Journal of Ethnopharmacology, 30 (1990) 265-279 Elsevier Scientific Publishers Ireland Ltd. 265 POSSIBLE REGENERATION OF THE ISLETS OF LANGERHANS IN STREPTOZOTOCIN-DIABETIC RATS GIVEN GYMAEMA SYLVESZ’RE LEAF EXTRACTS E.R.B. SHANMUGASUNDARAM, K. LEELA GOPINATH, K. RADHA SHANMUGASUNDARAM and V.M. RAJENDRAN Department of Biochemistry, Postgraduate Institute of Basic Medical Sciences, University of Madras, Taramani Campus Madras 600-119 (Indial (Accepted June 25, 1990) Summary Two water soluble extracts, GS, and GS,, obtained from the leaves of Gymnema sylvestre, were tested in streptozotocin treated rats for their effects on blood glucose homeostasis and pancreatic endocrine tissue. In the diabetic rats, fasting blood glucose levels returned to normal after 60 days of GS, and after 20 days of GS, oral administration. Blood collected during the conduct of oral glucose tolerance tests was used to assay for serum insulin. GS, and GS, therapy led to a rise in serum insulin to levels closer to normal fasting levels. In diabetic rat pancreas, both GS, and GS, were able to dou- ble the islet number and beta cell number. This herbal therapy appears to bring about blood glucose homeostasis through increased serum insulin lev- els provided by repair/regeneration of the endocrine pancreas. Introduction Gymnema sylvestre R.Br., a herb belonging to the Asclepiadaceae family, has been employed to control diabetes by traditional medical practitioners of India for several centuries. The hypoglycaemic effect of this herb was scien- tifically tested for the first time by Mhasker and Caius (1930). In alloxan- induced diabetes in rabbits and dogs, these investigators observed a lowering of blood glucose levels, while in pancreatectomised animals the leaves of G. sylvestre did not have any hypoglycaemic effect. They concluded that some residual pancreatic endocrine tissue is needed for this herb to be effective. In a couple of diabetic patients, administration of the dried leaves for several weeks led to reduction in glucose in the urine collected for 24 h, Correspondence to: Prof. K. Radha Shanmugasundaram. 0378.8741/$03.50 0 1990 Elsevier Scientific Publishers Ireland Ltd. Published and Printed in Ireland
POSSIBLE REGENERATION OF THE ISLETS OF LANGERHANS IN STREPTOZOTOCIN-DIABETIC RATS GIVEN GYMAEMA SYLVESZ’RE LEAF EXTRACTS
E.R.B. SHANMUGASUNDARAM, K. LEELA GOPINATH, K. RADHA SHANMUGASUNDARAM and V.M. RAJENDRAN
Department of Biochemistry, Postgraduate Institute of Basic Medical Sciences, University of Madras, Taramani Campus Madras 600-119 (Indial
(Accepted June 25, 1990)
Summary
Two water soluble extracts, GS, and GS,, obtained from the leaves of Gymnema sylvestre, were tested in streptozotocin treated rats for their effects on blood glucose homeostasis and pancreatic endocrine tissue. In the diabetic rats, fasting blood glucose levels returned to normal after 60 days of GS, and after 20 days of GS, oral administration. Blood collected during the conduct of oral glucose tolerance tests was used to assay for serum insulin. GS, and GS, therapy led to a rise in serum insulin to levels closer to normal fasting levels. In diabetic rat pancreas, both GS, and GS, were able to dou- ble the islet number and beta cell number. This herbal therapy appears to bring about blood glucose homeostasis through increased serum insulin lev- els provided by repair/regeneration of the endocrine pancreas.
Introduction
Gymnema sylvestre R.Br., a herb belonging to the Asclepiadaceae family, has been employed to control diabetes by traditional medical practitioners of India for several centuries. The hypoglycaemic effect of this herb was scien- tifically tested for the first time by Mhasker and Caius (1930). In alloxan- induced diabetes in rabbits and dogs, these investigators observed a lowering of blood glucose levels, while in pancreatectomised animals the leaves of G. sylvestre did not have any hypoglycaemic effect. They concluded that some residual pancreatic endocrine tissue is needed for this herb to be effective. In a couple of diabetic patients, administration of the dried leaves for several weeks led to reduction in glucose in the urine collected for 24 h,
Correspondence to: Prof. K. Radha Shanmugasundaram.
confirming its action. Prolonged administration (24 weeks) of the dried leaves of the herb to alloxan-diabetic rabbits brought about increased serum insulin levels both at fasting and after a glucose load (Shanmugasundaram et al., 19811.
In alloxan-induced diabetes, the insulin producing cells are damaged and, to bring about blood glucose and serum insulin homeostasis, either regenera- tion or repair of the beta cells in the islets of Langerhans would appear to be necessary. The data presented here provide evidence that G. sylvestre extracts do increase islets and beta cells in experimentally induced diabetes in albino rats. Streptozotocin was used to induce diabetes, rather than alloxan, since with this agent there is (i) no incidence of spontaneous rever- sion and (ii1 greater specificity for islets resulting in >90% of the rats becoming diabetic at the dosage used.
Two concentrates of the leaf extract were tested. GS, was obtained from GS, during purification and is a component of the latter.
