YabA and DnaD inhibit helix assembly of the DNA replicationinitiation protein DnaA
Graham Scholefield and Heath Murray*Centre for Bacterial Cell Biology, Institute for Cell andMolecular Biosciences, Newcastle University, NewcastleUpon Tyne NE2 4AX, UK.
Summary
Control of DNA replication initiation is essential for cellgrowth. A unifying characteristic of DNA replicationinitiator proteins is their distinctive AAA+ nucleotide-binding domains. The bacterial initiator DnaA assem-bles into a right-handed helical oligomer built uponinteractions between neighbouring AAA+ domains toform an active initiation complex. Recently we devel-oped a unique cross-linking assay that specificallydetects ATP-dependent DnaA helix assembly. Here wehave utilized this assay to show that two DnaA regu-latory proteins in Bacillus subtilis, YabA and DnaD,inhibit DnaA helix formation. These results, in combi-nation with our previous finding that the regulatoryfactor Soj/ParA also targets DnaA filament formation,highlight the critical importance of regulating DnaAhelix formation during the initiation reaction. Moreo-ver, these observations lead us to suggest that DnaAoligomerization may be the main regulatory step of theinitiator assembly pathway in B. subtilis, in contrastto the prevailing model of bacterial DNA replicationbased on Escherichia coli DnaA where ATP bindingappears to be the targeted activity.
Introduction
Accurate genome transmission is an absolute necessity forthe viability of all cells, and mutations affecting the regula-tion of DNA replication initiation can cause genetic insta-bility. In most cases DNA replication must be initiatedonce (and only once) per cell cycle to ensure preciseco-ordination of genome duplication and segregation.Throughout the three kingdoms of life specific proteins,DnaA in bacteria, Orc1/Cdc6 in archaea and ORC ineukaryotes, bind to DNA where they promote the initiationof DNA replication (Mott and Berger, 2007; Wigley, 2009;
Kawakami and Katayama, 2010). A unifying feature ofthese initiator proteins is their AAA+ nucleotide-bindingdomain, which is necessary for their structure and function(Tucker and Sallai, 2007).
The domain architecture of DnaA is critical for under-standing its role at the bacterial origin of replication (oriC).Domain IV, located at the C-terminus of DnaA, contains aclassical helix–turn–helix motif that specifically recognizes9 base-pair duplex DNA sequences (‘DnaA-boxes’)located within oriC (Fujikawa et al., 2003). Domain III iscomposed of the AAA+ motif that binds ATP and displaysa relatively low intrinsic ATPase activity (Neuwald et al.,1999; Nishida et al., 2002). ATP binding allows DnaA totransition from a monomer into a right-handed helicaloligomer (Erzberger et al., 2006). The distinguishingfeature of initiator AAA+ proteins is the presence of theinitiator-specific motif (ISM) within domain III, which con-sists of two α-helices that form a V-shaped steric wedgeand guide the DnaA oligomer into a non-planar helicalarrangement (Iyer et al., 2004; Erzberger et al., 2006).Additionally, the ISM forms a ‘shelf’ that interacts withsingle-stranded DNA (Ozaki et al., 2008; Duderstadt et al.,2011). Domain IV is connected to domain III by a semi-flexible α-helix that bends to accommodate the packing ofdomain IV into domain III of an adjacent DnaA proteinwithin the homo-oligomer (Erzberger et al., 2002; 2006;Duderstadt et al., 2010). Domain II tethers domains III/IVto domain I and is thought to primarily act as a flexiblelinker; however, there is also evidence to suggest that itmay be involved in helicase recruitment (Molt et al., 2009).Finally, domain I acts as a protein interaction hub thatpromotes DnaA dimerization and facilitates loading of thereplicative helicase (Weigel et al., 1999; Felczak et al.,2005; Abe et al., 2007).
Bacterial chromosomes are typically replicated bi-directionally from a single origin. oriC is composed of twofunctional elements, a set of DnaA-boxes that are used tolocalize DnaA at the origin and an intrinsically unstableAT-rich stretch of DNA (the DNA unwinding element orDUE) (Bramhill and Kornberg, 1988). Based largely onwork using Escherichia coli, it is thought that DnaA occu-pies the DnaA-boxes which most closely match theconsensus sequence (‘strong sites’) throughout the cellcycle (Samitt et al., 1989). These strong sites act as anchorpoints for the recruitment of additional ATP-bound DnaA
proteins onto DnaA-boxes which deviate from the consen-sus sequence (‘weak sites’) (Miller et al., 2009). This initialextended DnaA oligomer, which is bound to double-stranded DNAthrough domain IV, is thought to facilitate thesubsequent formation of a distinct compact DnaA-helixthat engages the upper strand of the DUE through residuesin domain III and promotes DNA duplex unwinding (Speckand Messer, 2001; Ozaki et al., 2008; Duderstadt et al.,2010; 2011; Ozaki and Katayama, 2012; Scholefield et al.,2012). The propensity of DnaA to assemble into a helicaloligomer in vivo has been shown to strongly correlate withDNA replication initiation frequency (Scholefield et al.,2012). In addition, amino acid substitutions in DnaA thatperturb helix formation in vitro inhibit replication originunwinding in vitro and functionality in vivo (Duderstadtet al., 2010). These results demonstrate the critical role forDnaA helix formation during DNA replication initiation.
Following DnaA assembly at oriC in Bacillus subtilis,two additional initiation proteins are sequentially recruited(DnaA → DnaD → DnaB) and are required to depositthe helicase loader:DNA helicase complex (DnaI:DnaCrespectively) onto the unwound DNA duplex (Smits et al.,2010). Both DnaD and DnaB proteins form higher-orderoligomeric complexes that are capable of binding andremodelling DNA (see Briggs et al., 2012). Interestingly,although DnaD directly interacts with DnaA and is requiredfor initiation of DNA replication, it has also been reportedto inhibit the cooperative DNA binding activity of DnaA(Ishigo-Oka et al., 2001; Bonilla and Grossman, 2012).These results suggest that DnaD may act as a switch toregulate DNA replication initiation. After helicase loadingand extension of the open complex, the replisome is con-structed and DNA polymerization commences.
The assembly of DnaA during bacterial DNA replicationinitiation is tightly regulated by multiple mechanisms (forreview see Katayama et al., 2010). DnaA binding to oriCcan be inhibited either by protein occlusion (SeqAin E. coli,Spo0A in B. subtilis and CtrA in Caulobacter crescentus),by spatial sequestration (YabA in B. subtilis), or by titration[datA in E. coli and DBCs in B. subtilis (Okumura et al.,2012)]. DnaA assembly at oriC can be either stimulated(DiaA in E. coli and HobA in Helicobacter pylori) orrepressed (SirA in B. subtilis) by the binding of regulatoryproteins to domain I. DnaA helix formation has also beenfound to be inhibited by monomeric Soj in B. subtilis(Scholefield et al., 2012). Lastly, DnaA is inactivated fol-lowing replisome formation through the stimulation of itsATP hydrolysis activity [Hda in E. coli and C. crescentus;datA in E. coli (Kasho and Katayama, 2013)].
In B. subtilis the negative regulator YabA was first iden-tified in a genome-wide yeast two-hybrid screen looking forfactors that interact directly with known replication pro-teins, and it was shown to form a ‘protein bridge’ betweenthe initiator DnaA and the DNA polymerase sliding clamp
processivity factor, DnaN (Noirot-Gros et al., 2002).Genetic analysis showed that deletion of yabA causeddramatic overinitiation of DNA replication and that YabAoverproduction inhibited DNA replication initiation, indicat-ing that YabA functions as a negative regulator of DnaA(Noirot-Gros et al., 2002; Goranov et al., 2009). Localiza-tion of YabA fused to fluorescent reporter proteins found itto be associated with the replisome during DNA replicationthrough its interaction with DnaN (Hayashi et al., 2005;Noirot-Gros et al., 2006; Goranov et al., 2009). Further-more, it has been shown that the YabA:DnaN complextethers DnaA to the replisome and that DnaA is enriched atoriC in the absence of YabA, suggesting that YabA acts byspatially sequestering DnaA away from the replicationorigin (Soufo et al., 2008; Merrikh and Grossman, 2011).
