Page 1
Journal of Vegetation Science 25 (2014) 534–545
Variation in tidal wetland plant diversity andcomposition within and among coastal estuaries:assessing the relative importance of environmentalgradients
Christopher N. Janousek & Christina L. Folger
Keywords
Hierarchical partitioning; NMDS; Path analysis;
Salt marsh; Soil salinity; Species accumulation
curves; Tidal elevation; Tidal swamp
Abbreviations
DCA = Detrended correspondence analysis;
MHHW =mean higher high water; MLLW =
mean lower low water; NMDS = Non-metric
multidimensional scaling; NIS = non-indigenous
species; NWI = National Wetlands Inventory;
SLR = sea level rise.
Nomenclature
Cook, T. and S. Sundberg. eds. (2011). Oregon
Vascular Plant Checklist, Oregon Flora Project.
(www.oregonflora.org/checklist.php).
Received 22 June 2012
Accepted 27 June 2013
Co-ordinating Editor: Rune Halvorsen
Janousek, C.N. (corresponding author,
[email protected] ) & Folger, C.L.
([email protected] ): Western Ecology
Division, Office of Research and Development,
US Environmental Protection Agency, 2111 SE
Marine Science Dr., Newport, OR, 97365, USA
Abstract
Questions:What is the relative importance of topographic (elevation), edaphic
(soil salinity, nitrogen and particle size) and hydrologic (estuarine river flow)
gradients for variation in tidal wetland plant composition and diversity?
Location: Four Oregon estuaries: a marine-dominated lagoon, two tidal-driven
bays, and a river-dominated site.
Methods:We surveyed species presence, cover and richness; and environmen-
tal factors (soil salinity, grain size, soil nitrogen and elevation) in plots in marsh
and swamp. We assessed patterns of community structure and the relative
importance of environmental gradients with hierarchical partitioning, ordina-
tion, species accumulation curves and path analysis.
Results: The relative importance of measured environmental gradients on plant
occurrence differed by species. Soil salinity or elevation explained the most vari-
ation in the majority of common species. Estuarine hydrology, soil nitrogen and
soil clay content were usually of secondary or minor importance. Assemblage
composition and species richness varied most strongly with tidal elevation. Local
soil salinity also affected composition, but differences in estuarine hydrology
had comparatively less effect on composition and richness. Higher-elevation
wetlands supported larger species pools and higher plot-level richness; fresher
wetlands had larger species pools than salt marsh but plot-level richness was rel-
atively invariant to differences in soil salinity.
Conclusions: Elevation and salinity tended to exert more influence on the veg-
etation structure of tidal wetlands than estuarine hydrology or other edaphic
variables. With relative sea-level rise expected to increase both flooding inten-
sity and salinity exposure in future wetlands, global climate change may lead to
changes in species distributions, altered floristic composition and reduced plant
species richness.
Introduction
Tidal wetlands are transitional habitats between terrestrial
ecosystems and marine-influenced tide flats and seagrass
meadows. These marshes and swamps provide important
ecosystem functions in coastal watersheds including nutri-
ent transformation and trophic support that lead to valued
ecosystem services (Barbier et al. 2011; Engle 2011). Vas-
cular plant abundance, composition and productivity play
key roles in these processes. Tidal wetland structure and
function are vulnerable to future climate change impacts,
because sea-level rise (SLR) or other changes may alter
intertidal stressors or resource availability (Parker et al.
2011; Stralberg et al. 2011).
Plant composition in coastal marshes and swamps is
determined in part by abiotic factors such as salinity and
tidal inundation (Engels & Jensen 2009; Watson & Byrne
2009). Elevation relative to tide level is an important driver
of plant composition in the Pacific Northwest (Eilers
1975), as it is in other geographic regions (Pennings et al.
2005; Silvestri et al. 2005). Disturbance, herbivory,
competition, facilitation and topographic heterogeneity
Journal of Vegetation Science534 Doi: 10.1111/jvs.12107© 2013 International Association for Vegetation Science
Published 2013. This article is a US Government work and is in the public domain in the USA.
Page 2
(e.g. due to the occurrence of tidal creeks) also influence
spatial patterns of plant composition and richness in these
ecosystems (Zedler et al. 1999; Rand 2000; Pennings et al.
2003; Gedan et al. 2009; Zedler 2010; Keammerer &
Hacker 2013).
Topographic, hydrologic and edaphic gradients may
overlap in complex ways in estuarine wetlands (Odum
1988; Cui et al. 2011; Davy et al. 2011) and have interac-
tive effects on vegetation structure (Silvestri et al. 2005).
For instance, wetland soil salinity can co-vary with tidal
elevation (though not always in similar ways in different
regions), but it also declines along the estuarine axis from
ocean to river (Engels & Jensen 2009). Because of the
overlap of multiple environmental gradients in these eco-
systems, it is important to disentangle these effects to better
understand the distribution of individual species and more
general patterns of composition and diversity. Quantifying
the relative sensitivity of the wetland flora to changes in
different abiotic factors is also critical to understanding the
mechanisms underlying vulnerability of coastal wetlands
to climate change. Which of the multitude of environmen-
tal changes expected with altered climate – greater flood-
ing, elevated air and water temperatures, increasing CO2
or dry season salinity (Watson & Byrne 2009; Parker et al.
2011) – are likely to impact wetland vegetation themost?
Tidal wetlands of the Pacific Northwest are a useful case
study for assessing which of the complex local and regional
environmental gradients are most important for plant
community composition. Estuaries in the region vary
greatly in size, watershed area and the relative degree of
freshwater inputs from coastal watersheds (Lee & Brown
2009). As in other temperate estuaries, there are sharp ele-
vation and salinity gradients. The region is also exceptional
because it has a diverse wetland flora (Jefferson 1975;
Weilhoefer et al. 2013). While detailed vegetation surveys
have been conducted (e.g. Eilers 1975; Liverman 1981),
there is little information on the relative importance of abi-
otic drivers of composition in the region. A recent study
across multiple Oregon estuaries by Weilhoefer et al.
(2013) noted the linkages between local land cover, chan-
nel salinity and plant composition, but did not examine
effects due to elevation or soil salinity.
In this study, we quantified variation in vascular plant
presence, cover, composition and species richness along
five environmental gradients within and among estuaries
in the Pacific Northwest. We assessed the relative strength
of tidal elevation, soil salinity, soil nitrogen, soil clay con-
tent and estuarine hydrology (degree of riverine input) on
community structure. We tested whether gradients that
best explained variation in individual species’ occurrences
(the term ‘explain’ used here in a statistical and not a cau-
sal sense) were also most important for explaining aggre-
gate composition and richness patterns. We then discuss
our findings in the context of temperate wetland vulnera-
bility to different aspects of global climate change, includ-
ing sea-level rise and estuary salinization.
Methods
Sampling design
Emergent marsh and woody tidal wetlands were sampled
in four outer coast estuaries that differ hydrologically:
Alsea, Coquille, Netarts and Yaquina (Table 1). Netarts
Bay in northern Oregon is a shallow bar-built marine
lagoon that receives little freshwater input (Lee & Brown
2009). The Yaquina and Alsea estuaries in central Oregon
and Coquille in southern Oregon are drowned river-
mouth estuaries. Each is strongly tidally influenced, with
increasing degrees of river dominance from Yaquina to
Alsea to Coquille. The majority of extant habitat is
emergent marsh, with small areas of tidal swamp and
shrub–scrub wetland.