Materials and Methods
Drug materials Collagenase (Type Vl and streptozotocin were obtained from Sigma Chemi-
cals Co., St. Louis, U.S.A. and radioimmunoassay reagents for insulin assay was from Bhabha Atomic Research Centre, Bombay, India.
Crushed dried leaves of authentic G. sylvestre were steeped in 50% ethanol and steam passed into the mixture for 3- 4 h. The resultant alco- holic extract was filtered before being concentrated under vacuum and acidi- fied to pH 3.0. The residue obtained (GS,, 7% yield w/w relative to starting material) was then further purified by dissolving in 0.1 N sodium hydroxide and reprecipitation with acid to give GS, (3% yield w/w relative to starting material). Both GS, and GS, were tested for their pharmacological effects in diabetic rats in separate experiments.
Animal stock Male albino rats (derived from the Wistar strain) weighing 120-150 g
were used. They were obtained from the inbred colonies maintained in the King’s Institute for Preventive Medicine, Madras. All animals were screened for detection of abnormalities in blood glucose homeostasis by subjecting them to an oral glucose tolerance test. Only those animals meeting the fol- lowing three criteria were used: (i) fasting blood glucose levels below 85 mgl dl, (ii) 60 min after glucose load, a blood glucose level below 150 mgldl and (iii) a return to fasting levels in 150 min.
Experimental diabetes Diabetes was induced in overnight fasted rats by intravenous (i.v.1 injec-
tion of 55 mg/kg streptozotocin using a 5% solution of freshly prepared streptozotocin in 0.1 M citrate buffer (pH 4.51. Control rats received citrate
267
buffer only. Fasting blood glucose was measured and glucosuria was detected in all the animals seven days after streptozotocin administration. All animals were given powdered food (Gold Mohur rat feed, Lipton Co., Bombay) and water ad libitum.
Five weeks (35 days) after the injection of citrate buffer (normal) or strep- tozotocin (diabetic), half of the normal and diabetic animals were adminis- tered the extracts, while the other half remained untreated for comparison. The dosage used in both cases (GS, and GS,) was 20 mgldaylrat. GS, and GS, in powder form was mixed with 3 g of powdered food and moistened with water to make a bolus which the rats ate in the mornings. Food trays were filled only after the drug-mixed rations were fully consumed. The presence of reducing sugar in the urine was tested using Benedict’s solution daily and fasting blood glucose was estimated every 5 days by the o-toluidine colour reaction (Cooper and McDaniel, 19701. In the case of GS,-administered dia- betic rats (DGS, group), the fasting blood glucose reached the normal range after 60 days (95 days after induction of diabetes), while with GS, therapy (DGS, group) the fasting glucose level was normal on the 20th day and GS, administration discontinued forthwith. The rats were maintained for a fur- ther period of 40 days on food ad libitum. On the 95th day a glucose toler- ance test (GTT) was performed on both treated groups (DGS, and DGS,) and their respective controls (N, NGS, and NGS,). Group N is the control group, while NGS, and NGS, are groups of non-diabetic rats under GS, and GS, therapy, respectively. Serum insulin levels were assayed in all the blood samples. Two days later, animals were sacrificed and pancreatic tissue was harvested.
Glucose tolerance test and insulin assay Oral glucose tolerance tests (OGTT) were carried out according to the
method of Cole and Harned (19381 as modified by Wexler and Fischer (1963). After an overnight fast, blood samples were collected and then 200 mg glu- cose in 2 ml solution administered at 30, 60, 120 and 180 min. Blood glucose levels were estimated by the o-toluidine method of Cooper and McDaniel (19701. Serum insulin levels were assayed by radioimmunoassay (Herbert et al., 19651 in all blood samples collected during OGTT.
Histologic and morphometric studies on the pancreas
Each animal was sacrificed by decapitation and the whole pancreas per- fused with formalin and removed immediately together with the spleen. The total pancreatic weight was recorded. The three regions of pancreas, viz. duodenal (head), gastric (body) and spleen (tail), were dissected according to the specifications of Jaffe (19511, cut into smaller fragments and fixed sepa- rately in Bouin’s fluid for 24 h. The segments were dehydrated with ethanol and embedded in paraffin-wax (56OC). Serial sections (5 pm1 were taken and stained with chrome-haematoxylin and phloxin according to Gomori (1941).
268
One hundred serial sections were studied for the number of islets and beta cell content in each one of the three regions of the pancreas for each rat. Beta cells stain deep purple and photographs were taken with green filter to obtain maximum contrast. Number of rats for which histological studies were made was 10 in each group.
Morphometric studies were made using stereological procedures according to Wiebel and Elias (1967) using a reticule. The reticule was mounted along the focal plane of the eye piece, and consisted of 100 squares. At 400 x mag- nification, the area covered by the 100 squares was 0.0625 mmz. The area covered by the section and diameters of the islets and beta cells were meas- ured by counting the number of squares in the reticule occupied by the structures. With the thickness of the sections at 5 pm, the volume of the sec- tion and islets was arrived at to express the percentage endocrine tissue.
In vitro studies on isolated islets of Langerhans From one set of normal and experimental rats, islets were isolated and
insulin release under stimulation with glucose and extract GS, was studied. The rats were sacrificed by decapitation and the pancreas was removed through an incision made in the ventral abdominal duct and placed in 3.0 ml Hank’s solution. The tissue taken from two rats was minced with a pair of scissors and incubated with (35 - 50 mg) collagenase (Type V) in 5.0 ml Hank’s solution at 3’7OC for 6- 12 min with constant stirring according to the method of Lacy and Kostianovsky (1967) as modified by James and Keith (1968). Aliquots of the mixture were examined periodically under a dissecting microscope to determine the point at which optimum separation of islets had occurred.
