.71
Betaine, mg/kg 0.00 0.00 0.00 569.98 1147.03
Lysine, % 1.08 1.08 1.08 1.08 1.08
Methionine, % 0.45 0.45 0.45 0.45 0.45
Methionine +Cysteine, % 0.77 0.77 0.77 0.77 0.77
Threonine, % 0.70 0.70 0.70 0.70 0.70
Fat, % 5.48 5.48 5.48 5.49 5.48
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 81
68
then eviscerated and the carcasses weighed. Immune organs (bursa of Fabricius, spleen and
thymus) were carefully dissected and weighed. The weight of each organ was calculated as a
percent of total body weight.
Corticosterone Radioimmunoassay
Radioimmunoassay was performed to determine corticosterone concentration in plasma.
Total plasma corticosterone was analyzed in duplicate using ImmuChemTM
Double Antibody
Corticosterone 125
RIA Kit4. The procedures described in the radioimmunoassay kit were
followed. Frozen plasma samples were thawed at room temperature, centrifuged to precipitate
the fat content and then vortexed for 5 seconds to remix the supernatant. Plasma was diluted 1:4
in order to line up with the standard curve for more accurate count. Precipitates were counted
using the Perkin Elmer 1470 Automated Gamma Counter5. The intra and inter coefficient of
variation (CVs) were 15.45 and 18.64% respectively for low (1.93 ng/ml) and 16.85 and 19.62%
respectively for high (10.61 ng/ml) corticosterone standards.
Statistical Analysis
The experimental design was a completely randomized design (CRD) split plot with
temperature in the whole plots (rooms) and diet in the subplots (pens). Data were analyzed using
mixed model analysis of variance procedure SAS 9.3 (SAS Institute, Cary, NC, USA) and least
squares means compared using protected LSD at 5% level of significance. The analysis of the
change in corticosterone levels over time was performed using repeated measures. Correlation
was used to study the relation between corticosterone and lymphoid organ weights.
4 MP Biomedicals, LLC
Diagnostics Division, 13 Mountain View Avenue Orangeburg, NY 10962 5 Perkin Elmer, Wallac Oy, P.O. Box 10
Turku, Finland
Page 82
69
Results
Bursa of Fabricius, Spleen and Thymus Weights
Bursa of Fabricius, spleen and thymus weights expressed as a percent of body weights
are presented in Table 15. There was no diet X temperature interaction (P > 0.10) for either of
the lymphoid organ weights. Spleen weight was not affected by treatments (P > 0.30). Thymus
weight was not affected by diet (P = 0.34), however it was decreased (P = 0.03) by heat stress
(0.22% HS vs 0.27% TN). Bursa of Fabricius weight was decreased (P = 0.03) by heat stress
(0.16% HS vs 0.13% TN). Diet tended (P = 0.06) to increase bursa weight with birds consuming
BET500 having higher weights (0.18%) compared to the rest of the diets.
Plasma Corticosterone
Data on plasma corticosterone concentrations have been log transformed, back
transformed means and standard errors are reported (Table 16 and Table 17). On day 22, prior to
the imposition of elevated temperature, corticosterone concentrations determined as base line
were statistically similar for both HS and TN birds. On day 45, there was no diet (P = 0.97) or
diet X temperature interaction (P = 0.16) on plasma corticosterone concentration. Corticosterone
level in HS (3.97 ng/ml) birds was significantly (P = 0.002) higher compared to TN (2.26 ng/ml)
birds (Table 16). There was temp X day interaction (P = 0.03) for corticosterone (Table 17). A
Pearson correlation among thymus, bursa, spleen weights and corticosterone day 45, showed a
weak and negative correlation between corticosterone and spleen (r = - 0.03, P = 0.78) whereas
thymus (r = - 0.33, P = 0.002) and bursa (r = -0.36, P = 0.0008) were negatively and moderately
related to corticosterone.
Page 83
70
Table 15. Effect of choline and betaine supplementation on lymphoid organ weights (% body
weight) 1
of heat-stressed broilers
Diet Temp.2
Bursa Thymus Spleen
Cntrl TN 0.14 0.32 0.07
HS 0.15 0.21 0.09
Cho500 TN 0.16 0.27 0.08
HS 0.12 0.27 0.08
Cho1000 TN 0.14 0.21 0.07
HS 0.12 0.24 0.07
Bet500 TN 0.19 0.29 0.08
HS 0.16 0.21 0.08
Bet1000 TN 0.16 0.26 0.07
HS 0.11 0.16 0.06
SEM 0.02 0.03 0.01
Effect P values
Diet 0.06 0.34 0.31
Temp 0.03 0.03 0.99
Diet x Temp 0.39 0.10 0.39
Main Effect Means
Diet
Cntrl 0.14 0.26 0.08
Cho500 0.14 0.27 0.08
Cho1000 0.13 0.22 0.07
Bet500 0.18 0.25 0.08
Bet1000 0.14 0.21 0.07
SEM 0.01 0.02 0.01
Temp
TN 0.16a
0.27a
0.08
HS 0.13b
0.22b
0.08
SEM 0.01 0.02 0.004 1 Data are least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Page 84
71
Table 16. Effect of choline and betaine supplementation on Corticosterone Concentration
(ng/ml) 1
of heat-stressed broilers on days 22 and 45
Diet Temp.2
Day 22 Day 45
Cntrl TN 3.63 2.97
HS 4.34 3.39
Cho500 TN 3.88 2.05
HS 3.10 4.99
Cho1000 TN 2.68 2.02
HS 6.68 4.00
Bet500 TN 4.41 2.19
HS 5.48 4.06
Bet1000 TN 3.69 1.92
HS 4.06 3.91
SEM 0.84 0.62
Effect P values
Diet 0.49 0.97
Temp 0.12 0.002
Diet x Temp 0.07 0.16
Main Effect Means
Diet
Cntrl 3.97 3.04
Cho500 3.47 3.05
Cho1000 4.23 2.85
Bet500 4.91 2.98
Bet1000 3.87 2.74
SEM
Temp
0.58 0.41
TN 3.61 2.16b
HS 4.57 3.97a
SEM 0.41 0.34 1 Data are back transformed least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Page 85
72
Table 17. Temp X Day interaction on corticosterone concentration (ng/ml) 1
of heat-stressed
broilers
Temp.2
Days Corticosterone
HS 22 4.57 a
HS 45 3.97 a
TN 22 3.61 a
TN
Pooled SEM
Effect
Temp x Day
45 2.16 b
0.35
P value
0.03 1 Data are least squares means.
2 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
Discussion
The results of his study suggest that during chronic HS corticosterone levels increased
and remained elevated compared to thermoneutral environment. This is consistent with the
results of a study by Quinteiro-Filho, et al. (2010). These researchers reported that broilers
subjected to 31 and 36oC for seven days had higher corticosterone levels compared to control
birds raised at 21oC. Similarly, Geraert, et al. (1996) reported that plasma corticosterone
increased and remained elevated in chickens exposed to 32oC for two weeks. Studies have also
shown positive effects related to increased corticosterone levels. An increase in plasma
corticosterone during acute HS enhances gluconeogenesis (Lin, et al., 2004a). Increased plasma
corticosterone following corticosterone feeding to mimic stress conditions also elicited an
increase in glucose absorption from the intestine (Nasir, et al., 1999). Generally, and as seen in
this study, prolonged high temperature or chronic HS causes plasma corticosterone to remain
elevated longer than normal and this could be detrimental to the immune system. In the absence
Page 86
73
of infection and external stressors other than high temperature, it appears that corticosterone
levels naturally decreases with age, in birds reared under TN environment. In the present study,
birds received a prophylactic dose of an anti-coccidia agent (Coban) in their diet, and no
infection was seen during the entire experiment. Hence, the sustained elevation of corticosterone
was due to HS.
