Page 1
R E S EA RCH AR T I C L E
Alkaline phosphatases in microbialites and bacterioplanktonfrom Alchichica soda lake, Mexico
Patricia M. Valdespino-Castillo1,2, Rocio J. Alc�antara-Hern�andez1,2, Javier Alcocer3,Mart�ın Merino-Ibarra4, Miroslav Macek3,5 & Luisa I. Falc�on2
1Posgrado en Ciencias del Mar y Limnolog�ıa, Universidad Nacional Aut�onoma de M�exico, Coyoac�an, Mexico; 2Laboratorio de Ecolog�ıa Bacteriana,
Instituto de Ecolog�ıa, Universidad Nacional Aut�onoma de M�exico, Coyoac�an, Mexico; 3Proyecto de Investigaci�on en Limnolog�ıa Tropical, FES
Iztacala, UNAM, Tlalnepantla, Estado de M�exico, M�exico; 4Unidad Acad�emica de Ecolog�ıa y Biodiversidad Acu�atica, Instituto de Ciencias del Mar
y Limnolog�ıa, Universidad Nacional Aut�onoma de M�exico, Coyoac�an, M�exico; and 5Academy of Sciences of the Czech Republic, Biology Centre
v. v. i. Institute of Hydrobiology, �Cesk�e Bud�ejovice, Czech Republic
Correspondence: Luisa I. Falc�on, Instituto de
Ecolog�ıa, Universidad Nacional Aut�onoma de
M�exico, Circuito exterior sn, Cd.
Universitaria, Coyoac�an 04510, Mexico.
Tel.: +52 55 5622 8222, ext. 46869;
fax: +52 55 5622 8995;
e-mail: [email protected]
Received 1 February 2014; revised 6 August
2014; accepted 7 August 2014.
DOI: 10.1111/1574-6941.12411
Editor: Gary King
Keywords
dissolved organic phosphorus utilization;
extracellular enzymes; low calcium
environment; microbial functional diversity.
Abstract
Dissolved organic phosphorus utilization by different members of natural com-
munities has been closely linked to microbial alkaline phosphatases whose affil-
iation and diversity is largely unknown. Here we assessed genetic diversity of
bacterial alkaline phosphatases phoX and phoD, using highly diverse microbial
consortia (microbialites and bacterioplankton) as study models. These micro-
bial consortia are found in an oligo-mesotrophic soda lake with a particular
geochemistry, exhibiting a low calcium concentration and a high Mg : Ca ratio
relative to seawater. In spite of the relative low calcium concentration in the
studied system, our results highlight the diversity of calcium-based metallo-
phosphatases phoX and phoD-like in heterotrophic bacteria of microbialites
and bacterioplankton, where phoX was the most abundant alkaline phosphatase
found. phoX and phoD-like phylotypes were more numerous in microbialites
than in bacterioplankton. A larger potential community for DOP utilization in
microbialites was consistent with the TN : TP ratio, suggesting P limitation
within these assemblages. A cross-system comparison indicated that diversity of
phoX in Lake Alchichica was similar to that of other aquatic systems with a
naturally contrasting ionic composition and trophic state, although no phylo-
types were shared among systems.
Introduction
Phosphorus is an essential element for cellular structure
and function. Its low availability in natural systems
(Wu et al., 2000) and its high turnover rates (Benitez-
Nelson & Buesseler, 1999; Benitez-Nelson, 2000; Ammer-
man et al., 2003) explain its demand by living systems and
correspond with a diverse metabolic network that has been
widely distributed in different life forms. We are starting
to recognize the identity of the organisms that harbour the
enzymatic tools for P transformations, among which
microbes, particularly heterotrophic bacteria (Dyhrman
et al., 2007; Cunha et al., 2010), play a central role (Kono-
nova & Nesmeyanova, 2002; White, 2009).
In oligotrophic waters, dissolved organic phosphorus
(DOP) typically constitutes the major fraction of total
dissolved P. Thus, regeneration of P from DOP has been
shown to play a significant role in supplying the P
required for biological production (Karl & Bj€orkman,
2001, 2002; Dyhrman et al., 2006; Young & Ingall, 2010),
particularly when the availability of dissolved inorganic
phosphorus (DIP) is low (e.g. in the upper layer of strati-
fied water bodies). Phosphomonoesters are an important
fraction of DOP in aquatic environments (Young &
Ingall, 2010). The degradation of these compounds
requires the action of phosphatases (Harke et al., 2012),
pH-dependent metalloenzymes (Jansson et al., 1988). The
catalytic activity of these enzymes is part of the initial
steps of organic matter degradation (Pinchuk et al.,
2008), where orthophosphate is cleaved hydrolytically
from organic molecules (Jansson et al., 1988). The
activity of these enzymes in aquatic systems has been
FEMS Microbiol Ecol && (2014) 1–16 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
MIC
ROBI
OLO
GY
EC
OLO
GY
Page 2
associated to bacterioplankton, phytoplankton (Jansson
et al., 1988; �Strojsov�a et al., 2003; Yamaguchi & Adachi,
2010) and zooplankton (Vidal et al., 2003), although their
diversity and distribution remain largely unknown
(Tringe et al., 2005; Cunha et al., 2010).
Recent studies, based on the amplification of genetic
regions associated to P-enzymes, have shown a differential
abundance and distribution of alkaline phosphatases (AP)
among microbial assemblages (Sakurai et al., 2008; Sebas-
tian & Ammerman, 2009). Alkaline phosphatase PhoA
has been widely described in cultured organisms such as
Escherichia coli. However, in an analysis of the Global
Ocean Survey dataset, Sebastian & Ammerman (2009)
proposed PhoX as a novel alkaline phosphatase more
broadly distributed in marine bacteria than the ‘classical
PhoA’. Soon after, Luo et al. (2009) reported that PhoD
was the most frequently AP found in marine environ-
ments. Along with these results, the availability of differ-
ent metal cofactors has been linked to the enzymatic
activity, and the potential presence and distribution of
AP (Monds et al., 2006; Wu et al., 2007; Luo et al., 2009;
Kathuria & Martiny, 2011). Ca+2 is a cofactor of
AP-PhoX and PhoD, whereas Mg+2 and Zn+2 are cofac-
tors of PhoA. In this sense, the low presence of phoA
genes could be related to low availability of Zn+2 in the
open ocean (sensu Luo et al., 2009). Although the infor-
mation about the presence and abundance of these genes
is growing, exploration of the diversity and dynamics of
AP in natural systems is still needed.
To gain insight into AP diversity, we used complex
microbial consortia (microbialites and bacterioplankton)
as study models from an oligo-mesotrophic (sensu Alco-
cer et al., 2000; Macek et al., 2009) environment with a
low phosphorus concentrations. The cationic proportions
of the saline alkaline Lake Alchichica (Fig. 1) differ signif-
icantly from those of seawater, particularly of Ca+2, Mg+2,
and Zn+2, displaying a peculiar chemistry characterized
by low calcium concentrations and a high Mg : Ca ratio
N
sM S3
400 m
sM S2
sM S4
sM S1
sM S5
sM S6
cM S3
cM S6
cM S5 cM S1,S4
cM S2 W (5, 25, 61 m)
Fig. 1. Microbialite and bacterioplankton sampling sites in Lake Alchichica, Puebla, highlands of Central Mexico. Outlined squares represent
spongy-type microbialites (subsample sites 1–6); white squares, columnar-type microbialites (subsamples 1–6) and the green circle shows the
water column bacterioplankton sampling sites (5, 25 and 61 m).
FEMS Microbiol Ecol && (2014) 1–16ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
2 P.M. Valdespino-Castillo et al.
Page 3
(Armienta et al., 2008; Ka�zmierczak et al., 2011; Mancil-
la-Villa et al., 2012), which resembles that of the ancient
soda ocean (sensu Kempe & Degens, 1985; Ka�zmierczak
et al., 2011).
Saline alkaline systems are habitats of a variety of
microorganisms, which have shown ecological and bio-
technological relevance (Antony et al., 2013). We
explored the distribution and diversity of functional genes
(alkaline metallophosphatases for DOP utilization) within
microbial communities (bacterioplankton and microbia-
lites) of Lake Alchichica to contribute to the understand-
ing of the interactions between the biogeochemical
phosphorus cycle (and therefore carbon cycle) and the
dynamics of other elements, as well as the identification
of AP-harbouring bacteria in this particular environment.
Materials and methods
Study area
Alchichica is a crater-lake located in the Central Mexican
Plateau (19°240N, 97°240W; 2340 m a.s.l.; Fig. 1) that
belongs to a cluster of six maar lakes on the easternmost
portion of the Trans-Volcanic Belt. The lake is saline
(8.5 g L�1), alkaline (pH 9.5) and deep (Zmean = 40.9 m
and Zmax = 62 m). The thermal regime of Lake Alchichi-
ca is warm-monomictic; the circulation period typically
occurs during the dry winter and the lake stratifies from
April to December. The ionic abundances of Lake Alchi-
chica are: Na+ > Cl� > Mg+2 > SO�24 > K+ > Ca+2 and
differ significantly from those of seawater, particularly
those of Ca+2, Mg+2, and Zn+2, because calcium concen-
tration is quite low (c. 0.3 mM) and there is a high
Mg : Ca ratio (c. 50; Armienta et al., 2008; Ka�zmierczak
et al., 2011). The study of this lake is now part of a
Long-Term Ecological Research program conducted by
UNAM (National Autonomous University of Mexico),
and its geological, hydrological and biological characteris-
tics have been described (Vilaclara et al., 1993; Lugo
et al., 1998; Alcocer et al., 2000).
