Page 1
Journal of General Virology
Detection of exosomal prions in blood by immunochemistry techniques--Manuscript Draft--
Manuscript Number: JGV-D-14-00185R2
Full Title: Detection of exosomal prions in blood by immunochemistry techniques
Short Title: Exosomal prions in blood
Article Type: Short Communication
Section/Category: TSE Agents
Corresponding Author: Maurizio PocchiariDepartment of Cell Biology and Neurosciences, Istituto Superiore di SanitàRoma, ITALY
First Author: Francesca Properzi
Order of Authors: Francesca Properzi
Mariantonia Logozzi
Hanin Abdel-Haq
Cristina Federici
Luana Lugini
Tommaso Azzarito
Ilaria Cristofaro
Daniela di Sevo
Elena Ferroni
Franco Cardone
Massimo Venditti
Marisa Colone
Emmanuel Comoy
Valérie Durand
Stefano Fais
Maurizio Pocchiari
Abstract: In most forms of prion diseases blood is infectious, but the detection byimmunochemistry techniques of the only available marker of infection (the misfoldedprion protein, PrPTSE) in blood remains elusive. We developed a novel method for thedetection of PrPTSE in blood of prion-infected rodents based on the finding thatPrPTSE is associated with plasma exosomes. However, further purification of exosomeon sucrose gradient was necessary for removing plasma immunoglobulins, whichinterfere with PrPTSE masking its detection by immunochemistry. Finally, we reportthat about 20% of plasma infectivity is associated with exosomes.
Powered by Editorial Manager® and ProduXion Manager® from Aries Systems Corporation
JGV Papers in Press. Published March 24, 2015 as doi:10.1099/vir.0.000117
Page 2
Detection of exosomal prions in blood by immunochemistry techniques 1
Running title: Exosomal prions in blood 2
Francesca Properzi, Mariantonia Logozzi, Hanin Abdel-Haq, Cristina Federici, Luana Lugini, Tommaso 3
Azzarito, Ilaria Cristofaro, Daniela di Sevo, Elena Ferroni, Franco Cardone, Massimo Venditti, Marisa Colone, 4
Emmanuel Comoy, Valérie Durand, Stefano Fais and Maurizio Pocchiari 5
Affiliations 6
Francesca Properzi, Hanin Abdel-Haq, Ilaria Cristofaro, Daniela di Sevo, Elena Ferroni, Franco Cardone, 7
Massimo Venditti, and Maurizio Pocchiari, Department of Cell Biology and Neurosciences, Istituto 8
Superiore di Sanità, Viale Regina Elena 299, 00161, Rome, Italy. 9
Mariantonia Logozzi, Cristina Federici, Luana Lugini, Tommaso Azzarito, and Stefano Fais, Department of 10
Therapeutic Research and Medicines Evaluation, Istituto Superiore di Sanità, 00161 Rome, Italy; 11
Marisa Colone, Department of Technologies and Health, Istituto Superiore di Sanità, 00161 Rome, Italy 12
Emmanuel Comoy and Valérie Durand, Institute of Emerging Diseases and Innovative Therapies, CEA, 13
Fontenay-aux-Roses, France 14
15
Corresponding author 16
Maurizio Pocchiari 17
Email: [email protected] 18
Telephone: +39 06 49903203 19
Fax: +39 06 49903805 20
21
22
23
Manuscript Including References (Word document)Click here to download Manuscript Including References (Word document): Properzi_text_revised_clean.docx
Page 3
2
Word Count Summary: 93 24
Word Count Text: 2411 25
Tables: 0 26
Figures: 3 27
Suppl. Figures: 4 28
29
Page 4
3
Summary paragraph 30
In most forms of prion diseases blood is infectious, but the detection by immunochemistry techniques of 31
the only available marker of infection (the misfolded prion protein, PrPTSE) in blood remains elusive. We 32
developed a novel method for the detection of PrPTSE in blood of prion-infected rodents based on the 33