Based on the result of the present study, there is a moderate and negative correlation
between corticosterone and lymphoid organ (thymus and bursa) weights. Heat stress decreased
thymus and bursa of Fabricius weights while spleen weight was unchanged compared to the TN
environment. This could be due to the increase in corticosterone level. These results are
consistent with those reported by Bartlett and Smith (2003) and Quinteiro-Filho, et al. (2010)
who found a decrease in thymus and bursa weights. However, in their studies spleen weight was
decreased by high temperature as well. The bursa of Fabricius and thymus are two gut associated
lymphoid tissues (John, 1994) involved in the adaptive immunity of chickens. Although this
study did not specifically determine and evaluate adverse effects of decreased lymphoid organ
weights on the immune functions, a reduction in their capacity to produce enough immune cells
for the adaptive immunity can be predicted.
The results of the study suggest that chronic HS causes an increase in corticosterone and
reduces lymphoid organ weights, the combination of which may impair the ability of heat-
stressed chickens to mount an appropriate immune response. Dietary supplementation of CHO
and BET did not prevent the increase of plasma corticosterone and the reduction of immune
organ weights in heat-stressed broilers.
Page 87
74
References
Bartlett, J. R., and M. O. Smith. 2003. Effects of different levels of zinc on the performance and
immunocompetence of broilers under heat stress. Poult. Sci. 82:1580-1588.
Beuving, G., and G. Vonder. 1978. Effect of stressing factors on corticosterone levels in the
plasma of laying hens. Gen. Comp. Endocrinol. 35:153-159.
Chrousos, G. P. 1995. The Hypothalamic–Pituitary–Adrenal Axis and Immune-Mediated
Inflammation. N. Engl. J. Med. 332:1351-1363. doi doi:10.1056/NEJM199505183322008
Cooper, M. A., and K. W. Washburn. 1998. The relationships of body temperature to weight
gain, feed consumption, and feed utilization in broilers under heat stress. Poult. Sci. 77:237-
242. doi 10.1093/ps/77.2.237
Edens, F. W., and H. S. Siegel. 1976. Modification of Corticosterone and Glucose Responses by
Sympatholytic Agents in Young Chickens During Acute Heat Exposure. Poult. Sci. 55:1704-
1712. doi 10.3382/ps.0551704
Edens, F. W., and H. t. Siegel. 1975. Adrenal responses in high and low ACTH response lines of
chickens during acute heat stress. Gen. Comp. Endocrinol. 25:64-73.
Etches, R. 1976. A radioimmunoassay for corticosterone and its application to the measurement
of stress in poultry. Steroids 28:763-773.
Garriga, C., R. R. Hunter, C. Amat, J. M. Planas, M. A. Mitchell, and M. Moretó. 2006. Heat
stress increases apical glucose transport in the chicken jejunum. Am. J. Physiol. Regul. Intgr.
Comp. Physiol. 290:R195-R201.
Geraert, P. A., J. C. F. Padilha, and S. Guillaumin. 1996. Metabolic and endocrine changes
induced by chronic heat exposure in broiler chickens: Biological and endocrinological
variables. Br. J. Nutr. 75:205-216. doi 10.1079/bjn19960125
John, J. L. 1994. The avian spleen: a neglected organ. Q. Rev. Biol.:327-351.
Kendall, M. D. 1980. Avian thymus glands: a review. Dev. Comp. Immunol. 4:191-209.
Lin, H., E. Decuypere, and J. Buyse. 2004a. Oxidative stress induced by corticosterone
administration in broiler chickens (Gallus gallus domesticus): 1. Chronic exposure. Comp.
Biochem. Physiol. B Mol. Integr. Physiol. 139:737-744.
Lin, H., E. Decuypere, and J. Buyse. 2004b. Oxidative stress induced by corticosterone
administration in broiler chickens (Gallus gallus domesticus): 2. Short-term effect. Comp.
Biochem. Physiol. B Mol. Integr. Physiol. 139:745-751.
McArthur, W., J. Chapman, and G. Thorbecke. 1971. Immunocompetent cells of the chicken I.
Specific surface antigenic markers on bursa and thymus cells. J. Exp. Med. 134:1036-1045.
Page 88
75
Nasir, A., R. Moudgal, and N. Singh. 1999. Involvement of corticosterone in food intake, food
passage time and in vivo uptake of nutrients in the chicken (Gallus domesticus). Br. Poult.
Sci. 40:517-522.
NRC. 1994. Nutrient Requirements of Poultry. 9th rev ed. Natl. Acad. Press, Washington, DC.
Post, J., J. Rebel, and A. Ter Huurne. 2003. Physiological effects of elevated plasma
corticosterone concentrations in broiler chickens. An alternative means by which to assess the
physiological effects of stress. Poult. Sci. 82:1313-1318.
Quinteiro-Filho, W., A. Ribeiro, V. Ferraz-de-Paula, M. Pinheiro, M. Sakai, L. Sá, A. Ferreira,
and J. Palermo-Neto. 2010. Heat stress impairs performance parameters, induces intestinal
injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 89:1905-1914.
Quinteiro-Filho, W., M. Rodrigues, A. Ribeiro, V. Ferraz-de-Paula, M. Pinheiro, L. Sa, A.
Ferreira, and J. Palermo-Neto. 2012a. Acute heat stress impairs performance parameters and
induces mild intestinal enteritis in broiler chickens: Role of acute hypothalamic-pituitary-
adrenal axis activation. J. Anim. Sci. 90:1986-1994.
Quinteiro-Filho, W. M., M. V. Rodrigues, A. Ribeiro, V. Ferraz-de-Paula, M. L. Pinheiro, L. R.
Sa, A. J. Ferreira, and J. Palermo-Neto. 2012b. Acute heat stress impairs performance
parameters and induces mild intestinal enteritis in broiler chickens: role of acute
hypothalamic-pituitary-adrenal axis activation. J. Anim. Sci. 90:1986-1994. doi
10.2527/jas.2011-3949
Sakomura, N. K., N. A. A. Barbosa, F. A. Longo, E. P. da Silva, M. A. Bonato, and J. B. K.
Fernandes. 2013. Effect of dietary betaine supplementation on the performance, carcass yield,
and intestinal morphometrics of broilers submitted to heat stress. Braz. J. Poultry Sci. 15:105-
111.
Selye, H. 1976. Forty years of stress research: principal remaining problems and misconceptions.
Can. Med. Assoc. J. 115:53.
Shini, S., P. Kaiser, A. Shini, and W. L. Bryden. 2008a. Biological response of chickens (Gallus
gallus domesticus) induced by corticosterone and a bacterial endotoxin. Comp. Biochem.
Physiol. B Mol. Integr. Physiol. 149:324-333.
Shini, S., P. Kaiser, A. Shini, and W. L. Bryden. 2008b. Differential alterations in ultrastructural
morphology of chicken heterophils and lymphocytes induced by corticosterone and
lipopolysaccharide. Vet. Immunol. Immunopathol. 122:83-93.
Page 89
76
Chapter V: Effect of dietary choline and betaine on choline metabolites and intestinal
morphology of heat-stressed broilers
Page 90
77
Abstract
The objective of this study was to evaluate the effects of supplemental choline (CHO) and
betaine (BET) on production of choline metabolites and on the intestinal morphology of broilers
under different temperature conditions, thermoneutral (TN: 23.9oC) and heat stress (HS: 28-
36oC). Eight hundred day-old mixed sex Cobb500 (female) x Hubbard (M99 male) chicks were
weighed, wing tagged and equally divided and placed into each of the two environments. Eight
replicates of ten chicks were randomly assigned to each of five dietary treatments in each
environment: treatment 1 (CON) the basal diet; treatment 2 (CHO500) basal diet plus 500 ppm
methyl equivalents added CHO; treatment 3 (CHO1000) basal diet plus 1000 ppm methyl
equivalents added CHO; treatment 4 (BET500) basal diet plus 500 ppm methyl equivalents
added BET and treatment 5 (BET1000) basal diet plus 1000 ppm methyl equivalents added BET.
The corn-soy bean basal diet was formulated to meet NRC requirements for broilers. On day 45,
blood was collected from eight birds per treatment while liver and intestinal samples were
collected on day 52. Samples were analyzed for CHO and CHO metabolites. Intestinal histology
was evaluated. Data were analyzed using mixed model analysis of variance (SAS 9.3, Cary, NC).
