j our na l ho me p age: www.elsev ier .com/ locate / t i ce
gg structure of Zorotypus caudelli Karny (Insecta, Zoraptera, Zorotypidae)
uta Mashimoa,∗, Ryuichiro Machidaa, Romano Dallaib, Marco Gottardob,avid Mercatib, Rolf G. Beutel c
Sugadaira Montane Research Center, University of Tsukuba, Sugadaira Kogen, Ueda, Nagano 386-2204, JapanDepartment of Evolutionary Biology, University of Siena, Via A. Moro 2, I-53100 Siena, ItalyInstitut für Spezielle Zoologie und Evolutionsbiologie mit Phyletischem Museum, Friedrich-Schiller-Universität Jena, Erbertstraße 1, 07743 Jena, Germany
r t i c l e i n f o
rticle history:eceived 12 March 2011eceived in revised form 3 April 2011ccepted 4 April 2011vailable online 7 May 2011
eywords:
a b s t r a c t
The structural features of eggs of Zorotypus caudelli Karny are described in detail. The egg is elliptic withlong and short diameters of 0.6 and 0.3 mm respectively, and creamy white. The egg shows a honeycombpattern on its surface, without any specialized structures for hatching such as an operculum or a hatchingline. The fringe formed by a fibrillar substance secreted after the completion of the chorion encircles thelateral surface. The egg layer is composed of an exochorion, an endochorion, and a vitelline envelope. Theexochorion and endochorion are electron-dense and homogeneous in structure. The exochorion shows
a perforation of numerous branching aeropyles. The exo- and endochorion are connected by numeroussmall columnar structures derived from the latter. The vitelline envelope is very thin and more electron-dense than the chorion. A pair of micropyles is present at the equator on the dorsal side of the egg.Originating at the micropyle, the micropylar canal runs through the chorion obliquely. The structuralfeatures of the eggs of Zoraptera were compared with those of other polyneopteran and paraneopteranorders.
araneoptera
. Introduction
With only ca. 40 fossil and extant species Zoraptera is one ofhe smallest hexapod orders and the small ground living insectsre inconspicuous and characterized by a cryptic life style. Never-heless, it is a fascinating group with alate and wingless morphs,regarious habits, and a wide distribution in warm regions (absentrom Australia). From the phylogenetic point of view, Zoraptera arendoubtedly the most enigmatic group of the entire Hexapoda. Not
ess than 10 different phylogenetic positions have been proposedsee Engel and Grimaldi, 2002) since the group was introducedy Silvestri (1913). In his groundbreaking work “Die Stammes-eschichte der Insekten” Hennig (1969) considered Zoraptera as theistergroup of Acercaria (Hemiptera, Thysanoptera, Psocoptera andhthiraptera), both forming together a monophyletic group Parane-ptera. This lineage is suggested by reductional features such as theeduced number of tarsomeres (2/3) and Malpghian tubules (6/4)nd the strongly condensed abdominal ganglionic chain (2/1 gan-lionic masses). Several more recent studies supported either their
istergroup relationship with Acercaria (Beutel and Weide, 2005)n agreement with Hennig (1969), or with Eumetabola (Beutel andorb, 2001), or Holometabola (Rasnitsyn, 1998). In contrast to these
concepts, other recent phylogenetic investigations suggested acloser relationship with polyneopteran groups, either as closest rel-ative of Dictyoptera (Wheeler et al., 2001), Embioptera (Yoshizawa,2007), or Dermaptera (Terry and Whiting, 2005). To highlight thehighly controversial systematic status of the enigmatic insect order,Beutel and Weide (2005) coined the term ‘Zoraptera problem’,modelled after the ‘Strepsiptera problem’ (Kristensen, 1991).
The unclear systematic position underlines that a more inten-sive investigation of different aspects and character systems ofthese mysterious insects is required. In the last 10 years severaldetailed morphological studies of Zoraptera have emerged, on theskeleto-muscular systems of the head (Beutel and Weide, 2005)and thorax (Friedrich and Beutel, 2008), the wing base structures(Yoshizawa, 2007), the postabdomen (Hünefeld, 2007), and thereproductive system (Dallai et al., in press-a, in press-b). How-ever, information on their embryogenesis is still totally lacking.We started an embryological study of Zoraptera using Zorotypuscaudelli Karny. In the present paper, as the first step of our study,we describe the egg structure of Z. caudelli and compare it with thecondition found in other neopteran lineages.
