Vacuolar and Plasma Membrane Proton Pumps Collaborateto Achieve Cytosolic pH Homeostasis in Yeast*
Received for publication, December 22, 2007, and in revised form, April 17, 2008 Published, JBC Papers in Press, May 23, 2008, DOI 10.1074/jbc.M710470200
Gloria A. Martınez-Munoz1 and Patricia Kane2
From the Department of Biochemistry and Molecular Biology, SUNY Upstate Medical University, Syracuse, New York 13210
Vacuolar proton-translocating ATPases (V-ATPases) play acentral role in organelle acidification in all eukaryotic cells. Toaddress the role of the yeast V-ATPase in vacuolar and cytosolicpH homeostasis, ratiometric pH-sensitive fluorophores specificfor the vacuole or cytosol were introduced into wild-type cellsand vma mutants, which lack V-ATPase subunits. Transientlyglucose-deprived wild-type cells respond to glucose additionwith vacuolar acidification and cytosolic alkalinization, andsubsequent addition of K� ion increases the pH of both the vac-uole and cytosol. In contrast, glucose addition results in anincrease in vacuolar pH in both vmamutants andwild-type cellstreatedwith theV-ATPase inhibitor concanamycinA.CytosolicpH homeostasis is also significantly perturbed in the vmamutants. Even at extracellular pH5, conditions optimal for theirgrowth, cytosolic pH was much lower, and response to glucosewas smaller in themutants. In plasmamembrane fractions fromthe vma mutants, activity of the plasma membrane protonpump, Pma1p, was 65–75% lower than in fractions from wild-type cells. Immunofluorescence microscopy confirmeddecreased levels of plasma membrane Pma1p and increasedPma1p at the vacuole and other compartments in the mutants.Pma1p was not mislocalized in concanamycin-treated cells, buta significant reduction in cytosolic pH under all conditions wasstill observed.We propose that short-term, V-ATPase activity isessential for both vacuolar acidification in response to glucosemetabolism and for efficient cytosolic pH homeostasis, andlong-term, V-ATPases are important for stable localization ofPma1p at the plasma membrane.
The importance of V-ATPases3 for acidification of the vacu-ole/lysosomes, Golgi apparatus, and endosomes of eukaryoticcells is well established (1, 2). Multiple cellular processes,including secondary transport of ions andmetabolites, matura-tion of iron transporters, endocytic and biosynthetic proteinsorting, and zymogen activation depend on compartment acid-
ification and have been linked to V-ATPase activity (1, 3). Insome cells such as macrophages, V-ATPases play specializedroles that clearly include regulation of cytosolic pH (4, 5). How-ever, although V-ATPases pump protons from the cytosol intoorganelles in all cells, they are not generally believed to play amajor role in cytosolic pH regulation.The yeast Saccharomyces cerevisiae has emerged as a major
model system for eukaryotic V-ATPases. One reason for this isthat yeast mutants lacking all V-ATPase activity (vmamutants)are viable, but loss of V-ATPase activity in eukaryotes otherthan fungi is lethal (6–9). Yeast vmamutants do exhibit a set ofdistinctive phenotypes, however, that includes the inability togrow at pH values lower than 3 or higher than 7 and sensitivityto high extracellular calcium concentrations (2). This Vma�
phenotype suggests a perturbation of pH homeostasis in thesecells that is not fully understood. It has been suggested that vmamutants survive at low extracellular pH (pH 5) by endocytosisof acidic extracellular fluid and transport to the vacuole (6, 10)or that they acidify the vacuole through diffusion of permeantacids (11). There have been few direct measurements of cyto-solic or vacuolar pH in the vmamutants under different extra-cellular conditions, however (11).pH homeostasis is critical for survival of yeast cells, as it is for
all eukaryotic cells. V-ATPases function in tandemwithPma1p,an essential P-type proton pump localized to the plasma mem-brane, to help control pH (12, 13). Glucose, the preferred car-bon source for S. cerevisiae, is metabolized through fermenta-tion and respiration. Enormous amounts of carbonic andorganic acids are produced by energy metabolism, and theseconstitute the main source of protons in yeast (13, 14). Despitemajor differences in localization and structure, there are a num-ber of similarities in regulation and function between Pma1pand V-ATPases. Both Pma1p and the V-ATPase use energyfrom ATP hydrolysis to pump protons out of the cytosol (13).Both pumps are regulated by extracellular glucose, althoughtheir regulatory mechanisms are distinct. Glucose stimulatesPma1 H�-ATPase activity by inducing phosphorylation (15),resulting in a decreased Km for ATP and an increased Vmax (16,17). V-ATPases are also activated by glucose, but they revers-ibly disassemble in response to glucose deprivation and readdi-tion, resulting in higher levels of assembled pumps and ATPaseactivity at the vacuole in the presence of glucose (18, 19). Theactivity of both pumps is sensitive to pH, indicating that theymay be tuned to respond to changes in cytosolic pH in vivo (20,21). Pma1p, in particular, is activated in response to cytosolicacidification (21), and this mode of regulation is likely to beimportant for maintenance of cytosolic pH in a narrow range,relatively independent of extracellular pH. Taken together,
* This work was supported, in whole or in part, by National Institutes of HealthGrant GM50322 (to P. M. K.). The costs of publication of this article weredefrayed in part by the payment of page charges. This article must there-fore be hereby marked “advertisement” in accordance with 18 U.S.C. Sec-tion 1734 solely to indicate this fact.
1 Current e-mail address: [email protected] To whom correspondence should be addressed: Dept. of Biochemistry and
Molecular Biology, SUNY Upstate Medical University, 750 East Adams St.,Syracuse, NY 13210; Tel.: 315-464-8742; Fax: 315-464-8750; E-mail:[email protected].
3 The abbreviations used are: V-ATPase, vacuolar proton-translocatingATPase; BCECF-AM, 2�,7�-bis(2-carboxyethyl)-5-(and-6)-carboxyfluores-cein, acetoxymethyl ester; MES, 2-[N-morpholino]ethanesulfonic acid;MOPS, 2-[N-morpholino]propanesulfonic acid; TEA, triethanolamine.
these data suggest that in addition to their parallel roles inremoval of protons from the cytosol, these two pumps areresponsive to similar metabolic stimuli.Both Pma1p and the V-ATPase are also electrogenic pumps.
Each establishes a pH gradient and membrane potential (��)critical for operation of other transporters in their respectivemembranes but also requires mechanisms for balancing thegeneration of membrane potential (22, 23). Potassium ion isprimarily responsible for balancing the plasma membranepotential in yeast (24). Transport of K� ion into the cell resultsin a depolarization of the plasma membrane, allowing stimula-tion of Pma1p (22) and consequent cytosolic alkalinization (25,26), and pH-responsive regulation of the Trk1p potassiumtransporter via phosphorylation provides additional means ofactivating Pma1p when cytosolic pH drops. The exact mecha-nisms for balancing the membrane potential at the vacuolarmembrane are less clear; many anion channels and other trans-porters are present in the vacuolar membrane that could con-tribute to the final potential, but their individual contributionsto membrane potential are not well defined.In addition to the primary proton pumps,multiple transport-
ers and buffers combine to determine the final cytosolic andvacuolar pH under various conditions. Both the plasma mem-brane and organellar membranes contain alkali cation/H�
exchangers that contribute to overall pH homeostasis (27, 28).In mammalian cells where there is no primary proton pumpcomparable with Pma1p; the plasma membrane NHE (Na�/H�) antiporters play a central role in pH homeostasis (29).Yeast cells also haveNa�/H� exchangers (Nha1p) at the plasmamembrane, but their function in pH homeostasis is clearly sec-ondary to that of Pma1p (27). In addition, bothmammalian andyeast cells contain organellar K�(Na�)/H� exchangers. Thefoundingmember of this class of exchangers, yeast Nhx1p (28),localizes to the yeast endosome and has clearly been shown toplay a role in regulating both vacuolar and cytosolic pH (30, 31).The existence of a distinct vacuolar K�(Na�)/H� exchangerwas inferred from physiological studies (32) and was recentlyproposed to be the VNX1 gene product (33). Both small mole-cule and protein buffers are also likely to contribute to pHhomeostasis in the cytosol and vacuole. In addition to thepotential role for permeant acids in the vma mutants, high-lighted by Plant et al. (11), it is notable that the vacuole containsvery high concentrations of polyphosphate that can providebuffering capacity (34, 35). It has also been proposed that vac-uolar proteins are well suited to buffer the vacuole to its typicalpH range (36).Several pieces of data suggest that the plasmamembrane and
vacuolar proton pumps Pma1 and V-ATPase are functionallyinterdependent. Bowman andBowman (37) isolatedmutants inNeurospora crassa that were resistant to the specific V-ATPaseinhibitor concanamycin A and discovered that they containedmutations in Pma1p. Mislocalization of Pma1 from the plasmamembrane to the endoplasmic reticulum (38) or the vacuole(39) in the vmamutants has been reported, but the physiolog-ical implications of the altered localization have not been clar-ified. To address a potential coordination of function betweenthe V-ATPase and Pma1p, we examined pHhomeostasis undervaried extracellular conditions in two vmamutants, vma2� and
vma3�. Both mutants lack all V-ATPase activity, but thevma2�mutation deletes a subunit of the peripheral V1 sector ofthe enzyme, whereas the vma3� mutation deletes the majorproteolipid subunit of the proton-pore containing theVo sector(2). Measurements of cytosolic and vacuolar pH indicate thatwild-type cells readjust pH in response to the addition of glu-cose and K� ion, as expected but that the vmamutations havedramatically perturbed pHhomeostasis in both the vacuole andthe cytosol. Mislocalization of Pma1p may account for some ofthe defects in the vma mutants, but even an acute loss ofV-ATPase activity in the presence of concanamycinA abolishesvacuolar pH responses and perturbs cytosolic pH homeostasis.These results suggest an unexpectedly large role for theV-ATPase in cellular pH homeostasis.
