Upper Paleocene-lower Eoceneradiolarian biostratigraphy of the San Francisco de Paula section, western Cuba: Regional and global comparisons Annika Sanfilippol and Donna M. Hull2 with a contribution by Emilio Flores Albin3 1 Scripps Institution of Oceanography, University of California at San Diego, La Jolla, CA 92093-0244 email: [email protected]2 The University of Texas at Dallas, Programs in Geosciences, P.O. Box 830688, Richardson, TX 75083-0688 email: [email protected]3 Centro de Investigacciones del Petroleo, CUPET ABSTRACT: In response to a growing needto definethe Paleocene/Eocene boundary and record the eventswhich immediately pre- cededand followed it within a 5 my span, the San Francisco de Paula section of western Cuba was evaluated forits potential as a bound- arystratotype. Radiolarians, reported in previous studies by Cuban geologists as a major component of the faunal assemblages in this section, havebeenrecollected and reanalyzed to determine their stratigraphic utility for recognition of thePaleocene/Eocene boundary in Cuba.On the basis of this and otherrecentmicrofossil studiesof the San Francisco de Paulasection,it is now knownthat the Paleocene/Eocene boundary in thissuccession is within anunconformity. Its placement herein is based on the highest occurrence of the planktonic foraminifera Morozovella velascoensis (Fernandez-Rodrguez et al. 1999, this volume). Radiolarians are present throughout the SanFrancisco de Paula succession withthe exception of a barren interval in themiddle of the section and two thin barren intervals nearthe top and the base. The most diverseradiolarian assemblages are from the shaley mudstones, whichcontain poorly preserved radiolarians that range from abundant to very rare. Other lithologies contain sparse, poorly preserved assemblages that appear strongly affected by dissolution. Thirty-five lowestand highest occurrences are noted within the sec- tion; however, because of poor preservation, several of thesedatums are higher or lower than expected when compared withtheknown global ranges of thesespecies. Theradiolarians from this study of theSan Francisco de Paula section canbe assigned to the stratigraphic interval from the upperPaleoceneBekomacampechensis to the lower Eocene Buryellaclinata Zones. Species typical of the B. campechensis Zone includeBekoma campechensis, B. demissa,Bekoma spp., Buryella pentadica, B. tetradica, B. foremanaeand Lamptonium pennatum. Faunas of the Bekoma bidartensis Zone contain Buryella tetradica, Giraffospyris lata, Phormocyrtis turgida, Podocyrtis (Podocyrtis) papalis, Pterocodon tenellus, Theocorys phyzella, Theocotyle nigriniae, Theocotylissa alpha, T auctor and Thyrsocyrtis (Thyrsocyrtis) hirsuta. The radiolarian assemblage representing the lower part of the Buryella clinata Zone is similar to thatof the Bekoma bidartensis Zone, andalso includes the marker speciesBuryella clinata. Assignment of the upper part of the San Francisco de Paula section to theBuryella clinata Zoneis tentative, because of poor preservation. Inthe San Francisco de Paula section, the preciselocation of the Paleocene/Eocene boundary cannot be determined on the basis of radiolarians. However, taken together, radiolarian and calcareous nannofossil evidence indicate that the Paleocene/Eocene boundary is within an unconformity between sam- ples SFP-22 andSFP-21 at the highest local occurrence of Morozovella velascoensis, equivalent to the top of planktonic foraminiferal Zone P5. In addition to tabulating the stratigraphic ranges of radiolarian species in western Cuba, this study presents a preliminary correla- tionbetween the lower Paleogene standard radiolarian low latitude zonation andother zonal schemes presented for the Caribbean re- gion andmid to high latitudes. Although the San Francisco de Paulasectiondoes possess some of the characteristics desirable in a stratotype for the Paleocene/Eocene boundary, its usefulness in terms of biostratigraphy is limited. INTRODUCTION Programme Project #308 was initiated to study Paleocene/Eo- The Paleocene/Eocene boundary interval is an interesting pe- cene boundary sections worldwide, including potential sections riod in Earth history, characterized by warm high latitudes previously documented in western Cuba (e.g., Bronnimann and (Stott and Kennett 1990; Kennett and Stott 1991) and changes Rigassi 1963). in oceanic circulation associated with deep water production at low latitudes (Miller et al. 1987; Kennett and Stott 1991; In contrast to calcareous microfossil groups for which a consid- Thomas 1990a, 1990b, 1993). Consequently, definition of the erable amount of lower Paleogene biostratigraphicinformation Paleocene/Eocene boundary takes on increasing significance as is available from land-based sequences in the Caribbean region it becomes necessary to place these climatic and as well as from Deep Sea Drilling Project (DSDP)/Ocean paleoceanographic events in their correct chronological succes- Drilling Program (ODP) cores, most biostratigraphic informa- sion. To meet this need. International Geological Correlation tion on Cenozoic radiolarians in the Caribbean region is from micropaleontology, volume 45, supplement 2, pp. 57-82, text-figures 1-3, plates 1-2, table 1, 1999 57
Transcript
Upper Paleocene-lower Eocene radiolarian biostratigraphy of the San Francisco de Paula section, western Cuba:
Regional and global comparisons
Annika Sanfilippol and Donna M. Hull2 with a contribution by Emilio Flores Albin3
1 Scripps Institution of Oceanography, University of California at San Diego, La Jolla, CA 92093-0244 email: [email protected]
2 The University of Texas at Dallas, Programs in Geosciences, P.O. Box 830688, Richardson, TX 75083-0688 email: [email protected]
3 Centro de Investigacciones del Petroleo, CUPET
ABSTRACT: In response to a growing need to define the Paleocene/Eocene boundary and record the events which immediately pre- ceded and followed it within a 5 my span, the San Francisco de Paula section of western Cuba was evaluated for its potential as a bound- ary stratotype. Radiolarians, reported in previous studies by Cuban geologists as a major component of the faunal assemblages in this section, have been recollected and reanalyzed to determine their stratigraphic utility for recognition of the Paleocene/Eocene boundary in Cuba. On the basis of this and other recent microfossil studies of the San Francisco de Paula section, it is now known that the Paleocene/Eocene boundary in this succession is within an unconformity. Its placement herein is based on the highest occurrence of the planktonic foraminifera Morozovella velascoensis (Fernandez-Rodrguez et al. 1999, this volume).
Radiolarians are present throughout the San Francisco de Paula succession with the exception of a barren interval in the middle of the section and two thin barren intervals near the top and the base. The most diverse radiolarian assemblages are from the shaley mudstones, which contain poorly preserved radiolarians that range from abundant to very rare. Other lithologies contain sparse, poorly preserved assemblages that appear strongly affected by dissolution. Thirty-five lowest and highest occurrences are noted within the sec- tion; however, because of poor preservation, several of these datums are higher or lower than expected when compared with the known global ranges of these species. The radiolarians from this study of the San Francisco de Paula section can be assigned to the stratigraphic interval from the upper Paleocene Bekoma campechensis to the lower Eocene Buryella clinata Zones. Species typical of the B. campechensis Zone include Bekoma campechensis, B. demissa, Bekoma spp., Buryella pentadica, B. tetradica, B. foremanae and Lamptonium pennatum. Faunas of the Bekoma bidartensis Zone contain Buryella tetradica, Giraffospyris lata, Phormocyrtis turgida, Podocyrtis (Podocyrtis) papalis, Pterocodon tenellus, Theocorys phyzella, Theocotyle nigriniae, Theocotylissa alpha, T auctor and Thyrsocyrtis (Thyrsocyrtis) hirsuta. The radiolarian assemblage representing the lower part of the Buryella clinata Zone is similar to that of the Bekoma bidartensis Zone, and also includes the marker species Buryella clinata. Assignment of the upper part of the San Francisco de Paula section to the Buryella clinata Zone is tentative, because of poor preservation. In the San Francisco de Paula section, the precise location of the Paleocene/Eocene boundary cannot be determined on the basis of radiolarians. However, taken together, radiolarian and calcareous nannofossil evidence indicate that the Paleocene/Eocene boundary is within an unconformity between sam- ples SFP-22 and SFP-21 at the highest local occurrence of Morozovella velascoensis, equivalent to the top of planktonic foraminiferal Zone P5.
In addition to tabulating the stratigraphic ranges of radiolarian species in western Cuba, this study presents a preliminary correla- tion between the lower Paleogene standard radiolarian low latitude zonation and other zonal schemes presented for the Caribbean re- gion and mid to high latitudes. Although the San Francisco de Paula section does possess some of the characteristics desirable in a stratotype for the Paleocene/Eocene boundary, its usefulness in terms of biostratigraphy is limited.
INTRODUCTION Programme Project #308 was initiated to study Paleocene/Eo- The Paleocene/Eocene boundary interval is an interesting pe- cene boundary sections worldwide, including potential sections riod in Earth history, characterized by warm high latitudes previously documented in western Cuba (e.g., Bronnimann and (Stott and Kennett 1990; Kennett and Stott 1991) and changes Rigassi 1963). in oceanic circulation associated with deep water production at low latitudes (Miller et al. 1987; Kennett and Stott 1991; In contrast to calcareous microfossil groups for which a consid- Thomas 1990a, 1990b, 1993). Consequently, definition of the erable amount of lower Paleogene biostratigraphic information Paleocene/Eocene boundary takes on increasing significance as is available from land-based sequences in the Caribbean region it becomes necessary to place these climatic and as well as from Deep Sea Drilling Project (DSDP)/Ocean paleoceanographic events in their correct chronological succes- Drilling Program (ODP) cores, most biostratigraphic informa- sion. To meet this need. International Geological Correlation tion on Cenozoic radiolarians in the Caribbean region is from
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
deep-sea drilling cores alone. Radiolarians from Cretaceous and lower Paleogene formations in western Cuba have been re- ported by Flores Albin (1983) and Flores Albin and Femaindez-Rodriguez (1985), and a small amount of additional information on Cenozoic radiolarians from Cuba, Barbados and Trinidad is summarized in Riedel and Sanfilippo (1971), and Sanfilippo and Riedel (1976). Maurrasse (1973, 1976, 1979) re- ported mainly on the paleoecologic and paleobiogeographic im- plications of radiolarian facies in Caribbean mid-Paleogene deep-sea sediments. DSDP Leg 4 collected useful Caribbean middle Eocene sequences with radiolarians at Site 29 (Riedel and Sanfilippo 1970). Leg 10 in the Gulf of Mexico sampled a similar sequence (Foreman 1973; Sanfilippo and Riedel 1973) and Leg 15 provided Caribbean equivalents of the sediment se- quences in the Gulf of Mexico (Riedel and Sanfilippo 1973) which permitted correlation with calcareous microfossils. Sub- sequent DSDP/ODP Legs (76-78, 96, 100-102 and 110) in the Caribbean region recovered only younger than middle Eocene radiolarian-bearing sediments. Although more numerous Paleocene and lower Eocene samples have been obtained from different parts of the world ocean, the recovery is still intermit- tent and radiolarian preservation commonly not adequate for detailed biostratigraphic work (Sanfilippo and Nigrini 1998a).
For these reasons it was anticipated that the discovery of a new land-based locality, south of the village of San Francisco de Paula in western Cuba (text-figure 1), would contain a suitable Paleocene/Eocene boundary section that would resolve at least some of the stratigraphic problems around the boundary and at the same time allow correlation between the calcareous and sili- ceous microfossil groups with the results from paleomagnetic and isotope investigations (M.-P. Aubry, person. communs., 1996, 1997; Femaindez-Rodriguez et al. 1999, this volume).
LOCATION AND LITHOSTRATIGRAPHY
The San Francisco de Paula section is located in western Cuba near the city of Habana (text-figure 1). It is 56m thick and con- sists of the Apolo Formation (Paleocene and lowermost Eocene) and the lower part of the overlying Capdevila Forma- tion (also known locally as the Alcava Formation; lower Eocene, for further explanation see FemaJndez-Rodriguez et al. 1999, this volume). The lower 40m of the section (text-figure 2), which includes the Paleocene/Eocene boundary interval, is composed of marly mudstones and soft shales which are weath- ered and, in places, largely covered by vegetation. The base of the section is more massively bedded with coarser-grained rocks and conglomeratic limestones. Forty-three meters above the base of the San Francisco de Paula section, the rock se- quence is marked by a distinctive lithologic boundary. The rocks above this boundary are much coarser-grained and are well indurated with distinctive graded bedding, suggesting storm or turbidite deposition. White marly clasts appear in lin- ear bands just above the lithologic boundary.
Fossils present in the San Francisco de Paula succession include planktonic and benthic foraminifera, large foraminifera, radio- larians, calcareous nannofossils, ostracodes, bryozoans, calcar- eous algae and mollusk fragments (Flores Albin and Fernandez-Rodriguez 1985; Fernmandez-Rodrfguez et al. 1999, this volume). Unfortunately, all fossil groups show varying de- grees of preservation, and none occur continuously throughout the sequence. Fernmandez-Rodrfguez et al. (1999, this volume), in their analysis of the planktonic foraminifera, suggest the presence of four unconformities in the sequence. Recognition of these unconformities is based in part on abrupt changes in
abundance, diversity, and preservation of the foraminiferal fau- nas. They relate these unconformities to the 58 to 54 Ma time span of active local tectonics which has been noted by other au- thors (Bralower and Ituralde-Vinent 1997; Lewis and Draper 1990).
METHODS AND MATERIALS
Initial discovery and collection of material from the San Fran- cisco de Paula section was made by E. Flores Albin, G. Femrnandez-Rodriguez and other Cuban colleagues shortly after this section was cut along a new road. Consequently, the origi- nal samples are relatively unweathered. The Cuban geologists measured the section, collected samples, described lithofacies, and analyzed the material principally for foraminifera and radiolarians (Flores Albin, Appendix 1; Femrnandez-Rodriguez et al. 1999, this volume).
In 1993, a group of scientists from the U.S., including A. Sanfilippo and D. Hull, joined the Cuban geologists to revisit the San Francisco de Paula section as well as other potential Paleocene/Eocene boundary sections in western Cuba on behalf of IGCP Project #308. The section was re-measured, described, and 54 samples were collected for studies of planktonic and benthonic foraminifera, radiolarians, and calcareous nanno- fossils; portions of these same samples were also set aside for carbon/oxygen isotope determinations.
The selection of procedures used to measure and sample the section during the 1993 excursion was based on prior knowl- edge of the Cuban scientists as to the approximate position of the Paleocene/Eocene boundary within the succession. Below a marked point within the sequence which represented the posi- tion of the Paleocene/Eocene boundary as determined by Cuban studies of the foraminifera (herein called the "boundary layer"), samples were collected at 10-cm intervals to a distance of 2.0m; beyond 2.0m, samples were collected from 0.5 to 6m apart, de- pending upon the lithologies and the extent of weathering. Above the marked point, samples were collected at intervals of 0.5m to 2.0m. These samples as well as the samples collected previously by Cuban scientists are shown on a stratigraphic col- umn in text-figure 2.
Sample SFE-0 is located within the succession (above the Paleocene/Eocene boundary layer) at a well-defined contact be- tween a hard, coarse sandstone and a mudstone/marl with white marly clasts which can be easily recognized by future investiga- tors; therefore, it was selected as a starting point for measuring the section. Sample SFP-13 was taken from the marked point, or "boundary layer" identified on the basis of planktonic foraminifera in earlier studies by Cuban geologists. By this defi- nition, samples SFP-14 through SFP-37 were presumed to be from the Paleocene part of the section. Samples SFE-0 to SFP-12 and SFE-1 through SFE-16 were presumed to be Eocene.