The microbial assemblages of Lake Alchichica have also
been studied. Macek et al. (2009) surveyed the water-
column picoplankton and showed that total and auto-
trophic picoplankton dynamics follow a regular pattern
linked to the hydrodynamic cycle of the system. Larger
picoplankton peaks occur during circulation and early
stratification (January–March) and low numbers occur
during late stratification. Hern�andez-Avil�es et al. (2010)
identified, by FISH, a vertical zonation of bacterioplank-
ton components associated to diverse biogeochemical
processes, among which denitrification emerged as a
major pathway for N loss from the water body. Bautista-
Reyes & Macek (2012) applied CARD-FISH to describe
bacterial taxon annual shifts in abundance patterns and
their selective feeding by ciliates and heterotrophic flagel-
lates throughout the water column.
A discontinuous ring of microbialites is located along
the perimeter of this 2.26-km2 lake, where two different
morphologies have been reported and described as
spongy-type (white) and columnar-type (brown) sensu
Tavera & Kom�arek (1996) and Ka�zmierczak et al. (2011;
Fig. 2). The mineralogy, texture, isotopic features and
microbial composition are described in Ka�zmierczak et al.
(2011) and Centeno et al. (2012). The high Mg : Ca ratio
Fig. 2. Morphology of microbialites studied in Lake Alchichica: spongy microbialite (upper) and columnar microbialite (lower). The external
morphology of the exposed (emerged) microbialites is shown on left; a section of submerged microbialites (at a depth c. 0.3 m) is shown on the
right.
FEMS Microbiol Ecol && (2014) 1–16 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Phosphorus utilization by complex communities 3
Page 4
(48.1 : 69.3) of Alchichica water is reflected in the micro-
bialite mineralogy, where uncommon hydromagnesite-
magnesite minerals are present (Ka�zmierczak et al., 2011).
Besides its structural complexity (Fig. 2), Alchichica
microbialites have shown important activity related to the
N cycle in a system that has been proposed to be limited
mainly by this element (Ram�ırez-Olvera et al., 2009;
Ardiles et al., 2012). Microbialites have shown high N2
fixation rates produced primarily by heterocystous
cyanobacteria (Falc�on et al., 2002; Beltr�an et al., 2012).
Sampling
Samples were taken during well established circulation
(February) and stratification (August) periods in 2011
from the two morphological types of microbialites that
coexist in the perimeter of Lake Alchichica. Microbialite
samples of c. 10 g were taken at a depth of c. 0.30 m
from six sampling sites distributed throughout the perim-
eter of the lake (Fig. 1). Six samples were collected for
each microbialite morphological type (columnar and
spongy) per triplicate and per site during well established
lake circulation and during stratification. Spongy micro-
bialites are more widely distributed than columnar micro-
bialites, which are concentrated around the W–SW third
of the lake shoreline (Fig. 1). Samples were frozen at
�20 °C until DNA extraction.
Bacterioplankton samples were taken at the centre of
the lake (Fig. 1) from three water depths: 5, 25 and
61 m, corresponding to the epilimnion, metalimnetic base
(during stratification) and bottom water (1 m above the
sediment), respectively. The same depths were sampled
during well established lake circulation. Triplicate 0.5-L
water samples were filtered through Osmonics (Poretics
Corp.) polycarbonate membranes (pore size 0.22 lm) per
triplicate for each depth. Filters were placed in DNA-free
2.0 mL tubes and kept frozen at �20 °C until analysis.
Environmental characterization
Physicochemical characterization of water column sam-
ples was performed to frame alkaline phosphatase poten-
tial. Temperature, dissolved oxygen (DO), pH and
conductivity of the water column and of the water sur-
rounding the microbialites, were recorded with a YSI
6600 multiparameter probe. Water samples for nutrient
analysis including soluble reactive phosphorus (SRP),
NO�2 , NO�
3 , dissolved inorganic nitrogen (DIN), total
phosphorus (TP), and total nitrogen (TN) were collected
in the same sites and kept in polypropylene containers
after filtration through 0.45 and 0.22 lm (HA Milli-
poreTM) mixed cellulose esters membranes. Samples were
kept frozen until their analysis (within 24–48 h) with a
segmented flow Autoanalyzer (Skalar San-plus) using the
standard methods adapted by Grasshoff et al. (1983) and
the circuits suggested by Kirkwood (1994). Organic P was
calculated as TP minus SRP.
A subsample of 1 cm2 of area (from the surface to
approximately first 2 cm depth) of each microbialite was
excised, lyophilized (Savant SpeedVac drier, Waltham,
MA), and then ground in an agate mortar and pestle for
further analysis. A homogeneous subsample of 0.1 g of
each microbialite sample was analysed to quantify total
phosphorus and total nitrogen after high-temperature
persulphate oxidation (Valderrama, 1981) and to assess
the nutrient limitation condition of the microbialite.
Although microbialite samples were heterogeneous, in
each case they comprised 1 cm2 of surface-living tissue
and a fraction of mineral matrix.
DNA extraction
DNA was extracted from the six different sites for each
type of microbialite (spongy and columnar) and pooled
to account for the global composition of each microbia-
lite type. Microbialite DNA extraction was carried out
using the protocol of Zhou et al. (1996), modified for
microbialite samples (Centeno et al., 2012). Approxi-
mately 5 g of each microbialite was pulverized along with
extraction buffer (100 mM Tris-HCl, 20 mM NaCl,
100 mM EDTA, pH 8) and 0.06 V of cetyl trimethylam-
monium bromide (CTAB) in liquid nitrogen. The
obtained mixture was incubated with lysozyme
(30 mg mL�1; Sigma Aldrich) for 30 min at 37 °C and
subsequently with proteinase K (10 mg mL�1; Sigma
Aldrich) and 0.1 V of sodium dodecyl sulphate (SDS; at
55 °C, overnight). Samples were then centrifuged at
1800 g, 20 min. The aqueous phase was recovered and
extracted twice with phenol : chloroform : isoamyl alco-
hol (25 : 24 : 1) and once with chloroform : isoamyl
alcohol (24 : 1). DNA was precipitated overnight at
�20 °C by adding 2 V of 2-propanol, 0.1 V of 3 M
sodium acetate (3M) and 2 lL of GlycoBlue (Ambion
Inc.). The precipitated DNA was recovered by centrifuga-
tion (8000 g, 15 min), washed twice with 80% ethanol
and resuspended in molecular grade water. The DNA
samples were then purified through Mini Spin columns
of DNeasy Blood & Tissue kit (QIAGEN, Alameda, CA)
according to the manufacturer’s instructions, and precipi-
tated again as mentioned above. Purified DNA was stored
at �20 °C until analysis.
Alkaline phosphatase amplification
A region of c. 400 bp of the phoD gene was PCR-ampli-
fied following the protocol of Sakurai et al. (2008) with
FEMS Microbiol Ecol && (2014) 1–16ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
4 P.M. Valdespino-Castillo et al.
Page 5
the primers ALPS-F730 (50-CAGTGGGACGACCACGAGGT-30) and ALPS-R1101 (50-GAGGCCGATCGGCATGTCG-30). PCR was performed using c. 10 ng of
DNA in a final reaction mixture of 25 lL, containing
2 lM of each primer, 2.5 lL 109 ViBuffer S (Vivantis,
Oceanside, CA), 0.2 mM of each dNTP and 0.5 U of Taq
DNA Polymerase (Vivantis). Amplification steps com-
prised an initial denaturation step at 94 °C for 3 min, 35
cycles of a denaturing step at 94 °C for 1 min, annealing
at 59 °C for 1 min, and extension at 72 °C for 2 min,
followed by a final extension at 72 °C for 7 min and
cooling at 4 °C.A fragment of c. 600 bp of the phoX gene was amplified
from environmental DNA using F/R primers phoX1, phoX2
and phoX3 (Sebastian & Ammerman, 2009): phoX1-F
(50-GARGARAAYTTYAACGGCTA-30) and phoX1-R (50-GCCAKSACRWAVAGATCC-30); phoX2-F (50-GARGAGAACWTCCACGGYTA-30) and phoX2-R (50-GATCTCGATGATRTGRCCRAAG-30); phoX3-F (50-GGGNACTTAYYT-MACBTGYGAA-30) and phoX3-R (50-GDCKATCCATBGKBGTTGC-30). PCR was performed using c. 10 ng of
DNA in a final reaction mixture of 25 lL, containing
0.4 lM of each primer, 2.5 lL 109 ViBuffer S (Vivantis),
0.2 mM of each dNTP and 0.5 U of Taq DNA Polymerase
(Vivantis). The PCR program comprised an initial dena-
turation at 94 °C for 5 min, 35 cycles of 94 °C for 30 s,
annealing at 52 °C for 30 s, extension at 72 °C for 1 min,
final extension of 72 °C for 10 min and cooling at 4 °C.PCR products from five reactions were pooled and then
gel-purified using QIAquick spin columns (Qiagen). Clone
libraries were constructed for each sample (per microbia-
lite morphology and water column sampling site) and sea-
son. Amplified phoD and phoX fragments were ligated to
pCR� 2.1 vector using the Original TA Cloning Kit (Invi-
trogen, Carlsbad, CA). Chemically competent E. coli TOP-
10 cells were transformed and selected using LB plates with
ampicillin (100 lg mL�1) and X-Gal (80 lg mL�1);
clones were then further screened by PCR, using M13
primers.
Sequence analysis
Sequence alignment for functional genes was done with
the predicted amino acid sequences aligned with CLUSTAL
W to which DNA sequences were imposed to avoid inser-
tions, deletions, internal stop codons or reading frame
shifts. A BLAST similarity search of problematic sequences
was used to exclude pseudogenes from the analysis. The
nucleotide sequences obtained from microbialites and
bacterioplankton were BLAST-searched using the BLASTN
tool (\http://www.ncbi.nlm.nih.gov/Blast.cgi) in the
National Center for Biotechnology Information (NCBI)
database. Sequences of the entire dataset were aligned
with SEQUENCHER 4.1.4 (Gene Codes Corp., Ann Arbor,
MI). Maximum likelihood branch lengths were fitted to
consensus trees with PHYML 3.0 (Guindon et al., 2010).
