Carbon trading for phosphorus gain: the balance betweenrhizosphere carboxylates and arbuscular mycorrhizalsymbiosis in plant phosphorus acquisitionpce_2547 2170..2180
M. H. RYAN1,3,4, M. TIBBETT2,4,6, T. EDMONDS-TIBBETT1,3, L. D. B. SURIYAGODA1,3,4,7, H. LAMBERS1,3,G. R. CAWTHRAY1,3 & J. PANG1,3,5
Schools of 1Plant Biology and 2Earth and Environment, 3Institute of Agriculture, 4Future Farm Industries CooperativeResearch Centre, The University of Western Australia, Crawley, WA 6009, 5CSIRO Plant Industry, Centre for Environmentand Life Sciences, Wembley, WA 6913, Australia, 6National Soil Resources Institute, Department of Environmental Scienceand Technology, School of Applied Science, Cranfield University, Cranfield, Bedfordshire MK43 0AL, UK and 7Faculty ofAgriculture, University of Peradeniya, Peradeniya 20400, Sri Lanka
ABSTRACT
Two key plant adaptations for phosphorus (P) acquisitionare carboxylate exudation into the rhizosphere and mycor-rhizal symbioses. These target different soil P resources,presumably with different plant carbon costs. We examinedthe effect of inoculation with arbuscular mycorrhizal fungi(AMF) on amount of rhizosphere carboxylates and plant Puptake for 10 species of low-P adapted Kennedia grown for23 weeks in low-P sand. Inoculation decreased carboxylatesin some species (up to 50%), decreased plant dry weight(21%) and increased plant P content (23%). There was apositive logarithmic relationship between plant P contentand the amount of rhizosphere citric acid for inoculated anduninoculated plants. Causality was indicated by experi-ments using sand where little citric acid was lost from thesoil solution over 2 h and citric acid at low concentrationsdesorbed P into the soil solution. Senesced leaf P concen-tration was often low and P-resorption efficiencies reached>90%. In conclusion, we propose that mycorrhizally medi-ated resource partitioning occurred because inoculationreduced rhizosphere carboxylates, but increased plant Puptake. Hence, presumably, the proportion of plant Pacquired from strongly sorbed sources decreased withinoculation, while the proportion from labile inorganic Pincreased. Implications for plant fitness under field condi-tions now require investigation.
Soil microbes may strongly impact plant resource partition-ing (Bever et al. 2010). This could occur through soil
microbes affecting plant functional traits such as those asso-ciated with nutrient uptake (Friesen et al. 2011). Phospho-rus (P) is often a key limiting resource in natural andmanaged ecosystems (Lambers et al. 2008) and, as a conse-quence, many plant species form an association with arbus-cular mycorrhizal fungi (AMF) (Brundrett 2009). Hyphaeof AMF enhance plant P uptake primarily through access-ing P from the soil labile inorganic P pool beyond rootP-depletion zones (Jakobsen, Abbott & Robson 1992; Yaoet al. 2001). However, plants also possess adaptations toenhance P uptake from other soil P resources. For instance,the release by roots of low-molecular-weight organic anions(carboxylates), as found in its most extreme form in speciesthat form cluster roots (Shane & Lambers 2005), can mobi-lize strongly sorbed P (Jones 1998).
Plant adaptations to acquire P may impose significantcarbon costs (Lynch & Ho 2004). Both formation of arbus-cular mycorrhizas and exudation of carboxylates into therhizosphere are thought to be carbon-demanding processes(Dinkelaker, Römheld & Marschner 1989; Graham 2000).A high carbon cost is consistent with both colonization byAMF and rhizosphere carboxylates decreasing as plant Pstatus increases (Ryan & Angus 2003; Pearse et al. 2006;Pang et al. 2010a), and with inoculation with AMF reducingroot exudation (Graham, Leonard & Menge 1981; Mada &Bagyaraj 1993).Thus, it is conceivable that carbon costs maylimit the ability of plants to simultaneously host AMF andexude large amounts of carboxylates into the rhizosphere.
Indeed, plant species that form cluster roots occur promi-nently on very low-P ancient soils in Australia or stronglyP-sorbing young soils in Chile and tend to be non-mycorrhizal (e.g. Australian Proteaceae) (Brundrett 2009;Lambers et al. 2012). However, plant species adapted to lessextreme environments appear to possess an intermediatestrategy. For instance, the Australian native perenniallegumes Lotus australis Andrews and Cullen australasicum(Schltdl.) J.W. Grime can become highly colonized by AMF(Tibbett et al. 2008), but also have high amounts of carboxy-lates in rhizosphere soil in the absence of cluster roots
(Pang et al. 2010a). Individual plants of such species couldperhaps alter the balance between colonization by AMFand exudation of carboxylates into the rhizosphere inresponse to P availability and presence of AMF, that is,exhibit phenotypic plasticity (see Ashton et al. 2010). Whilesuch plasticity could act to diminish carbon expenditure, itcould also allow partitioning of the soil P resource amongplants, due to AMF and rhizosphere carboxylates assistingplant access to different soil P resources (Lambers et al.2008).
We examined this idea using Kennedia, a genus compris-ing Australian perennial herbaceous legumes. Kennediaspecies were chosen as their propensity to show P-toxicitysymptoms at relatively low P-application rates (Pang et al.2010a,b) suggests they are well adapted to very low-P soils(Lambers et al. 2010). In addition, some species can becomehighly colonized by AMF (Denton et al. 2006). The 10Kennedia species used in the present experiment includesome with very wide distributions across a range of habitats(K. prorepens, K. prostrata, K. rubicunda) and some withvery restricted distributions in south-western Australia(K. beckxiana, K. lateritia, K. microphylla) (FloraBase2010; PlantNET 2010), a region where P is severely limiting(Lambers et al. 2010). We therefore expected that the 10Kennedia species would possess a range of adaptations tolow-P conditions. We hypothesized that the Kennediaspecies would reduce carboxylate exudation, and thusaccess different P resources, when an additional carbondrain was introduced through inoculation with AMF.
MATERIALS AND METHODS
Kennedia experiment
The experiment was a factorial randomized block designwith two factors, Species (10 Kennedia species, one repre-sented by two accessions) and Inoculation [plus inoculation(+Inoc) and minus inoculation (–Inoc) with AMF]. Therewere four replicates. Seed of K. stirlingii Lindl. was kindlysupplied by Dr Mike Ewing (The University of WesternAustralia). Seed of remaining species was purchased from acommercial supplier (Nindethana Australian Seeds). Thesespecies were K. beckxiana (F.Muell.) F.Muell., K. carinata(Benth.) Domin, K. coccinea Vent., K. eximia Lindl., K. la-teritia F.Muell., K. microphylla Meisn., K. nigricans Lindl.,K. prorepens (F.Muell.) F.Muell., K. prostrata R.Br. (acces-sion I and accession II) and K. rubicunda Vent. All speciesoccur naturally in south-western Australia, except K. rubi-cunda, which occurs along the eastern coast of Australia.