finding that PrPTSE is associated with plasma exosomes. However, further purification of exosome on 34
sucrose gradient was necessary for removing plasma immunoglobulins, which interfere with PrPTSE masking 35
its detection by immunochemistry. Finally, we report that about 20% of plasma infectivity is associated with 36
exosomes. 37
38
Page 5
4
Transmissible spongiform encephalopathy (TSE) or prion diseases are fatal neurodegenerative disorders 39
affecting humans and animals caused by unique infectious particles (prions), which are mainly composed 40
by misfolded forms of the cellular prion protein (PrPC). PrPC is constitutively expressed, but converts to the 41
misfolded noxious structure (PrPTSE) either spontaneously or upon exposure to prions (Weissmann, 2004). 42
Prions are efficiently transmitted via blood in most animal models of TSE diseases (Brown et al., 2001) 43
(Houston et al., 2008) and in humans with variant and possibly sporadic CJD (Llewelyn et al., 2004) (Wroe 44
et al., 2006) (Puopolo et al., 2011) (Douet et al., 2014). Prion infectivity in blood is associated with both 45
cellular and non cellular components (Gregori et al., 2006) with estimated titres in plasma ranging from 1 46
to 10 infectious unit/ml at disease onset (Brown et al., 2001) (Douet et al., 2014). Although plasma has 47
been shown to carry infectivity, the identification of PrPTSE in infected animals and humans has been elusive 48
and limited to the use of highly sensitive (Castilla et al., 2005) (Saá et al., 2006) (Pan et al., 2007) (Orrú et 49
al., 2011) or complex (Edgeworth et al., 2011) methodologies. Because blood components likely interfere 50
with PrPTSE detection (Hartwell et al., 2005) (Abdel-Haq, 2014), we thought to bypass this difficulty by 51
looking at plasma exosomes. Exosomes are nanovesicles that carry RNAs, proteins, lipids, other metabolites 52
(Théry et al., 2009) (Fais et al., 2013), PrPC and, in prion-infected hosts, PrPTSE and infectivity (Fevrier et al., 53
2004) (Alais et al., 2008) (Coleman et al., 2012) (Saá et al., 2014). 54
We first collected blood from intracerebrally 263K scrapie-infected hamsters (n=125) (Kimberlin and 55
Walker, 1977) or from uninfected and healthy animals as controls. Plasma was separated from blood cells 56
by centrifugation at 5,000 g for 30 minutes at room temperature. Single plasma fractions from infected 57
hamsters were pooled to a final volume of 250 ml and used for the measurement of infectivity and 58
purification of exosomes for the detection of PrPTSE by immunochemistry assays (Fig. 1). Bioassay was 59
performed by intracerebral inoculation of 124 six weeks-old female Syrian hamsters with 50 µl of thawed 60
plasma, under anesthesia with ketamine/xylazine (75 and 10 mg/kg respectively) mixture. Animals were 61
housed in accordance to the national guidelines in level-3 animal care facilities (agreement numbers A 92-62
032-02 for animal care facilities, 92-189 for animal experimentation, CEA, France) and regularly monitored. 63
Animals with clinical signs or with no disease up to 600-700 days were euthanized, the brain removed and 64
tested for the presence of PrPTSE by the ELISA test as previously described (Barret et al., 2003). Briefly, PrPTSE 65
Page 6
5
was extracted from brains by the Bio-Rad purification kit (USA), digested with proteinase K, and added into 66
anti-PrP SAF53 (Bertin Pharma, France) monoclonal antibody (mAb)-coated ELISA wells. Bound PrPTSE was 67