The solution was diluted to about 30 ml with Hank’s solution and the supernatant containing predominantly acinar tissue was removed. The resi- dual islets were washed eight times with Hank’s solution and the sedimenta- tion period was progressively reduced from 4 to 1 min. The islets sedimented under gravity. The washed islet suspension was poured into a Petri dish surrounded by crushed ice and examined under a dissecting micro- scope. The islets appeared as free, round or ovoid structures with greyish white to brownish red colour.
They were transferred in groups of 10 to the incubation mixture contain- ing 1.0 ml of ice cold Krebs-Hensleit buffer (KHB) with 3.3 mM glucose, capped and gassed with oxygen/carbon dioxide mix (95:5 v/v) for 3 min and incubated for 30 min in a Dubnoff metabolic shaker at 37OC. The buffer was removed and the islets were washed with physiological saline. They were used in the insulin secretion studies described below.
To study GS,-mediated release of insulin from the islets, two different levels of the crude drug (80 mg and 160 mg/lOO ml) were tested with glucose at 30, 150 and 300 mg/lOO ml at 37OC. In each case the reaction mixture con- tained 1.0 ml Krebs-Hensleit buffer containing the test substance and 10 islets. A O.l-ml sample of the fluid was removed at 0, 30 and 60 min and insu-
269
lin content was assayed by radioimmunoassay. The net insulin released was corrected for dilution and is expressed here as the amount of immunoreac- tive insulin (IRI) released per 10 islets. The in vitro studies on isolated islets were preferred over pancreatic perfusion, so as to assess the efficiency of a definite number of islets (in releasing insulin) from the normal and experi- mental animals before and after therapy. However, the possibility of missing small regenerating islets during isolation cannot be ruled out.
In a third set of rats (normal, NGS, and DGS, groups), pancreas tissue was excised at the end of the experimental period and homogenised with cold distilled water (4OCl. Amylase and lipase activities were assayed in the hom- ogenate according to the method of Bernfield (1955) and Cherry and Cran- dale (19321, respectively.
For statistical analysis, the diabetic rats (D group) were compared with the normals (N group), while those after GS, and GS, therapy were com- pared with their respective controls without the herbal therapy, i.e. the NGS, and NGS, groups were compared with group N, while the DGS, and DGS, groups were compared with group D. Student’s t-test was per- formed. Results
It can be seen from Table 1 that both G’S, and GS, are able to lower fast- ing blood glucose levels in the diabetic rats (DGS, and DGS, groups). With GS, administration, fasting glucose level was controlled at the end of 60 days (95th day), while almost the same degree of response was observed at the end of 20 days with GS, therapy. Mean blood glucose remained around 90 mg/dl, even after discontinuing GS, therapy. Pancreatic amylase and lipase activities lowered in the diabetic group (Dl are brought back to normal levels after GS, therapy. Glucosuria disappeared in the DGS, group on 75th day and in the DGS, group on 55th day.
From Table 2 it can be seen that the diabetic rats after GS, or GS, ther- apy had a near normal glucose tolerance curve. While the GTT was per- formed on GS, treated diabetics 40 days after termination of GS, therapy, both the fasting and postprandial blood glucose levels were under control and insulin levels in serum were near normal.
Such complete glucose homeostasis in streptozotocin diabetes suggests the repair or regeneration of the insulin secreting beta cells in the islets of Langerhans. When the herbal concentrates are administered to healthy rats (NGS, and NGSJ insulin secretion is clearly augmented (Table 2).
In control rats (NJ, the islets of Langerhans were distributed throughout the three regions of the pancreas, viz. duodenal (head), gastric (body) and splenic (tail) (Table 31. The islets of the duodenal and gastric regions were more or less of the same diameter, but in the splenic region the islets were larger and double the size. In the streptozotocin diabetic rats, the gastric region of the pancreas did not show any islets (Table 31 indicating that this is the major target for streptozotocin.
TA
BL
E
1
FAST
ING
B
LO
OD
GL
UC
OSE
(m
g/lO
O m
l) V
AL
UE
S D
UR
ING
T
HE
E
XPE
RIM
EN
T
Ani
mal
s of
gro
ups
N,
NG
S,
and
NG
S,
form
ed
one
grou
p un
til
the
35th
da
y w
hen
they
w
ere
divi
ded.
Si
mila
rly
the
diab
etic
an
imal
s (s
trep
tozo
to-
tin
inje
cted
) w
ere
divi
ded
into
th
ree
grou
ps
D,
DG
S, a
nd
DG
S,
on t
he
35th
da
y.
Eac
h ta
bula
r va
lue
is t
he
mea
n +
S.
E.M
. ob
tain
ed
in m
ore
than
30
rat
s in
eac
h gr
oup
stud
ied
in f
ive
batc
hes.
N
D
=
not
dete
rmin
ed.
Tes
t N
umbe
r of
day
s N
D
N
GS,
D
GS,
N
GS,
D
GS,
Bla
od
gluc
ose
Am
yIas
&
Lip
ased
Initi
al
76
r 0.
