Journal of Pathology J Pathol 2006; 209: 369–375 Published online 31 March 2006 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/path.1983 Original Paper Gastrointestinal stromal tumours overexpress fatty acid synthase S Rossi, 1 W Ou, 2 D Tang, 2 N Bhattacharya, 2 AP Dei Tos, 1 * JA Fletcher 2 and M Loda 2 1 Department of Pathology, Regional Hospital, Treviso, Italy 2 Department of Medical Oncology, Dana-Farber Cancer Institute and Pathology, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA, USA *Correspondence to: AP Dei Tos, Department of Pathology, Regional Hospital, Treviso, Italy. E-mail: [email protected]Received: 25 August 2005 Revised: 18 January 2006 Accepted: 23 January 2006 Abstract Fatty acid synthase (FASN), a key enzyme for de novo lipogenesis, is overexpressed in many malignant tumours and is associated with aggressive biological behaviour. FASN expression and its possible relationship with more aggressive behaviour in gastrointestinal stromal tumours (GISTs) have not been addressed to date. Here, FASN expression was assessed by immunohistochemistry in 60 primary GISTs (28 low/intermediate risk and 32 high risk) and seven metastatic GISTs. Sixteen smooth muscle gastrointestinal tumours were used as controls. FASN was overexpressed in 36 of 60 GISTs (60%): in 12 of 28 (42%) low/intermediate-risk GISTs and in 24 of 32 (75%) high-risk GISTs (p < 0.05). Two primary and seven metastatic GISTs and five GIST cell lines (GIST882, GIST430, GIST522, GIST62, and GIST48), analysed by western blot, showed variable FASN expression. Most metastatic samples expressed high levels of FASN protein. Additionally, seven of 60 GISTs showed a proliferation rate higher than 10% by Ki67 and all of them expressed FASN (p < 0.04). Finally, proliferation and apoptosis were investigated after FASN silencing in GIST882 cells, which displayed the highest FASN expression. siRNA-mediated FASN knock-down inhibited expression of the proliferation marker cyclin A, whereas no changes in p27 and cleaved PARP expression were seen. It is concluded that FASN is preferentially overexpressed in high-risk and metastatic GISTs, and that its overexpression likely contributes to cell proliferation. Copyright 2006 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd. Keywords: fatty acid synthase; gastrointestinal stromal tumour; tissue microarray; prolif- eration; siRNA Introduction Fatty acid synthase (FASN) is a cytosolic multifunc- tional enzyme that is involved in fatty acid de novo synthesis [1]. An additional FASN enzyme, encoded by a different nuclear gene and located in mitochon- dria, has also been characterized [2]. Under normal conditions, fatty acids are supplied by dietary fat and FASN is expressed at undetectable levels in most normal human tissues [1]. However, in proliferation- associated cellular processes, FASN plays an important role, allowing cancer cells to progress to S-phase [3] and protecting them from apoptosis [1,4,5,6]. Despite its low expression in normal tissues, FASN mRNA and protein are expressed at high levels in many epithelial tumours. In addition, FASN expression is often associ- ated with a poor prognosis [1]. FASN overexpression has been proposed as a prognostic indicator in mes- enchymal malignancies as well [7], but has not been examined in gastrointestinal stromal tumours (GISTs). GISTs represent the most common mesenchymal tumours of the gastrointestinal tract and their potential aggressive behaviour is currently assessed by tumour size and mitotic rate [8]. Because these often have marginal discriminatory power in terms of prognosis, there is a need for molecular markers that would allow more reliable prognostication. To this end, several molecules have been proposed as prognostic markers in GIST [9–17], but none of these appears to be sufficient, either alone or in combination, to add discriminatory power to the traditional prognostic indicators. Furthermore, they at best may represent biomarkers of behaviour and do not, to date, represent therapeutic targets. GISTs are characterized by strong expression of the KIT type III receptor tyrosine kinase and gain- of-function mutations of the KIT gene [18]. While the presence of KIT mutation has been associated with prognostic information [19,20], in a subset of GISTs KIT is wild type [21]. These instead often have platelet-derived growth factor receptor-A (PDGFR-A) mutations [22] with consequent activation of a cell- signalling pathway akin to that activated by KIT. Most metastatic GISTs undergo partial regression when treated with imatinib, a receptor tyrosine kinase Copyright 2006 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Transcript
