UvA-DARE is a service provided by the library of the University of Amsterdam (http://dare.uva.nl) UvA-DARE (Digital Academic Repository) Genetic basis of rare blood group variants Wigman, L. Link to publication Citation for published version (APA): Wigman, L. (2013). Genetic basis of rare blood group variants. General rights It is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons). Disclaimer/Complaints regulations If you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, stating your reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Ask the Library: https://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam, The Netherlands. You will be contacted as soon as possible. Download date: 07 Jan 2020
Transcript
UvA-DARE is a service provided by the library of the University of Amsterdam (http://dare.uva.nl)
UvA-DARE (Digital Academic Repository)
Genetic basis of rare blood group variants
Wigman, L.
Link to publication
Citation for published version (APA):Wigman, L. (2013). Genetic basis of rare blood group variants.
General rightsIt is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s),other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons).
Disclaimer/Complaints regulationsIf you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, statingyour reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Askthe Library: https://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam,The Netherlands. You will be contacted as soon as possible.
RHD and RHCE variant and zygosity genotyping via multiplex ligation–dependent probe amplification
Lonneke Haer-Wigman1
Barbera Veldhuisen1
Remco Jonkers1
Martin Lodén2
Tracey E. Madgett3
Neil D. Avent3
Masja de Haas1
C. Ellen van der Schoot1
1 Sanquin Research, Amsterdam and Landsteiner Laboratory, Academic Medical Centre, University of Amsterdam, Amsterdam, the Netherlands
2 MRC-Holland b.v., Amsterdam, the Netherlands3 School of Biomedical and Biological Sciences, Plymouth University,
Plymouth, United Kingdom.
Transfusion 2013;53(7):1559-74
Chapter 3
52
Abstract
Background: The presence of a D variant may hamper correct serologic D typing, which may result in D immunization. D variants can be determined via RHD genotyping. However, a convenient single assay to identify D variants is still lacking. We developed and evaluated a multiplex ligation-dependent probe amplification [MLPA] assay to determine clinically relevant RHD and RHCE variant alleles and RHD zygosity.
Methods: We analyzed 236 cases (73 normal and 163 selected samples) with the RH-MLPA assay, which is able to determine 79 RHD and 17 RHCE variant alleles and RHD zygosity. To confirm the results, mutations were verified by RHD and/or RHCE exon–specific sequencing and RHD zygosity was verified by quantitative real-time polymerase chain reaction for 18 cases.
Results: In 99% of the cases, the RH-MLPA assay correctly determined whether a person carried only wild-type RHD and RHCE alleles (n = 69) or (a) variant RHD allele(s) and/or (a) variant RHCE
allele(s) (n = 164). In only three cases, including two new RHD variant alleles, the variant allele was not identified, due to lack of detecting probes. These were RHD*16.02, a new partial RHD
allele, RHD*525A (p.Phe175Leu), and a new D- null allele, RHD*443G (p.Thr148Arg). All RHD (n = 175) and RHCE variant alleles (n = 79) indicated by the RH-MLPA assay were confirmed by sequencing. RHD zygosity was confirmed by quantitative PCR. Two hematopoietic chimeras were recognized.
Conclusion: The RH-MLPA genotyping assay is a fast, easy, and reliable method to determine almost all clinically relevant RHD and RHCE variant alleles, RHD zygosity, and RHD+/RHD- chimeras in blood donors, blood recipients, and pregnant women.
Chap
ter 3
Genotyping of RHD variants and zygosity
53
Introduction
The Rh blood group system is one of the most complex and immunogenic blood group systems in humans.1,2 Antibodies against the Rh antigens can give severe hemolytic transfusion reactions and/or hemolytic disease of the fetus and/or newborn.3,4 Therefore, it is crucial to correctly determine the Rh status in blood donors, blood recipients, and pregnant women to prevent immunization to the Rh antigens.5-7 The antigens of the Rh blood group system are encoded by two closely linked genes, RHD and RHCE that encode the homologous RhD and RhCE proteins, respectively. 8 The RhD protein carries the D antigen and the RhCE protein carries the C, c, E, and e antigens.9-13 A wide genetic diversity exists in RHD and RHCE, resulting in more than 250 RHD and RHCE
variant alleles.14 These variant alleles have been classified into four groups: the partial RHD
alleles, alleles encoding the weak D, and D-elution (Del) phenotypes, the D- null alleles and the RHCE variant alleles, based on their phenotypic expression (see Web resources). Individuals carrying a partial RHD allele lack one or more D epitopes and are prone to make alloantibodies against the epitopes they miss.2,15 Individuals carrying a weak D or Del allele express the RhD protein, but in low amounts.2,15 In theory, individuals with a weak D or Del variant express all D epitopes; hence they will not make anti-D.14,15 There have been, however, reports describing alloanti-D formation in individuals with weak D variants.16-18 Both weak D and Del can elicit an immune response in D- recipients.19,20 Individuals with a D- null allele lack the complete expression of the RhD protein.14,15 The D- phenotype is most often caused by the entire deletion of the RHD gene (the RHD*01N.01 allele) in the Caucasian population or the presence of the RHD*Ψ allele (RHD*Pseudogene) or the hybrid RHD*03N.01 allele, formerly called (C)ceS Type 1 in the black African population.21-23 Serology with monoclonal anti-D is currently the standard method to determine the Rh status of blood donors, recipients, and pregnant women.2,15,24 Serologic typing will, in most cases, only reveal the presence of an RhD variant, but cannot exactly determine which variant is present.2,5 Because transfusion policy, as well as preventive measurements during pregnancy, are based on the exact nature of the RhD variant, it is important to correctly determine which RhD variant is present.2,5,6,14 RHD and RHCE variant alleles can be determined via genotyping of the RHD and RHCE genes.5,25,26 Because molecular genotyping methods are increasingly applied for blood donor typing and for noninvasive fetal RHD genotyping, the number of cases with discrepant serologic and genotyping results that require further molecular analysis is expanding.5,25 A method to identify a broad range of clinically relevant Rh variants (all frequently occurring variants and all variants where the carrier is able to make alloanti-D) in a convenient and cheap assay is still lacking.14,25
Only pregnant women who are D- or express an RhD variant and who are carrying a D+ fetus are at risk for anti-D formation and subsequently of anti-D–mediated hemolytic disease of the fetus and/or newborn.3,6 Currently, the presence of a D+ fetus can be determined with
Chapter 3
54
noninvasive fetal RHD typing.27,28 If the father is homozygous for the RHD*01 allele (wild-type RhD) this test can be omitted.6,14 Upon serologic typing of the C, c, E, and e antigens of the father, a prediction of the paternal zygosity can be made based on the most frequently occurring RHD/RHCE haplotypes.6,29 Because the haplotype frequencies differ between ethnic populations, these predictions are not reliable without knowledge of the ethnic background.6,29 The exact paternal zygosity can be determined by genotype-based methods; however, the methods used at the moment are cumbersome and can be confounded by genetic alterations, especially in the black African population.30-33 We therefore developed a multiplex ligation–dependent probe amplification [MLPA] genotyping assay. This single assay detects mutations and copy number variation of the RHD and RHCE genes and is easy to use, as it only requires a thermocycler and capillary electrophoresis equipment. The aim of this study was to evaluate the performance of the RH-MLPA genotyping assay in the determination of clinically relevant RHD and RHCE variant alleles and RHD zygosity.
Material and methods
MaterialsDNA samples from 73 random blood donors with normal Rh serology were included after informed consent was given (including 11 D- and 62 D+ samples; 44 C+, 60 c+, 27 E+, and 68 e+ samples). Sanquin Diagnostics is the national reference laboratory for the analysis of the presence of Rh variants in blood recipients, blood donors, and pregnant women. For this study 97 cases were selected, which were analyzed between 2003 and 2011. This selection included preferentially a minimum of three samples for each known RHD variant allele and all samples of which the specific RHD variant was not yet determined. Twenty DNA samples were provided by other blood group centers and were selected for the presence of a specific RHD
variant allele. Forty-six cases positive for the RHD*Ψ allele were obtained from the screening program of D- pregnant women.
D serologySerologic D typing was routinely preformed with two anti-D reagents. A monoclonal anti-D reagent (immunoglobulin [Ig]M clone MS201, Sanquin Reagents) and a monoclonal blend reagent (IgM clone TH28 and IgG clone MS26, Sanquin Reagents) were used in a method with an immediate spin at room temperature. All D- samples were also tested with the monoclonal blend reagent in the indirect antiglobulin test. In case of a discrepancy between the results of the two reagents, the D-epitope pattern was determined with an in-house panel of anti-D reagents and/or the extended partial RhD typing set from Bio-Rad Laboratories B.V..
Chap
ter 3
Genotyping of RHD variants and zygosity
55
DNA isolationEthylenediaminetetraacetic acid–anticoagulated blood was collected and genomic DNA was isolated from white blood cells using a DNA extraction kit (QIAamp DNA blood mini kit).