In the biostratigraphic section we have followed Aubry's (1995, p. 215) concepts of lowest and highest occurrence, and that of first and last appearance datum, explained in the following way. A biozone is a stratigraphic unit defined by two or more paleontologic events that are lowest or highest occurrences (LO and HO, respectively). The temporal interval represented by a biozone varies regionally as diachrony occurs, or is unreliable due to preservational problems or differences in taxonomic con- cepts. The maximum temporal interval covered by a biozone corresponds to the life span of a given species in the case of a to-
58
Micropaleontology, vol. 45, supplement 2, 1999
80?
Straits of Florida
Atlantic Ocean
Caribbean Sea
0 50 100 150 km I I
I 1 i 0 50 1O mi
-Ill
TEXT-FIGURE 1 Location map of the San Francisco de Paula section in western Cuba.
tal range zone. The maximum temporal interval is referred to as the biochron, bounded by the first (temporal; evolutionary) ap- pearance datum (FAD) and the last (temporal; evolutionary) ap- pearance datum (LAD) of this species. In the geographic region where this species has its maximum stratigraphic range, its LO corresponds to its FAD, and its HO corresponds to its LAD. Outside the region, the LO and HO of this species differ from its FAD and LAD, respectively, and the time interval of that zone represents less than the biochron.
BIOSTRATIGRAPHY
Paleocene/Eocene boundary
Formal redefinition of the Paleocene/Eocene boundary cur- rently awaits the selection of a global stratigraphic section and a point within that section that will serve as the standard defini- tion of this boundary. While awaiting redefinition of the Paleocene/Eocene series boundary, many stratigraphers have recognized a stratigraphic interval between the top of the Thanetian Stage and the base of the Ypresian Stage in which to locate the Paleocene/Eocene series boundary. This interval, re- ferred to as "Paleocene/Eocene boundary interval", spans ap- proximately 2 million years (from latest Chron C25r to mid-Chron C24r). One of the sources of confusion in determin- ing this boundary is that many authors have equated the Paleocene/Eocene boundary with the NP9/NP10 calcareous nannofossil zonal boundary (55 Ma in Berggren and Aubry 1996; Aubry et al. 1996). It now appears that the age of 55 Ma applies to two horizons. The original 55 Ma age estimate in
Chron C24r in DSDP Site 550, which was also used as a calibra- tion point in the construction of the latest geomagnetic polarity time scale, lies at an unconformity separating stratigraphic lev- els low in Zone NP9 from those low in Zone NP10. The result is a younger (higher) NP9/NP10 zonal/chronal position in Chron C24r (Aubry et al. 1996). Thus, the age of 55 Ma applies to two distinct levels: 1) the 408 mbsf level in Site 550 (Swisher and Knox 1991) which corresponds to a NP9/NP10 biozonal con- tact; and 2) the NP9/NP10 biochronozonal boundary (Cande and Kent 1992). This has serious implications for correlation to the GPTS. Adding to this, there is a large uncertainty in the age estimates determined for events in Chron C24r; e.g., the uncer- tainty on the estimated age of the NP9/NP10 biochronal bound- ary is >0.5 my (For a discussion of the uncertainty on the numerical chronology in Chron C24r and the problems related to the accuracy of the radioisotopic ages, see Berggren and Aubry 1996.)
At present the series boundary is placed at the base of the Ypresian Stage (represented by the leper Clay Formation). The stratigraphic interval that is under consideration for redefining the Paleocene/Eocene boundary is between the top of the Thanet beds (-56.6 Ma, in latest Chron C25r; Zone NP8; Zone P4) and the base of the Ypresian Stage (-54.37 Ma, in mid-Chron C24r; at the NPlOa/b zonal boundary, and very close to the P6a/P6b, -54.0 Ma, zonal boundary [Berggren and Aubry 1998; Aubry 1998]). The younger age estimates for the Paleocene/Eocene boundary found in recent works, which differ from Berggren et al. (1985) and Aubry et al. (1988), are a result
59
84?
Gulf of Mexico
-24?
76 760
359
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
of the recalibration of the magnetochronologic framework by Cande and Kent (1992, 1995). In the San Francisco de Paula section, the boundary is placed at the LAD of Morozovella velascoensis (= base of Zone P6a in Berggren et al. 1995).
Text-figure 3 shows the stratigraphic correlation of six radio- larian biostratigraphic zonations for the Paleocene and lower Eocene to the revised integrated magnetobiochronologic scale of Berggren et al. (1995) via co-occurring planktonic foram- inifera, calcareous nannofossils and/or diatoms as reported by various authors, and converted to the presently applied concepts described in Berggren et al. (1995). For an explanation of how the various radiolarian zonal boundaries have been correlated to the Berggren et al. (1995) biochronologic scale see footnotes 1-37 in the caption to text-figure 3.
The ideal situation with completely cored and suitably fossiliferous sections spanning the Paleocene and lower Eocene does not exist as yet, and therefore we have had to use various approaches to complete the stratigraphic correlations diagram. It must be pointed out that the absence of a particular fauna does not necessarily mean that this interval is missing or was never deposited within a marine sedimentary sequence but rather, that up to the present time it has not been found for a variety of rea- sons. In DSDP sedimentary sequences, this interval may corre- spond to a short duration and thus be very thin, and may have been missed due to spot-coring, poor recovery, gaps in sam- pling, or, if it was in the older part of the stratigraphic column, lack of penetration. Dissolution of microfossils and paleogeo- graphic distributions are also significant factors to be consid- ered.
Foreman (1973) created the first lower Paleogene low latitude radiolarian zonation based on material from DSDP Leg 10 in the Gulf of Mexico. The zonation includes from oldest to youn- gest: the "unzoned" interval in the lower upper Paleocene, and the Bekoma bidartensis and Buryella clinata Zones. This zonation has been correlated to the Berggren et al. (1995) inte- grated magnetobiochronologic Cenozoic scale using data avail- able from co-occurring calcareous microfossils (Bukry 1973a; text-figure 3 herein).
The sediment cores recovered on DSDP Legs 10 and 15 were discontinuously cored. Thus many critical intervals were not observed until Nishimura (1987) tentatively established the up- per Paleocene Bekoma campechensis Zone based on material from DSDP Site 603 (lat. 35?29.66'N, long. 70?01.70'W, water depth 4634 meters). Subsequently she formally described and subdivided the B. campechensis Zone into three subzones (Nishimura 1992) based on a continuous sedimentary sequence from North Atlantic DSDP Site 384 (lat. 40?21.65'N, long. 51?39.80'W, water depth 3909 meters). The three subzones are, from oldest to youngest, Peritiviator(?) dumitricai, Orbula discipulus, and Stylotrochus nitidus-Pterocodon(?) poculum. Increased stratigraphic resolution also resulted in a revision of the lower boundary of the Bekoma bidartensis Zone. Age deter- minations based on co-occurring calcareous nannofossils (Tucholke et al. 1979; Okada and Thierstein 1979) have al- lowed correlation of these zones with the Berggren et al. (1995) scheme (text-figure 3 herein).
The standard low latitude radiolarian zonation, and its correla- tion to the geomagnetic polarity scale (text-figure 3) is taken from the Sanfilippo and Nigrini (1998a) re-examination of 24
DSDP sites and their correlation via calcareous nannofossils to the Berggren et al. (1995) time scale. None of the sedimentary sequences investigated by Sanfilippo and Nigrini contained radiolarian-bearing sediments exactly spanning the Paleocene/Eocene boundary interval. In spite of this, Sanfilippo and Nigrini were able to establish a sequence of 70 datums in a 10 my interval spanning the boundary from the upper Paleocene Bekoma campechensis Zone to the upper part of the Buryella clinata Zone in the lower Eocene. Further, they were able to re- late these datums via published data on co-occurring calcareous nannofossils to the calcareous nannofossil zonation. Using the definitions applied by Berggren et al. (1995) for determining the stratigraphic position of the Paleocene/Eocene boundary in terms of calcareous nannofossils and foraminifera (i.e., NP9/NP10 boundary, -55 Ma), they established six reliable lowest radiolarian occurrences which approximate the Paleocene/Eocene boundary: Calocycloma castum, Theocoty- lissa auctor, Lamptonium fabaeforme fabaeforme, Podocyrtis (Podocyrtis) papalis, Giraffospyris lata and Phormocyrtis turgida. The two major differences between this zonation and that of Foreman (1973) are: (1) the Sanfilippo and Nigrini (1998a) zonation combines radiolarian information from sev- eral sites with that of co-occurring calcareous microfossils, making it possible to correlate the radiolarian zonation to the geomagnetic polarity scale (and thus estimate ages for radiolarian datums); (2) the lower boundary of the Bekoma bidartensis Zone has been lowered as a result of Nishimura's (1992) recognition of the older Bekoma campechensis Zone.
The Caribbean radiolarian zonation proposed by Maurrasse (1973, unpublished thesis) is included herein because it was used by Flores Albin in his initial interpretation of the biostratigraphy in the San Francisco de Paula Section (see Ap- pendix 1 by Flores Albin; see also "Biostratigraphy of the San Francisco de Paula Section" below). This zonation consists of four zones in the upper Paleocene to lower Eocene. From oldest to youngest, they are: Theocorys(?) pseudophyzella, Eucyr- tidium cubense (=Phormocyrtis cubensis), Bekoma bidartensis, and Buryella clinata. We caution against using the Maurrasse zonation because the two "new" radiolarian zones proposed by Maurrasse, Eucyrtidium cubense and Theocorys(?) pseudo- phyzella, have never been formally described. Maurrasse (1973) correlated his zonal scheme to calcareous microfossil zones and to the radiolarian zonation of Foreman (1973) permitting us to correlate his zonal scheme to the current radiolarian zonation and the current geochronologic scale (Berggren et al. 1995).
Maurrasse did not define the base of the Theocorys(?) pseudophyzella zone. However, he considered the top of the zone approximately synchronous with the extinction of the nominate species and coincident with the base of the Eucyrtidium cubense zone. According to Maurrasse's correla- tions, this radiolarian zone spans approximately the upper part of the Globanomalina (=Globorotalia) pseudomenardii Zone (P4) and its uppermost limit is possibly in the G. velascoensis Zone (P5). The zone includes the LOs of Lamptonium pennatum, Buryella tetradica and Lychnocanoma auxilla.
In Maurrasse's original definition, the base of the Eucyrtidium cubense zone is defined by the LO of Eucyrtidium cubense (Phormocyrtis cubensis) which is approximately synchronous with the LO of Podocyrtis (Podocyrtis) papalis. This zone in- cludes the LOs of Phormocyrtis turgida, Lamptonium fabae- forme fabaeforme, and Theocorys acroria. The top coincides with the base of Foreman's Bekoma bidartensis Zone.
60
Micropaleontology, vol. 45, supplement 2, 1999
BIOZONES
lan ,.ton c LU i~~~~~~ ~~~~~ Benthc Calcareous Rar-ioar a LITHOLOGY trnifa BtcSAMPLES foramin.fea aria Radaia AGE ~~z :~~~~~~~~ LI l n?
l_^/OSA MPLESforaminifera narnofossa's sari ioo & emran.cez- .o& c'mz-Ab-
mRod',guez e: a, Az,~e's x"
_______trlis voi '-e * ^ *surr? :"ac:5^
_________
40-
~~~~~~~~~~. - . ' 1"1
.0-- - . .- . --;
..............
30 --' ^ -*^ *^ -^ ^ ^. .: ..
.._ ..._ , / - , ? j[
,~- ^ -
-- ~
<:.. .,
2 1
? ' T
0~~~~~~~~~~~~~'-~i-
J.E
5,
ao
u;
_ _
0 5 >
"u '5
c ET
C)
S .E
.;z
E-
'0 re
o
c *It (/; IL L
.c
z Li] 8 LU
-J
Lij
0
'5
C,..
5 c: rata B.
bidaa,tesns
F -0
t+.,
a- NPO z CZ CE 1-
8
NP6
Li
LU
Lii -I C
^r
all
Z:
Li
't;
LUJ
TEXT-FIGURE 2 Stratigraphic interpretation of the upper Paleocene-lower Eocene interval (Apolo Formation) of the San Francisco de Paula section with lithologic col- umn, showing thickness in meters and sampling intervals. Samples collected in previous field excursions by Cuban geologists are designated Cl through C62; samples collected in this study are indicated by the prefixes SFP and SFE. The Paleocene/Eocene boundary in this section has been recognized, at the unconformity between samples SFP-22 and SFP-21, on the basis of planktonic foraminifera (Fernmdndez-Rodrfguez et al. 1999, this volume) and is placed at the LAD Morozovella velascoensis, marker for the P5/P6a zonal boundary used in Berggren et al. (1995) to approximate the boundary. Plank- tonic foraminiferal biozones follow Berggren et al. 1995; benthic foraminiferal biozones Berggren and Miller 1989; and calcareous nannofossil biozones Martini 1971.
61
__
I I t7,:, .
- 1 - -
.! . : -.: h
. '. . : i : T-. -
.,! I' %':
- - -
v!r d
59
o:
a;4
-5'
<
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
As shown on the stratigraphic correlation diagram (text-figure 3), it is apparent that Maurrasse's E. cubense zone falls within the Bekoma bidartensis Zone as applied by Sanfilippo and Nigrini (1998a). Examination of his data reveals that many of the reported radiolarian events occur within the Bekoma bidartensis Zone. It is of interest to note that the E. cubense zone includes three of the LOs (Phormocyrtis turgida, Lamptonium fabaeforme fabaeforme and Podocyrtis (Podocyrtis) papalis) that Sanfilippo and Nigrini (1998a) use to approximate the Paleocene/Eocene boundary. However, recog- nition of B. bidartensis and Phormocyrtis cubensis in the Carib- bean sedimentary sequences is commonly hampered by preservation problems. In DSDP Site 152, for example, B. bidartensis is absent throughout the B. bidartensis Zone while the somewhat more robust P. cubensis is present in the upper part of the Bekoma campechensis Zone.
Maurrasse's recognition of the E. cubense zone between the up- per part of the B. campechensis and the lower part of the B. bidartensis Zones may be the result of a local difference in the stratigraphic range of E. cubense due to paleoenvironmental ef- fects, or the result of poor preservation. Furthermore, the ab- sence of P cubensis in the continuous sequence from North Atlantic DSDP Site 384 prevents recognition of the E. cubense zone at that site. However, Nishimura (1992) observed the evo- lutionary transition of B. bidartensis from B. campechensis at this site, resulting in a lowering of the base of the B. bidartensis Zone (Nishimura 1992), which removes the necessity of a zone, based on a paleoenvironmentally sensitive marker species, be- tween the B. campechensis and B. bidartensis Zones, at least as presently defined. Sanfilippo and Nigrini (1998a) observed the LO of P. cubensis in the upper part of the B. campechensis Zone prior to the LO of B. bidartensis in the Leg 10 material. This is only doubtfully considered as evidence for the presence of the E. cubense zone in the Caribbean because the Leg 10 sediment sequences are intermittently cored, and total radiolarian abun- dance is low and preservation poor. The lack of continuously cored material from duplicate sites prevents resolving the biostratigraphic problem of which event takes place first, and to what extent these events are effected by paleoenvironment and preservation. Correlation and standardization of the various radiolarian zonal schemes proposed for the Paleocene and lower Eocene will require much additional work using independent age control provided by isotope and/or paleomagnetic studies.
In contrast to the sequence of events documented in the tropics, the mid to high southern latitude pelagic sediments examined by Hollis (Strong et al. 1995) present a different succession. For example, there is a gap between the HO of Bekoma campe- chensis and the LO of B. bidartensis. In the Mead Stream sec- tion, this stratigraphic gap is represented by a 40 meter thick interval, containing a distinctive fauna lacking many of the tropi- cal taxa diagnostic of the B. bidartensis Zone (e.g., Calocycloma castum, Lamptonium fabaeforme fabaeforme, L. pennatum, Podocyrtis (Podocyrtis) papalis and Thyrsocyrtis tarsipes).This faunal assemblage was described by Hollis (1997) as a local zone, the Bekoma campechensis-bidartensis "gap" zone.