Journal of PathologyJ Pathol 2006; 209: 369–375Published online 31 March 2006 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/path.1983
Original Paper
Gastrointestinal stromal tumours overexpress fatty acidsynthaseS Rossi,1 W Ou,2 D Tang,2 N Bhattacharya,2 AP Dei Tos,1* JA Fletcher2 and M Loda2
1Department of Pathology, Regional Hospital, Treviso, Italy2Department of Medical Oncology, Dana-Farber Cancer Institute and Pathology, Brigham and Women’s Hospital, Harvard Medical School, Boston,MA, USA
*Correspondence to:AP Dei Tos, Department ofPathology, Regional Hospital,Treviso, Italy.E-mail: [email protected]
Received: 25 August 2005Revised: 18 January 2006Accepted: 23 January 2006
AbstractFatty acid synthase (FASN), a key enzyme for de novo lipogenesis, is overexpressed inmany malignant tumours and is associated with aggressive biological behaviour. FASNexpression and its possible relationship with more aggressive behaviour in gastrointestinalstromal tumours (GISTs) have not been addressed to date. Here, FASN expression wasassessed by immunohistochemistry in 60 primary GISTs (28 low/intermediate risk and 32high risk) and seven metastatic GISTs. Sixteen smooth muscle gastrointestinal tumourswere used as controls. FASN was overexpressed in 36 of 60 GISTs (60%): in 12 of 28 (42%)low/intermediate-risk GISTs and in 24 of 32 (75%) high-risk GISTs (p < 0.05). Two primaryand seven metastatic GISTs and five GIST cell lines (GIST882, GIST430, GIST522, GIST62,and GIST48), analysed by western blot, showed variable FASN expression. Most metastaticsamples expressed high levels of FASN protein. Additionally, seven of 60 GISTs showed aproliferation rate higher than 10% by Ki67 and all of them expressed FASN (p < 0.04).Finally, proliferation and apoptosis were investigated after FASN silencing in GIST882cells, which displayed the highest FASN expression. siRNA-mediated FASN knock-downinhibited expression of the proliferation marker cyclin A, whereas no changes in p27 andcleaved PARP expression were seen. It is concluded that FASN is preferentially overexpressedin high-risk and metastatic GISTs, and that its overexpression likely contributes to cellproliferation.Copyright 2006 Pathological Society of Great Britain and Ireland. Published by JohnWiley & Sons, Ltd.
Fatty acid synthase (FASN) is a cytosolic multifunc-tional enzyme that is involved in fatty acid de novosynthesis [1]. An additional FASN enzyme, encodedby a different nuclear gene and located in mitochon-dria, has also been characterized [2]. Under normalconditions, fatty acids are supplied by dietary fat andFASN is expressed at undetectable levels in mostnormal human tissues [1]. However, in proliferation-associated cellular processes, FASN plays an importantrole, allowing cancer cells to progress to S-phase [3]and protecting them from apoptosis [1,4,5,6]. Despiteits low expression in normal tissues, FASN mRNA andprotein are expressed at high levels in many epithelialtumours. In addition, FASN expression is often associ-ated with a poor prognosis [1]. FASN overexpressionhas been proposed as a prognostic indicator in mes-enchymal malignancies as well [7], but has not beenexamined in gastrointestinal stromal tumours (GISTs).
GISTs represent the most common mesenchymaltumours of the gastrointestinal tract and their potentialaggressive behaviour is currently assessed by tumour
size and mitotic rate [8]. Because these often havemarginal discriminatory power in terms of prognosis,there is a need for molecular markers that wouldallow more reliable prognostication. To this end,several molecules have been proposed as prognosticmarkers in GIST [9–17], but none of these appearsto be sufficient, either alone or in combination, toadd discriminatory power to the traditional prognosticindicators. Furthermore, they at best may representbiomarkers of behaviour and do not, to date, representtherapeutic targets.
GISTs are characterized by strong expression ofthe KIT type III receptor tyrosine kinase and gain-of-function mutations of the KIT gene [18]. Whilethe presence of KIT mutation has been associatedwith prognostic information [19,20], in a subset ofGISTs KIT is wild type [21]. These instead often haveplatelet-derived growth factor receptor-A (PDGFR-A)mutations [22] with consequent activation of a cell-signalling pathway akin to that activated by KIT.Most metastatic GISTs undergo partial regressionwhen treated with imatinib, a receptor tyrosine kinase
Copyright 2006 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
370 S Rossi et al
inhibitor, which blocks the ligand-independent con-stitutive KIT/PDGFR-A activation [22,23]. However,after prolonged therapy, secondary resistance mayoccur [24]. Furthermore, some GISTs do not respondto imatinib ab initio [22,25]. Thus, alternative thera-peutic options are needed.
