Diversity analysis of Lower Jurassic bivalves of the Andean Basin and the Pliensbachian-Toarcian mass extinction MARTIN ABERHAN AND FRANZ THEODOR FORSICH LETHAIA Aberhan, M. & Fiirsich, F.T. 1997 01 15: Diversity analysis of Lower Jurassicbivalves of the An- dean Basin and the Pliensbachian-Toarcian mass extinction. Lethaia, Vol. 29, pp. 181-195. Oslo. ISSN 0024-1 164. In the Andean Basin of southern South America marine bivalves show a marked decrease in diversity across the Pliensbachian-Toarcian boundary. This observation is in contrast to earlier statements, according to which a drop in diversity of marine organisms at the boundary is restricted to epicontinental seas of Western Europe. The decrease in bivalve diversity within the Andean Basin is largely due to the extinction of endemics and of some cosmopolitan species, less commonly to the Lazarus effect and local disappearances of taxa. As the only two regions for which detailed data are available (Andean Basin and Western Europe) exhibit similar diver- sity patterns across the boundary, the extinction event is not a regional feature, but appears to be global. In both regions the mass extinction correlates with sea-level highstand and wide- spread oxygen-poor deep-shelf environments suggesting a causative relationship. As shallow, largely well-aerated shelf environments in the Andean Basin also exhibit, though less markedly, a drop in bivalve diversity across the boundary, oxygen deficiency cannot be the sole cause of the mass extinction. Rather, in our model we assume sea-level changes and the resulting ocea- nographic and biotic changes to be the ultimately controlling factors of species diversity pat- terns. The apparent global character of the Pliensbachian-Toarcian mass extinction fits the periodicity hypothesis. However, as the mass extinction does not appear to have been cata- strophic and as there is no indication of impact(s) of extraterrestrial bodies, the oceanographic model proposed here is regarded as a more plausible (because it is more parsimonious) hypoth- esis. ODiversity, bivalves, Jurassic, mass extinction, Andean Basin. Martin Aberhan and Franz T. Fursich, Insritut fur Paliiontologie der Universitiit Wurzburg, Pleichenvall I, D-97070 Wurzburg, Germany; 20th April, 1995; revised 2lst December, 1995. Within the Phanerozoic, five major mass extinctions are generally recognized. Including additional, less spectacu- lar events, nine geologically brief periods of mass extinc- tion, among them the famous Cretaceous-Tertiary boundary event, have been defined for the Mesozoic and Cenozoic. These mass extinctions are characterized by a drastic decrease of generic diversity of marine inverte- brates (e.g., Raup & Sepkoski 1984; Sepkoski & Raup 1986; Sepkoski 1989). The nine mass extinctions form the basis for the periodicity hypothesis, according to which the events are separated by average periods of 26.2 Ma. The periodicity hypothesis implies a common cause for all mass extinctions, be it impacts of extraterrestrial bod- ies or increased activity of mantle plumes (Hallam 1987b). As the best studied, end-Cretaceous mass extinc- tion is, at least partially, explained by impact scenarios, extraterrestrial causes should under this hypothesis also account for the remaining mass extinctions. Two of these events occurred within the Jurassic: the Early Toarcian and end-Tithonian extinction events (Hallam 1986). Hallam (1986, 1987a), however, argued that these two events are only of regional extent and can be explained by severe reductions in habitat area caused either by marine regressions or spread of anoxic bottom waters. Hallam’s (1986, 1987a) data base for the Early Toarcian mass extinction consisted of genera of ammo- nites and species of bivalves, brachiopods, ostracodes, and foraminifers from northwestern Europe. The macrofau- nal groups exhibited drastic declines in diversity (Hallam 1987a), whereas no significant change in diversity of bivalve genera was recorded for the same time interval in South America (Hallam 1983). Hallam’s conclusions seriously undermine the perio- dicity hypothesis and throw doubts on a common cause of mass extinctions. This is the reason why we regard it as crucial to re-examine diversity changes throughout the Lower Jurassic in regions outside Europe. Based on exten- sive collections made by the authors and on an exhaustive survey of the literature, we present and discuss the changes in Lower Jurassic species diversity of the most
Transcript
Diversity analysis of Lower Jurassic bivalves of the Andean Basin and the Pliensbachian-Toarcian mass extinction MARTIN ABERHAN AND FRANZ THEODOR FORSICH
LETHAIA Aberhan, M. & Fiirsich, F.T. 1997 01 15: Diversity analysis of Lower Jurassic bivalves of the An- dean Basin and the Pliensbachian-Toarcian mass extinction. Lethaia, Vol. 29, pp. 181-195. Oslo. ISSN 0024-1 164.
In the Andean Basin of southern South America marine bivalves show a marked decrease in diversity across the Pliensbachian-Toarcian boundary. This observation is in contrast to earlier statements, according to which a drop in diversity of marine organisms at the boundary is restricted to epicontinental seas of Western Europe. The decrease in bivalve diversity within the Andean Basin is largely due to the extinction of endemics and of some cosmopolitan species, less commonly to the Lazarus effect and local disappearances of taxa. As the only two regions for which detailed data are available (Andean Basin and Western Europe) exhibit similar diver- sity patterns across the boundary, the extinction event is not a regional feature, but appears to be global. In both regions the mass extinction correlates with sea-level highstand and wide- spread oxygen-poor deep-shelf environments suggesting a causative relationship. As shallow, largely well-aerated shelf environments in the Andean Basin also exhibit, though less markedly, a drop in bivalve diversity across the boundary, oxygen deficiency cannot be the sole cause of the mass extinction. Rather, in our model we assume sea-level changes and the resulting ocea- nographic and biotic changes to be the ultimately controlling factors of species diversity pat- terns. The apparent global character of the Pliensbachian-Toarcian mass extinction fits the periodicity hypothesis. However, as the mass extinction does not appear to have been cata- strophic and as there is no indication of impact(s) of extraterrestrial bodies, the oceanographic model proposed here is regarded as a more plausible (because it is more parsimonious) hypoth- esis. ODiversity, bivalves, Jurassic, mass extinction, Andean Basin.
Martin Aberhan and Franz T. Fursich, Insritut fur Paliiontologie der Universitiit Wurzburg, Pleichenvall I , D-97070 Wurzburg, Germany; 20th April, 1995; revised 2lst December, 1995.
Within the Phanerozoic, five major mass extinctions are generally recognized. Including additional, less spectacu- lar events, nine geologically brief periods of mass extinc- tion, among them the famous Cretaceous-Tertiary boundary event, have been defined for the Mesozoic and Cenozoic. These mass extinctions are characterized by a drastic decrease of generic diversity of marine inverte- brates (e.g., Raup & Sepkoski 1984; Sepkoski & Raup 1986; Sepkoski 1989). The nine mass extinctions form the basis for the periodicity hypothesis, according to which the events are separated by average periods of 26.2 Ma. The periodicity hypothesis implies a common cause for all mass extinctions, be it impacts of extraterrestrial bod- ies or increased activity of mantle plumes (Hallam 1987b). As the best studied, end-Cretaceous mass extinc- tion is, at least partially, explained by impact scenarios, extraterrestrial causes should under this hypothesis also account for the remaining mass extinctions.
Two of these events occurred within the Jurassic: the Early Toarcian and end-Tithonian extinction events
(Hallam 1986). Hallam (1986, 1987a), however, argued that these two events are only of regional extent and can be explained by severe reductions in habitat area caused either by marine regressions or spread of anoxic bottom waters. Hallam’s (1986, 1987a) data base for the Early Toarcian mass extinction consisted of genera of ammo- nites and species of bivalves, brachiopods, ostracodes, and foraminifers from northwestern Europe. The macrofau- nal groups exhibited drastic declines in diversity (Hallam 1987a), whereas no significant change in diversity of bivalve genera was recorded for the same time interval in South America (Hallam 1983).
Hallam’s conclusions seriously undermine the perio- dicity hypothesis and throw doubts on a common cause of mass extinctions. This is the reason why we regard it as crucial to re-examine diversity changes throughout the Lower Jurassic in regions outside Europe. Based on exten- sive collections made by the authors and on an exhaustive survey of the literature, we present and discuss the changes in Lower Jurassic species diversity of the most
182 Martin Aberhan and Franz Theodor Fiirsich LETHAIA 29 (1996)
abundant and diverse group of benthos, the bivalves, from the Andean Basin. Apart from western North Amer- ica, this is the only area outside Europe for which Hallam (1983) produced bivalve data at the genus level. As we shall demonstrate, our data indicate that the Pliens- bachian-Toarcian mass extinction was not a regional phenomenon restricted to Europe but also left a distinct impact in both deep- and shallow-shelf faunas of the Andean region.
Environmental framework of the Lower Jurassic of the Andean Basin
'The Andean Basin of central and northern Chile and west-central Argentina (Fig. 1B) originated as a fault-con- trolled basin at the southwestern margin of the South American craton (e.g., Riccardi 1983; Dalziel 1986; Ricca- rdi et al. 1992). The sea entered parts of the basin in the Late Triassic. With the rise in sea-level during the Early Jurassic, much of the basin became flooded, and marine sediments extended eastwards, e.g., into what is known as the Neuquen embayment of west-central Argentina. Dur- ing the Early Jurassic, a magmatic arc formed to the west, separating the back-arc basin from the open Palaeo-Pacif- ic, without, however, severely restricting the interchange of faunas. Sea-level history of the basin is generally under eustatic control (e.g., Groschke et al. 1988; Hallam 1991) characterized by a rise during the Early Jurassic, reaching its peak in the Bajocian. Superimposed on this general trend are smaller-scale transgressive-regressive cycles with important Early Jurassic transgressive pulses in the Hettangian, Early and Late Sinemurian, and Late Pliensbachian to Early Toarcian, where the Early Jurassic sea-level highstand was reached (Hallam 1991).
The Lower Jurassic basin fill mainly consists of silt- stones, marlstones and mixed siliciclastic-carbonate sed- iments. Coarser-grained siliciclastics and pure carbonates are less common. The sediments represent shallow- to deep-shelf environments, ranging from above the fair weather wave base down to beyond storm influence.
Despite the general deepening trend during the Early Jurassic, some areas remained relatively shallow even dur- ing the Early Toarcian sea-level highstand (e.g., the El Transit0 area of northern Chile, east of Vallenar; Fig. lA), while in many other areas deep-shelf environments pre- vailed during that time interval.
Although we assembled data from the whole basin, most of the information comes from sections situated between 26' and 29"s. In this area, the Lower Jurassic environments are best explained as representing segments of a homoclinal ramp (Aberhan 1992, 1993). Along an onshore-offshore gradient the environments ranged from shallow nearshore sand bars, shallow-shelf mixed
siliciclastic-carbonate substrates to consolidated offshore mid-shelf carbonates, and finally to fine-grained deep- shelf sediments. The Pliensbachian-Toarcian time inter- val, on which we focus in this paper, is represented in deep-shelf areas by dark silty mudstones and marlstones, whereas this type of facies is only of subordinate impor- tance in coeval shallow- to mid-shelf areas.
Methods
A sound taxonomy is the prerequisite for producing meaningful diversity trends through time. Therefore all bivalve taxa have been identified down to species level. Data from the literature have been re-evaluated in order to subject data to a uniform species concept. This was facilitated by recent monographic revisions of early Jurassic bivalve faunas from Argentina (e.g., Damborenea 1987a, b; Leanza 1993) and northern Chile (Aberhan 1994a and unpublished). Additional information was obtained from Hillebrandt ( 1980), Hillebrandt & Schmidt-Effing (1981), Perez (1982), Hillebrandt & Westermann (1985), Damborenea (1990a, b, 1992,1993), Aberhan (1992), Damborenea & Mancenido (1992) and Perez & Reyes (1994). Information contained in range charts of Jurassic bivalves from South America (Damborenea 1990a, b) was also incorporated.
