Available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/watres Zebra mussel filtration and its potential uses in industrial water treatment Paul Elliott a, , David C. Aldridge a , Geoff D. Moggridge b a Department of Zoology, University of Cambridge, Downing Street, Cambridge CB2 3EJ, UK b Department of Chemical Engineering, University of Cambridge, Pembroke Street, Cambridge CB2 3RA, UK article info Article history: Received 22 July 2007 Received in revised form 15 October 2007 Accepted 16 October 2007 Available online 18 October 2007 Keywords: Dreissena polymorpha Zebra mussel Water treatment works Biofilter Water filtration Flow-through system abstract The zebra mussel (Dreissena polymorpha) is a notorious freshwater biofouling pest, and populations of the species can alter aquatic environments through their substantial filtration capabilities. Despite the ecological importance of zebra mussel filtration, many predictions of their large-scale effects on ecosystems rely on extrapolations from filtration rates obtained in static laboratory experiments, not accounting for natural mussel densities, boundary layer effects, flow rates or elevated algal concentrations. This study used large-scale industrial flume trials to investigate the influence of these factors on zebra mussel filtration and proposes some novel industrial applications of these findings. The flume trials revealed some of the highest zebra mussel clearance rates found to date, up to 574720 ml h 1 g 1 of wet tissue mass. Under low algal concentrations, chlorophyll a removal by zebra mussels was not proportional to mussel density, indicating that field rates of zebra mussel grazing may be much lower than previous studies have predicted. Increasing ambient velocities up to 100 ml s 1 (4 cm s 1 ) led to increased clearance rates by zebra mussels, possibly due to the replenishment of locally depleted resources, but higher velocities of 300 ml s 1 (12 cm s 1 ) did not lead to further significant increases in clearance rate. When additional algal cultures were dosed into the flumes, chlorophyll a removal increased approximately logarithmically with zebra mussel density and there were no differences in the clearance of three different species of alga: Ankyra judayi, Pandorina morum and Cyclotella meneghinia. Some novel industrial uses of these zebra mussel filtration studies are proposed, such as: (1) helping to inform models that predict the large- scale grazing effects of the mussels, (2) allowing estimates of zebra mussel densities in industrial pipelines, and (3) constructing large-scale biofilters for use in water clarification. & 2007 Elsevier Ltd. All rights reserved. 1. Introduction Much attention has been drawn to the industrial biofouling effects of zebra mussels around the world (Claudi and Mackie, 1994), but zebra mussels also have profound ecological effects on aquatic environments. Filter feeders, such as the zebra mussel, can be major consumers of phytoplankton, exerting significant top-down control on phytoplankton levels (Caraco et al., 1997). Because zebra mussels can reach densities of over 700,000 m 2 (Pathy, 1994), filter large volumes of water and retain a wide size range of particles (Sprung and Rose, 1988; Silverman et al., 1996), zebra mussel populations are capable of removing over 90% of organic matter from the water (MacIsaac, 1996). Since the zebra mussel invasion of North America, chlorophyll a concentration (an indicator of algal population density) has dropped by over 90% ARTICLE IN PRESS 0043-1354/$ - see front matter & 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.watres.2007.10.020 Corresponding author. Tel.: +44 1223336617; fax: +44 1223336676. E-mail address: [email protected] (P. Elliott). WATER RESEARCH 42 (2008) 1664– 1674
Transcript
ARTICLE IN PRESS
Available at www.sciencedirect.com
WAT E R R E S E A R C H 4 2 ( 2 0 0 8 ) 1 6 6 4 – 1 6 7 4
0043-1354/$ - see frodoi:10.1016/j.watres
�Corresponding autE-mail address: p
journal homepage: www.elsevier.com/locate/watres
Zebra mussel filtration and its potential uses in industrialwater treatment
Paul Elliotta,�, David C. Aldridgea, Geoff D. Moggridgeb
aDepartment of Zoology, University of Cambridge, Downing Street, Cambridge CB2 3EJ, UKbDepartment of Chemical Engineering, University of Cambridge, Pembroke Street, Cambridge CB2 3RA, UK
Table 1 – Results of two-way ANOVAs to test for the effectof sampling point and flow rate on chlorophyll aconcentration in flumes containing dead shells andliving zebra mussels
Factor F d.f. p
Dead shells
Sampling point 0.07 1 0.795
Flow 0.22 2 0.8
Position:flow 0.1 2 0.908
Living mussels
Sampling point 216.96 1 o0.001
Flow 9.96 2 o0.001
Fig. 3 – Influence of zebra mussel density in field-flumes on:
(a) percentage removal of chlorophyll a; (b) mass of
chlorophyll a removed; and (c) individual mussel clearance
rates.