EXPERIMENTAL PROCEDURES
Materials—2�,7�-Bis(2-carboxyethyl)-5-(and-6)-carboxy-fluorescein, acetoxymethyl ester (BCECF-AM) was purchasedfrom Invitrogen, and the yeast pHluorin plasmid was a gener-ous gift from Dr. Rajini Rao (Johns Hopkins University). Poly-clonal anti-Pma1pwas a generous gift fromDr. Ramon Serrano.Monoclonal antibody against Pma1p (40B7) was obtained fromAbcam, and monoclonal antibodies against Pep12p and alka-line phosphatase were purchased from Invitrogen. Alexa Fluor488 goat anti-mouse IgG used for immunofluorescence waspurchased from Invitrogen. Vanadate was purchased fromSigma and activated by heating at alkaline pH as described (40).Concanamycin A and other chemicals were purchased fromSigma.Strains and Culture Conditions—Cells were grown in YEPD
medium (1% yeast extract, 1% bactopeptone, 2% glucose) buff-ered to pH 5 or 7.5 with 50 mM potassium phosphate, 50 mMpotassium succinate as described (41). The BY4741 wild-type strain (MAT� his3�1 leu2�0 met15�0 ura3�0; OpenBiosystems) and the congenic vma2�::kanMX andvma3�::kanMX mutants were used throughout, except forthose experiments requiring a pep4mutants strain. For theseexperiments the SF838-1D� wild-type (MAT� ura3-53leu2-3, 112 his4-519 pep4-3 ade6) and the congenicvma2�::LEU2 and vma3�::URA3 mutants were used (42).Cytosolic and Vacuolar pH Measurements—Cytosolic pH
was measured using a pH-sensitive green fluorescent protein,yeast pHLuorin, as described by Brett et al. (31). BY4741 wild-type and vma mutant cells were transformed with the yeastpHLuorin-containing plasmid (31), and transformants wereselected in fully supplemented minimal medium lackingleucine (SD�leucine) and confirmed by fluorescence micros-copy (31). Cells containing the pHLuorin plasmid were grownto log phase (A600 � 0.5–0.6) in YEPD buffered to the desiredpH. Cells were collected by centrifugation and washed 2–3times in YEP (YEPDmediumwithout glucose) buffered at pH 5and suspended at 1 g of wet cell mass/ml in the same medium.For pH shift experiments, cells were grown to log phase asdescribed above, but cells were harvested at an absorbance of0.5, shifted toYEPD, pH7.5, and incubated for 3–4more hours.The cells were washed and resuspended in YEP pH 7.5. For pHmeasurements, 25 �l of cell suspension was added to 2 ml of 1mMMES/triethanolamine (TEA), pH 5, orMOPS/TEA, pH 7.5,
V-ATPases Contribute to Cytosolic pH Control in Yeast
20310 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 29 • JULY 18, 2008
and the mixture was stirred in a cuvette. Cytosolic pHresponses were recorded at 30 °C (a) after 5 min of stirring, (b)after the addition of glucose to 50 mM final concentration andan additional 5 min stirring, and (c) after the addition of KCl to50mM final and an additional 3min of incubation. Fluorescenceintensity at excitation wavelengths 405 and 485 nm was meas-ured in triplicate for each sample at a constant emission wave-length of 508 nm in a SPEX Fluorolog-3–21 fluorometer. Cali-bration of fluorescence with pH was carried out for each strainin each experiment as described (31). Calibration curves wereconstructed for every strain and included buffers titrated to 5,5.5, 6.0, 6.2, 6.5, 6.7, 7.0, and 7.5.Vacuolar pH measurement used the pH-sensitive ratiomet-
ric dye BCECF-AM as described (11, 30, 31). Briefly, cells weregrown to log phase in YEPD, pH 5 media, and collected bycentrifugation, and 100 mg of cells mass were resuspended in100 �l of the same medium. For experiments at extracellularpH 7.5, cells were washed and resuspended in YEP pH 7.5. Cellswere incubated in 50 �M BCECF-AM for 30 min at 30 °C withshaking. The cells were then washed 2–3 times with YEP, pH 5or 7.5 (depending on the experiment), to remove the dye andresuspended at the same density in the same media. For vacu-olar pHmeasurements, 25 �l of cell suspension was added to 2ml of 1 mM MES/TEA, pH 5, or MOPS/TEA, pH 7.5, and themixture was stirred in a cuvette. Response of vacuolar pH wasrecorded at 30 °C with stirring as described above, except thatfluorescence intensity at excitation wavelengths 450 and 490nm was measured in triplicate for each sample at a constantemission wavelength of 535 nm. Calibration of fluorescencewith pH was carried out for each strain as described (30, 31).Values are expressed as themean� S.E. for each condition, andstatistic significance was taken as p� 0.05 of identical means inthe two-sample t test for paired means.Proton Export—Proton pumping across the plasma mem-
brane was measured by recording extracellular pH changeswith a pH meter (Beckman Selection 2000) (43). Cells weregrown to log phase in YEPD, pH 5medium, washed, and resus-pended as described above. 25 �l of the cell suspension wasincubated in 15 ml of buffer (1 mM MES/TEA, pH 5), and theextracellular pH was monitored for 16 min at 30 °C with shak-ing. Extracellular pH was recorded manually every 30 s, andglucose (to 40mM final concentration) and KCl (to 40mM final)were added after 3 and 8 min of incubation, respectively.Isolation of Plasma Membrane—Preparation of total extract
and plasmamembrane from yeast was done according to Pana-retou and Piper (44) using wild-type and vma cells from theSF838-1D� strain background. One liter of cells in log phase(absorbance 0.6–0.8) were harvested by centrifugation (5 min,5000 � g), then resuspended in 80 ml of 0.4 M sucrose in bufferA (25 mM imidazole-HCl, pH 7, containing 1 �g/ml pepstatinA, 2�g/ml chymostatin, 1mMphenylmethylsulfonyl fluoride, 5�g/ml aprotinin, and 1 �g/ml leupeptin). The cell suspensionwas divided between two50-ml centrifuge tubes andpelleted bycentrifugation (5000� g for 10min). Glass beads were added totwice the pellet volume of cells followed by enough 0.4 Msucrose in buffer A to cover the cells and glass beads. Cells weresubjected to vortex mixing for three periods of 2.5 min each,with pauses of 30 s on ice. To pellet unbroken cells and glass
beads, suspension was diluted three times in 0.4 M sucrosebuffer A and centrifuged at 530 � g for 20 min. Supernatantfrom this step was then centrifuged at 22,000 � g for 30 min.The pellet was resuspended in 2 ml of buffer A by gentle vor-texing (30 s), and 1-ml aliquots of resuspended membraneswere loaded onto a discontinuous sucrose gradient (preparedby overlaying three 4-ml layers of 2.25, 1.65, and 1.1 M sucrose,all in buffer A) and centrifuged overnight (14 h) at 80,000� g inthe Beckman SW40Ti rotor. Membranes banding at the 2.25/1.65 M are enriched for plasma membrane. This band was col-lected, diluted 4 times with buffer A, and pelleted at 30,000 � gfor 40 min. The pellet was resuspended in buffer B (25 mMimidazole-HCl, pH 7, 50% (v/v) glycerol) containing the sameprotease inhibitors and stored at �20 °C.PlasmamembraneH�-ATPase activity was assayed inmem-
brane fractions with or without a 5-min preincubation with 50�M vanadate at room temperature with gentle shaking. ATPaseactivity was determined using the coupled enzyme assay (45),and vanadate-sensitive activity is reported. Typically 80% of thetotal ATPase activity in the plasma membrane fraction wasinhibited by vanadate, and there was very little concanamycinA-sensitive ATPase activity, suggesting minimal contamina-tion with vacuoles or endosomes. Protein concentrations weredetermined by the Lowry method (46).For assessment of protein levels of Pma1p, Pep12p, and alka-
line phosphatase, plasmamembrane fractions were solubilized,separated by SDS-PAGE, and transferred to nitrocellulose asdescribed (42), except that a portion of the samples to be blot-ted for Pma1p were solubilized in 100mMTris-HCl, pH 6.8, 4%SDS, 20% glycerol, and 10% �-mercaptoethanol rather thancracking buffer. Blots were probed with mouse monoclonalantibodies against Pep12p and alkaline phosphatase and rabbitpolyclonal antibodies against Pma1p.Western blot signals wererevealed with alkaline phosphatase-conjugated second anti-body (anti-mouse or anti-rabbit as appropriate), and the signalswere quantitated using ImageJ (National Institutes of Health).Equal protein concentrationswere loaded for each strain; 40�gwas loaded for detection of Pma1p, and 30 �g was loaded fordetection of alkaline phosphatase and Pep12p.Immunolocalization—Immunofluorescence staining of cells
was done essentially as described (47). For Pma1p staining, cellswere grown to a density of 1 A600/ml, rapidly fixed by directaddition of formaldehyde for 30 min, and then harvested andfixed overnight with 4.4% formaldehyde in 0.1 M potassiumphosphate, pH 6.5. Before staining, fixed cells were permeabi-lized with 2% SDS. They were then stained with mousemonoclonal anti-Pma1p (Abcam, 40B7 antibody) followedby incubation with goat anti-mouse IgG conjugated to AlexaFluor 488 (Invitrogen). Stained cells were visualized underfluorescein fluorescence optics on a Zeiss Axioplan 2 fluo-rescence microscope.
