Unusual structures of TTTA repeats in icaC gene of Staphylococcus aureus Pei Guo, Sik Lok Lam ⇑ Department of Chemistry, The Chinese University of Hong Kong, Shatin, New Territories, Hong Kong article info Article history: Received 10 February 2015 Revised 15 April 2015 Accepted 17 April 2015 Available online 29 April 2015 Edited by Christian Griesinger Keywords: TTTA repeat expansion DNA structure Slipped strand mispairing Nuclear magnetic resonance Staphylococcus aureus abstract One and two TTTA repeat expansions have been found in the coding region of icaC gene of Staphylococcus aureus variants which influence the expression of IcaC protein and alter the phenotype. Yet, the mechanism of these small-size TTTA repeat expansions remains unclear. In this study, we performed high-resolution nuclear magnetic resonance spectroscopic studies on TTTA repeats. Our results show that a DNA sequence containing three TTTA repeats can fold into dumbbell structures with a 3 0 or 5 0 -overhang. Exchange of these dumbbells makes the sequence behave like a 2-nt TT mini-loop at 25 °C. The occurrence of these mini-loop and dumbbell structures in the nascent strand during DNA replication provides possible mechanistic pathways which account for one and two repeat expansions. Ó 2015 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. 1. Introduction Staphylococcus aureus is one of the top four pathogens related to central line-associated bloodstream infections [1]. The high fre- quency and death rate of S. aureus infections are alarming. Therefore, it is important to develop effective vaccines for S. aureus infections. However, the antibiotic resistance due to the formation of a protective biofilm is a mounting problem [2,3]. In S. aureus, the two major components of its biofilm matrix are b-1,6-linked poly- N-acetylglucosamine (PNAG) and polysaccharide intercellular adhesin (PIA) which is a partially N-deacetylated form of PNAG [4]. PIA/PNAG molecules are synthesized by the proteins encoded in the intercellular adhesin gene (icaADBC) locus [5]. IcaA and IcaD produce the short PIA/PNAG oligomers, which are linked into longer polymer chains and translocated to cell surfaces by IcaC. Once there, deacetylation modifications of PIA/PNAG molecules will be carried out by IcaB [6,7]. Recently, three TTTA repeats were found in the coding region of icaC gene of S. aureus strain MN8m and one additional repeat was found in a S. aureus variant called JB12 (Fig. 1) [8]. The additional repeat shifts the reading frame of icaC, resulting in a premature stop codon and thus a shorter protein. Therefore, the variant phenotype is altered from PIA/PNAG-overproducing to PIA/PNAG- negative, and the formation of biofilm is inhibited. Besides, two other S. aureus clinical isolates, namely, NRS264 and NRS63, also show the PIA/PNAG-negative phenotype and their icaC genes have been found to contain one and two additional TTTA repeats, respectively (Fig. 1) [8]. Based on the results of icaC mutation assay, TTTA repeat expan- sion was suggested to occur likely through slipped strand mispair- ing [8]. As the formation of non-B structures of DNA repeats has been proposed to induce slipped strand mispairing [9–12], we speculate that TTTA repeats may also form some unusual sec- ondary structures. As a result, we performed high-resolution nuclear magnetic resonance (NMR) spectroscopic investigations to elucidate the solution structures of TTTA repeats in this study. Our results reveal that a DNA sequence containing three TTTA repeats is capable of forming two dumbbell conformers. At 25 °C, fast exchange of the two conformers makes the sequence behave like a 2-nt TT mini-loop in the second repeat. These unusual struc- tures provide possible mechanistic pathways for the occurrence of TTTA repeat expansions. http://dx.doi.org/10.1016/j.febslet.2015.04.031 0014-5793/Ó 2015 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. Abbreviations: PIA, polysaccharide intercellular adhesin; PNAG, b-1,6-linked poly-N-acetylglucosamine; NMR, nuclear magnetic resonance; DSS, 2,2-dimethyl- 2-silapentane-5-sulfonic acid; PAGE, polyacrylamide gel electrophoresis; 1D, one- dimensional; 2D, two-dimensional; NOESY, nuclear Overhauser effect spec- troscopy; HMBC, heteronuclear multiple-bond correlation; TOCSY, total correlation spectroscopy; HSQC, heteronuclear single-quantum coherence Author contributions: P.G. performed all the experiments. Both authors contributed in the design and analysis of the study, and writing of the manuscript. ⇑ Corresponding author. Fax: +852 2603 5057. E-mail address: [email protected](S.L. Lam). FEBS Letters 589 (2015) 1296–1300 journal homepage: www.FEBSLetters.org
Transcript
FEBS Letters 589 (2015) 1296–1300
journal homepage: www.FEBSLetters .org
Unusual structures of TTTA repeats in icaC gene of Staphylococcus aureus
http://dx.doi.org/10.1016/j.febslet.2015.04.0310014-5793/� 2015 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
Author contributions: P.G. performed all the experiments. Both authors contributedin the design and analysis of the study, and writing of the manuscript.⇑ Corresponding author. Fax: +852 2603 5057.