Here, we assess FASN expression by immunohis-tochemistry and western blot analysis in a series ofprimary GISTs and show that FASN expression ispreferentially found in high-risk GISTs and in GISTswith a high proliferation rate, as determined usingKi67. We also found FASN protein expression in allmetastatic tumours included in the study. In addi-tion, all of the GIST cell lines investigated, includingimatinib-resistant cell lines, expressed FASN, whileFASN gene silencing led to decreased expression ofthe cell proliferation marker cyclin A. These resultsmay have therapeutic implication in imatinib-resistantGISTs.
Materials and methods
Paraffin-embedded specimens
All mesenchymal tumours involving the oesophagus,stomach, and small and large bowel, coded as leiomy-omas, leiomyosarcomas, leiomyoblastomas, Schwan-nomas, and GISTs, were retrieved from the files ofthe Regional Hospital of Treviso from 1988 to 2003.The Institutional Review Board approved this study. Atotal of 78 cases were reviewed by two of the authors.A diagnosis of GIST was made or confirmed in 57cases, based on the combination of histological evalua-tion and KIT positivity. All positive cases showed dif-fuse strong cytoplasmic staining. Three cases, despitethe absence of KIT expression, were also defined asGIST and included in the study, based on morphol-ogy, CD34 positivity, and lack of expression of S100,desmin, and caldesmon [26]. Overall, therefore, 60cases were included in the study: 39 GISTs of thestomach; 18 GISTs of the small intestine; two of thecolon; and one of the oesophagus. The risk of aggres-sive behaviour was assessed in each case based on themitotic count in 50 consecutive high-power fields andthe tumour size, obtained from the original pathologyreports [8]. Twenty-five cases were scored as low-riskGISTs, three as intermediate-risk GISTs, and 32 ashigh-risk GISTs. In order to facilitate the statistical
comparison between cases with different risk, GISTswith intermediate risk were grouped with the caseswith low risk [27].
Six hepatic, five peritoneal, and one retroperitonealmetastases from seven of the 60 patients were alsoincluded: these were diagnosed between 1 and 6 yearsafter the diagnosis of the primary tumours. The sevenpatients with metastatic disease all belonged to thehigh-risk category.
In addition, 12 leiomyomas and four leiomyosarco-mas of the gastrointestinal tract were investigated forFASN expression.
Frozen specimensFrozen tissue from nine GISTs, two primary andseven metastatic, was obtained from the PathologyDepartment of Brigham and Women’s Hospital, Har-vard Medical School. The Institutional Review Boardapproved the use of these tissues for this study. Six ofthe seven patients from whom the metastatic sampleswere removed showed a partial response and one wasstable on imatinib.
Cell linesAll of the GIST cell lines (GIST882, GIST430,GIST522, GIST62, and GIST48) were obtained fromBrigham and Women’s Hospital, Harvard MedicalSchool. The GIST882 cell line shows a primaryimatinib-sensitive homozygous mutation in KIT exon13 (K642E) [28]. The GIST48 cell line shows a pri-mary imatinib-sensitive homozygous mutation in KITexon 11 (V560D) and a secondary imatinib-resistantheterozygous mutation in KIT exon 17 (D820A).GIST430 is a GIST cell line with a primary imatinib-sensitive heterozygous mutation in KIT exon 11 anda secondary imatinib-resistant heterozygous mutationin KIT exon 13 (V654A). The secondary muta-tions in both GIST48 and GIST430 are fairly com-mon imatinib-resistant mutations in GISTs [24]. BothGIST62 and GIST522 cell lines show a heterozygousKIT exon 11 mutation. In these cell lines, KIT pro-tein has been transcriptionally silenced. All data aboutGIST cell lines, except for GIST882, are unpublishedand are shown in Table 1.