RESULTS
vma Mutants Poorly Regulate Both Vacuolar and CytosolicpH—One of the defining phenotypes of vma mutants is theirsensitivity to external pH, but relatively little is known abouthow cytosolic and vacuolar pH are regulated in response tochanges in extracellular pH, even inwild-type cells. Ratiometric
V-ATPases Contribute to Cytosolic pH Control in Yeast
JULY 18, 2008 • VOLUME 283 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20311
fluorescent methods in living yeast cells have allowedmeasure-ments of vacuolar and cytosolic pH in cells grown under differ-ent conditions and in mutants lacking proteins implicated inpH homeostasis. We introduced the fluorescent dye BCECF-AM, which measures vacuolar pH in yeast (11, 30), or yeastpHluorin, a green fluorescent protein analog that measurescytosolic pH (31), into wild-type and vma mutant cells andstudied the dynamic adjustments of cytosolic and vacuolar pHin response to glucose and KCl addition. As described above,both glucose and potassium ion are strongly implicated in cyto-solic and vacuolar pH homeostasis (25, 26, 32, 48).Fig. 1A shows the pH responses of wild-type cells grown to
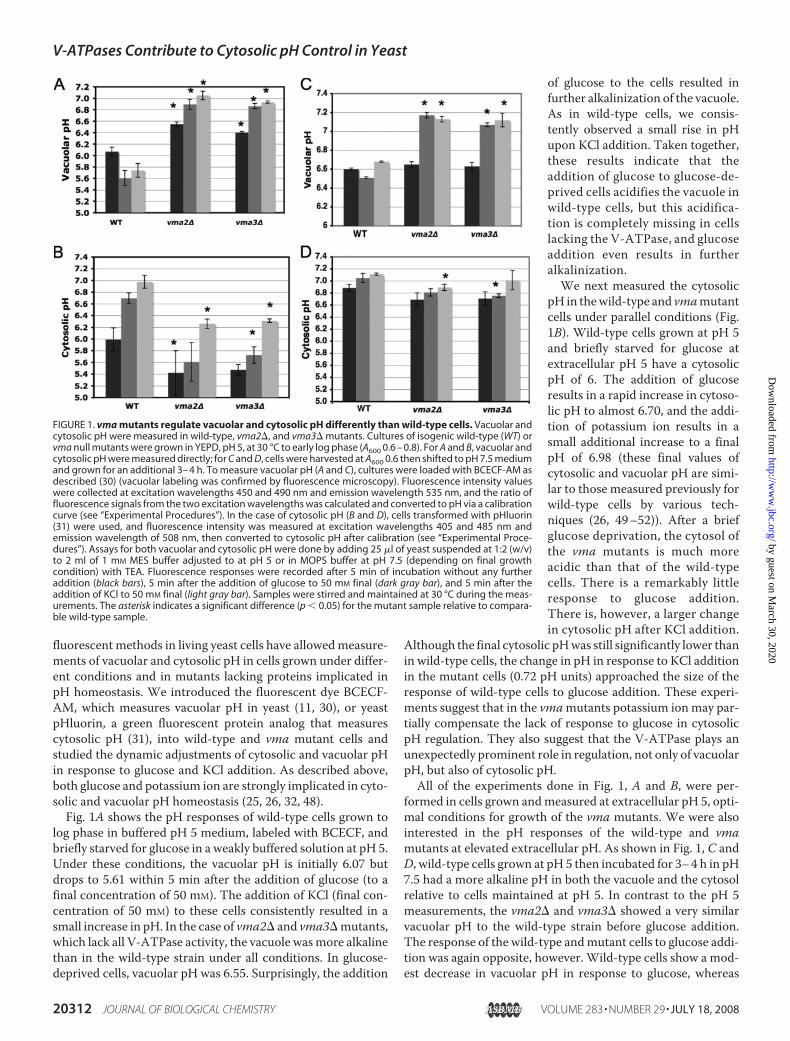
log phase in buffered pH 5 medium, labeled with BCECF, andbriefly starved for glucose in a weakly buffered solution at pH 5.Under these conditions, the vacuolar pH is initially 6.07 butdrops to 5.61 within 5 min after the addition of glucose (to afinal concentration of 50 mM). The addition of KCl (final con-centration of 50 mM) to these cells consistently resulted in asmall increase in pH. In the case of vma2� and vma3�mutants,which lack all V-ATPase activity, the vacuole wasmore alkalinethan in the wild-type strain under all conditions. In glucose-deprived cells, vacuolar pH was 6.55. Surprisingly, the addition
of glucose to the cells resulted infurther alkalinization of the vacuole.As in wild-type cells, we consis-tently observed a small rise in pHupon KCl addition. Taken together,these results indicate that theaddition of glucose to glucose-de-prived cells acidifies the vacuole inwild-type cells, but this acidifica-tion is completely missing in cellslacking the V-ATPase, and glucoseaddition even results in furtheralkalinization.We next measured the cytosolic
pH in thewild-type and vmamutantcells under parallel conditions (Fig.1B). Wild-type cells grown at pH 5and briefly starved for glucose atextracellular pH 5 have a cytosolicpH of 6. The addition of glucoseresults in a rapid increase in cytoso-lic pH to almost 6.70, and the addi-tion of potassium ion results in asmall additional increase to a finalpH of 6.98 (these final values ofcytosolic and vacuolar pH are simi-lar to those measured previously forwild-type cells by various tech-niques (26, 49–52)). After a briefglucose deprivation, the cytosol ofthe vma mutants is much moreacidic than that of the wild-typecells. There is a remarkably littleresponse to glucose addition.There is, however, a larger changein cytosolic pH after KCl addition.
Although the final cytosolic pHwas still significantly lower thanin wild-type cells, the change in pH in response to KCl additionin the mutant cells (0.72 pH units) approached the size of theresponse of wild-type cells to glucose addition. These experi-ments suggest that in the vmamutants potassium ionmay par-tially compensate the lack of response to glucose in cytosolicpH regulation. They also suggest that the V-ATPase plays anunexpectedly prominent role in regulation, not only of vacuolarpH, but also of cytosolic pH.All of the experiments done in Fig. 1, A and B, were per-
formed in cells grown andmeasured at extracellular pH 5, opti-mal conditions for growth of the vma mutants. We were alsointerested in the pH responses of the wild-type and vmamutants at elevated extracellular pH. As shown in Fig. 1, C andD, wild-type cells grown at pH 5 then incubated for 3–4 h in pH7.5 had a more alkaline pH in both the vacuole and the cytosolrelative to cells maintained at pH 5. In contrast to the pH 5measurements, the vma2� and vma3� showed a very similarvacuolar pH to the wild-type strain before glucose addition.The response of the wild-type andmutant cells to glucose addi-tion was again opposite, however. Wild-type cells show a mod-est decrease in vacuolar pH in response to glucose, whereas
FIGURE 1. vma mutants regulate vacuolar and cytosolic pH differently than wild-type cells. Vacuolar andcytosolic pH were measured in wild-type, vma2�, and vma3� mutants. Cultures of isogenic wild-type (WT) orvma null mutants were grown in YEPD, pH 5, at 30 °C to early log phase (A600 0.6 – 0.8). For A and B, vacuolar andcytosolic pH were measured directly; for C and D, cells were harvested at A600 0.6 then shifted to pH 7.5 mediumand grown for an additional 3– 4 h. To measure vacuolar pH (A and C), cultures were loaded with BCECF-AM asdescribed (30) (vacuolar labeling was confirmed by fluorescence microscopy). Fluorescence intensity valueswere collected at excitation wavelengths 450 and 490 nm and emission wavelength 535 nm, and the ratio offluorescence signals from the two excitation wavelengths was calculated and converted to pH via a calibrationcurve (see “Experimental Procedures”). In the case of cytosolic pH (B and D), cells transformed with pHluorin(31) were used, and fluorescence intensity was measured at excitation wavelengths 405 and 485 nm andemission wavelength of 508 nm, then converted to cytosolic pH after calibration (see “Experimental Proce-dures”). Assays for both vacuolar and cytosolic pH were done by adding 25 �l of yeast suspended at 1:2 (w/v)to 2 ml of 1 mM MES buffer adjusted to at pH 5 or in MOPS buffer at pH 7.5 (depending on final growthcondition) with TEA. Fluorescence responses were recorded after 5 min of incubation without any furtheraddition (black bars), 5 min after the addition of glucose to 50 mM final (dark gray bar), and 5 min after theaddition of KCl to 50 mM final (light gray bar). Samples were stirred and maintained at 30 °C during the meas-urements. The asterisk indicates a significant difference (p 0.05) for the mutant sample relative to compara-ble wild-type sample.