Bivalves were collected from more than 50 fossiliferous localities in northern Chile (Fig. 1A) by the authors and by A. von Hillebrandt, Berlin. The bivalves occur in a wide range of facies types, from fine- and coarse-grained silici- clastics to mixed carbonate-siliciclastics and calcite mud- stones to rudstones. Thus, both the areal extent and the range of facies are of the same order of magnitude as in northwestern Europe.
The data have been compiled in nine time slices repre- senting stages or subdivisions thereof: Hettangian, Early Sinemurian, Late Sinemurian, Early Pliensbachian, Late Pliensbachian, Early Toarcia?, Middle Toarcian, Late Toarcian, and Aalenian. The Aalenian, as the first stage of the Middle Jurassic, was included in our analysis to allow calculation of extinction and turnover rates for the Late Toarcian. The time slices are thought to be of roughly equal duration, although there is not yet a consensus on the exact time of duration of Jurassic stages (see time scales of Haq et al. 1988; Harland et al. 1990; Odin 1992; Gradstein et al. 1994). A finer, zonal, resolution would have been desirable but was rendered impossible due to the lack of more precise biostratigraphic information. On the other hand, these larger time units add to the fidelity of the data base, as potential errors, such as distortions caused by the Signor-Lipps Effect (the likelihood that the last representative of a species is not recovered; Raup 1986) are tuned down.
LETHAIA 29 (1996) Diversity of lurassic bivalves 183
14"
! S o
Antolagasta $& 24".
I 3 0' @ L~ marine Mesozoic
mainly Jurassic r - 2
volcanic rocks - L mainly Jurassic
- 70" 69"
Fig. I . CIA. Location ofsections (1-53) and outcrops of marine Mesozoic jediments and lurassic volcanics in northern Chile. For key ofand infor- mation on sections see Aberhan (1994a, p. 8). OB. Lower Jurassic palae- ogeography of the Andean Basin. Black triangles: volcanics; stippled: sedimentary basin fill. (Modified after Riccardi 1983.)
The data set has been analyzed in two ways: First, all data from the Andean Basin have been compiled for each time slice. For this purpose, both confirmed and assumed occurrences of taxa have been used. The latter group of taxa exhibit a discontinuous range, but are interpreted to have been present in the basin continuously from their first to their last appearance. In a second step, the data were analyzed separately for two sets of selected sections: those representing largely well-aerated, shallow- to mid- shelf environments throughout the Lower Jurassic and those in which initially shallow-water settings gave way to deep outer, commonly poorly oxygenated shelf environ-
184 Martin A berhan and Franz Theodor Fursich
Het
LETHAIA 29 (1996)
E ' L E ' L E ' M ' L Sin Plb Toa Aal
ments around the Pliensbachian-Toarcian boundary to be followed again by shallow-shelf conditions. In these cases, only confirmed occurrences were used.
The data base, consisting of the stratigraphic distribu- tion of Hettangian-Aalenian bivalves (a) from the Andean Basin as a whole (171 taxa), (b) from selected sec- tions characterized by poorly aerated deep-shelf facies during Late Pliensbachian to Early Toarcian times (90 taxa), and (c) from selected sections characterized by rel- atively shallow-water and well-aerated facies (86 taxa), has been deposited in the library of the Institut f i r Pala- ontologie der Universitat Wurzburg, Pleicherwall 1, 97070 Wiirzburg, Germany.
As a rule, diversity data were plotted for the total bivalve fauna. In a further step, the data for infaunal and epifaunal suspension-feeders have been presented sepa- rately, in order to evaluate whether diversity changes affect different life-habit groups selectively.
Results
Diversity trends
Diversity trends here represent trends in species richness expressed by the number of species. As the data base per investigated time slice is very large, the number of species is thought to be an accurate reflection of this aspect of diversity.
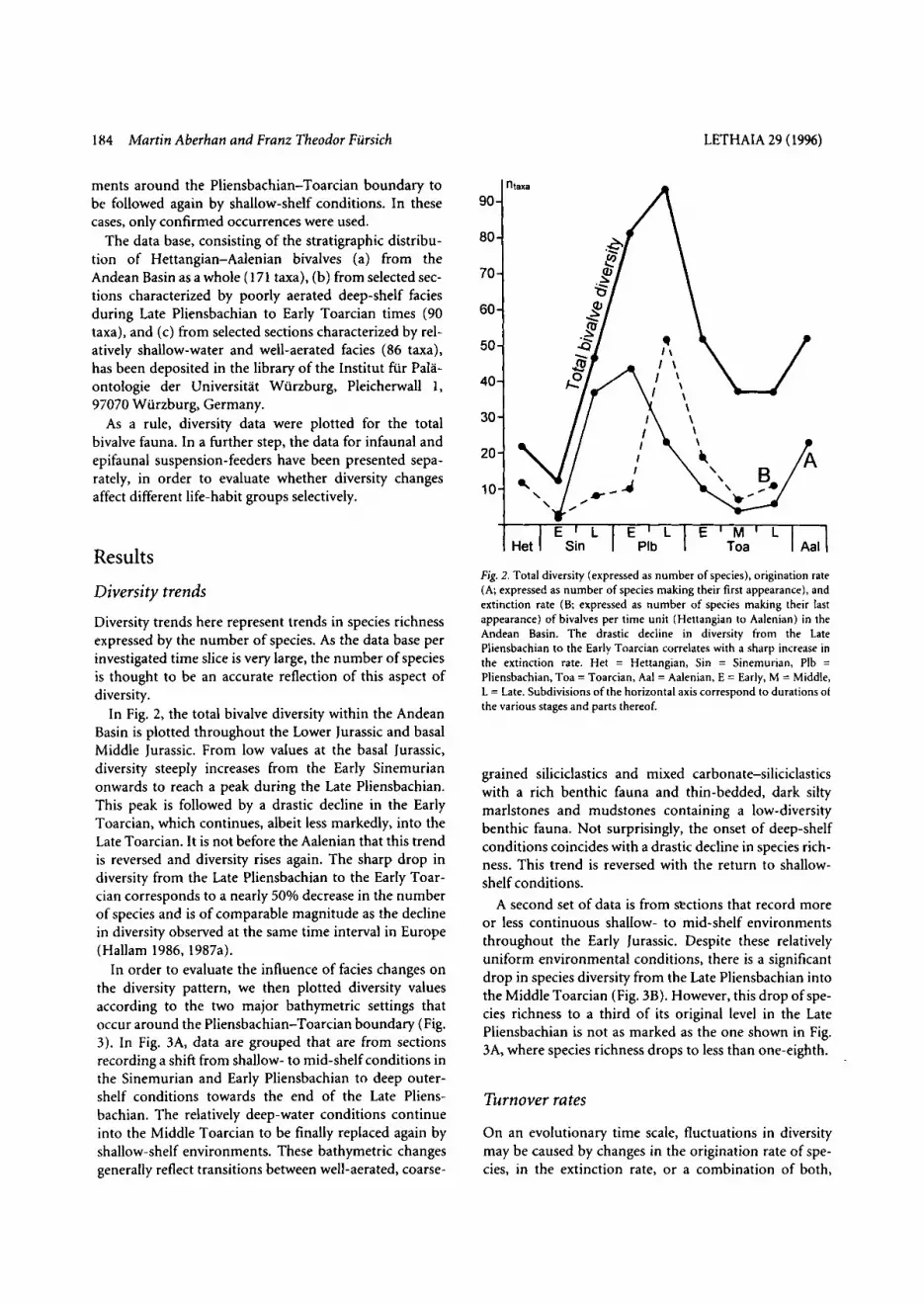
In Fig. 2, the total bivalve diversity within the Andean Basin is plotted throughout the Lower Jurassic and basal Middle Jurassic. From low values at the basal Jurassic, diversity steeply increases from the Early Sinemurian onwards to reach a peak during the Late Pliensbachian. This peak is followed by a drastic decline in the Early Toarcian, which continues, albeit less markedly, into the Late Toarcian. It is not before the Aalenian that this trend is reversed and diversity rises again. The sharp drop in diversity from the Late Pliensbachian to the Early Toar- cian corresponds to a nearly 50% decrease in the number of species and is of comparable magnitude as the decline in diversity observed at the same time interval in Europe (Hallam 1986, 1987a).
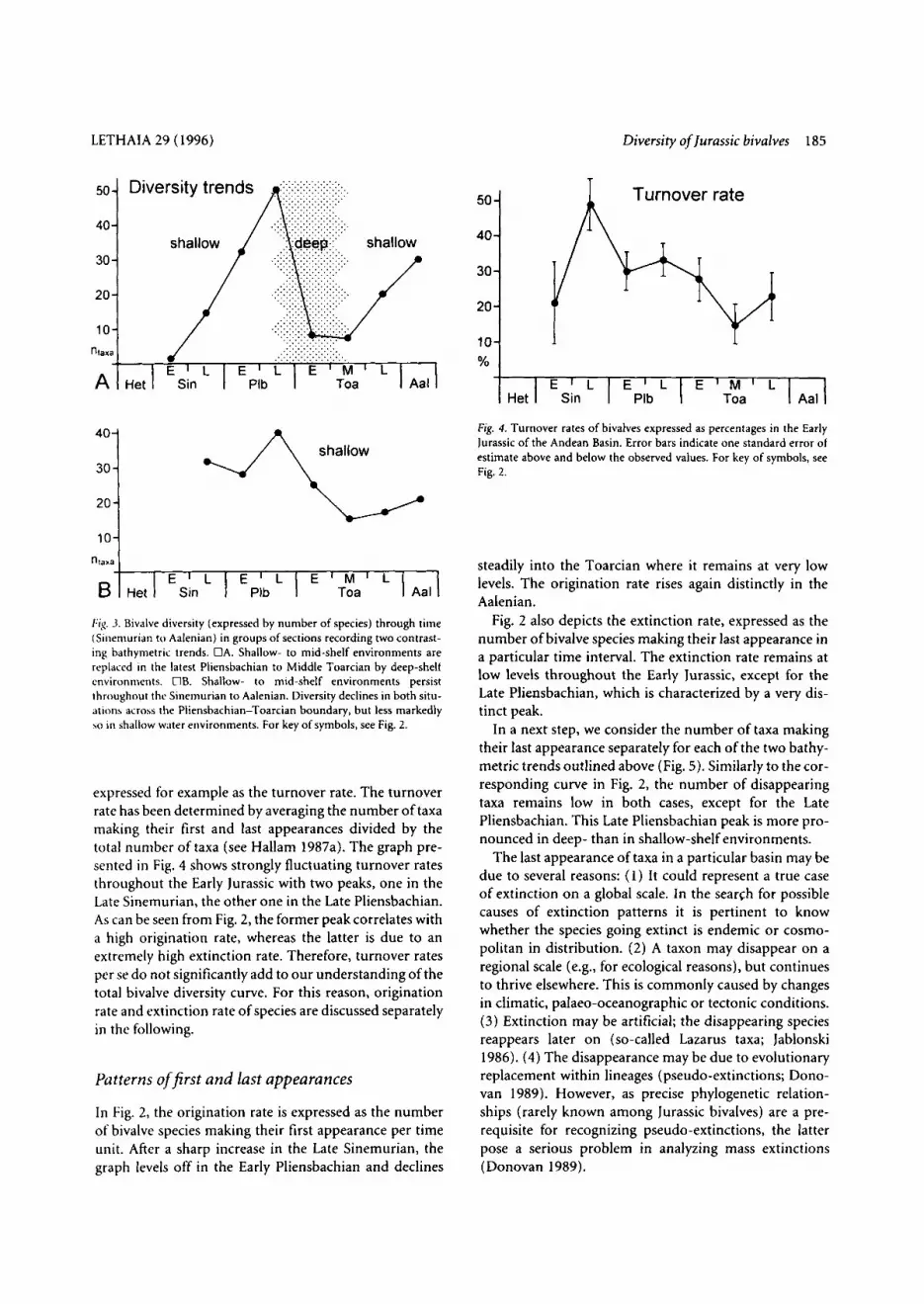
In order to evaluate the influence of facies changes on the diversity pattern, we then plotted diversity values according to the two major bathymetric settings that occur around the Pliensbachian-Toarcian boundary (Fig. 3). In Fig. 3A, data are grouped that are from sections recording a shift from shallow- to mid-shelf conditions in the Sinemurian and Early Pliensbachian to deep outer- shelf conditions towards the end of the Late Pliens- bachian. The relatively deep-water conditions continue into the Middle Toarcian to be finally replaced again by shallow-shelf environments. These bathymetric changes generally reflect transitions between well-aerated, coarse-
90
80
70
60
50
40
30
20
10
Fig. 2. Total diversity (expressed as number of species), origination rate (A; expressed as number of species making their first appearance), and extinction rate ( B expressed as number of species making their last appearance) of bivalves per time unit (Hettangian to Aalenian) in the Andean Basin. The drastic decline in diversity from the Late Pliensbachian to the Early Toarcian correlates with a sharp increase in the extinction rate. Het = Hettangian, Sin = Sinemurian, Plb = Pliensbachian, Toa = Toarcian, Aal = Aalenian, E = Early, M = Middle, L = Late. Subdivisions of the horizontal axis correspond to durations of the various stages and parts thereof.
