CELLULAR IMMUNOLOGY 114,257-27 1 (1988) CD8+ T Cells Lyse Autologous Monocytes in the Presence of Anti-CD3 Monoclonal Antibody: Association with Interleukin-1 Production’ KYOGO ITOH,*~~,’ CHARLES M. BALCH,* JOANN TRIAL,? AND CHRIS D. PLATSOUCAS~ Division of Surgery, *Department of General Surgery and the tDepartment ofImmunology, The University of Texas System Cancer Center, M. D. Anderson Hospital and Tumor Institute, Houston, Texas 77030 Received May IS, 1987; acceptedFebruary 19,1988 Human peripheral blood T cells lysed autologous or allogeneic monocytes, but not polymor- phonuclear cells (PMN) or lymphocytes, in the presence of anti-CD3 (OKT3 (IgG2,) or anti- Leu 4 (IgG,)) monoclonal antibody (mAb). Other mAbs such as OKT4 (IgG& OKT8 (IgG&, OKTl 1 (IgG,), and OKM 1 (IgGrJ did not mediate lysis of monocytes. Lysis of monocytes also did not occur in the presence of F(ab’)r fragments of OKT3 mAb. OKT3 mAb and control murine IgG2. mAb, but not F(ab’)r fragments of OKT3 mAb, were bound to the monocyte cell surface. Purified human IgG, and IgGr myeloma proteins, polyclonal human IgG, or Con A inhibited anti-CD3-dependent T-cell cytotoxicity against monocytes when added to the 4-hr ?Zr-release assay. Pretreatment of monocytes with an irrelevant murine IgG2. mAb also inhib- ited OKT3 mAb (&G&dependent lysis of these cells, but did not affect anti&u 4 mAb (IgG,)- dependent lysis, suggesting that two different Fc receptors were involved. These results strongly suggest that Fc IgG receptors on monocytes are a critical structure for anti-CD3dependent cytotoxicity. Lysis of monocytes was accompanied by interleukin-1 (ILl) production, which was detected in supematants from 4-hr cultures of T cells and monocytes in the presence of the OKT3 mAb. Both anti-CD3-dependent lysis of monocytes and IL- 1 production were severely decreased after treatment of T cells with either OKT3 or OKT8 mAb plus complement, but were not affected significantly by treatment with the OKT4 mAb plus complement. Purified CD8+ cells, prepared using the cell sorter, exhibited significant levels of anti-CD3dependent monocyte lysis (> 10%).In contrast, purified CD4+ cells did not exhibit significant levels of anti- CD3-dependent cytotoxicity (i 10%). Production of high concentrations of IL-l was observed in cultures of purified CD8+ cells and monocytes in the presence of anti-CD3 mAb. Only low concentrations of IL-1 were detected in cultures of purified CD4+ cells, monocytes, and OKT3 mAb. These results suggest that CD8+ cells are primarily responsible for lysis of monocytes, which is associated with IL-1 production. It appears that anti-CD3 mAb brings CD8+ T cells and monocytes into close proximity by binding to the CD3 antigen on T cells and to the Fc IgG receptor on monocytes. This interaction results in lysis of monocytes primarily by CD8+ cells, after bypassing any antigen recognition requirements that may be othetwise needed. Lysis of monocytes appears to be associated with IL- 1 release. o 1988 Academic FWSS, IIIC. ’ This work was supported in part by Grants CA-27 197, CA- 13148, CA-030 13, and CA-4 1699 from the National Institutes of Health, Grants IM-409D and CH-420 from the American Cancer Society, by a grant from the Eleanor Naylor Dana Charitable Trust, and by a grant from the Erwin Zaban Melanoma Research Fund. 257 0008-8749/88$3.00 Copyright 0 1988 by Academic F’ress, Inc. All rights of reproduction in any form reserved.
Transcript
CELLULAR IMMUNOLOGY 114,257-27 1 (1988)
CD8+ T Cells Lyse Autologous Monocytes in the Presence of Anti-CD3 Monoclonal Antibody: Association
with Interleukin-1 Production’
KYOGO ITOH,*~~,’ CHARLES M. BALCH,* JOANN TRIAL,? AND CHRIS D. PLATSOUCAS~
Division of Surgery, *Department of General Surgery and the tDepartment ofImmunology, The University of Texas System Cancer Center, M. D. Anderson Hospital
and Tumor Institute, Houston, Texas 77030
Received May IS, 1987; accepted February 19,1988
Human peripheral blood T cells lysed autologous or allogeneic monocytes, but not polymor- phonuclear cells (PMN) or lymphocytes, in the presence of anti-CD3 (OKT3 (IgG2,) or anti- Leu 4 (IgG,)) monoclonal antibody (mAb). Other mAbs such as OKT4 (IgG& OKT8 (IgG&, OKTl 1 (IgG,), and OKM 1 (IgGrJ did not mediate lysis of monocytes. Lysis of monocytes also did not occur in the presence of F(ab’)r fragments of OKT3 mAb. OKT3 mAb and control murine IgG2. mAb, but not F(ab’)r fragments of OKT3 mAb, were bound to the monocyte cell surface. Purified human IgG, and IgGr myeloma proteins, polyclonal human IgG, or Con A inhibited anti-CD3-dependent T-cell cytotoxicity against monocytes when added to the 4-hr ?Zr-release assay. Pretreatment of monocytes with an irrelevant murine IgG2. mAb also inhib- ited OKT3 mAb (&G&dependent lysis of these cells, but did not affect anti&u 4 mAb (IgG,)- dependent lysis, suggesting that two different Fc receptors were involved. These results strongly suggest that Fc IgG receptors on monocytes are a critical structure for anti-CD3dependent cytotoxicity. Lysis of monocytes was accompanied by interleukin-1 (ILl) production, which was detected in supematants from 4-hr cultures of T cells and monocytes in the presence of the OKT3 mAb. Both anti-CD3-dependent lysis of monocytes and IL- 1 production were severely decreased after treatment of T cells with either OKT3 or OKT8 mAb plus complement, but were not affected significantly by treatment with the OKT4 mAb plus complement. Purified CD8+ cells, prepared using the cell sorter, exhibited significant levels of anti-CD3dependent monocyte lysis (> 10%). In contrast, purified CD4+ cells did not exhibit significant levels of anti- CD3-dependent cytotoxicity (i 10%). Production of high concentrations of IL-l was observed in cultures of purified CD8+ cells and monocytes in the presence of anti-CD3 mAb. Only low concentrations of IL-1 were detected in cultures of purified CD4+ cells, monocytes, and OKT3 mAb. These results suggest that CD8+ cells are primarily responsible for lysis of monocytes, which is associated with IL-1 production. It appears that anti-CD3 mAb brings CD8+ T cells and monocytes into close proximity by binding to the CD3 antigen on T cells and to the Fc IgG receptor on monocytes. This interaction results in lysis of monocytes primarily by CD8+ cells, after bypassing any antigen recognition requirements that may be othetwise needed. Lysis of monocytes appears to be associated with IL- 1 release. o 1988 Academic FWSS, IIIC.
’ This work was supported in part by Grants CA-27 197, CA- 13 148, CA-030 13, and CA-4 1699 from the National Institutes of Health, Grants IM-409D and CH-420 from the American Cancer Society, by a grant from the Eleanor Naylor Dana Charitable Trust, and by a grant from the Erwin Zaban Melanoma Research Fund.
257
0008-8749/88$3.00 Copyright 0 1988 by Academic F’ress, Inc. All rights of reproduction in any form reserved.