Fig. 4 – Influence of flume flow speeds on (a) the reduction in
chlorophyll a concentration in flumes containing living and
dead zebra mussels and (b) the estimated clearance rates of
living zebra mussels.
WAT E R R E S E A R C H 4 2 ( 2 0 0 8 ) 1 6 6 4 – 1 6 7 41668
5-plate treatment, a mussel of length 22 mm would clear an
estimated 574720 ml h�1.
Position:flow 14.37 2 o0.001
3.2. Effects of flow speed on clearance rate
Chlorophyll a concentrations did not differ significantly
between the start and end of any flume containing dead
zebra mussel shells, no matter what the flow (Fig. 4a; Table 1).
In contrast, in flumes with living mussels, chlorophyll a
concentrations had significantly decreased by the end of the
flumes, with flow rate also having a significant effect on
chlorophyll a (Fig. 4a; Table 1).
Estimated clearance rates significantly differed across flow
comparisons showed that clearance rates were lowest at a
flow of 50 ml s�1 (po0.05), while the clearance rates at 100 and
300 ml s�1 did not differ significantly (p40.05).
3.3. Clearance rates with additional algae
After algal dosing, chlorophyll a concentration at the ends of
each flume followed a skewed unimodal pattern across time
(Fig. 5). Rather than compare each curve directly, the total
mass of chlorophyll a removed from each flume was
calculated using Eq. (2). The removal of different algal species
could not be compared directly because starting concentra-
tions of chlorophyll a in the different algal solutions were not
identical. Consequently, the total percentage of the dosed
chlorophyll a removed by the mussels was calculated. The
ARTICLE IN PRESS
Fig. 5 – Changes in chlorophyll a concentration with time in flumes exposed to 12 different combinations of experimental
treatments. Fifty millilitres of three different types of concentrated algal solution were dosed into flumes containing 0, 5, 10
and 15 plates of zebra mussels, flowing at 100 ml s�1 (the lines represent replicate measurements).
WA T E R R E S E A R C H 4 2 ( 2 0 0 8 ) 1 6 6 4 – 1 6 7 4 1669
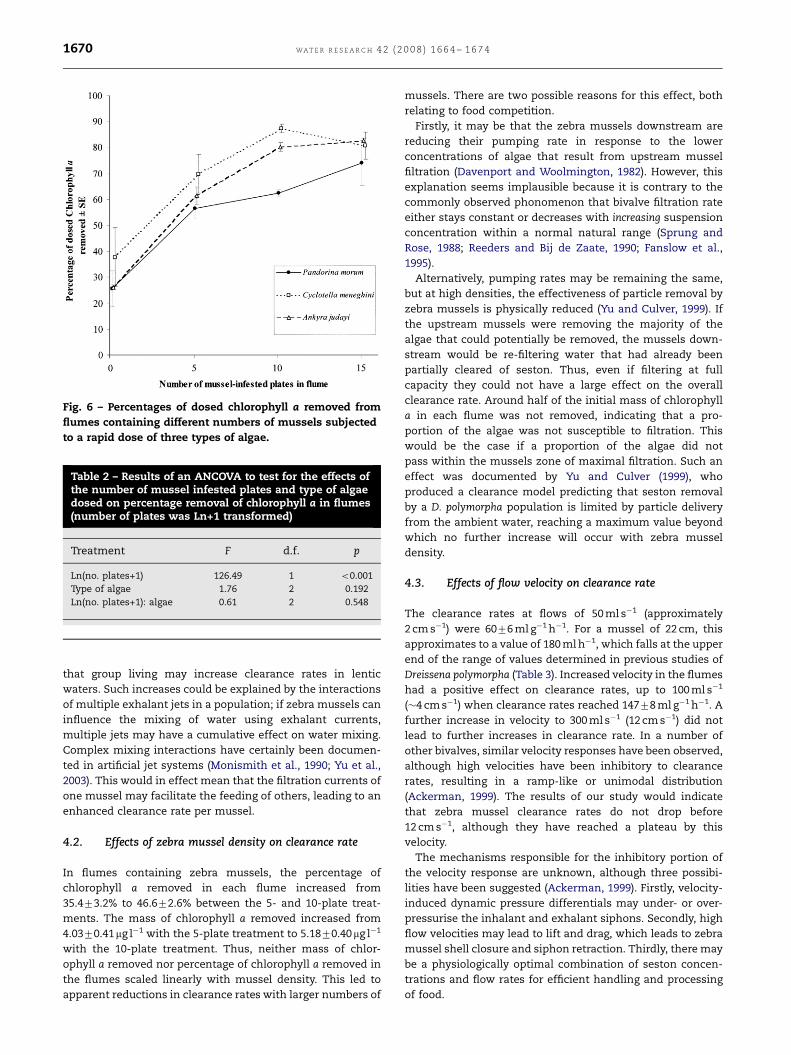
percentage of chlorophyll a removed increased significantly
with the natural logarithm of the number of plates of mussels
in the flume, but was not significantly affected by the algal
species used (Fig. 6; Table 2). There was no significant
interaction between the number of mussels in each flume
and the algal species used.
4. Discussion
4.1. Clearance rates in this study
The highest estimated clearance rate shown in this study