The applicability of Hollis's New Zealand high latitude zonation (paleolatitude 55?S; Hollis 1993, 1997; Strong et al. 1995) has not been tested in low latitudes due to the lack of radiolarian-bearing lower Paleocene sedimentary sequences, but it has been included here because of its relevancy and poten- tial use in other regions. Results of the integrated foraminiferal, radiolarian and dinoflagellate study by Strong et al.'s (1995)
provide a biostratigraphic and paleoenvironmental assessment for the uppermost Cretaceous to upper middle Eocene, spanning the Paleocene/Eocene transition in Marlborough, New Zealand. Hollis (1993) correlated his zones to the Berggren et al. (1985) time scale, using age control from co-occurring foraminifera (e.g., Strong 1977, 1981, 1984; Strong et al. 1987) and calcare- ous nannofossils (Burns et al. 1973). The South Pacific radiolarian zones have subsequently (Hollis 1997) been corre- lated to the Berggren et al.'s (1995) time scale, and included herein (text-figure 3) for a comparison with the zonal schemes developed for other regions. Nine of Hollis (1997) South Pacific zones are included herein, from oldest to youngest: Amphisphaera aotea [RP(SPac)l], Amphisphaera kina [RP(SPac)2], Buryella granulata [RP(SPac)3], Buryella formanae [RP(SPac)4], Buryella tetradica [RP(SPac)5], Bekoma campechensis [RP(SPac)6], Bekoma campechensis- Bekoma bidartensis [RP(SPac)7], Bekoma divaricata [RP(SPacO8] and Bekoma divaricata-Theocampe mongolfieri [RP(SPac)9].
The boreal zonal scheme resulting from investigations of lower Paleogene Russian sequences from the eastern side of the Urals, western Siberia, the central Volga and Caspian regions (Kozlova 1983, 1984, 1993) are also included because of their eventual future use. These zones, lacking many of the taxa oc- curring in tropical and subtropical regions (belonging to the genera Bekoma, Diclyoprora (Theocampe) and Thyrsocyrtis), were correlated to the tropical radiolarian zonation using epi- sodic occurrences of tropical taxa in the boreal assemblages. In some cases the boreal zones have been dated using co-occurring diatoms, or in the case of the assemblages from the Caspian re- gion by co-occurring foraminifera.
Biostratigraphy of the San Francisco de Paula Section
The Paleocene/Eocene boundary in this succession is within an unconformity, and has been placed at the HO of Morozovella velascoensis (Fernmandez-Rodriguez et al. 1999, this volume).
Weathered and indurated shales in the San Francisco de Paula section contain poorly preserved radiolarians which vary in abundance. Radiolarians from the soft shaley layers are some- what better preserved and present in moderate numbers. Age determinations are constrained by the presence of a few resis- tant "marker" species. In the middle part of the section (text-figure 4) there is a thick interval (approximately 16m) with poorly preserved, unidentifiable radiolarians, and adjacent, above and below, are two barren intervals (approximately 3.5m and 5m, respectively). The relative abundances of taxa and pres- ervation are reported in Table 1; a species list is presented in Appendix 2. Definitions of the radiolarian zones used in this pa- per can be found in Appendix 3.
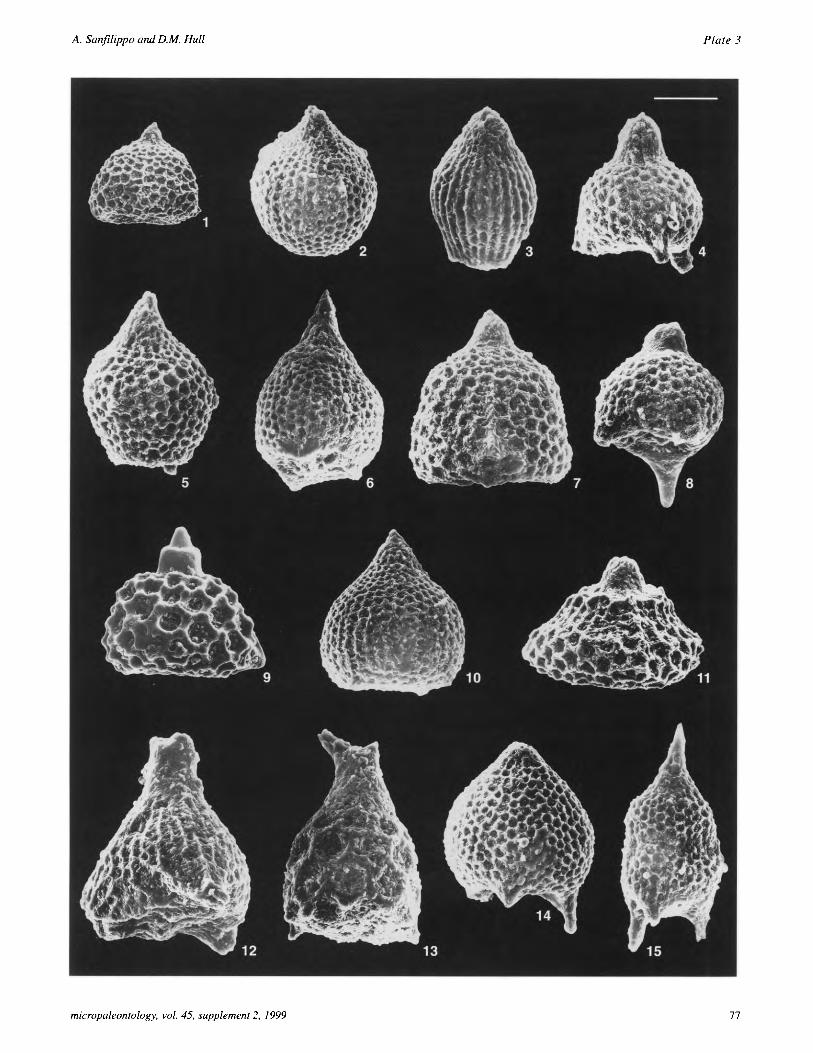
The material examined by us was too poorly preserved for de- tailed biostratigraphy. The marker species used in this strati- graphic interval have only been doubtfully identified (see Plates 1 and 2). Although numerous taxa of the genus Bekoma are present, they cannot be accurately identified at the species level because the characteristic feet, joined in some taxa, have not been observed intact. In spite of this we have made an attempt to distinguish B. campechensis from other co-occurring taxa of the genus Bekoma by using the characteristic shape of the cephalis for each taxon. Bekoma campechensis has a long dome-shaped to rectangular cephalis, whereas B. demissa has a cephalis with numerous pores that appears slightly depressed into the thorax. The cephalis and proximal elongated part of the thorax in B.
62
Micropaleontology, vol. 45, supplement 2, 1999
Nanno Plankt. Biochrons S fil & Kozlova, 1984;
TIME EPOCH Biram B y Biochrons Nigrini, 1998; in Maurrasse, 1973 Foreman, 1973 Nishimura, 1992 Central Volga, E.
N ew Zealand,
(Ma) EPOCH Biochrons 1973b, Martini, press Caribbean Gulf of Mexico N. Atlantic 40N .N e
Berggren et 1975; 1 Tropical . Ural, W. Siberia, South Pacific 50S al., 1995 Okada & 191 Tropical Eurasia
Bukry, 1980
Phormocyrtis striata striata
Phormocyrtis striata striata
Heliodiscus hexasteriscus
B. divaricata - T. mongolfieri
RP(SPac)9
29
Buryella clinata
, . , 22 , . . Spongotrochus paciferus
... - 23 - - - .
Petalospyris fiscella
Bekoma bidartensis Bekoma bidartensis
Eucyrtidium cubense
Theocorys (?) pseudophyzella
unzoned
- - 11 - - 15
Bekoma divaricata
RP(SPac)8
Petalospyris foveolata
- 16 , B. campechensis- bidartensis RP(SPac)7
BeKoma bidartensis
17
c
- 18
- 1 * b
a
aU a E CD a co E 0
Co Co
Tripodiscinus Bekoma
sengilensis campechensis
/ RP(SPac)6
Buryella 32 tetradica
i IInbUboI Idul d larnacium
- 20
CU
a)
co
-a 0
C...
0-
Eu 0 0
Buryella tetradica
RP(SPac)5
33
Buryella foremanae
RP(SPac)4
, 34
Buryella granulata
RP(SPac)3 - 35
A. Idna RP(SPac)2 36
A. aotea RP(SPac)1 37
TEXT-FIGURE 3 Stratigraphic correlation of radiolarian, calcareous nannofossil and planktonic foraminiferal zones to the geochronologic time scale of Berggren et al., 1995. The broad band spanning the interval between -54.6 to 55.5 Ma reflects current opinion on the position of the Paleocene /Eocene boundary. Planktonic foraminiferal specialists commonly use the P5/P6 boundary (-54.7 Ma) and calcareous nannoplankton specialists the NP9/NP10 (CP8/CP9) zonal bound- ary (-55.0 Ma) to represent the boundary for purpose of correlation (see also p. 20 comments on Berggren and Aubry 1998 and Aubry 1998). Hachured in- tervals represent intervals not represented by radiolarian-bearing sediments. Subdivision of Bekoma campechensis Zone: a = Peritivator (?) dumitricai; b = Orbula discipulus; c = Stylotrochus nitidus-Pterocodon (?) poculum; P*=Pax + P0. [See notes to text-figure 3 on following 2 pages.]
63
P9 CP11
P8
NP13 Phormocyrtis striata striata
an 1
u z LU 0 0 w
P7
c CU
OPi 0
b p 6
NP12
NP11
NP10
b
a
C 9
P5
50 -
51 -
52 -
53 -
54 -
55 -
56 -
57
58 -
59 -
60 -
61 -
62 -
63 -
64 -
65
b CP 8
a
CP7
CP6
CP5
CP4
P4
a) CU
NP9
NP8
NP7
NP6
NP5
c
b
a
b
Bekoma bidartensis
-4 -
c
C) 9,
0. E
E 0
in
P3
LU z LU
0 LU
<-i Qn
a
CP3 NP4
c
UJ wo P1 NP3
b
CP2
CP b
1 a a
NP CRET.
NP2
NP1 F, I
VI
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
NOTES TO TEXT-FIGURE 3 Ongoing research on the taxonomy and biostratigraphy of Paleogene planktonic foraminifers has necessitated substantial modifications in the numbering system for zones of the Paleocene to lower Eocene interval. Herein we have followed the modifications made by Berggren et al. (1995). For radiolarian zonal schemes that previously had been correlated to foraminiferal zonations older than the Berggren et al. (1995) scheme we have converted prior age assignments to follow the new framework. This is particularly important for the Paleocene/Eocene boundary interval, where Zone P5 of Berggren and Miller (1988) is no longer thought to exist because of an over- lap in the ranges of Globanomalina pseudomenardii and Morozovella subbotinae. To preserve the numerical continuity, a new Zone P5 was defined by Berggren et al. (1995) that is equivalent to P6a of Berggren and Miller (1988) minus a short overlap between G. pseudomenardii and M. subbotinae. For further discussion and emendations concerning the other zones see Berggren et al. (1995).
1. Follows correlation made by Foreman (1973) see footnote 12.
2, 3. In material recovered during DSDP Leg 10 the boundary be- tween Buryella clinata and Bekoma bidartensis Zones oc- curred in a stratigraphic gap between 94-32CC and 96-3-1. However, re-examination by Sanfilippo and Nigrini (1998a) of 24 DSDP cores in this stratigraphic interval determined that the uppermost part of the B. bidartensis Zone and the LO of B. clinata occurred in Cores 13 and 12 respectively at DSDP Site 313 equivalent to nannofossil Zone CP10 and in the Globorotalia subbotinae foraminiferal Zone (corresponding to the lower part of CP9b). Precise placement of the B. bidartensis/B. clinata zonal boundary is further hampered by the lack of radiolarian-bearing sediments in the interval equiv- alent to nannofossil Zone CP9, indicated by hachuring.
4. Sanfilippo and Nigrini (1998a) applied Foreman's (1973) con- cept of the B. bidartensis Zone, using the morphotypic first ap- pearance of B. bidartensis rather than the evolutionary transition applied by Nishimura (1992) resulting in a lower zonal boundary than that indicated by Nishimura (1992). The first appearance of B. bidartensis is approximated at the CP6/CP7 nannofossil zonal boundary.
5. For placement and correlation to calcareous nannofossil zones of the lower boundaries of Bekoma campechensis Zone and its subzones see footnotes 18-20.
6. Poor radiolarian coverage in this interval prevented Maurrasse (1973) from determining this zonal boundary and its relation- ship to calcareous microfossil zones. He suggested that the P. striata striata Zone correlated with the lower part of the Hantkenina aragonensis foraminiferal Zone (Zone P10), but that its correlation to the Globorotalia palmerae Zone (Zone P9) could not be determined.
7. Maurrasse implied that the lowermost part of the Buryella clinata Zone may partly correlate to the upper part of foraminiferal Zone Globorotalia subbotinae/Globorotalia wilcoxensis to Globorotalia aragonensis or Globorotalia palmerae Zone. Herein we have correlated the lower bound- ary of his Buryella clinata Zone with the upper half of nannofossil Zone CP9b.
8. Maurrasse correlated the Bekoma bidartensis Zone in his ma- terial with most of the foraminiferal Globorotalia edgari and Globorotalia subbotinae/Globorotalia wilcoxensis Zones and with the material used by Foreman (1973) which we were able to correlate to nannofossil Zone CP8b.
9. The Eucyrtidium cubense zone spans most of the Globorotalia velascoensis foraminiferal Zone and the lowest part of the Globorotalia edgari Zone. Its lower boundary has been ap- proximated at the nannofossil CP8a/CP8b boundary just below the base of foraminiferal Zone P5.
1Q lLTheocorys (?) phyzella zone spans the upper part of the Globanomalina pseudomenardii (P4) Zone. Its lower bound- ary is undefined. Maurrasse (1973) correlated this zone with the lower part of the 'unzoned interval' represented by DSDP core sections 86-7 and -8, and core section 96-5 (Foreman
1973; Sanfilippo and Riedel 1973). Herein we have given the zone a maximum interval, and thus interpret the base of the zone to lie within nannofossil Zone CP5 as suggested by Bukry's data (1973a).
12. DSDP Leg 10 drilled three discontinuous holes that did not quite overlap in the Paleocene-Eocene interval. Correlation with calcareous microfossil zones from this material [nannofossil zones from Bukry (1973a) and foraminiferal zones from McNeely (1973)] has made it possible to extrapo- late the radiolarian zonal boundaries to the geochronological scale. The lowermost sample (94-28CC) in the Phormocyrtis striata striata Zone lies in the Discoaster lodoensis nanno- fossil Zone (Zone CPll) and the Hantkenina aragonensis foraminiferal Zone (Zone P10). Because samples 94-29CC and 94-30CC in the underlying Buryella clinata Zone are in the foraminiferal Globorotalia aragonensis/formosa Zone (P8/7) Zone or nannofossil Zone CP1 1 we have provisionally placed the unrecovered boundary within the lower part of Zone CPU1.
13. This boundary falls in a long unrecovered interval between 94-31CC and 94-32CC within the Globorotalia rex foram- iniferal Zone. It was not possible to determine a nannofossil age based on the Leg 10 material for this boundary. We have tentatively placed the boundary in the top part of Zone P5 since the Bekoma bidartensis Zone is also within this zone.
14. The boundary between the B. bidartensis Zone and the un- zoned interval falls in an unrecovered interval between Cores 96-3 and 96-4, both in nannofossil Zone CP8b. To avoid con- fusion caused by the Berggren et al. (1995) modifications in the numbering system of the foraminiferal zones we have fol- lowed the nannofossil interpretation of Bukry (1973a) and ex- trapolated the placement of this radiolarian boundary to fall within Zone CP8b.