The inoculum was prepared as follows. Lancelin sand wassteamed at 80 °C for 1 h each on two successive days anddried overnight at 100 °C. The dried Lancelin sand con-tained 4 mg kg-1 bicarbonate-extractable P (Colwell 1963),7 mg kg-1 mineral N (Searle 1984) and had a pH (CaCl2) of5.0 (Rayment & Higginson 1992). A P-free complete nutri-ent solution (see below) was added to the Lancelin sand.The Lancelin sand was inoculated with a fungal isolate,DAOM 181602, which has been widely used as a model
fungus for mycorrhizal studies and was originally collectedfrom Pont Rouge, Québec, Canada (Stockinger, Walker &Schüßler 2009). Recently, this fungus has been moved to thenew taxon of Rhizophagus irregularis (Schüßler and Walker2010). Leek (Allium porrum L.) was used as a host plant.
White non-draining pots were filled with 1080 g ofwashed and pasteurized (as above) coarse river sand andeither 120 g of leek pot culture (sand, roots, hyphae andspores) (+Inoc) or 120 g of the sterile Lancelin sand(–Inoc). The dried, washed river sand contained 5 mg kg-1
mineral N (Searle 1984), 3 mg kg-1 bicarbonate-extractableP (Colwell 1963), 6 mg kg-1 total P (Allen & Jeffery 1990),and had a pH (CaCl2) of 6.3 (Rayment & Higginson 1992)and a P-retention index of 1.5 mL P g-1 (Allen & Jeffery1990).Analyses were performed by CSBP FutureFarm ana-lytical laboratories (Bibra Lake, Australia). The pots wereinitially wetted to 100% field capacity with a P-free nutrientsolution (described below); they were then returned to100% field capacity by weighing and watering with deion-ized water twice a week. Final concentrations of elementsadded were (mg kg-1 dry soil): N 30, K 100, S 50, Ca 24, Mg10, Cu 0.5, Zn 2, Mn 4, B 0.119, Mo 0.4, Fe 5 and Cl 23. Nwas added only at sowing as a mixture of NH4NO3 andCa(NO3)2 with a molar ratio of 1. Pots were inoculatedwith rhizobia isolated from K. prostrata and K. prorepensobtained from the Rutherglen Centre, Department ofPrimary Industries, Victoria, Australia. No P was added ini-tially to pots, but in response to signs of P deficiency,2 mg kg-1 of P as K2HPO4 was added at week 11 and againat week 18.
Kennedia seeds were placed in boiling water for 10 min.Any seeds that were then not imbibed had their seed coatscut with a scalpel and were allowed to imbibe. Multipleimbibed seeds were planted in pots on 10 August 2007.Germination and emergence in some species wereextremely slow, but the majority of species had emerged by10 September (week 4). Seedlings were later thinned to oneplant per pot. The experiment was carried out in a naturallylit glasshouse at The University of Western Australia, Perth.Average daily minimum and maximum temperatures foreach month of the experiment were: 13/27 °C August(winter), 13/27 °C September, 15/30 °C October, 18/33 °CNovember (spring), and 18/33 °C December and 21/32 °CJanuary (summer).
Plants were harvested on 20 January 2008 (week 23).Shoots were separated into stems, green leaves andsenesced leaves. Root systems were gently shaken toremove excess sand, with the sand that remained attachedto the roots defined as rhizosphere soil. The roots andattached sand were transferred to a beaker containing avolume of 0.2 mm CaCl2 adequate to allow immersion of theroot system; the use of CaCl2 ensured cell integrity wasmaintained. Roots were gently shaken to remove the rhizo-sphere soil as quickly as possible, with care taken to mini-mize root damage. A 1 mL subsample of the rhizosphereextract was filtered through a 0.22 mm syringe filter into ahigh-performance liquid chromatography (HPLC) vial,acidified with 5 mL orthophosphoric acid per mL of extract
and frozen at -20 °C until analysis. The root system wasthen thoroughly washed, fresh mass measured and a sub-sample of ~250 mg was taken and placed in 10% (w/v)KOH for staining for AMF. Root systems were carefullyexamined in a shallow water bath for the presence of clusterroots. Dry weights (DWs) of shoot components and remain-ing roots were determined after drying for 48 h at 70 °C.Root mass ratio was calculated as:
Root mass ratio Root DW Root DW Shoot DW= +( )
Rhizosphere extracts were analysed for low-molecular-weight carboxylates. Working standards of malic, malonic,iso-citric, shikimic, lactic, acetic, maleic, citric, succinic,fumaric, cis-aconitic and trans-aconitic acid (ICN Biomedi-cals Inc., Aurora, OH, USA) were used to identify carboxy-lates on an Alltima C-18 reverse phase column (250 ¥4.6 mm, Alltech, Deerfield, IL, USA) (Cawthray 2003).Oxalic acid was also screened, but no appreciable amountswere found. Root subsamples for determination of mycor-rhizal colonization levels were left in KOH for 5 d at ~25 °Cbefore being stained in a 5% (v/v) Sheaffer black ink/vinegar solution for 1 h and stored in lactoglycerol (Vier-heilig et al. 1998). The percentage of root length colonizedby AMF was calculated using the line intersect method(Giovannetti & Mosse 1980).
To determine P concentrations in plant components, a100 mg subsample of coarsely ground green leaves,senesced leaves, stems or roots was digested in a 3:1,HNO3:HClO4 solution and P was measured using a UV-Visspectrophotometer (Shimadzu Corporation, Kyoto, Japan)by the malachite green method (Motomizu, Wakimoto &Toei 1983). Senesced leaves were not present for all plants.P-resorption efficiency was calculated according to VanHeerwaarden, Toet, & Aerts (2003):
Two experiments were used to examine three aspects ofcarboxylate dynamics in washed river sand: (1) degradationover time of citric acid (with and without soil microbes); (2)effect of concentration of citric and malic acids on theirdegradation; and (3) effect of concentration of citric andmalic acids on P desorption into the soil solution. In the firstexperiment, based on Jones & Brassington (1998), a 200 mmcitric acid solution was prepared in 5 mm KCl and pHadjusted to 4.5 with KOH. This solution was divided intotwo equal volumes and 0.25% v/v chloroform was added toone. Next, 10 mL tubes were filled with 1 g of air-dried soiland 5 mL of either the (+) or (-) chloroform carboxylatesolution and rotated on an end-over-end shaker at320 r.p.m. for 0, 5, 10, 20, 30, 45, 60, 90 or 120 min. Therewere three replicates. Tubes were then centrifuged at16 000 g for 20 min, until the solution became clear enoughto remove the supernatant with a syringe. Supernatant wasfiltered through a 0.22 or 0.45 mm filter into an HPLC vial.Samples were acidified with 5 mL of concentrated HCl (i.e.
32%), to allow soil solution [P] to be determined, andquickly stored on ice before being frozen at -20 °C. Sampleswere later analysed for citric acid as described previously.For the second experiment, individual solutions of citric andmalic acids were prepared at 0, 1, 5, 10, 20, 50, 100, 200, 500and 1000 mm in 5 mm KCl, and pH was adjusted to 4.5 withKOH. There were four replicates. Chloroform was notadded. The experiment then proceeded as described previ-ously, except that the samples were rotated on the shakerfor 20 min only and supernatant was added to HPLC vials(for the determination of citric or malic acid concentration)and to Eppendorf tubes (for the estimation of P concentra-tion); both contained 5 mL of concentrated HCl. Malic andcitric acids were analysed as described previously and P wasdetermined using the malachite green method (Motomizuet al. 1983).