(based on wet mass extrapolations for 5 plates of mussels)
was 574720 ml h�1 g�1. This is a value far in excess of most
previous studies (Table 3, considering that mussels of length
19–20 mm weigh approximately 2 g). This may be because
previous studies using laboratory methods tend to under-
estimate filtration rates under natural conditions (Yu and
Culver, 1999). Many techniques that have been used in other
studies can negatively affect filtration rate, including artificial
diets, unnaturally high particle concentrations, short incu-
bation times, physical disturbance and chemical stress
(Mohlenberg and Riisgard, 1979; Reeders et al., 1989). The
values in this study more closely resemble the high values
for clearance found by Ackerman (1999) and Baldwin et al.
(2002), both of whom conducted experiments in a similar
flow-through apparatus; it appears that flow can increase
the effectiveness of the bivalve grazing. This may be because
flow generates increased mixing of the water, reducing
localised resource depletion. Peterson and Beal (1989) re-
viewed many marine studies and found that food particles
are often locally depleted around dense patches of suspen-
sion feeders.
In this study, the flow of the flumes would produce a
constant replenishment of algae around the mussels. This is
in contrast to many previous studies (commonly using the
chamber clearance method), which have often avoided
continuous mixing of the water to minimise disturbance of
the mussels (e.g. Sprung and Rose, 1988; Reeders et al., 1989).
As a consequence, the mussels in these other laboratory
experiments may undergo more re-filtration, reducing their
estimated clearance rates.
The high clearance rates observed in this study may also
result from the high densities of mussels that were used. It is
often assumed that the clearance rates of a population of
zebra mussels will be equal to the sum of the clearance rates
of constituent individuals (Stanczykowska, 1968; MacIsaac
et al., 1992; Bunt et al., 1993). Yu and Culver (1999) argue that
zebra mussel clearance will reach a maximum value beyond
which no further increase will occur with increased popula-
tion density due to localised resource depletion. The high
values for clearance found in the current investigation and a
similar study by Ackerman (1999) would certainly indicate
ARTICLE IN PRESS
Fig. 6 – Percentages of dosed chlorophyll a removed from
flumes containing different numbers of mussels subjected
to a rapid dose of three types of algae.