2. Materials and methods
About 300 adult and juvenile specimens Z. caudelliwere collected under the bark of decaying trees in tropicalforests in the Genting and Cameron Highlands, Malaysia, in
Y. Mashimo et al. / Tissue and Cell 43 (2011) 230– 237 231
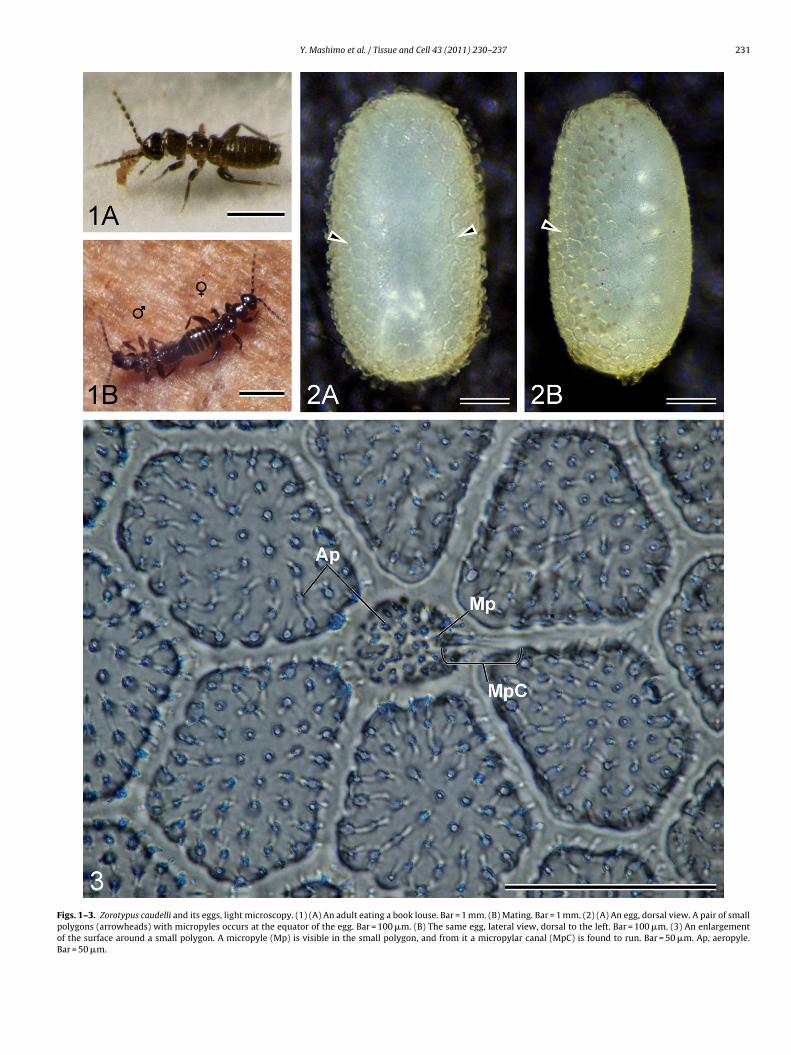
Figs. 1–3. Zorotypus caudelli and its eggs, light microscopy. (1) (A) An adult eating a book louse. Bar = 1 mm. (B) Mating. Bar = 1 mm. (2) (A) An egg, dorsal view. A pair of smallpolygons (arrowheads) with micropyles occurs at the equator of the egg. Bar = 100 �m. (B) The same egg, lateral view, dorsal to the left. Bar = 100 �m. (3) An enlargementof the surface around a small polygon. A micropyle (Mp) is visible in the small polygon, and from it a micropylar canal (MpC) is found to run. Bar = 50 �m. Ap, aeropyle.Bar = 50 �m.
232 Y. Mashimo et al. / Tissue and Cell 43 (2011) 230– 237
e and
F(twlic
et8(cficsc
oll3eeL
gTubete
Cwco
waaNwka
3
Mazlatd
a(
FaAa
Y. Mashimo et al. / Tissu
ebruary 2009 and April 2010. They were reared in a plastic case10 cm × 10 cm × 10 cm) containing wet tissue paper at roomemperature. They were fed on dry yeast, powdered dried silkorm pupae (commercially sold bait for fishing) and living book
ice (Liposcelis sp.) (Fig. 1A). Collected eggs deposited by femalesn the rearing cases were transferred to other plastic cases alsoontaining wet tissue paper and incubated at 25 ◦C.
Some eggs were fixed with Carnoy’s fixative (100%thanol:chloroform:acetic acid = 6:3:1) for 10 min, transferredo 80% ethanol, punctured with a fine needle, and stored in0% ethanol. Other eggs were soaked in Karnovsky’s fixative2% paraformaldehyde+2.5% glutaraldehyde 0.1 M HCl–sodiumacodylate buffer solution, pH 7.2) for 1 min, punctured with ane needle, and fixed for 1 h. After making a small opening in thehorion with fine forceps, the eggs were further fixed with theame fixative at 4 ◦C for 24 h and then stored in a 0.1 M HCl–sodiumacodylate buffer, pH 7.2, at 4 ◦C.
General features of the eggs were observed under a stere-microscope Leica MZ12. To observe the micropyles, the eggayers were mounted in a polyvinyl-lactophenol medium, Heinziquid (polyvinyl-alcohol 10 g + distilled water 80 ml + lactic acid5 ml + glycerin 10 ml + phenol 25 ml + chloral hydrate 20 g), andxamined under a biological microscope Nikon OPTIPHOTOquipped with Nikon Plan Apo objectives for light field images or aeica DM6000B for differential interference contrast images.