Statistical analyses
Statistical analyses were performed using Genstat version9.2 (Lawes Agricultural Trust, Rothamsted ExperimentalStation, UK, 2007). For the Kennedia experiment, the effectof Kennedia species and inoculation with AMF on eachvariable was examined using general analyses of variance(ANOVAs). Each ANOVA included the factors Species,Inoculation and Block. The Species ¥ Inoculation interac-tion was always examined. Normality of data was checkedand transformations were undertaken if required. Outlierswere carefully checked. All figures present the Species ¥Inoculation interaction (mean � SE). If this interaction wassignificant (P < 0.05), then the LSD at P = 0.05 is also pre-sented. If there was no significant interaction, but Inocula-tion had a significant effect (P < 0.05), then marginal meansfor Inoculation are presented in Table 1.
RESULTS
Kennedia experiment
ObservationsAfter the addition of P at weeks 11 and 18, plants under-went a short period of rapid growth, with inoculated plantsoften becoming noticeably larger than uninoculated plants.This difference then declined over time.At the final harvest,many plants showed signs of nutrient deficiency, that is,yellowing of leaves. Root clusters, classified by Adams, Bell& Pate (2002) as ‘medium-dense’ and ‘short-dense’, wereobserved on less than 10 plants, while denser clusters of thetype common in some Australian native plants (Shane &Lambers 2005) were never observed. Large nodules werepresent, but their occurrence was variable, with number perplant ranging from 0 to 24. Consistently nodulated specieswere K. eximia, K. nigricans and K. prostrata I, while K. ru-bicunda and K. stirlingii were not nodulated.
Inoculation with AMF
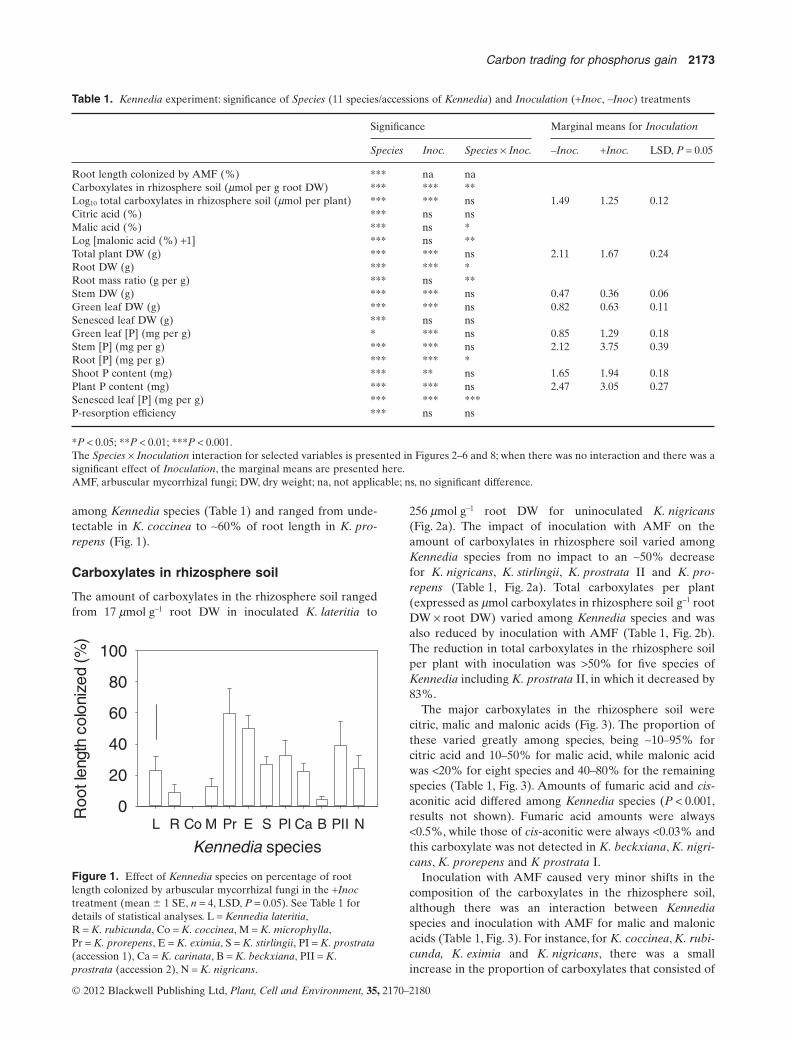
The uninoculated control plants were never colonized byAMF. Colonization by AMF in inoculated plants differed
among Kennedia species (Table 1) and ranged from unde-tectable in K. coccinea to ~60% of root length in K. pro-repens (Fig. 1).
Carboxylates in rhizosphere soil
The amount of carboxylates in the rhizosphere soil rangedfrom 17 mmol g-1 root DW in inoculated K. lateritia to
256 mmol g-1 root DW for uninoculated K. nigricans(Fig. 2a). The impact of inoculation with AMF on theamount of carboxylates in rhizosphere soil varied amongKennedia species from no impact to an ~50% decreasefor K. nigricans, K. stirlingii, K. prostrata II and K. pro-repens (Table 1, Fig. 2a). Total carboxylates per plant(expressed as mmol carboxylates in rhizosphere soil g-1 rootDW ¥ root DW) varied among Kennedia species and wasalso reduced by inoculation with AMF (Table 1, Fig. 2b).The reduction in total carboxylates in the rhizosphere soilper plant with inoculation was >50% for five species ofKennedia including K. prostrata II, in which it decreased by83%.
The major carboxylates in the rhizosphere soil werecitric, malic and malonic acids (Fig. 3). The proportion ofthese varied greatly among species, being ~10–95% forcitric acid and 10–50% for malic acid, while malonic acidwas <20% for eight species and 40–80% for the remainingspecies (Table 1, Fig. 3). Amounts of fumaric acid and cis-aconitic acid differed among Kennedia species (P < 0.001,results not shown). Fumaric acid amounts were always<0.5%, while those of cis-aconitic were always <0.03% andthis carboxylate was not detected in K. beckxiana, K. nigri-cans, K. prorepens and K prostrata I.
Inoculation with AMF caused very minor shifts in thecomposition of the carboxylates in the rhizosphere soil,although there was an interaction between Kennediaspecies and inoculation with AMF for malic and malonicacids (Table 1, Fig. 3). For instance, for K. coccinea, K. rubi-cunda, K. eximia and K. nigricans, there was a smallincrease in the proportion of carboxylates that consisted of
Table 1. Kennedia experiment: significance of Species (11 species/accessions of Kennedia) and Inoculation (+Inoc, -Inoc) treatments
Significance Marginal means for Inoculation
Species Inoc. Species ¥ Inoc. –Inoc. +Inoc. LSD, P = 0.05
Root length colonized by AMF (%) *** na naCarboxylates in rhizosphere soil (mmol per g root DW) *** *** **Log10 total carboxylates in rhizosphere soil (mmol per plant) *** *** ns 1.49 1.25 0.12Citric acid (%) *** ns nsMalic acid (%) *** ns *Log [malonic acid (%) +1] *** ns **Total plant DW (g) *** *** ns 2.11 1.67 0.24Root DW (g) *** *** *Root mass ratio (g per g) *** ns **Stem DW (g) *** *** ns 0.47 0.36 0.06Green leaf DW (g) *** *** ns 0.82 0.63 0.11Senesced leaf DW (g) *** ns nsGreen leaf [P] (mg per g) * *** ns 0.85 1.29 0.18Stem [P] (mg per g) *** *** ns 2.12 3.75 0.39Root [P] (mg per g) *** *** *Shoot P content (mg) *** ** ns 1.65 1.94 0.18Plant P content (mg) *** *** ns 2.47 3.05 0.27Senesced leaf [P] (mg per g) *** *** ***P-resorption efficiency *** ns ns
*P < 0.05; **P < 0.01; ***P < 0.001.The Species ¥ Inoculation interaction for selected variables is presented in Figures 2–6 and 8; when there was no interaction and there was asignificant effect of Inoculation, the marginal means are presented here.AMF, arbuscular mycorrhizal fungi; DW, dry weight; na, not applicable; ns, no significant difference.