detected by incubation with the 11C6 (Bertin Pharma, France) anti-PrP mAb coupled with 68
acetylcholinesterase, and revealed using the Ellman substrate at 414 nm optical wavelength. 69
Eighteen animals resulted positive for scrapie (time to death, mean, 265 days; s.e.m., 23 days) with a 70
corresponding infectivity titre of 3.1±0.7 infectious dose (ID) per ml (Supplementary Fig. 1), in line with 71
previously published results on plasma samples (Gregori et al., 2006) (Brown et al., 2001). Infectivity titre 72
(ID) was calculated by the limiting dilution method according to the Poisson distribution (Gregori et al., 73
2006). 74
From the same pool of plasma, exosomes were isolated from 5 ml aliquots of control and infectious plasma 75
pool by a previously published protocol (Caby et al., 2005). Briefly, exosome plasma pellets were prepared 76
by serial centrifugations; the resulting pellets were washed in PBS, filtered through a 0.22 μm filter 77
(Millipore, USA) and ultracentrifuged at 110,000g for 1 h. The exosome pellets were re-suspended in PBS, 78
lysis buffer or in 2.5 M sucrose buffer, for further analyses as indicated in Fig. 1. Antibodies used for 79
exosomal marker identification were the anti-Flotillin, anti-Tsg101, and anti-mouse CD81 (Santa Cruz, 80
Germany). For PrP identification the primary antibodies used were SAF84 and biotinylated-SAF84 (Bertin 81
Pharma, France), 3F4 (Millipore, Italy), and D18 (gift from R. A. Williamson, The Scripps, Florida). Western 82
blot analyses were performed as described (Lee et al., 2000) and the consistency of these preparations 83
confirmed by the detection of exosomal markers (Fig. 2a). Assuming a similar ratio between infectivity and 84
PrPTSE molecules in blood and in brain tissue (about 0.1 to 1 pg PrPTSE/infectious unit) (Brown et al., 2001), 85
we estimated that 25 µl of exosomal plasma pellets (EPPs), corresponding to 5 ml of the original plasma 86
volume and 15.5 IDs, were suitable for detecting PrPTSE by standard western blot analyses (Lee et al., 2000). 87
In EPPs from control and prion-infected samples, we found that anti-PrP antibodies recognised the Ig light 88
chain (26 kD), and the IgG (53 kD) and IgM (65 kD) heavy chains as previously reported (Hartwell et al., 89
2005). These bands were resistant to an amount (50 µg/ml) of proteinase K (PK) that is largely sufficient to 90
remove the bulk of tissue proteins in brain samples (Fig. 2). The atypical bands were seen irrespectively of 91
the anti-PrP primary antibody used (Supplementary Fig. 2) and were not detected in the absence of primary 92
Page 7
6
antibodies (Fig. 2a). They were specific to plasma (Fig. 2a) and observed in EPPs of other species including 93
humans (data not shown) and wild type mice (C57BL/10) (Fig. 2a and supplementary Fig. 2). They were also 94
reproduced in EPPs of PrP knock-out 129/Ola mice (gift from Dr Di Bari and Dr Agrimi, Istituto Superiore di 95
Sanità, Rome, Italy) (Fig. 2a and supplementary Fig. 2) clearly indicating a cross-reaction of the primary 96
antibodies with plasmatic PK-resistant material, unrelated to PrP epitopes. Finally, when primary anti-PrP 97
antibodies were pre-absorbed with an excess of Syrian hamster 23-231 recombinant PrP (recPrP) (5:1), 98
western blot membranes were negative, demonstrating that pre-absorbed primary antibodies are unable 99
to bind PK-resistant Igs (IgRES). These data suggest that the levels of endogenous PrPC in plasma (in the 100