MLPAAn MLPA assay specific for the analysis of RHD and RHCE, their variants and zygosity was developed. This RH-MLPA assay contains 17 RHD wild type, five RHCE wild type, 21 RHD
mutation, and two RHCE mutation probe combinations (Table 1). So-called wild type probe combinations are used to determine the copy number of wild-type RHD or RHCE sequence, while mutation probe combinations detect the presence of a mutated sequence. Twenty-five probe combinations were conventionally designed and consisted of two probe pieces and for the additional 20 probe combinations a three-piece design was chosen (Table 1). The RH-MLPA assay is able to distinguish between 51 RHD and 13 RHCE variant alleles. For another 28 RHD and four RHCE variant alleles the assay can identify the main type, but cannot discriminate between several subtypes, for example, RHD*12.01 or RHD*12.02 alleles (Table 2). Because probes hybridizing to the same regions will hamper each others’ binding, the probes of the RH-MLPA had to be divided into three pools, p401, p402, and p403 (Table 1). The MLPA reaction was performed according to the manufacturers’ protocol (MRC Holland) on a thermocycler (Biometra T1, Westerburg B.V. or Veriti, Applied Biosystems). In short, 5 μL containing 100 ng of DNA was denatured and 1.5 μL probe mix and 1.5 μL SALSA MLPA dilution buffer were added at 25°C. After hybridization at 60°C for between 16 and 20 hours, 1 μL of SALSA ligase-65, 1.5 μL of SALSA ligase Buffer A, and 1.5 μL SALSA ligase Buffer B were added and incubated for 15 minutes at 54°C. A polymerase chain reaction [PCR] was performed in a total volume of 50 μL containing 4 μL of SALSA dilution buffer, 2 μL of SALSA enzyme dilution buffer, 2 μL of universal primers, 0.5 μL of SALSA polymerase, and 10 μL of ligation sample. PCR conditions were as follows: 5 minutes at 95°C, 35 cycles of 30 seconds at 95°C, 30 seconds at 60°C and 1 minute at 72°C, followed by 20 minutes at 72°C. A mixture of 1.5 μL of MLPA sample, 8.5 μL of Formamide (Hi-Di, Applied Biosystems), and 0.5 μL of size standard (GeneScan 500-Liz, Applied Biosystems) was analyzed on a genetic analyzer (Model 3130, Applied Biosystems). Data analysis was performed using computer software (Genemarker, Version 1.85, Softgenetics). For each run two control samples hemizygously positive for the RHD*01 allele, as determined via RHD Exons 5- and 7-specific quantitative real-time PCR were used as reference samples to determine zygosity. Supplementary Figure S1 shows an example of the results of an MLPA reaction. The total RH-MLPA assay takes approximately 20 hours, including 16 hours of hybridization and 2 hours of hands-on time. Between 1 and 16 samples can be analyzed simultaneously in a single run consisting of all three RH-MLPA mixes.
Chapter 3
56
Tabl
e 1.
Per
form
ance
of R
HD
and
RH
CE-
spec
ific-
prob
es fr
om th
e RH
-MLP
A te
sted
in 2
36 c
ases
Prob
e na
me
Gen
eEx
on o
r In
tron
Prim
ary
ligat
ion
site
†Se
cond
ary
ligat
ion
site
‡Pr
obe
size
Use
of p
robe
/det
ectin
gPr
esen
t in
mix
Sign
al
dete
cted
in
test
ed c
ases
Mut
atio
n ve
rfied
by
seq
uenc
ing
D00
_-13
2ARH
D5’
UTR
c.-1
32A
c.-1
73C
217
RHD
*01
P401
222
MD
01_0
08G
RHD
Exon
1c.
8G14
9RH
D*0
1W.0
3P4
013
3
DC
E01_
048C
RHD
/RH
CEEx
on 1
c.48
C14
3RH
CEP4
0219
1
MD
01_0
48A
RHD
Exon
1c.
48A
144
RHD
*01N
.08
P403
33
MC
E01_
122G
RHCE
Exon
1c.
122G
156
RHCE
*02.
08.0
1P4
024
4
MD
02_1
86T
RHD
Exon
2c.
186T
c.22
5C24
1RH
D*0
3.01
, RH
D*0
3.04
, RH
D*0
4.01
, RH
D*0
3N.0
1P4
0114
5
DC
E02_
203G
RHD
/RH
CEEx
on 2
c.[2
01G
; 203
G]
224
RHD
*01
and
RHCE
*CP4
0322
0
MD
02_2
70A
RHD
Exon
2c.
270A
214
RHD
*01N
.10
P403
00
CE0
2_30
7CRH
CEEx
on 2
c.30
7Cc.
269G
218
RHCE
*cP4
0220
8
MD
02_3
29C
RHD
Exon
2c.
329C
158
RHD
*07.
01P4
036
6
DC
E02_
IVS1
-20G
RHD
/RH
CEIn
tron
2c.
336-
20G
180
RHD
*01
and
RHCE
*CP4
0222
1
CE0
2_in
s_C
RHCE
Intr
on 2
RHC
E*C
spe
cific
in
sert
ion
156
RHCE
*CP4
0112
3
D03
_380
TRH
DEx
on 3
c.38
0Tc.
405C
167
RHD
*01
P402
220
MD
03_4
10T
RHD
Exon
3c.
410T
c.38
3A19
2RH
D*0
3.01
, RH
D*0
3.04
, RH
D*0
3.06
, RH
D*0
4.01
, RH
D*1
2.03
, RH
D*0
3N.0
1P4
0113
9
MD
03_4
46A
RHD
Exon
3c.
446A
c.41
9T18
8RH
D*0
1W.0
5P4
034
3
D03
_455
ARH
DEx
on 3
c.45
5Ac.
486+
4T17
5RH
D*0
1P4
0220
7
MD
03_I
VS3+
1ARH
DIn
tron
3c.
486+
1A24
7RH
D*0
1EL.
02P4
014
4
MD
04_5
09C
RHD
Exon
4c.
509C
163
RHD
*12
P402
11
D04
_514
ARH
DEx
on 4
c.51
4Ac.
487-
5T18
7RH
D*0
1P4
0116
8
D04
_602
CRH
DEx
on 4
c.60
2C22
4RH
D*0
1P4
0219
6
MD
04_6
09A
RHD
Exon
4c.
609A
130
RHD
*ψP4
0146
5
D05
_667
TRH
DEx
on 5
c.66
7Tc.
639C
211
RHD
*01
P402
159
CE0
5_67
6GRH
CEEx
on 5
c.67
6Gc.
712A
205
RHCE
*eP4
0123
0
Chap
ter 3
Genotyping of RHD variants and zygosity
57
Tabl
e 1.
(Con
tinue
d)
Prob
e na
me
Gen
eEx
on o
r In
tron
Prim
ary
ligat
ion
site
†Se
cond
ary
ligat
ion
site
‡Pr
obe
size
Use
of p
robe
/det
ectin
gPr
esen
t in
mix
Sign
al
dete
cted
in
test
ed c
ases
Mut
atio
n ve
rfied
by
seq
uenc
ing
CE0
5_67
6CRH
CEEx
on 5
c.67
6Cc.
712A
211
RHCE
*EP4
0151
D05
_697
GRH
DEx
on 5
c.69
7G19
2RH
D*0
1P4
0319
5
MC
E05_
733G
RHCE
Exon
5c.
733G
c.78
7A20
4RH
CE*0
1.20
, RH
D*0
3N.0
1P4
0219
10
D05
_787
GRH
DEx
on 5
c.78
7Gc.
801+
17C
181
RHD
*01
P401
198
D05
_800
ARH
DEx
on 5
c.80
0A14
8RH
D*0
1P4
0320
2
MD
06_8
07G
RHD
Exon
6c.
807G
167
RHD
*01N
.18,
RH
D*ψ
P403
464
MD
06_8
09G
RHD
Exon
6c.
809G
192
RHD
*01W
.01
P402
245
MD
06_8
19A
RHD
Exon
6c.
819A
c.80
2-9T
169
RHD
*03.
01, R
HD
*03.
06,
RHD
*09.
03, R
HD
*09.
04,
RHD
*09.
05P4
015
3
MD
06_8
45A
RHD
Exon
6c.
845A
164
RHD
*15
P403
54
MD
06_8
72G
RHD
Exon
6c.
872G
132
RHD
*09.
05P4
020
0
MD
06_8
85T
RHD
Exon
6c.
885T
c.90
6G17
4RH
D*1
1P4
015
4
D06
_932
ARH
DEx
on 6
c.93
2A23
7RH
D*0
1P4
0220
3
D07
_941
GRH
DEx
on 7
c.94
1G17
4RH
D*0
1P4
0321
1
D07
_989
ARH
DEx
on 7
c.98
9A14
3RH
D*0
1P4
0121
1
MD
07_1
025C
RHD
Exon
7c.
1025
Cc.
992A
151
RHD
*04.
03, R
HD
*09.
01,
RHD
*09.
02, R
HD
*01W
.29
P402
74
D07
_106
1TRH
DEx
on 7
c.10
61T
181
RHD
*01
P403
206
MD
07_1
063A
RHD
Exon
7c.
1063
Ac.
1073
+25
T22
2RH
D*2
5P4
013
3
MD
08_1
136T
RHD
Exon
8c.
1136
T21
1RH
D*1
0P4
035
5
MD
09_1
154C
RHD
Exon
9c.
1154
Cc.
1999
A23
6RH
D*0
1W.0
2P4
018
5
D09
_119
3ARH
DEx
on 9
c.11
93A
c.12
27+
2T18
7RH
D*0
1P4
0221
9
MD
09_1
227A
RHD
Exon
9c.
1227
Ac.
1227
+46
C20
4RH
D*0
1EL.