Pei Guo, Sik Lok Lam ⇑Department of Chemistry, The Chinese University of Hong Kong, Shatin, New Territories, Hong Kong
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 February 2015Revised 15 April 2015Accepted 17 April 2015Available online 29 April 2015
Edited by Christian Griesinger
Keywords:TTTA repeat expansionDNA structureSlipped strand mispairingNuclear magnetic resonanceStaphylococcus aureus
One and two TTTA repeat expansions have been found in the coding region of icaC gene ofStaphylococcus aureus variants which influence the expression of IcaC protein and alter thephenotype. Yet, the mechanism of these small-size TTTA repeat expansions remains unclear. In thisstudy, we performed high-resolution nuclear magnetic resonance spectroscopic studies on TTTArepeats. Our results show that a DNA sequence containing three TTTA repeats can fold into dumbbellstructures with a 30 or 50-overhang. Exchange of these dumbbells makes the sequence behave like a2-nt TT mini-loop at 25 �C. The occurrence of these mini-loop and dumbbell structures in thenascent strand during DNA replication provides possible mechanistic pathways which account forone and two repeat expansions.� 2015 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
Staphylococcus aureus is one of the top four pathogens related tocentral line-associated bloodstream infections [1]. The high fre-quency and death rate of S. aureus infections are alarming.Therefore, it is important to develop effective vaccines for S. aureusinfections. However, the antibiotic resistance due to the formationof a protective biofilm is a mounting problem [2,3]. In S. aureus, thetwo major components of its biofilm matrix are b-1,6-linked poly-N-acetylglucosamine (PNAG) and polysaccharide intercellularadhesin (PIA) which is a partially N-deacetylated form of PNAG[4]. PIA/PNAG molecules are synthesized by the proteins encodedin the intercellular adhesin gene (icaADBC) locus [5]. IcaA andIcaD produce the short PIA/PNAG oligomers, which are linked intolonger polymer chains and translocated to cell surfaces by IcaC.Once there, deacetylation modifications of PIA/PNAG moleculeswill be carried out by IcaB [6,7].
Recently, three TTTA repeats were found in the coding region oficaC gene of S. aureus strain MN8m and one additional repeat wasfound in a S. aureus variant called JB12 (Fig. 1) [8]. The additionalrepeat shifts the reading frame of icaC, resulting in a prematurestop codon and thus a shorter protein. Therefore, the variantphenotype is altered from PIA/PNAG-overproducing to PIA/PNAG-negative, and the formation of biofilm is inhibited. Besides, twoother S. aureus clinical isolates, namely, NRS264 and NRS63, alsoshow the PIA/PNAG-negative phenotype and their icaC genes havebeen found to contain one and two additional TTTA repeats,respectively (Fig. 1) [8].
Based on the results of icaC mutation assay, TTTA repeat expan-sion was suggested to occur likely through slipped strand mispair-ing [8]. As the formation of non-B structures of DNA repeats hasbeen proposed to induce slipped strand mispairing [9–12], wespeculate that TTTA repeats may also form some unusual sec-ondary structures. As a result, we performed high-resolutionnuclear magnetic resonance (NMR) spectroscopic investigationsto elucidate the solution structures of TTTA repeats in this study.Our results reveal that a DNA sequence containing three TTTArepeats is capable of forming two dumbbell conformers. At 25 �C,fast exchange of the two conformers makes the sequence behavelike a 2-nt TT mini-loop in the second repeat. These unusual struc-tures provide possible mechanistic pathways for the occurrence ofTTTA repeat expansions.