Maximum likelihood bootstrap values were inferred from
1000 replicates. Phylogenetic topologies were visualized
and edited with FIGTREE (v.1.3.1). The determined AP par-
tial gene sequences were deposited in the GenBank data-
base under accession numbers KF891484–KF891515,KF891517–KF891828, KF891830–KF891882.
Operational taxonomic unit (OTU)-based analyses
from MOTHUR v.1.33.3 platform (Schloss et al., 2009)
were used to estimate diversity and coverage for the
selected genetic markers. Distance matrices and OTUs
for each set of sequences were calculated with the func-
tions ‘dist.seqs’ and ‘cluster’ using the furthest neighbour
algorithm. The cutoff value for phoX analysis was 0.04,
considered a reference criteria for defining OTUs of
functional genes (sensu Iwai et al., 2011). The phoD cut-
off value was 0.25, as estimated recently by Tan et al.
(2013). A nonparametric richness estimator (Chao) and
the Shannon diversity index (H0) were obtained for the
sets of sequences of Lake Alchichica but also for the sets
of sequences (phoX) found in different environmental
studies (see Table 1).
Results
A total of 398 AP novel sequences were recovered from
the environmental DNA extracted from microbialites and
bacterioplankton of Alchichica soda-lake. Water environ-
ment showed relatively high P concentrations and seemed
to be N limited, at least during the sampling dates
reported here (Table 2). In contrast, the TN : TP ratio of
microbialites, which integrates over a longer period,
points to P limitation in these consortia.
In this relatively low calcium system (high Mg : Ca
ratio), the abundance of calcium-based AP phoX was
higher overall than the phoD-like set of sequences, as can
be observed by the number of OTUs found (and also by
the number of clones; Table 1).
Physicochemical characterization and nutrient
status
Physicochemical parameters of the water surrounding mi-
crobialites (average of 12 sampled sites) are summarized
(Table 2), for circulation (February) and stratification
(August) periods along with environmental conditions in
the three water-column depths sampled for bacterioplank-
ton analysis.
Nutrients, TP and TN varied between stratification and
circulation (Table 2). Among the physicochemical parame-
ters measured, DO and pH showed a higher heterogeneity
FEMS Microbiol Ecol && (2014) 1–16 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Phosphorus utilization by complex communities 5
Page 6
Table
1.Bacterial
APphoD
andphoX
ofmicrobialassemblages
analysed
inthis
study:
microbialites
andbacterioplanktonofLake
Alchichica(duringcirculationan
dstratificationperiods)
and
environmen
taldatasetsfrom
differentphoXsurveys(Atlan
ticseaw
ater,mixed
water
andamicrocosm
experim
entin
Lake
Taihu,China)
System
Bacterial
assemblage
Season
phoD
phoX
No.sequen
ces
obtained
Unique
sequen
ces
OTU
s*Chao
Shan
non
(H0 )
Coverage
(%)
No.sequen
ces
obtained
Unique
sequen
ces
OTU
s†Chao
Shan
non
(H0 )
Coverage
(%)
Lake Alchichica‡
Microbialites
Circulation
39
15
20.0
2.22
92
50
16
30.0
2.28
88
Stratification
30
13
20.5
1.68
92
31
77.0
1.67
100
Bacterioplankton
Circulation
65
8.0
1.56
33
35
77.3
1.15
97
Stratification
19
77.3
1.44
93
11
616.0
1.04
72
Total
180
94
23§
68
2.16
94
192
120
27§
53.0
2.36
93
Sargasso
Seaan
d
Chesap
eake
Bay
¶Seaw
ater
––
––
–57
54
20
28.2
2.73
85
Lake
Taihu
Mixed
water**
––
––
–625
417
146
38.0
2.54
90
Microcosm
††
––
––
–1183
734
85
187.1
3.03
93
*phoDOTU
scutoff
=0.25sensu
Tanet
al.(2013).
†phoXOTU
scutoff
=0.05sensu
Iwai
etal.(2011).
‡Th
isstudy:
Lake
Alchichicaassembles:
Microbialites
=spongyan
dcolumnar
morphotype,
Bacterioplankton=dep
ths5,25an
d61m.
§OTU
sunshared
amongbacterial
assemblesan
dseasons.
¶ Seb
astian
&Ammerman
(2009):phoXfrom
theSargasso
Seaan
dChesap
eake
Bay
assemblages:seaw
ater
(dep
th=40m).
**Dai
etal.(2014):Lake
Taihuassemblages:mixed
water.
††Dai
(unpublished
):directGen
Ban
ksubmission.Lake
Taihuassemblage:
microcosm
experim
ent.
Table
2.Ph
ysicochem
ical
characterizationofLake
Alchichicalittoralzone(averageof12microbialitesamplingsites)
andwater
column(dep
ths5,25an
d61m
correspondto
water
column
bacterioplanktonsubsamples)
Dep
th
Circulation(Feb
ruary)
Stratification(August)
Littoralzone
Mean/ran
ge
5m
Epilimnion
25m
Metalim
nion
61m
Hypolim
nion
Water
column
Mean
Littoralzone
Mean/ran
ge
5m
Epilimnion
25m
Metalim
nion
61m
Hypolim
nion
Water
column
Mean
Temperature
(°C)
16.1
14.4
14.3
14.2
14.4
19.8
19.1
15.2
14.3
17.2
Conductivity(m
Scm
�1)
12.9
14.4
14.4
14.4
14.4
12.9
13.8
13.6
13.7
13.7
pH
7.6–9
.58.5
8.6
8.7
8.6
8.3–9
.58.5
8.5
8.5
8.5
DO
(mgL�
1)
6.4–1
0.1
6.6
5.9
3.9
5.7
5.9–7
.47.0
0.9
0.0
4.2
N�N
O� 3(lM)
0.66
0.19
0.67
0.53
0.49
0.59
4.85
8.04
4.32
5.92
DIN
(lM)
1.73
0.47
1.68
2.67
1.38
1.35
6.08
13.05
27.72
13.08
SRP(lM)
0.68
0.37
0.49
0.51
0.46
0.38
0.42
0.54
3.89
1.23
TP-SRP
2.3
5.8
5.8
6.5
5.9
2.4
5.6
6.2
7.7
6.3
TP(lM)
2.9
6.2
6.2
7.0
6.4
2.8
6.0
6.8
11.5
7.6
TN(lM)
38
72
64
67
69
40
65
70
82
69
DIN
:SR
P3
13
53
414
24
711
TN:TP
13
12
10
10
11
14
11
10
79
FEMS Microbiol Ecol && (2014) 1–16ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
6 P.M. Valdespino-Castillo et al.
Page 7
in the littoral zone, likely due to diel variability caused by
shallow depth and benthic macrophytes. Although the local
conditions for both types of microbialites varied, in general
they shared environmental conditions that distinguish the
littoral zone from the water column. In the littoral zone as
well as in the water column, nutrients, TP and TN varied
considerably between stratification and circulation, as can
be seen in Table 2.
SRP concentration was above P-limitation thresholds
(0.1 lM; Reynolds, 1999) in all sampling stations,
whereas DIN concentrations were below the N-limitation
threshold (6–7 lM; Ahlgren, 1989; Reynolds, 1999) dur-
ing circulation, and practically in the epilimnion during
the stratification. N : P ratios (DIN : SRP) were well
below 16 in most cases, pointing to the dominance of N
limitation in the Lake Alchichica water column. TN : TP
proportion also pointed to N limitation. Only during
stratification, and in particular at the 25 m depth, did the
N : P ratio suggest a condition of P limitation. In con-
trast, TN: TP ratios estimated for both microbialite types
(overall average 51 � 6; average � standard error;
spongy-type TN: TP = 54 � 7, columnar-type TN:
TP = 48 � 11) clearly indicated a strong phosphorus
limitation, marking a major functional difference between
bacterioplankton, where N-limitation is likely predomi-
nant, and microbialite assemblages, where P is limiting.
Concentrations of Ca+2, Mg+2 and Zn+2 reported for
Lake Alchichica were compared with those from seawa-
ter and Lake Taihu environmental phoX surveys (sum-
marized in Table 3). Ca+2 concentration in Alchichica is
quite low (0.27–0.37 mM), almost two orders of magni-
tude lower than Mg+2 (17.8–18.7 mM). Zn+2 concentra-
tion is high (0.459 mM, one order of magnitude above
seawater) particularly considering the salinity (8.5 g L�1)
of Lake Alchichica. The particularity of the proportions
of these metals, required by AP, and found in Lake Al-
chichica may help to widen our understanding of AP
diversity under these particular environmental condi-
tions.