01P4
033
3
D10
_137
5CRH
DEx
on 1
0c.
1375
C
162
RHD
*01
P401
223
† Po
sitio
n as
cou
nted
from
ATG
tran
slat
ion
star
t site
and
nuc
leot
ide
of th
e lig
atio
n si
te th
at is
spe
cific
for t
he n
ucle
otid
e de
tect
ed b
y th
e pr
obe
‡ Po
sitio
n as
cou
nted
from
ATG
tran
slat
ion
star
t site
and
nuc
leot
ide
of a
sec
ond
ligat
ion
site
whi
ch is
use
d to
mak
e th
e pr
obe
spec
ific
for R
HD
or R
HCE
Chapter 3
58
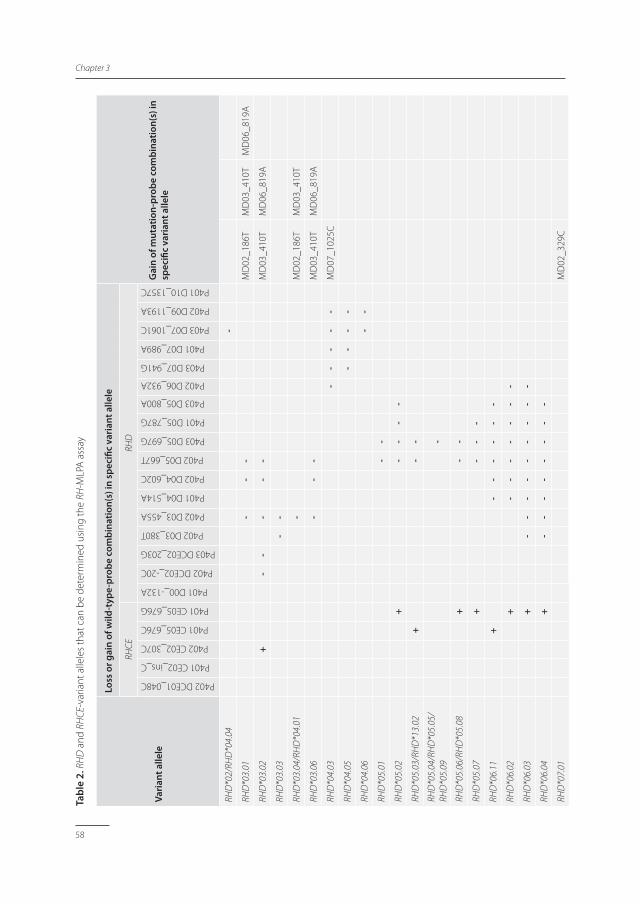
Tabl
e 2.
RH
D a
nd R
HCE
-var
iant
alle
les
that
can
be
dete
rmin
ed u
sing
the
RH-M
LPA
ass
ay
Vari
ant a
llele
Loss
or g
ain
of w
ild-t
ype-
prob
e co
mbi
natio
n(s)
in s
peci
fic v
aria
nt a
llele
Gai
n of
mut
atio
n-pr
obe
com
bina
tion(
s) in
sp
ecifi
c va
rian
t alle
le
RHCE
RHD
P402 DCE01_048C
P401 CE02_ins_C
P402 CE02_307C
P401 CE05_676C
P401 CE05_676G
P401 D00_-132A
P402 DCE02_-20C
P403 DCE02_203G
P402 D03_380T
P402 D03_455A
P401 D04_514A
P402 D04_602C
P402 D05_667T
P403 D05_697G
P401 D05_787G
P403 D05_800A
P402 D06_932A
P403 D07_941G
P401 D07_989A
P403 D07_1061C
P402 D09_1193A
P401 D10_1357C
RHD
*02/
RHD
*04.
04
-
RHD
*03.
01
-
--
M
D02
_186
TM
D03
_410
TM
D06
_819
A
RHD
*03.
02
+
-
-
-
--
M
D03
_410
TM
D06
_819
A
RHD
*03.
03
-
-
RHD
*03.
04/R
HD
*04.
01
-
M
D02
_186
TM
D03
_410
T
RHD
*03.
06
-
--
M
D03
_410
TM
D06
_819
A
RHD
*04.
03
-
--
--
M
D07
_102
5C
RHD
*04.
05
--
--
RHD
*04.
06
--
RHD
*05.
01
-
-
RHD
*05.
02
+
-
--
-
RHD
*05.
03/R
HD
*13.
02
+
-
-
RHD
*05.
04/R
HD
*05.
05/
RHD
*05.
09
-
RHD
*05.
06/R
HD
*05.
08
+
-
-
RHD
*05.
07
+
-
--
RHD
*06.
11
+
-
--
--
-
RHD
*06.
02
+
-
--
--
--
RHD
*06.
03
+
-
--
--
--
--
RHD
*06.
04
+
-
--
--
--
-
RHD
*07.
01
M
D02
_329
C
Chap
ter 3
Genotyping of RHD variants and zygosity
59
Tabl
e 2.
(Con
tinue
d)
Vari
ant a
llele
Loss
or g
ain
of w
ild-t
ype-
prob
e co
mbi
natio
n(s)
in s
peci
fic v
aria
nt a
llele
Gai
n of
mut
atio
n-pr
obe
com
bina
tion(
s) in
sp
ecifi
c va
rian
t alle
le
RHCE
RHD
P402 DCE01_048C
P401 CE02_ins_C
P402 CE02_307C
P401 CE05_676C
P401 CE05_676G
P401 D00_-132A
P402 DCE02_-20C
P403 DCE02_203G
P402 D03_380T
P402 D03_455A
P401 D04_514A
P402 D04_602C
P402 D05_667T
P403 D05_697G
P401 D05_787G
P403 D05_800A
P402 D06_932A
P403 D07_941G
P401 D07_989A
P403 D07_1061C
P402 D09_1193A
P401 D10_1357C
RHD
*07.
02
+
M
D02
_329
C
RHD
*09.
01
--
M
D07
_102
5C
RHD
*09.
02
--
-
M
D07
_102
5C
RHD
*09.
03/ R
HD
*09.
04
--
M
D06
_819
A
RHD
*09.
05
--
M
D06
_819
AM
D06
_872
G
RHD
*10.
00/R
HD
*10.
01/
RHD
*10.
02/R
HD
*10.
03/
RHD
*10.
06/R
HD
*10.
07
M
D08
_113
6T
RHD
*10.
04
-
M
D08
_113
6T
RHD
*10.
05
-
-
M
D08
_113
6T
RHD
*11
MD
06_8
85T
RHD
*12.
01/R
HD
*12.
02
-
M
D04
_509
C
RHD
*12.
03
-
M
D03
_410
TM
D04
_509
C
RHD
*13.
01
+
-
--
-
RHD
*14.
01
-
--
--
--
-
RHD
*14.
02
-
--
--
--
--
RHD
*15
MD
06_8
45A
RHD
*16.
01/R
HD
*30
-
RHD
*17.
01/R
HD
*17.
03
-
RHD
*17.
02
-
-
RHD
*25
-
MD
07_1
063A
RHD
*01W
.01
MD
06_8
09G
Chapter 3
60
Tabl
e 2.
(Con
tinue
d)
Vari
ant a
llele
Loss
or g
ain
of w
ild-t
ype-
prob
e co
mbi
natio
n(s)
in s
peci
fic v
aria
nt a
llele
Gai
n of
mut
atio
n-pr
obe
com
bina
tion(
s) in
sp
ecifi
c va
rian
t alle
le
RHCE
RHD
P402 DCE01_048C
P401 CE02_ins_C
P402 CE02_307C
P401 CE05_676C
P401 CE05_676G
P401 D00_-132A
P402 DCE02_-20C
P403 DCE02_203G
P402 D03_380T
P402 D03_455A
P401 D04_514A
P402 D04_602C
P402 D05_667T
P403 D05_697G
P401 D05_787G
P403 D05_800A
P402 D06_932A
P403 D07_941G
P401 D07_989A
P403 D07_1061C
P402 D09_1193A
P401 D10_1357C
RHD
*01W
.02
MD
09_1
154C
RHD
*01W
.14/
RHD
*01W
.40/
RHD
*01W
.51
-
RHD
*01W
.29
-
-
MD
07_1
025C
RHD
*01W
.41
-
RHD
*01E
L.01
MD
09_1
227A
RHD
*01E
L.02
MD
_IVS
3+1A
RHD
*01N
.01
RHD
*ψ
M
D04
_609
AM
D06
_807
G
RHD
*01N
.02
+
+
RHD
*01N
.03
+
+
RHD
*01N
.04
+
RHD
*01N
.05
+
RHD
*01N
.06
+
RHD
*01N
.07
+
RHD
*01N
.08
MD
01_0
48A
RHD
*01N
.10
MD
02_2
70A
RHD
*01N
.18
MD
06_8
07G
RHD
*01N
.20
RHD
*03N
.01
M
D02
_186
TM
D03
_410
TM
CE0
5_73
3G
RHCE
*01.
04/R
HCE
*01.
05+
++
Chap
ter 3
Genotyping of RHD variants and zygosity
61
Tabl
e 2.