Some eggs fixed with Karnovsky’s fixative were dehydrated in araded ethanol series and embedded in a methacrylate resin Kulzerechnovit 7100. Semithin sectioning (2 �m) was performed cutsing a Bio-Rad H-1500 microtome equipped with a tungsten car-ide steel knife (Meiwa Superhard Knife), as described by Machidat al. (1994a,b). Sections were stained with 1% Delafield’s haema-oxylin for 18 h, 0.5% eosin G for 1 h, and 0.5% fast green FCF 80%thanol solution for 1 min.
For scanning electron microscopy (SEM), some eggs treated witharnoy’s fixative were post-fixed with 1% OsO4 for 1 h. Fixed eggsere dehydrated in a graded ethanol series, dried at the criti-
al point (tousimis Samdri®-PVT-3D), coated with gold, and thenbserved under a TOPCON SM-300 SEM.
For transmission electron microscopy (TEM), eggs were fixedith Karnovsky’s fixative containing 1% tannic acid at 4 ◦C for 24 h
nd post-fixed with 1% OsO4 for 1 h. Fixed eggs were dehydrated in graded acetone series, embedded in water-miscible epoxy resin,isshin EM Quetol 651, and processed into sections 0.1 �m thickith an ultramicrotome, RMC MT-XL, equipped with a diamond
nife. Sections were stained with uranyl acetate and lead citratend observed under a TOPCON LEM-2000 TEM at 90 kV.
. Results
Pairs were often observed to mate under rearing conditions.ating in Z. caudelli is of the end-to-end type with the male supine
nd dragged around by the female (Fig. 1B), as reported for otherorapterans (Shetlar, 1978; Choe, 1997). Every few days, the femaleays only one egg on the substratum. To date, we have obtainedbout 3,000 eggs. In the present study, we designate the side ofhe egg facing the substratum as ventral, and the opposite side as
orsal.
Eggs of Z. caudelli are elliptic with long and short diameters ofbout 0.6 mm and about 0.3 mm respectively, and creamy whiteFigs. 2A, B, 4A and B). The surface shows a hexagonal or less fre-
igs. 4–8. Eggs of Zorotypus caudelli, scanning electron microscopy. (4) (A) An egg, ventnd a fringe, respectively. Bar = 100 �m. (B) The same egg, lateral view, dorsal to the lefn enlargement of the dorsal side. Bar = 50 �m. (7) An enlargement of a small polygon weropyle; Fr, fringe made of a fibrillar substance; Mp, micropyle.
Cell 43 (2011) 230– 237 233
quently pentagonal, honeycomb pattern made of an exochorionicridge. The single compartments measure about 50 �m in diame-ter (Figs. 3 and 5). The honeycomb pattern is more distinct on thedorsal side (Fig. 4B). Each honeycomb contains about 50 aeropy-les about 1 �m in diameter (Figs. 3, 5 and 6). A fringe formed bya fibrillar substance encircles the lateral surface of the egg a lit-tle biased dorsally (Figs. 4A, B and 6). The surface at both polesis featureless (Fig. 8A and B). At the equator on the dorsal sideof the egg, a pair of small polygons 20 �m in diameter is found(Figs. 3, 4A, B and 7): occasionally an additional polygon is present(Fig. 9A and B). Each small polygon contains 30–40 aeropyles andone micropyle about 2 �m diameter (Fig. 3); the aeropyles are notrecognisable in Fig. 7 as they are concealed by an extrinsic substancesecreted at the oviposition. In eggs with fringe structures devel-oped the small polygons are often covered by the fringe structureand difficult to observe.
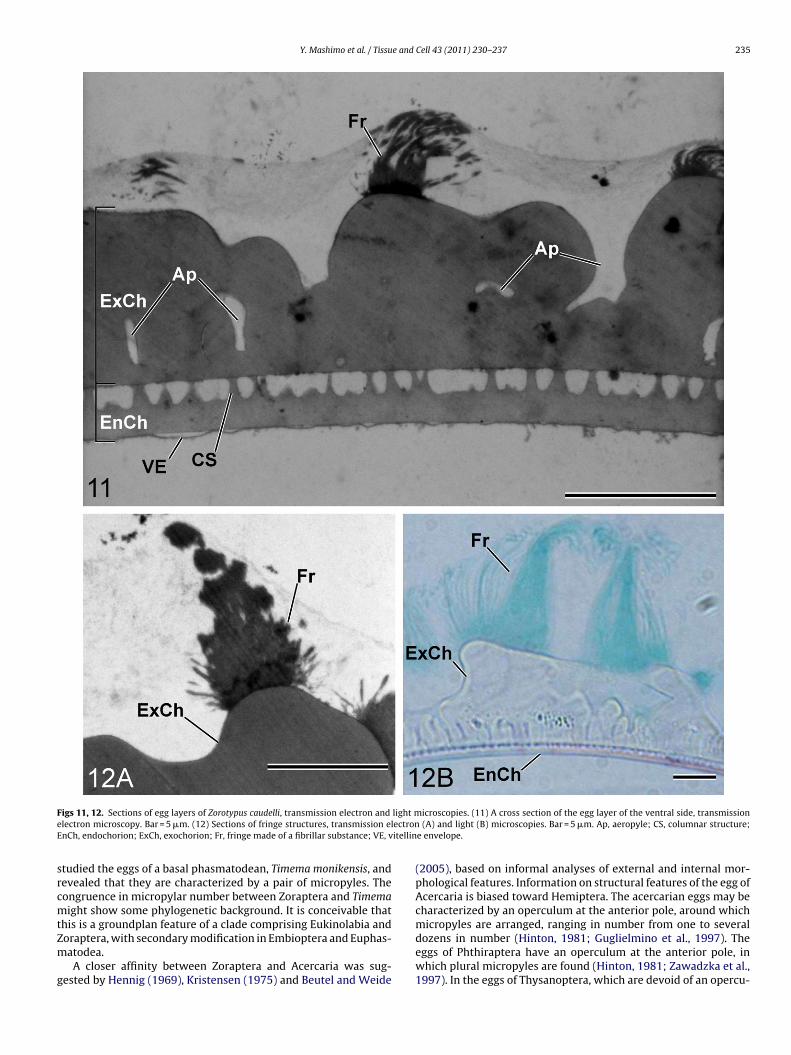