258 ITOH ET AL.
INTRODUCTION
Monoclonalantibodies (mAb)3 recognizing the CD3 T-cell differentiation antigen (anti-CD3 mAb) inhibit both specific T-cell-mediated cytotoxicity against allogeneic or autologous modified targets ( 1,2) and Con A-dependent nonspecific cytotoxicity against fresh solid tumor cells mediated by CD3+ CDS+ CD16 (Leu 1 l)- granular lymphocytes (3). The CD3 antigen may be associated with signal transfer or transduc- tion from the T-cell-receptor complex to structures inside the cells responsible for delivery of the lethal hit (4). Additionally, anti-CD3 mAbs mediate nonspecific lysis of cells from certain established human tumor cell lines, primarily by human CTL clones and CD3+ CD8+ Leu 7+ T cells (5-7). However, certain fresh melanoma tu- mor targets are not lysed by Leu 7+ T cells in the presence of OKT3 mAb (3). Murine CTL can also lyse target cells bearing Fc receptors in the presence of anti-T-cell- receptor mAb (8). Human CTL clones (9) and CD3+ CD8+ Leu 7+ T lymphocytes ( 10) also have the ability to lyse murine hybridomas producing OKT3 mAb, presum- ably by binding to the anti-CD3 mAb expressed on the surface of the target cells. Furthermore, CTL clones can lyse irrelevant target cells in the presence of heteroag- gregates containing anti-CD3 mAb covalently linked to anti-target cell antibodies ( 11, 12). These results suggest that presentation of the anti-CD3 mAb on the surface of the target cells through Fc receptors (5-8) heteroaggregate antibodies ( 11, 12), or other means (9, 10) allows the lysis of these targets by effector T cells expressing the CD3 antigen, apparently bypassing the requirement for specific antigen recognition.
We report here that peripheral blood T lymphocytes lyse autologous monocytes in the presence of anti-CD3 mAb. CD8+ cells were primarily responsible for this lysis, which was associated with high levels of IL- 1 production. In contrast, CD4+ cells did not exhibit significant levels of lysis of monocytes in the presence of anti-CD3 mAb, and low concentrations of IL- 1 were found in those cultures.
MATERIALS AND METHODS
Monoclonal antibodies. OKT3 (IgG2,), OKT4 (IgG&, OKT8 (IgG&, OKTl 1 (IgG&, and control murine IgG2, mAb were obtained from Ortho Diagnostic Co. (Raritan, NJ). Anti&u 4 (IgG,), anti-Leu M3, anti-transferrin receptor (IgG&, and anti&u 11 b mAb were obtained from Becton-Dickinson (Mountain View, CA). Anti&u 7 mAb (IgM) ( 13) ascites was prepared in our laboratory. F(ab’)* fragments of the OKT3 mAb were prepared by the method of Nisonoff et al. (14). Purified OKT3 mAb at a concentration of 5-10 mg/ml was treated with pepsin for 16 hr at 24°C in a buffer of pH 4.5 containing 0.07 M sodium acetate and 0.05 M sodium chloride. The mixture was neutralized with 1 NNaOH and fractionated on Sephadex G- 150. Analysis by SDS-PAGE revealed no detectable intact immunoglobulin mole-
2 To whom correspondence should be addressed at Department of General Surgery, M. D. Anderson Hospital and Tumor Institute, 15 15 Holcombe Blvd., HMB 178, Houston, TX 77030.
3 Abbreviations used: mAb, monoclonal antibody; CD3, cluster of differentiation 3; CD4, cluster of differentiation 4; CD8, cluster of differentiation 8; Con A, concanavalin A; NA lymphocytes, nylon-wool column-nonadherent lymphocytes; PMN, polymotphonuclear cells; LGL, large granular lymphocytes; CTL, cytotoxic T lymphocytes; IL- I, interleukin- 1; IL-2, interleukin-2; 2-ME, 2-mercaptoethanol; PBS, fetal bovine serum; PBMC, peripheral blood mononuclear cells; sIg, surface immunoglobulin; PHA, phy- tohemagglutin; ADCC, antibody-dependent cell-mediated cytotoxicity.
ANTI-CDfDEPENDENT LYSIS OF MONOCYTES 259
cules in the F(ab’)2 fragments. F(ab’h fragments of the OKT3 mAb and intact anti- body molecules stained similar proportions of peripheral blood T lymphocytes, dem- onstrating that the antigen-binding region of the digested mAb was intact and not affected by the F(ab’)* preparation procedure.
Endotoxin levels of reagents and culture medium. All media and reagents used in these experiments, with the exception of Percoll (Pharmacia), were free of endotoxin as determined by the Limulus amebocyte lysate assay (Associates of Cape Cod, Inc., Woodshole, MA) (sensitivity limit: 0.125 r&ml of endotoxin). Percoll contained very low levels of endotoxin (between 0.125 ng and 0.250 rig/ml) and it was not employed for the preparation of monocytes or in any of the experiments where IL- 1 determina- tions were carried out.
Preparation of lymphocytes. Heparinized peripheral blood or bulfy coats from adult healthy donors, free of hepatitis antigen and antibody, were supplied by the Gulf Coast Blood Center (Houston, TX). Heparinized blood or bufQ coats were di- luted with Hank’s Balanced Salt Solution (HBSS) and centrifuged on Ficoll-Hy- paque density cushions (Pharmacia, Piscataway, NJ) as previously described (14). Peripheral blood mononuclear cells (PBMC) were isolated from the interface. Non- adherent (NA)-lymphocytes were prepared by applying PBMC on nylon-wool col- umns as described previously ( 15). These NA-lymphocytes contained approximately 95% E-rosette-forming cells, as determined by rosetting with sheep erythrocytes, and less than 1% monocytes as determined by morphology, yeast-phagocytosis, and im- munofluorescence analysis using the anti-Leu M3 mAb. To isolate large granular lymphocytes (LGL) and T cells, NA-lymphocytes were applied on a gradient contain- ing five layers of different densities of Percoll solutions (Pharmacia) and centrifuged for 40 min as previously described ( 16). Cells isolated from the interface of the 40% Percoll layer were used as LGL-enriched lymphocytes. This fraction consisted of 70 + 5% LGL as determined by Giemsa staining. Cells isolated from the interface of the 50% Percoll layer were used as T-cell-enriched lymphocytes. This fraction consisted of ~99% T3+ lymphocytes as determined by immunofluorescence analysis. In the experiments in which IL-1 determinations were carried out, T cells were pre- pared from NA-lymphocytes by rosetting with sheep erythrocytes as previously de- scribed (4).
Preparation of monocytes, non-T lymphocytes, and polymorphonuclear cells (PAIN). Purified monocytes were prepared by adherence on fetal bovine serum (FBS) coated dishes as previously described (17). Cells prepared by this method consisted of more than 99% monocytes as determined by morphological criteria, more than 98% of cells phagocytizing yeast and more than 90% Leu M3+ cells. To prepare nylon- wool-adherent non-T lymphocytes, monocytes and PMN were removed by incubat- ing whole heparinized peripheral blood with carbonyl iron for 30 min at 37°C fol- lowed by centrifugation on a Ficoll-Hypaque density cushion. Monocyte-depleted lymphocytes were collected from the interface, applied on nylon-wool columns, and incubated for 45 min at 37°C. Nylon-fiber-retained cells were retrieved from the fiber by vigorous agitation in cold PBS, were mixed with neuraminidase-treated sheep erythrocytes for 1 hr at 4”C, and separated on Ficoll-Hypaque density cushions (4). E-rosette-negative nylon-wool-adherent cells were recovered from the interface and consisted of 80% sIg-positive cells. These cells contained less than 5% E-rosette-form- ing cells and less than 2% Leu M3+ cells.
260 ITOH ET AL.
Mixtures of PMN and red blood cells were recovered from the bottom of Ficoll- Hypaque density cushions after centrifugation of whole blood or bully coat cells (ap- propriately diluted) at 400g. These mixtures were applied on a 60% Percoll solution and centrifuged for 30 min. PMN were collected from the interface. Their purity was higher than 99% as determined by Giemsa staining.
Human IgG and myeloma proteins. Purified human IgG was obtained from Beh- ringwerke AG (Marburg, West Germany). Purified human y-globulin and Con A were obtained from Sigma Chemical Co. (St. Louis, MO). Purified IgG, , IgG2, IgG3, and IgG, human myeloma proteins were kindly provided by Dr. Donald Capra (UTHSC, Dallas, TX).