15. The oldest Paleocene material recovered during DSDP Leg 10 belongs to the foraminiferal Globanomalina pseudomenardii (Zone P4) and Heliolithus kleinpellii nannofossil Zone (= Zone CP5). The lowermost boundary of the unzoned interval is therefore placed at the lower boundary of Zone CP5.
16. Nishimura (1992) correlated her radiolarian zones to the cal- careous nannofossil zones reported for DSDP Site 384 by Okada and Thierstein (1979). No foraminiferal zones are available except for the lower part of the Paleogene section. At Site 384 coring was continuous between Cores 5 and 6 but the entire lower Eocene and the top part of the Paleocene are miss- ing. The section is interrupted in the lower part of a normal po- larity magnetochron which represents Chron C25n. The youngest part of the Bekoma bidartensis Zone recovered at Site 384 in Core 6, not necessarily the top part of the zone, is correlated with nannofossil Subzone Campylosphaera eodela (CP8b) and given maximum time equivalence.
17. The Bekoma campechensis/B. bidartensis zonal boundary is within nannofossil Zone CP7 arbitrarily placed in the lower part of the zone to accommodate radiolarian Subzone
64
Micropaleontology, vol. 45, supplement 2, 1999
Stylotrochus nitidus-Pterocodon (?) poculum which is corre- lated to nannofossil Zone CP6 and the lower part of Zone CP7.
18. The Orbula discipulus Subzone corresponds to nannofossil Zone CP5.
19. The Peritivator (?) dumitricai Subzone corresponds to nannofossil Zones CP3 and CP4.
20. The oldest Paleocene radiolarian-bearing sediments in Core 384-11 belong in foraminiferal Zone P2, and have been as- signed to nannofossil Zone CP3, thus the bottom of the Bekoma campechensis Zone has been arbitrarily placed at the lower boundary of Zone P2.
21. Kozlova (1984) suggested that it was possible to combine the two boreal radiolarian zones, Heliodiscus lentis and H. hexasteriscus, into a marker level that could be correlated with the two tropical Zones Buryella clinata and Phormocyrtis striata striata. She considered the bottom of the P. striata striata Zone to be within the H. hexasteriscus Zone (Kozlova 1984) and correlated the H. hexasteriscus Zone to the foraminiferal Zone Globorotalia aragonensis s.l. Krasheninnikov (1969). Herein the H. hexasteriscus Zone has been equated with the stratigraphic interval represented by the middle of nannofossil Zone CP10 through Zone CP12a. Foraminiferal zones of the southern USSR are from Krasheninnikov 1971.
22. Kozlova (1984) identified the presence of both the Heliodiscus lentis and Buryella clinata Zones in the upper part of the central Volga Formation, and thus it is possible to make an approximate correlation of the H. lentis zonal com- plex with the interval represented by the foraminiferal Globorotalia formosa (P7) to the lower part of Globorotalia aragonensis (P8) Zones, and to nannofossil Zone CP10.
23. Spongotrochus paciferus and H. lentis Zones are correlated to diatom Pyxilla gracilis Zone (Kozlova 1984) and the lower boundary herein approximated to the lower half of nannofossil Zone CP9b via Strelnikova (1987).
24. Based on co-occurring diatoms Kozlova (1984) correlated the Petalospyris fiscella Zone to Glezer's Coscinodiscus payeri Zone, which we in turn via Strelnikova (1987) have roughly interpreted to be time equivalent to the upper part of nannofossil Zone CP9.
25. According to Kozlova (1984) the Petalospyris foveolata Zone corresponds to diatom Coscinodiscus uralensis Zone equivalent to the lower Globorotalia subbotinae foram- iniferal (P6) Zone or nannofossil Zone CP9a.
26. The Tripodiscinus sengilensis and Buryella tetradica Zones are both correlated with diatom Trinacria ventriculosa Zone (Kozlova 1984) here interpreted as time equivalent to nannofossil Zone CP5 through CP7 (via Strelnikova 1987), and given equal time equivalence. The lower limit of B. tetradica is unknown because of the lack of material. How- ever, Kozlova (1984) compared her material with that recov- ered on DSDP Leg 10 which was correlated to foraminiferal Zone Globanomalina pseudomenardii P4 (see Foreman 1973).
27. The Thecosphaera larnacium Zone is correlated with foram- iniferal Globorotalia angulata s.l. Zone Krasheninnikov (Zone P3) and arbitrarily placed within the upper part of it to accommodate part of the Buryella (?) alifera Zone.
Foraminiferal zones of the southern USSR are from Krasheninnikov 1971.
28. The Buryella (?) alifera Zone has no tropical equivalent, however, Kozlova (1984) considered the assemblage similar in age to that described by Dumitrica (1973) and estimated as Danian in age (equivalent to foraminiferal Zones P1 and P2). The Cromyocarpus ovatus assemblage is an impoverished version of the B. (?) alifera zonal assemblage with which it is also equivalent in age. Co-occurring diatoms from the Trinacria heibergiana Zone make it possible to estimate the age as equivalent to nannofossil Zone CP4 through CPlb or lower. Radiolarians have not been found near the upper and lower limits of the C. ovatus and B. (?) alifera Zones.
29-31.Hollis (1997) calibrated his new radiolarian zones [RP(SPac)6-RP(SPac)9] with the revised Paleogene time scale (Berggren et al. 1995) by direct dating of radiolarian events using dates from co-occurring foraminifera and dinoflagellates at Mead Stream, Marlborough, New Zealand (Strong et al. 1995).
32. The primary reason for placing the RP(SPac)5/RP(SPac)6 boundary no lower than calcareous nannofossil zone CP5 in the New Zealand region is that the HO of the Buryella tetradica assemblage is within Zone CP5 at DSDP Site 208 (Hollis 1993). The top of Buryella tetradica Zone [RP(SPac)5] is defined by the LO of Bekoma campechensis. Thus, Hollis (1993) placed the upper boundary of the B. tetradica Zone at the base of foraminiferal Zone P4, based on age control for the earliest occurrences of Bekoma campechensis in the Gulf of Mexico (Foreman 1973), which, lacking further evidence, coincides with its lowest occur- rence in the South Pacific. However, the oldest Tertiary radiolarians recovered on Leg 10 are from sample 86-8-CC (dated by foraminifera as upper Paleocene Zone P4, and by calcareous nannofossils as Zone CP5), and no radiolarian samples were recovered below this sample. Therefore, the LO of Bekoma campechensis cannot be determined at this site. Subsequent work by Nishimura (1992) in the North At- lantic established the LO of Bekoma campechensis in this re- gion within the upper part of CP3, equivalent to foraminiferal Zone P2 (see comment 20) which is significantly lower than the LO in the Gulf of Mexico. Herein we have placed the top of the New Zealand Buryella tetradica Zone within nannofossil Zone CP5 as indicated by Hollis (1993, 1997).
33. At DSDP Site 208 the LO Buryella tetradica is in the middle of nannofossil Zone CP3 approximately at the lower/upper Paleocene boundary (Hollis 1993).
34,35 Reliable age control is lacking at DSDP Site 208 and in the Marlborough sections from New Zealand, and these events have therefore been approximated at the upper nannofossil Zones CPlb and lower CP2 respectively. The LOs of Buryella granulata and Buryella foremanae are within nannofossil Zones CPlb to CP2 at DSDP Site 208 but their location within this interval is unknown (see Strong et al. 1995; Hollis 1997).
36. The LO of Amphisphaera kina is below the base of nannofossil Zone CPlb but above the base of foraminiferal Zone Plb, and therefore placed in the uppermost part of CPla.
37. The LO Amphisphaera aotea is placed directly above the K/T boundary; see discussion in Hollis (1993, p. 308).
65
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
campechensis are poreless with a thick hyaline wall, whereas in B. bidartensis this feature is not so prominent. In B. divaricata the length of the cephalis and thorax, and the poreless part of the thorax is markedly shorter. Thus we are confident that in the lower part of the section we have identified Bekoma campechensis correctly, and separated it from other co-occurring taxa belonging to the genus Bekoma, which have been lumped in the category Bekoma spp. In the higher part of the section (SFP-14 - SFP-8) we have used the Bekoma spp. cat- egory to include all taxa belonging to the genus that cannot be positively identified. However, it is possible that a high propor- tion of the observed specimens are in fact B. bidartensis based on the fact that the HOs of B. campechensis, B. demissa and B. divaricata are usually below the LOs of Giraffospyris lata, Podocyrtis (Podocyrtis) papalis, Theocorys phyzella and Thyrsocyrtis (Thyrsocyrtis) hirsuta (Sanfilippo and Nigrini 1998a). Buryella clinata, the nominate species for the lower Eocene zone has been questionably identified in the upper part of the section (with exception of sample SFP-10) since we are unable to observe the characteristic diagonal pore alignment on the third segment (P1. 2, figs. 14, 15). However, remnants of the broad-based, ridged horn have been a useful morphological characteristic for identifying this taxon and aid in its identifica- tion in the absence of good preservation. Three assemblages representing the B. campechensis, B. bidartensis and B. clinata Zones were recognized as follows.
Biostratigraphic zones recognized in the San Francisco de Paula section
We have interpreted the co-occurrences of Bekoma campechensis, B. demissa, Bekoma spp., Buryella pentadica, B. tetradica, B. foremanae and Lamptonium pennatum to indicate that the lower part of the section (SFP-30A at 20.0 m below SFE-0 through SFP-37 at 43.44 m below SFE-0) is in the lower part of the Bekoma campechensis Zone. Evidence from Nishimura (1992) and Sanfilippo and Nigrini (1998a) confirm the HO of B. pentadica at the Bekoma campechensis/B. bidartensis zonal boundary. The HO of Buryellaforemanae has been documented in DSDP Site 208 and in sections from Mead Stream, Marlborough, New Zealand in the lower Paleocene (Hollis 1993; Strong et al. 1995), where it ranges from the B. foremanae Zone to the top of the New Zealand zonal equivalent of the Bekoma campechensis Zone. Comparison of the strati- graphic ranges of taxa present in the San Francisco de Paula section with those observed in the pelagic sediments from New Zealand has offered insight into the complexity of the problems involved in comparing stratigraphic ranges of taxa from differ- ent biogeographic regions. The faunas represented in the Mead Stream section are diverse, but many of the marker species used in the Cenozoic radiolarian zonation established for the low lat- itudes (Riedel and Sanfilippo 1978; Sanfilippo et al. 1985; Sanfilippo and Nigrini 1998a) and for the North Atlantic (Nishimura 1987, 1992) are either absent or occur only sporadi- cally, and vice versa (Strong et al. 1995). However, a sufficient number of age diagnostic taxa occur in both regions to attempt a preliminary correlation between the mid to high southern lati- tude zonal scheme developed by Hollis (1993, 1997; Strong et al. 1995) and that established for the low latitudes. As a result of this comparison we have found that some taxa encountered by Hollis in the Mead Stream section, e.g., Amphicraspedum prolixum, Bekoma bidartensis, B. campechensis, B. divaricata, Buryella tetradica, Lamptonium pennatum, Phormocyrtis
striata exquisita and Podocyrtis (Podocyrtis) papalis have their LOs higher, and for some of them (B. bidartensis and B. divaricata) their HOs lower, than in the tropics (see also discus- sion under the subheading "Lower Paleogene radiolarian zonations" above).
A second assemblage (SFP-11 at l.lm below SFE-0 through SFP-15 at 1.5m below SFE-0) composed of Theocotylissa al- pha, T auctor, Theocorys phyzella, Thyrsocyrtis (Thyrsocyrtis) hirsuta, Theocotyle nigriniae, Pterocodon tenellus, Podocyrtis (Podocyrtis) papalis and Phormocyrtis turgida is indicative of the Bekoma bidartensis Zone. The third assemblage includes many of the nassellarian taxa present in the B. bidartensis Zone and in addition rare Buryella clinata in sample SFP-10, as well as questionably identified B. clinata in SFP-9A, -9, -8 and SFE-14. Although preservation in most samples prevents posi- tive identification of the marker species (see above), we assign the entire sequence from SFP-10 through SFE-16 to the Buryella clinata Zone. This assignment is supported by evi- dence from co-occurring Podocyrtis (Podocyrtis) papalis and Theocotylissa alpha, represented by later morphotypes than those encountered in the underlying Bekoma bidartensis Zone, and by the fact that the Thyrsocyrtis (Thyrsocyrtis) hirsuta pop- ulation consists of a mixture of early, small morphotypes (PI. 2, fig. 12) together with the more typical later forms which are larger, with medianly thickened feet (P1. 2, figs. 10, 11).
Familiarity with evolving lineages may serve as a useful guide in determining stratigraphic position of one sample versus an- other by deciding where in the evolutionary development the morphotypes of a given lineage belong. If the phylogenetic characters of a morphotype can be considered closer (in resem- blance) to those of the ancestor, then the morphotype is said to be an "early form" indicating that it is from the early part of its lineage. Later morphotypes would thus bear a closer resem- blance to the descendant. For example, the genus Podocyrtis de- veloped from the genus Cryptocarpium (Sanfilippo and Riedel 1992, p. 8) in the upper Paleocene Bekoma bidartensis Zone as described by Sanfilippo and Riedel (1973, p. 531, pl. 36, figs. 1-3). The earliest morphotypes of Podocyrtis (Podocyrtis) papalis have the two cephalic lateral lobes separated from the unpaired eucephalic lobe by furrows that extend from the point of junction of the primary lattice bars with the lattice shell to a point just below that at which the apical spine leaves the ce- phalic surface, the apical bar extends freely within the cephalis. Thus early forms can be distinguished from later forms, in which the lateral furrows join the apical bar at a lower point, and the apical bar is incorporated in the cephalic wall. Furthermore, in the early forms the cephalis is uniformly porous while in later forms the eucephalic part of the cephalis is poreless or sparsely pored. Recognition of the various stages of the evolving mem- bers of the Theocotylissa lineage is important for interpreting the stratigraphy in the San Francisco de Paula section. In this lineage, Theocotylissa alpha evolved from T. auctor and gave rise to its descendant T. ficus by reduction of its three triangular feet and an increase in relative size of the abdomen (Sanfilippo and Riedel 1982). Thus one would refer to morphotypes of T. al- pha having a relatively large abdomen and a peristome bearing three undulations instead of three short feet as "later forms". An additional clue is the size of the horn, which in the "late forms" has been reduced to about half the size of the stout horn inher- ited from its ancestor T. auctor, to resemble the horn present in the descendant T. ficus.
66
Micropaleontology, vol. 45, supplement 2, 1999
PALEOCENE EOCENE EPOCH
Intervals barren of Radiolada (cross hachured); Dissolution
Intervals (dotted)
000000 n 20 0 0 00 n 5 Sample nos. (Cuban)
--n '1 l 'C .C . n -l1 ':n . cn s-
: , NP10 C z I o >l NP10d > > >'
F- D. multiradiatus ', O
1 R.cuspis J J - T. bramlettei <
- C. delus C O
T. contortus ( cn
-- T. orthostylus - D. lodoensis 1t
'on X wX-o _
G. pseudomenardii -I 0 < CD
M. velascoensis 3 0 CD..-
m ~
Eucyrtidium cubense
Bekoma ]i. campechensis :s1
_ _ ._S_.. id :
Bekoma bidartensis
Bekoma a bidartensis X
? Buryella clinata
Radiolaria Biozones Florez-Abin, Appendix 1
Radiolaria Biozones Sanfilippo & Hull, this chapter
Heliostylus spp.