However, the YabA spatial sequestration model predictsthat overexpression of DnaN should inhibit DNA replica-tion initiation by titrating YabA and DnaA away from oriC(Merrikh and Grossman, 2011). In contrast, although over-production of DnaN does result in decreased associationof YabA with oriC, it also caused YabA-dependent over-initiation of DNA replication (Goranov et al., 2009; Merrikhand Grossman, 2011). This result indicates that DnaNantagonizes YabA inhibition of DnaA. Consistent with thisnotion, it has been shown that recombinant YabA inhibitsATP-dependent cooperative DNA binding of DnaA at oriCin vitro (Merrikh and Grossman, 2011).
In this report we have investigated the mechanismthrough which YabA inhibits DnaA activity. Employing agenetic screen we identified mutations in dnaA that renderthe protein insensitive to YabA regulation in vivo. We thenutilized one of these DnaA variants to show that YabAspecifically inhibits DnaAhelix formation in vitro. Moreover,based on the striking similarity of our results here regard-ing YabA, our previously reported results regarding Soj(Scholefield et al., 2012), and a recent report regardingDnaD (Bonilla and Grossman, 2012), we have gone on toshow that DnaD also inhibits DnaA helix formation in vitro.Taken together, these results indicate that all of the knownregulators of B. subtilis DnaA present during vegetativegrowth specifically target DnaA helix formation during theinitiation reaction. These results highlight the critical impor-tance of regulating DnaA helix formation during the initia-tion reaction and lead us to suggest that DnaAoligomeriza-tion may be the key regulated step of the initiator assemblypathway in B. subtilis.
Results
YabA does not inhibit DnaA ATP binding or ATPhydrolysis activities
Previous studies indicated that YabA specifically inhibitsthe ATP-dependent cooperative DNA binding activity of
DnaA (Merrikh and Grossman, 2011). Because it has beensuggested that YabA interacts with the AAA+ domain ofDnaA (Cho et al., 2008), we tested whether YabA affectsthe ATP binding activity of DnaA. An ATP hydrolysisdeficient DnaA protein (DnaACC,R313A; note that all purifiedDnaA proteins used throughout this study harbourtwo cysteine substitutions, N191C and A198C, that arerequired for DnaA helix formation assays) was incubatedwith [α-P32]-ATP in the presence and absence of YabA; theDnaACC,R313A protein was utilized to prevent hydrolysis ofthe bound ATP during the experiment. After 10 min at 37°Cthe His-tagged proteins were separated from the reactionbuffer using magnetic nickel beads and washed to removeunbound ATP. The bound ATP was extracted and thenseparated by thin-layer chromatography. There was nosignificant difference between the amount of ATP bound toDnaACC,R313A in the presence or absence of YabA (Fig. 1A),indicating that YabAdoes not inhibit theATP binding activityof DnaACC,R313A.
We next investigated whether YabA affected theATPase activity of wild-type DnaA, since stimulation ofATP hydrolysis would be expected to decrease ATP-dependent cooperative DNA binding activity of DnaA.DnaACC ATPase activity was detected using a phosphaterelease assay and was observed to be fivefold above theATP hydrolysis deficient DnaACC,R313A protein (Fig. 1B).The amount of phosphate released in the presence ofYabA was the sum of the intrinsic ATPase activity ofDnaACC and the contaminating ATPase activity present inthe YabA preparation, indicating that YabA does not alterthe ATPase activity of wild-type DnaA (Fig. 1B). Takentogether, these results indicate that YabA does not affectthe ATP binding activity or ATP hydrolysis activity ofDnaA.
Mutations in dnaA disrupt YabA inhibition in vivo
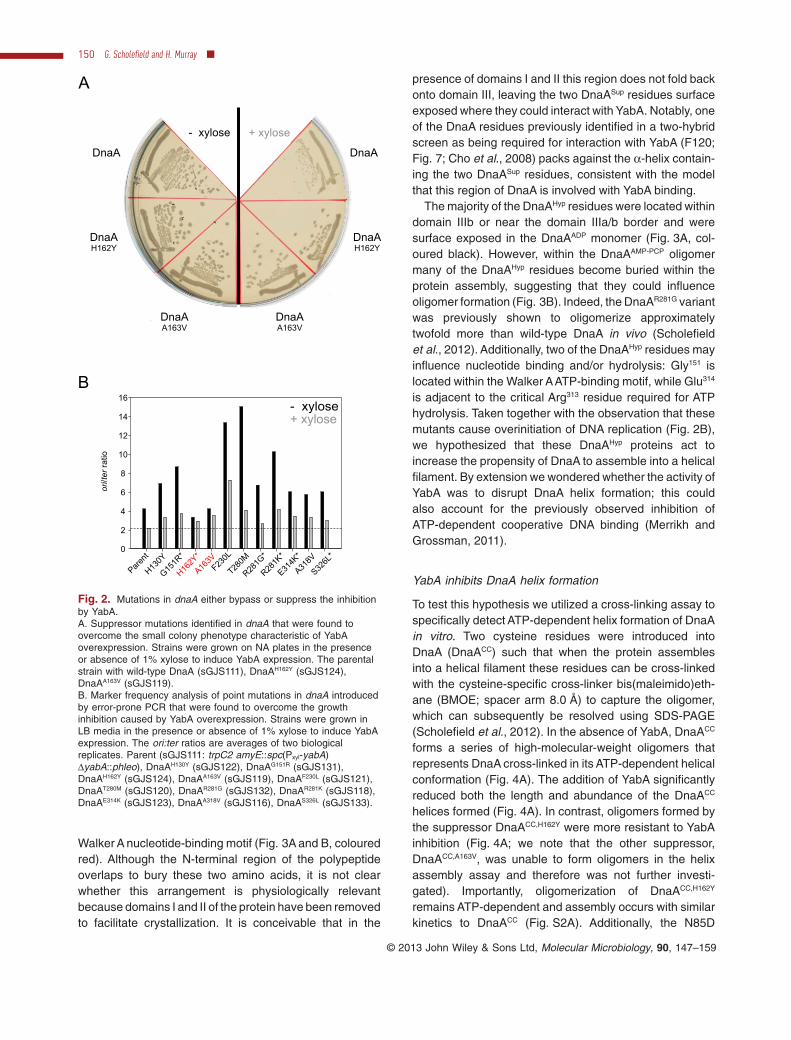
To try and understand how YabA was affecting DnaAactivity, we utilized a genetic approach to screen for dnaAmutants that could suppress the growth inhibition causedby overexpression of YabA (Fig. 2A and Fig. S1). A chlo-ramphenicol marker was integrated downstream of thednaAN operon and genomic DNAfrom this strain was usedas a substrate for error-prone PCR to generate pointmutations in dnaA. PCR products were transformed into astrain harbouring an inducible yabA allele and plated underYabA overexpression conditions to screen for large colo-nies. Genomic DNA from large colonies was backcrossedinto the parent strain to confirm that the mutation conferringfast growth was linked to dnaA. DNA sequencing identified11 distinct mutations that caused single amino acid substi-tutions within DnaA, all of which were located in domain III(the AAA+ motif).
To characterize the mutations in dnaA, marker frequencyanalysis (MFA) was used to measure the relative levels oforigin and terminus DNA, thereby generating a measure ofDNA replication initiation (Fig. 2B). The mutations withindnaA fell into two classes: hypermorphs that bypassedYabA inhibition (DnaAHyp proteins) by having a high basalrate of initiation, and suppressors that had an approxi-mately wild-type rate of initiation but were resistant toYabA inhibition (DnaASup proteins: DnaAH162Y, DnaAA163V;Fig. 2A). Strikingly, five of the identified residues werepreviously isolated in similar screens performed using thenegative regulator Soj (denoted by an asterisk in Fig. 2B)(Murray and Errington, 2008; Scholefield et al., 2012).