The egg layers are composed of a two-layered chorion compris-ing an exochorion and endochorion, and additionally an extremelythin vitelline envelope. The exochorion is about 5 �m thick,electron-dense and homogeneous in structure (Fig. 11). The exo-chorion forms a ridge which shows a honeycomb pattern on the eggsurface: the height of the ridge is various, e.g. about 3 �m on thedorsal side and less than 1 �m on the ventral side (Figs. 5 and 6). Thefringe is independent of the chorion, and probably secreted on tothe egg after the completion of the chorion: there is a discontinuitybetween the exochorion and fringe, and a difference in stainabil-ity, i.e. the latter shows greater affinity for fast green FCF and ismore electron-dense than the former (Fig. 12A and B). The aeropy-lar canals run through the exochorion, branching themselves, andreach the space formed between the exochorion and endochorion.
The endochorion is about 1 �m thick and homogeneous in struc-ture, with a slightly less electron-density than the exochorion(Fig. 11). The endochorion yields numerous short columnar struc-tures, about 1 �m in height and 0.2–0.4 �m in diameter, on its outersurface. The exochorion and endochorion are in contact with thesecolumns, being spaced by about 1 �m (Fig. 11). The vitelline enve-lope is an extremely thin layer adhering to the endochorion, lessthan 0.1 �m thick, and with a high electron density (Fig. 11).
From each micropyle a micropylar canal of about 15 �m lengthpenetrates the chorion and runs obliquely through it in lateraldirection (Fig. 10A). Around the micropylar canal, the small colum-nar structures on the outer surface of the endochorion are lacking,and the exochorion and endochorion are fused (Fig. 10C). At theinner opening of the micropylar canal the endochorion forms aflap, which covers the micropylar inner opening (Fig. 10B and C).The lumen of the micropylar canals is filled with a substance morebasophilic and electron-dense (Fig. 10A and C).
4. Discussion
The structural features of Zoraptera eggs have been described infive species, Zorotypus snyderi (Caudell, 1920), Zorotypus hubbardi(Gurney, 1938), Zorotypus brasiliensis (Silvestri, 1946), Zorotypusgurneyi and Zorotypus barberi (Choe, 1989). The information pro-vided is based on light and/or scanning electron microscopicobservations and is only fragmentary. We cannot find any dif-
ference between our observations of eggs of Z. caudelli and thestructural features of the eggs of the other zorapterans. The eggs ofZoraptera can be characterized as follows (new findings presentedhere indicated by italics): (1) elliptic in shape and creamy white
ral view. Arrowheads and arrows show a pair of small polygons with micropylest. Bar = 100 �m. (5) An enlargement of the ventral side of the egg. Bar = 50 �m. (6)ith a micropyle. Bar = 10 �m. (8) (A and B) Polar views of an egg. Bar = 100 �m. Ap,
234 Y. Mashimo et al. / Tissue and Cell 43 (2011) 230– 237
Figs. 9, 10. Eggs and micropylar structures of Zorotypus caudelli, light and scanning and transmission electron microscopies. (9) (A) An egg, dorsal view. Arrowheads showa pair of polygons and an additional one. Bar = 100 �m. (B) Enlargement. Bar = 50 �m. (10) (A) A longitudinal section of the micropylar canal (MpC), light microscopy. Anasterisk shows the basophilic substance filling the micropylar canal. Bar = 5 �m. (B) A flap (F) covering the inner opening of the micropylar canal, scanning electron microscopy.B An asA Ch, fum
ictnhodmtmloawfdi1fa
ZcE2ooomCeSsoa
ar = 5 �m. (C) A section of the micropylar canal, transmission electron microscopy.p, aeropyle; CS, columnar structure; EnCh, endochorion; ExCh, exochorion; Ex-Enicropylar canal.