Table 2 – Results of an ANCOVA to test for the effects ofthe number of mussel infested plates and type of algaedosed on percentage removal of chlorophyll a in flumes(number of plates was Ln+1 transformed)
Treatment F d.f. p
Ln(no. plates+1) 126.49 1 o0.001
Type of algae 1.76 2 0.192
Ln(no. plates+1): algae 0.61 2 0.548
WAT E R R E S E A R C H 4 2 ( 2 0 0 8 ) 1 6 6 4 – 1 6 7 41670
that group living may increase clearance rates in lentic
waters. Such increases could be explained by the interactions
of multiple exhalant jets in a population; if zebra mussels can
influence the mixing of water using exhalant currents,
multiple jets may have a cumulative effect on water mixing.
Complex mixing interactions have certainly been documen-
ted in artificial jet systems (Monismith et al., 1990; Yu et al.,
2003). This would in effect mean that the filtration currents of
one mussel may facilitate the feeding of others, leading to an
enhanced clearance rate per mussel.
4.2. Effects of zebra mussel density on clearance rate
In flumes containing zebra mussels, the percentage of
chlorophyll a removed in each flume increased from
35.473.2% to 46.672.6% between the 5- and 10-plate treat-
ments. The mass of chlorophyll a removed increased from
4.0370.41mg l�1 with the 5-plate treatment to 5.1870.40mg l�1
with the 10-plate treatment. Thus, neither mass of chlor-
ophyll a removed nor percentage of chlorophyll a removed in
the flumes scaled linearly with mussel density. This led to
apparent reductions in clearance rates with larger numbers of
mussels. There are two possible reasons for this effect, both
relating to food competition.
Firstly, it may be that the zebra mussels downstream are
reducing their pumping rate in response to the lower
concentrations of algae that result from upstream mussel
filtration (Davenport and Woolmington, 1982). However, this
explanation seems implausible because it is contrary to the
commonly observed phonomenon that bivalve filtration rate
either stays constant or decreases with increasing suspension
concentration within a normal natural range (Sprung and
Rose, 1988; Reeders and Bij de Zaate, 1990; Fanslow et al.,
1995).
Alternatively, pumping rates may be remaining the same,
but at high densities, the effectiveness of particle removal by
zebra mussels is physically reduced (Yu and Culver, 1999). If
the upstream mussels were removing the majority of the
algae that could potentially be removed, the mussels down-
stream would be re-filtering water that had already been
partially cleared of seston. Thus, even if filtering at full
capacity they could not have a large effect on the overall
clearance rate. Around half of the initial mass of chlorophyll
a in each flume was not removed, indicating that a pro-
portion of the algae was not susceptible to filtration. This
would be the case if a proportion of the algae did not
pass within the mussels zone of maximal filtration. Such an
effect was documented by Yu and Culver (1999), who
produced a clearance model predicting that seston removal
by a D. polymorpha population is limited by particle delivery
from the ambient water, reaching a maximum value beyond
which no further increase will occur with zebra mussel
density.
4.3. Effects of flow velocity on clearance rate
The clearance rates at flows of 50 ml s�1 (approximately
2 cm s�1) were 6076 ml g�1 h�1. For a mussel of 22 cm, this
approximates to a value of 180 ml h�1, which falls at the upper
end of the range of values determined in previous studies of
Dreissena polymorpha (Table 3). Increased velocity in the flumes
had a positive effect on clearance rates, up to 100 ml s�1
(�4 cm s�1) when clearance rates reached 14778 ml g�1 h�1. A
further increase in velocity to 300 ml s�1 (12 cm s�1) did not
lead to further increases in clearance rate. In a number of
other bivalves, similar velocity responses have been observed,
although high velocities have been inhibitory to clearance
rates, resulting in a ramp-like or unimodal distribution
(Ackerman, 1999). The results of our study would indicate
that zebra mussel clearance rates do not drop before
12 cm s�1, although they have reached a plateau by this
velocity.
The mechanisms responsible for the inhibitory portion of
the velocity response are unknown, although three possibi-
lities have been suggested (Ackerman, 1999). Firstly, velocity-
induced dynamic pressure differentials may under- or over-
pressurise the inhalant and exhalant siphons. Secondly, high
flow velocities may lead to lift and drag, which leads to zebra
mussel shell closure and siphon retraction. Thirdly, there may
be a physiologically optimal combination of seston concen-
trations and flow rates for efficient handling and processing
of food.