Next, the locations of these amino acids were mappedonto crystal structures of DnaA proteins. The two DnaASup
substitutions are located within the α-helix containing the
No Prot
einYab
A
DnaACC,R
313A
+Yab
A
DnaACC,R
313A
α-ATP
Density(AU)
6.2±0 .45
5.8±0 .48
DnaACC,R
313A
0
15
30
45
60
YabA
DnaACC
DnaACC +Y
abA
DnaACC +Y
abA*
A B
Mol
esof
Pi r
elea
sed/
Hou
r
Fig. 1. YabA does not affect the nucleotidebinding or hydrolysis activities of DnaA.A. YabA does not inhibit DnaA ATP binding.DnaACC,R313A (3 μM) and/or YabA (24 μM) wereincubated with [α-P32]-ATP before beingisolated from the reaction using magneticnickel beads and denatured with methanol.The released nucleotides were separated ona PEI cellulose TLC plate and visualized byphosphorimaging. Density values weremeasured using ImageJ (n = 3).B. Malachite green ATPase assay of DnaACC
and DnaACC,R313A in the presence and absenceof YabA. The asterisk (*) indicates theexpected ATP hydrolysis rate of a mixture ofDnaACC and YabA based on their respectiveindependent hydrolysis rates.
Walker A nucleotide-binding motif (Fig. 3A and B, colouredred). Although the N-terminal region of the polypeptideoverlaps to bury these two amino acids, it is not clearwhether this arrangement is physiologically relevantbecause domains I and II of the protein have been removedto facilitate crystallization. It is conceivable that in the
presence of domains I and II this region does not fold backonto domain III, leaving the two DnaASup residues surfaceexposed where they could interact with YabA. Notably, oneof the DnaA residues previously identified in a two-hybridscreen as being required for interaction with YabA (F120;Fig. 7; Cho et al., 2008) packs against the α-helix contain-ing the two DnaASup residues, consistent with the modelthat this region of DnaA is involved with YabA binding.
The majority of the DnaAHyp residues were located withindomain IIIb or near the domain IIIa/b border and weresurface exposed in the DnaAADP monomer (Fig. 3A, col-oured black). However, within the DnaAAMP-PCP oligomermany of the DnaAHyp residues become buried within theprotein assembly, suggesting that they could influenceoligomer formation (Fig. 3B). Indeed, the DnaAR281G variantwas previously shown to oligomerize approximatelytwofold more than wild-type DnaA in vivo (Scholefieldet al., 2012). Additionally, two of the DnaAHyp residues mayinfluence nucleotide binding and/or hydrolysis: Gly151 islocated within the Walker AATP-binding motif, while Glu314
is adjacent to the critical Arg313 residue required for ATPhydrolysis. Taken together with the observation that thesemutants cause overinitiation of DNA replication (Fig. 2B),we hypothesized that these DnaAHyp proteins act toincrease the propensity of DnaA to assemble into a helicalfilament. By extension we wondered whether the activity ofYabA was to disrupt DnaA helix formation; this couldalso account for the previously observed inhibition ofATP-dependent cooperative DNA binding (Merrikh andGrossman, 2011).
YabA inhibits DnaA helix formation
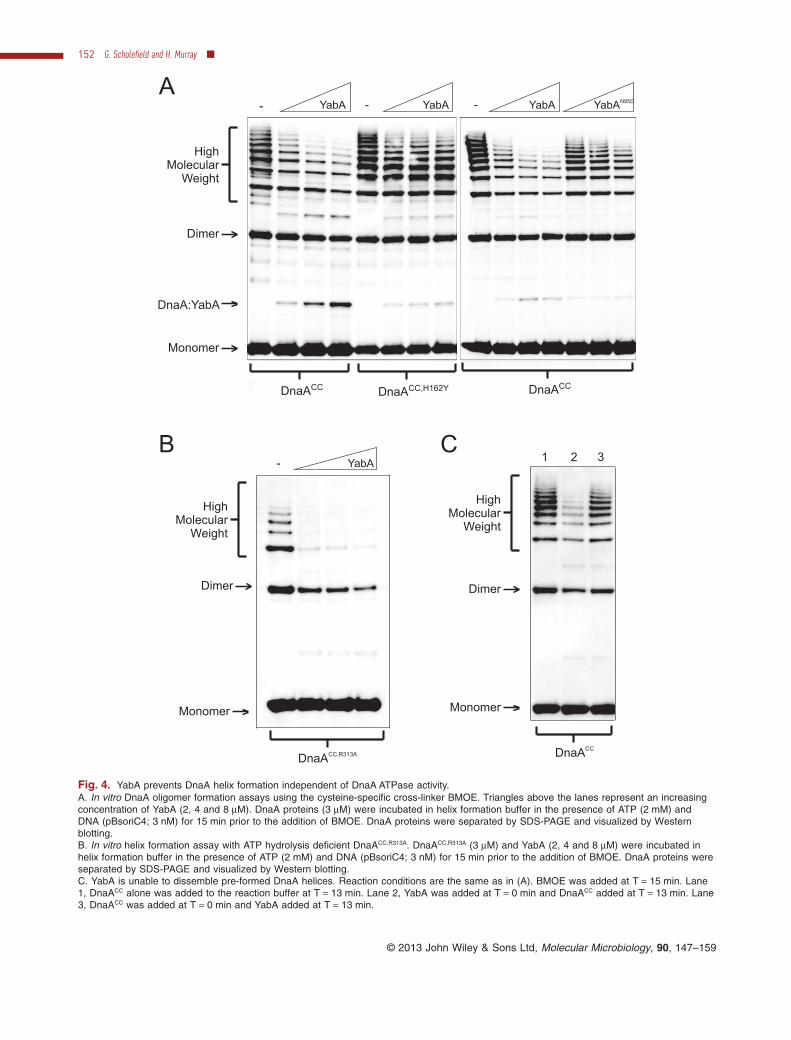
To test this hypothesis we utilized a cross-linking assay tospecifically detect ATP-dependent helix formation of DnaAin vitro. Two cysteine residues were introduced intoDnaA (DnaACC) such that when the protein assemblesinto a helical filament these residues can be cross-linkedwith the cysteine-specific cross-linker bis(maleimido)eth-ane (BMOE; spacer arm 8.0 Å) to capture the oligomer,which can subsequently be resolved using SDS-PAGE(Scholefield et al., 2012). In the absence of YabA, DnaACC
forms a series of high-molecular-weight oligomers thatrepresents DnaA cross-linked in its ATP-dependent helicalconformation (Fig. 4A). The addition of YabA significantlyreduced both the length and abundance of the DnaACC
helices formed (Fig. 4A). In contrast, oligomers formed bythe suppressor DnaACC,H162Y were more resistant to YabAinhibition (Fig. 4A; we note that the other suppressor,DnaACC,A163V, was unable to form oligomers in the helixassembly assay and therefore was not further investi-gated). Importantly, oligomerization of DnaACC,H162Y
remains ATP-dependent and assembly occurs with similarkinetics to DnaACC (Fig. S2A). Additionally, the N85D
Parent
0
2
4
6
8
ori/ter
ratio
- xylose+ xylose
10
12
14
16
H130Y
G151R
*
H162Y
*
A163V
F230L
T280M
R281G
*
R281K
*
E314K
*
A318V
S326L
*
B
A
- xylose + xylose
DnaA
DnaAH162Y
DnaAA163V
DnaA
DnaAH162Y
DnaAA163V
Fig. 2. Mutations in dnaA either bypass or suppress the inhibitionby YabA.A. Suppressor mutations identified in dnaA that were found toovercome the small colony phenotype characteristic of YabAoverexpression. Strains were grown on NA plates in the presenceor absence of 1% xylose to induce YabA expression. The parentalstrain with wild-type DnaA (sGJS111), DnaAH162Y (sGJS124),DnaAA163V (sGJS119).B. Marker frequency analysis of point mutations in dnaA introducedby error-prone PCR that were found to overcome the growthinhibition caused by YabA overexpression. Strains were grown inLB media in the presence or absence of 1% xylose to induce YabAexpression. The ori:ter ratios are averages of two biologicalreplicates. Parent (sGJS111: trpC2 amyE::spc(Pxyl-yabA)ΔyabA::phleo), DnaAH130Y (sGJS122), DnaAG151R (sGJS131),DnaAH162Y (sGJS124), DnaAA163V (sGJS119), DnaAF230L (sGJS121),DnaAT280M (sGJS120), DnaAR281G (sGJS132), DnaAR281K (sGJS118),DnaAE314K (sGJS123), DnaAA318V (sGJS116), DnaAS326L (sGJS133).