grained siliciclastics and mixed carbonate-siliciclastics with a rich benthic fauna and thin-bedded, dark silty marlstones and mudstones containing a low-diversity benthic fauna. Not surprisingly, the onset of deep-shelf conditions coincides with a drastic decline in species rich- ness. This trend is reversed with the return to shallow- shelf conditions.
A second set of data is from sections that record more or less continuous shallow- to mid-shelf environments throughout the Early Jurassic. Despite these relatively uniform environmental conditions, there is a significant drop in species diversity from the Late Pliensbachian into the Middle Toarcian (Fig. 3B). However, this drop of spe- cies richness to a third of its original level in the Late Pliensbachian is not as marked as the one shown in Fig. 3A, where species richness drops to less than one-eighth.
Turnover rates
On an evolutionary time scale, fluctuations in diversity may be caused by changes in the origination rate of spe- cies, in the extinction rate, or a combination of both,
LETHAIA 29 (1996)
Het
Diversity of Jurassic bivalves 185
E ’ L E ’ L E ’ M ’ L Sin Plb Toa Aal
“1 30
2oi shallow 4
Plb Toa Aa I
Fig. 3. Bivalve diversity (expressed by number of species) through time (Sineniurian t o Aalenian) in groups of sections recording two contrast- ing bathymetric trends. CIA. Shallow- to mid-shelf environments are replaced in the latest Pliensbachian to Middle Toarcian by deep-shelf environments, OB. Shallow- to mid-shelf environments persist throughout the Sinemurian to Aalenian. Diversity declines in both situ- ations across the Pliensbachian-Toarcian boundary, but less markedly so in shallow water environments. For key ofsymbols, see Fig. 2.
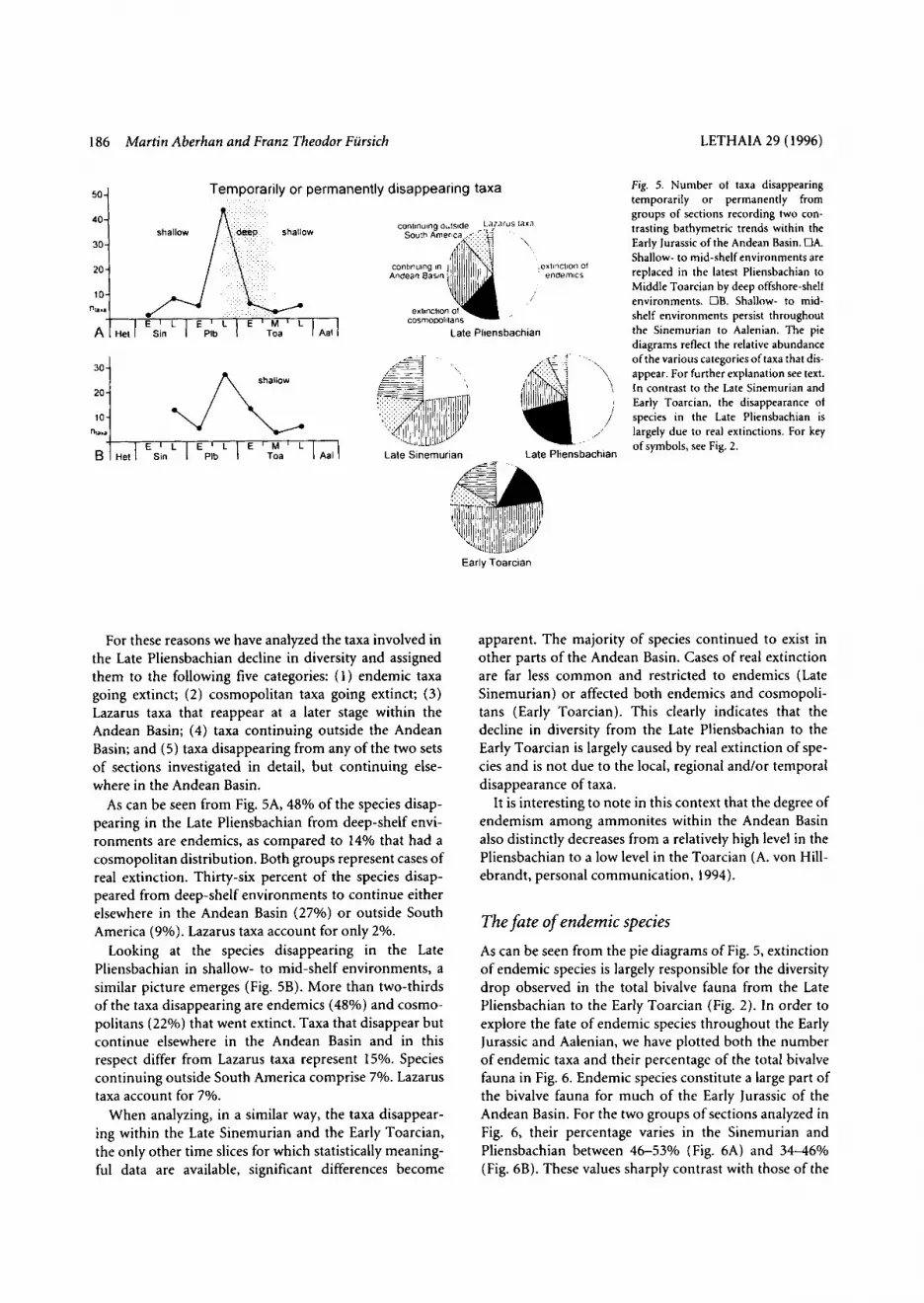
expressed for example as the turnover rate. The turnover rate has been determined by averaging the number of taxa making their first and last appearances divided by the total number of taxa (see Hallam 1987a). The graph pre- sented in Fig. 4 shows strongly fluctuating turnover rates throughout the Early Jurassic with two peaks, one in the Late Sinemurian, the other one in the Late Pliensbachian. As can be seen from Fig. 2, the former peak correlates with a high origination rate, whereas the latter is due to an extremely high extinction rate. Therefore, turnover rates per se d o not significantly add to our understanding of the total bivalve diversity curve. For this reason, origination rate and extinction rate of species are discussed separately in the following.
Patterns offirst and last appearances
In Fig. 2 , the origination rate is expressed as the number of bivalve species making their first appearance per time unit. After a sharp increase in the Late Sinemurian, the graph levels off in the Early Pliensbachian and declines
Turnover rate /h
Fig. 4. Turnover rates of bivalves expressed as percentages in the Early Jurassic of the Andean Basin. Error bars indicate one standard error of estimate above and below the observed values. For key of symbols, see Fig. 2.
steadily into the Toarcian where it remains at very low levels. The origination rate rises again distinctly in the Aalenian.
Fig. 2 also depicts the extinction rate, expressed as the number of bivalve species making their last appearance in a particular time interval. The extinction rate remains at low levels throughout the Early Jurassic, except for the Late Pliensbachian, which is characterized by a very dis- tinct peak.
In a next step, we consider the number of taxa making their last appearance separately for each of the two bathy- metric trends outlined above (Fig. 5). Similarly to the cor- responding curve in Fig. 2, the number of disappearing taxa remains low in both cases, except for the Late Pliensbachian. This Late Pliensbachian peak is more pro- nounced in deep- than in shallow-shelf environments.
The last appearance of taxa in a particular basin may be due to several reasons: ( 1 ) It could represent a true case of extinction on a global scale. In the search for possible causes of extinction patterns it is pertinent to know whether the species going extinct is endemic or cosmo- politan in distribution. (2) A taxon may disappear on a regional scale ( e g , for ecological reasons), but continues to thrive elsewhere. This is commonly caused by changes in climatic, palaeo-oceanographic or tectonic conditions. (3) Extinction may be artificial; the disappearing species reappears later on (so-called Lazarus taxa; Jablonski 1986). (4) The disappearance may be due to evolutionary replacement within lineages (pseudo-extinctions; Dono- van 1989). However, as precise phylogenetic relation- ships (rarely known among Jurassic bivalves) are a pre- requisite for recognizing pseudo-extinctions, the latter pose a serious problem in analyzing mass extinctions (Donovan 1989).
186 Martin Aberhan and Franz Theodor Fursich
50 - 40-
30 -
20 - 10-
nt...
A
LETHAlA 29 (1996)
Temporarily or permanently disappearing taxa
shallow shallow
Het Sin Plb Toa Aal
'\,
,exIIIICIIoIl 01 endemics
i
Late Pliensbachian
'\ ', shallow
/'
10
Plb Toa Aal Late Sinemurian bachian
For these reasons we have analyzed the taxa involved in the Late Pliensbachian decline in diversity and assigned them to the following five categories: (1 ) endemic taxa going extinct; (2) cosmopolitan taxa going extinct; (3) Lazarus taxa that reappear at a later stage within the Andean Basin; (4) taxa continuing outside the Andean Basin; and (5) taxa disappearing from any of the two sets of sections investigated in detail, but continuing else- where in the Andean Basin.
As can be seen from Fig. 5A, 48% of the species disap- pearing in the Late Pliensbachian from deep-shelf envi- ronments are endemics, as compared to 14% that had a cosmopolitan distribution. Both groups represent cases of real extinction. Thirty-six percent of the species disap- peared from deep-shelf environments to continue either elsewhere in the Andean Basin (27%) or outside South America (go/.). Lazarus taxa account for only 2%.
Looking at the species disappearing in the Late Pliensbachian in shallow- to mid-shelf environments, a similar picture emerges (Fig. 5B). More than two-thirds of the taxa disappearing are endemics (48%) and cosmo- politans (22%) that went extinct. Taxa that disappear but continue elsewhere in the Andean Basin and in this respect differ from Lazarus taxa represent 15%. Species continuing outside South America comprise 7%. Lazarus taxa account for 7%.