Kennedia speciesL R Co M Pr E S PI Ca B PII N
Roo
t len
gth
colo
nize
d (%
)
0
20
40
60
80
100
Figure 1. Effect of Kennedia species on percentage of rootlength colonized by arbuscular mycorrhizal fungi in the +Inoctreatment (mean � 1 SE, n = 4, LSD, P = 0.05). See Table 1 fordetails of statistical analyses. L = Kennedia lateritia,R = K. rubicunda, Co = K. coccinea, M = K. microphylla,Pr = K. prorepens, E = K. eximia, S = K. stirlingii, PI = K. prostrata(accession 1), Ca = K. carinata, B = K. beckxiana, PII = K.prostrata (accession 2), N = K. nigricans.
malonic acid in response to inoculation with AMF, while forK. microphylla the proportion decreased.
Plant growth
Total plant DW differed among Kennedia species and wasdecreased by an average of 21% by inoculation with AMF(Table 1, Fig. 4a). For root DW, the impact of inoculationwith AMF varied among Kennedia species, from a 69%increase in K. lateritia to a >50% reduction in K. prostrata II
and K. microphylla (Table 1, Fig. 4b). For root mass ratio,the impact of inoculation also differed among Kennediaspecies (Table 1, Fig. 4c). While root mass ratio of manyspecies was little affected by inoculation with AMF, it wasgreatly increased in K. stirlingii and K. lateritia, and greatlydecreased in K. prostrata II. Stem and green leaf DW dif-fered among Kennedia species and both were decreased byan average of 23% by inoculation with AMF (Table 1, datanot shown). Senesced leaf DW differed among Kennediaspecies, but was unaffected by inoculation with AMF(Table 1, data not shown).
Plant P uptake
All P-uptake related parameters varied among Kennediaspecies and all, except P-resorption efficiency, were alsoaffected by inoculation with AMF (Table 1). For allKennedia species when not inoculated with AMF, greenleaf [P] was close to 1 mg g-1 and stem [P] was ~2 mg g-1
(Fig. 5a,b). Green leaf and stem [P] increased with inocula-tion by an average of 52 and 76%, respectively. For root [P],there was an interaction between Kennedia species andinoculation with AMF, with the impact of inoculationranging from not significant to a >130% increase in K. pro-repens, K. prostrata II, K. coccinea and K. carinata (Fig. 5c).There was no relationship between root [P] and the amountof carboxylates in rhizosphere soil or total carboxylates perplant in the rhizosphere soil (results not shown).
Car
boxy
late
s (µ
mol
g–
1 roo
t DW
)
0
50
100
150
200
250
300- Inoc+ Inoc
Kennedia species
L R Co M Pr E S PI Ca B PII N0
50
100
150
200
Car
boxy
late
s (µ
mol
pla
nt–
1 )
(a)
(b)
Figure 2. Effect of Kennedia species and inoculation witharbuscular mycorrhizal fungi on (a) amount of carboxylates inrhizosphere soil and (b) total carboxylates in the rhizosphere soilper plant (mean � 1 SE, n = 4). The LSD at P = 0.05 is providedwhen there is an interaction between Species and Inoculation.Three outliers were removed in each of (a) and (b); the value ofthese in (a) were Co -Inoc = 439, Co +Inoc = 166, N -Inoc = 772.See Table 1 for details of statistical analyses. L = Kennedialateritia, R = K. rubicunda, Co = K. coccinea, M = K. microphylla,Pr = K. prorepens, E = K. eximia, S = K. stirlingii, PI = K. prostrata(accession 1), Ca = K. carinata, B = K. beckxiana, PII = K.prostrata (accession 2), N = K. nigricans. DW, dry weight.
Kennedia species (–Inoc, +Inoc)
L-IL+
IR-I
R+ICo-
ICo+
IM-I
M+IPr-IPr+
IE-I
E+I S-IS+IPI-I
PI+ICa-ICa+
IB-I
B+IPII-
IPII+
IN-I
N+I
Com
posi
tion
(%)
0
20
40
60
80
100
Citric acid
Malic acid
Malonic acid
Figure 3. Effect of Kennedia species and inoculation witharbuscular mycorrhizal fungi on the proportion of carboxylates inthe rhizosphere soil consisting of malonic, malic and citric acid(mean, n = 4). Combined, the proportion of iso-citric, shikimic,lactic, acetic, maleic, succinic, fumaric, cis-aconitic andtrans-aconitic acid was always less than 0.25% and these datahave not been included. See Table 1 for details of statisticalanalyses. L = Kennedia lateritia, R = K. rubicunda,Co = K. coccinea, M = K. microphylla, Pr = K. prorepens,E = K. eximia, S = K. stirlingii, PI = K. prostrata (accession 1),Ca = K. carinata, B = K. beckxiana, PII = K. prostrata (accession2), N = K. nigricans. -I = not inoculated, +I = Inoculated.
Shoot P content differed among Kennedia species andincreased on average by 18% when plants were inoculatedwith AMF (Table 1). Total plant (shoot plus root) Pcontent also differed among Kennedia species andincreased, on average, by 23% when plants were inocu-lated with AMF (Table 1, Fig. 6). This average increasereflected a large increase in a few species, notably K. lat-eritia, K. coccinea and K. eximia. There was a strong posi-tive logarithmic relationship between total plant P contentand the amount of carboxylates in rhizosphere soil forplants that were not inoculated (Fig. 7a). A similar, butstronger, relationship was evident between plant P contentand total carboxylates per plant in the rhizosphere soil forboth uninoculated and inoculated plants (Fig. 7b), withinoculated plants consistently having around 0.75 mg ofadditional P for a given amount of total carboxylates.There was no relationship between plant P content andeither malic acid or malonic acid (R2 < 0.14). However,
there was a positive logarithmic relationship between plantP content and the amount of citric in rhizosphere soil (notinoculated r2 = 0.46, P = 0.01; inoculated r2 = 0.44, P = 0.02)and, in particular, total citric acid in the rhizosphere soilper plant (not inoculated r2 = 0.73, P = 0.01; inoculatedr2 = 0.56, P = 0.005).