(Con
tinue
d)
Vari
ant a
llele
Loss
or g
ain
of w
ild-t
ype-
prob
e co
mbi
natio
n(s)
in s
peci
fic v
aria
nt a
llele
Gai
n of
mut
atio
n-pr
obe
com
bina
tion(
s) in
sp
ecifi
c va
rian
t alle
le
RHCE
RHD
P402 DCE01_048C
P401 CE02_ins_C
P402 CE02_307C
P401 CE05_676C
P401 CE05_676G
P401 D00_-132A
P402 DCE02_-20C
P403 DCE02_203G
P402 D03_380T
P402 D03_455A
P401 D04_514A
P402 D04_602C
P402 D05_667T
P403 D05_697G
P401 D05_787G
P403 D05_800A
P402 D06_932A
P403 D07_941G
P401 D07_989A
P403 D07_1061C
P402 D09_1193A
P401 D10_1357C
RHCE
*01.
07+
+
RHCE
*01.
08/R
HCE
*01.
09+
RHCE
*01.
20
M
CE0
5_73
3G
RHCE
*01.
20.0
6+
+
M
CE0
5_73
3G
RHCE
*01.
22
≥1
++
++
RHCE
*ce-
D(9
)-ce
+
RHCE
*02.
02
++
RHCE
*02.
04
≥1
++
++
RHCE
*02.
08.0
1
M
CE0
1_12
2G
RHCE
*02.
10.0
1
+
+
RHCE
*02.
10.0
2
++
+
RHCE
*02.
17
+
RHCE
*03.
03+
+
RHCE
*03.
04
+
Chapter 3
62
RHD multiplex PCRIn our institution all samples that show unexpected weak or negative reactions with D-typing reagents are currently analyzed using a multiplex PCR, simultaneously detecting RHD Exons 3, 4, 5, 6, 7, and 9 and an internal control.34 The RHD multiplex [RHD-MPX] PCR was performed on a DNA engine thermocycler (Dyad, Bio-Rad Laboratories B.V.) in a total volume of 50 μL, containing 10 to 1000 ng DNA, 5 μL of 10x FastStart Taq DNA polymerase buffer without MgCl2 (Roche), 3 μL of MgCl2 25 μM stock solution (Roche), 1 μL of PCR-grade nucleotide mix (Roche), 15 μL of primer mix (primer dilution between 0.1 and 1.2 μM; Eurogentec). PCR conditions were as follows: 5 minutes at 95°C, 33 cycles of 1 minute at 95°C, 1 minute at 56°C, and 1 minute at 72°C, followed by 5 minutes at 72°C. The presence or absence of the RHD
exons and internal control was analyzed by gel electrophoresis on a 1.5% (wt/vol) agarose gel (Invitrogen).
PCR and sequencingRHD and RHCE exon–specific sequencing was performed using RHD- and RHCE-specific primers flanking each exon. The PCR was performed on a thermocycler (Veriti, Applied Biosystems) in a total volume of 20 μL, containing 50 to 150 ng DNA, 10 μL of 2x PCR master mix (GeneAmp Fast, Applied Biosystems), 0.5 μM forward and reverse primer. PCR conditions were as follows: 10 seconds at 95°C, 35 cycles of 10 seconds at 95°C, and a specific annealing-elongation temperature and time for each primer set ranging from 61 to 68ºC, followed by 1 minute at 72°C. PCR products were purified using PCR product cleanup (ExoSAP-IT, GE Healthcare), according to manufacturers’ protocol. The sequence reaction was performed on a thermocycler (Veriti, Applied Biosystems) in a total volume of 20 μL, containing 1 μL of purified PCR product, 1 μL of 2.5x polymerase mix (BigDye Terminator v1.1, Applied Biosystems), 3.5 μL of 5x buffer (BigDye Terminator, Applied Biosystems), and 0.25 μM forward or reverse primer. Sequence conditions were as follows: 25 cycles of 15 seconds at 95°C, 10 seconds at 50°C, and 4 minutes at 60°C. Sequence products were analyzed on a genetic analyzer (3130, Applied Biosystems).
RHD Exons 5- and 7-specific quantitative real-time PCRZygosity was determined with an RHD Exons 5- and 7-specific quantitative real-time PCR. Primers were developed to detect ALB35 (quantification of input DNA), SRY36, and RHD Exons 527 and 737. The RHD Exons 5 and 7 primers and probes were synthesized by TIB MOLBIOL and the primers for SRY and ALB were synthesized by Invitrogen. The real-time PCR was performed on a sequence detection system (ABI PRISM 7000, Applied Biosystems) in a total volume of 25 μL, containing 25 ng of DNA, 12.5 μL of universal primer PCR master mix (Taqman, Applied Biosystems), 0.1 μM of the RHD Exon 5 and SRY probe or RHD Exon 7 and ALB probe and 0.3 and 0.9 μM of the RHD Exon 5 and SRY forward and reverse primers, respectively, or 0.3 μM of the
Chap
ter 3
Genotyping of RHD variants and zygosity
63
RHD Exon 7 and ALB forward and reverse primers. Real-time PCR conditions were as follows: 2 minutes at 50°C, 10 minutes at 95°C, and 50 cycles of 15 seconds at 95°C and 1 minute at 60°C. Data analysis was performed using computer software (Microsoft Office Excel 2003).
Short-tandem-repeat multiplex PCRThe presence of chimerism was determined with genomic DNA using short-tandem-repeat multiplex (Powerplex 16 HS system, Promega). The reaction was performed according to the manufacturers’ protocol.
ResultsPerformance of RH-MLPA assay with plasmid DNAFirst, all RH-MLPA wild-type (n = 22) and mutation probe combinations (n = 23) were analyzed using a cloned DNA plasmid sample, containing the genomic sequence of all probes, as a template. For all probes correct signals were detected (data not shown) and subsequently the performance of the RH-MLPA was evaluated with a set of genomic DNA samples.
Performance of RH-MLPA assay with genomic DNA
Wild-type probe combinationsNext, the performance of the RH-MLPA assay was analyzed with DNA from 73 healthy donors with normal Rh serology, representing all common Rh phenotypes. The RHD wild-type probe combinations are able to detect the copy number of all RHD exons, except that of Exon 8. No probe combination for RHD Exon 8 was included, because the sequence of RHD and RHCE Exon 8 is completely identical. The probe combinations detecting wild-type RHD Exon 2 also detect RHCE*C, because the RHD and RHCE*C sequence of Exon 2 is completely identical. RHCE*C genotyping is based on the detection of the RHCE*C specific 19-nucleotide insertion in Intron 2 and c.203G in Exon 2. RHCE*c genotyping is based on the detection of c.307C. RHCE*E and RHCE*e genotyping is based on the detection of c.676C and c.676G, respectively. To ensure RHCE specificity, the latter two probe combinations consist of three probe pieces, creating a second ligation site for the c.712A RHCE-specific nucleotide. In all D+ cases (n = 62) all RHD wild-type probe combination signals were present, while in the D- cases (n = 11) all RHD wild-type probe combination signals were absent. The RH-MLPA RHCE*01 (Rhce), RHCE*02 (RhCe), RHCE*03 (RhcE), and RHCE*04 (RhCE) genotyping results were completely concordant with serology. All possible common RHD/RHCE haplotypes were determined at least once (data not shown). In four cases one or more mutation probe combination signals were detected: two cases showed the presence of the RHD*07.02 allele next to an RHD*01 allele (wild-type RhD). Similarly one case had an RHD*Ψ allele next to an RHD*01 allele and one case carried an RHCE*02.08.01 allele (CeCw). Sequencing of the involved exons in these cases confirmed the presence of the variant alleles detected by the RH-MLPA.
Chapter 3
64
Mutation probe combinations
The mutation probe combinations of the RH-MLPA were selected to recognize, in combination with the wild-type probe combinations, the majority of clinically relevant RHD
variant alleles and most frequently occurring RHCE variant alleles in the Caucasian and black African populations. The performance of the mutation probe combinations was evaluated with DNA from 97 cases, for whom serology indicated the presence of an Rh variant and with DNA from 66 cases, for whom the presence and/or expressing of an Rh variant was already determined. With this set of samples 19 RHD mutation probe combination signals and two RHCE mutation probe combination signals were obtained at least once. To confirm the presence of a mutation indicated by one of the mutation probe combinations, the exons containing the respective mutations were sequenced for at least three independent samples (if available). All sequencing results were concordant with the results of the RH-MLPA (Table 1). For two RHD mutation probe combinations MD02_270A specific for the RHD*01N.10 allele and MD06_872G specific for the RHD*09.05 allele no genomic DNA was available. The RHCE mutation probe combination MCE05_733G was designed to determine whether an individual is positive (c.733G) or negative (c.733C) for the VS antigen. Of note, individuals carrying an RHCE*01.04 allele (ceAR) have the c.733G mutation, but are VS-.38 We designed the MCE05_733G with a second ligation site specific for the c.787A RHCE wild-type single-nucleotide polymorphism [SNP], which is present in all VS+ alleles and absent in the RHCE*01.04 allele. Six of the nine cases carrying a RHCE*01.04 allele were VS- and indeed these samples showed the absence of signal for MCE05_733G. In the three other cases the second RHCE allele was a VS+ allele and as expected the MCE05_733G signal was obtained.