Phylogenetic affiliation of phoX and phoD
A total of 218 and 180 sequences were obtained for
phoX and phoD genes, respectively. Sequences obtained
in this study were similar to AP from other environ-
mental studies and from genome databases. The
microbial consortia of Lake Alchichica (microbialites
and bacterioplankton) sequences for both phoX and
phoD were more similar between them than they were
to sequences from other environments, and were related
to Proteobacteria (mainly to Alphaproteobacteria) and
other heterotrophic groups such as Actinobacteria (Figs 3
and 4).
phoX sequences
Lake Alchichica AP phoX (from microbialites and bacte-
rioplankton) and reference sequences were organized in
five main clusters in the phylogenetic reconstruction
(Fig. 3). Cluster I (upper part of the tree) comprises
phoX-partial sequences from environmental studies along
with one sequence found only in the metalimnion of Lake
Alchichica (stratification 25 m), the single site where P
limitation was indicated by the N: P ratio. The geographi-
cal origin of the environmental sequences in Cluster I are
the Sargasso Sea surface water (Sebastian & Ammerman,
2009), and mixed water and sediments from Taihu Lake
(Dai et al., 2014). This cluster appears to be the most
unexplored phoX group, containing no sequences from
isolated strains. Cluster II comprises phoX sequences
exclusively from genome sections that belong to Betapro-
teobacteria and Actinobacteria, none of which were found
in the Alchichica microbial assemblages we studied. Clus-
ter III groups Alphaproteobacteria phosphatases or puta-
tive phosphatases and environmental phoX-partial
sequences. Most of Lake Alchichica phoX sequences
recovered from both microbialites and bacterioplankton,
were grouped here, although this clade also includes
phoX-partial sequences from marine waters and lake sedi-
ments. Alphaproteobacteria sequences in Cluster III are:
Sinorhizobium freddi (CP001389), Sinorhizobium meliloti
1021 (AL591688), Rhodobacter sphaeroides (CP000144),
Table 3. Trophic state and environmental conditions (concentration)
of major metal cofactors (Ca+2, Mg+2 and Zn+2) for bacterial AP. Data
include Lake Alchichica and environmental studies for alkaline
phosphatase gene surveys
System
Ion
Lake Alchichica
water column
Average
Sargasso Sea
Average (mM)
Lake Taihu
Mixed
water
Average Sediment
Ca+2 0.37†
0.27§10.3* 0.72¶ **
Mg+2 17.8†
18.7§53* 0.27¶ **
Zn+2 0.46‡ 0.05–9 9 10�3* 0.15¶ **
Mg : Ca 48.1†
69.3§5.30 0.38 0.88
Ca : Zn 806.1†‡
588.2§‡2 9 106 4.80 35.09
Trophic
state
Oligo-
mesotrophic
Oligotrophic Eutrophic –
*Millero (1996), †Armienta et al. (2008), ‡Mancilla-Villa et al. (2012),§Ka�zmierczak et al. (2011), and ¶Zhang et al. (2014).
Ion concentrations reported in **Wuenchuan et al. (2001) are
reported over dry mass (Ca+2 6387 mg kg�1, Mg+2 5633 mg kg�1,
Zn+2 182 mg kg�1), therefore only ionic ratios are included for com-
parison.
FEMS Microbiol Ecol && (2014) 1–16 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Phosphorus utilization by complex communities 7
Page 8
Ochrobactrum anthropi (CP000759), Starkeya novella
(CP002026), Mesorhizobium opportunistum (CP002279)
and Dinoroseobacter shibae DFL (CP0008830.1). Cluster
IV comprises a heterogeneous set of sequences: environ-
mental (Lake Alchichica) and genome sections from Al-
phaproteobacteria such as Paracoccus denitrificans PDI222
(CP000490.1), and members of the family Hyphomicrobia-
ceae such as Pelagibacterium halotolerans B2 (CP003075.1)
and Hyphomicrobium denitrificans, and Betaproteobacteria:
Ramlibacter tataouinensis TTB310 (CP000245) and Chlo-
roflexus aggregans (CP001337). Interestingly, most of the
sequences found in Alchichica were recovered only from
microbialites. Cluster V (bottom) is strongly separated
from the upper clades, grouping genomic phosphatase
sequences from Actinobacteria, such as Salinispora arenico-
la CNS-205 (CP000850.1), Salinispora tropica CNB-440
0.3
Alchichica lake microbialites (C)
KF891662, KF891663, KF891594
Alchichica microbialites (F)
CP001715.1 Candidatus Accumulibacter phosphatis
CP001964.1 Cellulomonas flavigena DSM 20109
KF891569, KF891574, KF891593, KF891674
Uncultured marine bacterium
30407155 Sinorhizobium meliloti 1021
KF891626, KF891635, KF891642, KF891646
CP002026.1 Starkeya novella DSM 506
KF891669
CP000830.1 Dinoroseobacter shibae DFL 12
Uncultured marine bacterium
KF891531, KF891532
CP000850.1 Salinispora arenicola CNS 205
Uncultured lake water and sediment bacterium
CP002279.1 Mesorhizobium opportunistum WSM2075
CP003563.1 Sinorhizobium fredii USDA 25
KF891519
CP001683.1 Saccharomonospora viridis DSM 43017
CP000667.1 Salinispora tropica CNB.440
Uncultured lake water and sediment bacterium
Alchichica microbialites and bacterioplankton (A)
Alchichica bacterioplankton (E)
328880049 Streptomyces venezuelae ATCC 10712
CP002896.1 Amycolaptopsis mediterranei S699
KF891566
Alchichica microbialites and bacterioplankton (D)
CP002083.1 Hyphomicrobium denitrificans ATCC 51888
CP003153.1 Dechlorosoma suillum PS
CP000759.1 Ochrobactrum anthropi ATCC 49188
KF891538, KF891541, KF891559, KF891636, KF891641
CP003075.1 Pelagibacterium halotolerans B2
CP001389.1 Sinorhizobium fredii NGR 234
CP001337.1 Chloroflexus aggregans DSM 9485
Alchichica bacterioplankton
Uncultured marine bacterium
KF891585, KF891591, KF891611, KF891616, KF891617
KF891650
CP001630.1 Actinosynnema nirum DSM 43827
KF891621, KF891647
Uncultured lake water and sediment bacterium
CP000144.1 Rhodobacter sphaeroides 2 4.1
CP000490.1 Paracoccus denitrificans PD1222
KF891620
Uncultured marine bacterium
KF891588
CP001867.1 Geodermatophilus obscurus DSM 43160
Alchichica microbialites and bacterioplankton (G)
15789340 Halobacterium sp. NRC.1
KF891643
CP000245.1 Ramlibacter tataounensis TTB310
Alchichica bacterioplankton (B)
751
989
997732
1000
996
908
819
1000
730
770
947
662
874
993
962
991
1000
851
1000
651
998
613
740
947
1000
996
1000
666
1000
658
840
820
991
603
603
927
569
1000
793
907
748
918
983
765
992
Cluster IV
Cluster I
Cluster II
Cluster III
Betaproteobacteria Actinobacteria
Uncultured lake (water and sediment) and marine bacterium
Actinobacteria Archaea
Alchichica microbialites and bacterioplankton
Alphaproteobacteria
Alchichica microbialites and bacterioplankton
Alphaproteobacteria
Fig. 3. Maximum likelihood topology for phoX alkaline phosphatase partial sequences. Magenta branches correspond to phoX sequences from
Alchichica microbialites (spongy morphological type and columnar-type) and bacterioplankton from three different depths (5, 25 and 61 m):
black branches and acquisition numbers correspond to phoX from bacterial genomes (predicted amino acid sequences domain COG3211:PhoX;
NCBI Conserved Domains) and colour branches correspond to uncultured bacterium phoX from lake water and sediments (green) and seawater
(blue). Sequences were retrieved from NCBI with BLASTN. Tree was constructed based on an alignment of c. 600 nucleotides; branches show
bootstrap testing over 1000 replicates (only bootstrap values > 60 are shown). The identity of Alchichica groups can be seen in Table S1
(Supporting Information).
FEMS Microbiol Ecol && (2014) 1–16ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
8 P.M. Valdespino-Castillo et al.
Page 9
(CP000667.1), Geodermatophilus obscurus DSM 43160
(CP001867.1), Streptomyces venezuelae ATCC 10712
(FR845719), Actinosynnema mirum DSM 43827
(CP001630.1) and the archaeon Halobacterium sp. NRC-1
(NC_002607). No sequences in this cluster were recov-
ered from Lake Alchichica samples.