9 76
2
0.9
76
k 0.
9 76
+
0.
9 76
f
0.9
76 +
0.
9
15
79
* 1.
3 19
0 -t
6.
8’
79
+
1.3
190
f 6.
8’
79 f
1.
3 19
0 f
6.8
35
81
f 1.
6 19
7 f
6.6’
80
-c
1.
6 19
9 f
5.5’
79
f
1.1
192
-+ 6
.8
55
76
-I-
2.0
200
2 5.
8’
74
f 1.
5 15
0 2
5.5
71 f
1.
1 93
f
2.0
75
75
f 1.
6 21
6 f
6.4c
70
-’
1.
6 10
3 -r
- 3.
3 72
+
1.5
90
f 1.
6 95
80
=
1.
8 22
0 2
6.2
68
2 0.
9 76
k
1.5
71 f
0.
9 92
f
2.0
96
685
zk 4
3 10
4 r
5 73
0 +-
37
641
f 46
N
.D.
N.D
. 96
29
.4
k 1.
4 13
.8 -
t 1.
0 38
.5 -
c 2.
1 39
.3 -
’ 1.
1 N
.D.
N.D
.
‘GS,
w
as a
dmin
iste
red
from
36
th t
o 95
th
days
. bG
S, w
as a
dmin
iste
red
from
36
th
to 5
5th
day
only
, al
thou
gh
the
anim
als
wer
e un
der
obse
rvat
ion
thro
ugh
the
95th
day
. c G
luco
suri
a pr
esen
t. d A
myl
ase
(mIU
/mg
prot
ein)
an
d lip
ase
(mIU
/mg
prot
ein)
w
ere
assa
yed
in p
ancr
eatic
ho
mog
enat
es.
TA
BL
E
2
BL
OO
D
SU
GA
R
AN
D
SE
RU
M
INS
UL
IN
LE
VE
LS
O
F
NO
RM
AL
A
ND
D
IAB
ET
IC
AN
IMA
LS
(B
EF
OR
E
AN
D
AF
TE
R
GS
, T
RE
AT
ME
NT
) D
UR
ING
OR
AL
GL
UC
OS
ET
OL
ER
AN
CE
T
ES
TS
C
AR
RIE
D
OU
T
AT
T
HE
E
ND
O
F T
HE
E
XP
ER
IME
NT
AL
P
ER
IOD
Th
e va
lues
of
gl
uco
se
and
insu
lin
ar
e ex
pres
sed
as
mg/
lOO
m
l bl
ood
and
pIU
/ml
seru
m
resp
ecti
vely
(m
ean
f
S.E
.M.
for
10 a
nim
als
in
each
gr
oup
).
NG
S,
and
NG
S,
indi
cate
n
orm
al
hea
lth
y an
imal
s on
G
S,
and
GS
, th
erap
y,
resp
ecti
vely
. D
GS
, an
d D
GS
, in
dica
te
diab
etic
an
imal
s af
ter
trea
tmen
t w
ith
G
S,
and
GS
,, re
spec
tive
ly.
Gro
up
F
asti
ng
+ 3
0 m
in
+ 6
0 m
in
+ 1
20 m
in
+ 1
80 m
in
N
NG
S,
NG
S,
D
DG
S,
DG
S,
Glu
cose
In
suli
n
Glu
cose
In
suli
n
Glu
cose
In
suli
n
Glu
cose
In
suli
n
Glu
cose
In
suli
n
Glu
cose
In
suli
n
77
* 1.
3 15
.9
+
0.9
68
f 1.
6 18
.6
f 0.
6 71
f
1.3
19.9
+
1.
6
217
f 4.
1 6.
3 +
0.
6 77
f
2.2
20.0
f
0.5
93
+
3.5
14.9
+
1.
0
137
f 1.
9 28
.6
f 0.
8
99
f 4.
1 33
.5
+
0.7
122
+
2.9
35.2
-c
0.
7 29
5 +
5.
1 9.
4 +
0.
7 11
2 f
5.1
29.0
f
0.5
156
f 4.
1 25
.2
” 0.
7
99
2 2.
5 21
.1
f 0.
8
97
+
2.9
25.5
f
0.6
88
f 2.
2 26
.4
f 1.
2
353
f 3.
8 9.
6 f
0.7
98
f 3.
8 27
.0
-c
0.6
133
f 2.
9 23
.6
-c
3.0
83
2 2.
2 16
.9
2 0.
6
77
2 1.
3 23
.8
f 0.
5 69
f
1.9
20
+
1.3
366
f 5.
7 9.
6 f
0.5
95
+
6.0
25.2
+
0.
5
123
f 3.
1 20
.2
f 0.
8
80
+
1.3
15.3
+
0.
7
69
+
1.3
19.1
f
0.8
67
f 1.
9 16
.1
+
1.1
396
f 5.
1 9.
6 +
0.
5 86
f
2.5
22.1
+
0.
7
99
f 2.
2 19
.9
+
0.8
TA
BL
E
3
ME
AN
N
UM
BE
R
f-(_
S.E
.M.)
A
ND
SI
ZE
O
F IS
LE
TS
OF
LA
NG
ER
HA
NS
AN
D
NU
MB
ER
O
F B
ET
A
CE
LL
S/IS
LE
T
IN
NO
RM
AL
A
ND
D
IA-
BE
TIC
, W
ITH
A
ND
W
ITH
OU
T
GS,
A
ND
G
S,
TH
ER
APY
The
nu
mbe
r is
ex
pres
sed
per
mm
3.