Determination of RHD and RHCE variant alleles using the RH-MLPA assayIn 160 of the above-described 163 individuals the RH-MLPA indicated the presence of one or more variant alleles. In these 163 cases a total of 172 RHD and 78 RHCE variant alleles or variant allele groups were determined as shown in Table 3. The assigned RHD and RHCE variant allele(s) were in all cases compatible with the initial serologic results obtained for the cases. In addition, in 99 of the 163 selected samples, we performed an RHD-MPX PCR, in which RHD Exons 3, 4, 5, 6, 7, and 9 are RHD specifically amplified. The results of the RHD-MPX combined with the results of the serology were concordant with the RHD variants concluded from the RH-MLPA, except for 11 cases. In these cases, carrying the RHD*05.05, RHD*10.02, RHD*10.03, RHD*11, or RHD*15 allele, the RHD-MPX results showed no aberrant pattern, suggesting the presence of weak D variants, while the RH-MLPA indicated the presence of partial RhD variants. Sequencing of the exons containing the mutations showed that in these cases the RH-MLPA assigned the correct RHD variant alleles (Table 3). In one case the RH-MLPA indicated the presence of a new partial RHD allele, containing the mutations of both RHD*03.02
and RHD*25 in one allele (c.150T>C, c.178A>C, c.201G>A, c.203G>A, c.307T>C, and c.1063G>A).
Chap
ter 3
Genotyping of RHD variants and zygosity
65
Tabl
e 3.
RH
D a
nd R
HCE
var
iant
alle
les
dete
cted
by
the
RH-M
LPA
ass
ay in
163
DN
A s
ampl
es
Sele
cted
don
ors
RH-M
LPA
con
clus
ion
Add
ition
al g
enot
ypin
g§Va
rian
t alle
le
dete
cted
in
test
ed c
ases
Sero
logy
and
RH
D-M
PX P
CR†
Sequ
ence
‡RH
DRH
CERH
DRH
CE
Part
ial D
alle
les
RH
D*0
3.01
RHD
*03.
01RH
CE*0
1.20
1
RHD
*03.
02N
ew R
HD
var
iant
(c.1
50T>
C;
c.17
8A>
C; c
.201
G>
A; c
.203
G>
A;
c.30
7T>
C; c
.106
3G>
A)
1
RHD
*03.
03RH
D*0
3.03
1
DIV
aRH
D*0
3.04
or R
HD
*04.
02RH
D*0
4.02
3
RHD
*03.
04RH
D*0
2 or
RH
D*0
4.04
RHD
*04.
041
DVI
t1RH
D*0
6.01
2
DVI
t2RH
D*0
6.02
7
DVI
IRH
D*0
7.01
4
DA
R††
RHD
*09.
01RH
CE*0
1.04
or R
HCE
*01.
05RH
CE*0
1.04
2
RHD
*09.
01RH
CE*0
1.04
or R
HCE
*01.
05 a
nd
RHCE
*01.
20RH
CE*0
1.04
2
RHD
*09.
01 (h
omoz
ygou
s)RH
CE*0
1.04
or R
HCE
*01.
05
(hom
ozyg
ous)
RHCE
*01.
04
(hom
ozyg
ous)
1
RHD
*09.
01 a
nd R
HD
*03N
.01
RHCE
*01.
04 o
r RH
CE*0
1.05
RHCE
*01.
041
RHD
*09.
02RH
CE*0
1.04
or R
HCE
*01.
05RH
CE*0
1.04
2
RHD
*01W
.29
1
RHD
*11
RHD
*11
3
RHD
*14.
02RH
D*1
4.02
1
RHD
*16.
01RH
D*1
6.01
1
DFR
RHD
*17.
011
RHD
*17.
022
RHD
*12.
01RH
D*1
2.01
or R
HD
*12.
02RH
CE*0
1.08
or R
HCE
*01.
09RH
D*1
2.01
RHCE
*01.
081
RHD
*25
RHD
*25
2
Part
ial D
var
iant
RHD
*01
RHD
*16.
011
New
var
iant
RH
D*5
25A
1
RHD
*05.
073
Chapter 3
66
Tabl
e 3.
(Con
tinue
d)
Sele
cted
don
ors
RH-M
LPA
con
clus
ion
Add
ition
al g
enot
ypin
g§Va
rian
t alle
le
dete
cted
in
test
ed c
ases
Sero
logy
and
RH
D-M
PX P
CR†
Sequ
ence
‡RH
DRH
CERH
DRH
CE
RH
D*1
0.05
1
RHD
*09.
03 o
r RH
D*0
9.04
RHCE
*01.
20RH
D*0
9.03
1
RHD
*09.
03 o
r RH
D*0
9.04
and
RH
D*0
3N.0
1RH
D*0
9.03
2
RHD
*09.
03 o
r RH
D*0
9.04
and
RH
D*0
3N.0
1RH
CE*0
1.20
RHD
*09.
031
Wea
k D
var
iant
RHD
*05.
04, R
HD
*05.
05 o
r RH
D*0
5.09
RHD
*05.
051
RHD
*10.
00, R
HD
*10.
01, R
HD
*10.
02,
RHD
*10.
03, R
HD
*10.
06 o
r RH
D*1
0.07
RHD
*10.
021
RHD
*10.
031
RHD
*10.
00, R
HD
*10.
01, R
HD
*10.
02,
RHD
*10.
03, R
HD
*10.
06 o
r RH
D*1
0.07
an
d RH
D*Ψ
RHCE
*ce-
D(9
)-ce
RHD
*10.
031
RHD
*11
2
RHD
*15
5
Wea
k D
and
Del
al
lele
s
Wea
k D
type
2
RHD
*01W
.02
7
RHD
*01W
.02
and
RHD
*03N
.01
1
RHD
*01W
.03
RHD
*01W
.03
1
RHD
*01W
.05
RHD
*01W
.05
1
Wea
k D
var
iant
RHD
*01W
.01
22
RHD
*01W
.01
and
RHD
*06.
021
RHD
*01W
.01
and
RHD
*03N
.01
1
RHD
*01W
.03
2
RHD
*01W
.05
3
RHD
*01E
L.01
RHD
*01E
L.01
3
RHD
*01E
L.02
RHD
*01E
L.02
2
Del
var
iant
RH
D*0
1EL.
02
2
D- n
ull a
llele
s
RHD
*ψRH
D*ψ
RHCE
*ce-
.D(9
)-ce
30
RHD
*ψRH
CE*c
e-D
(9)-
ce a
nd R
HCE
*01.
204
Chap
ter 3
Genotyping of RHD variants and zygosity
67
Tabl
e 3.
(Con
tinue
d)
Sele
cted
don
ors
RH-M
LPA
con
clus
ion
Add
ition
al g
enot
ypin
g§Va
rian
t alle
le
dete
cted
in
test
ed c
ases
Sero
logy
and
RH
D-M
PX P
CR†
Sequ
ence
‡RH
DRH
CERH
DRH
CE
RH
D*ψ
RHCE
*ce-
D(9
)-ce
and
RH
CE*0
1.04
or
RHCE
*01.
05RH
CE*0
1.04
1
RHD
*ψ (h
omoz
ygou
s)RH
CE*c
e-D
(9)-
ce (h
omoz
ygou
s)3
RHD
*ψ a
nd R
HD
*03N
.01
RHCE
*ce-
D(9
)-ce
5
RHD
*ψ a
nd R
HD
*03N
.01
RHCE
*ce-
D(9
)-ce
and
RH
CE*0
1.20
.06
1
RHD
*01N
.08
RHD
*01N
.08
3
D n
egat
ive‡
‡
RHD
*01
N
ew v
aria
nt
RHD
*443
G
1
RHD
chi
mer
a
Chi
mer
a
Chi
mer
a 50
% R
HD
1
Wea
k D
var
iant
C
him
era
30%
RH
D
1
RHCE
var
iant
al
lele
s
RH
CE*0
2.08
.01
RH
CE*0
2.08
.01
3
ceH
AR
RHCE
*01.
221
RhC
E va
riant
RHD
*10.
00, R
HD
*10.
01, R
HD
*10.
02,
RHD
*10.
03, R
HD
*10.
06 o
r RH
D*1
0.07
RHCE
*03.