Treatment of the cells with antibody plus complement. NA-lymphocytes were treated with either OKT3, OKT4, OKT8, anti&u 7, or anti-Leu 1 lb mAb for 45 min at 4’C, washed, and incubated with rabbit low toxic-H complement (Cedarlane Lab., Ontario, Canada) (15% v/v) for 50 min at 37°C as described previously (14). The cells were washed twice before further studies.
Preparation of CD4+ and CD8+ cells. Monocyte-depleted lymphocytes were stained for 30 min at 4°C with FITC-conjugated anti-Leu 2a (antiCD8) or FITC- conjugated anti-Leu 3a (anti-CD4) mAb (Becton-Dickinson). CD4+ or CD8+ cells were isolated using the cell sorter (EPICS Model C, Coulter Electronics, Hialeah, FL) as previously described (15).
Cytotoxic assay. Cytotoxicity was determined by a standard 4-hr “Cr-release assay as described previously (15). Monocytes, PMN, non-T lymphocytes, LGL-enriched lymphocytes, and T cells were incubated with 200 &i of Naz5’Cr0, (Amersham Corp., Arlington Heights, IL) for 60 min at 37°C. After washing three times, these cells were used as target cells. MAbs, human myeloma proteins, human IgG, or Con A was added to the effector cells immediately before the addition of the target cells, unless it is specified differently in the text. These agents did not affect spontaneous release. Spontaneous release was determined by incubating “Cr-labeled target cells in culture with medium alone, and it did not exceed 20% of the maximum release. The maximum release was determined by lysing target cells in 2 N HCl solution. Mean percentage lysis was calculated as follows:
maximum release (cpm) - spontaneous release (cpm) .
Detection of IgG Fc receptors. IgG Fc receptors were detected by immunofluores- cence. Purified monocytes, PMN, non-T lymphocytes, LGLenriched lymphocytes, or T lymphocytes were incubated for 30 min at 4’C with FITC-conjugated OKT3 mAb (IgGz,) or control FITC-conjugated murine IgGza (Ortho). Different aliquots of these cells were also incubated with F(ab’)* fragments of OKT3 mAb followed by staining with FITC-conjugated F(ab’)2 fragments of goat anti-mouse IgG (Cappel, Cochranville, PA). Immunofluorescence analysis was carried out using the cell sorter (Epics, Model C, Coulter Electronics).
Assaysfor IL-l and IL-2. A clone of DlO.G4.1 helper T cells was used for determi- nation of IL- 1 activity (19), and was kindly provided by Dr. L. Lachman of this insti- tution. DlO.G4.1 cells were stimulated by mitomycin C-treated C57BL/6 spleen cells every 2 weeks and maintained in RPM1 1640 medium supplemented with 10% FBS, 2 X lo-’ M 2-mercaptoethanol(2-ME) (complete medium), and 200 U/ml recombi- nant IL-2 (rIL-2). rIL-2 was kindly provided by Biogen Inc. (Cambridge, MA).
ANTI-CD3-DEPENDENT LYSIS OF MONOCYTES 261
TABLE 1
AntiCD3Dependent Cytotoxicity against Autologous Monocytes”
% Lysis of autologous target cells by NA-lymphocytes
mAb added IgG isotype Monocytes Non-T lymphocytes NA-lymphocytes
’ NA-lymphocytes were tested for cytotoxicity against autologous monocytes, other non-T lymphocytes, and NA-lymphocytes in 4-hr 5’Cr-release assays in the presence of mAb. Values represent mean ? SD of percentage lysis from six different donors at an E/T ratio of 50: 1.
b P < 0.00 1 as compared to percentage lysis of the others (Student’s two-tailed t test).
D 1 0.G4.1 cells were used for IL- 1 determination 16-20 days after their last stimula- tion with C57BL/6 spleen cells. Briefly, DlO cells were incubated for 24 hr with com- plete medium in the absence of rIL-2. Under these culture conditions, the D 10 cells do not exhibit a proliferative response in the presence of 1 U/ml of rIL-2. These cells (1 X lo4 cells per well) were incubated for 72 hr with twofold dilutions of the test sample in a total volume of 0.2 ml of complete medium and 2.5 &ml of Con A. Control DlO cell cultures were incubated with the test samples alone, with 2.5 &ml of Con A alone, or with medium alone for 72 hr. Positive controls were cultures of 1 X lo4 DlO cells in 0.2 ml complete medium, 2.5 pg/ml Con A, and various dilutions of recombinant IL- 1 p (Cistron Biotechnology, Pine Brook, CA) or rIL-2.
IL-2 activity was determined using the IL-Zdependent cloned NK8 cells as re- ported previously (20). Briefly, 1 X lo4 NK8 ceils in 0.2 ml of medium supplemented with 10% FBS and twofold dilutions of the test sample, or rIL-2, or rIL-1 were incu- bated for 24 hr. [3H]Thymidine (0.5 &i/well, 6.7 Ci/mmol, ICN, Irvine, CA) was added to the wells 6 hr before harvesting. The cells were harvested and [3H]thymidine incorporation was determined as previously described (4). Rabbit serum against re- combinant human IL- 1 was purchased from Cistron Biotechnology. Murine hybrid- omas producing anti-IL-2 mAb were kindly provided by Dr. K. Smith, Dartmouth Medical School (Hanover, NH) (2 1). Ascitic fluid of this mAb was prepared by stan- dard procedures.
RESULTS
Anti-CD3-Dependent Cytotoxicity against Monocytes
The ability of nonadherent lymphocytes to lyse autologous monocytes, other non- T lymphocytes, or NA-lymphocytes was evaluated in the presence of various mAbs in a 4-hr “Cr-release assay (Table 1). Significant levels of lysis of monocytes was
262 ITOH ET AL.
TABLE 2
AntXD3Dependent Cytotoxicity by Purified Populations of Leukocytes”
a Effector cells were tested for cytotoxicity against autologous target cells in the presence of OKT3 mAb (10’ dilution of ascitic fluid). Values represent mean + SD of percentage lysis from three donors.
b P < 0.001 as compared to percentage lysis caused by the other effector cells (Student’s two-tailed t test).
observed (P < 0.00 1) in the presence ofOKT3 mAb (IgG*,) or anti-Leu 4 mAb (IgG,). Both autologous (Table 1) or allogeneic (data not shown) monocytes were susceptible to antiCD3dependent cytotoxicity. In contrast, none of the other mAbs, including those of the IgGz, class, induced lysis of monocytes by the effector cells. Non-T lym- phocytes, NA-lymphocytes (Table l), purified LGL, or PMN (Table 2) were not sus- ceptible to anti-CD3-dependent cytotoxicity. Furthermore, monocytes were not lysed by NA-lymphocytes in the absence of the anti-CD3 mAb.