Fig. 1. Three TTTA repeats (underlined) have been identified in the coding region of icaC gene of S. aureus strain MN8m [8], showing PIA/PNAG-overproducing phenotype. Oneand two additional TTTA repeats (in red) were found in S. aureus isolates JB12, NRS264 and NRS63, showing PIA/PNAG-negative phenotype.
As the mutation site of icaC gene originally contains three TTTArepeats (Fig. 1) [8], firstly we prepared a sequence containing threeTTTA repeats and named it as ‘‘(TTTA)3’’. To facilitate the detailedstructural study of (TTTA)3, we also prepared a shorter sequencecontaining two TTTA repeats and named it as ‘‘(TTTA)2’’. Both(TTTA)3 and (TTTA)2 were synthesized using an AppliedBiosystems model 394 DNA synthesizer. They were purified bydenaturing polyacrylamide gel electrophoresis (PAGE) and diethy-laminoethyl Sephacel anion exchange column chromatography,and finally desalted using Amicon� Ultra-4 centrifugal filteringdevices. NMR samples were prepared by dissolving 0.5 lmol puri-fied DNA into 500 lL buffer solutions containing 10 mM sodiumphosphate (pH 7.0), and 0.1 mM 2,2-dimethyl-2-silapentane-5-sul-fonic acid (DSS).
2.2. NMR study
All NMR experiments were performed using two Bruker AV-500spectrometers operating at 500.30 and 500.13 MHz, respectively.For studying the labile protons, the samples were prepared in a90% H2O/10% D2O buffer solution. One-dimensional (1D) iminoand two-dimensional (2D) nuclear Overhauser effect spectroscopy(NOESY) spectra were acquired using the water suppression byexcitation sculpting [13]. For studying the non-labile protons, thesolvent was exchanged with a 99.96% D2O buffer solution and a2-s presaturation pulse was used to suppress the residual HDO sig-nal. 2D NOESY spectra were acquired with a data size of4096 � 512 and a mixing time of 300 ms unless otherwise speci-fied. The acquired data sets were zero-filled to give 4096 � 4096spectra with a cosine window function applied to both dimensions.For adenine H2 protons, the assignments were confirmed by1H-13C heteronuclear multiple-bond correlation (HMBC) experi-ments [14] with an evolution period of 65 ms for the long-rangecouplings. The 13C spectral width was set to 60 ppm and the carrierfrequency was centered at 140 ppm. Backbone 31P signals wereassigned using 2D total correlation spectroscopy (TOCSY) with amixing time of 75 ms and 1H-31P heteronuclear single-quantumcoherence (HSQC) experiments. The 31P spectral width was set to6 ppm and a data size of 4096 � 200 was collected. 13C and 31Pchemical shifts were indirectly referenced to DSS using the derivednucleus-specific ratio of 0.251449530 and 0.404808636, respec-tively [15].
To determine the thermal stability of (TTTA)2, variable temper-ature 1D 1H experiments were performed from 0 to 75 �C at a step
size of 2.5 �C. Thermodynamic parameters were determined by fit-ting the melting curves using MELTWIN version 3.5 software [16].
2.3. Gel mobility assay
25% polyacrylamide native gels were prepared to investigatethe oligomeric state of (TTTA)2 and (TTTA)3. These samples wereprepared in the same buffer solutions as for NMR studies. For thereference lane, a complementary 10-base pair duplex was preparedby hybridizing 50-CTGGCTGCGC-30 and 50-GCGCAGCCAG-30 in abuffer solution containing 10 mM sodium phosphate (pH 7.0)and 150 mM sodium chloride. PAGE was conducted at ambienttemperature and all DNA samples were kept at a concentrationof 0.4 mM. DNA bands were visualized by staining the gels withstains-all solution.