range of pg/ml) is not sufficient to replace the non-specific binding of largely represented plasma proteins 101
(in the range of mg/ml). 102
Stringent PK conditions (100 µg/ml), however, digested high molecular weight IgRES bands (Fig. 2a) 103
allowing the recognition of three bands of the same molecular weight of full length prion protein with a 104
glycoform ratio similar to that observed in hamster brain (Fig. 2a) (Meade-White et al., 2009). We then 105
further purified EPPs on a sucrose cushion starting from 15 ml of plasma to verify that these bands 106
effectively belong to full-length prion protein. Continuous sucrose gradient for the isolation and purification 107
of exosomes was performed as published (Raposo et al., 1996), with some modifications. Briefly, purified 108
exosomes were re-suspended in 2 ml of 2.5 M sucrose, 20 mM Hepes/NaOH, pH 7.2. A linear sucrose 109
gradient (2.0-0.25 M sucrose) was layered on top of the exosomes suspension in a UltraClear™ tube and 110
the sample was centrifuged at 110,000 g for 16 h at 4oC in a Beckman-Coulter (USA) SW41Ti rotor. Gradient 111
fractions (12 x 1 ml) were collected from the top of the tube and washed with PBS. Fractions density was 112
evaluated using an Abbé Refractometer (Carl Zeiss, Germany). 113
Silver staining after PAGE and quantification of bands using ImageJ (NIH, available at 114
http://imagej.nih.gov/ij/) showed that after sucrose gradient ultracentrifugation the bulk of the 115
immunoglobulins was retained at high-density fractions (1.64 to 1.70 g/ml), while exosomal fractions (1.11 116
to 1.13 g/ml) had significantly lower content of IgRES. On sucrose gradient purified EPPs, di-glycosylated 117
PrPC bands (37 kD) were detected at densities of 1.13 g/ml (Fig. 3) and disappeared after PK treatment (Fig. 118
3). This fraction was also positive on western blot to standard exosomal markers (Tsg101, Flotillin and 119
Page 8
7
CD81) and negative to ER and nucleus markers (calnexin and nucleoporin) indicating the purity of exosomal 120
preparation after sucrose gradient isolation (Fig. 3). As controls, ER and nucleus markers resulted positive in 121
the brain homogenate (Fig. 3). 122
In infectious EPPs, 37 kD PrP bands were observed in exosomal fractions at densities of 1.13 g/ml (Fig. 3). 123
After treatment with PK, PrP-positive bands showed the characteristic shift in molecular weights of PrPTSE 124
(27-30 kD) with a glycoform ratio similar to that observed in 263K-infected brain homogenate (Fig. 3). The 125
presence of PrPC on plasma exosomes was further ascertained by electron microscopy. Immunoelectron 126
microscopy was performed on isolated sucrose gradient fractions of hamster plasma exosomes. A drop of 127
exosome suspension was placed onto a formvar-carbon-coated grid. After primary antibody (SAF84) 128
incubation exosomes were labelled with anti-rabbit IgG or anti-mouse IgG 5nm diameter-gold particle-129
conjugated antibody (Sigma-Aldrich, USA). Grids were negatively stained with uranyl acetate solution and 130
examined with a Philips 208 transmission electron microscope (FEI Company, The Netherlands). In the 1.13 131
g/ml fraction there was presence of nanovesicles, which after immunogold staining, were PrP-positive 132
particles on the surface of nanovesicles (~50 nm) (Supplementary Fig. 3a and 3b). On these samples, 133
nanovesicles of the same size were positive to the standard exosomal marker Rab5b and no PrP staining 134