a b
of OKT3mAb
l Medium
I I I I I I I I I I] 0 10”10’010~ 108 10’ 106 105 104 103 102
Dilution of OKTBmAb lAscitesl
25 50 100
E/T Ratios
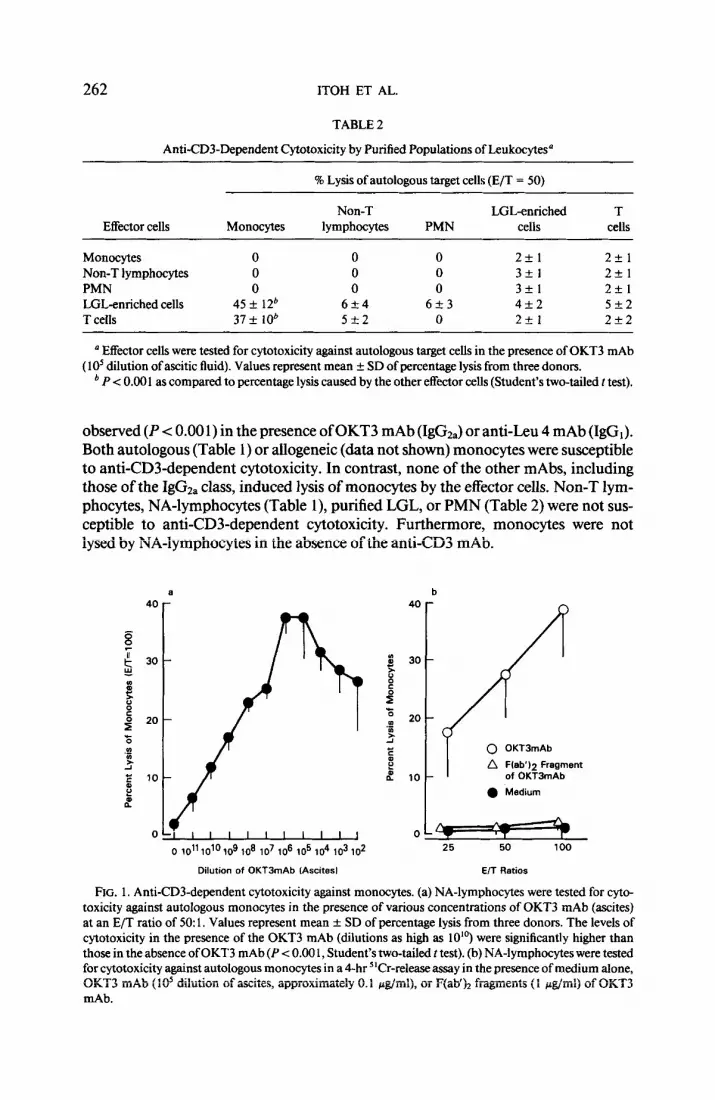
FIG. 1. Anti-CD3-dependent cytotoxicity against monocytes. (a) NA-lymphocytes were tested for cyto- toxicity against autologous monocytes in the presence of various concentrations of OKT3 mAb @cites) at an E/T ratio of 50: 1. Values represent mean + SD of percentage lysis from three donors. The levels of cytotoxicity in the presence of the OKT3 mAb (dilutions as high as 10”) were significantly higher than those in the absence of OKT3 mAb (P < 0.00 1, Student’s two-tailed t test). (b) NA-lymphocytes were tested for cytotoxicity against autologous monocytes in a 4-hr 5’Cr-release assay in the presence of medium alone, OKT3 mAb (10’ dilution of ascites, approximately 0.1 &ml), or F(ab’b fragments (1 &ml) of OKT3 mAb.
ANTI-CD3-DEPENDENT LYSIS OF MONOCYTES 263
1 Monocytes 1 LGL
t-4 control : (---I FITC-OKTS : (-1 FITWgG2a :
control :
FITC-lgG2a :
1% 88% 86%
Control : FtTC-OKT3 : FITC-lgG2a :
rx /--‘\ .’ %
T Lymphocytes
GJntrol : FITC-OKT3 :
.‘“\, FITC-IgGPa :
/ ‘\ *.-I’ ‘.
‘\
1% 62% 3%
1% 98% 1%
- Fluorescertce Intensity
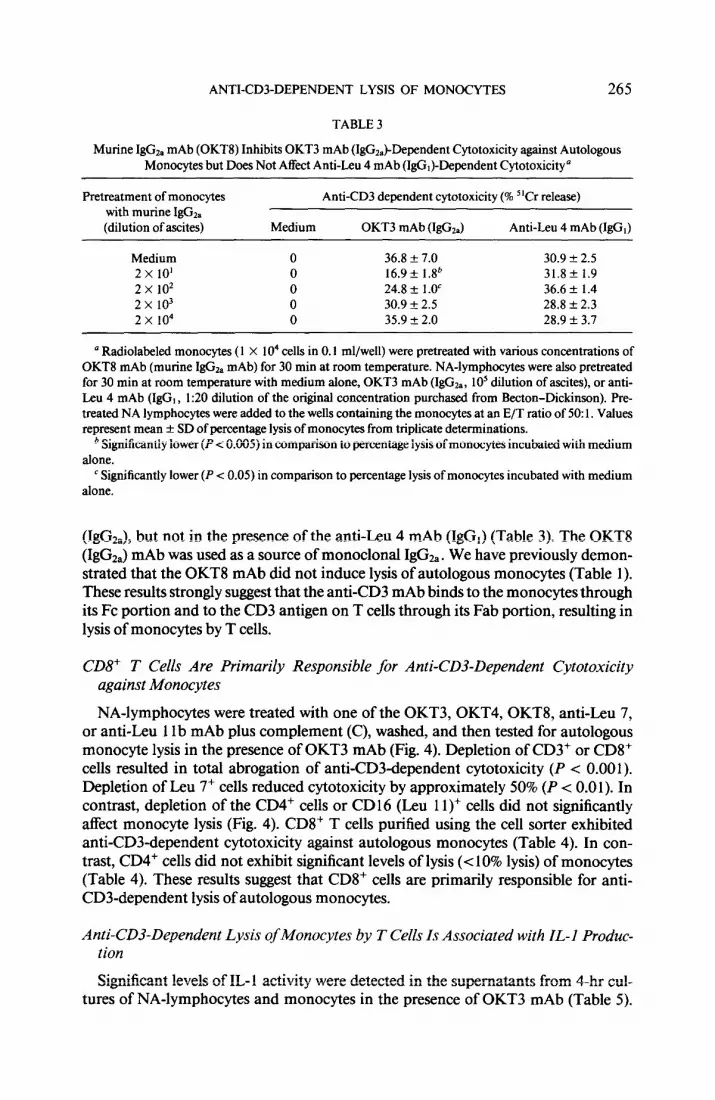
FIG. 2. Binding of OKT3 mAb or control murine I&&, mAb to monocytes. Purified cell fractions were incubated with FITC-conjugated OKT3 mAb or FITC-conjugated control monoclonal IgC& for 30 min at 4°C. Binding of mAb was analyzed using tlow cytometry (Epics Model C).
The levels of the anti-CD3 cytotoxicity were dependent on the concentration of the OKT3 mAb added to the “Cr-release assay (Fig. la). Significant levels of cytotox- icity (P < 0.001) were detectable at dilutions of OKT3 mAb (ascites) as low as 1: 10 lo. Maximum levels were reached at dilutions of 1: 1 O6 to 1: 1 05. A decrease in the cytotoxicity levels was observed at higher concentrations, most likely due to competi- tion for the CD3 antigen on the effector cells between the free antibody and the anti- body bound to the IgG Fc receptor on monocytes. Anti-CD3-dependent lysis of au- tologous monocytes was not observed in the presence of F(ab’)* fragments of the OKT3 mAb (Fig. lb). These results suggest that the Fc portion of the OKT3 mAb is required for anti-CD3-dependent cytotoxicity against monocytes and that the Fc receptor on monocy-tes is a structure necessary for this type of cytotoxicity.
Monocytes did not demonstrate cytotoxicity against “Cr-labeled NA-lymphocytes or purified T lymphocytes in the presence of OKT3 mAb (Table 2). Neither non-T lymphocytes nor PMN showed anti-CD3-dependent cytotoxicity against any target cells tested. In contrast, both LGL-enriched lymphocytes and T cells demonstrated anti-CD3-dependent cytotoxicity against monocytes but not against non-T lympho- cytes, PMN, LGL-enriched lymphocytes, or T cells (Table 2).