Ana
lysi
s ar
e ba
sed
on
10
anim
als
in
each
gr
oup
and
100
seri
al
sect
ions
in
ea
ch
anim
al
for
each
re
gion
.
Gro
up
Panc
reas
wei
ght
(g)
Duo
dena
l
Num
ber
of
isle
ts
Dia
met
er
(pm)
Gas
tric
Bet
a ce
ll/
isle
t
(N)
Num
ber
of
isle
ts
Sple
nic
Dia
met
er
Bet
a ce
ll/
(pm
) is
let
w)
Num
ber
of
isle
ts
Dia
met
er
(pm
)
Bet
a ce
ll/
isle
t
vv)
N
1.12
+
0.15
NG
S,
1.13
5
0.60
NG
S,
1.26
2
0.07
D
0.75
f
0.07
DG
S,
0.99
2
0.05
D
GS,
1.
03
k 0.
06
48
f 4.
4
53
2 2.
2
55
f 3.
5
28
2 2.
5++
57
f 2.
2-e
44
f 4.
4**
58
f 4.
4
65
f 5.
7
62
f 5.
4
41
2 4.
4’
53
+ 4.
7
33
r 2.
9
247
f 28
341
f 36
304
* 29
77
2 12
+++
144
k 18
**
74k
7
110
f 11
.1
120
-r-
6.0
109
k 6.
0
0
28
2 1.
9
36
2 4.
1
52
+ 8.
0 11
3 2
16
85
2 5.
7
59
2 4.
7 10
1 f
14
93
f 6.
0
55
2 6.
0 11
6 +
14
87
k 5.
7
0 0
34
f 4.
1+++
38
f 4.
1 82
2
10
73
2 4.
7-e
27
f 4.
1 52
f
13
82
+ 6.
6***
138
+ 11
.4
138
2 11
.7
133
-+
14.2
68
+ 1.
3+++
98
f 5.
4**
92
-+
6.6*
*
635
f 83
719
k 69
778
f 11
5
156
2 lo
+++
295
2 42
*
272
2 50
*
Sign
ific
ance
re
lativ
e to
G
roup
N
: +P < 0.05;
++P < 0.01;
+++P
< 0
.001
.
Sign
ific
ance
re
lativ
e to
co
rres
pond
ing
untr
eate
d gr
oup:
*P < 0.05;
**P < 0
.01;
***P < 0
.001
.
_
273
GS, and GS, treatment helped to maintain the weight of pancreas, number of islets and number of beta cells in the diabetic animals (Table 3).
It is interesting to note that with GS, and GS, therapy, the total pan- creatic weight increased by about 30% in diabetic animals, while the number of islets and beta cells were more than doubled. At the same time, the increases in number of islets and beta cells of the normal animals treated with GS, and GS, were not statistically significant, suggesting a regulated response of the pancreatic tissue to stimulation by the G. sylvestre leaf extracts.
The streptozotocin-diabetic animals had damage over the pancreatic acinar region as can be seen by reduced pancreatic amylase and lipases and these decreases are corrected after GS, therapy (Table 1).
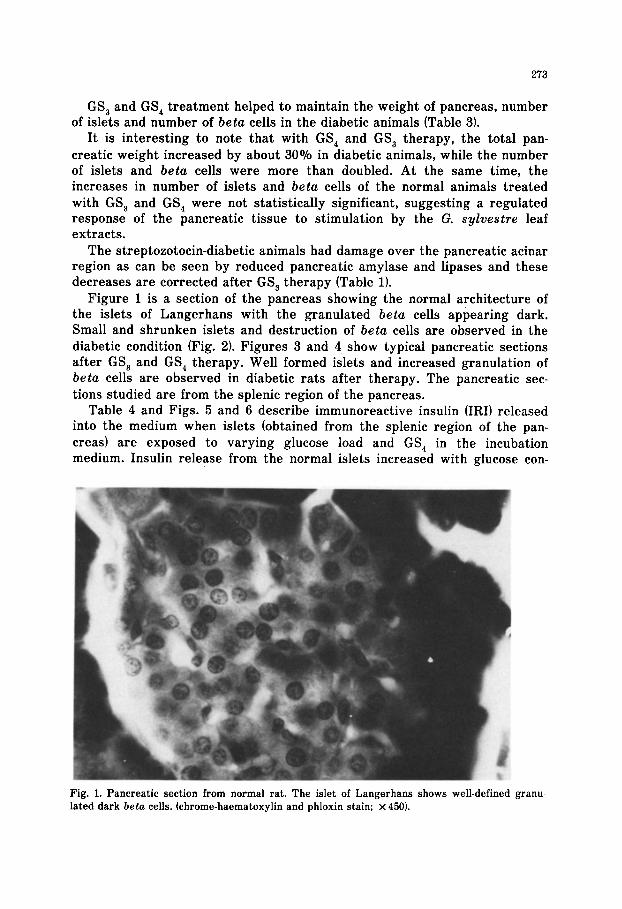
Figure 1 is a section of the pancreas showing the normal architecture of the islets of Langerhans with the granulated beta cells appearing dark. Small and shrunken islets and destruction of beta cells are observed in the diabetic condition (Fig. 21. Figures 3 and 4 show typical pancreatic sections after GS, and GS, therapy. Well formed islets and increased granulation of beta cells are observed in diabetic rats after therapy. The pancreatic sec- tions studied are from the splenic region of the pancreas.