Geographic information system layers of National Wet-
land Inventory (US Fish and Wildlife Service) wetland dis-
tribution (2009–2010) in each estuary were used to
conduct stratified random sampling. Strata comprised (1)
low estuarine emergent marsh (NWI code ‘E2EMN’); (2)
high estuarine emergent marsh (‘E2EMP’); and (3) several
tidally-influenced palustrine wetland classes (‘PEMR’,
‘PEMS’, ‘PFOR’, ‘PFOS’, ‘PSSR’, ‘PSSS’; Cowardin et al.
1979). Several additional plots in a mixed emergent
marsh/shrub–scrub/tidal swamp wetland along the Yaqu-
ina River were also randomly sampled to increase repre-
sentation of upstream tidal habitat, since it tended to be
Table 1. Estuary and wetland area, normalized freshwater flow index, tidal range (difference between MLLW and MHHW) and number of plots sampled by
estuary in this study. Estuaries are ordered from most river-dominated to most ocean-dominated. The normalized freshwater flow index represents the
total annual precipitation (m3�yr�1) in a given watershed divided by total estuarine area (m2). Tidal range data for Yaquina are from the lower estuary (South
Beach); the tide range in the upper estuary (Toledo) is 2.69 m. Data are from Lee & Brown (2009), NWI and tidesandcurrents.noaa.gov.
Estuary Estuary Type Estuary
Area (km2)
Total Wetland
Area (ha)
Tidal
Range (m)
Normalized Freshwater
Flow (m3�m�2�yr�1)
No. Plots
Sampled
Coquille Drowned river mouth 6.9 197 2.16 695 30
Alsea Drowned river mouth 12.5 252 1.97 211 26
Yaquina Drowned river mouth 20.0 265 2.54 63 86
Netarts Bar built 10.4 112 2.09 11 24
Journal of Vegetation ScienceDoi: 10.1111/jvs.12107© 2013 International Association for Vegetation SciencePublished 2013. This article is a US Government work and is in the public domain in the USA. 535
C.N. Janousek & C.L. Folger Environmental gradients in tidal wetlands
Page 3
poorly captured by NWI. Wetlands known to have been
recently restored were not sampled.
A subset of randomly selected points was sampled with
the goal of distributing plots widely among different geo-
graphic regions of each estuary and along the full elevation
and salinity gradients present at the sites. A GIS was used
to select points that were located in the field using sub-
meter accuracy hand-held GPS units (e.g. Trimble GeoXH).
At >80% of the GIS-selected points, up to two additional
plots were established at random distances upslope and
downslope of the initial point at distances ranging from
~3 m to ~100 m or more (depending on the width of the
wetland) to ensure that a broad range of elevations were
sampled. Rarely, random points were relocated a few
meters to the nearest spot of relatively level vegetated wet-
land because they fell onmudflat or disturbed habitat.
Vegetation data
The percentage cover of plant species, including any canopy
cover by shrubs or trees, was estimated between mid-May
and the end of September 2010 inside 166 1.0-m2 plots
(species with <1% cover were treated as zeros) to assess
dominant species. Only the uppermost layer of vegetation
was considered, so cover summed to 100%. Usually two
persons independently assessed percentage cover (gener-
ally to the nearest 1–5%) and these observations were then
averaged for the plot. To more accurately determine species
occurrence and species richness, including understorey spe-
cies, a 0.25-m2 quadrat was nested within the larger plot
and the presence of all rooted vascular plant taxa was
recorded. All taxa were included in richness estimates if at
least part of the plant was located inside the plot. Very small
juvenile plants (i.e. <2–3-cm tall) were ignored because
they were difficult to identity; however, this is believed to
have a negligible influence on richness or cover estimates.
Most plants were identified to species level, but some
were only determined to genus (Agrostis, Galium, Hordeum,
Spergularia, Stellaria; Appendix S1). Most or all bentgrasses
and chickweeds encountered probably could be assigned
to Agrostis stolonifera and Stellaria humifusa respectively, but
some morphological variation was present in the material
that might indicate the presence of additional species.
Hordeum specimens lacking an inflorescence could not be
confidently assigned to one of two species in the flora
(H. jubatum or H. brachyantherum) and were therefore
grouped. Triglochin plants were all treated as T. maritima,
but taxonomists sometimes recognize an additional shorter
species, T. concinna; plants of both morphologies were
observed. Our richness estimates may therefore slightly
underestimate true species richness in the plots, but are
consistent across the study. A small percentage (1.7%) of
plant occurrences (mostly grasses and thistle-like plants)
could not be identified confidently to genus or species
level. Voucher specimens collected for most species per
estuary were deposited at the US EPA (Newport, Oregon,
US); digital images were taken of all plots. Throughout the
remainder of the text, common species are referred to by
genus name only, except when necessary for clarity.
Canopy density
At each plot, light transmission through plant canopies (a
surrogate for canopy density) was estimated by measuring
light incidence above canopies and near the sediment sur-
face with a spherical PAR sensor (LI-COR, Inc., Lincoln,
NE, US). Measurements were made twice at two different
locations within each 0.25-m2 plot and then averaged.
Environmental measurements
Three 5-cm deep soil cores were pooled and then frozen to
assess edaphic characteristics at each plot. Later, after
thawing, samples were homogenized and sub-sampled to
evaluate summer porewater salinity, sediment particle size
and total organic nitrogen content (TON). Sediment grain
size was evaluated using a Coulter LS 100Q counter after
digesting organic matter with hydrogen peroxide and sus-
pending aliquots in a dispersant solution. Data used in this
study were percentage clay content (<3.9 lm). Total
organic nitrogen (%TON)wasmeasured with a Carlo Erba
elemental analyser after removing large roots (generally
>1-mm diameter) and drying and grinding samples. A
refractometer was used to measure pore water salinity by
adding thawed sediment to plastic syringes fitted with
paper filters and extruding water. Salinity is reported
herein as parts per thousand (ppt) even though it techni-
cally has no unit. A second researcher examined a subset
of samples and a majority (97%) of salinity values were
reproducible to�2 ppt. Wetland soil salinity can vary tem-
porally over short time scales, but we were able to obtain
only a single measure per plot during the summer season.
However, in a comparison of a subset of plots (n = 27) for
which a second measurement was collected during the
summer of 2011, the majority of stations, including more
saline ones (>25 ppt), varied by no more than 5 ppt
between years. While our salinity data are noisy, they cap-
ture the overall gradient (fresh to hypersaline) present in
the region during the dry season.
Plot elevations were determined with a survey-grade
Trimble� 4700 GPS receiver using fast static measurements
or by levelling from nearby GPS-surveyed positions. Raw
GPS data were differentially corrected using OPUS-RS soft-
ware (http://www.ngs.noaa.gov/OPUS) with Geoid09.