n color; (2) a honeycomb pattern in their surface; (3) egg layersomposed of an exochorion, an endochorion and a vitelline envelope;he exochorion is electron-dense and homogeneous in structure withumerous branching aeropyles; the endochorion is electron-dense,omogeneous, and bears numerous small columnar structures on itsuter surface; (4) a pair of small polygons at the equator on theorsal side of the egg; each polygon contains one micropyle; theicropylar canal runs obliquely through the chorion in lateral direc-
ion; the exochorion and endochorion are fused together around theicropylar canal; the endochorion at the inner opening of the micropy-
ar canal forms a flap; (5) no structures specialized for hatching (e.g.perculum, hatching line). The eggs of Z. caudelli are equipped with
fringe structure encircling the lateral surface. Similar featuresere documented in Choe’s (1989) scanning electron micrographs
or Z. barberi but not for Z. gurneyi. Such structures were neitherescribed in the study on Z. snyderi (Caudell, 1920) nor depicted
n those on Z. hubbardi and Z. brasiliensis (Gurney, 1938; Silvestri,946). The fringe is an extrinsic structure secreted onto the egg sur-ace after the completion of the egg, and the secretion may differmong the species.
As mentioned in the introduction, the phylogenetic position oforaptera is discussed very controversially. The main sistergroupandidates are the polyneopteran Dictyoptera, Dermaptera, andmbioptera, and alternatively Paraneoptera (Engel and Grimaldi,002; Yoshizawa, 2007). We will compare the structural featuresf the eggs of Zoraptera with those of these four groups. Dicty-ptera was suggested as the sister-group or at least a close relativef Zoraptera based on morphological evidence and analyses ofolecular or combined data sets (Silvestri, 1913; Caudell, 1918;
rampton, 1920; Weidner, 1969, 1970; Boudreaux, 1979; Wheelert al., 2001; Yoshizawa and Johnson, 2005; Ishiwata et al., 2011).
everal studies on the egg structure of Dictyoptera are available,uch as those of Iwaikawa et al. (1981) and Iwaikawa and Ogi (1982)n Mantodea, Hinton (1981) and Bellés et al. (1993) on Blattodea,nd Knower (1900), Mukerji (1970), Grandi (1990) and Grandi and
terisk shows the electron-denser substance filling the micropylar canal. Bar = 2 �m.sion of the exochorion and endochorion; F, flap covering the inner opening of the
Chicca (1999) on Isoptera. Although the information is fragmen-tary, the dictyopteran egg structure may be characterized by pluralmicropyles circularly arranged around the anterior or posteriorpole (circularly arranged at the level of equator of the egg in thetermite Zootermopsis nevadensis [Striebel, 1960]). A hatching fieldis developed in the eggs of Mantodea and Isoptera. In the eggsof Mantodea and Blattodea, which are protected by the ootheca,the chorion is very fragile, and the endochorion is laminal. Thus,we cannot find any common features of the egg structure sug-gesting phylogenetic affinities between Zoraptera and Dictyoptera.However, it has to be taken into consideration that a hypotheticalancestral condition is likely secondarily modified in extant mem-bers of the Dictyoptera due to the presence of the ootheca in thegroundplan of this lineage.
A sister-group relationship between Zoraptera and Dermapterawas proposed based on analyses of molecular and combined datasets (Carpenter and Wheeler, 1999; Terry and Whiting, 2005).According to Chauvin et al. (1991), the eggs of Dermaptera are char-acterized by a five-layered chorion, aeropyles arranged in a circle,and the presence of one micropyle at the anterior pole of the egg.None of these features is found in zorapteran eggs.
Engel and Grimaldi (2002) based on morphological and eco-logical features and Yoshizawa (2007) based on a comparison ofwing base suggested a sistergroup relationship between Zorapteraand Embioptera. Jintsu et al. (2007) and Jintsu and Machida (2009)characterized the eggs of Embioptera, describing a specializedmicropyle-related chorion structure, the micropylar tube, an oper-culum, and a polar mound. Again, all these features are absentin Zoraptera. However, there is one thing worth mentioning inthis context. Terry and Whiting (2005) suggested a close phyloge-netic affinity between Embioptera and Phasmatodea, both forming
a clade Eukinolabia, and this was also supported in recent morpho-logical studies on head structures (Wipfler et al., 2011; Friedemannet al., 2011). Jintsu et al. (2007) and Jintsu and Machida (2009) sup-ported Eukinolabia, based on features of the eggs. Jintsu et al. (2010)
Y. Mashimo et al. / Tissue and Cell 43 (2011) 230– 237 235
F light me ectronE itellin
srcmtZm
g
igs 11, 12. Sections of egg layers of Zorotypus caudelli, transmission electron and
lectron microscopy. Bar = 5 �m. (12) Sections of fringe structures, transmission elnCh, endochorion; ExCh, exochorion; Fr, fringe made of a fibrillar substance; VE, v
tudied the eggs of a basal phasmatodean, Timema monikensis, andevealed that they are characterized by a pair of micropyles. Theongruence in micropylar number between Zoraptera and Timemaight show some phylogenetic background. It is conceivable that
his is a groundplan feature of a clade comprising Eukinolabia and
oraptera, with secondary modification in Embioptera and Euphas-atodea.A closer affinity between Zoraptera and Acercaria was sug-
ested by Hennig (1969), Kristensen (1975) and Beutel and Weide
icroscopies. (11) A cross section of the egg layer of the ventral side, transmission (A) and light (B) microscopies. Bar = 5 �m. Ap, aeropyle; CS, columnar structure;
e envelope.