Buryella foremanae
Buryella pentadica
Buryella tetradica
Lamptonium pennatum
Phormocyrtis stnriata exquisita
Bekoma demissa
Bekoma campechensis
Bekoma spp.
Phormocyrtis turgida
Pterocodon tenellus
Amphicraspedum murrayanum
Phormocyrtis cubensis
Phormocyrtis striata striata
Spongodiscus cruciferus
Amphicraspedum prolixum
Spongodiscus quartus quartus
theocorys phyzella
Giraffospyris lata
Podocyrtis (Podocyrtis) papalis
tamptonium fab. fabaeforme
Theocotylissa auctor
Calocycloma castum
Theocotylissa alpha
Thyrsocyrtis (Thyrsocyrtis) hirsuta
Theonftvle nionriniae
Lithochytris archaea
Buryella clinata
Lamptonium sanfilippoae
Stylotrochus quadribrachiatus
TEXT-FIGURE 4 Stratigraphic range chart of radiolarians collected in 1993 from the San Francisco de Paula section. Radiolarian taxa arranged in approximate strati- graphic order of LO as suggested in Sanfilippo and Nigrini (1998a). LO of the six taxa marked in bold approximate the Paleocene/Eocene boundary. For stratigraphic position of the samples refer to text-figure 2 caption.
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
Interpretation of the data presented by E. Flores Albin (Appendix 1)
In addition to the interpretation based on our own data we pres- ent an explanation of the data given by E. Flores Albin (Appen- dix 1). (As noted previously, Flores Albin and his colleagues obtained samples from the San Francisco de Paula section shortly after the section was exposed. His assemblages are better preserved than those collected by us). In text-figure 5 we illustrate 35 radiolarian events recognized by Flores Albin (Ap- pendices 1, 2) within the San Francisco de Paula section. Spe- cies included in his range chart with last occurrences above the San Francisco de Paula section include: Lamptonium pennatum, Bekoma sp. cf. B. bidartensis, Spongodiscus americanus, Spongurus illepidus and Theocotylissa alpha. Species on his range chart with first occurrences below the section are: Bekoma campechensis, B. demissa, B. divaricata, Carpo- sphaera subbotinae, Hexacontium palaeocenicum, Lamp- tonium colymbus, L. incohatum, Lithomespilus mendosa, Orbula comitata, 0. discipulus, 0. ducalis and Theocosphaera larnacium. Taxa that have only single occurrences within the section and therefore are not included on the range chart are: Calocycloma ampulla, Giraffospyris lata, Lamptonium fabaeforme chaunothorax, Phormocyrtis striata exquisita, Phormocyrtis turgida, Spongodiscus quartus quartus, Stylo- sphaera goruna and Theocotylissa sp. cf. T fimbria. The strati- graphic position of these taxa within the San Francisco de Paula section are considered less important since their stratigraphic ranges are not restricted to the zones represented in the San Francisco de Paula section. However, their presence confirms the age assignment of the assemblage as a whole.
A comparison of the species ranges in the San Francisco de Paula section as observed by Flores Albin (text-figure 5) with those known from other sedimentary sequences (Foreman 1973; Sanfilippo and Nigrini 1998a) results in the following ob- servations. Most of the differences in the species ranges in the San Francisco de Paula section from those reported elsewhere are due to poor preservation.
(1) The LO of Theocotylissa auctor occurs much higher than the LO of Bekoma bidartensis in the San Francisco de Paula succession. It thus does not correspond to the LO of T auctor that is synchronous with the LO of B. bidartensis as postulated by Foreman (1973). However, it is consistent with the order of the events established by Sanfilippo and Nigrini (1998a);
(2) The evolutionary transition of Lamptonium pennatum to Lamptoniumfabaeforme fabaeforme, an evolutionary event oc- curring in the B. bidartensis Zone, cannot be determined in this section, where L. pennatum appears to have a much longer stratigraphic range than L. fabaeforme, contrary to evidence from many other localities;
(3) The HO of Rhopalocanium ornatum is older in this section than recorded elsewhere. It is known to occur throughout the Eocene in other sections worldwide;
(4) The HOs of Amphicraspedum murrayanum, A. prolixum and Eucyrtidium cubense (Phormocyrtis cubensis) are consid- ered low in this section. Globally these HOs are above the HO of Theocotylissa auctor (Sanfilippo and Nigrini 1998a);
(5) The LO of A. murrayanum is high; it is usually below the LO of Bekoma bidartensis: and.
(6) The LO of A. prolixum is above, rather than below (its usual position), the HO of B. campechensis.
Discussion of differences in zonal assignments
Although there is general agreement between our zonal assign- ments and those made by Flores Albin (Appendix 1), our strati- graphic interpretations differ in the following way (text-figures 2, 4, 5; Table 1):
Flores Albin, using Maurrasse' s zonation, recognized two radiolarian zones, the Eucyrtidium cubense and Bekoma bidartensis Zones. We are uncertain of the justification for his placement of the lower boundary of his Bekoma bidartensis Zone, which he places at the level of sample C23 (correspond- ing to our sample SFP-28). Sample C23 is from an approxi- mately 19 meter-thick part of the section in which the radiolarians are very rare and poorly preserved, or absent due to dissolution. We interpret this interval as a zone of uncertainty between the B. campechensis and B. bidartensis Zones.
Flores Albin (Appendix 1) assigned the upper part of the San Francisco de Paula section (his sample numbers C23-C62, our corresponding sample numbers SFP-28 to SFE-16) to the upper Paleocene/lower Eocene Bekoma bidartensis Zone. In contrast, based on the identification of Buryella clinata (in sample SFP-10) we divide the upper part of the section into two zones. We place the short interval (0.5 m) represented by our samples SFP-15 to SFP-11, just above a barren interval, in the B. bidartensis Zone, and the upper part of the section (our samples SFP-10 through SFE-16, see discussion above) in the lower Eocene Buryella clinata Zone. This age assignment is supported by calcareous nannofossil stratigraphy which places the interval between samples SFP-12 and SFE-16 in calcareous nannofossil Zone NP12 (Aubry 1999, this volume).
Correlation to foraminiferal zones
Four planktonic foraminiferal zones have been recognized in the San Francisco de Paula section (Femamndez-Rodriguez et al. 1999, this volume): Globonomalina pseudomenardii Total Range Zone (Zone P4), Morozovella velascoensis Interval Zone (Zone P5), Morozovella formosa formosalMorozovella lensiformis-Morozovella aragonensis Interval Subzone (Subzone P6b) and Morozovella aragonensis/Morozovella formosa formosa Concurrent Range Zone (Zone P7). According to initial correlations by E. Flores Albin and G. Fernafndez- Rodriguez (in Flores Albin, Appendix 1, and Fernandez- Rodriguez et al. 1999, this volume), in the San Francisco de Paula section, the Bekoma campechensis Zone (approximately equivalent to the informal Eucyrtidium cubense zone) can be correlated with the Globanomalina pseudomenardii Total Range (Zone P4) and the lower part of the Morozovella velascoensis Interval Zone (Zone P5). The Bekoma bidartensis Zone, as recognized by Flores Albin (Appendix 1), correlates with the remainder of the Morozovella velascoensis Interval Zone (Zone P5), the Morozovella formosa formosa/Morozo- vella lensiformis-Morozovella aragonensis Interval Subzone (Subzone P6b), an unzoned interval and the Morozovella aragonensis/Morozovella formosa formosa Concurrent Range Zone (Zone P7) (text-figure 2).
It should be noted that these correlations differ somewhat from those established by Maurrasse (1973), which were based on the tropical planktonic foraminiferal zonation as applied by authors working on the same DSDP samples as Maurrasse. He corre- lated the base of his Eucyrtidium cubense zone to the lower part
68
Micropaleontology, vol. 45, supplement 2, 1999
o eI RADIOLARIA
E y t a-v Datums from Florez-Abin, Appendix 1
C62 C61 C60 C59 C58 C57 3 C56 C55 6 C54 C53 C52
C49 , t C48 C47 EE?p
"
C46ttt 3F 1 C45 03 A cz2 ?g C44 T t . E i Q
c43 5 E , SZ C42
C39 0 C38 | S _ C37 ' <c C36 t C35 C34 C33 E C32 , C31 C C30 f C29 C28 C27 E c C26 2
O
C25E C24 ? E E C23 * .
C22 < 21 t
C20 8 *
C16 1) I:
c15 s
C14 0 i C13 . 0 C12 E C 1 2 I
C9 t C C8 I c- o C7 o C6 D 3 C5 LU
C04I C3 44 C12 c0
TEXT-FIGURE 5 Highest and lowest occurrences of radiolarian species recorded by E. Flores Albin (Appendix 1) in the San Francisco de Paula section. See text for dis- cussion.
69
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
of the Globorotalia (=Morozovella) velascoensis Zone, and the upper boundary to the lowermost part of the Globorotalia edgari Zone, which is not represented in Cuba. Further, Maurrasse correlated the base of his B. bidartensis Zone to the lower part of the Globorotalia edgari Zone, and the upper boundary to the upper part of the Globorotalia (=Morozovella) subbotinae/Globorotalia (=Morozovella) wilcoxensis Zone (and not higher into the Morozovella formosa formosa Zone).
Herein, we correlate the stratigraphic interval represented by the Bekoma campechensis radiolarian Zone with the Planorotalites pseudomenardii Total Range Zone (Zone P4). The short interval unequivocally representing the Bekoma bi- dartensis radiolarian Zone is correlated with the Morozovella formosa formosalMorozovella lensiformis-Morozovella aragon- ensis Interval Subzone (Subzone P6b). The interval between the B. campechensis and B. bidartensis Zones, represented by Zone P5 and part of Subzone P6b, cannot be assigned to either radiolarian zone. The uppermost part of the section, that is stratigraphically equivalent to the Buryella clinata Zone, is cor- related with the remainder of Subzone P6b, an unzoned interval and part of the Morozovella aragonensis/Morozovella formosa formosa Concurrent Range Zone (Zone P7).
Correlation with nannofossil zones
The following calcareous nannoplankton zones have been rec- ognized in the San Francisco de Paula section (Aubry 1999, this volume: NP6 in Sample SFP-34; NP10 in Samples SFP-13 through SFP-23; and NP12 in Samples SFE-16 through SFP-12 (Table 1, text-figures 2, 4). On the basis of calcareous nanno- fossil evidence, there is an unconformity between SFP-12 and SFP-13 (a 5 cm stratigraphic interval just below the previously identified Paleocene/Eocene boundary by Cuban geologists). In this part of the succession, calcareous nannofossils assemblages characteristic of Zone NP12 occur directly above assemblages characteristic of Subzone NPlOd. A second possible uncon- formity lies between Samples SFP-24 and SFP-25. Samples collected from other intervals within the section cannot be firmly dated on the basis of nannofossil biostratigraphy.
Sample SFP-34 has been assigned to Zone NP6, which corre- lates with the Bekoma campechensis radiolarian Zone. Samples SFP-13 through SFP-15, in the Bekoma bidartensis Zone, cor- relate with the uppermost part of Zone NP10 (Subzone NPlOd). Samples SFE-16 through SFP-12 in the Buryella clinata Zone correlate with Zone NP12.
Placement of the Paleocene/Eocene boundary in the San Francisco de Paula Section
As noted earlier, the Paleocene/Eocene boundary in this succes- sion is between samples SFP-22 and -21, and coincides with the NPlOa/NPlOd unconformable zonal contact (Aubry 1999, this volume). The HO of Morozovella velascoensis in the San Fran- cisco de Paula section coincides with an unconformity, and the Bekoma bidartensis radiolarian Zone extends above it. The LO of Buryella clinata, which marks the top of the Bekoma bidartensis Zone, is placed between samples SFP-10 and SFP-11 in Zone NP12. This supports the observations made by Sanfilippo and Nigrini (1998a) who determined that the LO of Buryella clinata at DSDP Site 313, and thus the uppermost part of the Bekoma bidartensis Zone are in Zone NP12 (= Zone CP10). The six radiolarian events used by Sanfilippo and Nigrini (1998a) to approximate the Paleocene/Eocene boundary occur in the San Francisco de Paula section although their pres- ence is somewhat patchy. Our data here show that the LOs of
Calocycloma castum, Lamptonium fabaeforme fabaeforme, Phormocyrtis turgida and Theocotylissa auctor are younger than the Paleocene/Eocene boundary (about 0.4 my younger than the boundary, based on the calcareous nannoplankton biochronology of Aubry et al. 1996). The LO's of Giraffospyris lata and Podocyrtis (Podocyrtis) papalis support placement of the boundary below sample SFP-15.
In Berggren et al. (1985), the Paleocene/Eocene boundary was placed at the boundary between calcareous nannofossil Zones NP9 and NP10. Detailed work by Berggren et al. (1998) and Aubry (1998) recalibrated this boundary and raised it into the middle part of Zone NP10. Although Zone NP10 has been rec- ognized in the San Francisco de Paula section, its upper bound- ary is unconformable and its lower boundary is uncertain (text-figures 2, 4 and Table 1). Thus, the placement of the Paleocene/Eocene boundary in this section on the basis of nannofossils is uncertain.
Based on previous studies, the Paleocene/Eocene boundary is known to fall within the Bekoma bidartensis radiolarian Zone. In the San Francisco de Paula section it can only be recognized in a short interval extending from Samples SFP-15 through SFP- 11. The lower extension of the Bekoma bidartensis Zone in the San Francisco de Paula section is uncertain due to poor pres- ervation and barren intervals. The recognition of its upper boundary is less uncertain, but still hampered by poor preserva- tion. Thus, the Paleocene/Eocene boundary cannot be deter- mined in this section on the basis of radiolarians.
CONCLUSIONS
(1) The San Francisco de Paula section in western Cuba con- tains three radiolarian zones for the upper Paleocene and lower Eocene: Bekoma campechensis (informally Eucyrtidium cubense zone), Bekoma bidartensis and Buryella clinata Zones;
(2) Radiolarian abundance varies from rare to abundant, diver- sity is low, and preservation is poor. This is likely due in part to weathering;
(3) Thirty-five datum events are noted within the section. Sev- eral of these datums are higher or lower than expected when compared with the known global ranges of these species; and
(4) Based on biostratigraphic data collected in this study, the San Francisco de Paula section is undesirable as a stratotype for the Paleocene/Eocene boundary.
ACKNOWLEDGMENTS
The authors would like to thank Richard H. Fluegeman, Ball State University, and the Center for Tropical Conservation for their support in this project. Partial funding has been provided by UNESCO. We thank Joseph J. Morley for his help in acquir- ing literature necessary for the completion of this paper, Florentin J.M.R. Maurrasse for helpful discussions, and John A. Barron for identification of diatoms. We would also like to thank Marie Pierre Aubry, William A. Berggren and Christo- pher J. Hollis who reviewed earlier versions of this article and offered many suggestions for its improvement. Special thanks to Kathryn L. Meyerhoff for drafting the locality map. This pa- per is a contribution to IGCP Project #308, Paleocene/Eocene Boundary Events in Space and Time. This work was supported in part by NSF grant number OCE91-23712 (to A. Sanfilippo).
70
Micropaleontology, vol. 45, supplement 2, 1999
TABLE 1 Radiolarian occurrences in samples collected in 1993 in the San Francisco de Paula section. LOs of the six radiolarian taxa, indicated in bold in the head- ing of the table, approximate the Paleocene/Eocene boundary. A=abundant; C=common; F=few; R=rare; VR=very rare; "+"=present; B=barren; "?"=questionable identification. M=moderate; P=poor; VP=very poor; D=dissolved and unrecognizable radiolarians. Abundances marked with an as- terisk in the Bekoma spp. column indicate probable presence of Bekoma bidartensis. For stratigraphic position of the samples refer to text-figure 2 cap- tion.