Results showed that neither diet nor temperature had any effect on villi height, crypt depth or
villi height:crypt depth ratio nor was there any diet X temperature interaction. Intestinal betaine
was affected by diet (P = 0.01) with CHO1000 (6.36 mg/100g) and BET1000 (6.40 mg/100g)
higher than the other treatments. The diet X temperature interaction for intestinal choline tended
(P = 0.06) to be significant while glycerophosphocholine (GPC), total lysophosphatidylcholine
(TLPC), total phosphatidylcholine (TPtCho), phosphocholine (PCho) and sphingomyelin (Sm)
were unaffected. Plasma BET was highest (P = 0.0001) for CHO1000 (233.59 umol/l) and
BET1000 (219.75 umol/l) while plasma CHO and PCho were unaffected.
Page 91
78
Glycerophosphocholine was highest for CON (18.60 umol/l). However, it was not statistically
different from CHO500 (11.53 umol/l). Plasma TPtCho was increased (P = 0.02) by temperature
in HS birds (11744.00 umol/l) compared to TN birds (5908.08 umol/l). There was a diet X
temperature interaction for plasma TLPC (P = 0.04) resulting from the CON TLPC decreasing
by 49.06 (82.83 TN to 33.77 umol/l HS) while BET1000 TLPC only decreased by 4.55 (47.9 TN
to 43.35 umol/l HS). Liver acetylcholine (Acho), TLPC, TPtCho and Sm contents were not
affected by diet, temperature or their interaction (P > 0.05). There was a diet X temperature
interaction (P = 0.03) for liver betaine content. Under HS, BET1000 (17.90 mg/100g) was
significantly higher than the other diets while in the TN environment, CHO1000 was the highest
(14.49 mg/100g). Liver CHO and GPC contents were affected by temperature which increased
(P = 0.01) CHO by 27.95% (7.14 mg/100g TN vs 9.91 mg/100 HS) and GPC (P = 0.02) levels
by 23.02% (3.88 mg/100g TN vs 5.04 mg/100g HS). In this study, CHO metabolites were
affected by temperature, while intestinal morphology was not.
Introduction
Choline is found in the form of phosphatidylcholine or sphingomyelin in most foods
(Combs Jr, 1991; McDowell, 1989) with the highest concentrations in wheat germ, dried
soybean, egg yolk, pork, and glandular meat such as beef and chicken liver (Zeisel, et al., 2003).
Choline is metabolized into different compounds (betaine, glycerophosphocholine, acetylcholine,
phosphatidylcholine among others) throughout the body. Betaine donates a methyl group to
homocysteine to form methionine (Borsook and Dubnoff, 1947) or serves as an osmolyte;
glycerophosphocholine also serves as an osmolyte (Nakanishi and Burg, 1989); acetylcholine is
involved in neurotransmission (Combs Jr, 1991; McDowell, 1989) whereas phosphatidylcholine
Page 92
79
is essential for cell membrane integrity (Plagemann, 1968) and transmembrane signaling (Zeisel,
1993). Choline plays important metabolic, physiological and structural functions in the body
therefore meeting its requirement at every step in the development is essential. Additionally, up
to eight weeks of age (broilers are processed at 6-7 weeks) chickens are unable to synthesize
enough endogenous choline to meet their requirements (Cook, et al., 1984; Molitoris and Baker,
1976) making dietary choline very crucial. It is important to identify and reduce factors that may
affect birds’ health and their ability to efficiently absorb and utilize choline.
Several stressors including osmotic stress and high temperature affect gut integrity. These
conditions may decrease feed intake and feed digestibility thus reducing choline absorption and
its distribution into different pathways. In birds, the contents of the small intestine are always
hypertonic to plasma osmolality (320 mOsmol) (Klasing, et al., 2002; Mongin, et al., 1976).
During heat stress, water excretion increases (Belay and Teeter, 1993) causing water imbalance.
Combined, these conditions exert substantial osmotic pressure on intestinal cells resulting in
decreased cell volume and perturbation of cells’ activity including enzymatic processes and
protein synthesis (Petronini, et al., 1992). To restore the osmotic balance, inorganic and organic
osmolytes are used by cells. Inorganic osmolytes are used in initial response to osmotic
imbalance but their excessive accumulation in cell can be detrimental (Petronini, et al., 1992). In
bacteria, at low osmotic pressure, potassium (K+) is preferably used to restore the balance.
However, when the external osmolarity is increased, proportional accumulation of K+ is
deleterious for cell activities and betaine is used in preference to K+
to restore cell turgor
(Sutherland, et al., 1986). Similarly, Petronini, et al. (1992) demonstrated that under
hyperosmotic conditions, unlike organic osmolytes, excessive accumulation of inorganic
osmolytes (Na+, Cl
- and K
+) disrupt enzymatic activities and protein synthesis.
Page 93
80
Among organic electrolytes such as polyols, sugar, amino acids and methylamines,
betaine has been shown to be the most effective in restoring cell volume, enzyme activity and
protein synthesis (Petronini, et al., 1992). Kettunen, et al. (2001a) suggested that betaine
accumulates in the intestinal cells, and plays an important role in osmoregulation in the small
intestine by controlling water movement across the intestinal epithelium. Several studies have
also proven the importance of betaine on the intestine of infected chickens. Betaine has been
shown to improve water balance and decrease the osmolarity in the duodenum of coccidia
infected chickens (Klasing, et al., 2002), stabilize intestinal mucosal structure and protect against
coccidia infections (Kettunen, et al., 2001b). The osmolytic properties of betaine could be
expected to improve osmotic pressure and protect the intestinal structure in broiler chickens
subjected to heat stress.
Similarly in mammals, glycerophosphocholine, predominantly formed from
phosphatidylcholine in the kidney, protects renal medullary cells from hyper tonicity of the
interstitial fluid (Nakanishi and Burg, 1989). Both, betaine and glycerophosphocholine are
choline metabolites and their relative importance in providing osmoprotectant benefits in the
intestine of chickens in unknown. The impact of heat stress on choline and choline metabolites in
the body have not been investigated. In addition, given the different functions of choline in the
body, evaluating the impact of heat stress on its different metabolites may be fundamental in
understanding the importance of supplemental choline and betaine to improve performance of
heat-stressed broilers. Therefore the objectives of this study were to evaluate the effects of
supplemental choline and betaine on choline metabolites, intestinal morphology and the relative
importance of betaine and glycerophosphocholine osmoprotectant properties on the intestine of
broilers under different temperature conditions, thermoneutral and heat stress.
Page 94
81
Materials and Methods
Birds and Housing
The procedures were reviewed and approved by the University of Tennessee Institutional
Animal Care and Use Committee (IACUC), and the research was conducted at the Johnson
Animal Research and Teaching Unit (JARTU). Two adjacent rooms, thermoneutral (TN) and
heat stress (HS) were used. Each room was divided into 40 pens equipped with individual gravity
feeders, water nipple line and bedding made of shredded paper. The rooms were equipped with
HOBO®6
data loggers to record daily temperature and humidity. In addition, a humidifier was
placed in the heat stress room to ensure that humidity remained above 50%. Eight hundred day-
old mixed sex Cobb500 (female) x Hubbard (M99 male) were obtained from Pilgrim’s Pride
hatchery in Cohutta, Georgia. They were vaccinated at the hatchery against Marek’s and
Gumboro diseases (VAXXITEK HVT-IBD); Escherichia coli, Salmonella typhimuriun and
Pseudomonas aeruginosa (GARASOL); Coccidiosis (COCCIVAC-B), Newcastle
(NEWHATCH C2-M) and Infectious bronchitis (MILDVAC ARK and GA 2008). Upon arrival,
the birds were weighed by groups of ten and assigned to one of each treatment.
Temperature Treatments
During the first 21 days, birds were brooded under similar conditions in the two rooms.