Protein extractionWhole-cell lysates from tumour specimens were pre-pared in 3.5 vol/wt of lysis buffer containing 10%
Table 1. Cell line data
Cell lines Clinical status Primary KIT mutationSecondary KIT
sucrose, 1% Nonidet P-40, 20 mM Tris–HCl (pH 8.0),137 mM NaCl, 10% glycerol, 2 mM EDTA, 10 mM
NaF, 1 mM Na3VO4, 1 mM phenylmethylsulphonylfluoride (PMSF; Sigma), 10 mg/ml soybean trypsininhibitor (Sigma), and 1 mg/ml leupeptin and apro-tinin (Roche Molecular Diagnostics, Mannheim, Ger-many). Tumour samples were cut into small pieces incold lysis buffer on dry ice and tissue was homoge-nized by a tissue tearer for a couple of seconds everytime for 3–5 times on ice and centrifuged at 12 000 gfor 15 min at 4 ◦C. The supernatants were collectedand stored at −80 ◦C. All the GIST cell lines wereharvested from T-75 cell culture flasks and lysed in100 ml of lysis buffer. The protein concentrations weremeasured using the Bio-Rad protein assay kit (Bio-RadLaboratories, Hercules, CA, USA).
Western blot analysis
Western blots were performed as previously described[29] using the following primary antibodies: anti-FASN (clone 23, 1 : 10 000; Transduction Laboratories,Lexington, KY, USA); anti-cyclin A (clone 6E6,1 : 500; Novocastra, Newcastle upon Tyne, UK); anti-p27 (clone 57, 1 : 5000; BD Transduction Laboratories,Lexington, KY, USA); anti-cPARP (clone ASP214;1 : 1000; Cell Signaling); and anti-actin (clone AC-40,1 : 10 000; Sigma, St Louis, MO, USA).
Immunohistochemistry
Six tissue microarrays (TMAs) were constructed fromthe 60 primary cases selected. For each case, onedonor block was identified. Two tissue cores (2 mmin diameter) were punched from two regions of thedonor block representative of the neoplasm and placedon six paraffin blocks to produce TMA blocks uti-lized for the immunohistochemistry (Figure 1A, a). Inorder to orientate the TMA blocks, two liver coreswere placed in the first two columns of the firstrow. For heat-mediated antigen retrieval (AR), 750 Wmicrowave oven (MW) pretreatment was performedfor 30 min in 10 mM citrate buffer (pH 6 or pH 7).All immunostains were performed in an automatedimmunostainer (Dako Autostainer, DakoCytomation,Glostrup, Denmark), using the following primary anti-bodies: c-Kit (1 : 100, no AR, polyclonal; DakoCy-tomation); FASN (1 : 500, pH 6 MW, clone 23; Trans-duction Laboratories); and Ki67 (1 : 100, clone Mib-1, pH 7 MW, DakoCytomation). Standardized 3,3′-diaminobenzidine (DAB) development times allowedaccurate comparison of all samples. Substitution ofphosphate buffered saline (PBS) for the primary anti-body served as a negative control. Liver cores wereused as a positive control for FASN immunostaining[1]. For the immunohistochemical evaluation, both tis-sue cores were examined. No differences in the FASNexpression level of the two representative duplicatetissue cores were observed in any of the cases. For themetastatic cases and the gastrointestinal smooth mus-cle tumours, a section representative of the neoplasm
was analysed. FASN, when expressed, was present inmore than 50% of the tumour cells with cytoplasmicstaining. The staining intensity ranged from weak tostrong. Cases were considered either positive or neg-ative. For Ki67, the percentage of positive nuclei wasobtained by counting 1000 neoplastic cells and both5% and 10% cut-offs were applied [10,11].
FASN siRNA transfection
For the siRNA (Qiagen-Xeragon, Germantown, MD,USA) experiment, 3 × 107 cells were first prepared in10 cm plates. According to the Nucleofection protocol,6 × 106 cells per well were transfected with either 150pmol of anti-FASN duplex oligoribonucleotide siRNA(5′ AAC CCU GAG AUC CCA GCG CUG dTdT3′) or 150 pmol of non-silencing control siRNA (3′dTdT UUG GGA CUC UAG GGU CGC GAC 5′)into 1 ml of pre-warmed (37 ◦C) normal medium insix-well plates, using an electroporation device (Nucle-ofector Device, Amaxa Biosystem) and Nucleofector
solution V (Cell Line Nucleofector Kit V, AmaxaBiosystems). This technology facilitates siRNA trans-port directly into the nucleus, allowing a very hightransfection efficiency to be achieved. Cells were lysedfor western blot analysis 48 h, 72 h or 96 h post-transfection.