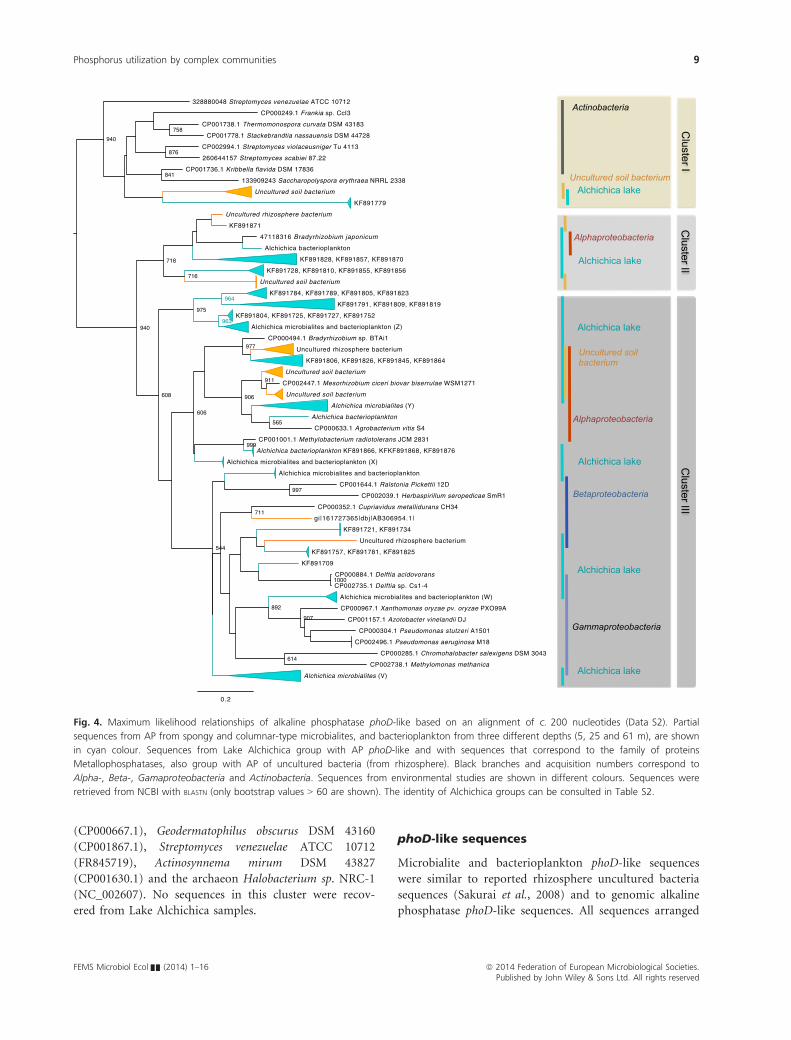
phoD-like sequences
Microbialite and bacterioplankton phoD-like sequences
were similar to reported rhizosphere uncultured bacteria
sequences (Sakurai et al., 2008) and to genomic alkaline
phosphatase phoD-like sequences. All sequences arranged
0.2
CP000967.1 Xanthomonas oryzae pv. oryzae PXO99A
CP001001.1 Methylobacterium radiotolerans JCM 2831
CP000884.1 Delftia acidovorans
CP000304.1 Pseudomonas stutzeri A1501
CP001736.1 Kribbella flavida DSM 17836
CP001778.1 Stackebrandtia nassauensis DSM 44728
CP001644.1 Ralstonia Pickettii 12D
47118316 Bradyrhizobium japonicum
CP002738.1 Methylomonas methanica
260644157 Streptomyces scabiei 87.22
Uncultured soil bacterium
Alchichica microbialites and bacterioplankton (Z)
KF891709
KF891828, KF891857, KF891870
Uncultured rhizosphere bacterium
Alchichica microbialites and bacterioplankton (X)
CP001157.1 Azotobacter vinelandii DJ
CP001738.1 Thermomonospora curvata DSM 43183
KF891871
KF891806, KF891826, KF891845, KF891864
Alchichica microbialites and bacterioplankton (W)
KF891791, KF891809, KF891819
CP002039.1 Herbaspirillum seropedicae SmR1
Uncultured rhizosphere bacterium
CP002496.1 Pseudomonas aeruginosa M18
CP000494.1 Bradyrhizobium sp. BTAi1
gi|161727365|dbj|AB306954.1|
CP002994.1 Streptomyces violaceusniger Tu 4113
CP000633.1 Agrobacterium vitis S4
CP002735.1 Delftia sp. Cs1-4
KF891779
CP000249.1 Frankia sp. CcI3
KF891784, KF891789, KF891805, KF891823
133909243 Saccharopolyspora erythraea NRRL 2338
Alchichica microbialites (V)
Alchichica microbialites and bacterioplankton
KF891804, KF891725, KF891727, KF891752
CP000352.1 Cupriavidus metallidurans CH34
Uncultured rhizosphere bacterium
Alchichica bacterioplankton
Alchichica microbialites (Y)
Uncultured soil bacterium
Uncultured soil bacterium
Alchichica bacterioplankton KF891866, KFKF891868, KF891876
Uncultured soil bacterium
328880048 Streptomyces venezuelae ATCC 10712
Alchichica bacterioplankton
KF891728, KF891810, KF891855, KF891856
KF891757, KF891781, KF891825
CP000285.1 Chromohalobacter salexigens DSM 3043
KF891721, KF891734
CP002447.1 Mesorhizobium ciceri biovar biserrulae WSM1271
565
975
606
963
608
997
940
906
940
907
999
964
876
977
711
544
718
892
1000
841
758
614
716
911
Cluster I
Cluster II
Cluster III
Alphaproteobacteria
Alphaproteobacteria
Betaproteobacteria
Gammaproteobacteria
Actinobacteria
Alchichica lake
Alchichica lake
Alchichica lake
Alchichica lake
Alchichica lake
Alchichica lake
Uncultured soil bacterium
Uncultured soil bacterium
Fig. 4. Maximum likelihood relationships of alkaline phosphatase phoD-like based on an alignment of c. 200 nucleotides (Data S2). Partial
sequences from AP from spongy and columnar-type microbialites, and bacterioplankton from three different depths (5, 25 and 61 m), are shown
in cyan colour. Sequences from Lake Alchichica group with AP phoD-like and with sequences that correspond to the family of proteins
Metallophosphatases, also group with AP of uncultured bacteria (from rhizosphere). Black branches and acquisition numbers correspond to
Alpha-, Beta-, Gamaproteobacteria and Actinobacteria. Sequences from environmental studies are shown in different colours. Sequences were
retrieved from NCBI with BLASTN (only bootstrap values > 60 are shown). The identity of Alchichica groups can be consulted in Table S2.
FEMS Microbiol Ecol && (2014) 1–16 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Phosphorus utilization by complex communities 9
Page 10
in a maximum likelihood tree are shown (Fig. 4), and
grouped in three clusters (Fig. 4). In Cluster I, sequences
from both types of Alchichica microbialites found during
lake circulation grouped with uncultured bacteria
sequences and phosphatases from Actinobacteria: Strepto-
myces venezuelae (gi.328880049), Streptomyces violaceusni-
ger Tu 4113 (CP002994.1), Streptomyces scabei 87.22(gi.
260644157), Frankia sp. CcI3 (CP000249.1), Thermomo-
nospora curvata DSM 43183 (CP001738.1) and Kribella
flavida DSM 17836 (CP001736.1). Cluster II comprised a
number of sequences of bacterioplankton and some
sequences from the microbialites grouped with uncul-
tured soil bacteria and the Alphaproteobacteria: Bradyrhiz-
obium japonicum (BA000040) alkaline phosphatase.
Cluster III included most of the recovered Alchichica
phoD-partial sequences. Microbialites and bacterioplank-
ton sequences group with some AP from uncultured soil
bacteria and with proteobacterial phosphatases in general,
and in particular with Alpha-, Beta- and Gammaproteobac-
teria as follows. Alphaproteobacteria: Bradyrhizobium sp.
Btail (CP000494.1), Mesorhizobium ciceri biovar biserrulae
WSM1271 (CP002447), Agrobacterium vitis S4 (CP000633.
1), Methylobacterium radiotolerans (CP001001.1). Betapro-
teobacteria sequences (middle clade) belonging to Ralsto-
nia pickettii 12D (CP001644.1), Herbaspirillum seropedicae
SmR1 (CP002039.1) and Cupriavidus metallidurans CH34
(CP000352.1). Gammaproteobacteria sequences from Delf-
tia acidovorans (CP000884.1), Delftia sp. Cs1–4(CP002735.1), Xantomonas oryzae pv oryzae PXO99A
(CP000967.1), Azotobacter vinelandii DJ (CP001157.1),
Pseudomonas stutzeri A1501 (CP000304.1) and Pseudomo-
nas aeruginosa M18 (CP002496.1), Chromohalobacter sa-
lexigens DSM 3043 (CP000285.1) and Methylomonas
methanica (CP002738.1). Similar to phoX sequences, there
are some clades that exhibit solely microbialite or bacterio-
plankton, circulation or stratification sequences (Fig. 4).
Diversity analysis
OTU numbers for microbialites and bacterioplankton, dur-
ing circulation and stratification, were slightly higher for
phoX (OTUs = 27, similarity cutoff = 0.05, sensu Iwai
et al., 2011 for functional genes) than for phoD
(OTUs = 23, similarity cutoff = 0.25 sensu Tan et al.,
2013; Table 1). Unique sequences constituted 62.5% of
total sequences for phoX and 52.2% for phoD. OTU-based
analyses were used to explore Lake Alchichica’s intrinsic
diversity of both AP studied. Microbialites and bacterio-
plankton from Alchichica shared six phoX OTUs and 10
phoD OTUs (equivalent to 22.2% and 43.5% of total OTUs,
respectively). For both markers (phoX and phoD), micro-
bialites showed higher diversity than bacterioplankton
samples (Table 1), except for the lake stratification period,
when bacterioplankton phoX was more diverse.
An OTU-based analysis was performed with four
uncultured bacterium-phoX datasets, using sequences
based on a c. 180 amino acids alignment (Data S1). These
sets of sequences are the results of environmental studies
that followed similar methods (e.g. using the same phoX
set of degenerated primers). The sources of these data
are: seawater (Sebastian & Ammerman, 2009), ‘mixed
water’ from shallow Lake Taihu (Dai et al., 2014) and a
microcosm experiment in Lake Taihu (J.Y. Dai, Effect of
microcystis bloom decomposition on genetic diversity of
bacterial phoX in an microcosm experiment, unpublished,
GenBank direct submission). Results are heterogeneous
and summarized in Table 1. The number of phoX OTUs
found was higher in the mixed water from Lake Tai-
hu > microcosm experiment in Lake Taihu > Lake Alchi-
chica > seawater. No phoX OTUs were shared among the
three systems (even extending the cutoff to 0.10); how-
ever, groups of Lake Taihu (mixed water and microcosm
experiment) shared 34 OTUs (cutoff = 0.05). To frame
the phoX genetic potential of each assemblage, we gath-
ered relevant environmental information on these systems
(Table 3).
Discussion
Potential for DOP utilization
Warm monomictic lakes (such as Alchichica) provide a
short-term (seasonal) changing scenario for their biota,
associated to the circulation–stratification annual cycle.
Bacterial alkaline phosphatase genes (phoX and phoD-like)
were present and widespread in the microbial consortia
of this soda system, although conductivity and species
richness have often shown an inverse relationship (Wil-
liams et al., 1990). Whereas conductivity is relatively
steady, both in the water column and in the littoral zone
of Alchichica, temperature, DO and pH are more variable
in the littoral zone, where microbialites are found
(Table 2, Fig. 5). Overall, pH conditions in this system
are in the optimal activity range for AP (Jansson et al.,
1988).