Table 4 and Figs. 5 and 6 describe immunoreactive insulin (IRI) released into the medium when islets (obtained from the splenic region of the pan- creas) are exposed to varying glucose load and GS, in the incubation medium. Insulin release from the normal islets increased with glucose con-
Fig. 1. Pancreatic section from normal rat. The islet of Langerhans shows well-defined granu-
lated dark beta cells. (chrome-haematoxylin and phloxin stain; x 450).
274
Fig. 2. Diabetic rat pancreas, showing imperfections. The islet is atrophied and hydropic ch are seen ( x 450).
,a
Fig. 3. Pancreatic section of a diabetic rat after GS, therapy. The islet is large and organised ( x 450).
275
Fig. 4. Islet from a diabetic rat after GS, therapy. The beta cells appear dark, well developed and granulated ( x 1000).
centration (Table 41. With islets obtained from control rats at the severely hyperglycaemic state of 300 mg glucoseldl, the maximum insulin released was 397 pIUnits/lO islets (i.e. 40 $Units/isletl. By increasing glucose level from 30 to 300 mg/dl (Table 41, insulin release was increased eight-fold and this can be taken as the reserve capacity of the pancreas available for secre- tion. Under similar conditions, diabetic rat islets released a maximum of 100 $Units/lO islets, while in the DGS, and DGS, groups (diabetes controlled by GS, and GS, therapy, respectively), glucose mediated insulin release was nearly 120 $Units/lO islets or 12 PIUnitslislet.
GS, stimulated insulin release from the normal islets to a very significant degree even with the hypoglycaemic level of 30 mgldl, but in hyperglycaemic conditions (150 and 300 mgldll, insulin secretory capacity and the maximum release was about 100 I.tIUnits/lO islets or 10 pIU/islet. In diabetic rats con- trolled by GS, and GS, therapy (DGS, and DGS, groups), insulin release as a response to glucose load was marginally higher than the diabetics at 30 min. When the islets were incubated with glucose at hyperglycaemic levels (300 mgldll, GS,-stimulated insulin release was 202 and 194 pIUnits/lO islets (or 20 $Units/isletl after GS, and GS, therapy, respectively. After therapy, the diabetic islets appear to have a reserve capacity for insulin release which is double that of the untreated diabetics, but only one half of the insulin release observed in the non-diabetic controls. However, at 300 mg/dl glucose, GS, induced insulin released from the islets appears to be reduced at the end
TA
BL
E
4
EFF
EC
T
OF
GL
UC
OSE
A
ND
G
S O
N
IN
VIT
RO
IN
SUL
IN
RE
LE
ASE
FR
OM
IS
LE
TS
OF
NO
RM
AL
(N
), U
NT
RE
AT
ED
D
IAB
ET
IC
(D),
AN
D
CO
NT
RO
LL
ED
-DIA
BE
TIC
(D
GS,
A
ND
D
GS,
) R
AT
S
Insu
lin
rele
ase
(pIU
/lO
isle
ts)
into
th
e m
ediu
m
at
the
end
of
30
and
60
min
is
gi
ven.
E
ach
incu
batio
n w
as
prec
eded
by
a
30.m
in
prei
ncub
atio
n w
ith
60 m
g gl
ucos
e/l0
0 m
l. T
he
resu
lts
repo
rt
the
mea
n of
fo
ur
obse
rvat
ions
of
ea
ch
pair
of
rat
s fo
r 5
pair
s of
ani
mal
s in
ea
ch
grou
p (m
ean
+ S.
E.M
.).
_ 30
30
60
- 15
0 30
60
GS
, (m
g/lO
O
ml)
_ 30
0 30
60
160
30
30
60
160
150
30
60
160
300
30
60
Glu
cose
tmg/
lOO
m
l)
Tim
e
(min
)
N
D
DG
S,
DG
S,
27
-t
2.5*
**
31
2 1.
5***
34
f 3.
0**
39
2 1.
5**
87
+ 7.
0*
116
2 3.
0*
41
2 2.
5**
39
2 2.
0**
96
+ 2.
5***
196
2 9.
0***
202
* 2.
0***
175
+ 13
.0**
*
29
2 1.
5
55
+ 3.
5
111
f 5.
0
249
2 17
.0
226
+ 8.
5
397
+ 15
.5
52
2 2.
5
89
2 2.
5
106
2 6.
0
206
+ 8.
0
235
+ 16
.0
190
2 9.
5
19
+ 1.
0+
22
+ 1.
0’
25
f 1.
5+
31
+ 3.
0+
79
f 7.
5t
100
k 3.
5’
26
+ 3.
5+
27
-t
3.5+
72
2 2.
5*
99
k 4.
5+
90
2 4.
0’
85
f 4.
5’
28
f 1.
5***
31
f 0.
5***
36
f 1.
5**
40
2 1.
0**
92
f 7.
5*
120
+ 4.
5*
35
2 2.
0**
37
2 2.
0**
94
f 3.
0***
136
+ 20
.0**
*
194
k 18
.0**
*
140
-r-
14.5
***
Sign
ific
ance
re
lativ
e to
G
roup
N
: +P
<
0.00
1.