Statistical analysis
FASN expression was compared between groups ofGISTs, using the χ2 test for qualitative data. A p valueof 0.05 was considered statistically significant.
Results
FASN is expressed in primary GIST tumours
Thirty-six of 60 primary tumours were positive forFASN (60%) by immunohistochemistry (Figure 1A,b). Gastric, intestinal, and oesophageal GISTs wereall represented in the FASN-expressing group. How-ever, the proportion of FASN-expressing tumours washigher among GISTs from the small intestine (14 of 18cases, 78%) than among gastric GISTs (20 of 39 cases,51%) (p = 0.051). To investigate whether FASN wasa marker of aggressive behaviour, GISTs were sub-divided into low/intermediate-risk versus high-riskgroups [27], according to established criteria [8]. Astatistically significant difference in the proportionof FASN-expressing tumours was found between thetwo groups. Specifically, 12 of 28 cases among thelow/intermediate-risk tumours (42%) and 24 of 32cases among the high-risk tumours (75%) (p < 0.05)expressed the FASN protein (Table 2). FASN expres-sion has been linked to increased proliferative potentialin soft tissue sarcomas [7]. To investigate a possiblerelationship between FASN expression and prolifera-tion, the GIST TMAs were also stained for Ki67. In13 of 60 cases, 11 high risk and two intermediate risk,
J Pathol 2006; 209: 369–375
372 S Rossi et al
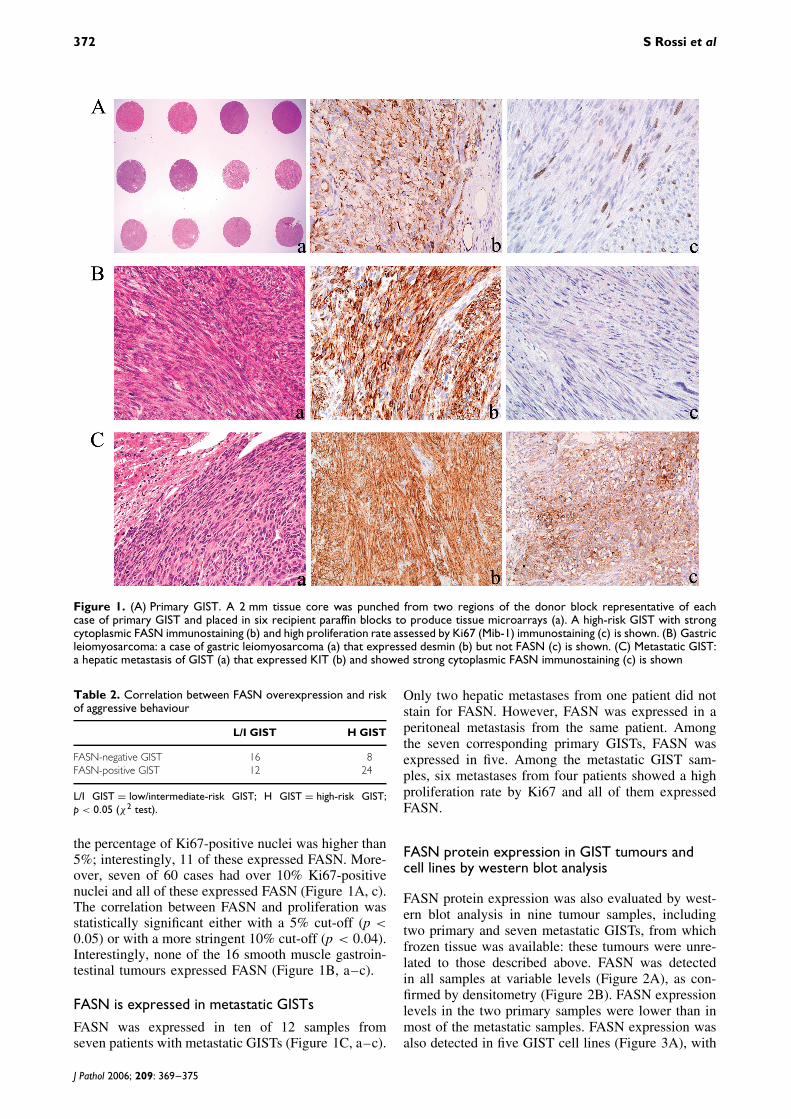
Figure 1. (A) Primary GIST. A 2 mm tissue core was punched from two regions of the donor block representative of eachcase of primary GIST and placed in six recipient paraffin blocks to produce tissue microarrays (a). A high-risk GIST with strongcytoplasmic FASN immunostaining (b) and high proliferation rate assessed by Ki67 (Mib-1) immunostaining (c) is shown. (B) Gastricleiomyosarcoma: a case of gastric leiomyosarcoma (a) that expressed desmin (b) but not FASN (c) is shown. (C) Metastatic GIST:a hepatic metastasis of GIST (a) that expressed KIT (b) and showed strong cytoplasmic FASN immunostaining (c) is shown
Table 2. Correlation between FASN overexpression and riskof aggressive behaviour
the percentage of Ki67-positive nuclei was higher than5%; interestingly, 11 of these expressed FASN. More-over, seven of 60 cases had over 10% Ki67-positivenuclei and all of these expressed FASN (Figure 1A, c).The correlation between FASN and proliferation wasstatistically significant either with a 5% cut-off (p <0.05) or with a more stringent 10% cut-off (p < 0.04).Interestingly, none of the 16 smooth muscle gastroin-testinal tumours expressed FASN (Figure 1B, a–c).