The available genetic potential for DOP utilization may
be relevant for communities that face important P avail-
ability changes in the short-term (e.g. SRP concentration
in the littoral zone and in surface waters approaches P
limitation thresholds during lake stratification). A greater
diversity of AP genes was found at depths where reminer-
alization is intense, such as the base of the metalimnion
and where inorganic P was abundant (e.g. the hypolim-
netic waters, close to the sediments). The depth distribu-
FEMS Microbiol Ecol && (2014) 1–16ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
10 P.M. Valdespino-Castillo et al.
Page 11
tion of DOP transformation genes has been explored
recently in marine waters. Luo et al. (2011) found that
phoX and phoD genes were distributed throughout the
water column (in the surface as well as in deep waters),
whereas phoA was found exclusively in deep waters. The
bacterioplankton assemblages of Lake Alchichica had a
widespread distribution of AP along the depth and nutri-
ent gradients. This observation agrees with Luo et al.’s
(2011) observation that AP may have a variety of func-
tions, such as contributing to the degradation of refrac-
tory organic matter (Luo et al., 2011) or nucleic acids, as
was pointed out by Pinchuk et al. (2008) for extracellular
PhoX.
The high TN: TP ratios (average 51) found in micro-
bialites suggest strong P limitation within these consortia
in contrast to bacterioplankton. As a group, AP have been
reported in microbialite metagenomes; a high abundance
of these genes was found in two microbialite metage-
nomes living in the Cuatro Ci�enegas Basin, which is also
a P-scarce environment in Northern Mexico (Breitbart
et al., 2009). The expected presence and diversity of DOP
transformation genes in complex assemblages (i.e. micro-
bialites) were verified here through the vast number of
AP sequences found and the fact that bacterial AP phoX
and phoD had not been described in detail in these
microbial consortia. It is notable that over half of the
phoX identified in Alchichica were recovered only from
microbialites, suggesting there may be a higher diversity
of AP in microbialite genomes than in other assemblages
(e.g. bacterioplankton) within the same system (Table 1).
Our results highlight the relevance of heterotrophic
bacteria, particularly Proteobacteria in DOP utilization by
AP. Proteobacteria is an abundant group of Alchichica mi-
crobialite bacterial composition (Centeno et al., 2012) as
well as of bacterioplankton during the stratification per-
iod, particularly Beta- and Gammaproteobacteria (Hern�an-
dez-Avil�es et al., 2010). This result is consistent with the
studies of Sebastian & Ammerman (2009) and Luo et al.
(2009), which found a great proportion of Alphaproteo-
bacteria harbouring phoX genes in marine and estuarine
samples, and databases. Other studies have highlighted
the role of these bacteria in organic matter utilization not
only through the use of AP but also through a diversity
of extracellular enzymes (Cunha et al., 2010).
Sebastian & Ammerman (2009) found a relevant pro-
portion of AP related to Planctomycetes in their study of
oceanic water. However, we did not find AP (phoX or
phoD) sequences related to Planctomycetes, in spite of
their relevant activity in the water column of Lake Alchi-
chica during the circulation period (sensu Hern�andez-
Avil�es et al., 2010). Further studies are needed to contrast
potential vs. functional participants in DOP utilization.
The strong genetic potential for DOP utilization found in
Lake Alchichica is consistent with the tendency of the
0 2 4
SRP ( µM )
0 10
TP ( µM )
0 10
NO3– ( µM )
0 20
DIN ( µM)
0 100
TN ( µM )
0 5 10
DO (mg L–1 )
0 5 10
pH –60
–50
–40
–30
–20
–10
0 5 15
Z (m
)
T (ºC)
Lake circulation Lake stratification
Fig. 5. General landscape of Lake Alchichica and physicochemical characterization of its water column (temperature, DO, pH); nutrient
concentration of soluble reactive phosphorus (SRP), nitrate (NO�3 ), DIN; TP and TN during well established circulation (February) and stratification
(August).
FEMS Microbiol Ecol && (2014) 1–16 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Phosphorus utilization by complex communities 11
Page 12
lake to mesotrophy, as seen by an abundance of microor-
ganisms similar to that in systems with a higher trophic
state (Bautista-Reyes & Macek, 2012) and with the pres-
ence of seasonal blooms, as previously described (Macek
et al., 2009).
Characterization of AP found in Lake Alchichica
Although the phylogeny of bacterial AP has not been
resolved yet, there have been advances in characterization
of these enzymes recently. Zaheer et al. (2009) identified
divergence in the structure of PhoX, and proposed two
groups of PhoX differing in having a conserved glycine
(PhoX-I) or asparagine (PhoX-II) next to their putative
catalytic Ca2+ binding site. Some of the Lake Alchichica
phoX sequences grouped to Alphaproteobacteria (P. deni-
trificans and S. meliloti), considered PhoX group II (sensu
Zaheer et al., 2009). The exploration of each of these
sequences in the Conserved Domains NCBI Database and
the HMMER platform (Finn et al., 2011) showed affilia-
tion to the protein family COG3211: PhoX (predicted
phosphatase, general function prediction only;
pfam05787: DUF839). Most of the sequences were recog-
nized as bacterial proteins of unknown function
(DUF839) that contain a predicted beta-propeller repeat.
Sinorhizobium meliloti 1021(Smc02634) PhoX has a Tat
signal peptide and Ca+2 requirement similar to that of
other PhoX proteins (Roy et al., 1982). The binding Ca+2
residues were recognized in all of the phoX sequences
generated in this study using the HMMER platform
search for amino acid sequences (residues E273 and E873,
sequence 3ZWU_A, Protein Data Bank).
Our results confirm that ALPS primers (Sakurai et al.,
2008) allow successful amplification of phoD-like
sequences, findings that are shared with Tan et al. (2013).
As seen in Fig. 3, phoD from Alchichica consortia and
their predicted amino acid sequences diversity were
related to different protein domains. BLAST searches found
that microbialite and bacterioplankton sequences were
related in general to Bacillus subtilis PhoD and related
proteins, including Metallophosphatases Domain
(cd07389 MPP_PhoD; NCBI, Conserved Domains). Other
sequences recovered were related to proteins of the Metal-
lophosphatase Superfamily, Metallophosphatase Domain
(cl13995 MPP_Superfamily; NCBI, Conserved Domains),
which are partial AP sequences of uncultured bacteria
from rhizosphere samples (JN388924 and JN388926; Chh-
abra et al., 2013), and uncultured soil bacteria
(AB306951, AB306956, AB306959, AB306960, AB306963,
AB306965, AB306967; Sakurai et al., 2008); this classifica-
tion is shared with Mesorhizobium loti MAFF303099
phosphatase (BA000012, Kaneko et al., 2000) and Delftia
acetivorans SPH-1 (CP000884, Schleheck et al., 2004).
Phosphatase sequences from Bradyrhizobium japonicum
USDA 110 are also classified in the MPP_PhoD domain.
Some of these protein domains have a recent assignation
and it is likely that the affiliation of AP will be expanded
and refined in the near future, as their structural com-
plexity has been related to their substrate non-specificity
(�Strojsov�a et al., 2003; Chr�ost & Siuda, 2002), substrate
promiscuity (O’Brien & Herschlag, 2001; Lassila & Hers-
chlag, 2008) and subcellular location (Luo et al., 2009).
Amplification with degenerated primers (Sakurai et al.,
2008) was very useful to find a large set of phoD-like AP
of diverse conformations, as revealed by the current clas-
sification of protein domains (in a large set of bacterial
phyla; see NCBI, INTERPRO 37.0). The calcium-binding site
of Bacillus subtilis PhoD (residues N215 and N216; 2YEQ,
Protein Data Bank) was present in 96% of the PhoD-like
translated sequences recovered from microbialites and
bacterioplankton of Lake Alchichica.
Alkaline phosphatase diversity
Overall, AP phoX were more abundant than phoD, as
revealed by the higher number of estimated phylotypes.
Microbialites had a higher abundance and diversity of
phoX and phoD AP compared with bacterioplankton
(Table 1). A higher potential for DOP utilization through
AP in microbialites is consistent with the clear evidence
of P limitation of microbialites (TN: TP = 51) vs. a less
defined nutrient limitation of bacterioplankton, as shown
by the environmental analyses (SRP, TP, organic P and
N: P ratios, Table 2).
OTU-based analyses using partial phoX and phoD
revealed that gene pools of these AP follow different dynam-
ics along the hydrological cycle of circulation–stratification(i.e. total OTUs of bacterioplankton in the circulation per-
iod are unique). A fraction of phoX and phoDOTUs (phylo-
types) were shared between circulation and stratification,
and also between microbialites and bacterioplankton, show-
ing that the same components of the communities poten-
tially use these AP for phosphorus utilization. However, the
potential for DOP utilization seems to be harboured by dif-
ferent members of the community over time.
phoX abundance and diversity in aquatic systems
(Table 1) represent the AP phoX potential in a broad gradi-
ent of environmental features: P-status (from oligotrophic
seawater to eutrophic conditions of Lake Taihu), commu-
nity structure (bacterioplankton, microbialites and a micro-
cosm experiment) and physicochemical and geochemical
characteristics, in which we highlighted calcium,magnesium
and zinc conditions (major metal cofactors of bacterial AP).
Many of Lake Alchichica sequences were similar to
those of Alphaproteobacteria, which constitute c. 30% of
the total composition of these microbialites (Centeno
FEMS Microbiol Ecol && (2014) 1–16ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
12 P.M. Valdespino-Castillo et al.
Page 13
et al., 2012), but primer characteristics may be constrain-
ing these results, too, underestimating phoD (Tan et al.,
2013), as well as phoX presence and diversity. Although
phoD has been reported to be the most abundant AP in
the ocean (Luo et al., 2009) and in agricultural soils (Tan
et al., 2013), in Lake Alchichica phoX was more abundant
than phoD. This is an interesting result that should be
weighed, considering the bacterial groups that our prim-
ers are able to identify (mainly Proteobacteria).