04RH
D*1
0.00
1
RH
CE*0
2.10
.01
1
† RH
D a
nd R
HCE
var
iant
alle
le d
eter
min
atio
n af
ter s
erol
ogy
and/
or R
HD
-MPX
-PC
R re
sults
‡ RH
D a
nd R
HCE
var
iant
alle
les
that
wer
e ch
osen
for t
heir
spec
ific
geno
type
§ Se
quen
cing
was
per
form
ed w
hen
the
RH-M
LPA
was
not
abl
e to
det
erm
ine
the
spec
ific
subt
ype
of th
e va
riant
alle
le
†† S
erol
ogy
and
the
RHD
-MPX
-PC
R ar
e no
t abl
e to
dis
tingu
ish
betw
een
the
DA
R1, D
AR2
and
Wea
k D
type
29
varia
nt a
llele
s‡‡
Thi
s pa
tient
was
incl
uded
in o
ur s
erie
s, be
caus
e he
r cur
rent
D s
erol
ogy
(D-)
was
dis
cord
ant w
ith a
pre
viou
sly
repo
rted
D+
phe
noty
pe b
y th
e bl
ood
bank
of A
zerb
aija
n
Chapter 3
68
Zygosity analysis showed the presence of one RHD allele and sequencing confirmed the presence of the mutations. Unfortunately, no red blood cells [RBCs] were available for further serologic typing. In 23 cases the RH-MLPA detected the presence of a variant allele, but could not discriminate between different subtypes (e.g., RHD*10 alleles and RHD*09.03 or RHD*09.04) and the samples were sequenced to assign the specific RHD or RHCE variant subtype (Table 3). In three cases in which serology indicated the presence of an Rh variant, the RH-MLPA indicated the presence of a single normal RHD*01 allele. Sequencing of all RHD exons revealed that one case concerned a hemizygously present RHD*16.02 allele (c.676G>C), for which no RHD-specific probe combination is included. In the two other cases two new RHD variant alleles were detected. One new RHD variant allele has the c.525C>A (p.Phe175Leu) mutation and the other RHD variant allele has the c.443C>G (p.Thr148Arg) mutation. The latter was determined as D- via the absorption-elution technique using a polyclonal anti-D (anti-D bromelain, Sanquin Reagentia). This patient was included in our series, because her current D serology (D-) was discordant with a previously reported D+ phenotype by the blood bank of Azerbaijan. Probably the donor was regarded as D+ because of the CE phenotype (RhCcee). The new RHD*525A (p.Phe175Leu) variant allele results in absence of expression of Epitope 1.2 (determined with monoclonal antibodies [MoAb] LHM174/102 and LHM70/45 of the extended partial RhD typing set of Bio-Rad Laboratories) and Epitope 2.2 (determined with MoAb 5C839). The LHM169/81 MoAb (extended partial RhD typing set), which detects Epitope 1.1 showed the same strength of reactivity for both the D+ control and the RBCs expressing the RHD*525A (p.Phe175Leu) allele.40 Similarly, all other D epitopes tested were normally present. The patient was typed positive for the C, c, and e antigens. In two other cases no RHD
variant allele was detected, but an aberrant RHD copy number of 0.3 and 0.5, respectively, was detected. These samples are further discussed below; see “RHD and RHCE zygosity” results. For three variant alleles a loss of signal of a wild-type probe combination was obtained, whereas presence of the RHD-specific nucleotide was shown by sequencing. In three cases carrying the RHD*25 or the new partial RHD allele (c.150T>C, c.178A>C, c.201G>A, c.203G>A, c.307T>C, c.1063G>A), the D07_1061T signal was absent. Apparently, the c.1063G>A mutation present in these alleles impaired the complete hybridization and ligation of D07_1061T. In the 46 cases carrying the RHD*Ψ allele, the D04_514A signal was absent. The 37-bp duplication in intron 3 (c.487-19) present in the RHD*Ψ allele prevents the hybridization of the D04_514A probe combination, because the insertion results in a shortening of the hybridization site to only 13 nucleotides instead of the normal 34 nucleotides. All 46 cases carrying the RHD*Ψ allele were also associated with an unexpected gain of the D09_1193A signal. The RHD and RHCE Exon 9 were therefore PCR amplified in three cases that were homozygous for the RHD*Ψ allele. In these cases RHCE Exon 9 could not be amplified, while RHCE Exons 8 and 10 were normally amplified and sequencing of these exons and surrounding introns showed RHCE wild-type sequence. We therefore hypothesize that in this RHCE variant allele, linked to
Chap
ter 3
Genotyping of RHD variants and zygosity
69
the RHD*Ψ allele, RHCE Exon 9 is replaced by RHD Exon 9 resulting in a RHCE*ce-D(9)-ce allele. This hybrid RHCE allele is normally expressed because these three homozygous cases were all serologically positive for the c and e antigens and also no abnormal reaction patterns have been described for D- black Africans. As no product was amplified in the RHCE Exon 9 PCR, the break points of the mutated RHCE allele are located outside the range of at least one of our RHCE Exon 9 primers. To confirm the presence of this hybrid allele in cases which have only one RHD*Ψ allele and hence are heterozygous for the RHCE*ce-D(9)-ce allele, we developed an MLPA probe combination for the RHCE specific c.1193T SNP. Seven cases hemizygous for the RHD*Ψ allele and one case heterozygous for this allele were tested with the CE09_1193T probe combination. In all tested samples the CE09_1193T showed a copy number of 1, indicating that these persons were indeed heterozygous positive for the RHCE*ce-D(9)-ce allele (data not shown).
RHD and RHCE zygosityRHD and RHCE zygosity were determined for all 236 DNA samples by RH-MLPA. Of these cases, 163 cases were found to be hemizygous for the RHD gene (Dd), 58 cases homozygous for the RHD gene (DD), and in 12 cases both RHD genes were deleted (dd). To confirm the zygosity determined by the RH-MLPA, 18 cases (seven DD and eleven Dd) were compared with zygosity typing using RHD Exons 5- and 7-specific quantitative real-time PCR. The results were concordant between both assays. For two cases the RH-MLPA detected an abnormal RHD copy number of 0.5 and 0.3 for all RHD-specific probe combinations, indicating the presence of, respectively, 50 and 30% hemizygously RHD*01 (RhD+). D serology showed a mixed field reaction in the case with an RHD-copy number of 0.5 and the case with a copy number of 0.3 was serologically determined as “weak D.” Further genetic analysis using short-tandem-repeat multiplex PCR showed that these samples were from individuals with a hematopoietic chimerism. To evaluate the sensitivity of the RH-MLPA to detect hematopoietic chimerism, DNA from a person hemizygous for RHD*01
(CcDdEe) was mixed with DNA from a person homozygous for RHD*01N.01 (ccddee). As shown in Figure 1 the presence of 0.25% hemizygous RHD*01 DNA is detected by two RHD wild-type probe combinations and the presence of 4% hemizygous RHD*01 DNA is detected by all RHD
wild-type probe combinations. When 2% or more hemizygous RHD*01 DNA is present the RH-
MLPA can reliably detect a hematopoietic chimerism and correct quantification of the amount of the RhD chimerism is possible when 8% or more hemizygous RHD*01 DNA is present. In 207 of the 236 cases (88%) an RHCE copy number of 2 was obtained for all RHCE-specific probe combinations. In the other cases, a copy number of 2 was obtained for all RHCE-specific probe combinations except for the CE05_676C and CE05_676G probe combinations that detect RHCE*E and RHCE*e, respectively. The abnormal RHCE*E/RHCE*e Exon 5 copy number of 0, 1, or 3 could be explained in all cases. All cases with a RHCE*01.04, RHCE*01.08, or RHCE*01.22
Chapter 3
70
ccddee
0.25%0.50% 1% 2% 4% 8%
16%32%
64%
CcDdEe
0.00
0.25
0.50
0.75
1.00
5'UTR
exon 2
exon 3
exon 4
exon 5
exon 6
exon 7
exon 9
exon 10
Legend
Percentage of CcDdEe DNA added to ccddee DNA
Prob
e si
gnal
(com
pare
d to
CcD
dEe
sam
ple)
Figure 1. RH-MLPA results obtained with a mixture of hemizygous RHD*01 D+ DNA from a donor pheno- and genotyped as (CcDdEe) mixed with homozygous D- RHD*01N.01 DNA typed as (ccddee). DNA was added in a dilution range from 64% to 0.25% RHD*01 in RHD*01N. The ratio of the RHD wild-type probe combinations is calculated using the hemizygous RHD*01 (CcDdEe) sample as a reference sample. For exons of which more than one set of RHD wild-type probe combinations is detected, the mean of the probes is given. Error bars indicate the standard deviation. Two RHD wild-type probe combinations detected the presence of 0.25% RHD*01. The presence of 2% RHD*01 is reliably detected by the RH-MLPA and the presence of 4% RHD*01 is detected by all RHD wild-type probe combinations.
allele have an RHCE*E/RHCE*e Exon 5 copy number of 1 (heterozygous variant allele) or 0 (homozygous variant allele), because they all contain the c.712A>G mutation, which impairs ligation of the second ligation site specific for the c.712A RHCE wild-type SNP of CE05_676C or CE05_676G. In 20 cases an elevated RHCE*E/RHCE*e Exon 5 copy number of 3 was obtained. All these cases carried next to their RHCE alleles an RHD-CE-D hybrid allele that contained a complete RHCE Exon 5. Therefore, the CE05_676G (including the RHD*05.07, RHD*06.02, and RHD*03N.01 allele) or CE05_676C (RHD*06.01) was able to bind to the hybrid allele, causing an elevated RHCE*E/ RHCE*e Exon 5 copy number.