(2005), based on informal analyses of external and internal mor-phological features. Information on structural features of the egg ofAcercaria is biased toward Hemiptera. The acercarian eggs may becharacterized by an operculum at the anterior pole, around whichmicropyles are arranged, ranging in number from one to several
dozens in number (Hinton, 1981; Guglielmino et al., 1997). Theeggs of Phthiraptera have an operculum at the anterior pole, inwhich plural micropyles are found (Hinton, 1981; Zawadzka et al.,1997). In the eggs of Thysanoptera, which are devoid of an opercu-
2 e and
l1ivWtppttsoecS
fetwpt
dafefalwdNtkZtshdsr
A
aitpFiSeCM
R
B
B
B
36 Y. Mashimo et al. / Tissu
um, a single micropyle is found near the posterior pole (Heming,979; Haga, 1985). The egg structure of Psocoptera was described
n detail by Seeger (1979) and some information was also pro-ided by Goss (1954) and Hinton (1981). An operculum is absent.ith an average thickness of 0.5 �m the chorion is exceptionally
hin, and correlated with this, micropyles and aeropyles are com-letely reduced (Seeger, 1979). Furrows, rows of impressions andapillae (5–10 �m) in Trogiomorpha were interpreted as forma-ions of the vitelline envelope by Seeger (1979). Desiccation of thehin walled eggs and negative mechanical impact are prevented bytrongly corrugated sculptures of the serosal cuticle, encrustationsf anal secretes or silk of the labial silk-glands (Seeger, 1979). Thexceptionally thin chorion and the absence of pylar openings wereonsidered as autapomorphies of Psocoptera by Seeger (1979).tructural affinities with the zorapteran eggs are not recognisable.
Apparently the available information on acercarian eggs is stillragmentary and the groundplan of the entire clade is far from beingstablished. What can be said at present is that the known struc-ural features of eggs occurring in acercarian lineages do not fit wellith what is found in Zoraptera. There is no apparent evidence sup-ortive of a lineage Paraneoptera including Zoraptera as the sisteraxon of Acercaria (e.g. Hennig, 1969).
As mention above the main purpose of this study was a detailedocumentation of features of the zorapteran egg. However, it ispparent that the presently available information is not sufficientor a reliable phylogenetic interpretation. The information on thegg structure of polyneopteran and acercarian groups is still tooragmentary for a formal character analysis. Moreover, it is conceiv-ble that potential synapomorphic egg features on an interordinalevel may be rarely preserved, due to secondary modifications
ithin the lineages. What should have priority is to screen andocument characters of the egg for a representative sample ofeoptera, with a specific focus on presumably basal representa-
ives of the orders (or more inclusive lineages) and presumptiveey taxa, (e.g. Mantoida [Mantodea], Timematodea [Phasmatodea],oraptera, Psocoptera). This will allow in future studies to includehis specific data set into an extensive matrix including a widepectrum of morphological features. An analysis of such a set ofolomorphological characters combined with extensive molecularata will likely allow a solid reconstruction of neopteran relation-hips, a well-founded placement of the enigmatic Zoraptera, and aeliable interpretation of the evolution of egg structures.
cknowledgments
We thank Drs. T. Uchifune, Y. Uchifune, M. Fukui, Messrs. Y. Nak-gaki, K. Sekiya, S. Shimizu, S. Tomizuka and M. Fujita for their helpn collecting material, and the director Prof. O. Numata and staff ofhe Sugadaira Montane Research Center, University of Tsukuba forroviding facilities. We also thank Messrs. Kim Hong Yap, Mohamadadil and Shahrul Nazly B. Mahmud for their assistance in collect-ng material in Malaysia. This study was supported by a Sasakawacientific Research Grant (22-504) from The Japan Science Soci-ty to YM and a Grant-in-Aid from the JSPS (Scientific Research: 21570089) to RM. This is the contribution from the Sugadairaontane Research Center, University of Tsukuba.
eferences
ellés, X., Cassier, P., Cerdá, X., Pascual, N., André, M., Rósso, Y., Piulachs, M.D., 1993.Induction of choriogenesis by 20-hydroxyecdysone in the German cockroach.Tissue Cell 25, 195–204.
eutel, R.G., Gorb, S.N., 2001. Ultrastructure of attachment specializations of
hexapods (Arthropoda): evolutionary patterns inferred from a revised ordinalphylogeny. J. Zool. Syst. Evol. Res. 39, 177–207.
eutel, R.G., Weide, D., 2005. Cephalic anatomy of Zorotypus hubbardi (Hexapoda:Zoraptera): new evidence for a relationship with Acercaria. Zoomorphology 124,121–136.
Cell 43 (2011) 230– 237
Boudreaux, H.B., 1979. Arthropod Phylogeny with Special Reference to Insects. JohnWiley and Sons, New York, 320 pp.
Carpenter, J.M., Wheeler, W.C., 1999. Cladística numérica, análisis simultáneo y filo-genia de hexápodos. Bol. Soc. Entomol. Aragon. 26, 333–346.