3. Results and discussion
In this study, we performed high-resolution 1H and 31P NMRspectroscopic investigations to reveal the solution structures of(TTTA)3 and (TTTA)2. Native gel analysis shows that both (TTTA)3
and (TTTA)2 behave as a monomeric species in the buffer solutions(Supporting Information, S1). Sequential assignments were madefrom the H6/H8-H10 fingerprint regions in 2D NOESY spectra inD2O and/or H2O using standard methods [17,18]. As the sequentialT3 H10-A4 H8 and T7 H10-A8 H8 NOEs were not observed in the 2DNOESY spectrum of (TTTA)2 at a mixing time of 300 ms, in order toassign the H8 signals of A4 and A8, the NOESY experiment wasrepeated at a longer mixing time of 800 ms and these sequentialNOEs were observed (Supporting Information, S2). For the assign-ments of adenine H2, they were confirmed by 1H-13C HMBC exper-iments [14] (Supporting Information, S3). Based on the H30
assignment results from TOCSY, the backbone 31P signals wereassigned using 1H-31P HSQC spectra (Supporting Information, S4and S5).
3.1. (TTTA)3 appears to form a 2-nt TT mini-loop in the second repeat
At 25 �C, (TTTA)3 appears to predominantly form a 2-nt TT mini-loop closed by a TA base pair in the second repeat (Fig. 2A). Thisloop was similar to the one found in the DNA hairpin ofd(ATCCTA TTTA TAGGAT) [19] as supported by the characteristic31P chemical shifts of downfield T6 (�3.77 ppm) and upfield T7(�4.62 ppm) [12,20] (Fig. 2A). These unusual shifts are due to thecharacteristic backbone features of the type II loop [12,20], inwhich T6 is located in the minor groove, positioning itself approx-imately perpendicular to the loop closing T5–A8, while T7 stacksover the base plane of T5–A8 (Fig. 2A). The stacking of T7 over
Fig. 2. (A) At 25 �C, the second repeat in (TTTA)3 predominantly forms a TT loop as supported by the unusually downfield T6 and upfield T7 31P signals. The TT loop adopts atype II loop in which T7 stacks over T5–A8 base pair and T6 is positioned in the minor groove. (B) The NOESY fingerprint region shows the unusually upfield shifted T7 H10 .Sequential NOEs of T6 H10-T7 H6 and T7 H10-A8 H8 were not observed (red dotted circles). (C) TA Hoogsteen base pair with the adenine in the syn orientation shows a shorterC10-C10 distance than that of Watson–Crick base pair [18], providing a better base plane for the stacked base. (D) The similar NOE intensities of A8 H8-H10 and A8 H8-H20/H20
support the syn orientation of A8 in T5–A8 Hoogsteen base pair. (E) The variable temperature aromatic and methyl proton regions of (TTTA)3. The aromatic protonassignments are shown in the 1D projection spectrum in (B). (F) The two proposed dumbbell structures formed by (TTTA)3 at lower temperatures.
T5–A8 also made the chemical shift of T7 H10 unusually upfield(Fig. 2B). Owing to the sharp turn in type II loop, the sequentialNOEs of T6 H10-T7 H6 and T7 H10-A8 H8 were not observed(Fig. 2B).
For the TT loop in the DNA hairpin [19], it was closed by a TAHoogsteen base pair of which the adenine adopts a syn orientation(Fig. 2C). In addition to the two hydrogen bonds, the base stackedon top of the closing Hoogsteen base pair also contributes to thestability of the mini-loop [20]. Owing to the shorter interstrandC10-C10 distance, the stacking interactions are more effective thanthose involving a closing Watson–Crick base pair [19,21]. In(TTTA)3, the base orientation of A8 also possesses the syn featureas evidenced by the similar NOE intensities of H8-H10 and H8-H20/H200 (Fig. 2D), suggesting T5–A8 also forms a Hoogsteen basepair. In the variable temperature imino proton spectra, signalswere observed only at �11 ppm at lower temperatures(Supporting Information, S6a), which are consistent with thechemical shifts of those belonging to TT loops [9,19]. No T5 iminosignal was observed in the range of 12–13 ppm [19,22], revealingthe T5–A8 Hoogsteen base pair was not as stable as the one withneighboring Watson–Crick base pairs in the stem region [19].