When analyzing, in a similar way, the taxa disappear- ing within the Late Sinemurian and the Early Toarcian, the only other time slices for which statistically meaning- ful data are available, significant differences become
Fig. 5. Number ot taxa disappearing temporarily or permanently from groups of sections recording two con- trasting bathymetric trends within the Early Jurassic of the Andean Basin. OA. Shallow- to mid-shelf environments are replaced in the latest Pliensbachian to Middle Toarcian by deep offshore-shelf environments. OB. Shallow- to mid- shelf environments persist throughout the Sinemurian to Aalenian. The pie diagrams reflect the relative abundance ofthe various categoriesoftaxa that dis- appear. For further explanation see text. In contrast to the Late Sinemurian and Early Toarcian, the disappearance of species in the Late Pliensbachian is largely due to real extinctions. For key of symbols, see Fig. 2.
Early Toarcian
apparent. The majority of species continued to exist in other parts of the Andean Basin. Cases of real extinction are far less common and restricted to endemics (Late Sinemurian) or affected both endemics and cosmopoli- tans (Early Toarcian). This clearly indicates that the decline in diversity from the Late Pliensbachian to the Early Toarcian is largely caused by real extinction of spe- cies and is not due to the local, regional and/or temporal disappearance of taxa.
It is interesting to note in this context that the degree of endemism among ammonites within the Andean Basin also distinctly decreases from a relatively high level in the Pliensbachian to a low level in the Toarcian (A. von Hill- ebrandt, personal communication, 1994).
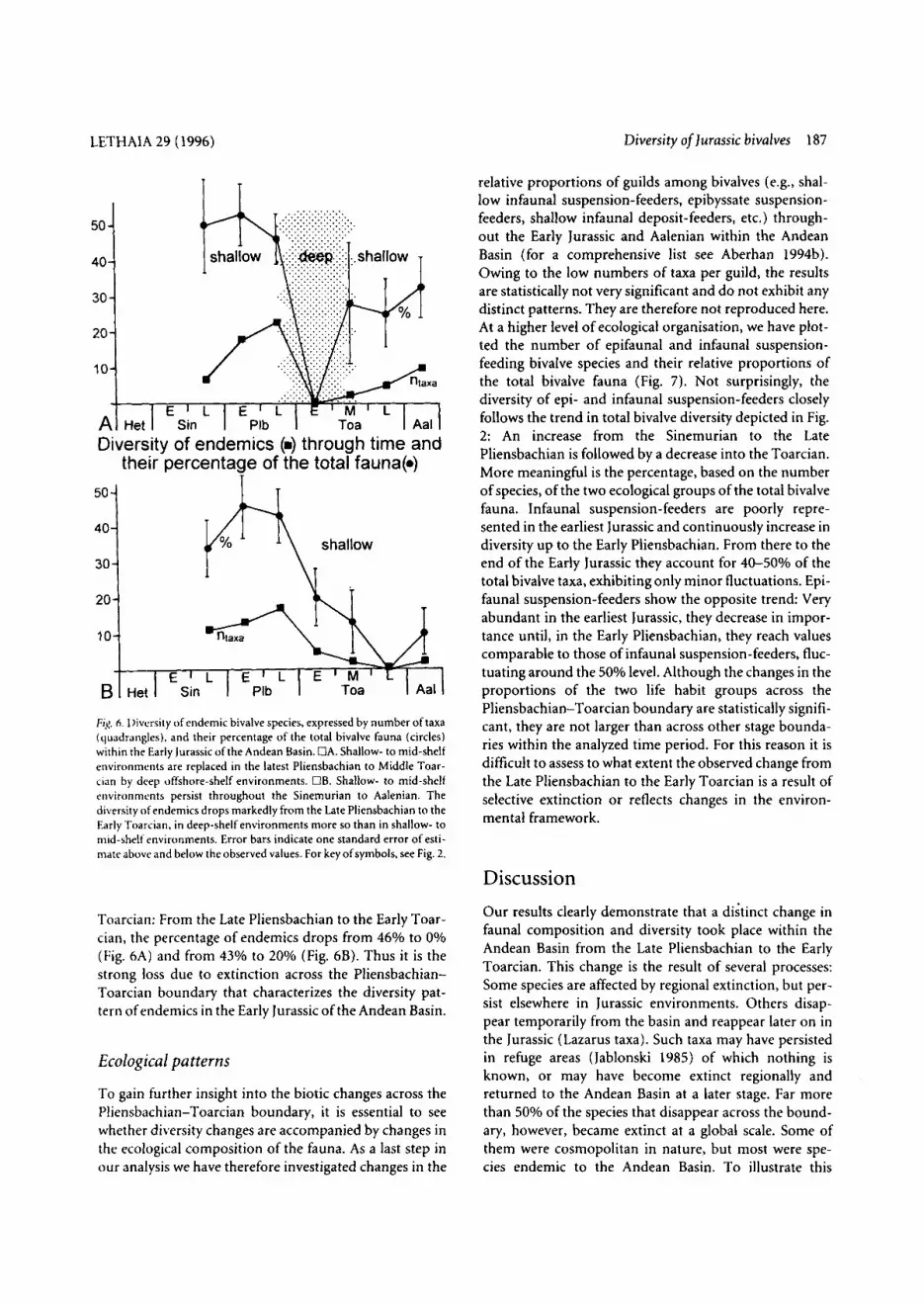
The fate of endemic species As can be seen from the pie diagrams of Fig. 5, extinction of endemic species is largely responsible for the diversity drop observed in the total bivalve fauna from the Late Pliensbachian to the Early Toarcian (Fig. 2). In order to explore the fate of endemic species throughout the Early Jurassic and Aalenian, we have plotted both the number of endemic taxa and their percentage of the total bivalve fauna in Fig. 6. Endemic species constitute a large part of the bivalve fauna for much of the Early Jurassic of the Andean Basin. For the two groups of sections analyzed in Fig. 6, their percentage varies in the Sinemurian and Pliensbachian between 4 6 5 3 % (Fig. 6A) and 3446% (Fig. 6B). These values sharply contrast with those of the
Diversity of endemics (m) through time and their percentage of the total fauna(.)
I T
20 -
FIaq. 6.I)iversity of endemic bivalve species, expressed by number oftaxa (quadrangles), and their percentage of the total bivalve fauna (circles) within the Early Jurassic of the Andean Basin. OA. Shallow- to mid-shelf environments are replaced in the latest Pliensbachian to Middle Toar- ciiin by deep offshore-shelf environments. DB. Shallow- to mid-shelf environments persist throughout the Sinemurian to Aalenian. The diversity of endemics drops markedly from the Late Pliensbachian to the Early Toarcian, in deep-shelf environments more so than in shallow- to mid-shelf environments. Error bars indicate one standard error of esti- mate above and below the observed values. For key of symbols, see Fig. 2.
Toarcian: From the Late Pliensbachian to the Early Toar- cian, the percentage of endemics drops from 46% to 0% (Fig. 6A) and from 43% to 20% (Fig. 6B). Thus it is the strong loss due to extinction across the Pliensbachian- Toarcian boundary that characterizes the diversity pat- tern of endemics in the Early Jurassic of the Andean Basin.
Ecological patterns
To gain further insight into the biotic changes across the Pliensbachian-Toarcian boundary, it is essential to see whether diversity changes are accompanied by changes in the ecological composition of the fauna. As a last step in our analysis we have therefore investigated changes in the
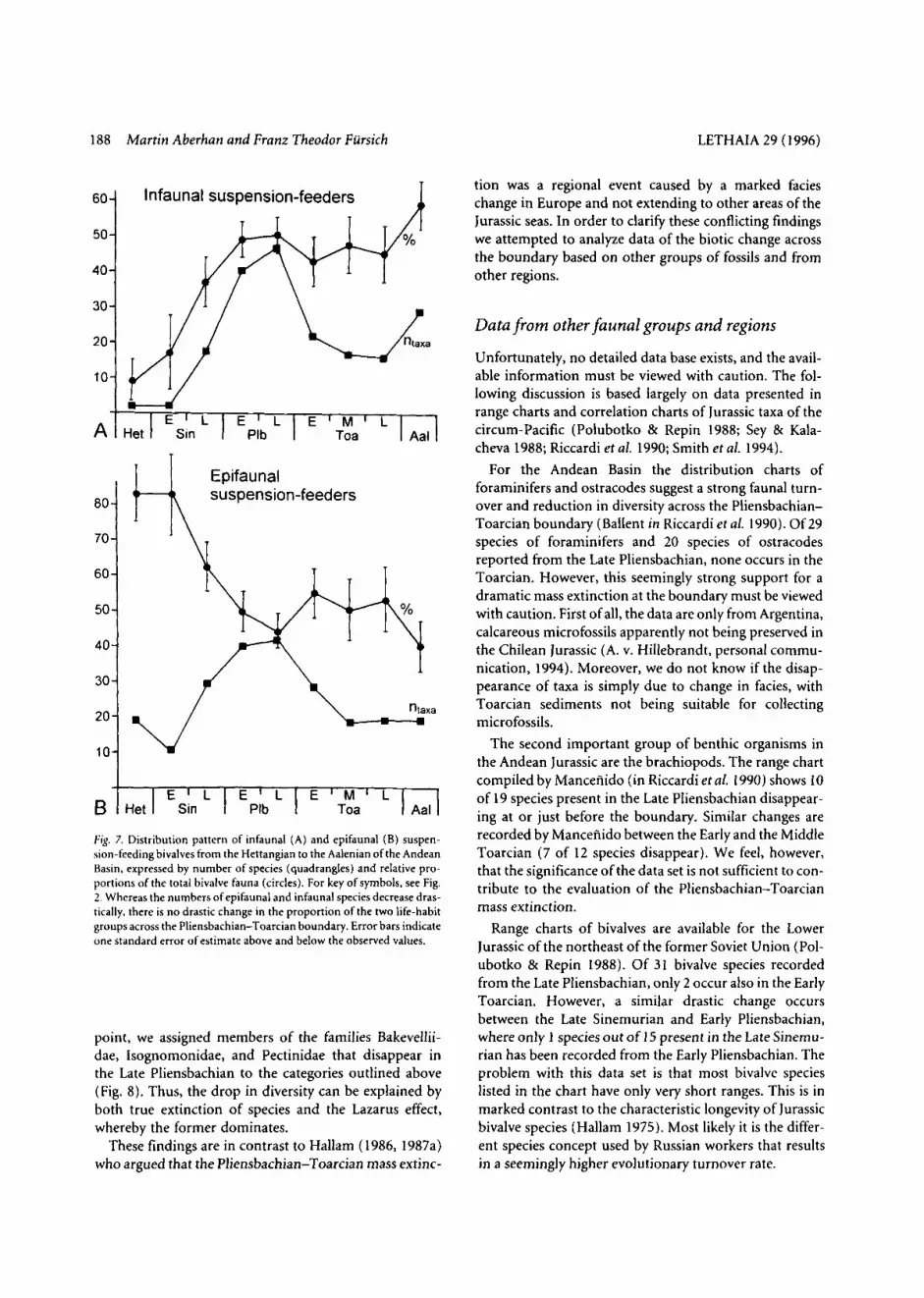
relative proportions of guilds among bivalves ( e g , shal- low infaunal suspension-feeders, epibyssate suspension- feeders, shallow infaunal deposit-feeders, etc.) through- out the Early Jurassic and Aalenian within the Andean Basin (for a comprehensive list see Aberhan 1994b). Owing to the low numbers of taxa per guild, the results are statistically not very significant and do not exhibit any distinct patterns. They are therefore not reproduced here. At a higher level of ecological organisation, we have plot- ted the number of epifaunal and infaunal suspension- feeding bivalve species and their relative proportions of the total bivalve fauna (Fig. 7). Not surprisingly, the diversity of epi- and infaunal suspension-feeders closely follows the trend in total bivalve diversity depicted in Fig. 2: An increase from the Sinemurian to the Late Pliensbachian is followed by a decrease into the Toarcian. More meaningful is the percentage, based on the number of species, of the two ecological groups of the total bivalve fauna. Infaunal suspension-feeders are poorly repre- sented in the earliest Jurassic and continuously increase in diversity up to the Early Pliensbachian. From there to the end of the Early Jurassic they account for 40-50% of the total bivalve taxa, exhibiting only minor fluctuations. Epi- faunal suspension-feeders show the opposite trend: Very abundant in the earliest Jurassic, they decrease in impor- tance until, in the Early Pliensbachian, they reach values comparable to those of infaunal suspension-feeders, fluc- tuating around the 50% level. Although the changes in the proportions of the two life habit groups across the Pliensbachian-Toarcian boundary are statistically signifi- cant, they are not larger than across other stage bounda- ries within the analyzed time period. For this reason it is difficult to assess to what extent the observed change from the Late Pliensbachian to the Early Toarcian is a result of selective extinction or reflects changes in the environ- mental framework.