Caudell, A.N., 1918. Zorotypus hubbardi, a new species of the order Zoraptera fromthe United States. Can. Entomol. 50, 375–381.
Caudell, A.N., 1920. Zoraptera not an apterous order. Proc. Entomol. Soc. Wash. 22,84–97.
Chauvin, G., Hamon, C., Vancassel, M., Vannier, G., 1991. The eggs of Forficula auricu-laria L. (Dermaptera, Forficulidae): ultrastructure and resistance to low and hightemperatures. Can. J. Zool. 69, 2873–2878.
Choe, J.C., 1989. Zorotypus gurneyi, new species, from Panama and redescription of Z.barberi Gurney (Zoraptera: Zorotypidae). Ann. Entomol. Soc. Am. 82, 149–155.
Choe, J.C., 1997. The evolution of mating systems in the Zoraptera: mating variationsand sexual conflicts. In: Choe, J.C., Cresbi, B.J. (Eds.), The Evolution of MatingSystems in Insects and Arachnids. Cambridge University Press, Cambridge, pp.130–145.
Crampton, G.C., 1920. Some anatomical details of the remarkable winged zorapteronZorotypus hubbardis [sic] Caudell, with notes on its relationships. Proc. Entomol.Soc. Wash. 22, 98–106.
Dallai, R., Mercati, D., Gottardo, M., Machida, R., Mashimo, Y., Beutel, R.G. Thefine structure of the female reproductive system of Zorotypus caudelli Karny(Zoraptera). Arthropod Struct. Dev., in press-a.
Dallai, R., Mercati, D., Gottardo, M., Machida, R., Mashimo, Y., Beutel, R.G. The malereproductive system of Zorotypus caudelli Karny (Zoraptera): sperm structureand spermiogenesis. Arthropod Struct. Dev., in press-b.
Engel, M.S., Grimaldi, D.A., 2002. The first Mesozoic Zoraptera (Insecta). Am. Mus.Novit. 3362, 1–20.
Friedemann, K., Wipfler, B., Bradler, S., Beutel, R.G., 2011. On the head morphologyof Phyllium and the phylogenetic relationships of Phasmatodea (Insecta). ActaZool., doi:10.1111/j.1463-6395.2010.00497.x.
Friedrich, F., Beutel, R.G., 2008. The thorax of Zorotypus (Hexapoda, Zoraptera) and anew nomenclature for the musculature of Neoptera. Arthropod Struct. Dev. 37,29–54.
Goss, R.J., 1954. Ovarian development and oogenesis in the book louse Liposcelisdivergens Badonnel (Psocoptera, Liposcelidae). Ann. Entomol. Soc. Am. 90,190–207.
Grandi, G., 1990. Oogenesis in Kalotermes flavicollis (Fabr.) (Isoptera, Kalotermiti-dae) III. Choriogenesis and corpus luteum formation in female supplementaryreproductives. Boll. Zool. 57, 97–107.
Grandi, G., Chicca, M., 1999. Oogenesis in supplementary reproductives of Retic-ulitermes lucifugus Rossi (Isoptera Rhinotermitidae): an ultrastructural study.Invert. Rep. Dev. 35, 65–79.
Guglielmino, A., Taddei, A.R., Carcupino, M., 1997. Fine structure of the eggshellof Ommatissus binotatus Fieber (Homoptera, Auchenorrhyncha, Tropiduchidae).Int. J. Insect Morphol. Embryol. 26, 85–89.
Gurney, A.B., 1938. A synopsis of the order Zoraptera, with notes on the biology ofZorotypus hubbardi Caudell. Proc. Entomol. Soc. Wash. 40, 57–87.
Haga, K., 1985. Oogenesis and embryogenesis of the idolothripine thrips, Bactrothripsbrevitubus (Thysanoptera, Phlaeothripidae). In: Ando, H., Miya, K. (Eds.), RecentAdvances in Insect Embryology in Japan. ISEBU, Tsukuba, pp. 45–106.
Heming, B.S., 1979. Origin and fate of germ cells in male and female embryos of Hap-lothrips verbasci (Osborn) (Insecta, Thysanoptera, Phlaeothripidae). J. Morphol.160, 323–344.
Hennig, W., 1969. Die Stammesgeschichte der Insekten. Waldemar Kramer, Frank-furt, 436 pp.
Hinton, H.E., 1981. Biology of Insect Eggs, vol. 2. Pergamon Press, Oxford, 304 pp.Hünefeld, F., 2007. The genital morphology of Zorotypus hubbardi Caudell, 1918
(Insecta: Zoraptera: Zorotypidae). Zoomorphology 126, 135–151.Ishiwata, K., Sasaki, G., Ogawa, J., Miyata, T., Su, Z.-H., 2011. Phylogenetic rela-
tionships among insect orders based on three nuclear protein-coding genesequences. Mol. Phyl. Evol. 58, 169–180.
Iwaikawa, Y., Ogi, K., 1982. Chorionic structures of the egg shells of mantis, Ten-odera aridifolia (Dictyoptera: Mantidae). Res. Bull. (Nat. Sci. Phychol.), Collageof General Education, Nagoya University 26, 69–83 (in Japanese with Englishsummary).