was observed in fractions of lower and higher densities (Supplementary Fig. 3c). 135
Finally, to further confirm the presence of prion in plasma exosomes we measured scrapie infectivity. 136
However, for this experiment we used the mouse-adapted 139A scrapie strain (Dickinson, 1976), because 137
139A, but not 263K infectivity can be quickly and accurately measured by using the End Point Scrapie Cell 138
Assay (SCEPA) (Klöhn et al., 2003). Susceptible CAD5 cells (gift from Prof. Charles Weissmann, The Scripps, 139
Florida, USA) were seeded in wells of 96-well plate and exposed to 12 or more aliquots of the 139A prion-140
containing samples for 3 days, grown to confluence and split 1:3 three times and 1:10 twice. When the cells 141
reached confluence, 25,000 cells were transferred to membranes of ELISpot plates and PrPTSE-containing 142
cells were identified as spots using the D18 antibody (Supplementary Fig. 4). A well is considered positive 143
when the following two criteria are fulfilled. Firstly, the number of positive spots must be higher then the 144
mean plus 5 times standard deviations of unseeded wells (background value). Secondly, prion propagation, 145
revealed by an increased number of positive spots after subsequent passages, must be evident. The ID was 146
Page 9
8
calculated considering the number of negative wells over positive by using a Poisson distribution. A titred 147
brain homogenate is used as positive control to monitor variations caused by antibodies. Nanovesicles 148
were isolated from 3 ml of plasma of 139A-infected mice at terminal stage. We found 1 of 6 positive wells 149
for exosomal samples and 4 of 48 positive wells for the 1:5 dilution of plasma supernatant. Accordingly to 150
the Poisson distribution for rare events, the IDs contained in the processed volume (3 ml) were 1.1 for the 151
exosomal pellet and 4.3 for plasma supernatant. These findings confirm that in vivo prions are associated to 152
nanovesicles similarly to what previously observed in vitro (Fevrier et al., 2004) (Alais et al., 2008) (Coleman 153
et al., 2012) (Fauré et al., 2006) (Vella et al., 2007) and that in plasma about 20% of total infectivity is 154
associated to exosomes leaving open the issue on how the remaining infectivity is distributed. 155
156
The most important finding of our work is the successful identification of PrPTSE in plasma of scrapie-157
infected hamsters by standard western blot technique performed on exosomal fractions. This result 158
enforces previous data showing the presence of PrPTSE in blood exosomes by the protein misfolding cyclic 159
amplification (PMCA) method (Saá et al., 2014), in plasma by the PMCA (Castilla et al., 2005) and RT-QuiC 160
assays (Orrú et al., 2011) and in human vCJD patients by the solid-state binding matrix assay (Edgeworth et 161
al., 2011), but adds the important novel information that PrPTSE, similarly to infectivity (Brown et al., 2001) 162
is detectable in blood without using amplification techniques. The detection of PrPTSE by the standard 163
western blot technique was only possible after reducing the interference of IgRES by exosomal pellet 164
fractionation on a sucrose gradient. IgRES compete for the primary antibody binding with endogenous PrP 165
epitopes crucially interfering with PrPTSE detection in blood. This result confirms that blood components 166
interfere with prion diagnosis (Hartwell et al., 2005) (Abdel-Haq, 2014) reducing the sensitivity and 167