Discussion Our results clearly demonstrate that a distinct change in faunal composition and diversity took place within the Andean Basin from the Late Pliensbachian to the Early Toarcian. This change is the result of several processes: Some species are affected by regional extinction, but per- sist elsewhere in Jurassic environments. Others disap- pear temporarily from the basin and reappear later on in the Jurassic (Lazarus taxa). Such taxa may have persisted in refuge areas (Jablonski 1985) of which nothing is known, or may have become extinct regionally and returned to the Andean Basin at a later stage. Far more than 50% of the species that disappear across the bound- ary, however, became extinct at a global scale. Some of them were cosmopolitan in nature, but most were spe- cies endemic to the Andean Basin. To illustrate this
188 Martin Aberhan and Franz Theodor Fursich LETHAIA 29 (1996)
1 604 lnfaunal suspension-feeders
50 -
40-
30 - 20 -
10-
E ' L E ' L E ' M ' L A Het I Sin Plb Toa Aal
T
Epifaunal suspension-feeders 80
60 -
50 - 40 -
30 -
20 -
Plb Toa Aa I
Fig. 7. Distribution pattern of infaunal (A) and epifaunal (B) suspen- sion-feeding bivalves from the Hettangian to the Aalenian of the Andean Basin. expressed by number of species (quadrangles) and relative pro- portions of the total bivalve fauna (circles). For key of symbols, see Fig. 2 . Whereas the numbers of epifaunal and infaunal species decrease dras- tically, there is no drastic change in the proportion of the two life-habit groups across the Pliensbachian-Toarcian boundary. Error bars indicate one standard error of estimate above and below the observed values.
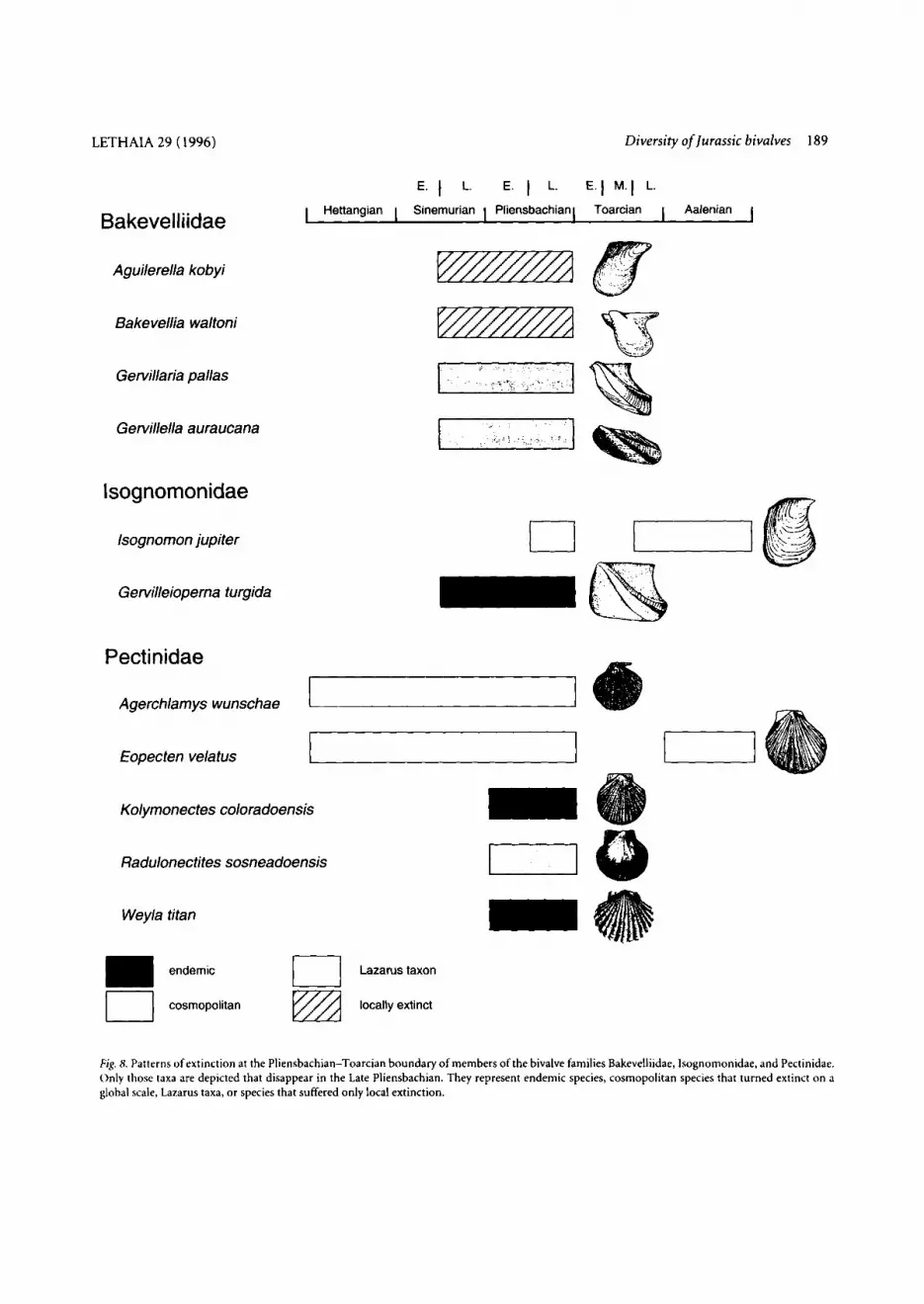
point, we assigned members of the families Bakevellii- dae, Isognomonidae, and Pectinidae that disappear in the Late Pliensbachian to the categories outlined above (Fig. 8). Thus, the drop in diversity can be explained by both true extinction of species and the Lazarus effect, whereby the former dominates.
These findings are in contrast to Hallam (1986, 1987a) who argued that the Pliensbachian-Toarcian mass extinc-
tion was a regional event caused by a marked facies change in Europe and not extending to other areas of the Jurassic seas. In order to clarify these conflicting findings we attempted to analyze data of the biotic change across the boundary based on other groups of fossils and from other regions.
Data from other faunal groups and regions
Unfortunately, no detailed data base exists, and the avail- able information must be viewed with caution. The fol- lowing discussion is based largely on data presented in range charts and correlation charts of Jurassic taxa of the circum-Pacific (Polubotko & Repin 1988; Sey & Kala- cheva 1988; Riccardi et al. 1990; Smith et al. 1994).
For the Andean Basin the distribution charts of foraminifers and ostracodes suggest a strong faunal turn- over and reduction in diversity across the Pliensbachian- Toarcian boundary (Ballent in Riccardi et al. 1990). Of 29 species of foraminifers and 20 species of ostracodes reported from the Late Pliensbachian, none occurs in the Toarcian. However, this seemingly strong support for a dramatic mass extinction at the boundary must be viewed with caution. First of all, the data are only from Argentina, calcareous microfossils apparently not being preserved in the Chilean Jurassic (A. v. Hillebrandt, personal commu- nication, 1994). Moreover, we do not know if the disap- pearance of taxa is simply due to change in facies, with Toarcian sediments not being suitable for collecting microfossils.
The second important group of benthic organisms in the Andean Jurassic are the brachiopods. The range chart compiled by Mancenido (in Riccardi et al. 1990) shows 10 of 19 species present in the Late Pliensbachian disappear- ing at or just before the boundary. Similar changes are recorded by Mancefiido between the Early and the Middle Toarcian (7 of 12 species disappear). We feel, however, that the significance ofthe data set is not sufficient to con- tribute to the evaluation of the Pliensbachian-Toarcian mass extinction.
Range charts of bivalves are available for the Lower Jurassic of the northeast of the former Soviet Union (Pol- ubotko & Repin 1988). Of 31 bivalve species recorded from the Late Pliensbachian, only 2 occur also in the Early Toarcian. However, a similar drastic change occurs between the Late Sinemurian and Early Pliensbachian, where only 1 species out of 15 present in the Late Sinernu- rian has been recorded from the Early Pliensbachian. The problem with this data set is that most bivalve species listed in the chart have only very short ranges. This is in marked contrast to the characteristic longevity of Jurassic bivalve species (Hallam 1975). Most likely it is the differ- ent species concept used by Russian workers that results in a seemingly higher evolutionary turnover rate.
LETHAIA 29 (1996) Diversity of Iurassic bivalves 189
E . ) L. E . 1 L. E . ) M . I L .
I Hettangian 1 Sinemurian I Pliensbachianl Toarcian I Aalenian 1 Ba kevel I i idae
Aguilerella kobyi
Bake vellia walfoni
Gervillaria pallas
Gervillella auraucana
lsognomonidae
lsognomon jupiter
Gervilleioperna turgida
Pecti nidae
Agerchlamys wunschae
Eopecfen velafus
Kolymonecfes coloradoensis
Radulonecfifes sosneadoensis I Weyla titan
Lazarus taxon
locally extinct
endemic
cosmopolitan
Fig. 8. Patterns of extinction at the Pliensbachian-Toarcian boundary of members of the bivalve families Bakevelliidae, Isognomonidae. and Pectinidae. Only those taxa are depicted that disappear in the Late Pliensbachian. They represent endemic species, cosmopolitan species that turned extinct on a global scale, Lazarus taxa, or species that suffered only local extinction.
190 Martin Aberhan and Franz Theodor Fiirsich LETHAIA 29 ( 1996)
The data set available for the Lower Jurassic of the Pacific margin of North America (Smith et al. 1994) appears to be relatively comprehensive, but most of the ranges given are uncertain and in many cases need to be revised (Aberhan, unpublished).
In summary, the available information about faunal changes across the Pliensbachian-Toarcian boundary from regions other than Europe and South America is, at present, not sufficient to add to the global picture of the events at the boundary. Nevertheless, the simultaneous decline in diversity in two regions separated as widely as Europe and the Andean Basin suggests a global causal fac- tor. In its extent, this decline greatly exceeds background fluctuations in diversity. This, its global nature, and its occurrence in several groups of organisms (Hallam 1986, 1987a) justify the term mass extinction.
The role of anoxia in the Pliensbachian-Toarcian mass extinction Hallam (1986, 1987a, 1989) explained the mass extinction at the Pliensbachian-Toarcian boundary in Europe by the spread of anoxic bottom waters in connection with a glo- bal sea-level rise in the Early Toarcian. In the same papers he discounted the presence of black shales and conse- quently anoxic conditions in the Lower Jurassic of the Andean Basin and failed to recognize the bivalve extinc- tion in this region. Jenkyns (1988), in contrast, assumed the anoxic event and the concomitant extinctions to be worldwide.