The temperature was set to 32.22oC the first week and decreased by 2.78
oC until it reached
23.9oC by the end of the third week. From day 22 to the end of the study, the temperature was
maintained at 23.9 oC in the TN room and cycled from 28
oC to 36
oC between 10:00 a.m. and
6 Onset Computer Corporation
470 MacArthur Blvd. Bourne, MA 02532
Page 95
82
8:00 p.m. in the HS room (Figure 4). The photoperiod regimen was 23 hours of light followed by
1 hour of dark in both environments. The humidity was maintained at 50% in the HS room.
Figure 4. Temperature treatment in TN (23.9 ±1°C) and HS (28-36±1°C) rooms.
10
15
20
25
30
35
40
Tem
p. °C
Time
HS
TN
Page 96
83
Dietary Treatments
Five dietary treatments were tested. The basal formulation with choline or betaine
supplementation was used to make the following dietary treatments:
Control (CON), basal formulation
Low choline (CHO500), basal diet plus 500 ppm methyl equivalents added choline
High choline (CHO1000), basal diet plus 1000 ppm methyl equivalents added choline
Low betaine (BET500), basal diet plus 500 ppm methyl equivalents added betaine
High betaine (BET1000), basal diet plus 1000 ppm methyl equivalents added betaine
The basal diet was formulated to meet broilers energy, protein, minerals and vitamins
requirements (NRC., 1994) except for methionine which was 20% lower. The birds were fed
starter diet (Table 18) from days 1-21, grower diet (Table 19) days 22-35 and finisher diet (Table
20) from days 36-49.
Blood Collection
On day 49, one bird was randomly selected from each pen in each room and blood was
collected from their brachial vein into heparinized tubes and stored on ice. Blood was
centrifuged at 2000 rpm for 10 min, and plasma stored in micro-centrifuge tubes at -80oC for
later choline metabolites analysis.
Processing of Birds
Prior to harvesting on day 52, birds were deprived of feed for 12 hours to allow the
emptying of the digestive track. Eighty birds (eight birds randomly selected from each treatment
per room), were weighed, hung on shackles, stunned with an electric knife, and then killed by
severing the jugular vein. They were allowed to bleed for 3 minutes then scalded in hot water
(59.44oC) for 40 seconds, and their feathers removed using a de-feathering machine.
Page 97
84
Table 18. Composition of experimental diet for starter period (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 62.86 62.86 62.86 62.86 62.86
Soybean meal 31.00 31.00 31.00 31.00 31.00
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.20 0.105 0.01 0.143 0.0853
Choline, mg/kg2 0.00 0.095 0.19 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.15 0.15 0.15 0.15 0.15
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 2.10 2.10 2.10 2.10 2.10
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 1.90 1.90 1.90 1.90 1.90
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3024.00 3024.00 3024.00 3024.00 3024.00
Crude protein, % 20.59 20.59 20.59 20.59 20.59
Calcium, % 0.98 0.98 0.98 0.98 0.98
Available Phosphorus, % 0.51 0.51 0.51 0.51 0.51
Potassium, % 0.81 0.81 0.81 0.81 0.81
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 92.99 92.99 92.99 92.99 92.99
Choline, mg/kg 1236.38 2186.44 3136.44 1236.38 1236.38
Betaine, mg/kg 0.00 0.00 0.00 585.01 1147.03
Lysine, % 1.18 1.18 1.18 1.18 1.18
Methionine, % 0.47 0.47 0.47 0.47 0.47
Methionine +Cysteine, % 0.81 0.81 0.81 0.81 0.81
Threonine, % 0.76 0.76 0.76 0.76 0.76
Fat, % 4.60 4.60 4.60 4.60 4.69
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 98
85
Table 19. Composition of experimental diet for grower period (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 62.32 62.32 62.32 62.32 62.32
Soybean meal 31.00 31.00 31.00 31.00 31.00
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.30 0.1884 0.0768 0.243 0.1853
Choline, mg/kg2 0.00 0.1116 0.2232 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.20 0.20 0.20 0.20 0.20
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 1.69 1.69 1.69 1.69 1.69
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 2.70 2.70 2.70 2.70 2.70
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3072.00 3074.00 3072.00 3072.00 3072.00
Crude protein, % 19.95 19.95 19.95 19.95 19.95
Calcium, % 0.89 0.89 0.89 0.89 0.89
Available Phosphorus, % 0.43 0.43 0.43 0.43 0.43
Potassium, % 0.80 0.80 0.80 0.80 0.80
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 92.48 92.49 92.48 92.48 92.48
Choline, mg/kg 1233.03 2349.37 3465.10 1233.02 1233.03
Betaine, mg/kg 0.00 0.00 0.00 585.01 1147.03
Lysine, % 1.18 1.18 1.18 1.18 1.18
Methionine, % 0.52 0.52 0.52 0.52 0.52
Methionine +Cysteine, % 0.85 0.85 0.85 0.85 0.85
Threonine, % 0.76 0.76 0.76 0.76 0.76
Fat, % 5.38 5.38 5.38 5.38 5.38
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 99
86
Table 20. Composition of experimental diet for finisher period (%)
Ingredients CON CHO500 CHO1000 BET500 BET1000
Corn, grain 66.00 66.00 66.00 66.00 66.00
Soybean meal 27.37 27.37 27.37 27.37 27.37
Vitamin premix1 0.0625 0.0625 0.0625 0.0625 0.0625
Filler, sand 0.30 0.1884 0.0768 0.243 0.1853
Choline, mg/kg2 0.00 0.1116 0.2232 0.00 0.00
Betaine, mg/kg2 0.00 0.00 0.00 0.057 0.1147
DL methionine 0.15 0.15 0.15 0.15 0.15
Salt 0.35 0.35 0.35 0.35 0.35
Limestone 1.10 1.10 1.10 1.10 1.10
Dical phosphate 1.69 1.69 1.69 1.69 1.69
Trace min. premix3 0.125 0.125 0.125 0.125 0.125
Fat, animal 2.70 2.70 2.70 2.70 2.70
Coban 0.0495 0.0495 0.0495 0.0495 0.0495
Lysine 0.10 0.10 0.10 0.10 0.10
Total 100.00 100.00 100.00 100.00 100.00
Nutrient Composition
ME, kcal/kg 3105.00 3107.00 3105.00 3108.00 3105.00
Crude protein, % 18.55 18.55 18.55 18.55 18.55
Calcium, % 0.88 0.88 0.88 0.88 0.88
Available Phosphorus, % 0.43 0.43 0.43 0.43 0.43
Potassium, % 0.74 0.74 0.74 0.74 0.74
Chloride, % 0.25 0.25 0.25 0.25 0.25
Sodium, % 0.16 0.16 0.16 0.16 0.16
Zinc, mg/kg 91.15 91.16 91.15 91.16 91.15
Choline, mg/kg 1156.71 2273.05 3388.78 1157.25 1156.71
Betaine, mg/kg 0.00 0.00 0.00 569.98 1147.03
Lysine, % 1.08 1.08 1.08 1.08 1.08
Methionine, % 0.45 0.45 0.45 0.45 0.45
Methionine +Cysteine, % 0.77 0.77 0.77 0.77 0.77
Threonine, % 0.70 0.70 0.70 0.70 0.70
Fat, % 5.48 5.48 5.48 5.49 5.48
Salt, % 0.53 0.53 0.53 0.53 0.53 1 Vitamin premix supplied per kg of feed: Vitamin A, 17636684 IU; Vitamin D3, 5952381 ICU; Vitamin
E, 35000 IU; Vitamin B12, 16 mg; Biotin, 176 mg; Menadione, 3858 mg; Thiamin, 3307 mg; Riboflavin,
15432 mg; Pantothenic acid, 24250 mg; Vitamin B6, 4409 mg; Niacin, 88180 mg; Folic acid, 1658 mg. 2
Choline chloride (60%), Betaine anhydrous, provided by Balchem Corporation and added at the expense
of the filler, sand.
3 Trace minerals premix supplied at 1.14 kg per ton of feed: Calcium, 3% min and 4% max; Manganese,
9.70%; Zinc, 7.50%; Copper, 3.53%; Iron, 4400 ppm; Iodine1586 ppm; Selenium, 240 ppm.