North American Vertical Datum 1988 (NAVD88) ortho-
metric heights obtained from OPUS-RS were converted to
Journal of Vegetation Science536 Doi: 10.1111/jvs.12107© 2013 International Association for Vegetation Science
Published 2013. This article is a US Government work and is in the public domain in the USA.
Environmental gradients in tidal wetlands C.N. Janousek & C.L. Folger
Page 4
local mean higher high water (MHHW) by measuring
NAVD88 elevations at permanent tidal benchmarks. GPS
vertical precision (SD) measured at a benchmark in South
Beach, OR, was 1.2 cm (n = 28). Repeated measurements
on a subset of plots in the field (n = 12) usually differed by
only 0–4 cm vertically (maximum difference = 5.7 cm).
Tidal benchmarks could not be located in Netarts Bay,
so the NAVD88–local MHHW relationship was determined
by using empirical water level data. Briefly, an Odyssey
water level logger (Dataflow Systems Pty, Christchurch,
NZ) was installed from 28 July to 22 Aug 2011 in a tidal
channel in south Netarts Bay to estimate MHHW for the
current 19-yr tidal epoch. The water level record was cor-
rected with the Garibaldi (Tillamook Bay) control tidal sta-
tion following standard computational methods (NOAA
2003). An estimated NAVD88 to MHHW correction factor
for Netarts was then obtained using the RTK-measured
geodetic elevation of the water level sensor (Trimble� R8
survey-grade GPS, n = 2).
Statistical analyses
All analyses (except path analysis) were conducted with R
(v. 2.14; R Foundation for Statistical Computing, Vienna,
AT). Presence–absence data were used for most analyses,
including hierarchical partitioning and species richness.
Analyses of vegetation composition were based on the per-
centage cover of the most commonly occurring species in
the 1.0-m2 plots (20 species). Due to occasional missing
environmental or species values, there was a small amount
of variation in sample size from analysis to analysis.
The relative effects of five environmental gradients (ele-
vation, soil salinity, estuarine flow, soil N and soil clay) on
the occurrence of 20 common species (each analysed sepa-
rately) were evaluated with logistic regression and hierar-
chical partitioning using the package ‘hier.part’ (Chevan &
Sutherland 1991). Hierarchical partitioning decomposes
variation that is uniquely and jointly accounted for by each
independent variable in the model. The statistical signifi-
cance of effects due to each environmental gradient was
assessed with a randomization test (n = 200). Hydrologic
differences among the four estuaries in the study were rep-
resented by the area-normalized flow index for each site
(Table 1). Sample sizes were n = 159 for all species except
Agrostis (n = 158). Hierarchical partitioning functions opti-
mally when predictor and response variables have mono-
tonic relationships (Heikkinen et al. 2005). We visually
checked for unimodal relationships between the 20 species
and elevation, salinity, TON and clay (flow was repre-
sented by too few values to visualize adequately) and
found that elevation was most likely to have some degree
of a unimodal response with species occurrence. Of the
20 species tested, responses between elevation and
occurrence of Triglochin, Cuscuta, Glaux and Plantago may
be the most strongly unimodal, and so hierarchical parti-
tioning may have underestimated the effect of elevation
relative to other variables in these particular models.
Plant composition was investigated with non-metric
multidimensional scaling (NMDS) using percentage cover
data for the 20 most frequently occurring species with the
package ‘vegan’ (Kruskal 1964a,b). The global NMDS was
run with function ‘metaMDS’ and Bray–Curtis dissimilari-
ties among plots (n = 152). Data were square-root trans-
formed and Wisconsin standardized prior to analysis. The
analysis was run with multiple restarts (n = 872) until two
convergent solutions were obtained, to avoid being
trapped in local minima. The solution was centred, rotated
on principle component axes, and scaled to half-change
units as per default settings with ‘metaMDS’. To graphi-
cally illustrate compositional heterogeneity among and
within different environmental classes, plots in the NMDS
ordination were alternatively coded by estuary (four clas-
ses), summer soil salinity (two classes: oligohaline to mes-
ohaline; polyhaline to hypersaline) or elevation (two
classes: <MHHW, >MHHW). Following the NMDS ordina-
tion, the function ‘envfit’ in ‘vegan’ was used to determine
relationships between composition and the five environ-
mental variables of interest in the study (tidal elevation,
soil salinity, estuary flow, soil N and soil clay content). The
robustness of the NMDS ordination was tested by perform-
ing a detrended correspondence analysis (DCA) on the
same data set with function ‘decorana’ in ‘vegan’. Plot
locations along DCA axes 1 and 2 were highly correlated
with their positions along the two axes in the NMDS ordi-
nation (r = 0.97 and r = 0.75, respectively), indicating that
the reported NMDS ordination axes extracted the main
gradient structure in the data (Økland 1996).
Species accumulation curves with 95% bootstrapped
confidence intervals were used to determine variation in
species richness among several gradients of interest (pack-
age ‘vegan’). Analyses were conducted by alternatively
classifying the same data set (n = 157) by estuary, by ele-
vation class and by salinity class (classes delineated as in
the NMDS comparisons mentioned above). Since a few
plots with missing data were not used (including a few
fresher Coquille plots with some unique species occur-
rences), the Coquille rarefaction analysis may have under-
estimated richness.
Finally, path analysis with maximum likelihood estima-
tion in AMOS (v.18.0.0; Crawfordville, FL, US) was used
to assess the importance of the five environmental factors
for plot-level plant species richness (n = 152; Arnold
1972). Variables were not transformed since univariate
andmultivariate kurtosis was acceptable (Byrne 2010). An
initial model was constructed that included plant canopy
density (measured as light transmission to the soil surface)
Journal of Vegetation ScienceDoi: 10.1111/jvs.12107© 2013 International Association for Vegetation SciencePublished 2013. This article is a US Government work and is in the public domain in the USA. 537
C.N. Janousek & C.L. Folger Environmental gradients in tidal wetlands
Page 5
as a sixth independent variable based on Grace & Pugesek’s
(1997) finding that this was an important predictor of plant
richness. However, an initial analysis showed it to have
essentially no relationship with richness, so this factor was
removed and not considered further. Thereafter a full
model of hypothesized relationships among five environ-
mental variables and plant richness was tested and non-
significant (P > 0.05) pathways were dropped from the
model iteratively. Model fit was initially poor (as suggested
by NFI, CFI, RMSEA, AIC, BCC and Hoelter indices; Byrne
2010), but improved markedly once estuarine flow was
completely removed from the model. In a final iterative
step, the pathway between MHHW and soil N (though
having a strong relationship) was also removed since the
pathway between soil N and richness had already been
eliminated earlier during model simplification. Standard-
ized and non-standardized path coefficients (the former
interpreted similarly to simple correlation coefficients) and
explained variance for exogenous variables in the final
model (R2) are reported.