specificity of the various tested assays. Moreover, these findings might explain the discrepancy between 168
the excellent performances of tests accomplished on spiked blood and their failure on endogenously 169
infected blood that is likely related to different interaction with plasma Igs between exogenous and 170
endogenous PrPTSE. 171
172
173
Page 10
9
Acknowledgments 174
We thank Dr Anna Ladogana for critical revision of the manuscript, Giuseppe Loreto for the preparations of 175
the figures, Dr Agnese Molinari for support with EM, Nick Verity for supplying the SCEPA samples, and Dr 176
Alessandra Garozzo for editorial assistance. 177
This work was partially supported by the Istituto Superiore di Sanità, Rome, Italy and by the Alliance 178
Biosecure Project “Plasmasecure”. 179
180
181
Page 11
10
REFERENCES 182
Abdel-Haq, H. (2014) Factors intrinsic and extrinsic to blood hamper the development of a routine blood 183
test for human prion diseases. J Gen Virol doi: 10.1099/vir.0.070979-0 184
Alais, S., Simoes, S., Baas, D., Lehmann, S., Raposo, G., Darlix, J. L. & Leblanc, P. (2008). Mouse 185
neuroblastoma cells release prion infectivity associated with exosomal vesicles. Biol Cell 100, 603–615. 186
Barret, A., Tagliavini, F., Forloni, G., Bate, C., Salmona, M., Colombo, L., De Luigi, A., Limido, L., Suardi, S. 187
& other authors. (2003). Evaluation of quinacrine treatment for prion diseases. J Virol 77, 8462–8469. 188
Brown, P., Cervenáková, L. & Diringer, H. (2001). Blood infectivity and the prospects for a diagnostic 189
screening test in Creutzfeldt-Jakob disease. J Lab Clin Med 137, 5–13. 190
Caby, M.-P., Lankar, D., Vincendeau-Scherrer, C., Raposo, G. & Bonnerot, C. (2005). Exosomal-like vesicles 191
are present in human blood plasma. Int Immunol 17, 879–887. 192
Castilla, J., Saá, P. & Soto, C. (2005). Detection of prions in blood. Nat Med 11, 982–985. 193
Coleman, B. M., Hanssen, E., Lawson, V. A. & Hill, A. F. (2012). Prion-infected cells regulate the release of 194
exosomes with distinct ultrastructural features. FASEB J 26, 4160–4173. 195
Dickinson, A.G. (1976) Scrapie in sheep and goats. In Slow Virus Diseases of Animals and Man. Frontiers of 196
Biology series, vol. 44, chapter 10, pp. 209-241. Edited by R. H. Kimberlin. Amsterdam and New York: North 197
Holland. 198
Douet, J.Y., Zafar, S., Perret-Liaudet, A., Lacroux, C., Lugan, S., Aron, N., Cassard, H., Ponto, C., Corbière, F. 199
& other authors. (2014). Detection of infectivity in blood of persons with variant and sporadic Creutzfeldt-200
Jakob disease. Emerg Infect Dis 20, 114-117. 201
Edgeworth, J. A., Farmer, M., Sicilia, A., Tavares, P., Beck, J., Campbell, T., Lowe, J., Mead, S., Rudge, P. & 202
other authors. (2011). Detection of prion infection in variant Creutzfeldt-Jakob disease: a blood-based 203
assay. Lancet 377, 487–493. 204
Page 12
11
Fais, S., Logozzi, M., Lugini, L., Federici, C., Azzarito, T., Zarovni, N. & Chiesi, A. (2013). Exosomes: the ideal 205
nanovectors for biodelivery. Biol Chem 394, 1–15. 206
Fauré, J., Lachenal, G., Court, M., Hirrlinger, J., Chatellard-Causse, C., Blot, B., Grange, J., Schoehn, G., 207
Goldberg, Y. & other authors. (2006). Exosomes are released by cultured cortical neurones. Mol Cell 208
Neurosci 31, 642–648. 209
Fevrier, B., Vilette, D., Archer, F., Loew, D., Faigle, W., Vidal, M., Laude, H. & Raposo, G. (2004). Cells 210
release prions in association with exosomes. Proc Natl Acad Sci USA 101, 9683–9688. 211