V-ATPases Contribute to Cytosolic pH Control in Yeast
20312 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 29 • JULY 18, 2008
vma mutants show a large increase in vacuolar pH. The addi-tion of KCl increased the vacuolar pH of the wild-type cellsfrom 6.5 to 6.7 but had relatively little effect on the vmamutants. In wild-type cells cytosolic pH was somewhat higherat extracellular pH 7.5 than at extracellular pH 5 both beforeand after glucose addition, again consistent with previousmeasurements (26). Cytosolic pH in the vma mutants washigher at extracellular pH 7.5 than at extracellular pH 5 andshowed less difference fromwild type. The pattern of responsesof bothwild-type and the vmamutant strain to glucose andKClwas different at elevated extracellular pH.All three strains showless response to both glucose and KCl addition at extracellularpH 7.5.Proton Export across the PlasmaMembrane Is Slowed in vma
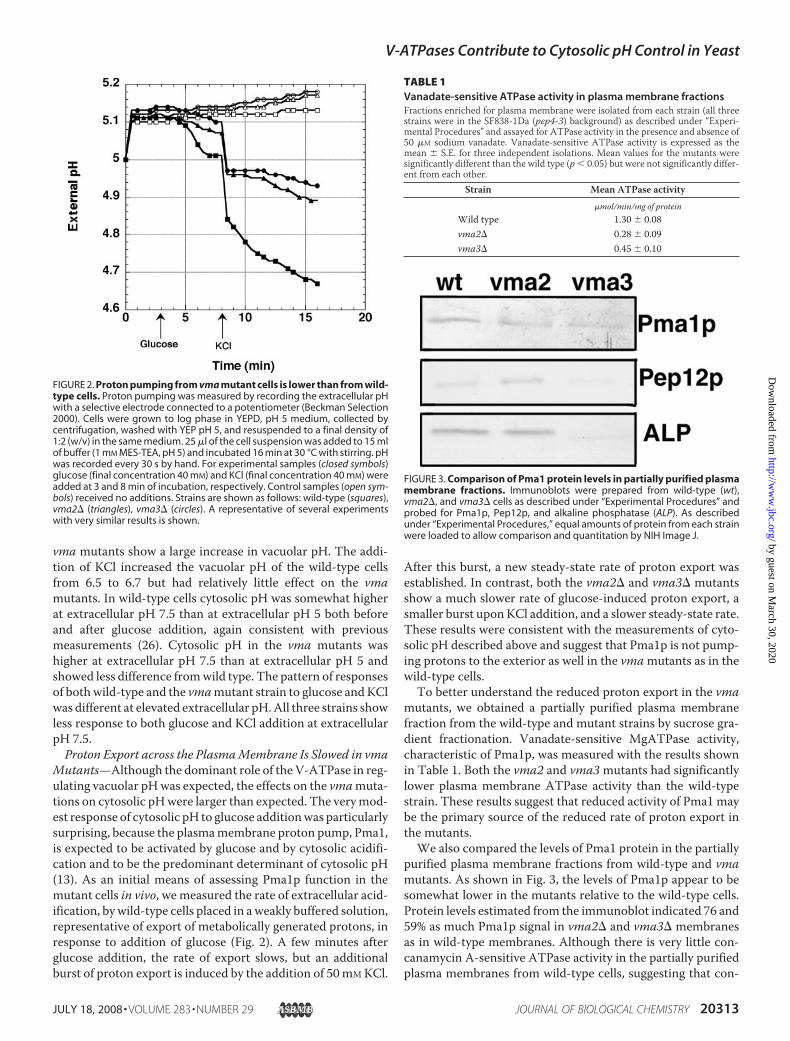
Mutants—Although the dominant role of the V-ATPase in reg-ulating vacuolar pHwas expected, the effects on the vmamuta-tions on cytosolic pHwere larger than expected. The verymod-est response of cytosolic pH to glucose additionwas particularlysurprising, because the plasmamembrane proton pump, Pma1,is expected to be activated by glucose and by cytosolic acidifi-cation and to be the predominant determinant of cytosolic pH(13). As an initial means of assessing Pma1p function in themutant cells in vivo, we measured the rate of extracellular acid-ification, bywild-type cells placed in aweakly buffered solution,representative of export of metabolically generated protons, inresponse to addition of glucose (Fig. 2). A few minutes afterglucose addition, the rate of export slows, but an additionalburst of proton export is induced by the addition of 50mMKCl.
After this burst, a new steady-state rate of proton export wasestablished. In contrast, both the vma2� and vma3� mutantsshow a much slower rate of glucose-induced proton export, asmaller burst uponKCl addition, and a slower steady-state rate.These results were consistent with the measurements of cyto-solic pH described above and suggest that Pma1p is not pump-ing protons to the exterior as well in the vmamutants as in thewild-type cells.To better understand the reduced proton export in the vma
mutants, we obtained a partially purified plasma membranefraction from the wild-type and mutant strains by sucrose gra-dient fractionation. Vanadate-sensitive MgATPase activity,characteristic of Pma1p, was measured with the results shownin Table 1. Both the vma2 and vma3mutants had significantlylower plasma membrane ATPase activity than the wild-typestrain. These results suggest that reduced activity of Pma1 maybe the primary source of the reduced rate of proton export inthe mutants.We also compared the levels of Pma1 protein in the partially
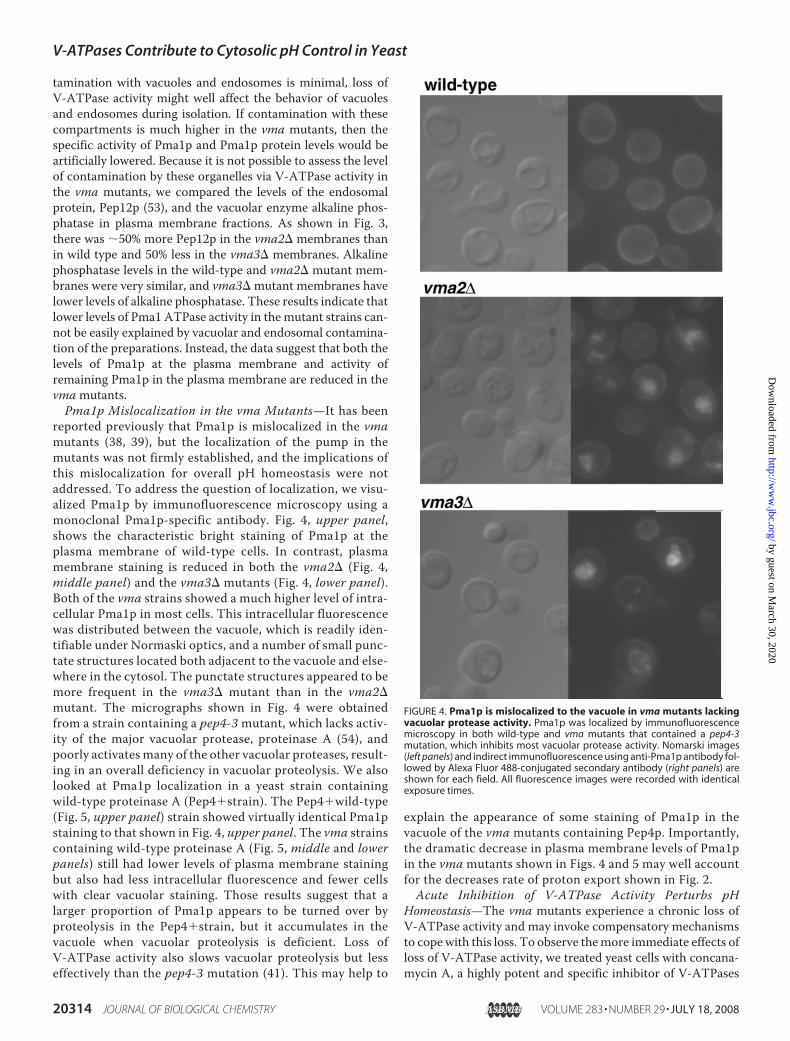
purified plasma membrane fractions from wild-type and vmamutants. As shown in Fig. 3, the levels of Pma1p appear to besomewhat lower in the mutants relative to the wild-type cells.Protein levels estimated from the immunoblot indicated 76 and59% as much Pma1p signal in vma2� and vma3� membranesas in wild-type membranes. Although there is very little con-canamycin A-sensitive ATPase activity in the partially purifiedplasma membranes from wild-type cells, suggesting that con-
FIGURE 2. Proton pumping from vma mutant cells is lower than from wild-type cells. Proton pumping was measured by recording the extracellular pHwith a selective electrode connected to a potentiometer (Beckman Selection2000). Cells were grown to log phase in YEPD, pH 5 medium, collected bycentrifugation, washed with YEP pH 5, and resuspended to a final density of1:2 (w/v) in the same medium. 25 �l of the cell suspension was added to 15 mlof buffer (1 mM MES-TEA, pH 5) and incubated 16 min at 30 °C with stirring. pHwas recorded every 30 s by hand. For experimental samples (closed symbols)glucose (final concentration 40 mM) and KCl (final concentration 40 mM) wereadded at 3 and 8 min of incubation, respectively. Control samples (open sym-bols) received no additions. Strains are shown as follows: wild-type (squares),vma2� (triangles), vma3� (circles). A representative of several experimentswith very similar results is shown.