Results
Overview of floral and environmental variability
Sixty-six taxa of vascular plants were identified to at least
genus level in 166 plots in the study (Appendix S2). Sarco-
cornia, Deschampsia, Juncus balticus subsp. ater, Distichlis, Jau-
mea, Agrostis, Triglochin and Potentillawere among the most
frequently occurring species, each having at least 1% cover
in ≥30% of all surveyed plots (Table 2). Plot elevations
ranged from �1.05 to +0.89 m relative to MHHW, or
approximately the upper half of the intertidal zone in the
region. Summer soil salinities ranged from nearly fresh
(0.5 ppt) to mildly hypersaline (44 ppt) conditions. Eleva-
tion was positively related to soil N content, unimodally
related to soil clay content and negatively correlated with
the amount of light reaching the sediment surface through
plant canopies (Appendix S3). Soil salinity was not
strongly related to any of these three variables.
Environmental gradients and species occurrences
The relative importance of the five environmental gradi-
ents in logistic regression models predicting plant occur-
rence varied by species (hierarchical partitioning; Fig. 1,
Appendix S4). For five of the 12 most common species
(Distichlis, Jaumea, Sarcocornia, Triglochin and Grindelia), soil
salinity was the most important variable (highest
explained model variance) and was positively related to
the likelihood of species occurrence. Elevation was the
most important variable for Hordeum and Atriplex, whereas
soil N was the most important variable for the occurrence
of Agrostis, Potentilla and Juncus balticus (all positively asso-
ciated with soil N). Clay was generally relatively unimpor-
tant except for Deschampsia and Carex lyngbyei. Estuarine
flow was generally either unimportant or of secondary
importance after soil salinity or another variable. Models
for most species left a large percentage of unexplained
Table 2. Common species in the data set. Frequency of occurrence is the percentage of 1-m2 plots in which the species had ≥1% cover.
Species and Abbreviation Frequency (%) Habit, Life History and Endemicity
Sarcocornia perennis (Sp) 44 Forb, perennial, native
Deschampsia cespitosa (Dc) 43 Grass, perennial, native
Juncus balticus subsp. ater (Jb) 40 Rush, perennial, native
Distichlis spicata (Ds) 39 Grass, perennial, native
Jaumea carnosa (Jc) 35 Forb, perennial, native
Agrostis spp., prob. usually A. stolonifera (As) 34 Grass, perennial, native & NIS
Triglochin maritima (Tm) 32 Forb, perennial, native
Potentilla anserina (Pa) 30 Forb, perennial, native
Carex lyngbyei (Cl) 27 Sedge, perennial, native
Hordeum spp. (H) 21 Grass, annual/perennial, native
Atriplex spp. (A) 19 Forb, all or mostly annual, inconclusive
Grindelia stricta (Gs) 16 Forb to subshrub, perennial, native
Carex obnupta (Co) 12 Sedge, perennial, native
Cuscuta pacifica (Cp) 11 Parasite, annual, native
Symphyotrichum subspicatum (Ss) 11 Forb, perennial, native
Spergularia spp. (S) 10 Forb, annual to perennial, native & NIS
Glaux maritima (Gm) 9 Forb, perennial, native
Galium spp. (G) 8 Forb, annual to perennial
Schenodorus arundinaceus (Sa) 7 Grass, perennial, NIS
Castilleja ambigua (Ca) 7 Forb, annual, native
NIS, non-indigenous species.
Habit, life history and endemicity data are from references found in Appendix S1.
Journal of Vegetation Science538 Doi: 10.1111/jvs.12107© 2013 International Association for Vegetation Science
Published 2013. This article is a US Government work and is in the public domain in the USA.
Environmental gradients in tidal wetlands C.N. Janousek & C.L. Folger
Page 6
variance, although total explained variance by all five vari-
ables was relatively high for Juncus balticus (52%), Sarcocor-
nia (39%) and Potentilla (36%).
Environmental gradients and assemblage composition
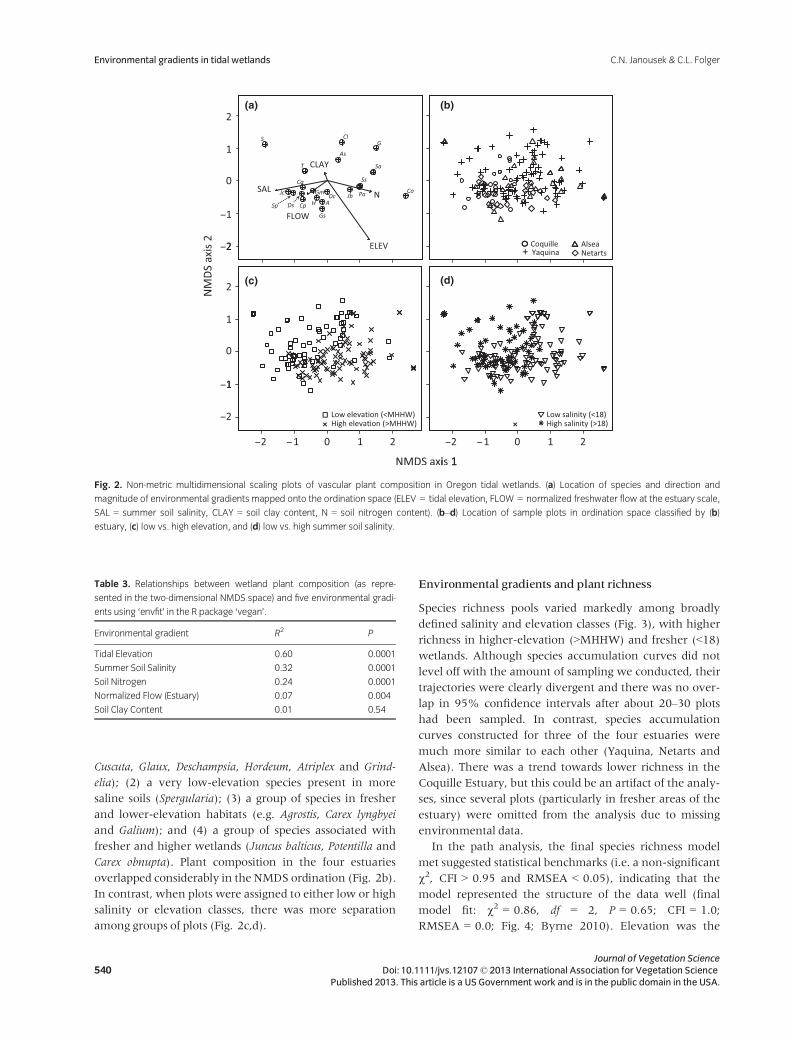
Differences in plant composition were adequately repre-
sented by a two-dimensional NMDS ordination
(stress = 0.11). Four of the five environmental gradients
tested were significantly correlated with species composi-
tion in NMDS space (Fig. 2a, Table 3). Of these gradients,
tidal elevation and soil salinity were most strongly related
to variation in plant composition. Common species tended
to group in a few clusters in ordination space: (1) a large
group of species occupying saline soils across a wide
range of elevations (Jaumea, Sarcocornia, Distichlis, Castilleja,
–5
0
5
10
15
20
25 Joint effectsIndependent effects
–5
0
5
10
15
20
25
–5
0
5
10
15
20
25
–5
0
5
10
15
20
25
Elev Elev ElevSal Flow Clay N Sal Flow Clay N Sal Flow Clay N
Perc
ent v
aria
nce
expl
aine
d*
*
*
* * *
*
*
*
*
*
*
* **
**
*
**
**
* *
*
* **
*
*
**
**
*
**
*
*
* **
**
Environmental gradient
Ds Jc Sp
Tm As Pa
Jb A Dc
H Gs Cl
+–++
+++
++
–++
+ –
++ ++ + +––
–
––
+
+
+++ –– + ++ +
+++ –
++– –
Fig. 1. Percentage variation in species occurrence accounted for by five environmental gradients in hierarchical partitioning of logistic regression models
(Elev = elevation; Sal = soil salinity; Flow = estuarine flow; Clay = percentage clay content of sediment; N = total organic nitrogen). Independent effects
of each factor on species occurrence are in black bars; joint effects of each factor (in combination with all other factors) are in grey bars. Plus and minus
signs below the bars indicate if associations between each statistically significant factor (P < 0.05; indicated by asterisk) and species presence in the
models were positive or negative. Species abbreviations follow codes in Table 2.