Fig. 3. The DnaA hypermorph and suppressor substitutions are located in domain III.A. A cartoon representation of monomeric DnaA from the Aquifex aeolicus crystal structure (PDB ID: 1L8Q) bound to ADP (stick) and Mg2+
(grey sphere). The hypermorph (black) and suppressor (red) residues are shown as space fill representations. The identity and positions ofthe B. subtilis amino acid substitutions are indicated above the corresponding residue of the A. aeolicus protein.B. A surface representation of the helical DnaA structure from A. aeolicus (PDB ID: 2HCB) bound to AMP-PCP. Individual DnaA monomersare coloured separately. The hypermorph (black) and suppressor (red) residues are indicated, and the positions of the B. subtilis amino acidresidues are indicated above the corresponding residue of the A. aeolicus protein.
Fig. 4. YabA prevents DnaA helix formation independent of DnaA ATPase activity.A. In vitro DnaA oligomer formation assays using the cysteine-specific cross-linker BMOE. Triangles above the lanes represent an increasingconcentration of YabA (2, 4 and 8 μM). DnaA proteins (3 μM) were incubated in helix formation buffer in the presence of ATP (2 mM) andDNA (pBsoriC4; 3 nM) for 15 min prior to the addition of BMOE. DnaA proteins were separated by SDS-PAGE and visualized by Westernblotting.B. In vitro helix formation assay with ATP hydrolysis deficient DnaACC,R313A. DnaACC,R313A (3 μM) and YabA (2, 4 and 8 μM) were incubated inhelix formation buffer in the presence of ATP (2 mM) and DNA (pBsoriC4; 3 nM) for 15 min prior to the addition of BMOE. DnaA proteins wereseparated by SDS-PAGE and visualized by Western blotting.C. YabA is unable to dissemble pre-formed DnaA helices. Reaction conditions are the same as in (A). BMOE was added at T = 15 min. Lane1, DnaACC alone was added to the reaction buffer at T = 13 min. Lane 2, YabA was added at T = 0 min and DnaACC added at T = 13 min. Lane3, DnaACC was added at T = 0 min and YabA added at T = 13 min.
substitution in YabA, which was previously shown todisrupt the interaction with DnaA, was less capable ofdisrupting oligomers formed by DnaACC (Fig. 4A;Noirot-Gros et al., 2006). We note that YabAN85D retainedthe ability to assemble into a tetramer in solution as judgedby gel filtration (data not shown), indicating that this proteinvariant is functional. Wild-type YabA was also found toinhibit helix formation of the ATP hydrolysis deficientDnaACC,R313A protein [Fig. 4B; note DnaACC,R313A requiresATP for oligomerization (Fig. S2B)], supporting the earlierobservation that YabA does not regulate the ATPase activ-ity of DnaA (Fig. 1B).
To determine whether YabA prevents DnaA helix for-mation and/or disassembles pre-formed DnaA helices, anorder of addition experiment was performed. Figure 4Cshows that DnaACC helices formed prior to YabA additionare highly resistant to YabA activity (lane 3) when com-pared with helices formed after YabA addition (lane 2),indicating that YabA acts by preventing DnaA helixformation.
Interaction of YabA with DnaA
The DnaAH162Y and DnaAA163V proteins were found to beresistant to YabA activity in vivo, and helix formation of theDnaACC,H162Y protein was less sensitive to YabA in vitro. Asnoted above, the α-helix containing these two adjacentresidues packs against a short α-helix containing residuePhe120 (Fig. 3A), which when changed to serine was pre-viously shown to disrupt the interaction between DnaAand YabA in a yeast two-hybrid assay (Cho et al., 2008).Based on these observations we investigated whetherDnaACC,H162Y could interact with YabA in vitro.
First, DnaA proteins were subjected to primary amine-specific cross-linking (BS3) with and without YabA underthe same experimental conditions used for the helix forma-tion assay (Fig. 5A). The appearance of two higher-ordercomplexes was observed in the presence of YabA, one ata molecular mass expected for a YabA:DnaA dimer(14 kDa + 54 kDa = 68 kDa) and another at a molecularmass expected for a YabA2:DnaA trimer (14 kDa + 14 kDa+ 54 kDa = 82 kDa). The formation of both heteromericcomplexes was decreased when DnaACC,H162Y was used inthis assay, and conversely formation of the homomericDnaACC,H162Y dimer was less reduced by YabA comparedwith DnaACC (Fig. 5A and B).
Second, surface plasmon resonance (SPR) was usedto test the interaction between DnaA and YabA. The DnaAproteins were immobilized on the sensor chip surfaceusing an amine coupling approach and YabA was theninjected over DnaACC and DnaACC,H162Y surfaces. In con-trast to the BS3 cross-linking assay, YabA was found tointeract with both DnaA proteins similarly using SPR(Fig. 5C). We do not currently understand the reason
underlying this apparent difference, but we note that thetwo interaction assays are performed under distinctexperimental conditions (see Discussion).
DnaD inhibits DnaA helix formation
During the course of this study it was reported that theDNA replication initiation protein DnaD inhibits the coop-erative binding of DnaA to DNA without affecting ATPbinding or hydrolysis activities (Bonilla and Grossman,2012). The similarity of the regulatory properties of YabAand DnaD suggested to us that DnaD might also regulateDnaA helix formation. To address this hypothesis DnaDwas purified and analysed using the DnaA helix formationassay in vitro. DnaD was found to inhibit the formation ofDnaACC helices to a greater extent than either the high-salt DnaD storage buffer or a control protein (BSA), indi-cating that this inhibition is likely specific (Fig. 6A). Underthe reaction conditions used in the helix formation assayDnaD did not interact stably with plasmid DNA as judgedby an electrophoretic mobility shift assay, suggesting thatit does not act by competing with DnaA for DNA (Fig. S3).Furthermore, an order of addition experiment showed thatpre-formed DnaACC helices are highly resistant to DnaDactivity (Fig. 6B), indicating that DnaD acts by preventingthe assembly of the DnaA oligomer.
A previous study using yeast two-hybrid analysis foundthat amino acid substitutions in DnaA that inhibited theinteraction with YabA also affected the binding of DnaD(Cho et al., 2008). Since it appears that DnaD inhibitsDnaA helix formation similarly to YabA, we determinedwhether the DnaACC,H162Y mutant would also suppressthe effect of DnaD. Interestingly, oligomers formed byDnaACC,H162Y were partially resistant to DnaD inhibition(Fig. 6C). This result suggests that the regulatory activitiesof DnaD and YabA on DnaA may overlap.
Discussion
YabA and DnaD inhibit DnaA helix formation
Here we report that the regulatory proteins YabAand DnaDboth inhibit the ability of the DNAreplication initiator proteinDnaA to assemble into a helical oligomer. These findingsare consistent with previous results indicating that YabAand DnaD inhibit cooperative binding of DnaA to DNA (i.e.the functional readout of DnaA helix formation) (Merrikhand Grossman, 2011; Bonilla and Grossman, 2012). Com-bined with our previous work where we identified Soj/ParAas a regulator of DnaA helix formation (Scholefield et al.,2012), it appears that all of the known vegetativelyexpressed DnaAregulatory proteins in B. subtilis target thesame step of the DnaA initiation pathway. These resultssuggest that regulation of DnaAactivity in B. subtilis occurs
at a different stage compared with regulation of DnaA inE. coli where nucleotide hydrolysis and exchange are thepredominant targets (Katayama et al., 2010).