Sign
ific
ance
re
lativ
e to
G
roup
D
: *P
<
0.05
; **
p <
0.
01;
***p
<
0.
001
277
150 mg Glucose
T m I
1
150mq Glucosetl60mg GS4 150 mg Glucose teomg GS,
30 60 30 60 30 60 30 60 30 60 30 60
Time (min)
f
30 60 30 60 30 60
Fig. 5. Insulin released in 30 and 60 min when islets from 0, normal; 0, diabetic; and W, DGS, groups were incubated with glucose at 160 mg/dl with and without GS, at 80 and 160 mg/dl. Mean insulin ( f S.E.M.) released into the media by 10 islets are represented.
of 60 min, when compared to the hormone in the medium at 30 min, which may be due to inactivation by GS, or reuptake into the islets.
Discussion
The data presented give evidence for increased insulin secretion and beta cell number after the administration of leaf extracts of G. sylvestre, suggest- ing possible regeneration or repair of the islets of Langerhans in streptozoto- tin-treated rats.
Experiments of Loubatieres (1944, 19461, first established that the proba- ble mode of action of sulphonylureas was by the release of pancreatic insulin, which was confirmed by Houssay and Penhos (1956). Unlike the sulphonylu- reas, GS, does not enhance insulin release in normal rats under normogly- caemic (100 mg/dl) conditions, but enhances hormone release in diabetic islets. It is here that G. sylvestre leaf extracts assume significance, because of their capacity to regenerate partially the damaged endocrine tissue, such that the insulin content/islet number increases by therapy.
Hitherto, it was believed that the destruction of beta cells in juvenile, maturity-onset and experimentally induced diabetes was irreversible and
278
LOO
z 300 al
5
0 < 3
3 200
I%
4
a”
.c- 3 ; 100 H
300 mg Glucose
300 mg Glucose + 80 mg GS4
300 mg Glucose + 160mg GS4
T TT
30 60 30 60 30 60 30 60 -30 60 30 60
Time (min)
30 60 30 60 30 60
Fig. 6. Insulin released in 30 and 60 min when isolated islets from q , normal; Ha, diabetic; and W. DGS, groups were incubated with glucose at 300 mgldl with and without GS, at 80 and 160 mg/ dl. Mean insulin ( + S.E.M.) released into the medium by 10 islets are represented.
this report is the first giving evidence of a reversal of the damage to the insulin producing cells by a drug. Mitotic studies are needed to confirm the appearance of new cells.
The chemical composition of GS,, the active beta cell regenerative portion of the leaves of G. sylvestre, remains unknown. The leaves have an antisac- charogenic property (suppression of sweet taste for several hours when the leaves are chewed) that has been recorded repeatedly in the literature (Cho- pra et al., 1928). Hooper (1889) isolated a crude alcohol-soluble extract having this antisaccharogenic property and named it as gymnemic acid. Gymnemic acid (a glycoside) was purified and characterised by Stocklin (1967) as a D glucuronide of a hexa-hydroxy A12-oleanene. However, this purified isolate did not show any antidiabetic activity. Mhasker and Caius (1930) analysed the leaves and reported the presence of 0.95% hentriacontane, in addition to chlorophylls, phytin, resins and anthraquinone derivatives which give a pur-
279
gative effect to the leaves. None of the isolates were found to have antidi- abetic activity, although the whole leaf powder was able to bring down blood glucose levels not only in experimental diabetes, but also in man (Mhaskar and Caius, 1930). However, in pancreatectomised animals, G. sylvestre leaves did not show any hypoglycaemic effect, suggesting that residual pancreatic function is essential for its antidiabetic activity. When these observations are viewed with the data presented in this paper, it may be concluded that the extract of G. sylvestre may bring down blood glucose levels by regenera- tion of the pancreatic islets and beta cells.
References
Bernfield, P. (1955) In: S.P. Colowich and N.P. Kaplan (Eds.1 Methods in Enzymology 1, 149- 165. 165.
Cherry, I.S. and Crandale, I.A. (19321 The specificity of pancreatic lipase: Its appearance in the blood after pancreatic injury. American Journal of Physiology 100,266-273.
Chopra, R.N., Bose, J.P. and Chatterjee, N.R. (19281 Indigenous drugs of India. Indian Journal of Medical Research 16,115- 124.
Cole, V.V. and Harned, B.K. (19381 Diabetic traits in a strain of rats. Endocrinology 23, 318-326.
Cooper, G.R. and McDaniel, V. (19701 In: R.P. MacDonald (Ed.1 Standard Methods of Clinical Chemistry, Vol. 6, Academic Press, London, p. 159.
Gomori, G. (19411 Observations with differential stains on human islets of Langerhans. American
Journal of Pathology 17, 395-406.
Herbert, V., Law, K.S. Gotilieb, C.N. and Bleicher, S.J. (19651 Coated charcoal immunoassay of insulin. Journal of Clinical Endocrinology 25, 1375- 1384.
Hooper, D. (18891 Gymnema sylvestre, R. Br., Chemical News 59, 159 and Chemical Zentrablatt I, 632.
Houssay, B.A and Penhos, J.C. (19561 Action of the hypoglycaemic sulphonyl compounds in hypo- physectomised, adrenalectomised and pancreatectomised animals. Metabolism 5, 727-738.