Binding of Murine IgG*, mAb to Human Monocytes
Purified monocytes bound FITC-conjugated control murine IgGzI mAb (86%) and FITC-conjugated OKT3 mAb (IgGz,) (86%) equally well, as determined by immuno- fluorescence (Fig. 2). In contrast, all the other cell populations (PMN, non-T cells, LGL, and T lymphocytes) did not bind FITC-conjugated control IgGza mAb. All T lymphocytes and 60% of the LGL-enriched lymphocytes bound the FITC-OKT3 mAb. A substantial proportion of the cells present in the LGL-enriched fraction ex- press the CD3 antigen (3). PMN or non-T lymphocytes did not bind OKT3 mAb
264 ITOH ET AL.
30 -
f 20-
@
H % .g
3 E E lo- L
lgG3
01 ” I I 1 0 2 20 200
Concentration of IgG (pglml)
(‘P<O.Ol, l * P<O.O2, SP<O.O5)
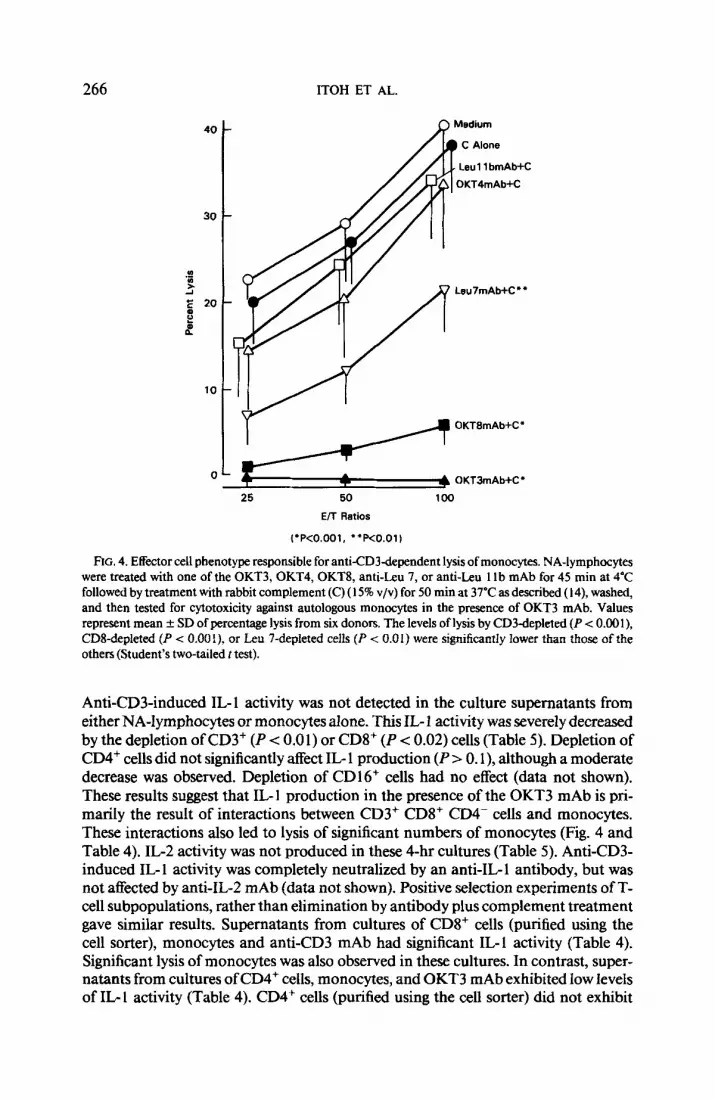
FIG. 3. Inhibition of antiCD3dependent cytotoxicity by purified human myeloma proteins and poly- clonal IgG. NA-lymphocytes were tested for cytotoxicity against autologous monocytes in a 4-hr “Cr- release assay in the presence of 10” dilution of OKT3 mAb (ascites) and 2 to 200 &ml of purilied human IgG, , IgGr , IgGs, or IgGd myeloma protein or polyclonal IgG. Cytotoxicity in the absence of OKT3 mAb was also determined as a negative control (M). Values represent mean f SD of percentage lysis from three donors at an E/T ratio of 100: I. Statistical analysis was determined by Student’s two-tailed t test and the results are shown in the figure.
(Fig. 2). Monocytes also bound FITC-conjugated anti&u 4 mAb (IgG,) as deter- mined using the cell sorter, but did not bind F(ab’h fragments of the OKT3 mAb (data not shown). The proportions of NA-lymphocytes stained with OKT3 mAb and F(ab’)z fragments of OKT3 mAb were identical (data not shown). These results sug- gest that monocytes, but not the other cell populations, bind murine IgG*, mAb through their IgG Fc receptor.
Inhibition of Anti-CD3-Dependent Cytotoxicity by Human IgG Myeloma Proteins and Murine Monoclonal IgGt
Addition of purified human IgGi or IgGj myeloma proteins directly to the cyto- toxic assay significantly inhibited anti-CD3-dependent cytotoxicity against mono- cytes (Fig. 3). IgGz and IgG4 myeloma proteins (up to 200 pg/ml) did not cause sig- nificant inhibition. Purified human IgG inhibited by 50% at a concentration of 200 &ml (Fig. 3) and by 85% at 1 mg/ml (data not shown). Addition of Con A directly to the cytotoxic assay also inhibited anti-CD3-dependent lysis of monocytes by 50% at a concentration of 10 pg/ml and by 90% at a concentration of 50 pg/ml. Addition of PHA failed to inhibit cytotoxicity (data not shown).
Additionally, we observed that pretreatment of monocytes with an unrelated mu- rine IgGz, mAb significantly inhibited their lysis in the presence of the OKT3 mAb
ANTI-CD3-DEPENDENT LYSIS OF MONOCYTES 265
TABLE 3
Murine IgGzs mAb (OKTS) Inhibits OKT3 mAb (IgG&Dependent Cytotoxicity against Autologous Monocytes but Does Not Affect Anti&u 4 mAb (IgGJ-Dependent Cytotoxicity”
Pretreatment of monocytes with murine IgGr. (dilution of ascites)
’ Radiolabeled monocytes ( 1 X lo4 cells in 0.1 ml/well) were pretreated with various concentrations of OKT8 mAb (murine IgGz, mAb) for 30 min at room temperature. NA-lymphocytes were also pretreated for 30 min at room temperature with medium alone, OKT3 mAb (IgGa, lo5 dilution of ascites), or anti- Leu 4 mAb (IgG, , 1:20 dilution of the original concentration purchased from Becton-Dickinson). Pre- treated NA lymphocytes were added to the wells containing the monocytes at an E/T ratio of 50: 1. Values represent mean + SD of percentage lysis of monocytes from triplicate determinations.
b Significantly lower (P < 0.005) in comparison to percentage lysis of monocytes incubated with medium alone.
‘Significantly lower (P < 0.05) in comparison to percentage lysis of moncqtes incubated with medium alone.
(IgG&, but not in the presence of the anti&u 4 mAb (IgG,) (Table 3). The OKT8 (IgG& mAb was used as a source of monoclonal IgGz, . We have previously demon- strated that the OKT8 mAb did not induce lysis of autologous monocytes (Table 1). These results strongly suggest that the anti-CD3 mAb binds to the monocytes through its Fc portion and to the CD3 antigen on T cells through its Fab portion, resulting in lysis of monocytes by T cells.