One reason for this variation may be the stability of theDnaA:ADP complexes in these distinct organisms. It hasbeen demonstrated that under identical experimental con-ditions the half-life of the E. coli DnaA:ADP complex is∼ 40 min, much longer than the complexes formed byeither the B. subtilis or Staphlococcus aureus DnaA pro-teins (5 or 2 min respectively; Kurokawa et al., 2009). If it isassumed that these in vitro assays faithfully represent thestability of different DnaA:ADP complexes in vivo and thatno other factors are involved, then it follows that themajority of DnaA proteins in organisms such as B. subtilisand S. aureus would be bound to the more abundant
nucleotide ATP and therefore would be competent toassemble into active initiation complexes [note that DnaAproteins from B. subtilis and E. coli have equally highaffinities for ATP; Kd = 20 nM and 30 nM respectively(Sekimizu et al., 1987; Fukuoka et al., 1990)]. In thissituation it would be necessary to regulate a step of theDnaA activity pathway downstream of nucleotide binding,and indeed targeting DnaA oligomerization would fulfilthis requirement. Interestingly, S. aureus contains geneshomologous to yabA, dnaD and soj, suggesting that DnaAhelix formation could likewise be directly regulated in thismedically pertinent organism. It will be highly informative todetermine how far the correlation between DnaA:ADPcomplex stability and regulation of DnaA helix formationextends through the bacterial kingdom.
80
40
120
RU
DnaACC
K = 0.68 ± 0.19 MD μ
0
RU
DnaACC,H162Y
K = 0.38 ± 0.11 MD μ
DnaA
DnaA2
20
YabA2:DnaAYabA:DnaA
DnaACC DnaACC,H162Y
DnaACC
DnaACC,H162Y
YabA:D
naA
40
60
80
100
0%of
inte
nsity
ofYa
bA:W
TD
naA
inte
ract
ion
band
0 200 400 600 800 1000Time (sec)
YabA 2:D
naA
A B
C
YabA
0 200 400 600 800 1000Time (sec)
020
60
100
- -+ +
80
40
120
20
60
100
Fig. 5. Interaction of YabA with DnaA.A. In vitro cross-linking assay using the primary amine-specific cross-linker BS3 (20 μM) in the presence of DNA (pBSoriC4; 3 nM) and ATP(2 mM). Protein complexes were separated by SDS-PAGE and DnaACC proteins were visualized by Western blotting. The identity of the DnaAproteins (3 μM) are indicated below and the presence of YabA (8 μM) is indicated above.B. Quantification of the YabA:DnaA interaction complexes from (A).C. Interaction of YabA with immobilized DnaACC proteins using surface plasmon resonance (SPR). DnaA proteins were bound to the SPR chipsurface via a standard amine coupling approach. A twofold serial dilution of YabA starting at 15 μM was injected over wild-type DnaACC andDnaACC,H162Y proteins.
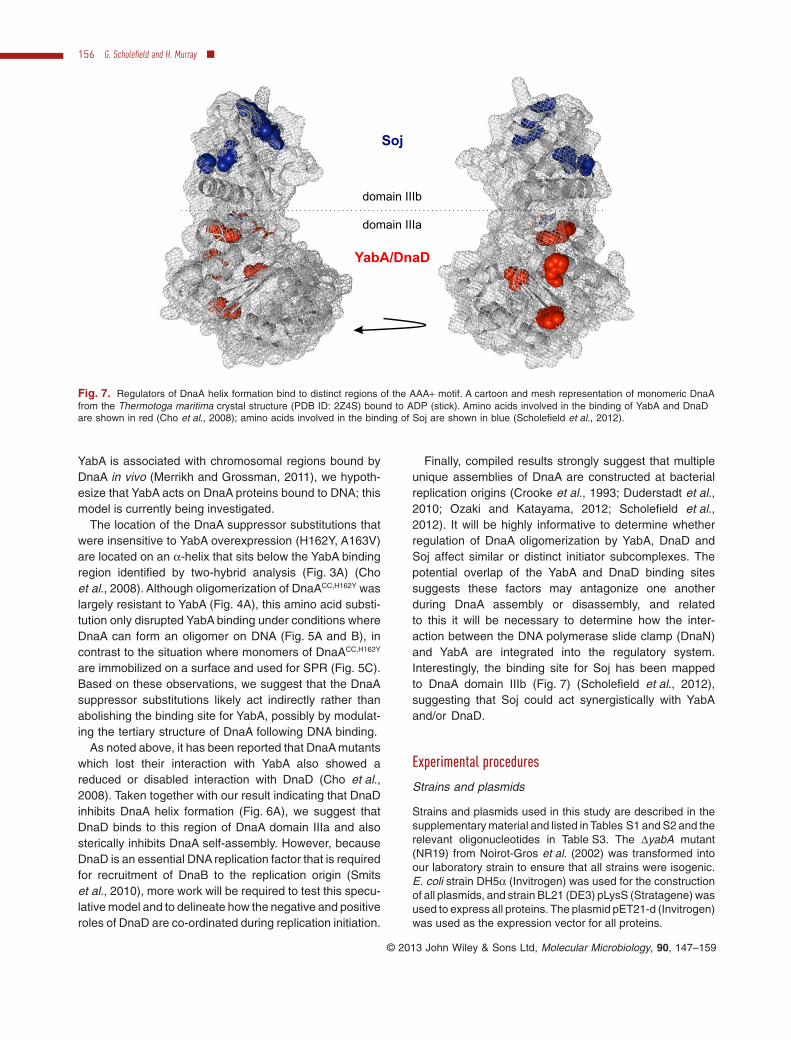
Yeast two-hybrid analysis strongly suggests that YabAinteracts with DnaA domain IIIa (Fig. 7) (Cho et al., 2008).A region encompassing two of the DnaA residues impli-
cated in the interaction with YabA (Tyr144, Thr271)appears to be buried within the DnaA:DnaA interface of thehelical oligomer. Therefore, the simplest model for regula-tion by YabA is that it binds to this region of DnaA domainIIIa and sterically inhibits DnaA self-assembly. Because
Monomer
Dimer
HighMolecular
Weight
-
DnaACC
bufferDnaDA B
BSA
Monomer
Dimer
HighMolecular
Weight
DnaACC
21 3
DnaACC DnaACC,H162Y
Monomer
Dimer
HighMolecular
Weight
- -DnaD DnaDC
Fig. 6. DnaD inhibits DnaA helix formation.A. In vitro DnaA oligomer formation assay using the cysteine-specific cross-linker BMOE. Triangles above the lanes represent an increasingconcentration of DnaD or BSA (8, 16 and 24 μM). DnaACC proteins (3 μM) were incubated in helix formation buffer in the presence of ATP(2 mM) and DNA (pBsoriC4; 3 nM) for 15 min prior to the addition of BMOE. DnaACC proteins were separated by SDS-PAGE and visualized byWestern blotting.B. DnaD is unable to dissemble pre-formed DnaA helices. Reaction conditions are the same as in (A). BMOE was added at T = 15 min. Lane1, DnaACC alone was added to the reaction buffer at T = 13 min. Lane 2, DnaD was added at T = 0 min and DnaACC added at T = 13 min.Lane 3, DnaACC was added at T = 0 min and DnaD added at T = 13 min.C. DnaACC,H162Y partially suppresses DnaD inhibition. DnaACC proteins (3 μM) and DnaD (8 and 16 μM) were incubated in helix formation bufferin the presence of ATP (2 mM) and DNA (pBsoriC4; 3 nM) for 15 min prior to the addition of BMOE. DnaA proteins were separated bySDS-PAGE and visualized by Western blotting.
YabA is associated with chromosomal regions bound byDnaA in vivo (Merrikh and Grossman, 2011), we hypoth-esize that YabA acts on DnaA proteins bound to DNA; thismodel is currently being investigated.