FIGURE 3. Comparison of Pma1 protein levels in partially purified plasmamembrane fractions. Immunoblots were prepared from wild-type (wt),vma2�, and vma3� cells as described under “Experimental Procedures” andprobed for Pma1p, Pep12p, and alkaline phosphatase (ALP). As describedunder “Experimental Procedures,” equal amounts of protein from each strainwere loaded to allow comparison and quantitation by NIH Image J.
TABLE 1Vanadate-sensitive ATPase activity in plasma membrane fractionsFractions enriched for plasma membrane were isolated from each strain (all threestrains were in the SF838-1Da (pep4-3) background) as described under “Experi-mental Procedures” and assayed for ATPase activity in the presence and absence of50 �M sodium vanadate. Vanadate-sensitive ATPase activity is expressed as themean � S.E. for three independent isolations. Mean values for the mutants weresignificantly different than the wild type (p 0.05) but were not significantly differ-ent from each other.
tamination with vacuoles and endosomes is minimal, loss ofV-ATPase activity might well affect the behavior of vacuolesand endosomes during isolation. If contamination with thesecompartments is much higher in the vma mutants, then thespecific activity of Pma1p and Pma1p protein levels would beartificially lowered. Because it is not possible to assess the levelof contamination by these organelles via V-ATPase activity inthe vma mutants, we compared the levels of the endosomalprotein, Pep12p (53), and the vacuolar enzyme alkaline phos-phatase in plasma membrane fractions. As shown in Fig. 3,there was 50% more Pep12p in the vma2� membranes thanin wild type and 50% less in the vma3� membranes. Alkalinephosphatase levels in the wild-type and vma2� mutant mem-branes were very similar, and vma3� mutant membranes havelower levels of alkaline phosphatase. These results indicate thatlower levels of Pma1 ATPase activity in themutant strains can-not be easily explained by vacuolar and endosomal contamina-tion of the preparations. Instead, the data suggest that both thelevels of Pma1p at the plasma membrane and activity ofremaining Pma1p in the plasma membrane are reduced in thevmamutants.Pma1p Mislocalization in the vma Mutants—It has been
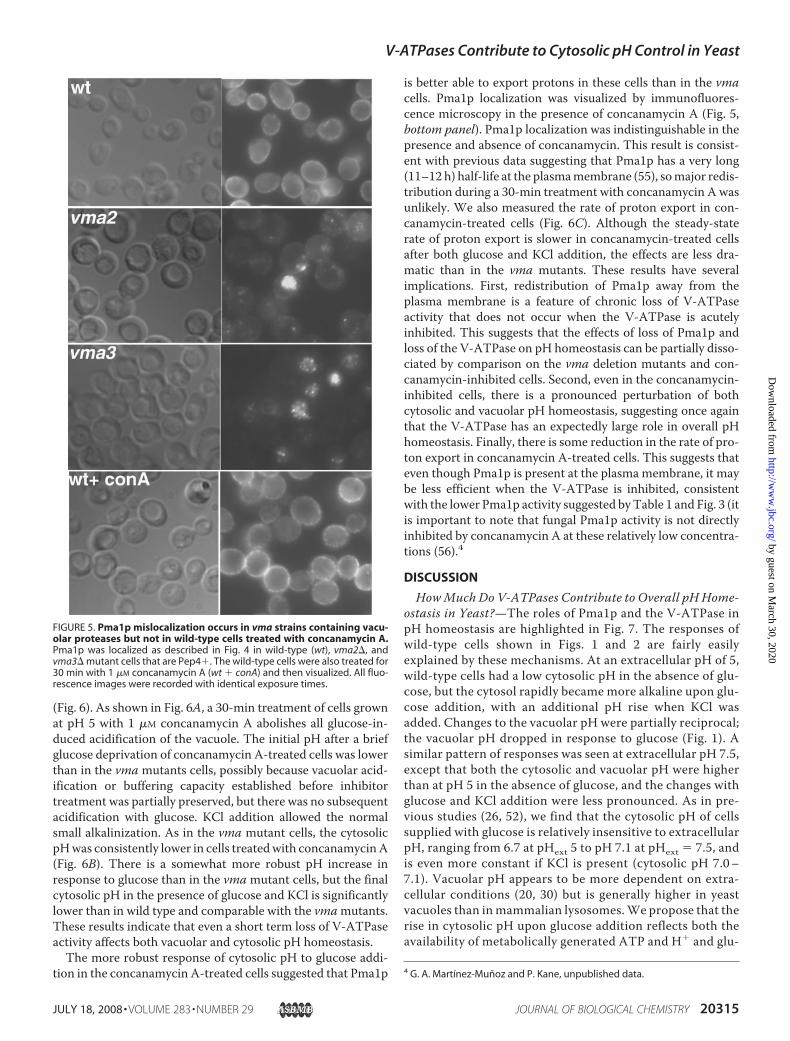
reported previously that Pma1p is mislocalized in the vmamutants (38, 39), but the localization of the pump in themutants was not firmly established, and the implications ofthis mislocalization for overall pH homeostasis were notaddressed. To address the question of localization, we visu-alized Pma1p by immunofluorescence microscopy using amonoclonal Pma1p-specific antibody. Fig. 4, upper panel,shows the characteristic bright staining of Pma1p at theplasma membrane of wild-type cells. In contrast, plasmamembrane staining is reduced in both the vma2� (Fig. 4,middle panel) and the vma3� mutants (Fig. 4, lower panel).Both of the vma strains showed a much higher level of intra-cellular Pma1p in most cells. This intracellular fluorescencewas distributed between the vacuole, which is readily iden-tifiable under Normaski optics, and a number of small punc-tate structures located both adjacent to the vacuole and else-where in the cytosol. The punctate structures appeared to bemore frequent in the vma3� mutant than in the vma2�mutant. The micrographs shown in Fig. 4 were obtainedfrom a strain containing a pep4-3mutant, which lacks activ-ity of the major vacuolar protease, proteinase A (54), andpoorly activates many of the other vacuolar proteases, result-ing in an overall deficiency in vacuolar proteolysis. We alsolooked at Pma1p localization in a yeast strain containingwild-type proteinase A (Pep4�strain). The Pep4�wild-type(Fig. 5, upper panel) strain showed virtually identical Pma1pstaining to that shown in Fig. 4, upper panel. The vma strainscontaining wild-type proteinase A (Fig. 5, middle and lowerpanels) still had lower levels of plasma membrane stainingbut also had less intracellular fluorescence and fewer cellswith clear vacuolar staining. Those results suggest that alarger proportion of Pma1p appears to be turned over byproteolysis in the Pep4�strain, but it accumulates in thevacuole when vacuolar proteolysis is deficient. Loss ofV-ATPase activity also slows vacuolar proteolysis but lesseffectively than the pep4-3 mutation (41). This may help to
explain the appearance of some staining of Pma1p in thevacuole of the vma mutants containing Pep4p. Importantly,the dramatic decrease in plasma membrane levels of Pma1pin the vma mutants shown in Figs. 4 and 5 may well accountfor the decreases rate of proton export shown in Fig. 2.Acute Inhibition of V-ATPase Activity Perturbs pH
Homeostasis—The vma mutants experience a chronic loss ofV-ATPase activity and may invoke compensatory mechanismsto copewith this loss. To observe themore immediate effects ofloss of V-ATPase activity, we treated yeast cells with concana-mycin A, a highly potent and specific inhibitor of V-ATPases
FIGURE 4. Pma1p is mislocalized to the vacuole in vma mutants lackingvacuolar protease activity. Pma1p was localized by immunofluorescencemicroscopy in both wild-type and vma mutants that contained a pep4-3mutation, which inhibits most vacuolar protease activity. Nomarski images(left panels) and indirect immunofluorescence using anti-Pma1p antibody fol-lowed by Alexa Fluor 488-conjugated secondary antibody (right panels) areshown for each field. All fluorescence images were recorded with identicalexposure times.