Journal of Vegetation ScienceDoi: 10.1111/jvs.12107© 2013 International Association for Vegetation SciencePublished 2013. This article is a US Government work and is in the public domain in the USA. 539
C.N. Janousek & C.L. Folger Environmental gradients in tidal wetlands
Page 7
Cuscuta, Glaux, Deschampsia, Hordeum, Atriplex and Grind-
elia); (2) a very low-elevation species present in more
saline soils (Spergularia); (3) a group of species in fresher
and lower-elevation habitats (e.g. Agrostis, Carex lyngbyei
and Galium); and (4) a group of species associated with
fresher and higher wetlands (Juncus balticus, Potentilla and
Carex obnupta). Plant composition in the four estuaries
overlapped considerably in the NMDS ordination (Fig. 2b).
In contrast, when plots were assigned to either low or high
salinity or elevation classes, there was more separation
among groups of plots (Fig. 2c,d).
Environmental gradients and plant richness
Species richness pools varied markedly among broadly
defined salinity and elevation classes (Fig. 3), with higher
richness in higher-elevation (>MHHW) and fresher (<18)wetlands. Although species accumulation curves did not
level off with the amount of sampling we conducted, their
trajectories were clearly divergent and there was no over-
lap in 95% confidence intervals after about 20–30 plots
had been sampled. In contrast, species accumulation
curves constructed for three of the four estuaries were
much more similar to each other (Yaquina, Netarts and
Alsea). There was a trend towards lower richness in the
Coquille Estuary, but this could be an artifact of the analy-
ses, since several plots (particularly in fresher areas of the
estuary) were omitted from the analysis due to missing
environmental data.
In the path analysis, the final species richness model
met suggested statistical benchmarks (i.e. a non-significant
v2, CFI > 0.95 and RMSEA < 0.05), indicating that the
model represented the structure of the data well (final
model fit: v2 = 0.86, df = 2, P = 0.65; CFI = 1.0;
RMSEA = 0.0; Fig. 4; Byrne 2010). Elevation was the
Table 3. Relationships between wetland plant composition (as repre-
sented in the two-dimensional NMDS space) and five environmental gradi-
ents using ‘envfit’ in the R package ‘vegan’.
Environmental gradient R2 P
Tidal Elevation 0.60 0.0001
Summer Soil Salinity 0.32 0.0001
Soil Nitrogen 0.24 0.0001
Normalized Flow (Estuary) 0.07 0.004
Soil Clay Content 0.01 0.54
(a) (b)
(c) (d)
Fig. 2. Non-metric multidimensional scaling plots of vascular plant composition in Oregon tidal wetlands. (a) Location of species and direction and
magnitude of environmental gradients mapped onto the ordination space (ELEV = tidal elevation, FLOW = normalized freshwater flow at the estuary scale,
SAL = summer soil salinity, CLAY = soil clay content, N = soil nitrogen content). (b–d) Location of sample plots in ordination space classified by (b)
estuary, (c) low vs. high elevation, and (d) low vs. high summer soil salinity.
Journal of Vegetation Science540 Doi: 10.1111/jvs.12107© 2013 International Association for Vegetation Science
Published 2013. This article is a US Government work and is in the public domain in the USA.
Environmental gradients in tidal wetlands C.N. Janousek & C.L. Folger
Page 8
strongest driver of plot-level species richness among the
variables retained in the final model. Richness was highest
in plots above MHHW (Fig. 5). Soil salinity and soil clay
content were also positively related to richness, but stan-
dardized path coefficients were about three times smaller
for these variables than for elevation. Soil N, although
strongly affected by elevation (e.g. Appendix S3), and estu-
arine flow were not significantly related to richness and
were dropped from the final model.
Discussion
Many abiotic factors and biological interactions among
species are known to affect plant composition and diversity
in estuarine wetlands (Engels & Jensen 2009; Keammerer
& Hacker 2013). Gradients of salinity, elevation and other
edaphic variables may co-occur in complex patterns
(Odum 1988). Our aim in this study was to quantify the
relative strength of relationships between major environ-
mental gradients and wetland plant composition and rich-
ness and use these results to inform projections about
future changes to tidal wetlands, particularly related to sea
level rise. Our analyses suggest that: (1) the environmental
gradients most strongly associated with plant distribution
differ to some extent among species; (2) tidal elevation is
the principal driver of aggregate assemblage composition
and diversity; and (3) hydrologic differences among estuar-
ies are generally of only secondary importance, relative to
other gradients, in shaping plant distribution and composi-
tion.
Environmental gradients and species occurrences
Soil salinity, tidal elevation and soil N content were alter-
natively the most important environmental correlates of
species occurrences for the common taxa in our study. Soil
salinity was the variable most strongly associated with the
distribution of five of the 12 most common species, all of
which were more common in more saline habitats. Wat-
son & Byrne (2009) also found that salinity was associated
with the distribution of many species across the San Fran-
cisco Estuary (positively correlated with Sarcocornia and
negatively associated with other species like Scirpus califor-
nicus). Our data cannot causally explain distribution, but
other research shows that halophytes may be limited to
more saline wetlands due to competitive displacement
(Crain et al. 2004; Engels & Jensen 2010; but see also Guo
& Pennings 2012). Occurrences of a few less common spe-
cies in our data set (Carex obnupta and Galium spp.) were
negatively related to soil salinity (Appendix S4).
0 20 40 60 800 20 40 60 800 20 40 60 80
0
10
20
30
40
50
60
Number of plots sampled
Spec
ies
rich
ness
Yaquina>MHHW
<MHHW
Coquille
Alsea
Netarts
>18 ppt
<18 ppt(a) (b) (c)
Fig. 3. Species accumulation curves for (a) each estuary in the study, (b) high vs. low tidal elevation plots, and (c) high vs. low salinity plots. Polygons show
95% confidence intervals.
Tidal elevation
Soil clay content
Soilsalinity
Soilnitrogen
Plantrichness
R2 = 0.32
Estuary normalized
-flow
–0.35 (–11.35)
0.20 (0.04)
0.19(0.08)
0.57 (3.63)
R2 = 0.12
Fig. 4. Path diagram of environmental gradient effects on vascular plant
richness. Pathways in the final model are shown in black with standardized
and non-standardized (in parentheses) coefficients. Path widths are scaled
by standardized path coefficients. Non-significant variables and pathways
are in grey (soil N was strongly correlated with tidal elevation, but this path
was not included in the final model since the N to richness path was
eliminated during model selection).