CD8+ T Cells Are Primarily Responsible for Anti-CD3-Dependent Cytotoxicity against Monocytes
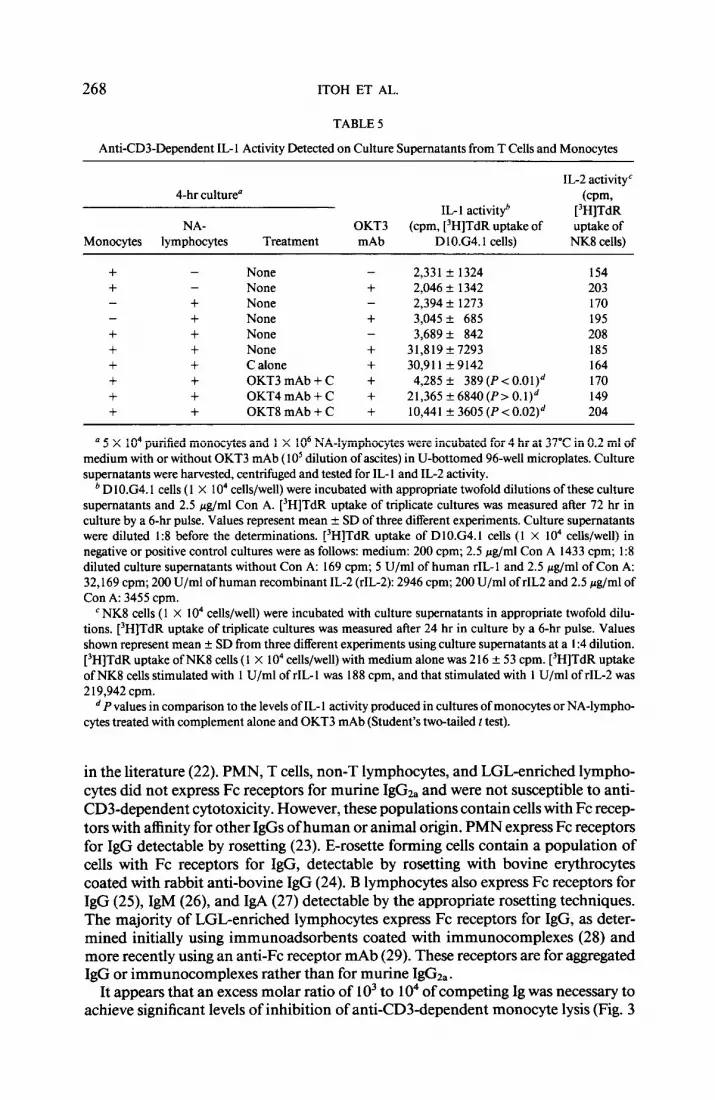
NA-lymphocytes were treated with one of the OKT3, OKT4, OKT8, anti&u 7, or anti&u 11 b mAb plus complement (C), washed, and then tested for autologous monocyte lysis in the presence of OKT3 mAb (Fig. 4). Depletion of CD3+ or CD8+ cells resulted in total abrogation of anti-CD3-dependent cytotoxicity (P < 0.001). Depletion of Leu 7+ cells reduced cytotoxicity by approximately 50% (P < 0.0 1). In contrast, depletion of the CD4+ cells or CD16 (Leu 1 l)+ cells did not significantly affect monocyte lysis (Fig. 4). CD8+ T cells purified using the cell sorter exhibited anti-CD3-dependent cytotoxicity against autologous monocytes (Table 4). In con- trast, CD4+ cells did not exhibit significant levels of lysis (< 10% lysis) of monocytes (Table 4). These results suggest that CD8+ cells are primarily responsible for anti- CD3-dependent lysis of autologous monocytes.
Anti-CD3-Dependent Lysis of Monocytes by T Cells Is Associated with IL-l Produc- tion
Significant levels of IL- 1 activity were detected in the supematants from 4-hr cul- tures of NA-lymphocytes and monocytes in the presence of OKT3 mAb (Table 5).
266 ITOH ET AL.
, Leu 11 bmAb+C
Leu7mAb+C**
25 50
E/l Ratios
100
~‘P<O.OOl, “RO.01)
FIG. 4. Effector cell phenotype responsible for antXD3dependent lysis of monocytes. NA-lymphocytes were treated with one of the OKT3,OKT4,OKT8, anti-Leu 7, or anti-Leu 1 lb mAb for 45 min at 4°C followed by treatment with rabbit complement (C) (15% v/v) for 50 min at 37’C as described ( 14), washed, and then tested for cytotoxicity against autologous monocytes in the presence of OKT3 mAb. Values represent mean + SD of percentage lysis from six donors. The levels of lysis by CD3depleted (P < 0.00 l), CDS-depleted (P < 0.00 i), or Leu 7-depleted cells (P < 0.0 1) were significantly lower than those of the others (Student’s two-tailed t test).
Anti-CD3-induced IL-1 activity was not detected in the culture supernatants from either NA-lymphocytes or monocytes alone. This IL 1 activity was severely decreased by the depletion of CD3+ (P < 0.0 1) or CDV (P < 0.02) cells (Table 5). Depletion of CD4+ cells did not significantly affect IL- 1 production (P 10. I), although a moderate decrease was observed. Depletion of CDl6+ cells had no effect (data not shown). These results suggest that IL-l production in the presence of the OKT3 mAb is pri- marily the result of interactions between CD3+ CDS’ CD4- cells and monocytes. These interactions also led to lysis of significant numbers of monocytes (Fig. 4 and Table 4). IL-2 activity was not produced in these 4-hr cultures (Table 5). Anti-CD3- induced IL- 1 activity was completely neutralized by an anti-IL- 1 antibody, but was not affected by anti-IL-2 mAb (data not shown). Positive selection experiments of T- cell subpopulations, rather than elimination by antibody plus complement treatment gave similar results. Supernatants from cultures of CDS+ cells (purified using the cell sorter), monocytes and anti-CD3 mAb had significant IL-1 activity (Table 4). Significant lysis of monocytes was also observed in these cultures. In contrast, super- natants from cultures ofCD4+ cells, monocytes, and OKT3 mAb exhibited low levels of IL- 1 activity (Table 4). CD4+ cells (purified using the cell sorter) did not exhibit
ANTI-CDZDEPENDENT LYSIS OF MONOCYTES 267
TABLE 4
Purified CD8 T Cells Arc Primarily Responsible for Anti-CD3Dependent Lysis of Monocytes: Association with IL- 1 Production”
Effector cells
OKT3 mAb in assay
% Lysis of autologous monocytes
IL-I activity (cpm, [‘H]TdR uptake of DlO.G4.1 cells)
CDS+ cells CDS+ cells
CD4+ cells CD4+ cells
- 0 2,045? 214 + 21.4 + 3.8 31,505 + 1954
- 0 2,165k 267 + 8.5 + 3.0b 8,501 f 1803b
’ Peripheral blood nonadherent lymphocytes were stained for 30 min at 4°C with FITC-conjugated anti- Leu 2a (CDS) or FITC-conjugated anti&u 3a (CD4) mAb. Fluorescent cells were separated using the cell sorter. The purity of the CD4+ cells was 98% and the purity of the CD8+ cells was 972, as determined by immunofluorescence analysis. These cells were tested for cytotoxicity against “Cr-labeled autologous monocytes at an E/T ratio of 10: 1 in the presence or absence of OKT3 mAb (10’ dilution of ascitic fluid). Simultaneously 2 X 10’ purified CD8+ or CD4+ cells were incubated with 5 X lo4 purified autologous monocytes in the presence or absence of OKT3 mAb. After incubation for 4 hr, supematants were har- vested, centrifuged, and tested for IL-l activity using DlO.G4.1 cells. Values represent mean & SD of triplicate samples.
b Significantly lower (P < 0.0 I) in comparison to the levels of cytotoxicity or IL- 1 activity produced in cultures of CD8+ cells, monocytes, and OKT3 mAb.
significant levels of lysis of monocytes (< 10%) (Table 4). IL-2 activity was not detect- able in these 4-hr culture supernatants (data not shown). These results suggest that CD8+ cells are primarily responsible for anti-CD3-induced lysis of autologous mono- cytes, which appears to be associated with IL- 1 production.
DISCUSSION
Peripheral blood CD8+ T lymphocytes are able to lyse autologous monocytes in the presence of the anti-CD3 mAb. This anti-CD3-dependent cytotoxicity was inhib- ited by purified serum human IgG (polyclonal), purified human IgG, or IgG3 my- eloma proteins, and murine monoclonal IgGza. Human IgGi and IgG3 myeloma proteins and human serum IgG were highly effective in inhibiting the proliferative responses to the OKT3 murine mAb (IgG&, whereas IgG2 and IgG4 myeloma pro- teins were less effective (22). Furthermore, the Fc portion of the OKT3 mAb was required for lysis. F(ab’)* fragments of the OKT3 mAb were ineffective in inducing anti-CD3-dependent cytotoxicity (Fig. 1 b). Pretreatment of monocytes with an unre- lated murine IgG2, mAb inhibited OKT3 (IgG&dependent lysis but not anti&u 4 (IgGJ-dependent lysis. These results strongly suggest that IgG Fc receptors on mono- cytes play a critical role in this type of cytotoxicity. It appears that the anti-CD3 mAb brings effector T cells and monocytes in close proximity by binding through its Fc portion to the Fc receptor on monocytes and via its Fab portion to the CD3 antigen on the T cells. Binding of the effector cells to the targets through the anti-CD3 mAb appears to be sufficient for triggering of the lethal hit. Antigen recognition require- ments appear to be bypassed.