Ionic conditions
Calcium-based AP (PhoX and PhoD-like) were diverse in
Lake Alchichica microbial communities even though the
Ca+2 concentration in this system is low in relation to
seawater (Table 3). PhoX distribution was related more
to Alphaproteobacteria (Fig. 3), and PhoD-like and AP
similar to the Metallophosphatases Superfamily of pro-
teins were more broadly distributed among bacterial
groups (Alpha-, Beta-, Gammaproteobacteria and Actino-
bacteria; Fig. 4). It is interesting that some of the Lake
Alchichica AP-partial sequences showed affiliation to
sequences from other environmental studies (marine,
estuarine and soil), whereas the metal composition (Ca+2,
Mg+2 and Zn+2) is substantially different from these sys-
tems (Table 3). Mg : Ca and Ca : Zn ratios are higher in
Lake Alchichica than in seawater. The high Mg : Ca ratio
found in Alchichica likely has a direct impact on the
composition of microbialites, which are among the few
composed mainly of hydromagnesite and hydromagne-
site-magnesite (Ka�zmierczak et al., 2011).
Conclusion
Lake Alchichica represents a variable environment in terms
of P concentration, with low N and relatively low P condi-
tions, but characterized by high Mg : Ca water ratios
relative to seawater. There is a high potential for DOP uti-
lization through microbial AP (phoX and phoD-like) in the
microbialites and the bacterioplankton. We hypothesize
that organic phosphorus is strongly utilized in Lake Alchi-
chica, which would be consistent with the presumably low
P inputs from groundwater (Ka�zmierczak et al., 2011) and
atmospheric deposition (Oseguera et al., 2010). The utili-
zation of DOP would be important to regenerate the inor-
ganic phosphorus required to sustain the relatively high
primary production during the mesotrophic stages of the
lake (Ram�ırez-Olvera et al., 2009; Macek et al., 2009). Due
to the high pH and the AP metal dependence, the condi-
tions of Lake Alchichica are potentially favourable for the
activity of AP that use Ca+2 (PhoX and PhoD), Mg+2 and
Zn+2 (PhoA) as metal cofactors. Saline alkaline systems
often exhibit unique ionic compositions and harbour
microbial communities whose metabolic potential should
be explored (sensu Antony et al., 2013). These systems may
offer an opportunity to gain insight into the elemental
dynamics related to the biogeochemistry of the planet. In
this sense, the interaction of metals that function as cofac-
tors or inhibitors of these enzymes (used for P transforma-
tions) offers an interesting study model. The role of
metallophosphatases and other P enzymes in aquatic sys-
tems is key to understanding P utilization and its link to
systems productivity, and ultimately to carbon dynamics
and plankton blooms.
Acknowledgements
This manuscript is part of the degree requirements of the
Posgrado en Ciencias del Mar y Limnolog�ıa, UNAM
(P.V.), who has a graduate student fellowship awarded by
CONACyT. We thank Luis Oseguera and the ‘Proyecto
de Investigaci�on en Limnolog�ıa Tropical’ (FES Iztacala,
UNAM) and Bernardo Valdespino for fieldwork support
in Lake Alchichica. Sergio Castillo of the Aquatic Biogeo-
chemistry Laboratory (ICMyL, UNAM) is acknowledged
for providing instruments for fieldwork and for develop-
ing chemical analyses. Additionally, we thank Osiris Ga-
ona and Antonio Cruz-Peralta for technical and
laboratory assistance. We acknowledge Marcos Merino
for the English revision of the document. Funding for this
project was granted by SEP-CONACyT No. 151796
(L.I.F.) and PAPIIT-UNAM No. IT100212-3 (L.I.F.). CO-
NACyT and UNAM funded L.I.F. with sabbatical leave
grants. All sampling was done under collector permit No.
PPF/DGOPA.033/2013 (L.I.F.).
References
Ahlgren I (1989) Lake Restoration by Reduction of Nutrient
Loading: Expectations, Experiences, Extrapolations. Academia
Verlag, Richarz, St. Augustin.
Alcocer J, Lugo A, Escobar E, S�anchez MR & Vilaclara G
(2000) Water column stratification and its implications in
the tropical warm monomictic Lake Alchichica, Puebla,
Mexico. Verh Int Ver Theor Angew Limnol 27: 3168–3169.Ammerman JW, Hood RR, Case DA & Cotner JB (2003)
Phosphorus deficiency in the Atlantic: an emerging
paradigm in oceanography. Eos 84: 165–170.Antony CP, Kumaresan D, Hunger S, Drake HL, Murrell JC &
Shouche YS (2013) Microbiology of Lonar Lake and other
soda lakes. ISME J 7: 468–476.Ardiles V, Alcocer J, Vilaclara G, Oseguera LA & Velasco L
(2012) Diatom fluxes in a tropical, oligotrophic lake
dominated by large-sized phytoplankton. Hydrobiologia 679:
77–90.Armienta MA, Vilaclara G, De la Cruz-Reyna S, Ramos S,
Ceniceros N, Cruz O, Aguayo A & Arcega-Cabrera F (2008)
FEMS Microbiol Ecol && (2014) 1–16 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Phosphorus utilization by complex communities 13
Page 14
Water chemistry of lakes related to active and inactive
Mexican volcanoes. J Volcanol Geoth Res 178: 249–258.Bautista-Reyes F & Macek M (2012) Ciliate food vacuole
content and bacterial community composition in the warm
monomictic crater Lake Alchichica, M�exico. FEMS Microbiol
Ecol 79: 85–97.Beltr�an Y, Centeno CM, Garc�ıa-Oliva F, Legendre P & Falc�on LI
(2012) N2 fixation rates and associated diversity (nifH) of
microbialite and mat-forming consortia from different aquatic
environments in Mexico. Aquat Microb Ecol 67: 15–24.Benitez-Nelson CR (2000) The biogeochemical cycling of
phosphorus in marine systems. Earth-Sci Rev 51: 109–135.Benitez-Nelson CR & Buesseler KO (1999) Variability of
inorganic and organic phosphorus turnover rates in the
coastal ocean. Nature 6727: 502–505.Breitbart M, Hoare A, Nitti A, Siefert J, Haynes M, Dinsdale
E, Edwards R, Souza V, Rohwer F & Hollander D (2009)
Metagenomic and stable isotopic analyses of modern
freshwater microbialites in Cuatro Ci�enegas, Mexico.
Environ Microbiol 11: 16–34.Centeno CM, Legendre P, Beltr�an Y, Alc�antara-Hern�andez RJ,
Lidstr€om UE, Ashby MN & Falc�on LI (2012) Microbialite
genetic diversity and composition relate to environmental
variables. FEMS Microbiol Ecol 82: 724–735.Chhabra S, Brazil D, Morrissey J, Burke J, O’Gara F &
Dowling DN (2013) Fertilization management affects the
alkaline phosphatase bacterial community in barley
rhizosphere soil. Biol Fert Soils 49: 31–39.Chr�ost RJ & Siuda W (2002) Ecology of microbial enzymes in
lake ecosystems. Enzymes in the Environment (Burns RG &
Dick RP, eds), pp. 35–72. Marcel Dekker, Inc., New York, NY.
Cunha A, Almeida A, Coelho FJRC, Gomes NCM, Oliveira V
& Santos AL (2010) Bacterial extracellular enzymatic activity
in globally changing aquatic ecosystems. Current Research,
Technology and Education Topics in Applied microbiology and
Microbial Biotechnology, Microbiology Series No. 2, Vol. 1
(M�endez-Vilas A, ed.), pp. 124–135. FORMATEX, Badajoz,
Spain.
Dai J, Chen D, Gao G, Tang X, Wu S, Wu X & Zhou J (2014)
Recovery of novel alkaline phosphatase-encoding genes
(phoX) from eutrophic Lake Taihu. Can J Microbiol 60:
167–171.Dyhrman ST, Chappell PD, Haley ST, Moffett JW, Orchard
ED, Waterbury JB & Webb EA (2006) Phosphonate
utilization by the globally important marine diazotroph
Trichodesmium. Nature 439: 68–71.Dyhrman ST, Ammerman JW & Van Mooy BAS (2007)
Microbes and the marine phosphorus cycle. Oceanography
20: 110–116.Falc�on LI, Escobar-Briones E & Romero D (2002) Nitrogen
fixation patterns displayed by cyanobacterial consortia
in Alchichica crater-lake, Mexico. Hydrobiologia 467:
71–78.Finn RD, Clements J & Eddy SR (2011) HMMER web server:
interactive sequence similarity searching. Nucleic Acids Res
39(suppl 2): W29–W37.
Grasshoff K, Kremling K & Ehrhardt M (1983) Methods of
Seawater Analysis. Verlag Chemie, Weinheim.
Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W
& Gascuel O (2010) New algorithms and methods to
estimate maximum-likelihood phylogenies: assessing the
performance of PhyML 3.0. Syst Biol 59: 307–321.Harke MJ, Berry DL, Ammerman JW & Gobler CJ (2012)
Molecular response of the bloom-forming cyanobacterium,
Microcystis aeruginosa, to phosphorus limitation. Microb Ecol
63: 188–198.Hern�andez-Avil�es JS, Macek M, Alcocer J, Lopez-Trejo B &
Merino-Ibarra M (2010) Prokaryotic picoplankton dynamics
in a warm-monomictic saline lake: temporal and spatial
variation in structure and composition. J Plankton Res 32:
1301–1314.Iwai S, Chai B, Jesus EDC, Penton CR, Lee TK, Cole JR &
Tiedje JM (2011) Gene-targeted metagenomics (GT
Metagenomics) to explore the extensive diversity of genes of
interest in microbial communities. Handbook of Molecular
Microbial Ecology I: Metagenomics and Complementary
Approaches (De Bruijn FJ, ed.), pp. 235–243. John Wiley &
Sons. Inc., Hoboken.