Gregori, L., Gurgel, P. V., Lathrop, J. T., Edwardson, P., Lambert, B. C., Carbonell, R. G., Burton, S. J., 212
Hammond, D. J. & Rohwer, R. G. (2006). Reduction in infectivity of endogenous transmissible spongiform 213
encephalopathies present in blood by adsorption to selective affinity resins. Lancet 368, 2226–2230. 214
Hartwell, R. C., Nelson, M. S., Kislan, M. M., Stenland, C. J., Miller, J. L. C., Pifat, D. Y., Petteway, S. R., Jr & 215
Cai, K. (2005). An improved Western blot assay to assess the clearance of prion protein from plasma-216
derived therapeutic proteins. J Virol Methods 125, 187–193. 217
Houston, F., McCutcheon, S., Goldmann, W., Chong, A., Foster, J., Sisó, S., González, L., Jeffrey, M. & 218
Hunter, N. (2008). Prion diseases are efficiently transmitted by blood transfusion in sheep. Blood 112, 219
4739–4745. 220
Kimberlin RH, Walker C. (1977). Characteristics of a short incubation model of scrapie in the golden 221
hamster. J Gen Virol 34, 295-304. 222
Klöhn, P.-C., Stoltze, L., Flechsig, E., Enari, M. & Weissmann, C. (2003). A quantitative, highly sensitive cell-223
based infectivity assay for mouse scrapie prions. Proc Natl Acad Sci USA 100, 11666–11671. 224
Lee, D. C., Stenland, C. J., Hartwell, R. C., Ford, E. K., Cai, K., Miller, J. L., Gilligan, K. J., Rubenstein, R., 225
Fournel, M. & Petteway, S. R., Jr. (2000). Monitoring plasma processing steps with a sensitive Western blot 226
assay for the detection of the prion protein. J Virol Methods 84, 77–89. 227
Page 13
12
Llewelyn, C. A., Hewitt, P. E., Knight, R. S. G., Amar, K., Cousens, S., Mackenzie, J. & Will, R. G. (2004). 228
Possible transmission of variant Creutzfeldt-Jakob disease by blood transfusion. Lancet 363, 417–421. 229
Meade-White, K. D., Barbian, K. D., Race, B., Favara, C., Gardner, D., Taubner, L., Porcella, S. & Race, R. 230
(2009). Characteristics of 263K scrapie agent in multiple hamster species. Emerging Infect Dis 15, 207–215. 231
Orrú, C. D., Wilham, J. M., Raymond, L. D., Kuhn, F., Schroeder, B., Raeber, A. J. & Caughey, B. (2011). 232
Prion disease blood test using immunoprecipitation and improved quaking-induced conversion. MBio 2, 233
e00078–00011. 234
Pan, T., Sethi, J., Nelsen, C., Rudolph, A., Cervenakova, L., Brown, P. & Orser, C. S. (2007). Detection of 235
misfolded prion protein in blood with conformationally sensitive peptides. Transfusion 47, 1418–1425. 236
Puopolo, M., Ladogana, A., Vetrugno, V. & Pocchiari, M. (2011). Transmission of sporadic Creutzfeldt-237
Jakob disease by blood transfusion: risk factor or possible biases. Transfusion 51, 1556–1566. 238
Raposo, G., Nijman, H. W., Stoorvogel, W., Liejendekker, R., Harding, C. V., Melief, C. J. & Geuze, H. J. 239
(1996). B lymphocytes secrete antigen-presenting vesicles. J Exp Med 183, 1161–1172. 240
Saá, P., Castilla, J. & Soto, C. (2006). Presymptomatic detection of prions in blood. Science 313, 92–94. 241
Saá, P., Yakovleva, O., De Castro, J., Vasilyeva, I., De Paoli, S. H., Simak, J. & Cervenakova, L. (2014). First 242
demonstration of PrPTSE in extracellular vesicles from plasma of mice infected with mouse-adapted variant 243
Creutzfeldt-Jakob disease by in vitro amplification of misfolded prion protein. J Biol Chem 289, 29247-244
29260. 245
Théry, C., Ostrowski, M. & Segura, E. (2009). Membrane vesicles as conveyors of immune responses. Nat 246
Rev Immunol 9, 581–593. 247