Page 100
87
Birds were then eviscerated and the carcasses weighed. Liver samples from both lobes were
collected, snap frozen with liquid nitrogen in cryogenic vials and stored at -80oC. The small
intestine was carefully separated, and two samples were collected: a 5 cm section was cut above
the Meckel’s diverticulum, cleaned in 10% saline buffer solution and snap frozen with liquid
nitrogen in cryogenic vials and stored at -80oC for later choline metabolites analysis, and another
5 cm section from the ileum was rinsed with 10% saline buffer and stored in formaldehyde at
room temperature for later histology.
Intestinal Histology
Histological sections from the intestinal samples were embedded in paraffin and stained
with hematoxylin and eosin. The tissue slides were observed under microscope EVOS® imaging
systems and photographs of three villi per slide and per bird showing complete villi and crypts
were taken (10x0.25 magnification). Villus length from the tip to the base of the villus and crypt
depth from the base of villus to the thin muscularis mucosae were measured in millimeter using
Imagej software.
Choline Metabolites Analysis by HILC-MS/MS
Plasma, liver and intestinal samples were extracted for choline and choline metabolites
analysis. The samples were kept on ice during the extraction process. First, 1 ml of extraction
solvent made of chloroform, methanol and water in a 1:2:0.8 ratio, was added to100 mg of
ground intestinal and liver tissues, 200 ul of plasma and 40 ul of standard solution. Samples were
centrifuged (16000rpm, 28620xg) at 4oC for 5min. The resulting supernatant was transferred into
separate glass vials. The extraction process was repeated twice for each sample and the
supernatant transferred into the same vial. The supernatant for each sample was dried under
constant nitrogen steam and the solid extract re-dissolved in 5 ml of methanol. The final
Page 101
88
solutions were analyzed for choline metabolites using the HILIC LC-MS/MS method adapted by
(Artegoitia, et al., 2014).
Statistical Analysis
The experimental design was a completely randomized design split plot with temperature
in the whole plots (rooms) and diet in the subplots (pens). Data were analyzed using mixed
model analysis of variance procedure SAS 9.3 (Cary, NC, USA) and least squares means
compared using protected LSD at 5% level of significance.
Results
Intestinal Histology
Intestinal villi height, crypts depth and villi height:crypt depth ratio are presented in
Table 21. There was no diet X temperature interaction for villi height, crypt depth and villi
height:crypt depth ratio (P > 0.2). Neither diet nor temperature affected crypt and villi:crypt
ratio, however, HS tended (P = 0.07) to decrease villi length (1.39 mm TN vs 1.26 mm HS).
Intestinal Choline and Choline Metabolites
Choline and CHO metabolites concentrations in the intestine are reported in Table 22.
Intestinal GPC, TLPC, TPtCho, PCho and Sm were not affected (P > 0.15) by temperature, diet
and diet X temperature interaction. There was no diet or diet X temperature interaction effect (P
> 0.1) on Acho however, temperature tended (P = 0.07) to increase it (0.83 in HS vs 0.62
mg/100g in TN).
There was no diet X temperature interaction for intestinal BET (P = 0.57), nor was there a
temperature effect (P = 0.34). However, intestinal BET content was affected by diet (P = 0.02).
Page 102
89
Table 21. Effect of choline and betaine supplementation on intestinal villi height, crypts depth
(mm) 1
and villi height:crypt depth of heat-stressed broilers
Diet Temp.2 Villi Crypt Villi:Crypt
CON TN 1.55 0.16 10.63
HS 1.28 0.13 9.97
CHO500 TN 1.34 0.14 10.34
HS 1.21 0.14 9.22
CHO1000 TN 1.34 0.14 9.74
HS 1.39 0.15 10.07
BET500 TN 1.35 0.16 8.88
HS 1.23 0.13 10.64
BET1000 TN 1.35 0.16 9.17
HS 1.21 0.14 9.35
Pooled SEM 0.30 0.02 0.81
Effect P values
Diet 0.25 0.96 0.85
Temp 0.07 0.10 0.87
Diet x Temp 0.22 0.58 0.56
Main Effect Means
Diet
CON 1.41 0.15 10.30
CHO500 1.28 0.14 9.78
CHO1000 1.36 0.15 9.91
BET500 1.29 0.14 9.76
BET1000 1.28 0.15 9.26
Pooled SEM 0.06 0.01 0.63
Temp
TN 1.39 0.15 0.38
HS 1.26 0.14 0.42
Pooled SEM 0.04 0.01 0.40 1
Data are least squares means. 2TN = Thermoneutral environment, HS = Heat stress environment.
Page 103
90
Table 22. Effect of choline and betaine supplementation on intestinal choline and choline
metabolites (mg/100g) 1
of heat-stressed broilers
Metabolites2
Diet Temp3
BET CHO GPC Acho4
TLPC4
TPtCho4
PCho Sm
CON TN 3.89 11.23 3.42 0.76 12.40 18473.75 24.94 144.47
HS 4.96 11.47 4.83 1.06 15.49 21863.73 25.00 161.79
CHO500 TN 5.24 9.51 4.27 0.52 13.52 12177.63 25.45 177.74
HS 4.27 9.94 4.06 0.85 14.37 14887.65 27.73 169.13
CHO1000 TN 7.02 9.16 3.97 0.49 9.56 12829.29 24.71 168.43
HS 5.71 10.38 4.03 0.69 9.72 12900.75 25.29 157.29
BET500 TN 5.02 12.19 4.23 0.51 10.44 13301.08 26.86 163.73
HS 4.59 9.30 4.27 0.79 8.43 10447.52 26.32 177.41
BET1000 TN 6.73 14.43 3.85 0.90 13.50 15600.19 27.38 179.34
HS 6.08 13.25 4.09 0.80 15.79 23656.70 28.42 176.55
Pooled SEM 0.75 1.09 0.55 0.15 3.24 4547.97 1.38 16.01
Effect P values
Diet 0.02 0.10 0.98 0.14 0.24 0.18 0.17 0.53
Temp 0.34 0.71 0.38 0.07 0.78 0.63 0.48 0.88
Diet x Temp 0.57 0.06 0.61 0.58 0.93 0.79 0.87 0.84
Main Effect Means
Diet
CON 4.43b
11.35 4.12 0.90 13.86 20097.39 24.97 153.13
CHO500 4.75b
9.72 4.17 0.66 13.94 13464.63 26.60 173.43
CHO1000 6.36a
9.77 4.00 0.58 9.64 12864.97 25.00 162.86
BET500 4.81b
10.74 4.25 0.64 9.38 11788.27 26.59 170.57
BET1000 6.40a
11.84 3.97 0.85 14.60 19210.65 27.90 177.95
Pooled SEM 0.53 0.77 0.39 0.11 2.28 3186.17 0.98 11.32
Temp
TN 5.58 10.50 3.95 0.62 11.77 14304.33 25.87 166.74
HS 5.12 10.87 4.26 0.83 12.36 15967.12 26.55 168.43
Pooled SEM 0.34 0.69 0.25 0.08 1.48 2422.05 0.66 7.98 1 Data are least squares means.
2 Metabolites: betaine (BET), free choline (CHO), glycerophosphocholine (GPC), acetylcholine (Acho),
total lysophosphatidylcholine (TLPC), total phosphatidylcholine (TPtCho), phosphocholine (PCho),
sphingomyelin (Sm). 3 TN = Thermoneutral environment, HS = Heat stress environment.
a,b Means within same column followed by different letters differ significantly (P < 0.05).
4 Data were log transformed and back transformed means reported.
Page 104
91
Both BET1000 and CHO1000 are statistically similar (6.36 and 6.40 mg/100g) but higher than
the CON (4.43 mg/100g), CHO500 (4.75 mg/100g) and BET500 (4.81 mg/100g).