In many sections of the Andean Lower Jurassic, the lat- est Pliensbachian and Toarcian is represented by thin- bedded, dark silty marlstones and mudstones. Compared with earlier sediments, these mud- and marlstones con- tain only a low-diversity bivalve and brachiopod fauna dominated either by epifaunal opportunists, in particular so-called flat clams, or by infaunal deposit-feeders, mainly nuculids. This benthic fauna represents the relics of four former communities, the Propeamussium pumilum, the Posidonotis cancellata [ = P. semiplicata], the Nuculana cf. ovum, and the Gryphaea sp. A association (Aberhan 1992, 1993). These associations have been interpreted as oxygen-controlled to a variable degree, with the redox boundary characteristically staying below the sediment-water interface except in the case of the Posidonotis cancellata association. There, the position of the redox boundary is assumed to have been at or imme- diately above the interface, excluding virtually all benthic organisms except for the extreme opportunist Posidono- tis. This biofacies persisted for considerable periods of time and represents the most severe reduction in oxygen (Aberhan 1992, 1993).
Although no classical black shales are known from the Early Jurassic of the Andean Basin both sediments and
benthic fauna strongly suggest oxygen-depleted deeper- shelf environments at the Pliensbachian-Toarcian boundary. Therefore we interpret the marked drop in diversity shown in Figs. 2 and 3A as due to poorly oxygen- ated environments existing for an extended period of time and restricting the benthic fauna to opportunists and to specialists able to cope with the adverse conditions. How- ever, as can be seen from Figs. 3B and 5B, a similar, although less pronounced, drop in diversity occurs in shallow- to mid-shelf environments well-aerated for most of the time elsewhere in the basin during the same time interval. Solely within the uppermost Pliensbachian, intervals of dark silty mudstones occur in several of the sections representing shallow to mid-shelf environments, which, judging from their faunal content, experienced some degree of oxygen deficiency. Thus, low oxygen sup- ply alone is not sufficient to explain the observed diversity pattern of bivalves within the Andean Basin. In analogy, spread of anoxia and the corresponding reduction in hab- itat area might not have been the sole factor for the decrease in bivalve diversity across the boundary in Europe. Late Pliensbachian and Toarcian shallow-water environments in Europe are little known, and their faunal content has been poorly documented. Hallam’s (1986, 1987a) approach to analyze diversity trends in the Early Jurassic suffers from failing to differentiate between largely well-aerated and poorly oxygenated environments at the boundary. We believe that in this way anoxia inev- itably would be identified as the main cause of the mass extinction and that its role was probably overestimated. In the following we propose an alternative model for the mass extinction at the Pliensbachian-Toarcian boundary, in which anoxia plays some role but in our view should be regarded as product of a global palaeo-oceanographic sit- uation that caused severe reduction in marine diversity.
A palaeo-oceanographic model for the Pliensbachian-Toarcian extinction event
Any model for the mass extinction at the Pliensbachian- Toarcian boundary must explain the following two obser- vations: (1 1 Extinction takes place not only in poorly aer- ated deep-shelf environments, but also in well-aerated shallow- to mid-shelf settings and (2) extinction affected largely endemic species.
In addition, both the Andean Basin and the epiconti- nental seas of Europe experienced a general eustatic rise in sea-level since the Hettangian, reaching a peak in the Early to Middle Toarcian (e.g., Hallam 1988). Moreover, oxygen-impoverished environments were widespread in both regions. Therefore, sea-level and anoxia should be integrated in any oceanographic model explaining the mass extinction. With Hallam (1989), we envisage changes in sea-level as the initiating causal factor for the
LETHAIA 29 ( 1996) Diversity of Jurassic bivalves 19 1
-
I \ , t I
Diversity increase1 [Diversity decrease1
Early transgression A B Peak transgression
Fig. Y. Model explaining increase in marine diversity during early trans- gression ( A ) and decrease during peak transgression (B) . For explana- tion. see text.
mass extinction at the Pliensbachian-Toarcian boundary, but we would like to stress the associated climatic changes. In Figs. 9 and 10 we have illustrated this relation- ship and the numerous interconnected pathways. The model in Fig. 9 has been inspired by the model recently proposed by Follmi et al. (1994) to explain the phospho- genesis in the Lower Cretaceous of the northern Tethyan margin and the model by Jeppsson (1990) and Aldridge et al. (1993) to explain oceanographic cycles in the Silurian. These models are modifications and variations of the glo- bal climatic and oceanographic models proposed by Fischer & Arthur (1977) and Fischer (1984).
Our model differentiates between early transgression, when marine species diversity actually increases (Fig. 9A), and peak transgression, during which diversity decreases (Fig. 9B). Extrinsic perturbations, such as increased activ- ity at mid-oceanic ridges leads to a eustatic sea-level rise. The resulting larger shelf areas provide an increased hab- itat heterogeneity. At the same time, larger shelf areas are thought to reduce the degree of environmental stress, with salinity and temperature fluctuation becoming rela- tively less important. The eustatic sea-level rise leads to intensified greenhouse conditions. In this context, the more equable climate with a decreased seasonality enhances overall environmental stability, decreases envi- ronmental stress, and finally promotes K-selection (e.g., Johnson 1974; Hallam 1978, 1989). Both an increase in habitat heterogeneity and K-selection increase marine species diversity during early transgression (Fig. 9A).
The intensified greenhouse conditions also result in intensified continental weathering, which in turn increases the nutrient supply to the oceans. In the same way, increased humidity promotes vegetation of conti-
nental areas. With the accelerated water cycle characteris- tic of greenhouse conditions much of this biomass is transported to the sea, and at least part of it adds to the nutrient supply of marine organisms. As a result, produc- tivity of the epicontinental seas increases. In combination with a reduced thermohaline circulation in the ocean due to more equable climates, high primary productivity pro- duces widespread anoxia or at least oxygen-deficiency at the sediment-water interface. This may culminate in the formation of black shales (e.g., Arthur & Sageman 1994; Wignall 1994). Widespread anoxia decreases habitat het- erogeneity and increases the degree of environmental stress, leading to the extinction of K-selected specialists and favouring broad-niched r-selected generalists.
With continued transgression, formerly isolated shelf areas become connected, facilitating faunal exchange. Within a particular basin, the relative importance of endemic taxa is diminished by immigration of cosmopol- itan species. In addition to preemptive competition (Hal- lam 1990) caused by anoxia, competitive replacement of endemic species by cosmopolitans (displacive competi- tion of Hallam 1990) might have decreased their relative proportion of the fauna. On a global scale, the spread of cosmopolitan species at the expense of endemic species reduces the species diversity. Thus extinction caused by decreased habitat diversity and increase in environmental stress, probably enhanced by extinction of endemics due to competitive replacement, lead to a global decrease in diversity.
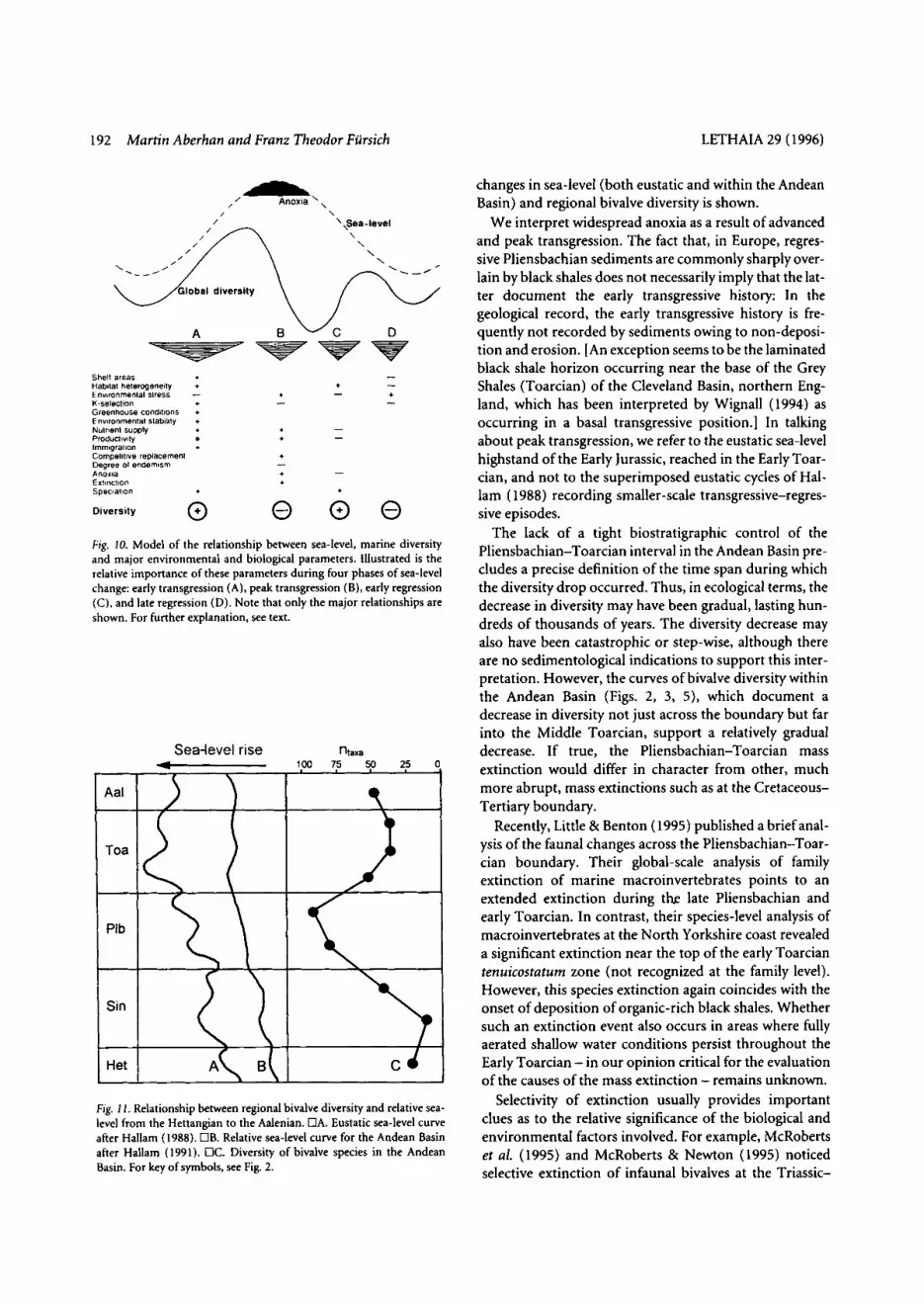
The increase in diversity during early transgression fol- lowed by a marked decrease during peak transgression is also shown in Fig. 10. In addition, the diversity trends during ensuing regression and their relationship to some important biological and environmental parameters are depicted. During fall of eustatic sea-level, the significance of many of the interconnected parameters of Fig. 9 changes. In particular, nutrient supply and therefore pro- ductivity decrease and ocean circulation improves. Con- sequently, anoxia disappears, environmental conditions ameliorate, and habitat heterogeneity increases again. This leads to an increase in species diversit);, which is aug- mented by increasing speciation due to progressive geo- graphic isolation. With advancing fall in sea-level, how- ever, diversity trends are again reversed: Severe habitat restriction and increasing environmental stress lead to extinction of K-selected taxa and reduce species diversity. The importance of this second drop in diversity, coincid- ing with sea-level lowstand, depends on the magnitude of sea-level fall.
In our opinion, the model fits the general environmen- tal history of Early Jurassic shelf seas and in particular explains the increase in diversity during the Sinemurian and Pliensbachian and the following decline in diversity across the Pliensbachian-Toarcian boundary. This is illustrated in Fig. 11, where the relationship between
192 Martin Aberhan and Franz Theodor Fursich LETHAIA 29 (1996)
0 0 Fig. 10. Model of the relationship between sea-level, marine diversity and major environmental and biological parameters. IUustrated is the relative importance of these parameters during four phases of sea-level change: early transgression (A), peak transgression (B), early regression (C). and late regression (D). Note that only the major relationships are shown. For further explanation, see text.