FASN is expressed in metastatic GISTs
FASN was expressed in ten of 12 samples fromseven patients with metastatic GISTs (Figure 1C, a–c).
Only two hepatic metastases from one patient did notstain for FASN. However, FASN was expressed in aperitoneal metastasis from the same patient. Amongthe seven corresponding primary GISTs, FASN wasexpressed in five. Among the metastatic GIST sam-ples, six metastases from four patients showed a highproliferation rate by Ki67 and all of them expressedFASN.
FASN protein expression in GIST tumours andcell lines by western blot analysis
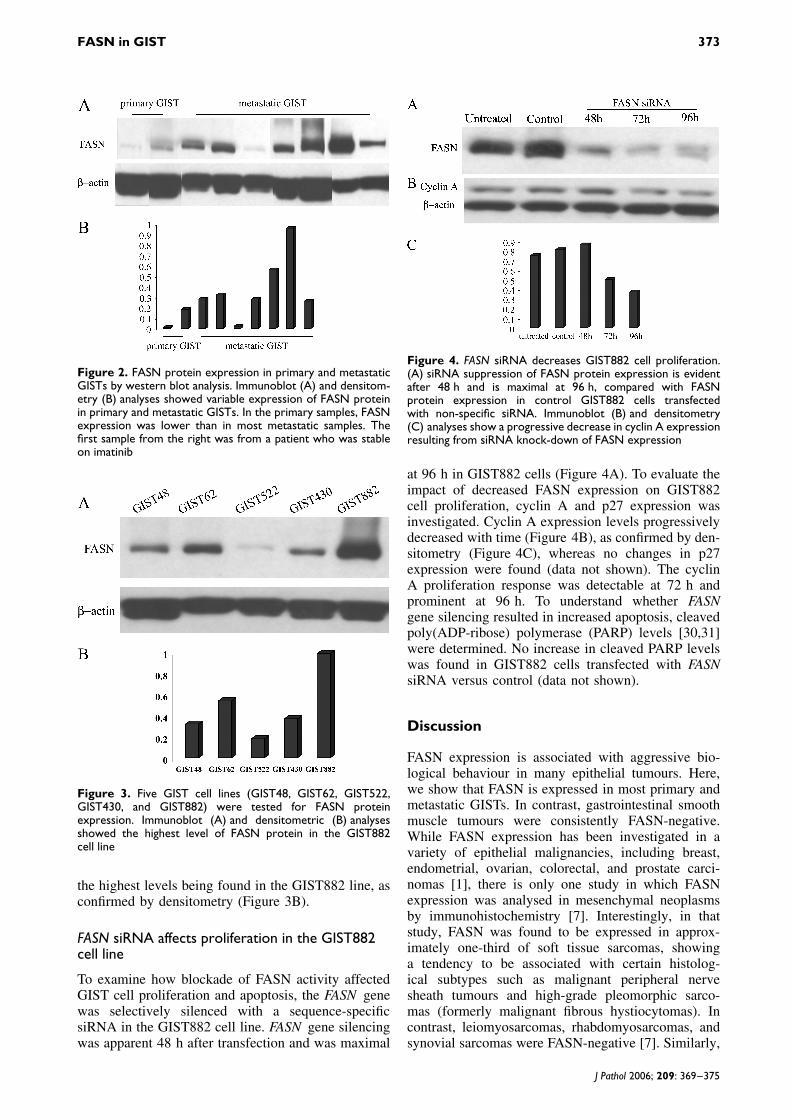
FASN protein expression was also evaluated by west-ern blot analysis in nine tumour samples, includingtwo primary and seven metastatic GISTs, from whichfrozen tissue was available: these tumours were unre-lated to those described above. FASN was detectedin all samples at variable levels (Figure 2A), as con-firmed by densitometry (Figure 2B). FASN expressionlevels in the two primary samples were lower than inmost of the metastatic samples. FASN expression wasalso detected in five GIST cell lines (Figure 3A), with
J Pathol 2006; 209: 369–375
FASN in GIST 373
Figure 2. FASN protein expression in primary and metastaticGISTs by western blot analysis. Immunoblot (A) and densitom-etry (B) analyses showed variable expression of FASN proteinin primary and metastatic GISTs. In the primary samples, FASNexpression was lower than in most metastatic samples. Thefirst sample from the right was from a patient who was stableon imatinib
Figure 3. Five GIST cell lines (GIST48, GIST62, GIST522,GIST430, and GIST882) were tested for FASN proteinexpression. Immunoblot (A) and densitometric (B) analysesshowed the highest level of FASN protein in the GIST882cell line
the highest levels being found in the GIST882 line, asconfirmed by densitometry (Figure 3B).
FASN siRNA affects proliferation in the GIST882cell line
To examine how blockade of FASN activity affectedGIST cell proliferation and apoptosis, the FASN genewas selectively silenced with a sequence-specificsiRNA in the GIST882 cell line. FASN gene silencingwas apparent 48 h after transfection and was maximal
Figure 4. FASN siRNA decreases GIST882 cell proliferation.(A) siRNA suppression of FASN protein expression is evidentafter 48 h and is maximal at 96 h, compared with FASNprotein expression in control GIST882 cells transfectedwith non-specific siRNA. Immunoblot (B) and densitometry(C) analyses show a progressive decrease in cyclin A expressionresulting from siRNA knock-down of FASN expression