Iwaikawa, Y., Ogi, K., Ito, M., 1981. Structures of the egg shells of mantis, Tenoderaaridifolia (Dictyoptera: Mantidae). Zool. Mag. 90, 652 (in Japanese).
Jintsu, Y., Machida, R., 2009. TEM observations of the egg membranes of a web-spinner, Aposthonia japonica (Okajima) (Insecta: Embioptera). Proc. Arthropod.Embryol. Soc. Jpn. 44, 19–24.
Jintsu, Y., Uchifune, T., Machida, R., 2007. Egg membranes of a web-spinner, Apos-thonia japonica (Okajima) (Insecta: Embioptera). Proc. Arthropod. Embryol. Soc.Jpn. 42, 1–5.
Jintsu, Y., Uchifune, T., Machida, R., 2010. Structural features of eggs of the basalphasmatodean Timema monikensis Vickery and Sandoval, 1998 (Insecta: Phas-matodea: Timematidae). Arthropod Syst. Phyl. 68, 71–78.
Knower, H.M., 1900. The embryology of a termite. J. Morphol. 16, 505–568.Kristensen, N.P., 1975. The phylogeny of hexapod “orders”. A critical review of recent
accounts. J. Zool. Syst. Evol. 13, 1–44.Kristensen, N.P., 1991. Phylogeny of extant hexapods. In: CRISO (Ed.), The Insects of
Australia, vol. 1, 2nd ed. Melbourne University Press, Carlton, pp. 125–140.Machida, R., Nagashima, T., Ando, H., 1994a. Embryonic development of the jumping
bristletail Pedetontus unimaculatus Machida, with special reference to embry-onic membranes (Hexapoda: Microcoryphia, Machilidae). J. Morphol. 220,147–165.
e and
M
M
R
S
S
S
S
Y. Mashimo et al. / Tissu
achida, R., Nagashima, T., Yokoyama, T., 1994b. Mesoderm segregation of a jump-ing bristletail, Pedetontus unimaculatus Machida (Hexapoda, Microcoryphia),with a note on an automatic vacuum infiltrator. Proc. Arthropod. Embryol. Soc.Jpn. 29, 23–24 (in Japanese).
ukerji, D., 1970. Embryology of termites. In: Krishma, K., Weesner, F.M.(Eds.), Biology of Termites, vol. 2. Academic Press, New York/London,pp. 37–72.
asnitsyn, A.P., 1998. On the Taxonomic position of the insect order Zorotyp-ida = Zoraptera. Zool. Anz. 237, 185–194.
eeger, W., 1979. Spezialmerkmale an eihüllen und embryonen von Psocoptera imvergleich zu anderen Paraneoptera (Insecta); Psocpotera als monophyletischegruppe. Stutt. Beit. Natur. Ser. A (Biol.) 329, 1–57.
hetlar, D.J., 1978. Biological observations on Zorotypus hubbardi Caudell (Zoraptera).
Entomol. News 89, 217–223.
ilvestri, F., 1913. Descrizione di un nuovo ordine di insetti. Boll. Lab. Zool. Gen. Agr.Portici 7, 193–209.
ilvestri, F., 1946. Descrizione di due specie neotropicali di Zorotypus (Insecta,Zoraptera). Boll. Lab. Entomol. Agr. Portici 7, 1–12.
Cell 43 (2011) 230– 237 237
Striebel, H., 1960. Zur Embryonalentwicklung der Termiten. Acta Trop. 17, 193–260.Terry, M.D., Whiting, M.F., 2005. Mantophasmatodea and phylogeny of the lower
neopterous insects. Cladistics 21, 240–257.Weidner, H., 1969. Die Ordnung Zoraptera oder Bodenläuse. Entomol. Z. 79, 29–51.Weidner, H., 1970. Zoraptera (Bodenläuse). In: Helmcke, J.-G., Starck, D., Wermuth,
H. (Eds.), Handbuch der Zoologie, vol. 4. Walter de Gruyter, Berlin, pp. 1–12.Wheeler, W.C., Whiting, M., Wheeler, Q.D., Carpenter, J.M., 2001. The phylogeny the
extant hexapod orders. Cladistics 17, 113–169.Wipfler, B., Machida, R., Müller, B., Beutel, R.G., 2011. On the head morphology of
Grylloblattodea (Insecta) and the systematic position of the order - ,with a newnomenclature for the head muscles of Dicondylia. Syst. Entomol. 36, 241–266.
Yoshizawa, K., 2007. The Zoraptera problem: evidence for Zoraptera + Embiodeafrom the wing base. Syst. Entomol. 32, 197–204.
Yoshizawa, K., Johnson, K.P., 2005. Aligned 18S for Zoraptera (Insecta): phylogeneticposition and molecular evolution. Mol. Phyl. Evol. 37, 572–580.
Zawadzka, M., Jankowska, W., Bilinski, S.M., 1997. Egg shells of mallophagans andanoplurans (Insecta: Phthiraptera): morphogenesis of specialized regions andthe relation to F-actin cytoskeleton of follicular cells. Tissue Cell 29, 665–673.