Fig. 11 . Relationship between regional bivalve diversity and relative sea- level from the Hettangian to the Aalenian. OA. Eustatic sea-level curve after Hallam (1988). OB. Relative sea-level curve for the Andean Basin after Hallam (1991). OC. Diversity of bivalve species in the Andean Basin. For key of symbols, see Fig. 2.
changes in sea-level (both eustatic and within the Andean Basin) and regional bivalve diversity is shown.
We interpret widespread anoxia as a result of advanced and peak transgression. The fact that, in Europe, regres- sive Pliensbachian sediments are commonly sharply over- lain by black shales does not necessarily imply that the lat- ter document the early transgressive history: In the geological record, the early transgressive history is fre- quently not recorded by sediments owing to non-deposi- tion and erosion. [An exception seems to be the laminated black shale horizon occurring near the base of the Grey Shales (Toarcian) of the Cleveland Basin, northern Eng- land, which has been interpreted by Wignall (1994) as occurring in a basal transgressive position.] In talking about peak transgression, we refer to the eustatic sea-level highstand of the Early Jurassic, reached in the Early Toar- cian, and not to the superimposed eustatic cycles of Hal- lam ( 1988) recording smaller-scale transgressive-regres- sive episodes.
The lack of a tight biostratigraphic control of the Pliensbachian-Toarcian interval in the Andean Basin pre- cludes a precise definition of the time span during which the diversity drop occurred. Thus, in ecological terms, the decrease in diversity may have been gradual, lasting hun- dreds of thousands of years. The diversity decrease may also have been catastrophic or step-wise, although there are no sedimentological indications to support this inter- pretation. However, the curves of bivalve diversity within the Andean Basin (Figs. 2, 3, 5), which document a decrease in diversity not just across the boundary but far into the Middle Toarcian, support a relatively gradual decrease. If true, the Pliensbachian-Toarcian mass extinction would differ in character from other, much more abrupt, mass extinctions such as at the Cretaceous- Tertiary boundary.
Recently, Little & Benton (1995) published a brief anal- ysis of the faunal changes across the Pliensbachian-Toar- cian boundary. Their global-scale analysis of family extinction of marine macroinvertebrates points to an extended extinction during thrr late Pliensbachian and early Toarcian. In contrast, their species-level analysis of macroinvertebrates at the North Yorkshire coast revealed a significant extinction near the top of the early Toarcian tenuicostatum zone (not recognized at the family level). However, this species extinction again coincides with the onset of deposition of organic-rich black shales. Whether such an extinction event also occurs in areas where fully aerated shallow water conditions persist throughout the Early Toarcian - in our opinion critical for the evaluation of the causes of the mass extinction - remains unknown.
Selectivity of extinction usually provides important clues as to the relative significance of the biological and environmental factors involved. For example, McRoberts et al. (1995) and McRoberts & Newton (1995) noticed selective extinction of infaunal bivalves at the Triassic-
LETHAIA 29 (1996)
Jurassic transition, Infaunal bivalves apparently are less efficient filter feeders relative to their metabolic demands than epifaunal filter feeders. A likely cause for the end- Triassic mass extinction was therefore a strong reduction in primary productivity, which affected infaunal organ- isms more strongly than epifaunal ones. In the Early Jurassic of the Andean Basin there is a statistically signifi- cant change in the relative proportion of infaunal and epi- faunal suspension-feeding bivalves across the Pliens- bachian-Toarcian boundary (Fig. 7). As similar or even larger changes took place across other stage and substage boundaries, it is difficult to evaluate to what extent lim- ited supply of food may have contributed to the decrease in diversity.
In our model, sea-level fluctuations play an important role in controlling diversity patterns. It is, however, obvi- ous that our model is not applicable to smaller-scale regressive-transgressive couplets of the Jurassic sea-level curve of Hallam (1988). Obviously, the order of magni- tude is critical. In the Early Toarcian, the eustatic sea-level reaches its peak within the Early Jurassic. Similar peaks are reached in the Early Callovian, Middle Oxfordian, and Early Tithonian (Hallam 1988) without being associated with black shales and distinct declines in diversity. Appar- ently, threshold effects play a decisive role in triggering the chain of environmental and biological changes (Fig. 9), which finally lead to mass extinction. The fact that the other sea-level highstands mentioned above are not asso- ciated with mass extinctions can be explained in several ways. For example, the opening of the so-called Hispanic Corridor, connecting the Palaeo-Pacific with the Western Tethys, in the Pliensbachian (e.g., Damborenea & Man- cenido 1979; Smith &Tipper 1986) enabled, at least inter- mittently, free migration of marine organisms. Conse- quently it most likely contributed to the extinction of endemic species in the Andean Basin. Another important point is that regional tectonic events may have overridden eustatic sea-level rise and therefore counteracted the oce- anographic and climatic effects of the latter. Thus, the Jurassic relative sea-level curve of the Andean Basin dras- tically deviates for several time slices from the eustatic sea- level curve (Hallam 1991). Finally the effects of eustatic sea-level rise also depend on continental hypsometry and thus on the relative position of sea-level prior to the eustatic rise (Wyatt 1987).
In the lurassic, there is a second, apparently regional, extinction event at the end of the Tithonian (Hallam 1986). Associated with a distinct regression in Europe, this event could be explained by the diversity curve shown in Fig. 10, where loss of habitat heterogeneity and increas- ing environmental stress lead to a decrease in species diversity (see also Hallam 1986).
Diversity of Jurassic bivalves 193
Conclusions In contrast to earlier opinions, diversity of marine bivalve species drastically decreases within the Andean Basin across the Pliensbachian-Toarcian boundary. A similar decrease in diversity had been observed earlier on in epicontinental seas of Western Europe and was regarded as a regional phenomenon (e.g., Hallam 1986, 1987a). The decrease in bivalve diversity within the Andean Basin is due to the extinction of cosmopolitan and, above all, endemic species, more rarely to local extinc- tions and the Lazarus effect. The diversity patterns observed in Europe and the Andean Basin suggest a global cause and justify the term mass extinction. Time of extinction within the Andean Basin correlates, as in Europe, with sea-level highstand and oxygen- deficient deep-shelf environments. Spread of anoxia cannot, however, be the sole reason for the drop in diversity, as shallow-shelf areas are also affected, albeit less severely. The Pliensbachian-Toarcian mass extinction is explained as the result of competitive replacement of endemic taxa by cosmopolitan taxa and the loss of habitat heterogeneity in connection with spread of oxygen-poor bottom waters. Most likely it was not a catastrophic event but may have taken hundreds of thousands of years. A palaeo-oceanographic model is proposed, which relates diversity changes to eustatic sea-level changes. The lack of mass extinctions at similar sea-level high- stands in the Jurassic suggests that threshold effects are important for triggering mass extinctions. Although the global character of the Pliensbachian- Toarcian mass extinction appears to strengthen the periodicity hypothesis, there is no indication of an extraterrestrial cause. The decrease in marine species diversity can be explained by an oceanographic sce- nario in which sea-level changes are regarded as the ultimate cause.
Acknowledgments. -We would like to thank A. von Hillebrandt, Berlin, for making his bivalve collections from Chile available to us and C. McRoberts, Wiirzburg, for commenting on the manuscript and for use- ful suggestions. We are grateful to A. Hallam and I. Kfii. for editorial reviews of the manuscript.
References Aberhan, M. 1992: Palokologie und zeitliche Verbreitung benthischer
Faunengemeinschaften im Unterjura von Chile. Beringeria 5, 1-174. Aberhan, M. 1993: Benthic macroinvertebrate associations on a carbon-
ate-clastic ramp in segments of the Early Jurassic back-arc basin of northern Chile (26-29”s). Revisfa Geoldgica de Chile 20, 105-136.
Aberhan, M. 1994a: Early Jurassic Bivalvia of northern Chile. Part 1. Subclasses Palaeotaxodonta, Pteriomorphia, and Isofilibranchia. Ber- ingeria 13.3-1 15.
194 Martin Aberhan and Franz Theodor Fursich LETHAIA 29 (1996)
Aberhan, M. 1994b Guild-structure and evolution of Mesozoic benthic shelfcommunities. Palaios 9, 516-545.
Aldridge, R.J., Jeppsson, L. & Dorning, K.J. 1993: Early Silurian oceanic episodes and events. Journal of the ~eo /og ica / Society London 150, 501 -51 3.
Arthur, M.A. & Sageman, B.B. 1994: Marine black shales: depositional mechanisms and environments of ancient deposits. Annual Reviews ofEarth and Planetary Sciences 22,499-551.
Chong, G.D. & Hillebrandt, A. von 1985: El Triasico preandino de Chile entre 10s 23'30' y 2600' de lat. sur. IV Congreso geolbgico Chileno. Tomo I , 162-210. Universidad del Norte, Antofagasta.
Dalziel, I.W.D. 1986: Collision and Cordilleran orogenesis: an Andean perspective. In Coward, M.P. & Ries, A.C. (eds.): Collision tectonics. Geological Society Special Publication 19, 389-404.
Damborenea, S.E. 1987a: Early Jurassic Bivalvia of Argentina. Part 1: Stratigraphical introduction and superfamilies Nuculanacea, Arca- cea, Mytilacea and Pinnacea. Palaeonrographica A I99,23-111.
Damborenea, S.E. 1987b: Early Jurassic Bivalvia ofArgentina. Part 2: Su- perfamilies Pteriacea, Buchiacea and part of Pectinacea. Palaeonro- graphica A 199, 113-216.
Damborenea, S.E. 1990a: Bivalves. In Riccardi, A.C., Damborenea, S.E. & Manceiiido, M.O.: Lower Jurassic of South America and Antarctic Peninsula. Newletters on Stratigraphy 21,8590.
Damborenea, S.E. 1990b Bivalves. In Riccardi, A.C., Westermann, G.E.G., & Damborenea, S.E.: Middle Jurassic of South America and Antarctic Peninsula. Newletters on Stratigraphy 21, 114, 1 1 6 - 1 19.
Damborenea, S.E. 1992: South American Jurassic bivalves. In Wester- mann, G.E.G. (ed.): The Jurassic ofthe circum-Pacific, Atlas, PIS. 115- 1 19. Cambridge University Press, Cambridge.
Damborenea, S.E. 1993: Early Jurassic South American pectinaceans and circum-Pacific palaeobiogeography. Palaeogeography, Palaeocli- matology, Palaeoecology 100, 109-123.
Damborenea, S.E. & Manceiiido, M.O. 1979: On the palaeogeographical distribution of the pectinid genus Weyla (Bivalvia, Lower Jurassic). Palaeogeography, Palaeoclimatology, Palaeoecology 27,85- 102.
Damborenea, S.E. & Manceiiido, M.O. 1992: A comparison of Jurassic marine benthonic faunas from South America and New Zealand. Journal ofthe Royal Society ofNew Zealand 22, 131-152.
Dedi6s, P. 1967 Cuadrangulo Vicuiia, Provincia de Coquimbo. Instituro de lnvestigaciones Geolbgicas, Carta Geolbgica de Chile 16, 1-65.
Donovan, S.K. 1989: Palaeontological criteria for the recognition of mass extinction. In Donovan, S.K. (ed.) Mass extinctions. Processes and Evidence, 19-36. Belhaven, London.
Fischer, A.G. 1984: The two Phanerozoic supercycles. In Berggren, W.A. & Van Couvering, J.A. (eds.): Carasrrophies and Earth History, 129- 150. Princeton University Press, Princeton, N.J.