Immunofluorescence studies revealed that only monocytes from all the cell popu- lations examined expressed Fc receptors for murine IgGza, in agreement with reports
268 ITOH ET AL.
TABLE 5
Anti-CD3Dependent IL-I Activity Detected on Culture Supernatants from T Cells and Monocytes
IL-2 activityC 4-hr culture“ (cpm,
IL- 1 activityb [‘HJTdR NA- OJA. (cpm, [‘H]TdR uptake of uptake of
’ 5 X lo4 purified monocytes and 1 X lo6 NA-lymphocytes were incubated for 4 hr at 37’C in 0.2 ml of medium with or without OKT3 mAb (1 O5 dilution of ascites) in U-bottomed 96-well microplates. Culture supernatants were harvested, centrifuged and tested for IL- 1 and IL-2 activity.
’ DlO.G4.1 cells (1 X 10’ cells/well) were incubated with appropriate twofold dilutions of these culture supernatants and 2.5 &ml Con A. [‘H]TdR uptake of triplicate cultures was measured after 72 hr in culture by a 6-hr pulse. Values represent mean f SD of three different experiments. Culture supematants were diluted I:8 before the determinations. [‘H]TdR uptake of DlOG4.1 cells (1 X lo4 cells/well) in negative or positive control cultures were as follows: medium: 200 cpm; 2.5 &ml Con A 1433 cpm; 1:8 diluted culture supematants without Con A: 169 cpm; 5 U/ml of human rIL- 1 and 2.5 &ml of Con A: 32,169 cpm; 200 U/ml of human recombinant IL-2 (rIL-2): 2946 cpm; 200 U/ml of rIL2 and 2.5 &ml of Con A: 3455 cpm.
c NK8 cells (1 X lo4 cells/well) were incubated with culture supematants in appropriate twofold dilu- tions. [‘H]TdR uptake of triplicate cultures was measured after 24 hr in culture by a 6-hr pulse. Values shown represent mean + SD from three different experiments using culture supematants at a I :4 dilution. [‘H]TdR uptake of NK8 cells (I X lo4 cells/well) with medium alone was 2 16 f 53 cpm. [‘H]TdR uptake of NK8 cells stimulated with 1 U/ml of rIL- 1 was 188 cpm, and that stimulated with 1 U/ml of rIL-2 was 2 19,942 cpm.
d P values in comparison to the levels of IL- 1 activity produced in cultures of monocytes or NA-lympho- cytes treated with complement alone and OKT3 mAb (Student’s two-tailed t test).
in the literature (22). PMN, T cells, non-T lymphocytes, and LGL-enriched lympho- cytes did not express Fc receptors for murine IgGz, and were not susceptible to anti- CD3-dependent cytotoxicity. However, these populations contain cells with Fc recep- tors with affinity for other IgGs of human or animal origin. PMN express Fc receptors for IgG detectable by rosetting (23). E-rosette forming cells contain a population of cells with Fc receptors for IgG, detectable by rosetting with bovine erythrocytes coated with rabbit anti-bovine IgG (24). B lymphocytes also express Fc receptors for IgG (25), IgM (26), and IgA (27) detectable by the appropriate rosetting techniques. The majority of LGL-enriched lymphocytes express Fc receptors for IgG, as deter- mined initially using immunoadsorbents coated with immunocomplexes (28) and more recently using an anti-Fc receptor mAb (29). These receptors are for aggregated IgG or immunocomplexes rather than for murine IgGza.
It appears that an excess molar ratio of 1 O3 to 1 O4 of competing Ig was necessary to achieve significant levels of inhibition of anti-CD3-dependent monocyte lysis (Fig. 3
ANTI-CD3-DEPENDENT LYSIS OF MONOCYTES 269
and Table 3). Several explanations may account for the need of high Ig concentrations for inhibition. The OKT3 mAb concentration used in these experiments was a 1: 10’ dilution of ascitic fluid (approximately 0.1 pg/ml). However, significant lysis (P < 0.001) of monocytes occurs at much lower concentrations of OKT3 mAb (as low as 1: 10” of ascitic fluid, Fig. la). Therefore, it appears that the presence of very few OKT3 molecules on the surface of the monocytes is sufficient for their lysis by the effector cells. These few OKT3 mAb molecules may remain on the cell surface after the replacement of the vast majority of the OKT3 mAb by the competing Igs. Altema- tively, the affinity of the OKT3 mAb bound on the surface of T cells for the Fc recep- tor on monocytes may be higher than that of competing Igs.
Anti-CD3-dependent cytotoxicity is mediated primarily by CD3+ CD8+ CD4- cells. These CD8+ T cells have been found to be responsible for lectin-dependent cytotoxicity against targets from human tumor cell lines (3, 7) and from fresh solid tumor cells (3). These CD3+ CD8+ cells can also lyse irrelevant target cells in the presence of heteroaggregates containing anti-CD3 mAb covalently linked to antitar- get antibody (30). In addition, these cells lyse OKT3 mAb-expressing murine hybrid- omas (10). The lysis of all these targets is inhibited by OKT3 mAb (3, 7, 10). Our results and those of others suggest that these CD3+ CD8+ cells may be specific CTL that have been generated in vivo against autologous modified cells such as virus-in- fected cells or tumor cells (3, 10). This is supported by the expression of the CD3 antigen on these cells and by the fact that they do not exhibit cytotoxic activity unless their antigen recognition requirements are bypassed by the anti-CD3 mAb. CTL gen- erated in vivo are thought to play an important role for the control of the development of tumors and viral infections (3 l-35).
Anti-CD3-induced lysis is inhibited by Con A at the effector phase. It has been suggested that Con A may compete with the anti-CD3 mAb for binding to the CD3 antigen on the surface of T cells (36). Experiments using 51Cr-labeled T cells revealed that monocytes did not lyse these target cells in the presence of the OKT3 mAb in an antibody-dependent cell-mediated cytotoxicity (ADCC). These results suggest that the mechanisms involved in ADCC by monocytes are different from those involved in anti-CD3-dependent cytotoxicity by T cells.
Blood monocytes are able to release IL- 1 into culture supernatants after lethal stim- ulation by toxic dust particles (silica) (37). It has been suggested that IL-1 produced by monocytes in the presence of anti-CD3 mAb induces IL-2 receptor expression on T cells stimulated with anti-CD3 mAb (38). Monocytes are required for anti-CD3 mAb-dependent IL-2 production and proliferation (36, 39). Their primary function appears to be the presentation of the anti-CD3 mAb to T cells through their Fc recep- tors (40). We report here that presentation by monocytes of anti-CD3 mAb to CD8+ T cells resulted in lysis of monocytes, and that it is associated with the release of IL- 1 activity into the culture supernatant. Depletion of CD3+ or CD8+ cells resulted in significant reduction of the levels of monocyte lysis and of IL- 1 production. In con- trast, depletion of CD4+ cells did not significantly affect the levels of lysis of mono- cytes or IL-l production. Purified CD8+ T cells exhibited significant levels of anti- CD3-dependent cytotoxicity against autologous monocytes. Supematants from cul- tures of CD8+ cells and monocytes in the presence of anti-CD3 mAb contained high concentrations of IL-l activity (Table 4). In contrast, CD4’ cells did not cause sig- nificant levels of monocyte lysis in the presence of anti-CD3 mAb and supematants from cultures of CD4+ cells and monocytes in the presence of anti-CD3 mAb had
270 ITOH ET AL.
low levels of IL- I activity. Supematants from cultures of CD&depleted lymphocytes, monocytes, and anti-CD3 mAb exhibited significantly lower IL- 1 activity, in compar- ison to that produced in cultures containing NA-lymphocytes, monocytes, and anti- CD3 (lo,44 1 cpm versus 30,9 11 cpm; P c 0.02; Table 5). Depletion of CD8+ cells using mAb and complement treatment did not completely abolish the proliferation of the DlO.G4.1 cells. It is possible that these cpm may not be due to the production of IL-1 alone. It may be attributed to the response of DlO cells to other cytokines such as IL-4 or to mixtures of these cytokines with low concentrations of IL- 1. It has been reported (4 1) that IL-4 significantly enhances the proliferation of D 10.G4.1 cells in the presence of IL- 1. Similar explanations may account for the low but significant proliferation of D 10.G4.1 cells in response to supematants from cultures of purified CD4+ cells and monocytes (Table 4; 850 1 f 1803 cpm versus 2 165 f 267 cpm of the controls). It appears that CD8+ CD4- cells are primarily responsible for antiCD3- dependent lysis of monocytes and that this lysis is associated with IL- 1 production. Similarly, CD3+ CD4- CD8+ T lymphocytes have been shown to be responsible for lysis of murine hybridomas expressing the OKT3 mAb on the cell surface (10). In contrast, CD4+ CD8- T cells appear to be primarily responsible for IL2 production in response to these OKT3 hybridomas (42).