The location of the DnaA suppressor substitutions thatwere insensitive to YabA overexpression (H162Y, A163V)are located on an α-helix that sits below the YabA bindingregion identified by two-hybrid analysis (Fig. 3A) (Choet al., 2008). Although oligomerization of DnaACC,H162Y waslargely resistant to YabA (Fig. 4A), this amino acid substi-tution only disrupted YabA binding under conditions whereDnaA can form an oligomer on DNA (Fig. 5A and B), incontrast to the situation where monomers of DnaACC,H162Y
are immobilized on a surface and used for SPR (Fig. 5C).Based on these observations, we suggest that the DnaAsuppressor substitutions likely act indirectly rather thanabolishing the binding site for YabA, possibly by modulat-ing the tertiary structure of DnaA following DNA binding.
As noted above, it has been reported that DnaA mutantswhich lost their interaction with YabA also showed areduced or disabled interaction with DnaD (Cho et al.,2008). Taken together with our result indicating that DnaDinhibits DnaA helix formation (Fig. 6A), we suggest thatDnaD binds to this region of DnaA domain IIIa and alsosterically inhibits DnaA self-assembly. However, becauseDnaD is an essential DNA replication factor that is requiredfor recruitment of DnaB to the replication origin (Smitset al., 2010), more work will be required to test this specu-lative model and to delineate how the negative and positiveroles of DnaD are co-ordinated during replication initiation.
Finally, compiled results strongly suggest that multipleunique assemblies of DnaA are constructed at bacterialreplication origins (Crooke et al., 1993; Duderstadt et al.,2010; Ozaki and Katayama, 2012; Scholefield et al.,2012). It will be highly informative to determine whetherregulation of DnaA oligomerization by YabA, DnaD andSoj affect similar or distinct initiator subcomplexes. Thepotential overlap of the YabA and DnaD binding sitessuggests these factors may antagonize one anotherduring DnaA assembly or disassembly, and relatedto this it will be necessary to determine how the inter-action between the DNA polymerase slide clamp (DnaN)and YabA are integrated into the regulatory system.Interestingly, the binding site for Soj has been mappedto DnaA domain IIIb (Fig. 7) (Scholefield et al., 2012),suggesting that Soj could act synergistically with YabAand/or DnaD.
Experimental procedures
Strains and plasmids
Strains and plasmids used in this study are described in thesupplementary material and listed in Tables S1 and S2 and therelevant oligonucleotides in Table S3. The ΔyabA mutant(NR19) from Noirot-Gros et al. (2002) was transformed intoour laboratory strain to ensure that all strains were isogenic.E. coli strain DH5α (Invitrogen) was used for the constructionof all plasmids, and strain BL21 (DE3) pLysS (Stratagene) wasused to express all proteins. The plasmid pET21-d (Invitrogen)was used as the expression vector for all proteins.
YabA/DnaD
Soj
domain IIIb
domain IIIa
Fig. 7. Regulators of DnaA helix formation bind to distinct regions of the AAA+ motif. A cartoon and mesh representation of monomeric DnaAfrom the Thermotoga maritima crystal structure (PDB ID: 2Z4S) bound to ADP (stick). Amino acids involved in the binding of YabA and DnaDare shown in red (Cho et al., 2008); amino acids involved in the binding of Soj are shown in blue (Scholefield et al., 2012).
Nutrient agar (NA; Oxoid) was used for routine selection andmaintenance of both B. subtilis and E. coli strains. For experi-ments in B. subtilis cells were routinely grown in either Luria–Bertani (LB) medium or casein hydrolysate (CH) medium.Supplements were added as required: 20 μg ml−1 tryptophan,5 μg ml−1 chloramphenicol, 2 μg ml−1 kanamycin, 50 μg ml−1
spectinomycin. For plasmid and protein expression in E. colicells were grown in LB medium or Nutrient Broth (Oxoid) andsupplemented with 30 μg ml−1 (for low-copy-number plas-mids) or 75 μg ml−1 ampicillin and 10 μg ml−1 chlorampheni-col. Unless otherwise stated all chemicals and reagents wereobtained from Sigma-Aldrich.
ATP binding assay
Samples were prepared and treated in an identical manner tothe oligomer formation assay up until the addition of BMOE. Atthis point 5 μl of magnetic nickel beads (Promega) were addedand incubated at room temperature for 2 min. Proteins werethen collected using a magnet and washed twice with 1 ml ofhelix formation buffer supplemented with imidazole (10 mM).After the final wash the beads were resuspended in 10 μlmethanol to denature the proteins and release the boundnucleotide. Denatured proteins collected using a magnet and1 μl of the supernatant was then spotted onto PEI cellulosepaper (Merck) and subjected to thin-layer chromatography(TLC) using a solvent composed of 5% formic acid and 0.5 MLiCl. TLC plates were exposed onto phosphoimager screensand nucleotide was detected using a Typhoon imager.
ATPase assay
The malachite green assay solution was created by mixing0.0812% (w/v) malachite green, 2.23% (w/v) polyvinylalcohol, 5.72% (w/v) ammonium molybdate in HCl (6 M) andH2O in a ratio of 2:1:1:2. The solution was incubated for 2 hwith gentle agitation (during this time the solution turned froma muddy brown colour to a golden yellow). Reactions withand without YabA (24 μM) were diluted into oligomer forma-tion buffer supplemented with ATP (5 mM), pBSoriC4 (3 nM).DnaA (3 μM) was then added and the reaction incubated fora total of 6 h. Ten microlitres of samples were taken every5 min for 20 min (including time zero) and mixed with 160 μlof malachite green assay solution, followed by the addition of20 μl sodium citrate (34%). The colour was allowed to stabi-lize for 10 min before the absorbance was detected at620 nm. A standard curve was created using a serial dilutionof sodium phosphate (3–500 mM). The A620 was convertedinto mole Pi produced using a phosphate standard.
Screen for dnaA mutants that suppress growthinhibition of YabA
Error-prone PCR was performed using Phire DNA polymer-ase (Finnzymes) on chromosomal DNA from strain HM387with primers oHM56 and oQPCR33 (Scholefield et al., 2012).Twenty 50 μl reactions were independently run to increasethe diversity of mutations. Reactions were denatured at 98°Cfor 30 s, followed by 20 cycles of 98°C for 30 s, 60°C for 30 s
and 72°C for 8 min. The reactions were purified using a PCRPurification Kit (Qiagen) and pooled. Mutagenized DNA wastransformed into sGJS111 and plated onto nutrient agarplates at 37°C in the presence of chloramphenicol and 1%xylose (to induce expression of yabA). Large colonies wereselected (100 from a starting pool of ∼ 20 000) and streakedonto nutrient agar plates at 37°C in the presence or absenceof 1% xylose. Chromosomal DNA from 11 candidates thatgrew similarly in the presence and absence of xylose wasisolated and backcrossed into sGJS111 to confirm that thesuppressor mutation was linked to the chloramphenicolmarker. The oriC region of these suppressors was amplifiedusing primers oHM254 and oHM255, followed by sequencingto identify the mutations.
Marker frequency analysis
Starter cultures were grown in liquid medium at 30°C overnightand then diluted 1:100 into fresh medium (with inducer whereindicated). Sodium azide (0.5%; Sigma) was added to expo-nentially growing cells (A600 = 0.2–0.4) to prevent furthermetabolism. Chromosomal DNA was isolated using a DNeasyBlood and Tissue Kit (Qiagen). Rotor-Gene SYBR Green PCRKit was used for PCR reactions (Qiagen). Q-PCR was per-formed in a Rotor-Gene Q Instrument (Qiagen). By use ofcrossing points (CT) and PCR efficiency a relative quantifica-tion analysis (ΔΔCT) was performed using Rotor-Gene Soft-ware version 2.0.2 (Qiagen) to determine the ori/ter ratio ofeach sample. The results were normalized to the ori/ter ratio ofa DNA sample from B. subtilis spores which only contain onechromosome and thus have an ori/ter ratio of 1.