For senesced leaf [P], there was an interaction betweenKennedia species and inoculation with AMF, with inocula-tion causing large increases in only a few species (Table 1,Fig. 8a). Treatments with the highest senesced leaf [P], thatis, inoculated K. coccinea, K. stirlingii and K. beckxiana,were those that had the highest leaf [P] (Fig. 5a).P-resorption efficiency varied among Kennedia species andwas not affected by inoculation (Table 1, Fig. 8b).K. stirlingii, K. coccinea and K. beckxiana had the lowest
Tot
al p
lant
DW
(g)
0
1
2
3
Roo
t DW
(g)
0.0
0.2
0.4
0.6
0.8
1.0
Kennedia speciesL R Co M Pr E S PI Ca B PII N
Roo
t mas
s ra
tio (
g g
–1 )
0.0
0.1
0.2
0.3
0.4
0.5
– Inoc+ Inoc
(a)
(b)
(c)
Figure 4. Effect of Kennedia species and inoculation witharbuscular mycorrhizal fungi on (a) total plant DW, (b) root DWand (c) root mass ratio [mean � 1 SE, n = 4, 1 outlier removed in(b) and (c)]. The LSD at P = 0.05 is provided when there is aninteraction between Species and Inoculation. See Table 1 fordetails of statistical analyses. L = Kennedia lateritia,R = K. rubicunda, Co = K. coccinea, M = K. microphylla,Pr = K. prorepens, E = K. eximia, S = K. stirlingii, PI = K. prostrata(accession 1), Ca = K. carinata, B = K. beckxiana, PII = K.prostrata (accession 2), N = K. nigricans. DW, dry weight.
Ste
m P
(m
g g–
1 )
0
2
4
6
Kennedia species
L R Co M Pr E S PI Ca B PII N
Roo
t P (
mg
g–1 )
0
1
2
3
4
Gre
en le
af P
(m
g g–
1 )
0
1
2
3
4 – Inoc+ Inoc
(a)
(b)
(c)
Figure 5. Effect of Kennedia species and inoculation witharbuscular mycorrhizal fungi on (a) green leaf [P], (b) stem [P]and (c) root [P] [mean � 1 SE, n = 4, two outliers removed in (a)and one in (c)]. The LSD at P = 0.05 is provided when there is aninteraction between Species and Inoculation. See Table 1 fordetails of statistical analyses. L = Kennedia lateritia,R = K. rubicunda, Co = K. coccinea, M = K. microphylla,Pr = K. prorepens, E = K. eximia, S = K. stirlingii, PI = K. prostrata(accession 1), Ca = K. carinata, B = K. beckxiana, PII = K.prostrata (accession 2), N = K. nigricans.
P-resorption efficiencies at ~60%, while K. lateritia, K. pros-trata I and K. prostrata II had the highest at >90%.
Carboxylate dynamics in washed river sand
When 200 mm citric acid was added to the sand, citric acid inthe soil solution decreased only slightly over 2 h witharound 95% remaining when chloroform was added and90% remaining when chloroform was not added (Fig. 9a).In view of this response, 20 min of incubation was used inthe subsequent experiment. Loss of citric and malic acidfrom the soil solution after 20 min followed a linear rela-tionship with concentration (Fig. 9b). When 200 mm citricacid was added to soil, ~25–30% was lost from the soilsolution; losses were minimal at concentrations >200 mm.Malic acid had no effect on the concentration of P in the soilsolution after 20 min (Fig. 9c). However, an addition of5–10 mm citric acid increased the concentration of P in the
soil solution by ~0.10 mg kg-1 and an addition of 100–1000 mm citric acid increased it by ~0.15 mg kg-1.
DISCUSSION
Our results support the hypothesis that Kennedia specieswill reduce the amount of carboxylates in rhizosphere soil ifan additional carbon drain due to inoculation with AMF ispresent. Because inoculation increased plant P content, italso appears that inoculation resulted in plants gaining agreater proportion of their P from labile inorganic P. Theimplications of these findings and other points of interestare discussed next.
Carboxylates in rhizosphere soil and theireffects on total plant P
The amount of carboxylate in the rhizosphere soil was>50 mmol g-1 root DW for five species of Kennedia andreached 256 mmol g-1 root DW for K. nigricans. These car-boxylate concentrations are consistent with previous glass-house studies of Australian and exotic legumes grown inlow-P sand (Pearse et al. 2006; Pang et al. 2010a). Veneklaaset al. (2003) reported carboxylates in rhizosphere soil to beup to ~200 mmol g-1 root DW for chickpea (Cicer arietinumL.) and up to ~275 mmol g-1 root DW for white lupin
Kennedia species
L R Co M Pr E S PI Ca B PII N
Pla
nt P
con
tent
(m
g)
0
1
2
3
4
5– Inoc+ Inoc
Figure 6. Effect of Kennedia species and inoculation witharbuscular mycorrhizal fungi on plant (root plus shoot) P content(mean � 1 SE, n = 4, two outliers removed). There was nointeraction between Species and Inoculation. See Table 1 fordetails of statistical analyses. L = Kennedia lateritia,R = K. rubicunda, Co = K. coccinea, M = K. microphylla,Pr = K. prorepens, E = K. eximia, S = K. stirlingii, PI = K. prostrata(accession 1), Ca = K. carinata, B = K. beckxiana, PII = K.prostrata (accession 2), N = K. nigricans.
(a)
Log carboxylates (µmol g–1 root DW)
1.0 1.4 1.8 2.2
Pla
nt P
con
tent
(m
g)
1
2
3
4
5
Log carboxylates (µmol plant–1)
0.6 1.0 1.4 1.8 2.2
– Inoc+ Inoc
(b)
Figure 7. Relationship between plant (root plus shoot) Pcontent and (a) amount of carboxylates in rhizosphere soil (notinoculated r2 = 0.51, P = 0.009; inoculated r2 = 0.38, P > 0.05) (b)total carboxylate in the rhizosphere soil per plant (not inoculatedr2 = 0.49, P = 0.01; inoculated r2 = 0.53, P = 0.008). DW, dry weight.
Kennedia species
L R Co M Pr E S PI Ca B PII N
P-r
esor
ptio
n ef
ficie
ncy
(%)
40
60
80
100
Sen
esce
dle
af P
(m
g g–
1 )
0.0
0.2
0.4
0.6
0.8
1.0
1.2 – Inoc+ Inoc
(a)
(b)
Figure 8. Effect of Kennedia species and inoculation witharbuscular mycorrhizal fungi on (a) senesced leaf [P] and (b)P-resorption efficiency (mean � 1 SE, n = 4). The LSD at P = 0.05is provided when there is an interaction between Species andInoculation. See Table 1 for details of statistical analyses.L = Kennedia lateritia, R = K. rubicunda, Co = K. coccinea,M = K. microphylla, Pr = K. prorepens, E = K. eximia,S = K. stirlingii, PI = K. prostrata (accession 1), Ca = K. carinata,B = K. beckxiana, PII = K. prostrata (accession 2),N = K. nigricans.
(Lupinus albus L.), the latter with considerable cluster-rootformation, grown in a range of soils.