Fischer, A.G. & Arthur, M.A. 1977: Secular variations in the pelagic realm. In Cook, H.E. & Enos, P. (eds.): Deep-Water Carbonate Envi- ronments. Society of Economic Paleontologists and Mineralogists, Spe- cial Publication 25, 19-50.
Fiillmi, K.B., Weissert, H., Bisping, M. & Funk, H. 1994: Phosphogene- sis, carbon-isotope stratigraphy, and carbonate-platform evolution along the Lower Cretaceous northern Tethyan margin. Geological So- ciety ofAmerica Bulletin 106,729-746.
Gradstein, F.M., Agterberg, F.P., Ogg, J.G., Hardenbol, J., van Veen, P. Thierry. I . & Huang Zehui 1994: A Mesozoic time scale. Journal of Geophysical Research 99, No. B12,24051-24074.
Groschke, M., Hillebrandt, A. von, Prinz, P., Quinzio. L.A. & Wilke, H.- C. 1988: Marine Mesozoic paleogeography in northern Chile between 2lo-26"S. In Bahlburg, H., Breitkreuz, C. & Giese, P. (eds.): The southern central Andes. Lecture Notes in Earth Sciences 17, 105-1 17.
Hallam, A. 1975: Evolutionary size increase and longevity in Jurassic bi- valves and ammonites. Nature 258,493496.
Hallam. A. 1978: How rare is phyletic gradualism? Evidence from Jurassic bivalves. Paleobiology 4 , 16-25.
Hallam, A. 1983: Early and mid-Jurassic molluscan biogeography and the establishment of the central Atlantic seaway. Palaeogeography, Palaeoclimatology, Palaeoecology 43, 18 1-193.
Hallam, A. 1986 The Pliensbachian and Tithonian extinction events. Nature 31 9,765-768.
Hallam, A. 1987a: Radiations and extinctions in relation to environmen- tal change in the marine Lower Jurassic of northwest Europe. Palaeo- biology 13, 152-168.
Hallam, A. 1987b End-Cretaceous mass extinction event: argument for terrestrial causation. Science 238, 1237-1242.
Hallam, A. 1988 A reevaluation of Jurassic eustacy in the light of new data and the revised Exxon curve. In Wilgus, C.K., Hasting, B.S. Kendall, C.G.St.C., Posamentier, H.B. Ross, C.A. & Van Wagoner, J.C. (eds.): Sea-level changes - an integrated approach. Society of Ecomonic Paleontologists and Mitteralogists Special Publications 42, 261-273.
Hallam, A. 1989 The case for sea-level change as a dominant causal fac- tor in mass extinction of marine invertebrates. Philosophical Transac- tions ofthe Royal Society of London B 325,437-455.
Hallam, A. 1990: Biotic and abiotic factors in the evolution of Early Mes- ozoic marine molluscs. In Ross, R.M. & Allmon, W.D. (eds.): Causes ofEvoluhon. A Paleontological Perspective, 249-269, The University of Chicago Press, Chicago, Ill.
Hallam, A. 1991: Relative importance of regional tectonics and eustacy for the Mesozoic of the Andes. In Macdonald, D.I.M. (ed.): Sedimen- tation, Tectonics and Eustacy. International Association of Sedimentol- ogists Special Publications 12, 189-200.
Haq, B.U., Hardenbol, 1. & Vail, P.R. 1988: Mesozoic and Cenozoic chronostratigraphy and eustatic cycles. In Wilgus, C.K., Hasting, B.S. Kendall,C.G.St.C., Posamentier, H.B. Ross, C.A. &Van Wagon- er, J.C. (eds.): Sea-level changes - an integrated approach. Society of Ecomonic Paleontologisis and Mineralogists Special Publications 42.
Harland, W.B., Cox, A.V., Llewellyn, P.G., Pickton, C.A.G., Smith, A.G. & Walters, R. 1990: A Geologic Time Scale, 1989. 131 pp. Cambridge University Press, Cambridge.
Hillebrandt, A. von 197 I: Der Jura in der chilenisch-argentinischen Hochkordillere (25' bis 32'30's). Miittstersche Forschungen zur Geol- ogie und Palaontologie 20/21,63-87.
Hillebrandt, A. von 1973: Neue Ergebnisse iiber den Jura in Chile und Argentinien. Miinstenchhe Forschungen zur Geologie utid Paliiontologie
Hillebrandt, A. von 1977: Ammoniten aus dem Bajocien (Jura) von Chile (Sudamerika). Neue Arten der Gattungen Stephanoceras und Domeykoceras n. gen. (Stephanoceratidae). Mitteilungen der Bayer- ischen Staatssammlung fur Palaontologie und historische Ceologie 17, 3569.
Hillebrandt, A. von 1980 Palaeozoogeografia de Jurhsico marino (Lias hasta Oxfordiano) en Suramerica. In Zeil, W. (ed.): Nuevos Resultados de la lnvestigacibn Geocienrifica Alemana en Latinoanikrica, 132-134. Deutsche Forschungsgemeinschaft, Bonn, and lnstituto de Colabo- raci6n Cientifica, Tubingen.
Hillebrandt, A. von 1990 The TriassiclJurassic boundary in northern Chile. Cahiers de Universitk Catholique Lyon, Series des Sciences 3,
Hillebrandt, A. von & Schmidt-Effing, R. 1981: Ammoniten aus dem Toarcium (Jura) von Chile (Sudamerika). Zineliatia 6, 3-74.
Hillebrandt, A. von & Westermann, G.E.G., 1985: Aalenian (Jurassic) ammonite faunas and zones of the Southern Andes. Zitteliana 12, 3-55.
Hillebrandt, A. von, Groschke, M., Prinz, P. & Wilke. H.-G. 1986 Ma- rines Mesozoikum in Nordchile zwischen 21" und 26's. Berliner Ge- owissenschafiliche Abhandlungen A 66, 169- 190.
Jablonski, D. 1985: Marine regressions and mass extinctions: a test using modern biota. In Valentine, J.W. (ed.): Phanerozoic diversitypatterns, 335-354. Princeton University Press, Princeton, New Jersey.
Jablonski, D. 1986: Causes and consequences of mass extinctions: a comparative approach. In Elliott, D.K. (ed.): Dynamics of Extitiction, 183-229. John Wiley, New York. N.Y.
71-108.
31/32,167-199.
27-53.
LETHAIA 29 (1996) Diversity of Jurassic bivalves 195
Jenkyns, H.C. 1988: The Early Toarcian (Jurassic) anoxic event: strati- graphic, sedimentary, and geochemical evidence. American Journal of Science 288, 10 1-1 5 I .
Jeppsson, L. 1990: An oceanic model for lithological and faunal changes tested on the Silurian record. Journal ofthe Geological Society, London 147.663474.
Johnson, J.G. 1974: Extinction of perched faunas. Geology2,479482. Leanza, H.A. 1993: Jurassic and Cretaceous trigoniid bivalves from west-
central Argentina. Bulletins ofAmerican Paleontology 105,5-95. Little, C.T.S. & Benton, M.J. 1995 Early Jurassic mass extinction: a do-
bal long-term event. Geology 23,495-498. McRoberts, C.A. & Newton, C.R. 1995: Selective extinction among end-
Triassic European bivalves. Geology 23, 102-104. McRoberts, C.A.. Newton, C.R. & Allasinaz, A. 1995: End-Triassic bi-
Mpodozis, A,, Rivano, S. & Vicente, J.C. 1973: Resultados preliminares del estudio geol6gico de la Aka Cordillera de Ovalle entre 10s rios Grande y Los Molles (Prov. de Coquirnbo, Chile). Actas Quinto Con- greso Geologico Argentino, Tomo 4, 117-132.
Naranjo, J.A. & l’uig, A. 1984: Hojas Taltal y Chaiiaral, regiones de An- tofagasta y Atacama. Servicio Nacional de Geologia y Minerfa, Carta Geolbgica de Chile 62-63, 1-140.
Odin, G.S. 1992: Numerical time scale in 1988. In Westermann, G.E.G., (ed.): The Jurassic ofthe Circum-Pacific, 3-1 1. Cambridge University Press, Cambridge.
Perez, E. d’A. 1982: Bioestratigrafia del Jurasico de Quebrada Asientos, Norte de Potrerillos, Regidn de Atacarna. Servicio Nacional de Ge- ologia y Mineria - Chile, Boletin 37, 1-149.
Perez, E. &A. & Levi, B. 1961: Relaci6n estratigrafica entre la formaci6n Moctezuma y el granito subyacente, Calama, Provincia de Antofagas- ti. Minerales 74,3948.
Perez, E. d’A. & Reyes, R. B. 1994: CatPlogo de ejemplares tipo. conser- vados en la coleccidn paleontol6gica del Servicio Nacional de Ge- ologia y Mineria, Chile, desde 1953 a 1993. Servicio Nacional de Ge- ologia y Mineria - Chile, Boletin 46, 1-99.
Polubotko, 1.V. & Repin, Yu.S. 1988: Lower and Middle Jurassic of the North-East. Newsletters on Stratigraphy 19, 1-17.
Quinzio, L.A. 1987: Stratigraphische Untersuchungen im Unterjura des Siidteils der Provinz Antofagasta in Nord-Chile. Berliner geowissen- schafiliche Abhandlungen A 87, 1-106.
Raup, D.M. 1986: Biological extinction in earth history. Science 231, 1528-1533.
Raup, D.M. & Sepkoski, J.J., Jr. 1984: Periodicity ofextinctions in thege- ologic past. Proceedings of the National Academy ofsciences USA 81,
Riccardi, A.C. 1983: The Jurassic of Argentina and Chile. In Moullade, M. & Nairn, A.E.M. (eds.): The Phanerozoic Geology ofthe World 11. The Mesozoic, B, 201-263. Elsevier, Amsterdam.
Riccardi, A.C., Gulisano, C.A., Mojica, J., Palacios, O., Schubert, C. & Thomson, M.R.A. 1992 Western South America and Antarctica. In Westermann, G.E.G., (ed.): The Jurassic of the Circum-Pacific, 122- 161. Cambridge University Press, Cambridge.
Riccardi, A.C., Damborenea, S.E. & Manceiiido, M.O. 1990: Lower Jurassic of South America and Antarctic Peninsula. Newsletters on Stratigraphy21,75-103.
Sepkoski, JJ., Jr. 1989 Periodicity in extinction and the problem ofcat- astrophism in the history of life. Journal ofthe Geological Society Lon-
Sepkoski, J.J., Jr. & Raup, D.M. 1986 Periodicity in marine extinction events. In Elliott, D.K. (ed.): Dynamics of .Extinction, 3-36. Wiley, New York, N.Y.
Sepulveda, P. & Naranjo, J.A. 1982 Geologia de la hoja carrera Pinto, Regidn de Atacama. Servicio Nacional de Geologia y Mineria, Carta Geolbpca de Chile 53, 1-60,
Sey, 1.1. & Kalacheva, E.D. 1988: Ammonites and bivalves of the Far East. Newsletters on Stratigraphy 19,35455.
Smith, P.L. & Tipper, H.W. 1986: Plate tectonics and paleobiogeogra- phy: Early Jurassic (Pliensbachian) endemism and diversity. Palaios I , 399412.
Smith, P.L., Beyers, J.M., Carter! E.S. Jakobs, G.K., Pal@, J.. Pessagno, E. & Tipper, H. E. 1994: Lower Jurassic. Newsletters on Stratigraphy 31,
Wignall, P.B. 1994 Black Shales. Oxford Monographs on Geology and
Wyatt, A.R. 1987: Shallow water areas in space and time. Journal ofthe