For the two ends of the mini-loop in (TTTA)3, no imino protonsignal and unusual NMR features were observed for T1 to A4 inthe first repeat and T9 to A12 in the third repeat at 25 �C. In
addition to A4 and A12 adopting the anti orientation as supportedby the weaker H8-H10 NOE than H8-H20/H200 NOEs (Fig. 2D), all 31Pchemical shifts of the residues in the first and third repeats (�3.91to �4.19 ppm) were found to fall within the range for random coilDNAs (�3.80 to �4.20 ppm) [23], suggesting there is no secondarystructure at both ends of the mini-loop. Interestingly, instead ofgetting sharper 31P signals of T6 and T7 due to a further stabiliza-tion of the TT loop at lower temperatures, these signals were sur-prisingly broadened (Fig. 2A). Moreover, most of the aromatic andmethyl proton signals were also seriously broadened at tempera-tures below 10 �C (Fig. 2E). Therefore, we suspected that the firstand third repeats also possess the ability to fold into mini-loops,forming exchangeable dumbbell structures with a 30 and 50-over-hang, respectively (Fig. 2F). In order to determine if the formationof such small dumbbell structures is feasible, we investigated thesolution structures of (TTTA)2.
3.2. (TTTA)2 forms a mini dumbbell structure composed of two TT loops
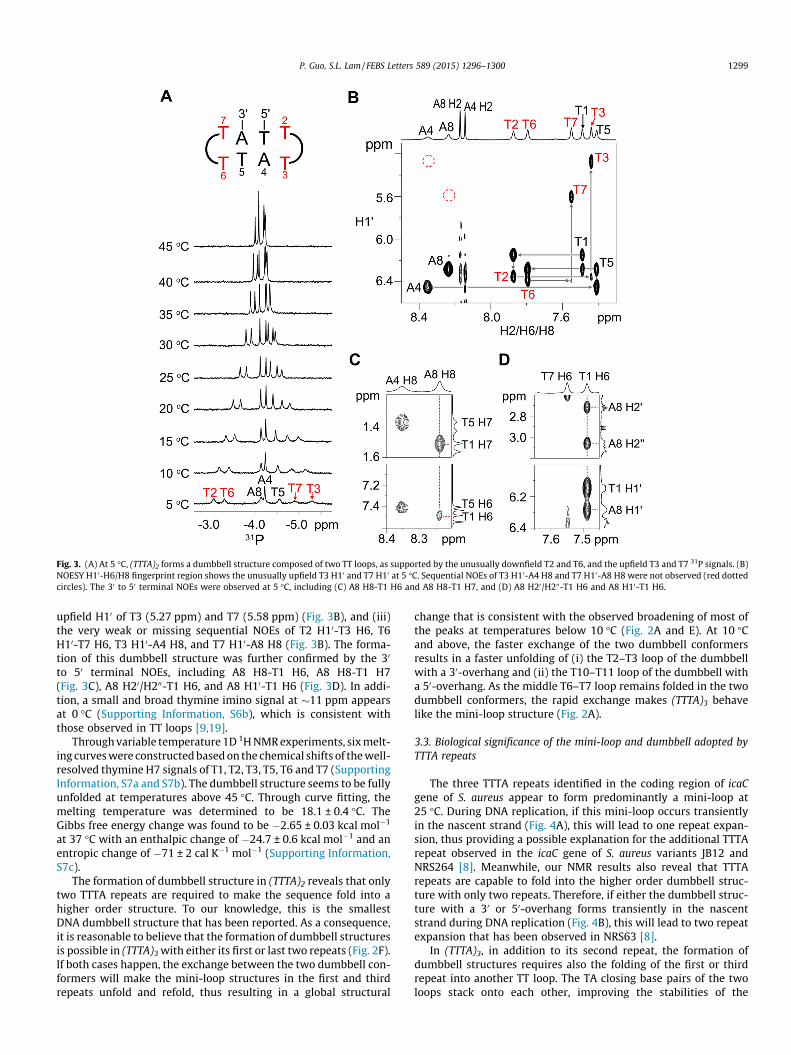
The NMR spectral features of (TTTA)2 support it adopts a dumb-bell structure composed of two TT loops at 5 �C (Fig. 3A). Theseinclude (i) the unusually downfield 31P signals of T2 (�3.06 ppm)and T6 (�3.30 ppm) and the unusually upfield 31P signals of T3(�5.30 ppm) and T7 (�4.91 ppm) (Fig. 3A), (ii) the unusually
Fig. 3. (A) At 5 �C, (TTTA)2 forms a dumbbell structure composed of two TT loops, as supported by the unusually downfield T2 and T6, and the upfield T3 and T7 31P signals. (B)NOESY H10-H6/H8 fingerprint region shows the unusually upfield T3 H10 and T7 H10 at 5 �C. Sequential NOEs of T3 H10-A4 H8 and T7 H10-A8 H8 were not observed (red dottedcircles). The 30 to 50 terminal NOEs were observed at 5 �C, including (C) A8 H8-T1 H6 and A8 H8-T1 H7, and (D) A8 H20/H200-T1 H6 and A8 H10-T1 H6.