V-ATPases Contribute to Cytosolic pH Control in Yeast
20314 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 29 • JULY 18, 2008
(Fig. 6). As shown in Fig. 6A, a 30-min treatment of cells grownat pH 5 with 1 �M concanamycin A abolishes all glucose-in-duced acidification of the vacuole. The initial pH after a briefglucose deprivation of concanamycin A-treated cells was lowerthan in the vma mutants cells, possibly because vacuolar acid-ification or buffering capacity established before inhibitortreatment was partially preserved, but there was no subsequentacidification with glucose. KCl addition allowed the normalsmall alkalinization. As in the vma mutant cells, the cytosolicpHwas consistently lower in cells treatedwith concanamycinA(Fig. 6B). There is a somewhat more robust pH increase inresponse to glucose than in the vmamutant cells, but the finalcytosolic pH in the presence of glucose and KCl is significantlylower than in wild type and comparable with the vmamutants.These results indicate that even a short term loss of V-ATPaseactivity affects both vacuolar and cytosolic pH homeostasis.The more robust response of cytosolic pH to glucose addi-
tion in the concanamycin A-treated cells suggested that Pma1p
is better able to export protons in these cells than in the vmacells. Pma1p localization was visualized by immunofluores-cence microscopy in the presence of concanamycin A (Fig. 5,bottom panel). Pma1p localization was indistinguishable in thepresence and absence of concanamycin. This result is consist-ent with previous data suggesting that Pma1p has a very long(11–12 h) half-life at the plasmamembrane (55), somajor redis-tribution during a 30-min treatment with concanamycin A wasunlikely. We also measured the rate of proton export in con-canamycin-treated cells (Fig. 6C). Although the steady-staterate of proton export is slower in concanamycin-treated cellsafter both glucose and KCl addition, the effects are less dra-matic than in the vma mutants. These results have severalimplications. First, redistribution of Pma1p away from theplasma membrane is a feature of chronic loss of V-ATPaseactivity that does not occur when the V-ATPase is acutelyinhibited. This suggests that the effects of loss of Pma1p andloss of the V-ATPase on pH homeostasis can be partially disso-ciated by comparison on the vma deletion mutants and con-canamycin-inhibited cells. Second, even in the concanamycin-inhibited cells, there is a pronounced perturbation of bothcytosolic and vacuolar pH homeostasis, suggesting once againthat the V-ATPase has an expectedly large role in overall pHhomeostasis. Finally, there is some reduction in the rate of pro-ton export in concanamycin A-treated cells. This suggests thateven though Pma1p is present at the plasma membrane, it maybe less efficient when the V-ATPase is inhibited, consistentwith the lower Pma1p activity suggested byTable 1 and Fig. 3 (itis important to note that fungal Pma1p activity is not directlyinhibited by concanamycin A at these relatively low concentra-tions (56).4
DISCUSSION
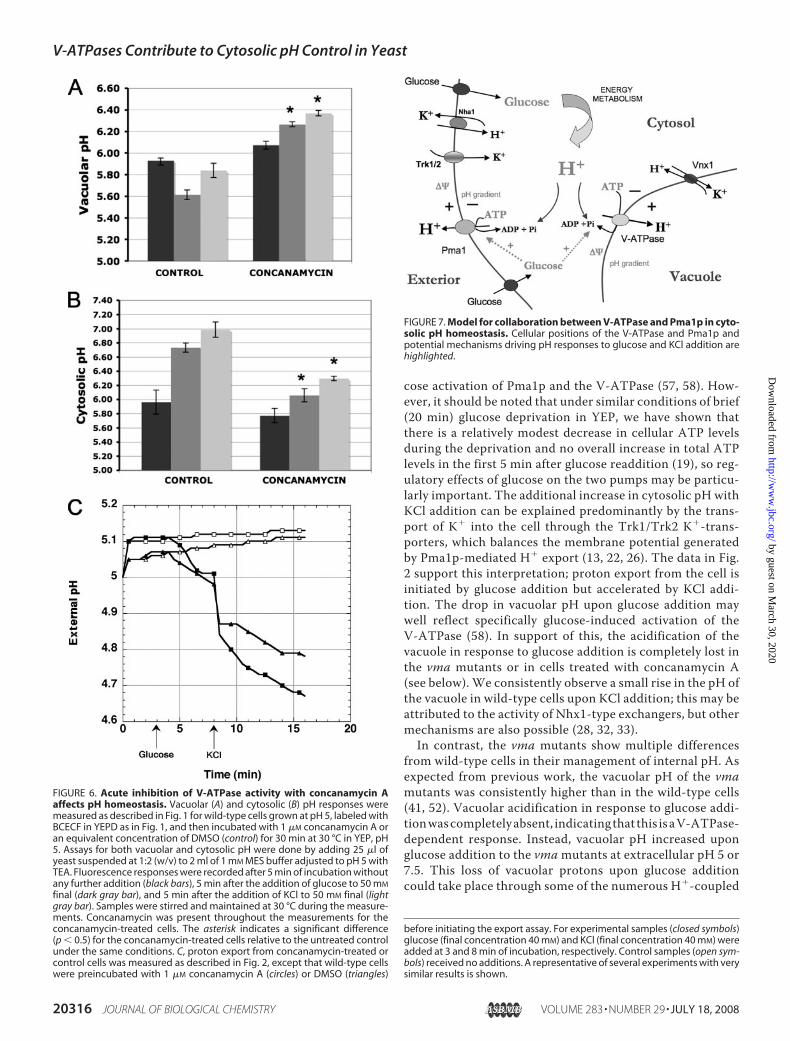
HowMuch Do V-ATPases Contribute to Overall pH Home-ostasis in Yeast?—The roles of Pma1p and the V-ATPase inpH homeostasis are highlighted in Fig. 7. The responses ofwild-type cells shown in Figs. 1 and 2 are fairly easilyexplained by these mechanisms. At an extracellular pH of 5,wild-type cells had a low cytosolic pH in the absence of glu-cose, but the cytosol rapidly became more alkaline upon glu-cose addition, with an additional pH rise when KCl wasadded. Changes to the vacuolar pH were partially reciprocal;the vacuolar pH dropped in response to glucose (Fig. 1). Asimilar pattern of responses was seen at extracellular pH 7.5,except that both the cytosolic and vacuolar pH were higherthan at pH 5 in the absence of glucose, and the changes withglucose and KCl addition were less pronounced. As in pre-vious studies (26, 52), we find that the cytosolic pH of cellssupplied with glucose is relatively insensitive to extracellularpH, ranging from 6.7 at pHext 5 to pH 7.1 at pHext � 7.5, andis even more constant if KCl is present (cytosolic pH 7.0–7.1). Vacuolar pH appears to be more dependent on extra-cellular conditions (20, 30) but is generally higher in yeastvacuoles than inmammalian lysosomes.We propose that therise in cytosolic pH upon glucose addition reflects both theavailability of metabolically generated ATP and H� and glu-
4 G. A. Martınez-Munoz and P. Kane, unpublished data.
FIGURE 5. Pma1p mislocalization occurs in vma strains containing vacu-olar proteases but not in wild-type cells treated with concanamycin A.Pma1p was localized as described in Fig. 4 in wild-type (wt), vma2�, andvma3� mutant cells that are Pep4�. The wild-type cells were also treated for30 min with 1 �M concanamycin A (wt � conA) and then visualized. All fluo-rescence images were recorded with identical exposure times.
V-ATPases Contribute to Cytosolic pH Control in Yeast
JULY 18, 2008 • VOLUME 283 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20315
cose activation of Pma1p and the V-ATPase (57, 58). How-ever, it should be noted that under similar conditions of brief(20 min) glucose deprivation in YEP, we have shown thatthere is a relatively modest decrease in cellular ATP levelsduring the deprivation and no overall increase in total ATPlevels in the first 5 min after glucose readdition (19), so reg-ulatory effects of glucose on the two pumps may be particu-larly important. The additional increase in cytosolic pH withKCl addition can be explained predominantly by the trans-port of K� into the cell through the Trk1/Trk2 K�-trans-porters, which balances the membrane potential generatedby Pma1p-mediated H� export (13, 22, 26). The data in Fig.2 support this interpretation; proton export from the cell isinitiated by glucose addition but accelerated by KCl addi-tion. The drop in vacuolar pH upon glucose addition maywell reflect specifically glucose-induced activation of theV-ATPase (58). In support of this, the acidification of thevacuole in response to glucose addition is completely lost inthe vma mutants or in cells treated with concanamycin A(see below). We consistently observe a small rise in the pH ofthe vacuole in wild-type cells upon KCl addition; this may beattributed to the activity of Nhx1-type exchangers, but othermechanisms are also possible (28, 32, 33).In contrast, the vma mutants show multiple differences
from wild-type cells in their management of internal pH. Asexpected from previous work, the vacuolar pH of the vmamutants was consistently higher than in the wild-type cells(41, 52). Vacuolar acidification in response to glucose addi-tionwascompletelyabsent, indicatingthat this isaV-ATPase-dependent response. Instead, vacuolar pH increased uponglucose addition to the vmamutants at extracellular pH 5 or7.5. This loss of vacuolar protons upon glucose additioncould take place through some of the numerous H�-coupled
FIGURE 6. Acute inhibition of V-ATPase activity with concanamycin Aaffects pH homeostasis. Vacuolar (A) and cytosolic (B) pH responses weremeasured as described in Fig. 1 for wild-type cells grown at pH 5, labeled withBCECF in YEPD as in Fig. 1, and then incubated with 1 �M concanamycin A oran equivalent concentration of DMSO (control) for 30 min at 30 °C in YEP, pH5. Assays for both vacuolar and cytosolic pH were done by adding 25 �l ofyeast suspended at 1:2 (w/v) to 2 ml of 1 mM MES buffer adjusted to pH 5 withTEA. Fluorescence responses were recorded after 5 min of incubation withoutany further addition (black bars), 5 min after the addition of glucose to 50 mM
final (dark gray bar), and 5 min after the addition of KCl to 50 mM final (lightgray bar). Samples were stirred and maintained at 30 °C during the measure-ments. Concanamycin was present throughout the measurements for theconcanamycin-treated cells. The asterisk indicates a significant difference(p 0.5) for the concanamycin-treated cells relative to the untreated controlunder the same conditions. C, proton export from concanamycin-treated orcontrol cells was measured as described in Fig. 2, except that wild-type cellswere preincubated with 1 �M concanamycin A (circles) or DMSO (triangles)
before initiating the export assay. For experimental samples (closed symbols)glucose (final concentration 40 mM) and KCl (final concentration 40 mM) wereadded at 3 and 8 min of incubation, respectively. Control samples (open sym-bols) received no additions. A representative of several experiments with verysimilar results is shown.