Jansson M, Olsson H & Petterson K (1988) Phosphatases;
origin, characteristics and function in lakes. Hydrobiologia
170: 157–175.Kaneko T, Nakamura Y, Sato S, et al. (2000) Complete
genome structure of the nitrogen-fixing symbiotic bacterium
Mesorhizobium loti. DNA Res 7: 331–338.Karl DM & Bj€orkman K (2001) Phosphorus cycle in seawater:
dissolved and particulate pool inventories and selected
phosphorus fluxes. Methods Microbiol 30: 239–270.Karl DM & Bj€orkman K (2002) Dynamics of DOP.
Biochemistry of Marine Dissolved Organic Matter (Hansell D
& Carlson C, eds), pp. 246–366. Academic Press, San Diego,
CA.
Kathuria S & Martiny AC (2011) Prevalence of a
calcium-based alkaline phosphatase associated with the
marine cyanobacterium Prochlorococcus and other ocean
bacteria. Environ Microbiol 13: 74–83.Ka�zmierczak J, Kempe S, Kremer B, Lopez-Garcia P, Moreira
D & Tavera R (2011) Hydrochemistry and microbialites of
the alkaline crater Lake Alchichica, Mexico. Facies 57:
543–570.Kempe S & Degens ET (1985) An early soda ocean? Chem
Geol 53: 95–108.Kirkwood DS (1994) Sanplus Segmented Flow Analyzer and
Its Applications. Seawater Analysis. Skalar, Amsterdam.
Kononova SV & Nesmeyanova MA (2002) Phosphonates and
their degradation by microorganisms. Biochemistry 67: 220.
Lassila JK & Herschlag D (2008) Promiscuous sulfatase activity
and thio-effects in a phosphodiesterase of the
alkaline phosphatase superfamily. Biochemistry 47:
12853–12859.Lugo A, Alcocer J, Sanchez M & Escobar E (1998) Littoral
protozoan assemblages from two Mexican hyposaline lakes.
Hydrobiologia 381: 9–13.
FEMS Microbiol Ecol && (2014) 1–16ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
14 P.M. Valdespino-Castillo et al.
Page 15
Luo H, Bennera R, Long RA & Hu J (2009) Subcellular
localization of marine bacterial alkaline phosphatases. P Natl
Acad Sci USA 106: 21219–21223.Luo HW, Zhang HM, Long RA & Benner R (2011) Depth
distributions of alkaline phosphatase and phosphonate
utilization genes in the North Pacific Subtropical Gyre.
Aquat Microb Ecol 62: 61–69.Macek M, Alcocer J, Lugo-V�azquez A, Mart�ınez-P�erez ME,
Peralta Soriano L & Vilaclara Fatj�o G (2009) Long term
picoplankton dynamics in a warm-monomictic, tropical
high altitude lake. J Limnol 68: 183–192.Mancilla-Villa OR, Ortega-Escobar HM, Ram�ırez-Ayala C,
Uscanga-Mortera E, Ramos-Bello R & Reyes-Ortigoza AL
(2012) Metales pesados totales y ars�enico en el agua para
riego de Puebla y Veracruz, M�exico. Rev Int Contam Ambie
28: 39–48.Millero FJ (1996) Chemical Oceanography. CRC Press, Boca
Raton.
Monds RD, Newell PD, Schwartzman JA & O’Toole GA
(2006) Conservation of the Pho regulon in
Pseudomonas fluorescens Pf0-1. Appl Environ Microbiol 72:
1910–1924.O’Brien PJ & Herschlag D (2001) Functional interrelationships
in the alkaline phosphatase superfamily: phosphodiesterase
activity of Escherichia coli alkaline phosphatase. Biochemistry
40: 5691–5699.Oseguera LA, Alcocer J & Vilaclara G (2010) Relative
importance of dust inputs in aquatic biological
production as sources of lake sediments in an
oligotrophic lake in a semi-arid area. Earth Surf Proc
Land 36: 419–426.Pinchuk GE, Ammons C, Culley DE, Li SMW, McLean JS,
Romine MF, Nealson KH, Fredrickson JK & Beliaev AS
(2008) Utilization of DNA as a sole source of
phosphorus, carbon, and energy by Shewanella
spp.: ecological and physiological implications for
dissimilatory metal reduction. Appl Environ Microbiol 74:
1198–1208.Ram�ırez-Olvera MA, Alcocer J, Merino-Ibarra M & Lugo A
(2009) Nutrient limitation in a tropical saline lake: a
microcosm experiment. Hydrobiologia 626: 5–13.Reynolds CS (1999) Non-determinism to probability, or N : P
in the community ecology of phytoplankton: nutrient ratios.
Arch Hydrobiol 146: 23–35.Roy NK, Ghosh RK & Das J (1982) Monomeric alkaline
phosphatase of Vibrio cholerae. J Bacteriol 150: 1033–1039.Sakurai M, Wasaki J, Tomizawa Y, Shinano T & Osaki M
(2008) Analysis of bacterial communities on alkaline
phosphatase genes in soil supplied with organic matter. Soil
Sci Plant Nutr 54: 62–71.Schleheck D, Knepper TP, Fischer K & Cook AM (2004)
Mineralization of individual congeners of linear
alkylbenzenesulfonate by defined pairs of heterotrophic
bacteria. Appl Environ Microbiol 70: 4053–4063.Schloss PD, Westcott SL, Ryabin T, et al. (2009) Introducing
mothur: Open-source, platform-independent,
community-supported software for describing and
comparing microbial communities. Appl Environ Microbiol
75: 7537–7541.Sebastian M & Ammerman JW (2009) The alkaline
phosphatase PhoX is more widely distributed in
marine bacteria than the classical PhoA. ISME J 3: 563–572.�Strojsov�a A, Vrba J, Nedoma J, Kom�arkov�a J & Znachor P
(2003) Seasonal study of extracellular phosphatase
expression in the phytoplankton of a eutrophic reservoir.
Eur J Phycol 38: 295–306.Tan H, Barret M, Mooij MJ, Rice O, Morrissey JP, Dobson
A, Griffiths B & O’Gara F (2013) Long-term
phosphorus fertilisation increased the diversity of the total
bacterial community and the phoD phosphorus
mineraliser group in pasture soils. Biol Fert Soils 49:
661–672.Tavera R & Kom�arek J (1996) Cyanoprokaryotes in the
volcanic lake of Alchichica. Puebla State, Mexico. Algal Stud
83: 511–538.Tringe S, Von Mering C, Kobayashi A, Salamov A, Chen K,
Chang H, Podar M, Short J, Mathur E & Detter J (2005)
Comparative metagenomics of microbial communities.
Science 308: 554–557.Valderrama JC (1981) The simultaneous analysis of total
nitrogen and total phosphorus in natural waters. Mar Chem
10: 109–122.Vidal M, Duarte CM, Agust�ı S, Gasol JM & Vaqu�e D (2003)
Alkaline phosphatase activities in the central Atlantic Ocean
indicate large areas with phosphorus deficiency. Mar Ecol
Prog Ser 262: 43–53.Vilaclara G, Ch�avez M, Lugo A, Gonz�alez H & Gayt�an M
(1993) Comparative description of crater- lakes basic
chemistry in Puebla State, Mexico. Verh Int Ver Theor
Angew Limnol 25: 435–440.White AE (2009) New insights into bacterial acquisition of
phosphorus in the surface ocean. P Natl Acad Sci USA 106:
2013–2014.Williams WD, Boulton AJ & Taaffe RG (1990) Salinity as a
determinant of salt lake fauna: a question of scale.
Hydrobiologia 197: 257–266.Wu J, Sunda W, Boyle EA & Karl DM (2000) Phosphate
depletion in the western North Atlantic Ocean. Science 289:
759–762.Wu J-R, Shien J-H, Shien HK, Hu C-C, Gong S-R, Chen
L-Y & Chang P-C (2007) Cloning of the gene and
characterization of the enzymatic properties of
the monomeric alkaline phosphatase (PhoX) from
Pasteurella multocida strain X-73. FEMS Microbiol Lett
267: 113–120.Wuenchuan Q, Dickman M & Sumin W (2001) Multivariate
analysis of heavy metal and nutrient concentrations in
sediments of Taihu Lake, China. Hydrobiologia 450:
83–89.Yamaguchi H & Adachi M (2010) The utilization of organic
phosphorus by eukaryotic phytoplankton in marine
environments (Review). Bull Plankton Soc Japan 57: 1–12.
FEMS Microbiol Ecol && (2014) 1–16 ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
Phosphorus utilization by complex communities 15
Page 16
Young CL & Ingall ED (2010) Marine dissolved organic
phosphorus composition: insights from samples recovered
using combined electrodialysis/reverse osmosis. Aquat
Geochem 16: 563–574.Zaheer R, Morton R, Proudfoot M, Yakunin A & Finan TM
(2009) Genetic and biochemical properties of an alkaline
phosphatase PhoX family protein found in many bacteria.
Environ Microbiol 11: 1572–1587.
Zhang Y, Chen L, Dai J & Yu T (2014) Predict metal toxicity
and water quality criteria of different types of water into
Taihu Lake, China using biotic ligand model. Environ Nat
Resour Res 4: 10–18.
Zhou J, Bruns MA & Tiedje JM (1996) DNA recovery from
soils of diverse composition. Appl Environ Microbiol 62:
316–322.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Table S1. Identity of phoX sequences of Lake Alchichica
arranged in a maximum likelyhood topology (Fig. 3).
Table S2. Identity of phoD sequences of Lake Alchichica
arranged in a maximum likelyhood topology (Fig. 4).
Data S1. Amino acid alignment of phoD primers with
genomic references and Alchichica sequences obtained in
this study.
Data S2. Nucleotide alignment of phoD primers with
genomic references and Alchichica sequences obtained in
this study.
FEMS Microbiol Ecol && (2014) 1–16ª 2014 Federation of European Microbiological Societies.Published by John Wiley & Sons Ltd. All rights reserved
16 P.M. Valdespino-Castillo et al.