upfield H10 of T3 (5.27 ppm) and T7 (5.58 ppm) (Fig. 3B), and (iii)the very weak or missing sequential NOEs of T2 H10-T3 H6, T6H10-T7 H6, T3 H10-A4 H8, and T7 H10-A8 H8 (Fig. 3B). The forma-tion of this dumbbell structure was further confirmed by the 30
to 50 terminal NOEs, including A8 H8-T1 H6, A8 H8-T1 H7(Fig. 3C), A8 H20/H200-T1 H6, and A8 H10-T1 H6 (Fig. 3D). In addi-tion, a small and broad thymine imino signal at �11 ppm appearsat 0 �C (Supporting Information, S6b), which is consistent withthose observed in TT loops [9,19].
Through variable temperature 1D 1H NMR experiments, six melt-ing curves were constructed based on the chemical shifts of the well-resolved thymine H7 signals of T1, T2, T3, T5, T6 and T7 (SupportingInformation, S7a and S7b). The dumbbell structure seems to be fullyunfolded at temperatures above 45 �C. Through curve fitting, themelting temperature was determined to be 18.1 ± 0.4 �C. TheGibbs free energy change was found to be �2.65 ± 0.03 kcal mol�1
at 37 �C with an enthalpic change of �24.7 ± 0.6 kcal mol�1 and anentropic change of �71 ± 2 cal K�1 mol�1 (Supporting Information,S7c).
The formation of dumbbell structure in (TTTA)2 reveals that onlytwo TTTA repeats are required to make the sequence fold into ahigher order structure. To our knowledge, this is the smallestDNA dumbbell structure that has been reported. As a consequence,it is reasonable to believe that the formation of dumbbell structuresis possible in (TTTA)3 with either its first or last two repeats (Fig. 2F).If both cases happen, the exchange between the two dumbbell con-formers will make the mini-loop structures in the first and thirdrepeats unfold and refold, thus resulting in a global structural
change that is consistent with the observed broadening of most ofthe peaks at temperatures below 10 �C (Fig. 2A and E). At 10 �Cand above, the faster exchange of the two dumbbell conformersresults in a faster unfolding of (i) the T2–T3 loop of the dumbbellwith a 30-overhang and (ii) the T10–T11 loop of the dumbbell witha 50-overhang. As the middle T6–T7 loop remains folded in the twodumbbell conformers, the rapid exchange makes (TTTA)3 behavelike the mini-loop structure (Fig. 2A).
3.3. Biological significance of the mini-loop and dumbbell adopted byTTTA repeats
The three TTTA repeats identified in the coding region of icaCgene of S. aureus appear to form predominantly a mini-loop at25 �C. During DNA replication, if this mini-loop occurs transientlyin the nascent strand (Fig. 4A), this will lead to one repeat expan-sion, thus providing a possible explanation for the additional TTTArepeat observed in the icaC gene of S. aureus variants JB12 andNRS264 [8]. Meanwhile, our NMR results also reveal that TTTArepeats are capable to fold into the higher order dumbbell struc-ture with only two repeats. Therefore, if either the dumbbell struc-ture with a 30 or 50-overhang forms transiently in the nascentstrand during DNA replication (Fig. 4B), this will lead to two repeatexpansion that has been observed in NRS63 [8].