Citric, malic and malonic acids were the main carboxy-lates in the rhizosphere of the Kennedia species examined,and all are known to aid plants to access strongly sorbed soilP (Jones 1998; Veneklaas et al. 2003; Oburger et al. 2009).However, in our experiment the driver for the positive rela-tionship between total plant P content and the amount of
carboxylates (Fig. 7) was citric acid. Support for this rela-tionship being causal comes from several sources. Firstly, inthe carboxylate dynamics experiments, only a low propor-tion of added citric acid was lost from the soil solutionirrespective of concentration (Fig. 9a,b) and 0.10–0.15 mg kg-1 of P was released into the soil solution over20 min when citric acid was added (Fig. 9c). Secondly, thelogarithmic nature of the relationship between total plant Pcontent and rhizosphere carboxylates (Fig. 7) is consistentwith concentrations of added citric acid as low as 10 mmresulting in significant P desorption into the soil solution,while little change in the amount of P desorbed into the soilsolution occurred when 100–1000 mm of citric acid wasadded (Fig. 9c). However, this could also reflect plants accu-mulating P, perhaps due to N deficiency limiting growth dueto poor nodulation. Finally, a positive relationship betweenplant P content and amount of rhizosphere carboxylateswas also found by Veneklaas et al. (2003) for chickpea, withplant P content higher for a given amount of carboxylatewhen soils had a lower P retention index (i.e. <10). The Pretention index of the washed river sand used in the currentstudy was very low at 1.5.
While malonic acid did not appear to influence plant Puptake, it is worth noting that it is a strong inhibitor ofmitochondrial (Ikuma & Bonner 1967) and bacterial(Hirsch et al. 1963) respiration. High concentrations may beimportant for processes such as enabling build-up of highconcentrations of citrate prior to its release in an exudativeburst (Playsted et al. 2006), but so far experimental evi-dence to support this is lacking (Oburger et al. 2009).
Inoculation with AMF reduced the amount ofcarboxylate in the rhizosphere soil, yetincreased plant P uptake
The impact of inoculation with AMF on the amount ofcarboxylate in rhizosphere soil varied among Kennediaspecies, from no impact to around a ~50% decrease for fourspecies. The large reduction in total carboxylate in rhizo-sphere soil per plant with inoculation reflects a large declinein root DW or carboxylate concentration or both. In bothcases, the reduction was particularly notable for specieswith the highest amounts of carboxylate in rhizosphere soil(Fig. 2). The mechanism for the reduction in rhizospherecarboxylates with inoculation is unknown. It could be adirect effect of less carbon availability or a result of higherplant [P] in inoculated plants (Fig. 5). Increased P availabil-ity tends to reduce the amount of carboxylate in rhizo-sphere soil (Pearse et al. 2006; Pang et al. 2010a) and adecrease in root exudates (reducing sugars and aminoacids) of Sorghum vulgare Pers. with inoculation corre-sponded to an increase in root [P] and a decrease in rootmembrane permeability (Graham et al. 1981). However, inthe present experiment there was no relationship betweenroot [P] and amount of carboxylates.
Inoculation with AMF increased average plant P content(Table 1) and consistently supplied an additional ~0.75 mgof P for a given amount of carboxylates in rhizosphere soil
0
0.05
0.1
0.15
0.2
0.25
0.3
0 1 5 10 20 50 100 200 5001000
Acid concentration (µM)
Citric acidMalic acidWater
0.50
0.60
0.70
0.80
0.90
1.00
0 30 60 90 120
Time (min)
Citr
ic a
cid
rem
aini
ng in
soi
l sol
utio
n (%
)
Chl – Chl + Mean
0
50
100
150
200
0 100 200
0
200
400
600
800
1000
200 400 600 800 1000
Citric acidMalic acid
Fin
al c
once
ntra
tion
of a
cid
in s
oil s
olut
ion
(µM
)
P in
soi
l sol
utio
n (m
g kg
–1)
Initial concentration of acid added (µM)
1:1
1:1
(a)
(b)
(c)
Figure 9. (a) The proportion of added citric acid (200 mm)remaining in the soil solution of washed river sand over 120 mineither without (Chl -) or with (Chl +) the addition of chloroformto remove soil microbes (mean � 1 SE, n = 6). (b) Therelationship between the concentrations in the soil solution ofcitric and malic acid, and the concentration of these acids addedto washed river sand after 20 min (no added chloroform)(mean � 1 SE, n = 4, the 1:1 relationship is also shown). (c) Theconcentration of P in the soil solution of washed river sand20 min after a range of concentrations of citric and malic acidwas added (no added chloroform) (mean � 1 SE, n = 4, a watercontrol is also shown).
(Fig. 7).These trends strongly reflect large increases in totalplant P with inoculation in Kennedia species with lowamounts of carboxylates (Fig. 6). In a soil higher in stronglysorbed P, but with a similar low level of labile inorganic P,perhaps the impact of inoculation with AMF on plant Pcontent would be less, and maybe even negative, due toreduction in rhizosphere carboxylates.
An increase in rhizosphere carboxylates when AMF isabsent, that is, a shift towards greater use of strongly sorbedsoil P, may confer advantages under field conditions forperennial species in low-P environments. Perennials arelikely to be highly colonized by AMF in the cooler, wetterwinter months (Denton et al. 2006; Tibbett & Cairney 2007;Tibbett et al. 2008). However, colonization might be low orabsent in summer, especially in Mediterranean climates,due to drought (Armstrong, Helyar & Christie 1992; Ryan& Ash 1996) and high soil temperatures (Braunberger,Abbott & Robson 1997). Drought also reduces movementof P through soil to roots and decreases the release ofinorganic P from P bound in organic matter, and therebyreduces plant P uptake (Sardans & Peñuelas 2007). Undersuch circumstances, if plants are not dormant, enhancedexudation of carboxylates might be advantageous. Indeed,Suriyagoda et al. (2010) recently observed increasedamounts of carboxylates in rhizosphere soil under dry con-ditions for perennial legumes grown without AMF. Theimplications of the phenotypic plasticity shown by theKennedia species now require investigation under field con-ditions; it may confer a competitive advantage over coex-isting species (Ashton et al. 2010).
Kennedia coccinea was not colonized by AMF, but itsresponse to inoculation with AMF was similar to that ofother species; a small decrease in plant DW and a largeincrease in P uptake. We have no explanation for thisanomaly, but note that similar situations have beenreported elsewhere (Fonseca, Berbara & Daft 2001;O’Connor, Smith & Smith 2002). Dennett et al. (2011)found that inoculation of Solanum centrale, an Australiandesert sub-shrub, growing in coarse river sand caused sub-stantial colonization and a large increase in growth when noP was added. However, under low- and high-P conditions,inoculation resulted in no colonization, but root [P] wasincreased and the root–shoot ratio decreased. Dennett et al.(2011) discuss possible mechanisms for this phenomenon.As this is not the focus of the current paper, readers arereferred to Dennett et al. (2011) for more information.
Inoculation with AMF has little impact on rootmass ratio
There was no consistent change in root mass ratio withinoculation. A decrease in root mass ratio in response toinoculation with AMF is often reported for annual cropspecies, that is, relatively short-lived species adapted tomoist environments (Gavito et al. 2000; Guo et al. 2006).Such species have generally been domesticated in prehis-toric times and may have increased their root mass ratioduring the domestication process (Weeden 2007). Many
Australian perennial legumes have a relatively low rootmass ratio and an even distribution of roots with depth; thismay enhance their performance in environments both lowin P and prone to water stress (Denton et al. 2006; Suriya-goda et al. 2010; Pang et al. 2011).Thus, it may not be advan-tageous for plants such as Kennedia to reduce their rootmass ratio in response to colonization by AMF.