71
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
REFERENCES
AUBRY, M.-P., 1995. From chronology to stratigraphy: Interpreting the lower and middle Eocene stratigraphic record in the Atlantic Ocean. In Berggren, W. A., Kent, D. V., Aubry, M.-P. and Hardenbol, J. (Eds.), Geochronology, time scales and global stratigraphic correla- tion. SEPM (Society for Sedimentary Geology), Publication N? 54:213-274.
, 1998. Stratigraphic (dis)continuity and temporal resolution of geological events in the upper Paleocene-lower Eocene deep-sea re- cord. In Aubry, M.-P., Lucas, S. and Berggren, W. A. (Eds.), Late Paleocene - Early Eocene climatic and biotic evolution. New York: Columbia University Press, 37-66.
, 1999. Late Paleocene-Early Eocene Sedimentary History in Western Cuba: Implications for the LPTM and for Regional Tectonic History. In Fluegeman, R., and Aubry, M.-P., (Eds.), Lower Paleogene Biostratigraphy of Cuba. Micropaleontology, volume 45, supplement 2, pp. 5-18.
AUBRY, M.-P., BERGGREN, W. A., KENT, D. V., FLYNN, J. J., KLITGORD, K. D., OBRADOVICH, J. D. and PROTHERO, R. R., 1988. Paleogene geochronology: An integrated approach. Paleoceanography, 3:707-742.
AUBRY, M.-P., BERGGREN, W. A., STOTT, L. D. and SINHA, A., 1996. The upper Paleocene-lower Eocene stratigraphic record and the Paleocene/Eocene boundary carbon isotope excursion. In: Knox, R. W. O'B., Corfield, R. and Dunnay, R. E. (Eds.), Correlation of the Early Paleogene in Northwest Europe. London, Geological Society of London Special Publication N? 101:353-380.
BERGGREN, W. A., and AUBRY, M.-P., 1996. A late Paleocene-early Eocene NW European and North Sea magnetobiostratigraphic corre- lation network: A sequence stratigraphic framework. In: Knox, R.W. O'B., Corfield, R. and Dunnay, R.E. (Eds.), Correlation of the Early Paleogene in Northwest Europe. London, Geological Society of London Special Publication N? 101:309-352.
, 1998. The Paleocene/Eocene Epoch/Series boundary: Chronostratigraphic framework and estimated geochronology. In:
Aubry, M.-P., Lucas, S. and Berggren, W. A. (Eds.), Late Paleocene - Early Eocene climatic and biotic evolution. New York: Columbia University Press, 18-36.
BERGGREN, W. A., and MILLER, K. G., 1988. Paleogene tropical planktonic foraminiferal biostratigraphy and magnetobio- chronology. Micropaleontology, 34:362-380.
BERGGREN, W. A., KENT, D. V. and FLYNN, J. J., 1985. Paleogene geochronology and chronostratigraphy. In: N. J. Snelling (Ed.), The chronology of the geological record. London, Geological Society of London Memoir N? 10:141-195.
BERGGREN, W. A., KENT, D. V., SWISHER, C. C. and AUBRY, M.-P., 1995. A revised Cenozoic geochronology and chronostrati- graphy. In: Berggren, W. A., Kent, D. V. and Hardenbol, J. (Eds.), Geochronology, Time Scales and Global Stratigraphic Correlations: A Unified Temporal Framework for an Historical Geology. Society of Economic Paleontologists and Mineralogists Special No. 54:129-212.
BORISENKO, N. N., 1958. Radiolyarii paleotsena zapadnoi Kubani (Paleocene Radiolaria of Western Kubanj): Trudy Vsesoyuznyi Neftegazovyi Nauchno-Issledovatelskii Institut (VNII), Krasno- darskii Filial, N0. 17:81-100.
BRALOWER, T. J., AND ITURALDE-VINENT, M. A., 1997. Micropaleontological dating of the collision between the North American Plate and the Greater Antilles Arc in western Cuba. Palaios, 12:133-150.
BRANDT, R., 1935. Die Mikropalaeontologie des Heiligenhafener, Kieseltones (Ober Eozan) Radiolarien; Systematik. In: Wetzel, E. 0. (Ed.), Jahresbericht des Niedersachsischen geologischen Vereins. Hannover, Germany, 27:48-59.
BRONNIMANN, P. and RIGASSI, D., 1963. Contribution to the geol- ogy and paleontology of the area of the City of La Habana and its sur- roundings. Eclogae Geologica Helvetiae, 56(7): 192-480.
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
BUKRY, D., 1973a. Coccolith stratigraphy, Leg 10 - Deep Sea Drilling Project. In: Worzel, J. L. Bryant, W. et al. (Eds.), Initial Reports of the Deep Sea Drilling Project, Volume 10: 385-406. Washington, D.C.: U.S. Government Printing Office.
BUKRY, D., 1973b. Low latitude coccolith biostratigraphic zonation. In: Edgar, N. T., Saunders, J. B. et al. (Eds.), Initial reports of the Deep Sea Drilling Project, 15: 685-703. Washington, D.C.: U.S. Government Printing Office.
BUKRY, D., 1975. Coccolith and silicoflagellate stratigraphy, north- western Pacific Ocean, Deep Sea Drilling Project Leg 32. In: Larson, R. L., Moberly, R. et al. (Eds.), Initial reports of the Deep Sea Drilling Project, 32: 685-677. Washington, D.C.: U.S. Government Printing Office.
BURNS, R. E., ANDREWS, J. E. et al., 1973. Initial Reports of the Deep Sea Drilling Project. In: Bums, R. E., Andrews, J. E. et al. (Eds.), Ini- tial reports of the Deep Sea Drilling Project, 21: 1-931. Washington, D.C.: U.S. Government Printing Office.
CANDE, S. C. and KENT, D. V., 1992. Anew geomagnetic polarity time scale for the Late Cretaceous and Cenozoic. Journal of Geophysical Research, 97(B10):13917-13951.
CANDE, S. C. and KENT, D. V., 1995. Revised calibration of the geo- magnetic polarity time scale for the late Cretaceous and Cenozoic. Journal of Geophysical Research, 100:6093-6095.
CLARK, B. L. and CAMPBELL, A. S., 1942. Eocene radiolarian faunas from the Monte Diablo area, California. Geological Society of America, Special Paper 39:1-112, 9 pl.
DUMITRICA, P., 1973. Paleocene Radiolaria, DSDPLeg 21. In: Bums, R. E., Andrews, J. E. et al. (Eds.), Initial reports of the Deep Sea Drilling Project, 21: 787-817. Washington, D.C.: U.S. Government Printing Office.
EHRENBERG, C. G., 1847. Uber die mikroskopischen kieselschaligen Polycystinen als miichtige Gebirgsmasse von Barbados und iiber das Verhaltniss der aus mehr als 300 neuen Arten bestehenden ganz
eigenthimlichen Formengruppe jener Felsmasse zu den jetzt lebenden Thieren und zur Kreidebildung. Eine neue Anregung zur Erforschung des Erdlebens. K6nigliche Preussische Akademie der Wissenschaften zu Berlin, Bericht, Jahre 1847:40-60.
, 1854. Mikrogeologie. Das Erden und Felsen schaffende Wirken des unsichtbar kleinen selbststiindigen Lebens auf der Erde. Leopold Voss, Leipzig, Germany, p. xxviii + 374 p., Atlas, 31 p., 41 pl.
--- , 1873. Gr6ssere Felsproben des Polycystinen-Mergels von Bar- bados mit weiteren Erliuterungen. K6nigliche Preussische Akademie der Wissenschaften zu Berlin, Monatsberichte, Jahre 1873:213-263.
, 1875. Fortsetzung der mikrogeologischen Studien als Gesammt-Uebersicht der mikroskopischen Palaontologie gleichartig analysirter Gebirgsarten der Erde, mit specieller Rticksicht auf den Polycystinen-Mergel von Barbados. K6nigliche Akademie der Wissenschaften zu Berlin, Abhandlungen, Jahre 1875:1-225, 30 pl.
FERNANDEZ-RODRIGUEZ, G., BLANCO-BUSTAMENTE, S., and FLUEGEMAN, R.H., 1999. Paleocene-Eocene planktonic foraminiferal biostratigraphy of western Cuba. In: Fluegeman, R., and Aubry, M.-P., (Eds.), Lower Paleogene Biostratigraphy of Cuba. Micropaleontology, 45, supplement 2, pp. 27-42.
FLORES ALBIN, E., 1983. Radiolarios de algunas formaciones del Cretacico Paleogeno Inferior de Cuba occidental (Radiolarians from Cretaceous and lower Paleogene formations in western Cuba). Ciencias de la Tierra y del Espacio, 7:3-36.
FLORES ALBIN, E. and FERNANDEZ-RODRIGUEZ, G., 1985. Acereca de la bioestratigrafia del limite entre los depositos del paleoceno-eoceno inferior (formaciones Alkazar y Capdevila) en la provincia Ciudad Habana (Biostratigraphy of the limit between Paleocene - Lower Eocene sediments (Alkazar and Capdevila forma- tions) in Havana City Province). Revista Tecnologica, 15, Serie: Geologia, N0 1:8-18.
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
FLUEGEMAN, R. H., 1999. Preliminary study of the benthic foraminifera of the San Francisco de Paula section (upper Paleocene-lower Eocene), Ciudad de la Habana, Cuba. In: Fluegeman, R., and Aubry, M.-P., (Eds.), Lower Paleogene Biostratigraphy of Cuba, Micropaleontology, volume 45, supple- ment 2, pp. 43-56.
FOREMAN, H. P., 1973. Radiolaria of Leg 10 with systematics and ranges for the families Amphipyndacidae, Artostrobiidae, and Theoperida. In: Worzel, J. L. Bryant, W. et al. (Eds.), Initial Reports of the Deep Sea Drilling Project, 10: 407-474. Washington, D.C.: U. S. Government Printing Office.
, 1975. Radiolaria from the North Pacific, Deep Sea Drilling Project, Leg 32. In: Larson, R. L., Moberly, R. et al. (Eds.), Initial re- ports of the Deep Sea Drilling Project, 32: 579-676. Washington, D.C.: U.S. Government Printing Office.
GOLL, R. M., 1969. Classification and phylogeny of Cenozoic Trissocyclidae (Radiolaria) in the Pacific and Caribbean basins. Part II. Journal of Paleontology, 43(2):322-339.
HAECKEL, E., 1881. Entwurf eines Radiolarien-Systems auf Grund von Studien der Challenger-Radiolarien. Jena. Z. Med. Naturwiss. 15 (new ser., 8)(3):1-418p.
, 1887. Report on the Radiolaria collected by H.M.S. Chal- lenger during the years 1873-1876. Report on the Scientific Results of the Voyage of the H.M.S. Challenger, Zoology, v. 18, 2 parts, At- las, clxxxviii + 1803 p., 140 pl.
HOLLIS, C. J., 1993. Latest Cretaceous to Late Paleocene radiolarian biostratigraphy: A new zonation from the New Zealand region. Ma- rine Micropaleontology, 21(4): 295-327.
, 1997. Cretaceous-Paleocene Radiolaria from eastern Marlborough, New Zealand. Institute of Geological and Nuclear Sci- ences, Monograph 17:1-152
KENNETT, J. P. and STOTT, L. D., 1991. Abrupt deep-sea warming, paleoceanographic changes and benthic extinctions at the end of the Palaeocene. Nature, 353:225-229.
KOZLOVA, G. E., 1983. Rasprostraneniye radiolyariyevykh zon Atlantiki v paleogene Povolzh'ya (Distribution of radiolarian zones of the Atlantic in the Paleogene of the Volga region). Akademiya Nauk SSSR, Seriya Geologicheskaya (Proceedings of the USSR Academy of Sciences, Geological Series), 3:46-51.
1984. Zonalnoe podrazdelenie borealnogo paleogena po radiolyariyam (Zonal subdivision of the boreal Paleogene by radio- larians). In: Petrushevskaya, M. G. and Stepanjants, S. D. (Eds.), Morfologiya, ekologiya i evolutsiya Radiolyarii. Materialy IV simpoziuma evropeiskikh radiolyaristov EURORAD IV (Morphol- ogy, ecology, and evolution of radiolarians. Material from the IV symposium of European radiolarists EURORAD IV): 196-210. Le- ningrad, USSR: Akademiya Nauk SSSR, Zoological Institute.
1993. Radiolarian zonal scale of the boreal Paleogene. In: Blueford, J. R. and Murchey, B. L. (Eds.), Radiolaria of giant and subgiant fields in Asia. Nazarov Memorial Volume. Micropaleontology Special Publication N? 6:90-93. New York, USA: Micropaleontology Press, American Museum of Natural His- tory.
KOZLOVA, G. E. and Gorbovets, A. N., 1966. Radiolyarii verkhnemelovykh i verkhneeotsenovykh otlozhenii Zapadno-Sibirskoi nizmennosti (Radiolarians of the Upper Creta- ceous and Upper Eocene deposits of the West Siberian Lowland). Trudy vsesoyuznogo neftyanogo nauchno-issledovatelskogo geologorazvedochnogo instituta (VNIGRI) (Proceedings of the All Union Petroleum Scientific Research Institute for Geological Survey (VNIGRI)), N? 248:1-159.
KRASHENINNIKOV, V. A., 1960. Nekotorye Radiolyarii Nizhnego i Srednego Eotsena Zapadnogo Predkavkazya (Some radiolarians of the Lower and Middle Eocene of the Western Pre-Caucasus). Mineralogicko-Geologicka i Okhrana Nedr SSSR Vsesoyuznogo Nauchno-Issledovatelskogo Geologorazved Neftyanogo Instituta, 16:271-308.
, 1969. Geographical and stratigraphical distribution of plank- tonic foraminifers in Paleogene deposits of tropical and subtropical areas. Akademy Nauk SSSR Geol. Inst. Trudy, 202:1-190.
PLATE 3 Scanning electron micrographs of radiolarians collected from the San Francisco de Paula section by Emilio Flores Albin
(Appendix 1). Sample locations unknown. Identifications by A. Sanfilippo; photographs were taken by D. Hull at the University of Texas at Dallas. Scale (upper right) = number of microns cited for each figure.
LEWIS, J. F., and DRAPER, G., 1990. Geology and tectonic evolution of the northern Caribbean margin. In: Dengo, G., and Case, J. E., (Eds.), The Caribbean region. Geological Society of America, The Geology of North America, H:77-140.
MARTINI, E., 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation. In: Farinacci, A. (Ed.) Proceedings of the Second Planktonic Conference, Roma, 1970: Roma, Tecnoscienza, 2:739-785.
MAURRASSE, F. J. M. R., 1973. Biostratigraphy, paleoecology, biofacies variations of middle Paleogene sediments in the Caribbean Deep-Sea. Ph.D. dissertation, Columbia University: 1-484
, 1976. Paleoecologic and paleoclimatic implications of radiolarian facies in Caribbean Paleogene deep-sea sediments. In: Causse, R. (Ed.), Transactions de la VII Conference Geologique des Caraibes du 30 juin au 12 juillet 1974, St. Francois, Guadeloupe, pp. 185-204.
MCNEELY, B. W., 1973. Biostratigraphy of the Mesozoic and Paleocene pelagic sediments of the Campeche Embankment area. In: Worzel, J. L. Bryant, W. et al. (Eds.), Initial Reports of the Deep Sea Drilling Project, 10: 679-695. Washington, D.C.: U.S. Government Printing Office.
MILLER, K. G., JANECEK, T. R., KATZ, M. E. and KEIL, D. J., 1987. Abyssal circulation and benthic foraminiferal changes near the Paleocene/Eocene boundary. Paleoceanography, 2:741-761.