In (TTTA)3, in addition to its second repeat, the formation ofdumbbell structures requires also the folding of the first or thirdrepeat into another TT loop. The TA closing base pairs of the twoloops stack onto each other, improving the stabilities of the
Fig. 4. A brief scheme shows the possible pathways for TTTA repeat expansions.Formation of (A) mini-loop structure and (B) dumbbell structures on the nascentstrand can lead to one and two TTTA repeat expansion, respectively.
unusual structure and thereby enhancing the propensity of TTTArepeats to slip during DNA replication. Similar behaviors have alsobeen observed in DNA sequences containing CCTG repeats [12].The formation of dumbbell structure with two CT loops has beensuggested to stabilize the slipped hairpin structure, therebyincreasing the unusual structure formation propensity of CCTGrepeats.
At present, expansions of short tandem repeats have been foundto associate with at least 15 human genetic diseases such as CAGrepeats in Huntington’s disease, CTG repeats in myotonic dystro-phy type 1 and CCTG repeats in myotonic dystrophy type 2 [24].The formation of non-B structures can lead to strand slippageand thereby repeat expansion during DNA replication [25]. In addi-tion to the non-B DNA hairpins [9], triplexes [26], and tetraplexes[11] formed by short tandem repeats, the dumbbell structuresobserved in the present study also serve as a structural intermedi-ate resulting from strand slippage, providing insights into theunderlying molecular mechanisms of repeat expansion diseases.
Acknowledgments
We would like to thank Professor H.N.C. Wong for his continu-ing support on the research activities of our group. The workdescribed in this paper was partially supported by a GeneralResearch Fund (Project No. CUHK14302114) and SpecialEquipment Grant (Project No. SEG/CUHK09) from the ResearchGrants Council of the Hong Kong Special Administrative Region,and a direct Grant from CUHK Research Committee Funding(Project No. 4053053).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.febslet.2015.04.031.
References
[1] Popoola, V.O. and Milstone, A.M. (2014) Decolonization to preventStaphylococcus aureus transmission and infections in the neonatal intensivecare unit. J. Perinatol. 34, 805–810.
[2] van Hal, S.J. and Fowler Jr., V.G. (2013) Is it time to replace vancomycin in thetreatment of methicillin-resistant Staphylococcus aureus infections? Clin.Infect. Dis. 56, 1779–1788.
[3] Hoiby, N., Bjarnsholt, T., Givskov, M., Molin, S. and Ciofu, O. (2010) Antibioticresistance of bacterial biofilms. Int. J. Antimicrob. Agents 35, 322–332.
[4] Rohde, H., Frankenberger, S., Zahringer, U. and Mack, D. (2010) Structure,function and contribution of polysaccharide intercellular adhesin (PIA) toStaphylococcus epidermidis biofilm formation and pathogenesis of biomaterial-associated infections. Eur. J. Cell Biol. 89, 103–111.
[5] Heilmann, C., Schweitzer, O., Gerke, C., Vanittanakom, N., Mack, D. and Gotz, F.(1996) Molecular basis of intercellular adhesion in the biofilm-formingStaphylococcus epidermidis. Mol. Microbiol. 20, 1083–1091.
[6] Gerke, C., Kraft, A., Sussmuth, R., Schweitzer, O. and Gotz, F. (1998)Characterization of the N-acetylglucosaminyltransferase activity involved inthe biosynthesis of the Staphylococcus epidermidis polysaccharide intercellularadhesin. J. Biol. Chem. 273, 18586–18593.
[7] Vuong, C., Kocianova, S., Voyich, J.M., Yao, Y., Fischer, E.R., DeLeo, F.R. and Otto,M. (2004) A crucial role for exopolysaccharide modification in bacterial biofilmformation, immune evasion, and virulence. J. Biol. Chem. 279, 54881–54886.
[8] Brooks, J.L. and Jefferson, K.K. (2014) Phase variation of poly-N-acetylglucosamine expression in Staphylococcus aureus. PLoS Pathog. 10,e1004292.
[9] Chi, L.M. and Lam, S.L. (2005) Structural roles of CTG repeats in slippageexpansion during DNA replication. Nucleic Acids Res. 33, 1604–1617.