Jaffe, F.A. (19511 A quantitative study of the islets of Langerhans in the rabbit. Anatomic Recu- rie 111,109-121.
James, E.V. and Keith, D.B. (19681 Effect of glucose concentration on insulin and glucagon from isolated islets of Langerhans of rats. Diabetes 17, 187- 193.
Lacy, P.E. and Kostianovsky. M. (19671 Method for the isolation of intact islets from the rat pan- creas. Diabetes 16,35-40.
Loubatieres, A. (19441 Analyse de mechanisma del’action hypoglycaemisule due p-aminobenzene sulphamidothiodiazol(2254 R.P.1, Comptes Rendus des Seances de la Societe de Biologie et de ses Filiales (Paris) 138,766 - 778.
Loubatieres, A. (19461 Etude physiologique et pharmacodynamique de certains derives fulfam- ides hypoglycaemiant. Archives Internationales de Physiologie 54, 174-184.
Mhaskar, K.S. and Caius J.F. (19301 A study of Indian medicinal plants. II: Gymnema sylvestre R. Br. Indian Medical Research Memoirs 16, 2- 75.
Shanmugasundaram, K.R. Panneerselvam, C., Samudram, P. and Shanmugasundaram, E.R.B. (19811 Insulinotropic activity of Gymnema sylvestre R.Br., an Indian medicinal herb used in controlling diabetes mellitus. Pharmacological Research Communications 13, 475-485.
Stocklin, W., Weiss, E. and Raichstein, T. (19671 Gymnema Saure, das antizaccharin Prinzip von G. sylvestre, R. Br. Helvetica Chimica Acta 50, 474-490.
Weibel, E.R. and Elias, H. (19671 General Stereologic Principles. Quantitative Methods in Mor- phometry, Springer Verlag, Berlin.
Wexler, B.C. and Fischer, C.T. (19631 Abnormal glucose tolerance in repeatedly bred rats and with atherosclerosis. Nature 200, 133- 136.















![Untitled Document []€¦ · Title: Untitled Document Created Date: 191000309095417](https://static.documents.pub/doc/80x56/600101983d1deb13132b1db7/untitled-document-title-untitled-document-created-date-191000309095417.jpg)
![Untitled Document [docs.oracle.com] · Title: Untitled Document Created Date: 191000801172643](https://static.documents.pub/doc/80x56/603a60be0019ab59b214b59a/untitled-document-docs-title-untitled-document-created-date-191000801172643.jpg)
![Untitled-2 [about.gitlab.com] · Title: Untitled-2 Created Date: 20161031172420Z](https://static.documents.pub/doc/80x56/5aecde047f8b9a66258f266b/untitled-2-about-untitled-2-created-date-20161031172420z.jpg)
![Untitled Document [] · Title: Untitled Document Created Date: 191050201155549](https://static.documents.pub/doc/80x56/5fb77709eb118919875133dc/untitled-document-title-untitled-document-created-date-191050201155549.jpg)
![Untitled-6 []...Untitled-6 Author: turtle Subject: Untitled-6 Created Date: 3/5/2003 8:41:27 PM ...](https://static.documents.pub/doc/80x56/602bf9e5400fe51bf2284894/untitled-6-untitled-6-author-turtle-subject-untitled-6-created-date-352003.jpg)

![Untitled-2 [] · Title: Untitled-2 Created Date: 20100707113029Z](https://static.documents.pub/doc/80x56/5f8bab75572d02710a49e309/untitled-2-title-untitled-2-created-date-20100707113029z.jpg)
![Untitled-3 []Title: Untitled-3 Created Date: 20050330105657Z](https://static.documents.pub/doc/80x56/60347471aeac2f25265309a6/-untitled-3-title-untitled-3-created-date-20050330105657z.jpg)
![Untitled-1 [amadas.com] · Title: Untitled-1 Author: mikee Subject: Untitled-1 Created Date: 191000407142803](https://static.documents.pub/doc/80x56/5f91617c574a7c792f76c110/untitled-1-title-untitled-1-author-mikee-subject-untitled-1-created-date.jpg)
![Untitled-8 []...Title: Untitled-8 Created Date: 20181121103711Z](https://static.documents.pub/doc/80x56/5f535b10acd546526243f6d6/-untitled-8-title-untitled-8-created-date-20181121103711z.jpg)

![Untitled-4 [] · Title: Untitled-4 Author: Marie Created Date: 20171127161813Z](https://static.documents.pub/doc/80x56/5aecde857f8b9a90318ed907/untitled-4-untitled-4-author-marie-created-date-20171127161813z.jpg)
![Untitled-2 [] · Title: Untitled-2 Author: Utilizador Created Date: 20141128161636Z](https://static.documents.pub/doc/80x56/5f4bc13c9b9d42176052304d/untitled-2-title-untitled-2-author-utilizador-created-date-20141128161636z.jpg)
![Untitled-5 []Title Untitled-5 Created Date 20140415073242Z](https://static.documents.pub/doc/80x56/5ffd33023c19df0499476daa/-untitled-5-title-untitled-5-created-date-20140415073242z.jpg)

![Untitled-23 []...Title Untitled-23 Created Date 20170127153519Z](https://static.documents.pub/doc/80x56/6035343495251f126a22f79d/-untitled-23-title-untitled-23-created-date-20170127153519z.jpg)