Purification of YabA-His6
Escherichia coli BL21 (DE3) pLysS (Stratagene) containingthe appropriate yabA-his6 plasmid (see Table S2) was inocu-lated into LB containing ampicillin and chloramphenicol andgrown overnight at 37°C. The culture was diluted 1:100 into LBcontaining ampicillin and incubated at 37°C until the A600
reached ∼ 0.6. Cultures were then supplemented with 1 mMIPTG and shifted to 30°C for 3 h to induce protein expression.Cells were collected by centrifugation at 5000 g (4°C) for10 min and then resuspended in YabA purification buffer(50 mM HEPES pH 7.6, 300 mM NaCl and 40 mM imidazole).One Complete EDTA-free protease inhibitor tablet (Roche)and lysozyme (32 mg ml−1) were added and cells were incu-bated on ice for 1 h with gentle agitation. Cells were lysedusing a Vibra-Cell sonicator (Sonics) on ice for approximately5 min (power setting 80; 4 s pulse, 1 s rest). Cell debris wasremoved by centrifugation at 31 000 g (4°C) for 45 min. Thesupernatant was applied to a 1 ml HisTrap FF column (GEHealthcare), washed with 20 ml YabA purification buffer andthen eluted with a linear gradient of YabA purification buffersupplemented with 1 M imidazole. Fractions were analysed bySDS-PAGE and relevant fractions were pooled and applied toa HiLoad Superdex 75 gel filtration column pre-equilibratedwith YabA gel filtration buffer (30 mM HEPES pH 7.6, 250 mMNaCl, 1 mM DTT and 20% sucrose) and run overnight at0.3 ml min−1. Fractions were then analysed by SDS-PAGE,relevant fractions pooled, frozen in liquid nitrogen and storedat −80°C.
Escherichia coli BL21 (DE3) pLysS containing the dnaD-his6
plasmid was inoculated, grown and induced as describedabove for YabA-His6. Cells were collected by centrifugation at5000 g (4°C) for 10 min and then resuspended in DnaD puri-fication buffer (50 mM HEPES pH 7.6, 500 mM NaCl, 30 mMimidazole, 0.1 mM EDTA and 20% sucrose) such that theywere concentrated 25-fold. Cells were lysed as describedabove for YabA-His6. The supernatant was diluted twofoldwith DnaD dilution buffer (50 mM HEPES pH 7.6, 30 mMimidazole, 0.1 mM EDTA and 20% sucrose), applied to a 1 mlHisTrap FF column (GE Healthcare), washed with 20 mlDnaD purification buffer (also diluted twofold with DnaD dilu-tion buffer) and then eluted with a linear gradient of DnaDelution buffer (50 mM HEPES pH 7.6, 250 mM NaCl, 500 mMimidazole, 0.1 mM EDTA and 20% sucrose). Fractions wereanalysed by SDS-PAGE, pooled and dialysed in DnaD dialy-sis buffer (50 mM HEPES pH 7.6, 500 mM NaCl, 0.5 mMEDTA, 1 mM DTT and 20% sucrose). Dialysed protein wasfrozen in liquid nitrogen and stored at −80°C.
In vitro helix formation assay
This assay was performed as previously described(Scholefield et al., 2012).
Surface plasmon resonance
Experiments were conducted on the Proteon XPR36 systemusing a GLC sensor chip. Protein solutions were preparedat room temperature and held within the Proteon at 4°C. Theinteraction analysis was performed at 25°C. The system wasprimed with chip preparation buffer (25 mM HEPES pH 7.6,200 mM NaCl, 10 mM MgCl2, 1 mM EDTA and 0.005% P20)and the chip surface was pre-prepared with concurrent 60 sinjections of 0.5% SDS, 50 mM NaOH and 100 mM HCl. Thesurface was activated with a 210 s injection of a 50:50 mix of0.2 M EDC and 50 mM NHS (Bio-Rad). Different DnaAligandswere diluted into binding buffer (25 mM sodium acetatepH 4.5, 500 mM NaCl and 0.005% P20) and injected over fivechannels until ∼ 1000 RU had been immobilized. BSA wasdiluted in the same buffer and immobilized to ∼ 1000 RU usingthe remaining channel to act as a reference. Unreacted esterswere then blocked with a 300 s injection of 1 M ethanolamine.The system was then primed with helix formation buffer(25 mM HEPES pH 7.6, 200 mM NaCl, 100 mM potassiumglutamate and 10 mM MgCl2) supplemented with 5 mM ATPand 0.005% P20. YabA was exchanged (PD-10 column, GEHealthcare) into helix formation buffer and then systemicallyinjected over the protein surfaces for a contact time of 300 s.The surface was regenerated between each injection with an8 s injection of regeneration buffer (50 mM NaOH, 200 mMNaCl, 1 mM EDTA and 0.005% P20). The data reported is thedifference between the response observed in the channelcontaining the immobilized DnaA ligand and the responseobserved in the channel containing BSA.
Primary amine cross-linking assay
This assay was performed as previously described(Scholefield et al., 2012).
DnaD electrophoretic mobility shift assay (EMSA)
DnaD was serially diluted into helix formation buffer andpBsoriC4 plasmid DNA (120 fmol) was then added. The reac-tion was left to proceed for 15 min at 37°C and nucleoproteincomplexes were then separated on a 1% agarose gel (boththe running buffer and the gel contained 0.5× Tris-borate). Tovisualize the DNA the gel was post-stained in running buffercontaining ethidium bromide (1 μg ml−1) for 30 min and thende-stained for 10 min in water before being imaged on aTyphoon 8600 fluoroimager (GE Healthcare).
Acknowledgements
We thank J. Errington, A. Koh and N. Rostami for usefuldiscussions. We also thank the anonymous reviewers forstimulating comments and suggestions. This work was sup-ported by a BBSRC studentship to G.S. and by a RoyalSociety University Research Fellowship to H.M.
References
Abe, Y., Jo, T., Matsuda, Y., Matsunaga, C., Katayama, T.,and Ueda, T. (2007) Structure and function of DnaAN-terminal domains: specific sites and mechanisms ininter-DnaA interaction and in DnaB helicase loading onoriC. J Biol Chem 282: 17816–17827.
Bonilla, C.Y., and Grossman, A.D. (2012) The primosomalprotein DnaD inhibits cooperative DNA binding by the rep-lication initiator DnaA in Bacillus subtilis. J Bacteriol 194:5110–5117.
Bramhill, D., and Kornberg, A. (1988) Duplex opening by dnaAprotein at novel sequences in initiation of replication at theorigin of the E. coli chromosome. Cell 52: 743–755.
Briggs, G.S., Smits, W.K., and Soultanas, P. (2012) Chromo-somal replication initiation machinery of low-G+C-contentFirmicutes. J Bacteriol 194: 5162–5170.
Cho, E., Ogasawara, N., and Ishikawa, S. (2008) The func-tional analysis of YabA, which interacts with DnaA andregulates initiation of chromosome replication in Bacillussubtilis. Genes Genet Syst 83: 111–125.
Crooke, E., Thresher, R., Hwang, D.S., Griffith, J., andKornberg, A. (1993) Replicatively active complexes of DnaAprotein and the Escherichia coli chromosomal originobserved in the electron microscope. J Mol Biol 233: 16–24.
Duderstadt, K.E., Mott, M.L., Crisona, N.J., Chuang, K.,Yang, H., and Berger, J.M. (2010) Origin remodeling andopening in bacteria rely on distinct assembly states of theDnaA initiator. J Biol Chem 285: 28229–28239.
Duderstadt, K.E., Chuang, K., and Berger, J.M. (2011) DNAstretching by bacterial initiators promotes replication originopening. Nature 478: 209–213.
Erzberger, J.P., Pirruccello, M.M., and Berger, J.M. (2002)The structure of bacterial DnaA: implications for generalmechanisms underlying DNA replication initiation. EMBO J21: 4763–4773.