NISHIMURA, A., 1987. Cenozoic Radiolaria in the western North At- lantic, Site 603, Leg 93 of the Deep Sea Drilling Project. In: van Hinte, J. E., Wise, S. W., Jr. et al. (Eds.), Initial reports of the Deep Sea Drilling Project, 93: 713-737. Washington, D.C.: U.S. Govern- ment Printing Office.
, 1992. Paleocene radiolarian biostratigraphy in the northwest Atlantic at Site 384, Leg 43, of the Deep Sea Drilling Project. Micropaleontology, 38(4):317-362.
OKADA, H. and BUKRY, D., 1980. Supplementary modification and introduction of code numbers to the low-latitude coccolith biostratigraphic zonation (Bukry, 1973: 1975). Marine Micropaleontology, 5:321-325.
OKADA, H. and THIERSTEIN, H. R., 1979. Calcareous nannoplankton - Leg 43, Deep Sea Drilling Project. In: Tucholke, B. E., Vogt, P. R. et al. (Eds.), Initial reports of the Deep Sea Drilling Project, 43: 507-573. Washington, D.C.: U.S. Government Printing Office.
PETRUSHEVSKAYA, M. G., 1977. Novye vidy radiolayarii otryada Nassellaria (New species of the radiolarians of the order Nassellaria). Issledovaniya fauny moreii - Novye vidy i rody morskikh bespozvonochnykh (New species and genera of marine in- vertebrates). Zool. Inst. Acad. Sci. USSR, Leningrad, 21(24): 10-19.
RIEDEL, W. R. and SANFILIPPO, A., 1970. Radiolaria, Leg 4, Deep Sea Drilling Project. In: Bader, R. G., Gerard, R. D. et al. (Eds.), Ini- tial Reports of the Deep Sea Drilling Project, 4: 503-574. Washing- ton, D.C.: U. S. Government Printing Office.
, 1971. Cenozoic Radiolaria from the western tropical Pacific, Leg 7. In: Winterer, E. L., Riedel, W. R. et al. (Eds.), Initial Reports of
the Deep Sea Drilling Project, 7:1529-1672. Washington, D.C.: U. S. Government Printing Office.
, 1973. Cenozoic Radiolaria from the Caribbean, Deep Sea Drilling Project, Leg 15. In: Edgar, N. T., Saunders, J. B. et al. (Eds.), Initial reports of the Deep Sea Drilling Project, 15: 705-751. Wash- ington, D.C.: U.S. Government Printing Office.
, 1978. Stratigraphy and evolution of tropical Cenozoic radiolar- ians. Micropaleontology, 23(1):61-96.
SANFILIPPO, A. and NIGRINI, C. A., 1998a. Upper Paleocene - lower Eocene deep-sea radiolarian stratigraphy and the Paleocene/Eocene series boundary. In: Aubry, M.-P., Lucas, S. G. and Berggren, W. A. (Eds.), Late Paleocene - early Eocene climatic and biotic evolution. New York: Columbia University Press, 244-276.
, 1998b. Code numbers for Cenozoic low latitude radiolarian biostratigraphic zones and GPTS conversion tables. Marine Micropaleontology, 33:109-156.
SANFILIPPO, A. and RIEDEL, W. R., 1973. Cenozoic Radiolaria (ex- clusive of theoperids, artostrobiids and amphipyndacids) from the Gulf of Mexico, DSDP Leg 10. In: Worzel, J. L., Bryant, W., et al. (Eds.), Initial Reports of the Deep Sea Drilling Project, 10: 475-611. Washington, D.C.: U.S. Government Printing Office.
,1976. Radiolarian occurrences in the Caribbean Region. Publi- cation de la VII conference geologique des Caraibes du 30 juin au 12 juillet 1974 (VII Confdrence G6ologique des Caraibes, Cayenne, Departement Franqais de la Guyane), 145-168.
, 1982. Revision of the radiolarian genera Theocotyle, Theoco- tylissa and Thyrsocyrtis. Micropaleontology, 28(2): 170-188.
,1992. The origin and evolution of Pterocorythidae (Radiolaria): A Cenozoic phylogenetic study. Micropaleontology, 38(1): 1-36.
SANFILIPPO, A., WESTBERG-SMITH, M. J. and RIEDEL, W. R., 1985. Cenozoic Radiolaria. In: Bolli, H. M., Perch-Nielsen, K. and Saunders, J. B. (Eds.), Plankton stratigraphy. Cambridge, UK: Cam- bridge University Press, 631-712.
STOTT, L. D. and KENNETT, J. P., 1990. Antarctic Paleogene plank- tonic foraminifer biostratigraphy: ODP Leg 113, Sites 689-690. In: Barker, P. F., Kennett, J. P. et al. (Eds.), Proceedings of the Ocean Drilling Program, Scientific Results, 113: 849-863. College Station, TX: Ocean Drilling Program.
STRELNIKOVA, N. I., 1987. Paleogenovye diatomovye vodorosli vysokikh shirot severnogo i yuzhnogo polusharii. (Paleogene dia- toms of the high latitudes of the northern and southern hemispheres). Ministry of High and Middle specialist organization, Leningrad, Or- der of Lenin and Order of Red Workers State University. Doctoral thesis in biological sciences, Leningrad.
STRONG, C. P., 1977. Cretaceous-Tertiary boundary at Woodside Creek, north-eastern Marlborough. New Zealand Journal of Geology and Geophysics, 20:687-696.
,1981. Cretaceous-Tertiary boundary at Woodside Creek revis- ited. Geological Society of New Zealand, Newsletter, 51:4-5.
, 1984. Cretaceous-Tertiary boundary at Chancet Rocks Scien- tific Reserve, Northeast Marlborough. New Zealand Geological Sur- vey, Rec., 3:47-51.
STRONG, C. P., BROOKS, R. R., WILSON, S. M., REEVES, R. D., ORTH, C. J., MAO, X.-Y., QUINTANA, L. R. and ANDERS, E., 1987. A new Cretaceous-Tertiary boundary site at Flaxbourne River, New Zealand: biostratigraphy and geochemistry. Geochemica Cosmochemica Acta, 51:2769-2777.
78
Micropaleontology, vol. 45, supplement 2, 1999
STRONG, C. P., HOLLIS, C. J. and WILSON, G. J., 1995. Foraminiferal, radiolarian and dinoflagellate biostratigraphy of Late Cretaceous to Middle Eocene pelagic sediments (Muzzle Group), Mead Stream, Marlborough, New Zealand. New Zealand Journal of Geology and Geophysics, 38:171-212.
SWISHER, C. C. III and KNOX, R. W. O'B., 1991. The age of the Paleocene/Eocene boundary: 40Ar/39Ar dating of the lower part of NP10, North Sea Basin and Denmark. In: IGCP Project 308 (Paleocene/Eocene boundary events), International Annual Meeting and Field Conference, Brussels, 2-6 December 1991, Abstracts with Program, 16.
THOMAS, E., 1990a. Late Cretaceous-early Eocene mass extinctions in the deep sea. In: Sharpton, V. L. and Wards, P. (Eds.), Global catas- trophes. Geological Society of America, Special Paper N? 247:481-495.
, 1990b. Late Cretaceous through Neogene deep-sea benthic foraminifers (Maud Rise, Weddell Sea, Antarctica). In: Baker, P. F., Kennett, J. P. et al. (Eds.), Proceedings of the Ocean Drilling Pro- gram, Scientific Results, 113:571-594. College Station, TX: Ocean Drilling Program.
,1993. Cenozoic deep sea circulation: evidence from deep sea benthic foraminifera. In: Kennett, J. P. and Warnke, D. (Eds.), The Antarctic paleoenvironment: A perspective on global change. AGU Antarctic Research Series N? 56:141-165.
TUCHOLKE, B. E., VOGT, P. R. et al., 1979. Site 384: The Creta- ceous/Tertiary boundary, Aptian reefs, and the J-Anomaly Ridge. In: Tucholke, B. E., Vogt, P. R. et al. (Eds.), Initial Reports of the Deep Sea Drilling Project, 43: 107-132. Washington, D.C.: U.S. Govern- ment Printing Office.
APPENDIX 1
Biostratigraphical Analysis of Planktonic Foraminifera and Radiolaria at the Paleocene/Eocene Boundary in the San Francisco de Paula sec- tion, Western Cuba, contributed by Emilio Flores Albin. [ Data shown on text-figures 2, 5; Plate 3.]
Two radiolarian zones were recognized in early studies of the San Francisco de Paula section: the Eucyrtidium cubense zone and the Bekoma bidartensis Zone (upper Paleocene-lower Eocene; text-figures 2, 3, 5). The Eucyrtidium cubense zone was proposed as an informal partial range zone by Maurrasse (1973, unpublished thesis) for the Caribbean region, the Atlan- tic and part of the Pacific Ocean. Its base is defined by the earli- est morphotypic appearance of E. cubense. The top of this zone is recognized by the first occurrence of Bekoma bidartensis, which is in the upper Paleocene. The Bekoma bidartensis Zone was defined by Foreman (1973). Its base is recognized by the earliest morphotypic appearance of Bekoma bidartensis; the top, which has not been recognized in the San Francisco de Paula section, is placed at the evolutionary bottom of the Buryella clinata interval-chronozone (Riedel and Sanfilippo 1978).
Paleocene-Eocene radiolarian assemblages in the San Francisco de Paula section are characterized by low species diversity and variable abundance. However, the number of diagnostic species is greater in this section than in other Cuban sections of Paleocene and Eocene age (Flores Albin and Fernandez- Rodriguez 1985). Some of these species are listed below, and their ranges in the San Francisco de Paula section shown on text-figure 5. Some forms are illustrated on Plate 3. [For a com- plete species list, additional photographs and taxonomic refer- ences: see Appendix 2 and Plates 1, 2].
The most common radiolarians recognized in the Paleocene as- semblages by this author (EFA) are: Bekoma campechensis Foreman Bekoma(?) demissa Foreman Bekoma divaricata Foreman Clathrocycloma capitaneum Foreman Diploplegma somphum Sanfilippo and Riedel Hexacontium palaeocenicum Sanfilippo and Riedel Lamptonium(?) colymbus Foreman Lamptonium(?) incohatum Foreman Lithomespilus mendosa (Krasheninnikov) Orbula comitata Foreman Orbula discipulus Foreman Orbula ducalis Foreman
Paleocene species which extend into the lower and middle Eocene and are present in the succession include: Bekoma bidartensis Riedel and Sanfilippo Eucyrtidium cubense Riedel and Sanfilippo [= Phormocyrtis
cubensis (Riedel and Sanfilippo) Foreman] Lamptonium pennatum Foreman Phormocyrtis striata exquisita (Kozlova) Spongodiscus quartus quartus (Borisenko) Stylosphaera goruna Sanfilippo and Riedel Theocotylissa auctor Foreman
Species which appear for the first time in lower Eocene sedi- ments are: Lamptoniumfabaeforme chaunothorax Riedel and Sanfilippo Lamptonium fabaeformefabaeforme (Krasheninnikov) Theocotylissa alpha Foreman Theocotylissa fimbria Foreman
Four planktonic foraminiferal zones are recognized in the San Francisco de Paula section: Globanomalina pseudomenardii, Morozovella velascoensis, Morozovella subbotinae and Moro- zovella formosa formosa Zones (Fernandez-Rodriguez et al. 1999, this volume). In the San Francisco de Paula section, the Eucyrtidium cubense zone can be correlated with the Globa- nomalina pseudomenardii Total Range Zone (Zone P4) and the lower part of the Morozovella velascoensis Interval Zone (Zone P5). The Bekoma bidartensis Zone correlates with the remain- der of the Morozovella velascoensis Zone (Zone P5), the Morozovella formosa formosalMorozovella lensiformis- Morozovella aragonensis Interval Subzone (Subzone P6b), an unzoned interval and the Morozovella aragonensis/Morozovella formosa formosa Concurrent Range Zone (Zone P7)
79
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
APPENDIX 2 Species List (Species indicated by an asterisk are from Flores Albin's tabulation, text-figure 5).
*Amphicraspedum murrayanum Haeckel 1887, p. 523, pl. 44, fig. 10; Sanfilippo and Riedel 1973, p. 524, pl. 10, figs. 3-6, pl. 28, fig. 1. [Plate 1, figure 2]
*Amphicraspedum prolixum Sanfilippo and Riedel 1973, p. 524, pl. 10, figs. 7-11, pl. 28, figs. 3, 4.
Bekoma bidartensis Riedel and Sanfilippo 1971, p. 1592, pl. 7, figs. 1-7; Foreman 1973, p. 432, pl. 3, figs. 20-21, pl.10, fig. 6; Sanfilippo et al. 1985, p. 667, figs. 13.1a, b; Nishimura 1992, p. 332, pl. 5, figs. 8, 9.
*Bekoma sp. cf. B. bidartensis. We do not know how Flores Albin' s concept differs from the original description.
*Bekoma campechensis Foreman 1973, p. 432, pl. 3, fig. 24, pl. 10, figs. 1-2, 4; Nishimura 1992, p. 332, pl. 5, figs. 1-4, pl. 13, fig. 1. [Platel, figures. 12, 13]. Not a single specimen belonging to the genus Bekoma was observed with feet. In spite of this we have made an attempt to distinguish B. campechensis from other co-occurring taxa of the genus Bekoma by using the char- acteristic shape of the cephalis for each taxon. Bekoma campechensis has a long dome-shaped to rectangular cephalis, while B. demissa has a cephalis with numerous pores that appears slightly depressed into the thorax. The cephalis and proximal elongated part of the thorax in B. campechensis are poreless with a thick hyaline wall, while in B. bidartensis this feature is not so prominent. In B. divaricata the length of the cephalis and thorax, and the poreless part of the thorax is markedly shorter.
*Bekoma(?) demissa Foreman 1973, p. 432, pl. 3, fig. 22, pl. 10, figs. 5, 6. See comment for B. campechensis above. [Plate 1, figure 11; Plate 3, figure 14]
*Bekoma divaricata Foreman 1973, p. 433, pl. 3, fig. 23, pl. 10, figs. 3, 4.
*Bekoma sp. A. This species is from Flores Albin' s original study. We have no further information.
Bekoma spp. In this category we have included two-segmented forms with typical Bekoma-type cephalis and traces of three feet that are circular in cross section that are incom- plete due to poor preservation. [Plate 1, figures 14, 16, 17]
Buryella clinata Foreman 1973, p. 433, pl. 8, figs. 1-3, pl. 9, fig. 19. Foreman 1975, p. 620, pl. 9, figs. 35-36; Sanfilippo et al. 1985, p. 668, figs. 14. la, b. [Plate 2, fig- ures 14, 15]. Due to the poor preservation we are unable to observe the characteristic diagonal pore alignment on the third segment, and we have therefore doubtfully as- signed the illustrated specimens to Buryella clinata. Rem- nants of the broad-based, ridged horn have been observed.
Buryella ?foremanae Petrushevskaya 1977, p. 16, pl. 3d, e; Hollis 1993, p. 322, PI. I, fig. 12.
Buryella pentadica Foreman 1973, p. 433, pl. 8, fig. 8, pl. 9, figs. 15-16. [Plate 1, figures 9, 10]
Buryella tetradica Foreman 1973, p. 433, pl. 8, figs .4-5, pl. 9, figs. 13-14; Sanfilippo et al. 1985, p. 668, figs. 14.3a, b. [Plate 1, figures 19, 20; Plate 3, figure 3]
*Calocycloma ampulla Ehrenberg 1854, pl. 36, figs. 15a-c; 1873, p. 225; 1875, p. 70, pl. 10, figs. 11-12a, b; Foreman 1973, p. 434, pl. 1, figs. 1-5, pl. 9, fig. 20.
Calocycloma castum (Haeckel). Calocyclas casta Haeckel 1887, p. 1384, pl. 73, fig. 10.