Journal of Vegetation ScienceDoi: 10.1111/jvs.12107© 2013 International Association for Vegetation SciencePublished 2013. This article is a US Government work and is in the public domain in the USA. 541
C.N. Janousek & C.L. Folger Environmental gradients in tidal wetlands
Page 9
Tidal elevation was the dominant environmental driver
for only a few common species we tested, including Atri-
plex and Hordeum. However, although not the single most
important gradient, elevation was also relatively strongly
related to the distribution of Potentilla, Deschampsia and
Grindelia. All five of these species tend to prefer higher
marshland. Very little variation in the distribution of Jau-
mea, Triglochin or Agrostis was explained by tidal elevation.
However, the relative importance of elevation on the
occurrence of Sarcocornia, Distichlis, Jaumea, Agrostis, Grind-
elia, and particularly Triglochin, Glaux and Plantago, might
have been underestimated since these species might have
some degree of unimodal relationships with elevation,
while hierarchical partitioning requires linear relationships
between predictor and independent variables (Heikkinen
et al. 2005).Agrostis occurred across a broad range of eleva-
tions, suggesting that it is tolerant of wide variation in
flooding intensity.
The distribution of a few species was correlated princi-
pally with other environmental gradients. For instance,
Juncus balticus had a strong affinity for N-rich soils. Tjepk-
ema & Evans (1976) observed N fixation associated with
this species in an estuary in northern Oregon - a potential
cause of this association. Davy et al. (2011) found that ele-
vation, soil redox conditions, and soil salinity have differ-
ent magnitudes of effect on species cover or occurrence in
a restored salt marsh in England, depending on the species
under consideration.
Environmental gradients and plant composition and
richness
While various environmental variables were most strongly
correlated with individual species distributions, tidal eleva-
tion appeared to be the dominant driver of overall plant
composition and plot-level richness. Tidal elevation was
the strongest correlate of plant composition in the NMDS
ordination (Table 3) and of species richness in the path
analysis (Figs 4 and 5). Tidal elevation itself is tightly cou-
pled to a variety of environmental variables, including
flooding frequency and duration and soil redox potential
(Davy et al. 2011). Increased submergence at lower eleva-
tions is known to negatively affect many wetland species,
potentially limiting diversity in these wetlands (Grace &
Pugesek 1997). Elevation was also related to other envi-
ronmental gradients such as soil N content (Fig. 4, Appen-
dix S3). In the path analysis, elevation affected plant
richness both through a strong direct effect and an indirect
effect via soil salinity.
We found maximum plant richness at elevations above
MHHW and much lower richness in frequently flooded
plots. Salinity and elevation have both been considered as
limiting plant richness in tidal wetland ecosystems (Odum
1988; Gough et al. 1994; Engels & Jensen 2009) by reduc-
ing the pool of species tolerant to salinity or flooding stress
(Grace & Pugesek 1997). Our results are consistent with
studies that show a positive relationship between plant
richness and tidal elevation (Gough et al. 1994; Grace &
Pugesek 1997; Kunza & Pennings 2008; Engels & Jensen
2009; Moeslund et al. 2011).
Soil salinity had a weak positive relationship with plot-
level plant richness in the path analysis, in contrast to
other research showing that fresher wetlands are more
diverse (Garc�ıa et al. 1993; Gough et al. 1994; Grace &
Pugesek 1997; Engels & Jensen 2009; Sharpe & Baldwin
2009; Watson & Byrne 2009;Wiezski et al. 2010). In simple
regression, plant richness was neither significantly related
to summer salinity nor to winter salinity measured in the
following season at the same plots (data not shown). Weil-
hoefer et al. (2013) also only found a weak negative salin-
ity–diversity relationship in the Oregon estuarine marsh
flora. While our species accumulation results show that
fresher wetlands do have a larger overall species pool, plot-
level richness did not differ between fresher and more
–1.0 –0.5 0.0 0.5 1.0 0 10 20 30 40 0 5 10 15 20 25 30
12
10
8
6
4
2
0
Elevation above MHHW (m) Soil salinity (ppt) Sediment clay content (%)
Plan
t ric
hnes
s
Fig. 5. Relationships between species richness and elevation, summer soil salinity and soil clay content. Plot symbols differ by estuary as in Fig 2b.
Journal of Vegetation Science542 Doi: 10.1111/jvs.12107© 2013 International Association for Vegetation Science
Published 2013. This article is a US Government work and is in the public domain in the USA.
Environmental gradients in tidal wetlands C.N. Janousek & C.L. Folger
Page 10
saline assemblages. Higher beta diversity in fresher wet-
lands may explain this discrepancy: assemblages in more
saline habitats can have high richness, but show little
variation in composition from plot-to-plot, whereas assem-
blages in fresher wetlands may differ more composition-
ally. In summary, salinity appears to have a relatively
strong effect on composition across Oregon estuaries (e.g.
Weilhoefer et al. 2013) and on landscape-scale diversity,
but not on plot-level richness.
Estuary-scale variation in hydrology
Our results suggest that hydrologic differences among estu-
aries play only a secondary role in structuring plant abun-
dance, composition or species richness. Some rarer species
in our data set were found in only one or a few estuaries,
but common taxa were usually present in all estuaries,
regardless of hydrologic condition (the absence of Grindelia
in Coquille is a notable exception). While hydrology may
be expected to greatly impact salinity regimes for lower-
elevation estuarine habitats such as seagrass beds, high
tidal wetlands may be relatively less impacted by variation
in the degree of river dominance because they are rarely
flooded. Groundwater inputs or precipitation may be com-
paratively more important determinants of salinity. For
instance, even during the summer in Netarts Bay (a site
expected overall to be very saline), wetlands at very high
tidal elevations had surprisingly low salinities, perhaps due
to groundwater inputs or retention of spring precipitation.
Plant composition, such as the presence of a large swath of
salt-intolerant Carex obnuptamarsh at the estuarine–upland
margin, reflected these edaphic conditions. Conversely,
many low-marsh plots in the river-dominated Coquille
estuary resembled low marshes in the other estuaries pop-
ulated by succulent halophytes, probably because they
experience frequent tidal inundation and are situated geo-
graphically close to themouth of the estuary.
While our results suggest that watershed-level hydrol-
ogy may not be a major driver of plant composition in Ore-
gon tidal wetlands, additional data are needed on linkages
between watershed hydrology and overall wetland species
richness and composition. In a study of the deltas of 17
estuaries in the Puget Sound region of Washington and
British Columbia, Hutchinson (1988) found a fairly high
degree of correspondence between marsh assemblages and
estuary-level environmental metrics (e.g. river discharge/
near-shore wave power). He suggested that salinity differ-
ences among estuaries drive these patterns, with fresher
estuaries having increased dominance of species such as
Carex lyngbyei, Triglochin and Schoenoplectus americanus, and
more saline estuaries having higher abundance of Sarcocor-
nia, Distichlis, Juncus balticus and Potentilla. Few other stud-
ies appear to examine relationships between wetland plant
composition and estuarine hydrology at regional scales.