Neither diet nor temperature affected intestinal CHO content (P > 0.10). However, their
interaction tended to have a significant (P = 0.06) impact. This was caused by BET1000
decreasing intestinal CHO content by 1.18 (14.43 mg/100g TN to 13.25 mg/100g HS) whereas
CHO500 increased it by 0.43 mg/100g (9.51 mg/100g TN to 9.94 mg/100g HS).
Liver Choline and Choline Metabolites
Liver choline and choline metabolites concentration are reported in Table 23. There was
no temperature, diet or diet X temperature interaction on liver Acho, TLPC, TPtCho and Sm
content (P > 0.05). However, there was diet X temperature interaction for betaine (P = 0.03).
This was caused by BET1000 increasing liver betaine content by 6.17 (11.74 mg/100g TN to
17.91 mg/100g HS) whereas both CHO1000 and CON decreased it by 1.27 (14.49 mg/100g to
13.22 mg/100g) and 0.68 (5.98 mg/100g TN to 5.30 mg/100g HS) respectively. Similarly, PCho
was affected by the interaction between diet and temperature (P = 0.04). This was due to CON
and BET1000 increasing liver PCho content by 8.39 (17.57 mg/100g TN to 25.96 mg/100g HS)
and 7.47 (15.69 mg/100g TN to 23.16 mg/100g HS) respectively whereas CHO500 and
CHO1000 decreased liver PCho by 0.58 (20.08 mg/100g TN to 19.50 mg/100g HS) and 1.32
(21.41 mg/100g to 20.09 mg/100g HS) respectively.
Liver CHO and GPC were not affected by diet and there was no diet X temperature
interaction (P > 0.05). However, the CHO content was significantly higher (P = 0.01) in HS birds
(9.91 mg/100) compared to TN birds (7.14 mg/100g). Similarly, GPC content for HS birds (5.04
mg/100g) was significantly higher (P = 0.02) than that of TN birds (3.88 mg/100g).
Page 105
92
Table 23. Effect of choline and betaine supplementation on liver choline and choline metabolites
(mg/100g) 1
of heat-stressed broilers
Metabolites2
Diet Temp3
BET CHO GPC Acho TLPC TPtCho PCho Sm4
CON TN 5.98de
7.99 4.78 0.70 8.17 18475.00 17.57bcd
140.88
HS 5.30e
8.81 5.94 0.90 8.96 26361.00 25.96a
143.46
CHO500 TN 6.72de
6.31 3.00 0.58 7.20 13523.00 20.08bcd
126.69
HS 8.07de
9.49 4.89 0.92 8.51 21738.00 19.50bcd
140.34
CHO1000 TN 14.49ab
6.58 3.72 0.60 9.00 17322.00 21.41abc
130.61
HS 13.22b
9.80 4.59 0.78 7.94 17865.00 20.09bcd
145.95
BET500 TN 8.95cd
7.78 3.12 0.59 8.21 12877.00 16.37cd
140.71
HS 8.20cd
8.95 5.42 0.77 8.36 18387.00 19.37bcd
142.78
BET1000 TN 11.74bc
7.06 5.17 0.61 7.34 15684.00 15.69d
137.30
HS 17.91a
2.49 4.37 0.97 8.85 18713.00 23.16ab
441.19
Pooled SEM 1.22 1.17 0.75 0.15 0.58 4679.46 2.03 93.52
Effect P values
Diet 0.001 0.46 0.28 0.76 0.74 0.66 0.35 0.86
Temp 0.16 0.01 0.02 0.12 0.14 0.09 0.04 0.34
Diet x Temp 0.03 0.21 0.30 0.89 0.18 0.92 0.04 0.93
Main Effect Means
Diet
CON 5.64 8.40 5.36 0.80 8.57 22418.00 21.76 142.17
CHO500 7.39 7.90 3.95 0.75 7.85 17631.00 19.79 133.51
CHO1000 13.85 8.19 3.95 0.69 8.47 17593.00 20.75 138.28
BET500 9.07 8.37 4.27 0.68 8.29 15632.00 17.88 141.74
BET1000 14.82 9.77 4.67 0.79 8.09 17199.00 19.43 289.24
Pooled SEM 0.86 0.82 0.53 0.11 0.41 5515.43 1.43 66.13
Temp
TN 9.58 7.14b
3.88b
0.61 7.98 15576.00 18.23 135.24
HS 10.74 9.91a
5.04a
0.87 8.52 20613.00 21.62 202.74
Pooled SEM 0.56 0.67 0.34 0.11 0.26 2093.11 1.06 42.28 1 Data are least squares means.
2 Metabolites: betaine (BET), free choline (CHO), glycerophosphocholine (GPC), acetylcholine (Acho),
total lysophosphatidylcholine (TLPC), total phosphatidylcholine (TPtCho), phosphocholine (PCho),
sphingomyelin (Sm). 3 TN = Thermoneutral environment, HS = Heat stress environment.
a,b,c,d Means within same column followed by different letters differ significantly (P < 0.05).
4 Data were rank transformed and untransformed means reported.
Page 106
93
Plasma Choline and Choline Metabolites
Plasma choline and choline metabolites concentration are presented in Table 24. There
was no diet or temperature effect or their interaction on PCho and Cho (P > 0.05). There was no
diet X temperature interaction (P = 0.10) on plasma GPC. However, both diet and temperature
significantly (P < 0.05) affected plasma GPC. Chickens consuming the CON diet (18.60 umol/l)
and BET500 (11.53 umol/l) had significantly higher plasma GPC concentration compared to the
rest of the diets (7.89 to 10.20 umol/l). Heat-stressed birds had significantly lower plasma GPC
(8.58 umol/l) compared to TN birds (13.87 umol/l).
Plasma TPtCho was not affected by diet, nor was there a diet X temperature interaction
(P > 0.05) whereas temperature significantly (P = 0.02) decreased TPtCho by 5835.82 umol/l
(11744.00 TN vs 5908.08 umol/l HS). There was no diet or diet X temperature interaction for
plasma Sm (P > 0.05) but temperature tended (P = 0.05) to increase it (339.94 TN vs 369.29
umol/l HS).
Betaine was not affected by temperature, nor was there a diet X temperature interaction
(P > 0.05). However, betaine level was significantly (P = 0.0001) decreased by diet. Birds
consuming BET1000 (219.75 umol/l) and CHO1000 (233.59 umol/l) had significantly higher
betaine concentration compared to the rest of the diets (99.29 to 166.67 umol/l). There was a diet
X temperature interaction for plasma TLPC (P = 0.04). This was caused by the CON decreasing
plasma TLPC by 49.06 (82.83 TN to 33.77 umol/l HS) while BET1000 decreased only by 4.55
(47.9 TN to 43.35 umol/l HS).
Page 107
94
Table 24. Effect of choline and betaine supplementation on plasma choline and choline
metabolites (umol/l) 1
of heat-stressed broilers
Metabolites2
Diet Temp3
BET CHO GPC TLPC TPtCho PCho Sm
CON TN 98.31 15.48 21.23 82.83a
15873.00 2.99 315.98
HS 100.27 14.16 15.97 33.77c
7404.34 2.90 375.07
CHO500 TN 161.98 15.10 10.19 66.09ab
9552.04 2.64 304.64
HS 160.05 15.49 5.59 36.09c
6404.87 2.37 354.08
CHO1000 TN 245.71 17.99 12.16 47.15bc
12329.00 3.20 337.32
HS 221.47 14.94 8.25 39.20c
5420.88 2.31 384.88
BET500 TN 187.28 13.94 14.59 47.75bc
8727.52 2.93 361.63
HS 146.06 16.30 8.48 32.43c
5761.40 3.57 380.85
BET1000 TN 213.08 16.18 11.16 47.90bc
12239.00 2.24 380.14
HS 226.43 13.50 4.62 43.35bc
6548.90 3.02 351.11
Pooled SEM 14.75 1.65 3.89 8.39 2318.04 0.51 23.49
Effect P values
Diet 0.0001 0.79 0.045 0.174 0.07 0.59 0.39
Temp 0.35 0.56 0.04 0.005 0.02 0.93 0.05
Diet x Temp 0.32 0.32 0.10 0.04 0.67 0.40 0.34
Main Effect Means
Diet
CON 99.29c
14.82 18.60a
58.30 11638.00 2.94 345.53
CHO500 161.02b
15.29 7.89b
51.09 7978.45 2.51 329.36
CHO1000 233.59a
16.46 10.20b
43.18 8874.92 2.76 361.10
BET500 166.67b
15.12 11.53ab
40.09 6244.46 3.25 371.24
BET1000 219.75a
14.84 7.89b
45.62 9393.94 2.63 365.63
Pooled SEM 10.43 1.17 2.74 5.94 1319.79 0.37 16.61
Temp
TN 181.27 15.74 13.87a
58.34 11744.00a
2.80 339.94
HS 170.86 14.88 8.58b
36.97 5908.08b
2.83 369.20
SEM 7.67 1.01 1.74 4.53 1585.91 0.28 10.51 1 Data are least squares means.