Chap
ter 3
Genotyping of RHD variants and zygosity
71
Discussion
In this study we showed that the RH-MLPA genotyping assay is a reliable method to predict the Rh-phenotype and to determine the majority of RHD and RHCE variant alleles as well as RHD zygosity. The MLPA technique is easy to use, as it only requires a thermocycler and capillary electrophoresis equipment.First, the specificity of the RHD and RHCE wild-type and RHD and RHCE mutation probe combinations was determined with plasmid DNA and subsequently with genomic DNA. All RHD and RHCE wild-type and mutation probe combinations detected the wild-type or mutated sequence according to their design. We showed two exceptions of wild-type probe combinations in which the loss of signal was not caused by mutation at the ligation site, but due to a mutation close by the ligation site: absence of D07_1061T in variant alleles with c.1063G>A mutation and absence of D04_514A in RHD*Ψ alleles.To evaluate the accuracy of the RH-MLPA assay its performance was tested with a set of 163 cases that were previously serologically typed for Rh and in some cases already typed with an RHD-MPX PCR or by RHD exon–specific sequencing. In 160 cases (98%) the RH-MLPA assigned correctly if a wild-type and/or variant allele(s) was (were) present. In one case sequencing revealed the presence of a partial RHD allele (RHD*16.02), for which no probe was included in the RH-MLPA because it is extremely rare.41 It is still possible to add an extra mutation probe for detection of the RHD*16.02 allele. In the other two cases, two new variant RHD
alleles were found: a D- null allele RHD*443G (p.Thr148Arg) and a partial RHD allele RHD*525A
(p.Phe175Leu). We therefore conclude that the RH-MLPA can be used to determine an RHD
and/or RHCE variant in the vast majority of samples analyzed in a reference laboratory. Only in few samples follow-up analysis is needed to show the presence of either a very rare or a new variant allele. In all 254 variant alleles detected by the RH-MLPA the correct variant or variant group was assigned. In one case the RH-MLPA could even determine a new partial RHD allele (c.150T>C, c.178A>C, c.201G>A, c.203G>A, c.307T>C, and c.1063G>A). In 26 of the 254 variant alleles (10%) detected by the RH-MLPA, additional sequencing was necessary to determine the specific subgroup. Only in the RHD*10 variant group (n = 4) determination of the subtype is clinically relevant, because the RHD*10.00 allele, with normal RhD expression, should be discriminated from the other RHD*10 alleles.A new RHD*443G (p.Thr148Arg) allele was detected in this study. This seems to be a D- null allele, caused by a mutation resulting in the substitution of a polar amino acid (threonine) with a positively charged amino acid (arginine) in the fifth transmembrane region of the RhD protein. To confirm this unexpected profound effect an RhD expression model should be performed.42 For the first time it was recognized that the RHD*Ψ allele is linked (in all 46 persons tested) to a hybrid RHCE*ce variant allele containing RHD Exon 9 (RHCE*ce-D(9)-ce). This could be determined with the RH-MLPA, because it determines the RHD copy number of Exon 9. This
Chapter 3
72
hybrid allele contains two mutations c.1170C>T and c.1193T>A, encoding for one amino acid change p.Glu398Val at the intracellular C-terminal tail of the Rhce protein. Serologic D and CE typing in persons homozygous positive for the RHD*Ψ RHCE*ce-D(9)-ce haplotype showed the normal presence of the c and e antigens and the absence of all D epitopes. The amino acid change is present in the intracellular tail of the protein and thus far no effect on the expression of the RHCE epitopes has been shown.21
The RH-MLPA assay is very accurate in determining gene copy number variation. In the RH-MLPA the RHD copy number is based on the signals derived from 17 RHD wild-type probe combinations. This makes the RH-MLPA more suitable for RHD and RHCE zygosity determination than real-time quantitative PCR30 or amplification of the hybrid Rh box.31-33 Clinically, it is important to determine RHD zygosity in fathers, to assess the need for fetal RHD typing in RhD alloimmunized D- women. Furthermore, because the RH-MLPA is highly accurate in the determination of exon copy number, this is the first high-throughput assay that is able to recognize hybrid RHD/RHCE alleles next to a normal RHD gene. Next to the “normal” zygosity scores of 0, 1, and 2, the RH-MLPA is also able to reliably determine the presence of hematopoietic chimerism of just 2% RHD*01 (RhD+) in an RHD*01N.01 (RhD–) background. It is important to detect RHD+/RHD- chimeras, because it has been shown that transfusion of RBCs from a donor with a hematopoietic chimerism of 6% RHD+ DNA was able to immunize two D- recipients.43 The RH-MLPA can also reliably genotype patients who received multiple non-leukoreduced RBC units. After massive transfusions a (weak) signal derived from donor white blood cells might be detected with the RH-MLPA.44 Because the MLPA is a quantitative method, it will be readily recognized if a signal is obtained from a minor population of transfused white blood cells.Some variants were correlated with a loss or gain of the RHCE*E/RHCE*e Exon 5 copy number. In the RHCE*01.04, RHCE*01.05, RHCE*01.08, and RHCE*01.22 alleles the correct binding of the CE05_676G and CE05_676C probe combination is inhibited and therefore it is not possible to determine whether these variant alleles contain the RHCE*E or RHCE*e variation. Because the RHCE*01.04, RHCE*01.05, RHCE*01.08, and RHCE*01.22 alleles result in partial e antigen expression, an individual carrying one of these alleles should always be determined as RHCE*e+, hence Rhe+.45-47 For the RHD*06 and RHD*03N.01 hybrid alleles in which the RH-
MLPA suggest an RHCE*E/RHCE*e Exon 5 copy number of three it is known that the hybrid alleles result in the expression of the e antigen (RHD*06.02 and RHD*03N.01 allele) or the E antigen (RHD*06.01 allele).48 For these cases the presence of the RHCE*e and/or RHCE*E SNP present in the hybrid allele should be taken into account when determining the RhEe status. For the precise determination of variant RHCE alleles and the identification of rare RHCE variant alleles an additional MLPA has to be developed, but as listed in Table 2 the most frequently occurring RHCE variant alleles are recognized with the current RH-MLPA.
Chap
ter 3
Genotyping of RHD variants and zygosity
73
In conclusion, the RH-MLPA genotyping assay correctly determines clinically relevant RHD
and RHCE variant alleles. Therefore, it can be used as a single and rapid assay to facilitate comprehensive typing of RHD and RHCE variants in blood recipients and blood donors, thereby providing a safe basis for blood transfusion. The need for extensive RHD genotyping, by for instance the RH-MLPA assay, of blood recipients, blood donors, and pregnant women is still growing, because molecular genotyping methods are increasingly applied and therefore the number of cases with discrepant serologic and genotyping results that require further molecular analysis will increase.
Acknowledgements
We thank Peter Ligthart (Sanquin Diagnostic Services, Amsterdam, the Netherlands) for his technical assistance, Karel de Groot (MRC Holland, Amsterdam, the Netherlands) for assistance in designing the MLPA probe mixes, Christof Weinstock (Institute for Clinical Transfusion Medicine and Immunogenetics Ulm, Ulm, Germany), Inge von Zabern (Institute for Clinical Transfusion Medicine and Immunogenetics Ulm, Ulm, Germany), Geoff Daniels (Bristol Institute for Transfusion Sciences, National Health Service Blood and Transplant, Bristol, UK),Martin Pisacka (Institute of Haematology and Blood Transfusion, Prague, Czech Republic), and Yanli Ji (Guangzhou Blood Center, Guangzhou Guangdong, China) for providing DNA samples with specific RHD genotypes.
Web Resources
ISBT Working Party on Red Cell Immunogenetics and Blood Group Terminology http://www.isbtweb.org/working-parties/red-cell-immunogenetics-and-bloodgroup-terminology/blood-group-terminology/bloodgroup-allele-terminology/ Accessed at 05-10-2012.
Chapter 3
74
References1. Daniels G, Reid ME. Blood groups: the past 50 years. Transfusion 2010;50:281-9.
2. Westhoff CM. Rh complexities: serology and DNA genotyping. Transfusion 2007;47 Suppl:17S-22S.
3. Urbaniak SJ, Greiss MA. RhD haemolytic disease of the fetus and the newborn. Blood Rev 2000;14:44-61.
4. Daniels G, Poole J, de Silva M, Callaghan T, MacLennan S, Smith N. The clinical significance of blood group antibodies. Transfus Med 2002;12:287-95.
5. Avent ND, Martinez A, Flegel WA, Olsson ML, Scott ML, Nogues N, Pisacka M, Daniels GL, Muniz-Diaz E, Madgett TE, Storry JR, Beiboer S, Maaskant-vanWijk PM, von Zabern I, Jiménez E, Tejedor D, López M, Camacho E, Cheroutre G, Hacker A, Jinoch P, Svobodova I, van der Schoot E, de Haas M. The Bloodgen Project of the European Union, 2003-2009. Transfus Med Hemother 2009;36: 162-7.
6. Moise KJ. Red blood cell alloimmunization in pregnancy. Semin Hematol 2005;42:169-78.
7. Wiener AS, Unger LJ. Rh factors related to the Rho factor as a source of clinical problems; diagrammatic representation of their reactions and prediction of still undiscovered Rh factors. J Am Med Assoc 1959;169:696-9.
8. Cherif-Zahar B, Mattei MG, Le Van Kim C, Bailly P, Cartron JP, Colin Y. Localization of the human Rh blood group gene structure to chromosome region 1p34.3-1p36.1 by in situ hybridization. Hum Genet 1991;86:398-400.
9. Arce MA, Thompson ES, Wagner S, Coyne KE, Ferdman BA, Lublin DM. Molecular cloning of RhD cDNA derived from a gene present in RhD-positive, but not RhD-negative individuals. Blood 1993;82:651-5.
10. Cherif-Zahar B, Bloy C, Le Van Kim C, Blanchard D, Bailly P, Hermand P, Salmon C, Cartron JP, Colin Y. Molecular cloning and protein structure of a human blood group Rh polypeptide. Proc Natl Acad Sci U S A 1990;87:6243-7.
11. Le Van Kim C, Mouro I, Cherif-Zahar B, Raynal V, Cherrier C, Cartron JP, Colin Y. Molecular cloning and primary structure of the human blood group RhD polypeptide. Proc Natl Acad Sci U S A 1992;89:10925-9.