[10] Gacy, A.M. et al. (1998) GAA instability in Friedreich’s Ataxia shares acommon, DNA-directed and intraallelic mechanism with other trinucleotidediseases. Mol. Cell 1, 583–593.
[11] Fojtik, P. and Vorlickova, M. (2001) The fragile X chromosome (GCC) repeatfolds into a DNA tetraplex at neutral pH. Nucleic Acids Res. 29, 4684–4690.
[12] Lam, S.L., Wu, F., Yang, H. and Chi, L.M. (2011) The origin of genetic instabilityin CCTG repeats. Nucleic Acids Res. 39, 6260–6268.
[13] Stott, K., Stonehouse, J., Keeler, J., Hwang, T.L. and Shaka, A.J. (1995) Excitationsculpting in high-resolution nuclear magnetic resonance spectroscopy:application to selective NOE experiments. J. Am. Chem. Soc. 117, 4199–4200.
[14] van Dongen, M.J., Wijmenga, S.S., Eritja, R., Azorin, F. and Hilbers, C.W. (1996)Through-bond correlation of adenine H2 and H8 protons in unlabeled DNAfragments by HMBC spectroscopy. J. Biomol. NMR 8, 207–212.
[15] Markley, J.L., Bax, A., Arata, Y., Hilbers, C.W., Kaptein, R., Sykes, B.D., Wright,P.E. and Wuthrich, K. (1998) Recommendations for the presentation of NMRstructures of proteins and nucleic acids – (IUPAC Recommendations 1998).Pure Appl. Chem. 70, 117–142.
[16] McDowell, J.A. and Turner, D.H. (1996) Investigation of the structural basis forthermodynamic stabilities of tandem GU mismatches: solution structure of(rGAGGUCUC)2 by two-dimensional NMR and simulated annealing.Biochemistry 35, 14077–14089.
[17] Wijmenga, S.S. and van Buuren, B.N.M. (1998) The use of NMR methods forconformational studies of nucleic acids. Prog. Nucl. Magn. Reson. Spectrosc.32, 287–387.
[18] Feigon, J., Wright, J.M., Leupin, W., Denny, W.A. and Kearns, D.R. (1982) Use oftwo-dimensional NMR in the study of a double-stranded DNA decamer. J. Am.Chem. Soc. 104, 5540–5541.
[19] Blommers, M.J., van de Ven, F.J., van der Marel, G.A., van Boom, J.H. andHilbers, C.W. (1991) The three-dimensional structure of a DNA hairpin insolution two-dimensional NMR studies and structural analysis ofd(ATCCTATTTATAGGAT). Eur. J. Biochem. 201, 33–51.
[20] Ippel, H.H., van den Elst, H., van der Marel, G.A., van Boom, J.H. and Altona, C.(1998) Structural similarities and differences between H1- and H2-familyDNA minihairpin loops: NMR studies of octameric minihairpins. Biopolymers46, 375–393.
[21] van Dongen, M.J., Wijmenga, S.S., van der Marel, G.A., van Boom, J.H. andHilbers, C.W. (1996) The transition from a neutral-pH double helix to a low-pH triple helix induces a conformational switch in the CCCG tetraloop closinga Watson-Crick stem. J. Mol. Biol. 263, 715–729.
[22] Yang, H., Zhan, Y., Fenn, D., Chi, L.M. and Lam, S.L. (2008) Effect of 1-methyladenine on double-helical DNA structures. FEBS Lett. 582, 1629–1633.
[23] Ho, C.N. and Lam, S.L. (2004) Random coil phosphorus chemical shift ofdeoxyribonucleic acids. J. Magn. Reson. 171, 193–200.
[24] Yang, H.D., Ho, D.H. and Seol, W. (2012) Unstable repeat expansion inneurodegenerative dementias: mechanisms of disease. Dement.Neurocognitive Disord. 11, 1–12.
[25] Mirkin, S.M. (2007) Expandable DNA repeats and human disease. Nature 447,932–940.
[26] Gacy, A.M. et al. (1998) GAA instability in Friedreich’s Ataxia shares acommon, DNA-directed and intraallelic mechanism with other trinucleotidediseases. Mol. Cell 1, 583–593.