FIGURE 7. Model for collaboration between V-ATPase and Pma1p in cyto-solic pH homeostasis. Cellular positions of the V-ATPase and Pma1p andpotential mechanisms driving pH responses to glucose and KCl addition arehighlighted.
V-ATPases Contribute to Cytosolic pH Control in Yeast
20316 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 29 • JULY 18, 2008
antiporters present in the vacuolar membrane (3), but a lossof buffering power in the vacuole may also contribute. Vac-uolar polyphosphates play an important buffering role (35),and vma mutants have very low levels of vacuolar polyphos-phate (34, 52). One significant implication of the vacuolarpH measurements is that the vma mutants do not appear toacidify of the vacuole through endocytic uptake of extracel-lular fluid (6, 10). As shown in Fig. 1A, at external pH 5 in thepresence of glucose and KCl, the vacuolar pH in the vmamutants is 7.0. This result implies that the synthetic lethal-ity between the vma and the end mutants (10) must haveanother explanation and is entirely consistent with the con-clusions of Plant et al. (11), who found that in the absence ofpermeant acids the vmamutants had a near-neutral vacuolarpH.The prominent effect of the vma mutations on cytosolic
pH, particularly at pHext 5, was initially surprising. Underconditions of cytosolic acidification, Pma1p activity is stim-ulated, and this stimulation is regarded as central to overallpH homeostasis (21, 59). Remarkably, even in the presence ofglucose and KCl, the cytosolic pH of the vma mutants was0.6 pH units lower than that of the wild-type cells. Thefailure of the vma mutants to respond to glucose additionwith an alkalinization of the cytosol was particularly note-worthy. Coupled with the evidence that there is little activa-tion of proton export upon glucose addition (Fig. 2), theseresults strongly suggested that Pma1p, rather than compen-sating for cytosolic acidification arising from loss ofV-ATPase activity with increased activity, was actually lessactive in the mutants. This was confirmed biochemically(Table 1). In contrast to the glucose response, the cytosolicpH of the vmamutants was more responsive to KCl addition,and KCl addition also strongly stimulated proton export.This may reflect in part stimulation of Trk1 by acidic cyto-solic pH (59) and the tight interdependence of pH and K�
homeostasis. Further experiments will be necessary to fullydetermine why pH homeostatic mechanisms fail to achievecontrol of cytosolic pH in the vma mutant cells.After a shift to alkaline extracellular pH (Fig. 1B), vma
mutants were able to achieve a cytosolic pH very similar to thatof wild-type cells, along with amuchmore alkaline vacuolar pH(Fig. 1, C andD). The vmamutants do not grow at pHext � 7.5,but the source of their growth defect may not be directlyrelated to pH homeostasis. It has been suggested that copperand iron limit growth of yeast cells at alkaline pH (60), andDavis-Kaplan et al. (61) have demonstrated that the vmamutants are constitutively iron-deprived. Transcriptionalprofiling of the vma mutants also suggests that they experi-ence iron deprivation, even at pH 5 (62). Other factors suchas iron deprivation may well be the direct cause of the pH-dependent conditional lethality.Mislocalization of Pma1p in the vma Mutant Cells—Our
data indicate that Pma1 protein is present at lower levels atthe plasmamembrane of the vmamutants and in the absenceof vacuolar protease activity accumulates in the vacuole.When normal levels of vacuolar protease are present, Pma1pappears to be rapidly turned over at the vacuole, althoughother Pma1p-containing intracellular membranes are visible
in a population of the vma mutant cells (Figs. 3 and 4). Weknow neither how nor why Pma1p is mistargeted to the vac-uole in the vma mutants. Pma1p is normally a long-livedprotein, but a number of factors, including mutations inPma1p itself, altered recruitment to rafts, or altered ubiquiti-nation, can intervene to change its longevity or divert it awayfrom the plasma membrane to the vacuole (63–65). There isalso evidence of signal-induced alterations in Pma1p stabil-ity; specifically, activation of the calcium-activated proteinphosphatase calcineurin results in lower Pma1p levels (66).The vmamutants rely on constitutive calcineurin activationfor viability (67), so this may contribute to the lowering ofPma1p levels. Further experiments will be necessary todetermine whether Pma1p is routed directly to the vacuolein the mutants or is retrieved from the plasma membrane byendocytosis. Perzov et al. (38) demonstrated that anotherplasma membrane protein, Gas1, is localized normally in thevma mutants, but Davis-Kaplan et al. (61) reported thatFet3/Ftr1, a plasma membrane transporter complex respon-sible for high affinity iron transport, is present at low levels inthe plasma membrane of vma mutants, like Pma1p. There-fore, it is still unclear whether the mutants have a generaldefect in targeting and/or stability of a subclass of plasmamembrane proteins or whether there is a specific mislocal-ization of Pma1p.The question of why Pma1p is mistargeted is inextricably
linked to the question of how it reaches the vacuole.Although the idea of populating the vacuole with an alterna-tive proton pump in the vmamutants as a means of compen-sating for loss of V-ATPase activity is attractive, vacuolesfrom the vma mutant strain have almost no vanadate-sensi-tive ATPase activity and no ATP-driven proton pumping(41, 68), and overexpression of Pma1p does not suppress thevma mutant phenotypes (11). It is still possible that Pma1pactivity in earlier compartments, such as endosomes and/orGolgi, contributes to compartment acidification, and a needfor populating endosomes with Pma1p could account for thesynthetic lethality of the end and vma mutants (10). How-ever, the cost of mislocalizing Pma1p, in terms of pH homeo-stasis and reduced activity of other plasma membrane trans-porters that rely on the pH gradient and membrane potential,would appear to be quite high. In fact, some of thewide range ofphenotypes of the vmamutants (69)might arise indirectly fromreduced Pma1p activity rather than directly from loss ofV-ATPase activity. Nevertheless, In N. crassa, mutations thatreduced Pma1p activity suppress concanamycin sensitivity(70), suggesting that reduced Pma1p activity may help cells tosurvive in the absence of a functional V-ATPase by a mecha-nism that is not yet clear.Vacuolar and Plasma Membrane Proton Pumps Collabo-
rate in pH Homeostasis—Reduced Pma1p function does notfully account for the perturbed pH homeostasis in the vmamutants, however. Although concanamycin-treated wild-type cells show normal plasmamembrane staining of Pma1p,they still show pronounced perturbations in pH homeosta-sis. Not only is glucose-induced vacuolar acidification elim-inated after 30 min of concanamycin A treatment, but cyto-solic pH is significantly lower under all conditions. We
V-ATPases Contribute to Cytosolic pH Control in Yeast
JULY 18, 2008 • VOLUME 283 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20317
propose that V-ATPases influence cytosolic pH homeostasison at least two levels. First, they have a direct effect that islikely to involve sequestration of protons into the vacuoleand other organelles and is lost in the concanamycin-treatedcells. It is interesting, however, that the rate of proton exportfrom the cell in response to glucose is actually lower in theconcanamycin-treated cells. Thus loss of V-ATPase activitydecreases Pma1p activity even when it is properly localizedand could indicate a “cross-talk” or coordinate regulationbetween the V-ATPase and Pma1p. Both of these pumps arehighly regulated at a number of levels, and their regulation isnot completely understood (2, 13, 59). Second, V-ATPasesappear to be critical for efficient localization of Pma1p to theplasma membrane. Taken together, the data indicate thatPma1p and V-ATPases are not independent players workingtoward a common goal of pH homeostasis but, rather, arehighly interdependent pumps that are coordinately regu-lated on multiple levels.
Acknowledgments—We thank Dr. Rajini Rao, Johns Hopkins Univer-sity for the pHluorin plasmid and for helpful discussions. We alsothankDr. Ramon Serrano for providing polyclonal antibodies againstPma1p andDr. Antonio Pena for providing cyanine dye that was usedin background experiments important to this work.