Sharpe & Baldwin (2009) found that plant composition
varied more among wetlands from different salinity
regimes that were located relatively close geographically
than between wetlands of similar salinity classes located
on opposite sides of the Chesapeake Bay estuary. Kunza &
Pennings (2008) suggested that higher diversity in Texas
vs. Georgia salt marshes may be due to differences in tidal
regimes.
Implications for climate change
Future coastal wetlands are likely to experience a suite of
environmental changes, including higher temperatures,
elevated CO2, sea level rise and changes in estuarine
hydrology (Parker et al. 2011). Our data on vegetation
relationships with some of these gradients suggess that
plant composition may be particularly sensitive to changes
in relative flooding intensity in estuarine marshes. Modest
degrees of relative SLR may have a large effect on species
composition and diversity by changing flooding regimes
and soil biogeochemistry (Baldwin et al. 2001). Sea level
rise modelling by Moeslund et al. (2011) and Stralberg
et al. (2011) suggests that higher tidal wetlands may be
particularly vulnerable. Sea level rise may also result in
higher salinities in wetland soils in more inland regions of
estuaries (Callaway et al. 2007). Changes in the timing of
coastal precipitation, such as intensified seasonality, may
also change salinity, although changes in snowfall-to-pre-
cipitation ratios are unlikely to impact salinity because
snowfall is not a major source of freshwater in the Oregon
coast range. Elevated salinity may have secondary effects
on composition by favouring halophytes such as Sarcocor-
nia, Distichlis, Grindelia and Jaumea at the expense of less
salt-tolerant species.
Future SLR is dependent on multiple factors that cannot
yet be accurately projected. Local sediment supply, in situ
organic matter production or sediment compaction, and
global SLR (itself affected by future rates of anthropogenic
CO2 emissions, ice sheet melting, etc.), are all important
components for projecting the futuremagnitude of relative
SLR. Despite these uncertainties, if wetland accretion rates
cannot match relative SLR, the direction of climate change
effects on estuaries is clear: flooding and salinity stresses on
plants will increase. Our data suggest that increasing these
factors will change plant composition and lower vascular
plant richness.
Conclusions
Plant composition and richness varied across a suite of
environmental gradients in the studied wetlands. Relative
gradient importance depended in part on which metrics of
Journal of Vegetation ScienceDoi: 10.1111/jvs.12107© 2013 International Association for Vegetation SciencePublished 2013. This article is a US Government work and is in the public domain in the USA. 543
C.N. Janousek & C.L. Folger Environmental gradients in tidal wetlands
Page 11
plant composition were under investigation, but elevation
and salinity consistently emerged as important drivers of
vegetation structure. To the extent that future SLR
increases submergence times and salinity exposure in Paci-
fic Northwest tidal marshes and swamps, our results sug-
gest that vegetation composition will change and wetland
plant richness will decline. Altered vegetation structure in
these important coastal ecosystems may in turn lead to
changes in ecosystem function.
Acknowledgements
We thank D. Beugli, H. Brunner, V. Goldsmith, J. Saari-
nen, K. Marko, J. Stecher, T. Ernst, M. Frazier, L. Brophy,
R. Loiselle, T. MochonCollura, S. Cline and M. Armstrong
for various lab, field, technical and statistical assistance. P.
Clinton kindly provided GIS support. B. Watson, M. Fra-
zier, R. Halvorsen and two anonymous reviewers made
helpful comments on the manuscript. Sampling was made
possible by the cooperation of numerous landholders
including the US Fish andWildlife Service (Bandon NWR),
BLM, US Forest Service, Oregon State Parks, Oregon State
University, Port of Toledo, Port of Bandon, The Wetlands
Conservancy, van Eck Foundation and other private own-
ers. The information in this publication has been funded
by the U.S. Environmental Protection Agency. It has been
subjected to review by the National Health and Environ-
mental Effects Research Lab and approved for publication.
Approval does not signify that the contents reflect the
views of the Agency, nor does mention of trade names or
commercial products constitute endorsement or recom-
mendation for use.
References
Arnold, S.J. 1972. Species densities of predators and their prey.
The American Naturalist 106: 220–236.
Baldwin, A.H., Egnotovich, M.S. & Clarke, E. 2001. Hydrologic
change and vegetation of tidal freshwater marshes: field,
greenhouse, and seed-bank experiments. Wetlands 21:
519–531.
Barbier, E.B., Hacker, S.D., Kennedy, C., Koch, E.W., Stier, A.C.
& Silliman, B.R. 2011. The value of estuarine and coastal
ecosystem services. Ecological Monographs 81: 169–193.
Byrne, B.M. 2010. Structural equation modeling with AMOS, 2nd
edn. 396 pp. Routledge, New York, NY, US.
Callaway, J.C., Parker, V.T., Vasey, M.C. & Schile, L.M. 2007.
Emerging issues for the restoration of tidal marsh ecosystems
in the context of predicted climate change. Madro~no 54:
234–248.
Chevan, A. & Sutherland, M. 1991. Hierarchical partitioning.
The American Statistician 45: 90–96.
Cowardin, L.M., Carter, V., Golet, F.C. & LaRoe, E.T. 1979. Classi-
fication of wetlands and deepwater habitats of the United States. US
Fish and Wildlife Publication FWS/BSB-79/31, Washington,
DC, US.
Crain, C.M., Silliman, B.R., Bertness, S.L. & Bertness, M.D.
2004. Physical and biotic drivers of plant distribution across
estuarine salinity gradients. Ecology 85: 2539–2549.
Cui, B.-S., He, Q. & An, Y. 2011. Community structure and abi-
otic determinants of salt marsh plant zonation vary across
topographic gradients. Estuaries and Coasts 34: 459–469.
Davy, A.J., Brown, M.J.H., Mossman, H.L. & Grant, A. 2011.
Colonization of a newly developing salt marsh: disentangling
independent effects of elevation and redox potential on halo-
phytes. Journal of Ecology 99: 1350–1357.
Eilers, H.P. 1975. Plants, plant communities, net production and tide
levels: the ecological biogeography of the Nehalem salt marshes, Til-
lamook County, Oregon. Ph.D. thesis, Oregon State University,
Corvallis, OR, US.
Engels, J.G. & Jensen, K. 2009. Patterns of wetland plant diver-
sity along estuarine stress gradients of the Elbe (Germany)
and Connecticut (USA) rivers. Plant Ecology and Diversity 2:
301–311.
Engels, J.G. & Jensen, K. 2010. Role of biotic interactions and
physical factors in determining the distribution of marsh
species along an estuarine salinity gradient. Oikos 119:
679–685.
Engle, V.D. 2011. Estimating the provision of ecosystem services
by Gulf ofMexico coastal wetlands.Wetlands 31: 179–193.
Garc�ıa, L.V., Mara~n�on, T., Moreno, A. & Clemente, L. 1993.
Above-ground biomass and species richness in a Mediterra-
nean salt marsh. Journal of Vegetation Science 4: 417–424.