ARTICLE IN PRESS
Table 3 – Literature review of the clearance rates of Dreissena polymorpha
Reference Testing device Food Size of mussel(mm)
Temperature(1C)
Clearance rate(ml mussel h�1)
Reeders et al. (1989) Stirred vessel Lake water 20–22 13–17.7 30–170
Claudi, R., Evans, D.W., 1993. Chemical addition strategies forzebra mussel control in once-through service water systems.In: Nalepa, T.F., Schloesser, D.W. (Eds.), Zebra Mussels: Biology,Impacts and Control. Lewis, Boca Raton, FL, pp. 563–574.
Claudi, R., Mackie, G.L., 1994. Practical Manual for Zebra MusselMonitoring and Control. Lewis, Boca Raton, FL.
Davenport, J., Woolmington, A.D., 1982. A new method ofmonitoring ventilatory activity in mussels and its use in astudy of the ventilatory patterns of Mytilus edulis-L. J. Exp. Mar.Biol. Ecol. 62, 55–67.
Elliott, P., Aldridge, D.C., Moggridge, G.D., 2005. The increasingeffects of zebra mussels on water installations in England.Water Environ. Manage. Dec, 367–375.
Fanslow, D.L., Nalepa, T.F., Lang, G.A., 1995. Filtration rates of thezebra mussel (Dreissena polymorpha) on natural seston fromSaginaw Bay, Lake Huron. J. Great Lakes Res. 21, 489–500.
Heath, R.T., Fahnenstiel, G.L., Gardner, W.S., Cavaletto, J.F., Hwang,S.J., 1995. Ecosystem-level effects of zebra mussels (Dreissenapolymorpha): An enclosure experiment in Saginaw Bay, LakeHuron. Journal of Great Lakes Research. 21, 501–516.
Jochimsen, E.M., Carmichael, W.W., An, J.S., Cardo, D.M., Cookson,S.T., Holmes, C.E.M., Antunes, M.B.D., de Melo, D.A., Lyra, T.M.,Barreto, V.S.T., Azevedo, S., Jarvis, W.R., 1998. Liver failure anddeath after exposure to microcystins at a hemodialysis centerin Brazil. N. Engl. J. Med. 338, 873–878.
Kryger, J., Riisgard, H.U., 1988. Filtration rate capacities in 6species of European fresh-water bivalves. Oecologia 77, 34–38.
Lei, J., Payne, B.S., Wang, S.Y., 1996. Filtration dynamics of thezebra mussel, Dreissena polymorpha. Can. J. Fish. Aquat. Sci. 53,29–37.
MacIsaac, H.J., 1996. Potential abiotic and biotic impacts of zebramussels on the inland waters of North America. Am. Zool. 36,287–299.
MacIsaac, H.J., Sprules, W.G., Johannsson, O.E., Leach, J.H., 1992.Filtering impacts of larval and sessile zebra mussels (Dreissenapolymorpha) in western Lake Erie. Oecologia (Berlin) 92, 30–39.
Mikheev, V.P., 1967. Filtration nutrition of the Dreissena. In:Voprosy prudovogo rybovodstva, 15, Pischevaya Promyshien-nost Publisher, Moscow, pp. 117–129 (in Russian with Englishsummary).
Mohlenberg, F., Riisgard, H.U., 1979. Filtration rate, using a newindirect technique. Mar. Biol. 54, 143–148.
Monismith, S.G., Koseff, J.R., Thompson, J.K., Oriordan, C.A., Nepf,H.M., 1990. A study of model bivalve siphonal currents.Limnol. Oceanogr. 35, 680–696.
Morton, B.S., 1971. Studies on the biology of Dreissena polymorphaPall; V. Some aspects of filter feeding and the effects of micro-organisms upon the rate of filtration. Proc. Malacol. Soc.London 39, 289–301.
Nicholls, K.H., Hopkins, G.J., 1993. Recent changes in LakeErie (north shore) phytoplankton: cumulative impacts of
ARTICLE IN PRESS
WAT E R R E S E A R C H 4 2 ( 2 0 0 8 ) 1 6 6 4 – 1 6 7 41674
phosphorous loading reductions and the zebra mussel intro-duction. J. Great Lakes Res. 19, 637–647.