Vella, L. J., Sharples, R. A., Lawson, V. A., Masters, C. L., Cappai, R. & Hill, A. F. (2007). Packaging of prions 248
into exosomes is associated with a novel pathway of PrP processing. J Pathol 211, 582–590. 249
Weissmann, C. (2004). The state of the prion. Nat Rev Microbiol 2, 861–871. 250
Page 14
13
Wroe, S. J., Pal, S., Siddique, D., Hyare, H., Macfarlane, R., Joiner, S., Linehan, J. M., Brandner, S., 251
Wadsworth, J. D. F. & other authors. (2006). Clinical presentation and pre-mortem diagnosis of variant 252
Creutzfeldt-Jakob disease associated with blood transfusion: a case report. Lancet 368, 2061–2067. 253
254
Page 15
14
Figure Legends 255
Figure 1 Diagram of the main experimental procedure 256
Infectivity of plasma pool was measured by bioassay. Positive animals were identified by ELISA detection of 257
PrPTSE on brain samples. Prion infected exosomal plasma pellets (EPPs) were prepared, treated with or 258
without proteinase K (PK) and analysed by western blot before and after sucrose gradient fractionation. IC, 259
intracerebral 260
261
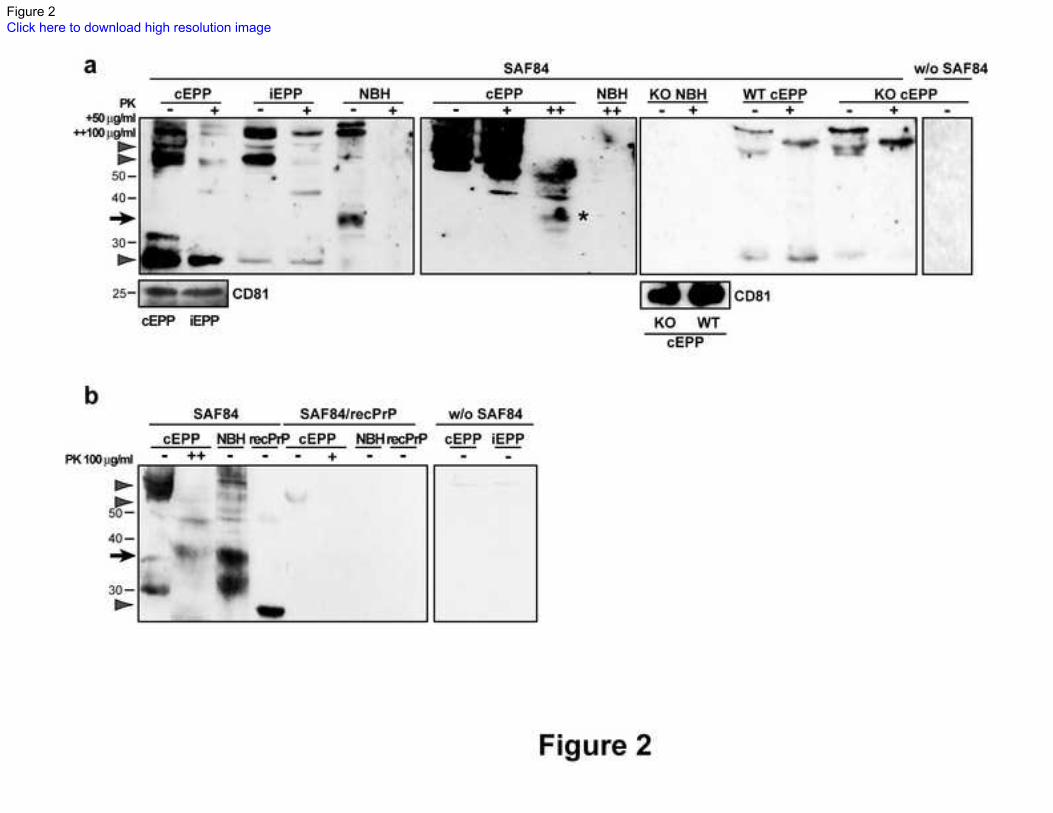
Figure 2. Western blots of PrP in infected and control exosomal plasma pellets 262
a PrP positive immunoglobulin (Ig) bands were resistant to standard (50µg/ml; +) proteinase K (PK) 263
digestion (arrowheads) in both exosomal plasma pellets from control (cEPP) and infected (iEPP) 264
samples. The same conditions digested 37 kD PrP (arrow) of normal brain homogenate (NBH). After 265
stringent treatment with 100 µg/ml of PK (++), PrP-like bands were also seen, possibly due to 266
degradation (asterisk). EPPs of PrP knock-out (KO) mouse are positive to anti PrP SAF84 antibody. CD81 267
positive cEPP of both wild type (WT) and KO mice contain PrP positive Ig bands (arrowheads) that are 268
resistant to PK. No staining is observed without (W/O) the primary SAF84 antibody and in NBH. The 269
exosomal marker CD81 is shown in the lower panels. (n=3). 270
b Pre-absorption of anti-PrP SAF84 antibody with recombinant PrP (recPrP) (5:1) removes the recPrP 271
band, PrP bands (arrows) of NBH, and Ig bands of EPPs (arrowheads). No staining was observed in 272
absence of the primary SAF84 antibody. (n=3). 273
Numbers on the left indicate molecular weights in kD. 274
275
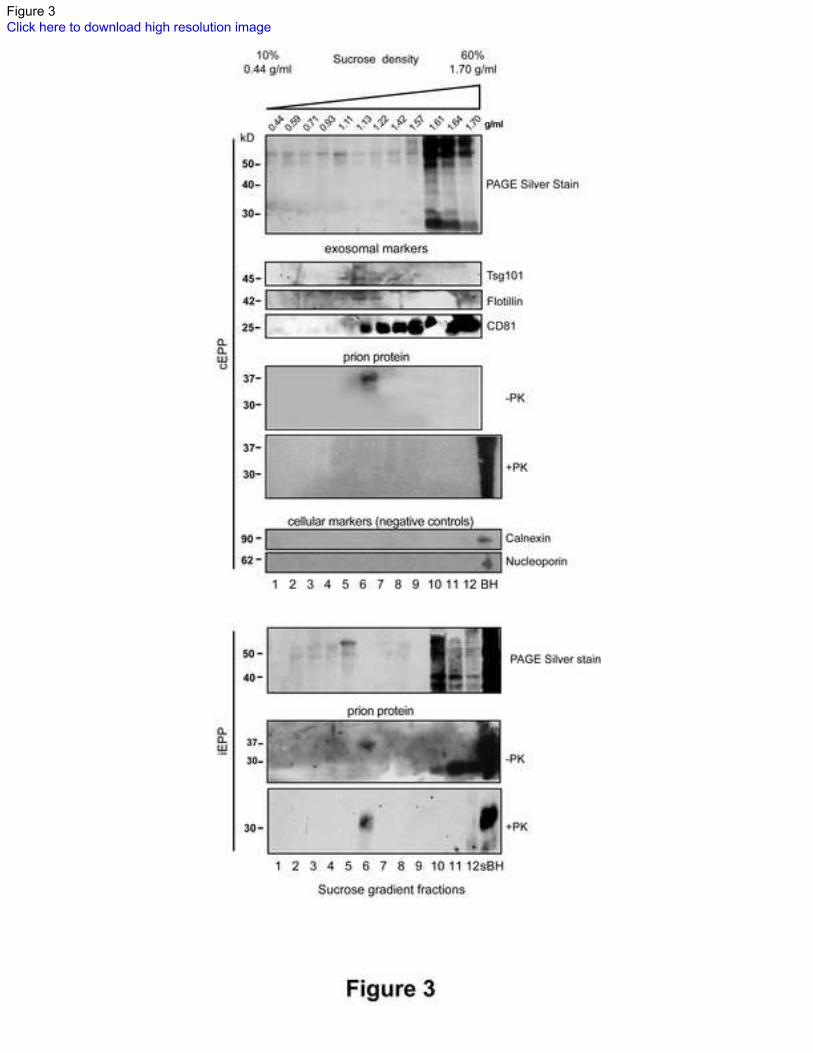
Figure 3 Silver staining and western blots of sucrose gradient fractions of infected and control 276
exosomal plasma pellets 277
PAGE silver staining reveals that the bulk of plasma proteins are retained in the high-density fractions of 278
both exosomal plasma pellets from control (cEPP) and infected (iEPP) samples. 279
Western blots with SAF84 antibody show the presence of PrPC in fraction 6 (1.13 g/ml) of cEPPs (n=3), 280
which is abolished by the treatment with proteinase K (PK). PK resistant PrPTSE is shown in fraction 6 of 281
Page 16
15
iEPPs. Western blots of sucrose fractions for exosomal markers (Tsg101, flotillin and CD81) show that 282
exosomes float in fractions 5 to 8. Absence of calnexin and nucleoporin indicates no cellular debris 283
contamination. nBH, normal hamster brain homogenate; sBH, scrapie-infected hamster brain homogenate. 284
Page 17
Figure 1Click here to download high resolution image
Page 18
Figure 2Click here to download high resolution image
Page 19
Figure 3Click here to download high resolution image
Page 20
Supplementary Material FilesClick here to download Supplementary Material Files: Supplementary Figure Legends.pdf