12. Simsek S, de Jong CA, Cuijpers HT, Bleeker PM, Westers TM, Overbeeke MA, Goldschmeding R, van der Schoot CE, von dem Borne AE. Sequence analysis of cDNA derived from reticulocyte mRNAs coding for Rh polypeptides and demonstration of E/e and C/c polymorphisms. Vox Sang 1994;67:203-9.
13. Avent ND, Ridgwell K, Tanner MJ, Anstee DJ. cDNA cloning of a 30 kDa erythrocyte membrane protein associated with Rh (Rhesus)-blood-group-antigen expression. Biochem J 1990;271:821-5.
14. Flegel WA. Molecular genetics and clinical applications for RH. Transfus Apher 2011;44:81-91.
15. Daniels G. Human blood groups. 2nd ed. Oxford: Blackwell Science; 2002.
18. McGann H, Wenk RE. Alloimmunization to the D antigen by a patient with weak D type 21. Immunohematology 2010;26:27-9.
19. Denomme GA, Wagner FF, Fernandes BJ, Li W, Flegel WA. Partial D, weak D types, and novel RHD alleles among 33,864 multiethnic patients: implications for anti-D alloimmunization and prevention. Transfusion 2005;45:1554-60.
20. Wagner T, Kormoczi GF, Buchta C, Vadon M, Lanzer G, Mayr WR, Legler TJ. Anti-D immunization by DEL red blood cells. Transfusion 2005;45:520-6.
21. Singleton BK, Green CA, Avent ND, Martin PG, Smart E, Daka A, Narter-Olaga EG, Hawthorne LM, Daniels G. The presence of an RHD pseudogene containing a 37 base pair duplication and a nonsense mutation in Africans with the Rh D-negative blood group phenotype. Blood 2000;95:12-8.
22. Colin Y, Cherif-Zahar B, Le Van Kim C, Raynal V, Van Huffel V, Cartron JP. Genetic basis of the RhD-positive and RhD-negative blood group polymorphism as determined by Southern analysis. Blood 1991;78:2747-52.
23. Faas BH, Beckers EA, Wildoer P, Ligthart PC, Overbeeke MA, Zondervan HA, von dem Borne AE, van der Schoot CE. Molecular background of VS and weak C expression in blacks. Transfusion 1997;37:38-44.
24. Reid ME, Lomas-Francis C. The blood group antigen facts book. 2nd ed. San Diego (CA): Academic Press; 2003.
25. Veldhuisen B, van der Schoot CE, de Haas M. Blood Group genotyping: from patient to high-throughput donor screening. Vox Sang 2009;97:198-206.
26. Avent ND. Large scale blood group genotyping. Transfus Clin Biol 2007;14:10-5.
Chap
ter 3
Genotyping of RHD variants and zygosity
75
27. Finning KM, Martin PG, Soothill PW, Avent ND. Prediction of fetal D status from maternal plasma: introduction of a new noninvasive fetal RHD genotyping service. Transfusion 2002;42:1079-85.
28. Scheffer PG, van der Schoot CE, Page-Christiaens GC, de Haas M. Noninvasive fetal blood group genotyping of rhesus D, c, E and of K in alloimmunised pregnant women: evaluation of a 7-year clinical experience. BJOG 2011;118:1340-8.
29. Pirelli KJ, Pietz BC, Johnson ST, Pinder HL, Bellissimo DB. Molecular determination of RHD zygosity: predicting risk of hemolytic disease of the fetus and newborn related to anti-D. Prenat Diagn 2010;30:1207-12.
30. Krog GR, Clausen FB, Dziegiel MH. Quantitation of RHD by real-time polymerase chain reaction for determination of RHD zygosity and RHD mosaicism/chimerism: an evaluation of four quantitative methods. Transfusion 2007;47:715-22.
31. Grootkerk-Tax MG, Maaskant-VanWijk PA, van Drunen J, van der Schoot CE. The highly variable RH locus in nonwhite persons hampers RHD zygosity determination but yields more insight into RH-related evolutionary events. Transfusion 2005;45:327-37.
32. Wagner FF, Flegel WA. RHD gene deletion occurred in the Rhesus box. Blood 2000;95:3662-8.
34. Maaskant-VanWijk PA, Faas BH, de Ruijter JA, Overbeeke MA, von dem Borne AE, van Rhenen DJ, van der Schoot CE. Genotyping of RHD by multiplex polymerase chain reaction analysis of six RHD-specific exons. Transfusion 1998;38:1015-21.
35. Pongers-Willemse MJ, Verhagen OJ, Tibbe GJ, Wijkhuijs AJ, de Haas V, Roovers E, van der Schoot CE, van Dongen JJ. Real-time quantitative PCR for the detection of minimal residual disease in acute lymphoblastic leukemia using junctional region specific TaqMan probes. Leukemia 1998;12:2006-14.
36. Lo YM, Tein MS, Lau TK, Haines CJ, Leung TN, Poon PM, Wainscoat JS, Johnson PJ, Chang AM, Hjelm NM. Quantitative analysis of fetal DNA in maternal plasma and serum: implications for noninvasive prenatal diagnosis. Am J Hum Genet 1998;62:768-75.
37. Rijnders RJ, Christiaens GC, Bossers B, van der Smagt JJ, van der Schoot CE, de Haas M. Clinical applications of cell-free fetal DNA from maternal plasma. Obstet Gynecol 2004;103:157-64.
38. Daniels GL, Faas BH, Green CA, Smart E, Maaskant-Van Wijk PA, Avent ND, Zondervan HA, von dem Borne AE, van der Schoot CE. The VS and V blood group polymorphisms in Africans: a serologic and molecular analysis. Transfusion 1998;38:951-8.
39. Lomas C, McColl K, Tippett P. Further complexities of the Rh antigen D disclosed by testing category DII cells with monoclonal anti-D. Transfus Med 1993;3:67-9.
40. Scott M. Section 1A: Rh serology. Coordinator’s report. Transfus Clin Biol 2002;9:23-9.
41. Flegel WA, von Zabern I, Doescher A, Wagner FF, Vytiskova J, Pisacka M. DCS-1, DCS-2, and DFV share amino acidsubstitutions at the extracellular RhD protein vestibule. Transfusion 2008;48:25-33.
42. Smythe JS, Avent ND, Judson PA, Parsons SF, Martin PG, Anstee DJ. Expression of RHD and RHCE gene products using retroviral transduction of K562 cells establishes the molecular basis of Rh blood group antigens. Blood 1996;87:2968-73.
43. Wagner FF, Frohmajer A, Flegel WA. RHD positive haplotypes in D negative Europeans. BMC Genet 2001;2:10.
44. Rozman P, Dovc T, Gassner C. Differentiation of autologous ABO, RHD, RHCE, KEL, JK, and FY blood group genotypes by analysis of peripheral blood samples of patients who have recently received multiple transfusions. Transfusion 2000;40:936-42.
45. Hemker MB, Ligthart PC, Berger L, van Rhenen DJ, van der Schoot CE, Wijk PA. DAR, a new RhD variant involving exons 4, 5, and 7, often in linkage with ceAR, a new Rhce variant frequently found in African blacks. Blood 1999;94:4337-42.
46. Noizat-Pirenne F, Lee K, Pennec PY, Simon P, Kazup P, Bachir D, Rouzaud AM, Roussel M, Juszczak G, Menanteau C, Rouger P, Kotb R, Cartron JP, Ansart-Pirenne H. Rare RHCE phenotypes in black individuals of Afro-Caribbean origin: identification and transfusion safety. Blood 2002;100:4223-31.
47. Beckers EA, Faas BH, von dem Borne AE, Overbeeke MA, van Rhenen DJ, van der Schoot CE. The R0Har RH:33 phenotype results from substitution of exon 5 of the RHCE gene by the corresponding exon of the RHD gene. Br J Haematol 1996;92:751-7.
48. Avent ND, Finning KM, Liu W, Scott ML. Molecular biology of partial D phenotypes. Transfus Clin Biol 1996;3:511-6.
Chapter 3
76
Chap
ter 3
Genotyping of RHD variants and zygosity
77
Supporting informationChapter 3
Chapter 3
78
Supplementary Figure S1. Example of analysis of mix P401. a) The blue peaks indicate the signals from the analyzed sample and red peaks indicate signals from a reference sample (CcDdEe genotype). The black arrows indicate the control-probe-combination signals and are used for normalization of both the analyzed and reference sample. The signals of the RHD-specific-probe combinations of the analyzed sample (green arrows) show the same fluorescence intensity as the signals of the reference samples, indicating that the tested sample has the same RHD copy number as the reference sample (Dd). In contrast, the signal obtained for the RHCE*e specific-probe combination (light blue arrow) shows twice the amount of fluorescence intensity compared to the reference sample, therefore the copy number of RHCE*e is double of that obtained for the reference sample, therefore the copy number of RHCE*e is 2n in the analyzed sample. The RHCE*C and RHCE*E specific-probe-combination signals (dark blue arrows) are absent in the analyzed sample, while the reference sample shows signals, indicating that these alleles are absent in the analyzed sample. The red arrow indicates the presence of the MD_819A signal in the analyzed sample and thus the presence of the RHD*09.03, RHD*09.04 or RHD*09.05 allele. b) The results are shown in the fourth column as ratios of the signals obtained for the analyzed sample compared to signals obtained for the reference sample. The Bin Size indicates the length of the probe. Interpretation of the results is the same as in a).