Interactions between CD8+ or CD4+ cells and monocytes in the presence of anti- CD3 mAb may provide a model for investigating interactions between T-cell subsets and monocytes in general and for the release of factors as a result of these interactions. Stimulation of T cells by anti-CD3 mAb requires monocytes (36, 39), and it may be considered as a nonspecific monocytedependent model for T-cell activation and proliferation, analogous to that of specific antigen presentation to T cells by mono- cytes. Furthermore, administration of the OKT3 mAb in vivo is effective in reversing acute rejection of renal allografts (43,44). It is possible that in these patients mono- cytes may present the OKT3 mAb to effector T cells in vivo, resulting in monocyte lysis and IL- 1 production.
ACKNOWLEDGMENTS
We thank Ms. Caryl Lane for technical assistance in preparing the F(ab’)r fragments of the OKT3 mAb, Mr. Philip Barren for operating the cell sorter, and Ms. Millie Young and Ms. Helen Farr for assistance in preparing this manuscript.
REFERENCES
1. Platsoucas, C. D., and Good, R. A., Proc. Nati. Acad. Sci. USA 78,4500,198 1. 2. Chang, T. W., Kung, P. C., Gingas, S. P., and Goldstein, G., Proc. Natl. Acad. Sci. USA 78, 1805,
1981. 3. Itoh, K., Platsoucas, C. D., Tilden, A. B., Pollock, R. E., and Balch, C. M., Cell. Immunol. lB8,283,
1987. 4. Platsoucas, C. D., Eur. J. Immunol. 14,566, 1984. 5. Mentzer, S. J., Barbosa, J. A., and Burakoff, S. J., J. Immunol. 135,34, 1985. 6. Spits, H., Yssel, H., Leeuwenberg, K., and devries, J., Eur. J. Zmmunol. l&88, 1986. 7. Phillips, J. H., and Lanier, L. L., J. ImmunoI. 136, 1579, 1986. 8. Staem, U. D., Kanagawa, O., and Bevan, M. J., Nature (London) 314,628,1985. 9. Hoffman, R. W., Bluestone, J. A., Leo, O., and Shaw, S., J. Immunol. 135,5, 1985.
10. Itoh, K., Balch, C. M., and Platsoucas, C. D., Ceil. Zmmunol. 108,313, 1987. 11. Perez, P., Hoffman, R. W., Shaw, S., Bluestone, J. A., and &gal, D. M., Nature (London) 315, 354,
1985. 12. Karpovsky, B., Titus, J. A., Stephany, D. A., and Segal, D. M., J. Exp. Med. 160,1686, 1984.
ANTI-CD3-DEPENDENT LYSIS OF MONGCYTES 271
13. Abo, T., and Balch, C. M., J. Immunol. 127,1024, 198 1. 14. Nisonoff, A., Wissler, F. C., Lipman, F. C., and Woemile, D. L., Arch. B&hem. Biophys. 89, 230,
1960. 15. Itoh, K., Tilden, A. B., and BaIch, C. M., J. Immunol. 136,3910, 1986. 16. Arai, S., Yamamoto, H., Itoh, K., and Kumagai, K., J. Immunol. 131,65 1, 1983. 17. Kumagai, K., Itoh, K., Hinuma, S., and Tada, M., J. Immunol. Methods 29, 17, 1979. 18. Itoh, K., and Kumagai, K., J. Immunol. 124,1830, 1980. 19. Kaye, J., and Janeway, C. A., LymphokineRes. 3, 175, 1984. 20. Suzuki, R., Handa, K., Itoh, K., and Kumagai, K., J. Immunol. 135,1714,1983. 21, Smith, K. A., Favata, M. F., and Oroszlan, S., J. Immunol. 131, 1808, 1983. 22. Tsoukas, C. D., Lambris, J., Lotz, M., Vallentine, M. A., Vaughan, J. H., and Carson, D. A., Cell.
Immunol. 89,66, 1985. 23. Ehlenberg, A. G., and Nussenzweig, V., J. Exp. Med. 145,357, 1977. 24. Platsoucas, C. D., Good, R. A., and Gupta, S., Proc. Natl. Acad. Sci. USA 76,1972, 1979. 25. Dickler, H. B., J. Exp. Med. 140,508, 1974. 26. Pichler, W. J., and Knapp, W., &and. J. Immunol. 1, 105, 1978. 27. Gupta, S., Platsoucas, C. D., and Good, R. A., Proc. Natl. Acad. Sci. USA 76,4025, 1979. 28. West, W. H., Cannon, G. B., Kay, H. D., Bonnard, G. D., and Herberman, R. B., J. Immunol. 118,
355,1977. 29, Perussia, B., Starr, S., Abraham, S., Fanning, V., and Trinchieri, G., J. Immunol. 130,2133, 1983. 30. Perez, P., Hoffman, R. W., Titus, J. A., and Segal, D. M., J. Exp. Med. 103,100, 1986. 3 1. Zarling, J. M., Dierckins, M. S., Sevenich, E. A., and Clouse, K. A., J. Immunol. 127,2118, 1981. 32. Bennink, J. R., Yowdell, J. W., Smith, G. L., Moller, C., and Moss, B., Nature (London) 311, 578,
1984. 33. Zarling, J. M., Eichberg, J. W., Moran, P. A., McClure, J., Sridhar, P., and Hu, S-L., J. Immunol. 139,
988,1987. 34. McMichael, A. J., Michie, C. A., Gotch, F. M., Smith, G. L., and Moss, B., J. Gen. Virol. 67, 7 19,
1986. 35. Kannagi, M., Sugamura, K., Kinoshita, K. I., Uchino, H., and Hinuma, Y., J. Immunol. 133, 1037,
1984. 36. Palacios, R., J. Immunol. 128,337, 1982. 37. Schwab, R., Crow, M. K., Russo, C. R., and Weksler, M. D., J. Immunol. 135,17 14,1985. 38. Rola-Pleszczynski, M., and Lemaire, I., J. Immunol. 135,3958, 1985. 39. Van Waune, J. P., DeMey, J. R., and Goossens, J. G., J. Immunol. 124,2708, 1980. 40. Kaneoka, H., Perez-Rojas, G., Sasasuki, T., Benike, C. J., and Engleman, E. G., J. Immunol. 131, 158,
1983. 41, Kupper, T., Horowitz, M., Lee, F., Robb, R., and Flood, P. M., J. Immunol. 138,4280, 1987. 42. Itoh, K., Balch, C. M., and Platsoucas, C. D., Cell. Immunol., in press. 43. Cosimi, A. B., Colvin, R. B., Burton, R. C., Rubin, R. H., Goldstein, G., Kung, P. C., Hansen, P.,
Delmonico, F. L., and Russell, P. S., N. Engl. J. Med. 305,308, 1981. 44. Ortho Multicenter Transplant Study Group, N. Engl. J. Med. 313,337, 1985.