2 Metabolites: Betaine (BET), free choline (CHO), glycerophosphocholine (GPC), total
lysophosphatidylcholine (TLPC), total phosphatidylcholine (TPtCho),phosphocholine (PCho),
sphingomyelin (Sm). 3 TN = Thermoneutral environment, HS = Heat stress environment.
a,b,c Means within same column followed by different letters differ significantly (P < 0.05).
Page 108
95
Discussion
The villi height, crypt depth and villi:crypt ratio have been used in numerous studies to
assess the overall digestive and absorptive capacity of the small intestine. An increase in
villi:crypt ratio was shown to be positively correlated to the intestinal digestive and absorptive
capacity in pigs (McDonald, et al., 2001), in dogs (Kuzmuk, et al., 2005) and in chickens
(Burkholder, et al., 2008). The results of the present study showed that villus height, crypt depth
and villi height:crypt depth were unchanged in response to diet. However, there was a trend for
high temperature to decrease villi height in the ileum. This could decrease the digestive and
absorptive capacity of heat-stressed broilers. These results are consistent with the findings of
Quinteiro-Filho, et al. (2012) who reported no changes in villus height, crypt depth and villus
height:crypt depth ratio. However the results are not consistent with those reported by Mitchell
and Carlisle (1992) who found that villi height decreased by 19% in birds subjected to 35oC for
14 days. Acute heat stress has also been shown to decrease crypt depth as reported by
Burkholder, et al. (2008), in birds subjected to 30oC for 24h. These differences are probably due
to the fact that in the current experiment, birds were exposed to cycling temperature for longer
period of time (28 vs 14 days and 24h respectively) and might have adapted to the stress
condition.
In the present study, heat stress had no impact on most choline metabolites in the liver
except that it increased choline and GPC. Heat stress has been shown to increase de novo
lipogenesis which occurs primarily in the liver of chickens and to enhance fat deposition
(Geraert, et al., 1996; He, et al., 2015). Choline is involved in the synthesis of TPtCho needed for
the formation of very low density lipoprotein (VLDL), which is necessary for triglycerides
export from the liver (Kouba, et al., 1995). With an increase in lipogenesis during heat stress, the
Page 109
96
increase in choline level seen in the present study may be facilitating fat removal from the liver.
Similarly, an increase in GPC in the liver could be explained by its osmolytic property.
According to Kwon, et al. (1995), GPC is an osmolyte mainly known to protect renal cells
against hypertonicity. Therefore its increase in the liver cells could be seen as a mechanism to
restore or maintain osmotic balance. Unlike GPC, liver betaine was not affected by temperature
which suggests that GPC was preferably used in heat stress conditions to maintain osmotic
balance in the liver. Conversely, in previous studies, betaine has been shown to protect
hepatocytes kupffer cells from hyper osmosis in rat (Zhang, et al., 1996). However, there was a
diet X temperature interaction for liver betaine content with BET1000 increasing it by 34.45 %
under heat stress environment compared to thermoneutral. This is consistent with the results
reported by Saarinen, et al. (2001) who found that dietary betaine was efficient in increasing
hepatic betaine concentration. However, in their study birds were not exposed to stress
conditions. Similarly, Kettunen, et al. (2001b) reported that betaine accumulates in liver when
the diet is supplemented with higher levels of betaine. Therefore dietary supplementation of
betaine at 1000 ppm can be used to increase liver betaine content in broilers under heat stress
conditions.
Choline metabolites GPC, TPtCho, PCho, TLPC and Sm in the intestinal tissue were not
affected by diet, temperature or by their interaction in the current study. Similarly, there was no
temperature or diet X temperature interaction on intestinal betaine. However, it was affected by
diet. Intestinal betaine was increased by dietary supplementation of betaine and choline at 1000
ppm. This is consistent with the results reported by Kettunen, et al. (2001a) who found that
betaine accumulates in the small intestine and helps retain water in hyperosmotic saline solution.
In the current study, betaine did not accumulate in the intestinal tissue of heat-stressed broilers.
Page 110
97
This could be explained by the fact that heat stress condition did not severely increase osmotic
pressure on intestinal cells. Previous studies on heat stress impact on broilers did not evaluate
intestinal betaine content. However, when broilers were challenged with coccidia, (Kettunen, et
al., 2001b) reported an accumulation of betaine in the intestinal tissues. This difference may be
due to the fact that unlike heat stress, coccidiosis increases water loss through diarrhea resulting
in a more severe increase in osmotic pressure. Because heat stress did not affect betaine and GPC
in the present study it was not possible to determine their relative importance in providing
osmotic benefits on the intestine of heat-stressed broilers. Even though there was not a
significant diet X temperature interaction on intestinal choline content, the interaction showed an
increasing trend. All the diets except BET500 and BET1000 increased intestinal choline content
in heat-stressed chickens.
Plasma choline and PCho were not affected by diet and temperature or by their
interaction while plasma betaine content was increased only by diet. During heat stress there was
a large decrease in plasma GPC content compared to thermoneutral environment. This could be
explained by the fact that GPC was retained in the liver to serve as an osmolyte as shown by the
higher amount of GPC in the liver. Plasma TLPC was decreased in broilers reared under HS
conditions compared to those reared under TN conditions. The largest difference was seen
between TN and HS for the CON compared to the difference between TN and HS for CHO1000
or BET1000. This could suggest that during heat stress, supplementation with CHO1000,
BET1000 could limit the decrease in plasma TLPC. This is important for the body to maintain its
ability to prevent hemorrhaging since TLPC is one to the major components in blood clotting
(Xu, et al., 2013) and could thus have an impact on carcass quality. Plasma TPtCho was not
affected by diet and there was no diet X temperature interaction but was decreased by heat stress.
Page 111
98
References
Artegoitia, V. M., J. L. Middleton, F. M. Harte, S. R. Campagna, and M. J. de Veth. 2014.
Choline and choline metabolite patterns and associations in blood and milk during lactation in
dairy cows. PLoS One 9:e103412.
Belay, T., and R. Teeter. 1993. Broiler water balance and thermobalance during thermoneutral
and high ambient temperature exposure. Poult. Sci. 72:116-124.
Borsook, H., and J. W. Dubnoff. 1947. Methionine formation by transmethylation in vitro. J.
Biol. Chem. 169:247-258.
Burkholder, K., K. Thompson, M. Einstein, T. Applegate, and J. Patterson. 2008. Influence of
stressors on normal intestinal microbiota, intestinal morphology, and susceptibility to
Salmonella enteritidis colonization in broilers. Poult. Sci. 87:1734-1741.
Combs Jr, G. F. 1991. The vitamins: fundamental aspects in nutrition and health. Am. J. Clin.
Nutr. 53:755-763.
Cook, M., W. Springer, and J. Hebert. 1984. Enhanced incidence of leg abnormalities in reovirus
WVU 2937-infected chickens fed various dietary levels of selected vitamins. Avian Dis.:548-
561.