Erzberger, J.P., Mott, M.L., and Berger, J.M. (2006) Structuralbasis for ATP-dependent DnaA assembly and replication-origin remodeling. Nat Struct Mol Biol 13: 676–683.
Felczak, M.M., Simmons, L.A., and Kaguni, J.M. (2005) Anessential tryptophan of Escherichia coli DnaA protein func-
tions in oligomerization at the E. coli replication origin. JBiol Chem 280: 24627–24633.
Fujikawa, N., Kurumizaka, H., Nureki, O., Terada, T.,Shirouzu, M., Katayama, T., and Yokoyama, S. (2003)Structural basis of replication origin recognition by theDnaA protein. Nucleic Acids Res 31: 2077–2086.
Fukuoka, T., Moriya, S., Yoshikawa, H., and Ogasawara, N.(1990) Purification and characterization of an initiationprotein for chromosomal replication, DnaA, in Bacillus sub-tilis. J Biochem (Tokyo) 107: 732–739.
Goranov, A.I., Breier, A.M., Merrikh, H., and Grossman, A.D.(2009) YabA of Bacillus subtilis controls DnaA-mediatedreplication initiation but not the transcriptional response toreplication stress. Mol Microbiol 74: 454–466.
Hayashi, M., Ogura, Y., Harry, E.J., Ogasawara, N., andMoriya, S. (2005) Bacillus subtilis YabA is involved in deter-mining the timing and synchrony of replication initiation.FEMS Microbiol Lett 247: 73–79.
Ishigo-Oka, D., Ogasawara, N., and Moriya, S. (2001) DnaDprotein of Bacillus subtilis interacts with DnaA, the initiatorprotein of replication. J Bacteriol 183: 2148–2150.
Iyer, L.M., Leipe, D.D., Koonin, E.V., and Aravind, L. (2004)Evolutionary history and higher order classification of AAA+ATPases. J Struct Biol 146: 11–31.
Kasho, K., and Katayama, T. (2013) DnaA binding locus datApromotes DnaA-ATP hydrolysis to enable cell cycle-coordinated replication initiation. Proc Natl Acad Sci USA110: 936–941.
Katayama, T., Ozaki, S., Keyamura, K., and Fujimitsu, K.(2010) Regulation of the replication cycle: conserved anddiverse regulatory systems for DnaA and oriC. Nat RevMicrobiol 8: 163–170.
Kawakami, H., and Katayama, T. (2010) DnaA, ORC, andCdc6: similarity beyond the domains of life and diversity.Biochem Cell Biol 88: 49–62.
Kurokawa, K., Mizumura, H., Takaki, T., Ishii, Y., Ichihashi,N., Lee, B.L., and Sekimizu, K. (2009) Rapid exchange ofbound ADP on the Staphylococcus aureus replicationinitiation protein DnaA. J Biol Chem 284: 34201–34210.
Merrikh, H., and Grossman, A.D. (2011) Control of the repli-cation initiator DnaA by an anti-cooperativity factor. MolMicrobiol 82: 434–446.
Miller, D.T., Grimwade, J.E., Betteridge, T., Rozgaja, T.,Torgue, J.J., and Leonard, A.C. (2009) Bacterial originrecognition complexes direct assembly of higher-orderDnaA oligomeric structures. Proc Natl Acad Sci USA 106:18479–18484.
Molt, K.L., Sutera, V.A., Jr, Moore, K.K., and Lovett, S.T.(2009) A role for nonessential domain II of initiator protein,DnaA, in replication control. Genetics 183: 39–49.
Mott, M.L., and Berger, J.M. (2007) DNA replication initiation:mechanisms and regulation in bacteria. Nat Rev Microbiol5: 343–354.
Murray, H., and Errington, J. (2008) Dynamic control of theDNA replication initiation protein DnaA by Soj/ParA. Cell135: 74–84.
Neuwald, A.F., Aravind, L., Spouge, J.L., and Koonin, E.V.(1999) AAA+: a class of chaperone-like ATPases associ-ated with the assembly, operation, and disassembly ofprotein complexes. Genome Res 9: 27–43.
Nishida, S., Fujimitsu, K., Sekimizu, K., Ohmura, T., Ueda, T.,
and Katayama, T. (2002) A nucleotide switch in theEscherichia coli DnaA protein initiates chromosomal repli-cation: evidence from a mutant DnaA protein defective inregulatory ATP hydrolysis in vitro and in vivo. J Biol Chem277: 14986–14995.
Noirot-Gros, M.F., Dervyn, E., Wu, L.J., Mervelet, P.,Errington, J., Ehrlich, S.D., and Noirot, P. (2002) Anexpanded view of bacterial DNA replication. Proc Natl AcadSci USA 99: 8342–8347.
Noirot-Gros, M.F., Velten, M., Yoshimura, M., McGovern, S.,Morimoto, T., Ehrlich, S.D., et al. (2006) Functional dissec-tion of YabA, a negative regulator of DNA replication initia-tion in Bacillus subtilis. Proc Natl Acad Sci USA 103: 2368–2373.
Okumura, H., Yoshimura, M., Ueki, M., Oshima, T.,Ogasawara, N., and Ishikawa, S. (2012) Regulation ofchromosomal replication initiation by oriC-proximal DnaA-box clusters in Bacillus subtilis. Nucleic Acids Res 40:220–234.
Ozaki, S., and Katayama, T. (2012) Highly organized DnaA-oriC complexes recruit the single-stranded DNA for repli-cation initiation. Nucleic Acids Res 40: 1648–1665.
Ozaki, S., Kawakami, H., Nakamura, K., Fujikawa, N.,Kagawa, W., Park, S.Y., et al. (2008) Acommon mechanismfor the ATP-DnaA-dependent formation of open complexesat the replication origin. J Biol Chem 283: 8351–8362.
Samitt, C.E., Hansen, F.G., Miller, J.F., and Schaechter, M.(1989) In vivo studies of DnaA binding to the origin ofreplication of Escherichia coli. EMBO J 8: 989–993.
Scholefield, G., Errington, J., and Murray, H. (2012) Soj/ParAstalls DNA replication by inhibiting helix formation of theinitiator protein DnaA. EMBO J 31: 1542–1555.
Sekimizu, K., Bramhill, D., and Kornberg, A. (1987) ATPactivates dnaA protein in initiating replication of plasmidsbearing the origin of the E. coli chromosome. Cell 50:259–265.
Smits, W.K., Goranov, A.I., and Grossman, A.D. (2010)Ordered association of helicase loader proteins with theBacillus subtilis origin of replication in vivo. Mol Microbiol75: 452–461.
Soufo, C.D., Soufo, H.J., Noirot-Gros, M.F., Steindorf, A.,Noirot, P., and Graumann, P.L. (2008) Cell-cycle-dependent spatial sequestration of the DnaA replicationinitiator protein in Bacillus subtilis. Dev Cell 15: 935–941.
Speck, C., and Messer, W. (2001) Mechanism of originunwinding: sequential binding of DnaA to double- andsingle-stranded DNA. EMBO J 20: 1469–1476.
Tucker, P.A., and Sallai, L. (2007) The AAA+ superfamily – amyriad of motions. Curr Opin Struct Biol 17: 641–652.
Weigel, C., Schmidt, A., Seitz, H., Tungler, D., Welzeck, M.,and Messer, W. (1999) The N-terminus promotes oli-gomerization of the Escherichia coli initiator protein DnaA.Mol Microbiol 34: 53–66.



















![Antibodies to PhnD Inhibit Staphylococcal to PhnD Inhibit Staphylococcal Biofilms ... phils]) were purified from ... Antibodies to PhnD Inhibit Staphylococcal Biofilms](https://static.documents.pub/doc/80x56/5ac950907f8b9aa1298cfe6f/antibodies-to-phnd-inhibit-staphylococcal-to-phnd-inhibit-staphylococcal-biolms.jpg)