Gedan, K.B., Crain, C.M. & Bertness, M.D. 2009. Small-mammal
herbivore control of secondary succession in New England
tidal marshes. Ecology 90: 430–440.
Gough, L., Grace, J.B. & Taylor, K.L. 1994. The relationship
between species richness and community biomass: the
importance of environmental variables.Oikos 70: 271–279.
Grace, J.B. & Pugesek, B.H. 1997. A structural equation model of
plant species richness and its application to a coastal wetland.
American Naturalist 149: 436–460.
Guo, H. & Pennings, S.C. 2012. Mechanismsmediating plant dis-
tributions across estuarine landscapes in a low-latitude tidal
estuary. Ecology 93: 90–100.
Heikkinen, R.K., Luoto, M., Kuussaari, M. & P€oyry, J. 2005.
New insights into butterfly–environment relationships using
partitioning methods. Proceedings of the Royal Society, Series B
272: 2203–2210.
Hutchinson, I. 1988. The biogeography of the coastal wetlands of
the Puget Trough: deltaic form, environment, and marsh
community structure. Journal of Biogeography 15: 729–745.
Jefferson, C.A. 1975. Plant communities and succession in Oregon
coastal salt marshes. PhD dissertation, Oregon State University,
Corvallis, OR, US.
Keammerer, H.B. & Hacker, S.D. 2013. Negative and neutral
marsh plant interactions dominate in early life stages and
across physical gradients in an Oregon estuary. Plant Ecology
214: 303–315.
Journal of Vegetation Science544 Doi: 10.1111/jvs.12107© 2013 International Association for Vegetation Science
Published 2013. This article is a US Government work and is in the public domain in the USA.
Environmental gradients in tidal wetlands C.N. Janousek & C.L. Folger
Page 12
Kruskal, J.B. 1964a. Multidimensional scaling by optimizing
goodness of fit to a nonmetric hypothesis. Psychometrika 29:
1–27.
Kruskal, J.B. 1964b. Nonmetric multidimensional scaling: a
numerical method. Psychometrika 29: 115–129.
Kunza, A.E. & Pennings, S.C. 2008. Patterns of plant diversity in
Georgia and Texas salt marshes. Estuaries and Coasts 31: 673–
681.
Lee, H. II & Brown, C.A. (eds.) 2009. Classification of regional pat-
terns of environmental drivers and benthic habitats in Pacific
Northwest estuaries. US EPA, Office of Research and Develop-
ment, Newport, OR, US.
Liverman, M.C. 1981. Multivariate analysis of a tidal marsh ecosys-
tem at Netarts Spit, Tillamook County, Oregon. MS Thesis Oregon
State University, Corvallis, OR, US.
Moeslund, J.E., Arge, L., Bøcher, P.K., Nygaard, B. & Svenning,
J.-C. 2011. Geographically comprehensive assessment of
salt–meadow vegetation–elevation relations using LiDAR.
Wetlands 31: 471–482.
National Oceanic and Atmospheric Administration. 2003. Com-
putational techniques for tidal datums handbook. NOAA Special
Publication NOS CO-OPS 2, US Department of Commerce,
Silver Spring, MD, US.
Odum, W.E. 1988. Comparative ecology of tidal freshwater and
salt marshes.Annual Review of Ecology and Systematics 19: 147–
176.
Økland, R.H. 1996. Are ordination and constrained ordination
alternative or complementary strategies in general ecological
studies? Journal of Vegetation Science 7: 289–292.
Parker, V.T., Callaway, J.C., Schile, L.M., Vasey, M.C. & Herbert,
E.R. 2011. Climate change and San Francisco Bay-Delta tidal
wetlands. San Francisco Estuary andWatershed Science 9: 1–15.
Pennings, S.C., Selig, E.R., Houser, L.T. & Bertness, M.D. 2003.
Geographic variation in positive and negative interactions
among salt marsh plants. Ecology 84: 1527–1538.
Pennings, S.C., Grant, M.-B. & Bertness, M.D. 2005. Plant zona-
tion in low-latitude salt marshes: disentangling the roles of
flooding, salinity and competition. Journal of Ecology 93: 159–
167.
Rand, T.A. 2000. Seed dispersal, habitat suitability and the distri-
bution of halophytes across a salt marsh tidal gradient. Jour-
nal of Ecology 88: 608–621.
Sharpe, P.J. & Baldwin, A.H. 2009. Patterns of wetland plant spe-
cies richness across estuarine gradients of Chesapeake Bay.
Wetlands 29: 225–235.
Silvestri, S., Defina, A. & Marani, M. 2005. Tidal regime, salinity
and salt marsh plant zonation. Estuarine Coastal Shelf Science
62: 119–130.
Stralberg, D., Brennan, M., Callaway, J.C., Wood, J.K., Schile,
L.M., Jongsomjit, D., Kelly, M., Parker, V.T. & Crooks, S.
2011. Evaluating tidal marsh sustainability in the face of sea-
level rise: a hybrid modeling approach applied to San Fran-
cisco Bay. PLoS One 6: e27388.
Tjepkema, J.D. & Evans, H.J. 1976. Nitrogen fixation associated
with Juncus balticus and other plants of Oregon wetlands. Soil
Biology and Biochemistry 8: 505–509.
Watson, E.B. & Byrne, R. 2009. Abundance and diversity of tidal
marsh plants along the salinity gradient of the San Francisco
Estuary: implications for global change ecology. Plant Ecology
205: 113–128.
Weilhoefer, C.L., Nelson, W.G., Clinton, P. & Beugli, D.M. 2013.
Environmental determinants of emergent macrophyte vege-
tation in Pacific Northwest estuarine tidal wetlands. Estuaries
and Coasts 36: 377–389.
Wiezski, K., Guo, H., Craft, C.B. & Pennings, S.C. 2010. Ecosys-
tem functions of tidal fresh, brackish, and salt marshes on
the Georgia coasts. Estuaries and Coasts 33: 161–169.
Zedler, J.B. 2010. How frequent storms affect wetland vegeta-
tion: a preview of climate-change impacts. Frontiers in Ecology
and the Environment 8: 540–547.
Zedler, J.B., Callaway, J.C., Desmond, J.S., Vivian-Smith, G.,
Williams, G.D., Sullivan, G., Brewster, A.E. & Bradshaw,
B.K. 1999. California salt-marsh vegetation: an improved
model of spatial pattern. Ecosystems 2: 19–35.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Appendix S1. Floristic and natural history references
used in the study.
Appendix S2. Relationships between soil N, soil clay
content, PAR transmission, tidal elevation and soil salinity.
Appendix S3. Percent variation in minor species’
occurrence accounted for by five environmental gradients
in hierarchical partitioning of multivariate logistic regres-
sion models (Elev, elevation; Sal, soil salinity; Flow, estua-
rine flow; Clay, percent clay content of sediment; N, total
organic nitrogen).
Appendix S4. List of species, by estuary, located
inside 0.25m2 plots in the study
Journal of Vegetation ScienceDoi: 10.1111/jvs.12107© 2013 International Association for Vegetation SciencePublished 2013. This article is a US Government work and is in the public domain in the USA. 545
C.N. Janousek & C.L. Folger Environmental gradients in tidal wetlands