REFERENCES1. Forgac, M. (2007) Nat. Rev. Mol. Cell Biol. 8, 917–9292. Kane, P. M. (2006) Microbiol. Mol. Biol. Rev. 70, 177–1913. Klionsky, D. J., Herman, P. K., and Emr, S. D. (1990) Microbiol. Rev. 54,
266 –2924. Swallow, C. J., Grinstein, S., Sudsbury, R. A., and Rotstein, O. D. (1993)
J. Cell. Physiol. 157, 453– 4605. Swallow, C. J., Grinstein, S., and Rotstein, O. D. (1990) J. Biol. Chem. 265,
7645–76546. Nelson, H., and Nelson, N. (1990) Proc. Natl. Acad. Sci. U. S. A. 87,
3503–35077. Bowman, E. J., Kendle, R., and Bowman, B. J. (2000) J. Biol. Chem. 275,
167–1768. Davies, S. A., Goodwin, S. F., Kelly, D. C., Wang, Z., Sozen, M. A., Kaiser,
K., and Dow, J. A. (1996) J. Biol. Chem. 271, 30677–306849. Sun-Wada, G., Murata, Y., Yamamoto, A., Kanazawa, H., Wada, Y., and
Futai, M. (2000) Dev. Biol. 228, 315–32510. Munn, A. L., and Riezman, H. (1994) J. Cell Biol. 127, 373–38611. Plant, P. J., Manolson, M. F., Grinstein, S., and Demaurex, N. (1999) J. Biol.
Chem. 274, 37270 –3727912. Serrano, R., Kielland-Brandt, M. C., and Fink, G. R. (1986) Nature 319,
689 – 69313. Serrano, R. (1991) in The Molecular and Cellular Biology of the Yeast
Saccharomyces: Genome Dynamics, Protein Synthese, and Energetics(Broach, J. R., Pringle, J. R., and Jones, E. W., eds) Vol. 1, pp. 523–585, ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY
14. Conway, E. J., and Brady, T. G. (1950) Biochem. J. 47, 360 –36915. Lecchi, S., Nelson, C. J., Allen, K. E., Swaney, D. L., Thompson, K. L., Coon,
J. J., Sussman, M. R., and Slayman, C. W. (2007) J. Biol. Chem. 282,35471–35481
16. Eraso, P., Mazon, M. J., and Portillo, F. (2006) Biochim. Biophys. Acta1758, 164 –170
17. Portillo, F., Eraso, P., and Serrano, R. (1991) FEBS Lett. 287, 71–7418. Kane, P. M. (1995) J. Biol. Chem. 270, 17025–1703219. Parra, K. J., and Kane, P. M. (1998) Mol. Cell. Biol. 18, 7064 –707420. Padilla-Lopez, S., and Pearce, D. A. (2006) J. Biol. Chem. 281,
10273–1028021. Eraso, P., and Gancedo, C. (1987) FEBS Lett. 224, 187–192
22. Seto-Young, D., and Perlin, D. S. (1991) J. Biol. Chem. 266, 1383–138923. Grabe, M., and Oster, G. (2001) J. Gen. Physiol. 117, 329 –34424. Madrid, R., Gomez, M. J., Ramos, J., and Rodriguez-Navarro, A. (1998)
J. Biol. Chem. 273, 14838 –1484425. Pena, A., Cinco, G., Gomez-Puyou, A., and Tuena, M. (1972) Arch. Bio-
chem. Biophys. 153, 413– 42526. Calahorra, M., Martinez, G. A., Hernandez-Cruz, A., and Pena, A. (1998)
Yeast 14, 501–51527. Banuelos, M. A., Sychrova, H., Bleykasten-Grosshans, C., Souciet, J. L., and
Potier, S. (1998) Microbiology 144, 2749 –275828. Nass, R., and Rao, R. (1998) J. Biol. Chem. 273, 21054 –2106029. Orlowski, J., and Grinstein, S. (1997) J. Biol. Chem. 272, 22373–2237630. Ali, R., Brett, C. L., Mukherjee, S., and Rao, R. (2004) J. Biol. Chem. 279,
4498 – 450631. Brett, C. L., Tukaye, D. N., Mukherjee, S., and Rao, R. (2005)Mol. Biol. Cell
16, 1396 –140532. Martinez-Munoz, G. A., and Pena, A. (2005) Yeast 22, 689 –70433. Cagnac, O., Leterrier, M., Yeager, M., and Blumwald, E. (2007) J. Biol.
Chem. 282, 24284 –2429334. Freimoser, F. M., Hurlimann, H. C., Jakob, C. A., Werner, T. P., and Am-
rhein, N. (2006) Genome Biology 7, R10935. Pick, U., Bental, M., Chitlaru, E., and Weiss, M. (1990) FEBS Lett. 274,
15–1836. Brett, C. L., Donowitz, M., and Rao, R. (2006) FEBS Lett. 580, 717–71937. Bowman, E. J., and Bowman, B. J. (2000) J. Exp. Biol. 203, 97–10638. Perzov, N., Nelson, H., and Nelson, N. (2000) J. Biol. Chem. 275,
40088 – 4009539. Hirata, R., and Takatsuki, A. (2001) RIKEN Review 41, 90 –9140. Gordon, J. A. (1991) Methods Enzymol. 201, 477– 48241. Yamashiro, C. T., Kane, P. M., Wolczyk, D. F., Preston, R. A., and Stevens,
T. H. (1990) Mol. Cell. Biol. 10, 3737–374942. Kane, P. M., Kuehn, M. C., Howald-Stevenson, I., and Stevens, T. H. (1992)
J. Biol. Chem. 267, 447– 45443. Pena, A. (1975) Arch Biochem. Biophys. 167, 397– 40944. Panaretou, B., and Piper, P. (2006) Methods Mol. Biol. 313, 27–3245. Lotscher, H. R., deJong, C., and Capaldi, R. A. (1984) Biochemistry 23,
4140 – 414346. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951) J. Biol.
Chem. 193, 265–27547. Roberts, C. J., Raymond, C. K., Yamashiro, C. T., and Stevens, T. H. (1991)
Methods Enzymol. 194, 644 – 66148. Pena, A., Cinco, G., Puyou, A. G., and Tuena, M. (1969) Biochim. Biophys.
Acta 180, 1– 849. Pena, A., Ramirez, J., Rosas, G., and Calahorra, M. (1995) J. Bacteriol. 177,
1017–102250. Preston, R. A., Murphy, R. F., and Jones, E. W. (1989) Proc. Natl. Acad. Sci.
U. S. A. 86, 7027–703151. Haworth, R. S., and Fliegel, L. (1993) Mol. Cell. Biochem. 124, 131–14052. Beauvoit, B., Rigoulet, M., Raffard, G., Canioni, P., and Guerin, B. (1991)
Biochemistry 30, 11212–1122053. Gerrard, S. R., Levi, B. P., and Stevens, T. H. (2000) Traffic 1, 259 –26954. Ammerer, G., Hunter, C. P., Rothman, J. H., Saari, G. C., Valls, L. A., and
Stevens, T. H. (1986) Mol. Cell. Biol. 6, 2490 –249955. Benito, B., Moreno, E., and Lagunas, R. (1991) Biochim. Biophys. Acta
1063, 265–26856. Bowman, E. J., Siebers, A., and Altendorf, K. (1988) Proc. Natl. Acad. Sci.
U. S. A. 85, 7972–797657. Serrano, R. (1983) FEBS Lett. 156, 11–1458. Kane, P. M. (2000) FEBS Lett. 469, 137–14159. Yenush, L., Merchan, S., Holmes, J., and Serrano, R. (2005) Mol. Cell. Biol.
25, 8683– 869260. Serrano, R., Bernal, D., Simon, E., and Arino, J. (2004) J. Biol. Chem. 279,
19698 –1970461. Davis-Kaplan, S. R., Ward, D. M., Shiflett, S. L., and Kaplan, J. (2004) J. Biol.
Chem. 279, 4322– 432962. Milgrom, E., Diab, H., Middleton, F., and Kane, P. M. (2007) J. Biol. Chem.
282, 7125–713663. Bagnat, M., Chang, A., and Simons, K. (2001) Mol. Biol. Cell 12,
V-ATPases Contribute to Cytosolic pH Control in Yeast
20318 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 283 • NUMBER 29 • JULY 18, 2008
4129 – 413864. Liu, Y., and Chang, A. (2006) J. Cell Sci. 119, 360 –36965. Wang, Q., and Chang, A. (2002) Proc. Natl. Acad. Sci. U. S. A. 99,
12853–1285866. Withee, J. L., Sen, R., and Cyert, M. S. (1998) Genetics 149, 865– 87867. Tanida, I., Hasegawa, A., Iida, H., Ohya, Y., and Anraku, Y. (1995) J. Biol.
Chem. 270, 10113–1011968. Umemoto, N., Yoshihisa, T., Hirata, R., and Anraku, Y. (1990) J. Biol.
Chem. 265, 18447–1845369. Kane, P. M. (2007) J. Bioenerg. Biomembr. 39(5– 6):415–2170. Bowman, E. J., O’Neill, F. J., and Bowman, B. J. (1997) J. Biol. Chem. 272,
14776 –14786
V-ATPases Contribute to Cytosolic pH Control in Yeast
JULY 18, 2008 • VOLUME 283 • NUMBER 29 JOURNAL OF BIOLOGICAL CHEMISTRY 20319
Vacuolar and plasma membrane proton pumpscollaborate to achieve cytosolic pH homeostasis in yeast.Gloria A. Martínez-Munoz and Patricia Kane
The E-mail address for Dr. Martínez-Munoz has been updated [email protected]. The footnote should now read asfollows, “To whom correspondence may be addressed. E-mail:[email protected].”