P-resorption proficiency and efficiency
Very high P-resorption proficiency (i.e. low senesced leaf[P]) and P-resorption efficiency in Kennedia was an unex-pected finding of our study (Fig. 7). In their natural habitatof low-P soils, woody Australian perennials in the genusBanksia have less variable and often higher P-resorptionproficiencies than did the Kennedia species (Dentonet al. 2007), but some species in both studies had veryhigh P-resorption proficiencies indeed (i.e. senesced leaf[P] < 0.1 mg g-1). P-resorption efficiency reached 85–99%for six Kennedia species. Such high values are unusual, yetconsistent with our assumption that many Kennedia speciesare highly adapted to very low-P environments (Lamberset al. 2010). The species with the highest senesced leaf [P]had the lowest P-resorption efficiencies, suggesting that Premobilization is responsive to plant P status. The Banksiaspecies studied by Denton et al. (2007) had P-resorptionvalues of 27–82%. These values are lower than in thecurrent study, primarily due to much lower green leaf [P](~0.2 mg g-1) for the Banksia species.
In conclusion, this study is the first to show that inocula-tion with AMF can reduce the amount of carboxylates inrhizosphere soil. However, while the concentration of citricacid in the rhizosphere was positively correlated with plantP uptake, inoculation also generally increased plant Puptake. Thus, mycorrhizally mediated partitioning of thesoil P resource appeared to occur, as inoculated plants pre-sumably accessed a smaller proportion of their P fromstrongly sorbed soil P sources. The presence among theseperennial Kennedia species of such phenotypic plasticity inrelation to soil P resource use may represent adaptation toseasonal changes in soil P resources and the ability of AMFto colonize roots.
ACKNOWLEDGMENTS
This research was supported through an ARC-linkage grantfrom the Australian Research Council in partnership withthe Department of Agriculture and Food Western Australia,Heritage Seeds, the Chemistry Centre of Western Australia,the Facey Group and the Mingenew Irwin Group. L.D.B.S.appreciates the SIRF/UIS Scholarship awarded by TheUniversity of Western Australia and further scholarshipsupport from the late Frank Ford.
REFERENCES
Adams M.A., Bell T.L. & Pate J.S. (2002) Phosphorus sources andavailability modify growth and distribution of root clusters and
nodules of native Australian legumes. Plant, Cell & Environment25, 837–850.
Allen D.C. & Jeffery R.C. (1990) Methods for the Analysis ofPhosphorus in Western Australian Soils. Chemistry Centre ofWestern Australia, Perth,WA,Australia. Report on InvestigationNo 37.
Armstrong R.D., Helyar K.R. & Christie E.K. (1992) Vesicular-arbuscular mycorrhiza in semi-arid pastures of south-eastQueensland and their effect on growth responses to phosphorusfertilisers by grasses. Australian Journal of Agricultural Research43, 1143–1155.
Ashton I.W., Miller A.E., Bowman W.D. & Suding K.N. (2010)Niche complementarity due to plasticity in resource use: plantpartitioning of chemical N forms. Ecology 91, 3252–3260.
Bever J.D., Dickie I.A., Facelli E., Facelli J.M., Klironomos J.,Moora M., Rillig M.C., Stock W.D., Tibbett M. & Zobel M.(2010) Rooting theories of plant community ecology in micro-bial interactions. Trends in Ecology & Evolution 25, 468–478.
Braunberger P.G., Abbott L.K. & Robson A.D. (1997) Earlyvesicular-arbuscular mycorrhizal colonisation in soil collectedfrom an annual clover-based pasture in a Mediterranean envi-ronment: soil temperature and the timing of autumn rains.Australian Journal of Agricultural Research 48, 103–110.
Brundrett M.C. (2009) Mycorrhizal associations and other meansof nutrition of vascular plants: understanding the global diversityof host plants by resolving conflicting information and develop-ing reliable means of diagnosis. Plant and Soil 320, 37–77.
Cawthray G.R. (2003) An improved reversed-phase liquid chro-matographic method for the analysis of low-molecular massorganic acids in plant root exudates. Journal of ChromatographyA 1011, 233–240.
Colwell J.D. (1963) The estimation of the phosphorus fertiliserrequirements of wheat in southern NSW by soil analysis. Aus-tralian Journal of Experimental Agriculture and Animal Hus-bandry 3, 190–197.
Dennett A.L., Burgess L.W., McGee P.A. & Ryder M.H. (2011)Arbuscular mycorrhizal associations in Solanum centrale (bushtomato), a perennial sub-shrub from the arid zone of Australia.Journal of Arid Environments 75, 688–694.
Denton M.D., Sasse C., Tibbett M. & Ryan M.H. (2006) Rootdistributions of Australian herbaceous perennial legumes inresponse to phosphorus placement. Functional Plant Biology 33,1091–1102.
Denton M.D., Veneklaas E.J., Freimoser F.M. & Lambers H. (2007)Banksia species (Proteaceae) from severely phosphorus-impoverished soils exhibit extreme efficiency in the use andre-mobilization of phosphorus. Plant, Cell & Environment 30,1557–1565.
Dinkelaker B., Römheld V. & Marschner H. (1989) Citric acidexcretion and precipitation of calcium citrate in the rhizosphereof white lupin (Lupinus albus L.). Plant, Cell & Environment 12,285–292.
FloraBase (2010) The Western Australian Flora. http://www.florabase.dec.wa.gov.au/ (accessed 28 July 2010).
Fonseca H., Berbara R.L.L. & Daft M.J. (2001) Shoot delta N-15and delta C-13 values of non-host Brassica rapa change whenexposed to +/- Glomus etunicatum inoculum and three levels ofphosphorus and nitrogen. Mycorrhiza 11, 151–158.
Friesen M.L., Porter S.S., Stark S.C., von Wettberg E.J., Sachs J.L. &Martinez-Romero E. (2011) Microbially mediated plant func-tional traits. Annual Review of Ecology, Evolution, and System-atics 42, 23–46.
Gavito M.E., Curtis P.S., Mikkelsen T.N. & Jakobsen I. (2000)Atmospheric CO2 and mycorrhiza effects on biomass allocationand nutrient uptake of nodulated pea (Pisum sativum L.) plants.Journal of Experimental Botany 51, 1931–1938.
Giovannetti M. & Mosse B. (1980) An evaluation of techniques formeasuring VAM infection in roots. New Phytologist 84, 489–500.
Graham J.H. (2000) Assessing costs of arbuscular mycorrhizal sym-biosis in agroecosystems. In Current Advances in MycorrhizaeResearch (eds G.K. Podila & D.D. Douds), pp. 111–126. APSPress, St Paul, MN, USA.
Graham J.H., Leonard R.T. & Menge J.A. (1981) Membrane-mediated decrease in root exudation responsible for phosphorusinhibition of vesicular-arbuscular mycorrhiza formation. PlantPhysiology 68, 548–552.
Guo T., Zhang J.L., Christie P. & Li X.L. (2006) Influence of nitro-gen and sulfur fertilizers and inoculation with arbuscular mycor-rhizal fungi on yield and pungency of spring onion. Journal ofPlant Nutrition 29, 1767–1778.