Pathy, D.A., 1994. The life history and demography of the zebramussel, Dreissena polymorpha, populations in Lake St. Clair,Lake Erie, and Lake Ontario. Master abstracts 33/02, 489.
Peterson, C.H., Beal, B.F., 1989. Bivalve growth and higher orderinteractions: importance of density, site and time. Ecology 70,1390–1404.
Reed-Andersen, T., Carpenter, S.R., Padilla, D.K., Lathrop, R.C.,2000. Predicted impact of zebra mussel (Dreissena polymorpha)invasion on water clarity in Lake Mendota. Can. J. Fish. Aquat.Sci. 57, 1617–1626.
Reeders, H.H., Bij de Zaate, A.B., 1990. Zebra mussels (Dreissenapolymorpha)—a new perspective for water-quality manage-ment. Hydrobiologia 200, 437–450.
Reeders, H.H., DeVaate, A.B., Slim, F.J., 1989. The filtration rate ofDreissena polymorpha (Bivalvia) in 3 Dutch lakes with referenceto biological water quality management. Freshwater Biol. 22,133–141.
Reeders, H.H., Bij de Vaate, A., Noordhuis, R., 1993. Potential of thezebra mussel for water quality management. In: Nalepa, T.F.,Schloesser, D.W. (Eds.), Zebra Mussels: Biology, Impacts andControl. Lewis, Boca Raton, FL, pp. 439–454.
Riisgard, H.A., 2001. On measurement of filtration rates inbivalves—the stony road to reliable data: review and inter-pretation. Mar. Ecol. Prog. Ser. 211, 275–291.
Silverman, H., Achberger, E.C., Lynn, J.W., Dietz, T.H., 1995.Filtration and utilization of laboratory-cultured bacteria byDreissena polymorpha, Corbicula fluminea, and Carunculina tex-asensis. Biol. Bull. 189, 308–319.
Silverman, H., Lynn, J.W., Dietz, T.H., 1996. Particle capture by thegills of Dreissena polymorpha: structure and function of latero-frontal cirri. Biol. Bull. 191, 42–54.
Smith, T.E., Stevenson, R.J., Caraco, N.F., Cole, J.J., 1998. Changes inphytoplankton community structure during the zebra mussel(Dreissena polymorpha) invasion of the Hudson River (NewYork). J. Plankton Res. 20, 1567–1579.
Sprung, M., Rose, U., 1988. Influence of food size and foodquantity on the feeding of the mussel Dreissena polymorpha.Oecologia 77, 526–532.
Stanczykowska, A., 1968. The filtration capacity of populationsof Dreissena polymorpha in different lakes as a factor affectingcirculation of matter in the lake. Ekologia Polska B. 14,265–270.
Stanczykowska, A., 1984. Role of bivalves in the phosphorus andnitrogen budget in lakes. Verh. Inter. Ver. Theor. Angew.Limnol. 22, 982.
Vanderploeg, H.A., Strickler, J.R., Liebig, J.R., Nalepa, T.F., Fahnen-stiel, G.L., Gardener, W.S., Cavaletto, J.F., Fanslow, D.L., Johen-gen, T.H., 1995. Do zebra mussels promote blue-green andmetaphyton blooms on Sanginaw Bay, and do these bloomsaffect the mussels? In: Proceedings of the 38th Conferenceof the International Association of Great Lakes Research,p. 113.
Vanderploeg, H.A., Liebig, J.R., Nalepa, T.F., Johengen, T.H.,Strickler, J.R., 1996. Zebra mussels may be promoting Micro-cystis blooms in Sanginaw bay and Lake Erie through selectivefiltering and nutrient excretion. In: The Sixth InternationalZebra Mussel and Other Aquatic Nuisance Species Confer-ence, Dearborn, MI, March 1996.
Yu, D., Ali, M.S., Lee, J.H.W., 2003. Experiments on interaction ofmultiple jests in a crossflow. In: 16th ASCE EngineeringMechanics Course. University of Washington, Seattle.
Yu, N., Culver, D.A., 1999. Estimating the effective clearance rateand refiltration by zebra mussels, Dreissena polymorpha, in astratified reservoir. Freshwater Biol. 41, 481–492.