Calocycloma castum (Haeckel) Foreman 1973, p. 434, pl. 1, figs. 7, 9, 10; Sanfilippo et al. 1985, p. 669, fig. 14.4. [Plate 1, figure 21]
*Carposphaera subbotinae (Borisenko). Cenosphaera subbotinae Borisenko 1958, p. 85, pl. 23, figs. 4, 5; Sanfilippo and Riedel 1973, p. 490, pl. 4, fig. 3, pl. 23, figs. 4, 5.
*Clathrocycloma capitaneum Foreman 1973, p. 434, pl. 2, fig. 15, pl. 11, fig. 11. [Plate 3, figure 13]
*Diploplegma somphum Sanfilippo and Riedel 1973, p. 491, pl. 4, fig. 5, pl. 23, figs. 7, 8
*Eucyrtidium cubense Riedel and Sanfilippo [= Phormocyrtis cubensis (Riedel and Sanfilippo) Foreman], Eucyrtidium cubense Riedel and Sanfilippo 1971, p. 1594, pl. 7, figs. 10-11. Phormocyrtis cubensis (Riedel and Sanfilippo) Foreman 1973, p. 438, pl. 7, figs. 11, 12, 14. [Plate 2, fig- ures 1, 2]
*Giraffospyris lata Goll 1969, p. 334, pl. 58, figs. 22, 24-26; Sanfilippo and Riedel 1973, p. 529, pl. 18, figs. 3-7, pl. 33, fig. 4. [Plate 1, figure 18]
Heliostylus spp. Haeckel 1881, p. 457; Sanfilippo and Riedel 1973, p. 522, pl. 8, figs. 1-7, pl. 26, figs. 10-12, pl. 27, fig. 1.
*Hexacontium palaeocenicum Sanfilippo and Riedel 1973, p. 492, pl. 4, fig. 2, pl. 24, fig. 4.
*Lamptonium(?) colymbus Foreman 1973, p. 435, pl. 6, fig. 2, pl. 11, figs. 15, 19. [Plate 3, figure 2]
*Lamptonium fabaeforme chaunothorax Riedel and Sanfilippo. Lamptonium(?) fabaeforme(?) chaunothorax Riedel and Sanfilippo 1970, p. 524, pl. 5, figs. 8-9; Sanfilippo et al. 1985, p. 673, fig. 18.3.
*Lamptonium fabaeforme fabaeforme (Krasheninnikov). [?]Cyrtocalpisfabaeformis Krasheninnikov 1960, p. 296, pl. 3, fig. 11. Lamptonium (?) fabaeforme fabaeforme (Krasheninnikov) Riedel and Sanfilippo 1970, p. 523, pl. 5, fig. 6; Foreman 1973, p. 436, pl. 6, figs. 6-9; Sanfilippo et al. 1985, p. 674, fig. 18.2. [Plate 1, figures 7, 8; Plate 3, figure 5]
*Lamptonium(?) incohatum Foreman 1973, p. 436, pl. 6, fig. 1, pl. 11, fig. 18. [P1. 3, fig. 10]
*Lamptonium pennatum Foreman 1973, p. 436, pl. 6, figs. 3-5, pl. 11, fig. 13; Sanfilippo et al. 1985, p. 674, fig. 18.1. [Plate 1, figures 4-6]
Lamptonium sanfilippoae Foreman 1973, p. 436, pl. 6, figs. 15, 16, pl. 11, figs. 16, 17.
Lithochytris archaea Riedel and Sanfilippo 1970, pl. 9, fig. 8; Foreman 1973, p. 436, pl. 2, figs. 4, 5.
*Lithomespilus mendosa (Krasheninnikov). Ellipsidium (?) mendosum Krasheninnikov 1960, p. 281, pl. 1, fig. 14; Sanfilippo and Riedel 1973, p. 517, pl. 4, figs. 6, 7, pl. 24, figs. 10, 11.
*Orbula comitata Foreman 1973, p. 437, pl. 3, fig. 11, pl. 10, figs. 5, 7, 8. [Plate 3, figure 9]
*Orbula discipulus Foreman 1973, p. 438, pl. 3, fig. 10, pl. 10, figs. 9, 10. [Plate 3, figures 1, 7]
*Orbula ducalis Foreman 1973, p. 438, pl. 3, fig. 12, pl. 10, figs. 11, 12. [Plate 3, figure 11]
Phormocyrtis cubensis (Riedel and Sanfilippo). Eucyrtidium cubense Riedel and Sanfilippo 1971, p. 1594, pl. 7, figs. 10-11. Phormocyrtis cubensis (Riedel and Sanfilippo) Foreman 1973, p. 438, pl. 7, figs. 11, 12, 14. [Plate 2, fig- ures 1, 2]
*Phormocyrtis striata exquisita (Kozlova). Podocyrtis exquisita Kozlova in Kozlova and Gorbovets 1966, p. 106, pl. 17, fig. 2. Phormocyrtis striata exquisita (Kozlova) Foreman 1973, p. 438, pl. 7, figs. 1-4, 7, 8, pl. 12, fig. 5 (with synonymy); Sanfilippo et al. 1985, p. 678, figs. 20.2a-c. [Plate 2, figures 3, 4]
Phormocyrtis striata striata Brandt. Phormocyrtis striata Brandt 1935, p. 55, pl. 9, fig. 12; Riedel and Sanfilippo 1970, p. 532, pl. 10, fig. 7; Foreman 1973, p. 438, pl. 7, figs. 5, 6, 9; Sanfilippo et al. 1985, p. 679, figs. 21.1a, b. [Plate 2, figure 5]
*Phormocyrtis turgida (Krasheninnikov). Lithocampe turgida Krasheninnikov 1960, p. 301, pl. 3, fig. 17. Phormocyrtis
80
Micropaleontology, vol. 45, supplement 2, 1999
turgida (Krasheninnikov) Foreman 1973, p. 438, pl. 7, fig. 10; pl. 12, fig. 6. [Plate 2, figure 6]
Podocyrtis (Podocyrtis) papalis Ehrenberg. Podocyrtis papalis Ehrenberg 1847, p. 55, fig. 2; 1854, pl. 36, fig. 23; 1873, p. 251; Riedel and Sanfilippo 1970, p. 533, pl. 11, fig. 1; Sanfilippo and Riedel 1973, p. 531, pl. 20, figs. 11-14, pl. 36, figs. 2, 3, figs. 5, 6; Sanfilippo and Riedel 1992, pl. 3, figs. 9, 10. [Plate 2, figure 22]
Pterocodon(?) tenellus Foreman 1973, p. 439, pl. 5, fig. 7; pl. 12, fig. 4. [Plate 2, figure 13]
*Rhopalocanium ornatum Ehrenberg 1847, fig. 3; 1854, pl. 36, fig. 9; 1873, p. 256; 1875, p. 82, pl. 17, fig. 8; Fore- man 1973, p. 439, pl. 2, figs. 8-10, pl. 12, fig. 3.
*Spongodiscus americanus Kozlova, in Kozlova and Gorbovets 1966, p. 88, pl. 14, figs. 1, 2; Sanfilippo and Riedel 1973, p. 524, pl. 11, figs. 9-13, pl. 27, fig. 11, pl. 28, fig. 9.
Spongodiscus cruciferus (Clark and Campbell). Spongasteriscus cruciferus Clark and Campbell 1942, p. 50, pl. 1, figs. 1-6, 8, 10, 11, 16-18. Spongodiscus cruciferus (Clark and Campbell) Sanfilippo and Riedel 1973, p. 524, pl. 11, figs. 14-17, pl. 28, figs. 10, 11. [Plate 1, figure 1]
*Spongodiscus quartus quartus (Borisenko). Staurodictya quartus Borisenko 1958, p. 96, pl. 2, fig. 5. Spongodiscus quartus quartus in Sanfilippo and Riedel 1973, p. 525, pl. 12, figs. 6, 7; pl. 29, figs. 5, 6. [Plate 1, figure 15]
*Spongurus illepidus Krasheninnikov 1960, p. 283, pl. 2, fig. 2.
*Stylosphaera goruna Sanfilippo and Riedel 1973, p. 521, pl. 1, figs. 20-22; pl. 25, figs. 9, 10.
Stylotrochus quadribrachiatus quadribrachiatus Sanfilippo and Riedel 1973, p. 526, pl. 14, figs. 3, 4, pl. 31, fig. 1. [Plate 1, figure 3]
Theocorys(?) phyzella Foreman 1973, p. 440, pl. 5, fig. 8; pl. 12, fig. 1. [Plate 2, figures 20, 21]
*Theocosphaera larnacium Sanfilippo and Riedel 1973, p. 521, pl. 3, figs. 4-6, pl. 25, figs. 13, 14.
Theocotyle nigriniae Riedel and Sanfilippo. Theocotyle cryptocephala(?) nigriniae Riedel and Sanfilippo 1970, p. 525, pl. 6, fig. 5 (non 6); Theocotyle nigriniae Riedel and Sanfilippo, Sanfilippo and Riedel 1982, p. 178, pl. 2, figs. 1-3 (with synonymy); Sanfilippo et al. 1985, p. 685, figs. 25. la, b. [Plate 2, figures 16, 17]
*Theocotylissa alpha Foreman. Theocotyle (Theocotylissa) al- pha Foreman 1973, p. 441, pl. 4, figs. 13-15 (non 14), pl. 12, fig. 16; Foreman 1975, p. 621; Theocotylissa alpha Foreman, Sanfilippo and Riedel 1982, p. 179, pl. 2, figs. 16-17 (with synonymy); Sanfilippo et al. 1985, p. 686, fig. 25.6. [Plate 2, figures 18, 19; plate 3, figure 6]
*Theocotylissa auctor Foreman. Theocotyle (Theocotylissa) auctor Foreman 1973, p. 441, pl. 4, figs. 8-10, pl. 12, fig. 13. Theocotylissa auctor Foreman, Sanfilippo and Riedel 1982, p. 180, pl. 2, figs. 14,15; Sanfilippo et al. 1985, p. 686, figs. 25.5a, b. [Plate 2, figures 7-9]
*Theocotylissa sp. cf. T fimbria Foreman 1973, p. 441, pl. 5, figs. 1, 2, pl. 12, fig. 21.
Thyrsocyrtis (Thyrsocyrtis) hirsuta (Krasheninnikov). Podocyrtis hirsutus Krasheninnikov 1960, p. 300, pl. 3, fig. 16. Thyrsocyrtis (Thyrsocyrtis) hirsuta (Krasheninnikov) Sanfilippo and Riedel 1982, p. 173, pl. 1, figs. 3-4 (with synonymy); Sanfilippo et al. 1985, p. 687, fig. 26.2. [Plate 2, figures 10-12; plate 3, figure 15]
APPENDIX 3 The definitions of the radiolarian zones applied herein are from Sanfilippo and Nigrini (1998b).
Phormocyrtis striata striata Interval Zone (Foreman 1973; emend. Riedel and Sanfilippo 1978)
TOP: evolutionary transition from Theocotyle nigriniae to Theocotyle cryptocephala; coincident with the lower limit of the Theocotyle cryptocephala Zone.
BASE: morphotypic LO of Theocorys anaclasta; coincident with the upper limit of the Buryella clinata Zone.
Events included in the zone are: Spongatractus balbis -> Spongatractus pachystylus LAD Lamptonium sanfilippoae FAD Thyrsocyrtis (Thyrsocyrtis) rhizodon FAD Podocyrtis (Podocyrtoges) diamesa
The lower limit of the zone is approximately synchronous with FAD Lamptonium fabaeforme constrictum; Phormocyrtis striata exquisita -> Phormocyrtis striata striata; FAD Podocyrtis (Lampterium) acalles; LAD Phormocyrtis cubensis; FAD Lychnocanoma bellum.
Buryella clinata Interval Zone (Foreman 1973; emend. Fore- man 1975; emend. Riedel and Sanfilippo 1978)
TOP: morphotypic LO of Theocorys anaclasta; coincident with the lower limit of the Phormocyrtis striata striata Zone.
BASE: evolutionary transition from Pterocodon ?anteclinata to Buryella clinata; coincident with the upper limit of the Bekoma bidartensis Zone.
The lower limit of the zone is approximately synchronous with FAD Spongatractus balbis; FAD Lamptonium sanfilippoae; FAD Theocotyle nigriniae; FAD Thyrsocyrtis (Thyrsocyrtis) hirsuta
Remarks: This zone corresponds to the stratigraphic range of B. clinata (above its evolutionary transition from P. (?) anteclinata) below the LO of T. anaclasta.
Bekoma bidartensis Interval Zone (Foreman 1973; emend. Nishimura 1992; emend. Sanfilippo and Nigrini 1998b)
TOP: evolutionary transition from Pterocodon ? anteclinata to Buryella clinata; coincident with the lower limit of the Buryella clinata Zone.
BASE (redefined in Sanfilippo and Nigrini 1998b): morphotypic LO of Bekoma bidartensis; coincident with the up- per limit of the Bekoma campechensis Zone. Nishimura (1992) revised the definition of the base of this zone to the evolutionary
81
A. Sanfilippo and D.M. Hull: Upper Paleocene - Lower Eocene radiolarian biostratigraphy of the San Francisco de Paula Section, western Cuba
transition from Bekoma campechensis to Bekoma bidartensis. Sanfilippo and Nigrini (1998b) rejected this emendation be- cause, in practice, they found that recovered sediment se- quences of this age are short and discontinuous and thus unsuitable for recognizing evolutionary transitions. The defini- tion was therefore emended back to the original definition of Foreman 1973.
The Paleocene/Eocene boundary lies within this zone (Sanfilippo and Nigrini 1998a)
Bekoma campechensis Interval Zone (Nishimura 1987; emend. 1992; emend. Sanfilippo and Nigrini 1998b)
TOP (redefined in Sanfilippo and Nigrini 1998b): the morphotypic LO of Bekoma bidartensis. The definition is emended (ibid.) to coincide with the lower limit of the Bekoma bidartensis Zone.
Stylotrochus nitidus - Pterocodon(?) poculum Interval Sub- zone (Nishimura 1992; emend. Sanfilippo and Nigrini 1998b)
TOP (redefined in Sanfilippo and Nigrini 1998b): the morphotypic LO of Bekoma bidartensis. The definition is emended (ibid.) to coincide with the lower limit of the Bekoma bidartensis Zone.
BASE: morphotypic LO of Stylotrochus nitidus and Pterocodon(?) poculum; coincident with the upper limit of the Orbula discipulus Interval Subzone.
Orbula discipulus Interval Subzone (Nishimura 1992; emend. Sanfilippo and Nigrini 1998b)
TOP (defined in Sanfilippo and Nigrini 1998b): morphotypic LO of Stylotrochus nitidus and Pterocodon(?) poculum; coinci- dent with the lower limit of Stylotrochus nitidus -Pterocodon(?) poculum Interval Subzone.
BASE (defined in Sanfilippo and Nigrini 1998b): morphotypic HO of Peritiviator(?) dumitricai; coincident with the upper limit of Peritiviator(?) dumitricai Interval Subzone.
Remarks: Nishimura (1992) defined this zone "by the occur- rence of Orbula discipulus. The subzone is established on only three samples and appears to be an interval between Subzones Stylotrochus nitidus - Pterocodon(?) poculum and Peritivi- ator(?) dumitricai." However, 0. discipulus is present in only the middle one of the three stratigraphically sequential samples which Nishimura places in her subzone. Hence, the subzone does not represent the known range of 0. orbula. For that reason Sanfilippo and Nigrini (1998b) have chosen to treat her subzone as an Interval Zone defined by the established limits of the subzones above and below it.
Peritiviator(?) dumitricai Concurrent Range Subzone (Nishi- mura 1992)
TOP: morphotypic HO of Peritiviator(?) dumitricai; coincident with the lower limit of Orbula discipulus Interval Subzone.