Hirsch C.A., Rasminsky M., Davis B.D. & Lin E.C.C. (1963) Afumarate reductase in Escherichia coli distinct from succinatedehydrogenase. Journal of Biological Chemistry 238, 3770–3774.
Ikuma H. & Bonner W.D. Jr (1967) Properties of higher plantmitochondria. III. Effects of respiratory inhibitors. Plant Physi-ology 42, 1535–1544.
Jakobsen I., Abbott L.K. & Robson A.D. (1992) External hyphaeof vesicular-arbuscular mycorrhizal fungi associated with Trifo-lium subterraneum L. 1 Spread of hyphae and phosphorus inflowinto roots. New Phytologist 120, 371–380.
Jones D.L. (1998) Organic acids in the rhizosphere – a criticalreview. Plant and Soil 205, 25–44.
Jones D.L. & Brassington D.S. (1998) Sorption of organic acids inacid soils and its implications in the rhizosphere. EuropeanJournal of Soil Science 49, 447–455.
Lambers H., Brundrett M.C., Raven J.A. & Hopper S.D. (2010)Plant mineral nutrition in ancient landscapes: high plant speciesdiversity on infertile soils is linked to functional diversity fornutritional strategies. Plant and Soil 334, 11–31.
Lambers H., Bishop J.G., Hopper S.D., Laliberté E. & Zúñiga-FeestA. (2012) Phosphorus-mobilisation ecosystem engineering: theroles of cluster roots and carboxylate exudation in youngP-limited ecosystems. Annals of Botany in press.
Mada R.J. & Bagyaraj D.J. (1993) Root exudation from Leucaenaleucocephala in relation to mycorrhizal colonisation. WorldJournal of Microbiology & Biotechnology 9, 342–344.
Motomizu S., Wakimoto T. & Toei K. (1983) Spectrophotometricdetermination of phosphate in river waters with molybdate andmalachite green. The Analyst 108, 361–367.
O’Connor P.J., Smith S.E. & Smith F.A. (2002) Arbuscular mycor-rhizas influence plant diversity and community structure in asemiarid herbland. New Phytologist 154, 209–218.
Oburger E., Kirk G.J.D., Wenzel W.W., Puschenreiter M. & JonesD.L. (2009) Interactive effects of organic acids in the rhizo-sphere. Soil Biology & Biochemistry 41, 449–457.
Pang J., Ryan M.H., Tibbett M., Cawthray G.R., Siddique K.H.M.,Bolland M.D.A., Denton M.D. & Lambers H. (2010a) Variationin morphological and physiological parameters in herbaceousperennial legumes in response to phosphorus supply. Plant andSoil 331, 241–255.
Pang J., Tibbett M., Denton M.D., Lambers H., Siddique K.H.M.,Bolland M.D.A., Revell C.K. & Ryan M.H. (2010b) Variation inseedling growth of 11 perennial legumes in response to phospho-rus supply. Plant and Soil 328, 133–143.
Pang J., Yang J., Ward P., Siddique K.H.M., Lambers H., Tibbett M.& Ryan M. (2011) Contrasting responses to drought stress inherbaceous perennial legumes. Plant and Soil 348, 299–314.
Pearse S.J., Veneklaas E.J., Cawthray G.R., Bolland M.D.A. &Lambers H. (2006) Carboxylate release of wheat, canola and 11grain legume species as affected by phosphorus status. Plant andSoil 288, 127–139.
PlantNET (2010) NSW Flora Online. http://plantnet.rbgsyd.nsw.gov.au/ (accessed 28 July 2010).
Playsted C.W.S., Johnston M.E., Ramage C.M., Edwards D.G., Caw-thray G.R. & Lambers H. (2006) Functional significance ofdauciform roots: exudation of carboxylates and acid phos-phatase under phosphorus deficiency in Caustis blakei (Cyper-aceae). New Phytologist 170, 491–500.
Rayment G.E. & Higginson F.R. (1992) Australian LaboratoryHandbook of Soil and Water Chemical Methods. Inkata Press,Melbourne, Vic., Australia.
Ryan M.H. & Angus J.F. (2003) Arbuscular mycorrhizae in wheatand field pea crops on a low P soil: increased Zn-uptake but noincrease in P-uptake or yield. Plant and Soil 250, 225–239.
Ryan M.H. & Ash J.E. (1996) Colonisation of wheat in southernNSW by VA-mycorrhizal (VAM) fungi is significantly reducedby drought. Australian Journal of Experimental Agriculture 36,563–569.
Sardans J. & Peñuelas J. (2007) Drought changes phosphorus andpotassium accumulation patterns in an evergreen Mediterraneanforest. Functional Ecology 21, 191–201.
Schüßler A. & Walker C. (2010) The Glomeromycota: A SpeciesList with New Families and Genera. A Schüßler & C. Walker,Gloucester, UK.
Searle P.L. (1984) The berthelot or indophenol reaction and its usein the analytical chemistry of nitrogen. The Analyst 109, 549–568.
Shane M.W. & Lambers H. (2005) Cluster roots: a curiosity incontext. Plant and Soil 274, 101–125.
Stockinger H., Walker C. & Schüßler A. (2009) Glomus intraradi-ces DAOM197198′, a model fungus in arbuscular mycorrhiza
research, is not Glomus intraradices. New Phytologist 183, 1176–1187.
Suriyagoda L.D.B., Ryan M.H., Renton M. & Lambers H. (2010)Multiple adaptive responses of Australian native perenniallegumes with pasture potential to grow in phosphorus- andmoisture-limited environments. Annals of Botany 105, 755–767.
Tibbett M. & Cairney J.W.G. (2007) The cooler side of mycorrhizas:their occurrence and functioning at low temperatures. CanadianJournal of Botany 85, 51–62.
Tibbett M., Ryan M.H., Barker S.J., Chen Y.L., Denton M.D.,Edmonds-Tibbett T. & Walker C. (2008) The diversity of arbus-cular mycorrhizas of selected Australian Fabaceae. Plant Biosys-tems 142, 420–427.
Van Heerwaarden L.M., Toet S. & Aerts R. (2003) Nitrogen andphosphorus resorption efficiency and proficiency in six sub-arcticbog species after 4 years of nitrogen fertilization. Journal ofEcology 91, 1060–1070.
Veneklaas E.J., Stevens J., Cawthray G.R., Turner S., Grigg A.M. &Lambers H. (2003) Chickpea and white lupin rhizosphere car-boxylates vary with soil properties and enhance phosphorusuptake. Plant and Soil 248, 187–197.
Vierheilig H., Coughlan A.P., Wyss U. & Piche Y. (1998) Ink andvinegar, a simple staining technique for arbuscular-mycorrhizalfungi. Applied and Environmental Microbiology 64, 5004–5007.
Weeden N.F. (2007) Genetic changes accompanying the domesti-cation of Pisum sativum: is there a common genetic basis to the‘Domestication Syndrome’ for Legumes? Annals of Botany 100,1017–1025.
Yao Q., Li X.L., Feng G. & Christie P. (2001) Mobilization ofsparingly soluble inorganic phosphates by the external myceliumof an abuscular mycorrhizal fungus. Plant and Soil 230, 279–285.
Received 2 February 2011; accepted for publication 9 May 2012