Geraert, P. A., J. C. F. Padilha, and S. Guillaumin. 1996. Metabolic and endocrine changes
induced by chronic heat exposure in broiler chickens: Biological and endocrinological
variables. Br. J. Nutr. 75:205-216. doi 10.1079/bjn19960125
He, S., S. Zhao, S. Dai, D. Liu, and S. G. Bokhari. 2015. Effects of dietary betaine on growth
performance, fat deposition and serum lipids in broilers subjected to chronic heat stress.
Anim. Sci. J. doi 10.111/asj.12372
Kettunen, H., S. Peuranen, and K. Tiihonen. 2001a. Betaine aids in the osmoregulation of
duodenal epithelium of broiler chicks, and affects the movement of water across the small
intestinal epithelium in vitro. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 129:595-603.
Kettunen, H., K. Tiihonen, S. Peuranen, M. Saarinen, and J. Remus. 2001b. Dietary betaine
accumulates in the liver and intestinal tissue and stabilizes the intestinal epithelial structure in
healthy and coccidia-infected broiler chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol.
130:759-769.
Klasing, K., K. Adler, J. Remus, and C. Calvert. 2002. Dietary betaine increases intraepithelial
lymphocytes in the duodenum of coccidia-infected chicks and increases functional properties
of phagocytes. J. Nutr. 132:2274-2282.
Kouba, M., D. Hermier, and M.-A. Bernard-Griffiths. 1995. Comparative study of hepatic VLDL
secretion in vivo in the growing turkey (Meleagris gallopavo) and chicken (Gallus
domesticus). Comp. Biochem. Physiol. B Mol. Integr. Physiol. 110:47-55.
Page 112
99
Kuzmuk, K. N., K. S. Swanson, K. A. Tappenden, L. B. Schook, and G. C. Fahey. 2005. Diet
and age affect intestinal morphology and large bowel fermentative end-product concentrations
in senior and young adult dogs. J. Nutr. 135:1940-1945.
Kwon, E. D., K. Jung, L. C. Edsall, H. Kim, A. Garcia-Perez, and M. Burg. 1995. Osmotic
regulation of synthesis of glycerophosphocholine from phosphatidylcholine in MDCK cells.
Am. J. Physiol-Cell. Ph. 268:C402-C412.
McDonald, D., D. Pethick, B. Mullan, and D. Hampson. 2001. Increasing viscosity of the
intestinal contents alters small intestinal structure and intestinal growth, and stimulates
proliferation of enterotoxigenic Escherichia coli in newly-weaned pigs. Br. J. Nutr. 86:487-
498.
McDowell, L. R. 1989. Choline. Pages 565-596 in Vitamins in Animal and Human Nutrition.
2nd ed.
Mitchell, M., and A. Carlisle. 1992. The effects of chronic exposure to elevated environmental
temperature on intestinal morphology and nutrient absorption in the domestic fowl (Gallus
domesticus). Comp. Biochem. Physiol. Part A Physiol. 101:137-142.
Molitoris, B. A., and D. H. Baker. 1976. The choline requirement of broiler chicks during the
seventh week of life. Poult. Sci. 55:220-224.
Mongin, P., M. Larbier, N. Carbobaptista, D. Licois, and P. Coudert. 1976. Comparison of
osmotic pressures along digestive-tract of domestic-fowl and rabbit. Br. Poult. Sci. 17:379-
382. doi 10.1080/00071667608416291
Nakanishi, T., and M. B. Burg. 1989. Osmoregulation of glycerophosphorylcholine content of
mammalian renal cells. Am. J. Physiol. 257:C795-801.
NRC. 1994. Nutrient Requirements of Poultry. 9th rev ed. Natl. Acad. Press, Washington, DC.
Petronini, P. G., E. De Angelis, P. Borghetti, A. Borghetti, and K. P. Wheeler. 1992. Modulation
by betaine of cellular responses to osmotic stress. Biochem. J 282:69-73.
Plagemann, P. G. 1968. Choline metabolism and membrane formation in rat hepatoma cells
grown in suspension culture: I. Incorporation of choline into phosphatidylcholine of
mitochondria and other membranous structures and effect of metabolic inhibitors. Arch.
Biochem. Biophys. 128:70-87.
Quinteiro-Filho, W., A. Gomes, M. Pinheiro, A. Ribeiro, V. Ferraz-de-Paula, C. Astolfi-Ferreira,
A. Ferreira, and J. Palermo-Neto. 2012. Heat stress impairs performance and induces
intestinal inflammation in broiler chickens infected with Salmonella Enteritidis. Avian Pathol.
41:421-427.
Saarinen, M. T., H. Kettunen, K. Pulliainen, S. Peuranen, K. Tiihonen, and J. Remus. 2001. A
novel method to analyze betaine in chicken liver: effect of dietary betaine and choline
Page 113
100
supplementation on the hepatic betaine concentration in broiler chicks. J. Agric. Food Chem.
49:559-563.
Sutherland, L., J. Cairney, M. Elmore, I. Booth, and C. Higgins. 1986. Osmotic regulation of
transcription: induction of the proU betaine transport gene is dependent on accumulation of
intracellular potassium. J. Bacteriol. 168:805-814.
Xu, H., N. Valenzuela, S. Fai, D. Figeys, and S. A. L. Bennett. 2013. Targeted lipidomics -
advances in profiling lysophosphocholine and platelet-activating factor second messengers.
Febs J. 280:5652-5667. doi 10.1111/febs.12423
Zeisel, S. 1993. Choline phospholipids: signal transduction and carcinogenesis. FASEB J. 7:551-
557.
Zeisel, S. H., M.-H. Mar, J. C. Howe, and J. M. Holden. 2003. Concentrations of choline-
containing compounds and betaine in common foods. J. Nutr. 133:1302-1307.
Zhang, F., U. Warskulat, M. Wettstein, and D. Haussinger. 1996. Identification of betaine as an
osmolyte in rat liver macrophages (Kupffer cells). Gastroenterology 110:1543-1552.
Page 114
101
Chapter VI: Summary and Conclusion
Page 115
102
The results of this study indicate that heat stress reduced feed intake and body weight
gain while increasing mortality, corticosterone levels and reducing thymus and bursa of
Fabricius weights. The supplementation of choline and betaine to heat-stressed broilers did not
improve growth performance or suppress the negative impact of high environmental temperature
on immune organ weights and corticosterone levels. Heat stress increased lightness and reduced
yellowness of breast meat color. Intestinal morphology showed no impact of heat stress on crypt
depth and villi height:crypt depth ratio. However heat stress tended to increase villi height.
Dietary supplementation of choline and betaine to heat-stressed broilers did not affect intestinal
villi height and crypt depth. Most choline metabolites were unaffected by diet or temperature or
by their interaction in the intestine, liver and plasma. However, supplemental choline and betaine
increased intestinal betaine contents. There was no effect of high temperature or diet by
temperature interaction for intestinal betaine and glycerophosphocholine contents however liver
choline and glycerophosphocholine were increased by heat stress. Adding choline and betaine to
provide 500 and 1000 ppm methyl equivalents respectively could increase increased liver betaine
contents in heat-stressed broilers. Plasma glycerophosphocholine was decreased by temperature
and supplementing with choline or betaine did not increase its content compared to the CON.
Meat color and drip loss are important qualities that might influence consumers’ choices.
However, further studies are needed including determination of the meat pH, cook loss and shear
force to fully appreciate the effect of heat stress and the supplementation of choline and betaine
on the meat quality. In this study, supplementation of choline and betaine did not improve
growth performance of heat-stressed broilers, however they improved foot heath and decreased
meat drip loss four days post slaughter.
Page 116
103
Vita
Kouassi R. Kpodo was born in Kissidougou (Guinea). He obtained a degree in Agronomy/option
animal production from the College of Agronomy in Togo. After graduation, he worked for non-
governmental agencies where he managed and implemented developmental programs that
focused on the integration of animal husbandry and leguminous plants to improve soil fertility in
rural areas. Kouassi emigrated in the USA in 2008 and decided to further his education in animal
science. In 2012, he received a bachelor degree in animal science from the University of
Tennessee, and was offered the opportunity to pursue a Master’s degree with Dr. Michael Smith
as mentor.