at 96 h in GIST882 cells (Figure 4A). To evaluate theimpact of decreased FASN expression on GIST882cell proliferation, cyclin A and p27 expression wasinvestigated. Cyclin A expression levels progressivelydecreased with time (Figure 4B), as confirmed by den-sitometry (Figure 4C), whereas no changes in p27expression were found (data not shown). The cyclinA proliferation response was detectable at 72 h andprominent at 96 h. To understand whether FASNgene silencing resulted in increased apoptosis, cleavedpoly(ADP-ribose) polymerase (PARP) levels [30,31]were determined. No increase in cleaved PARP levelswas found in GIST882 cells transfected with FASNsiRNA versus control (data not shown).
Discussion
FASN expression is associated with aggressive bio-logical behaviour in many epithelial tumours. Here,we show that FASN is expressed in most primary andmetastatic GISTs. In contrast, gastrointestinal smoothmuscle tumours were consistently FASN-negative.While FASN expression has been investigated in avariety of epithelial malignancies, including breast,endometrial, ovarian, colorectal, and prostate carci-nomas [1], there is only one study in which FASNexpression was analysed in mesenchymal neoplasmsby immunohistochemistry [7]. Interestingly, in thatstudy, FASN was found to be expressed in approx-imately one-third of soft tissue sarcomas, showinga tendency to be associated with certain histolog-ical subtypes such as malignant peripheral nervesheath tumours and high-grade pleomorphic sarco-mas (formerly malignant fibrous hystiocytomas). Incontrast, leiomyosarcomas, rhabdomyosarcomas, andsynovial sarcomas were FASN-negative [7]. Similarly,
J Pathol 2006; 209: 369–375
374 S Rossi et al
all gastrointestinal leiomyomas and leiomyosarcomaslacked FASN expression by immunohistochemistry.
Since FASN protein expression has been reportedpreferentially in less differentiated carcinomas [32–34]and in more advanced tumours [35,36], we exploredthe link between FASN expression and risk of aggres-sive behaviour in primary GISTs. FASN expressionwas preferentially detected in 75% of high-risk GISTs,but in only 42% of low/intermediate-risk GISTs, rais-ing the possibility of an association between FASN andtumour aggressiveness in GIST. Furthermore, FASNwas expressed in a higher proportion of GISTs of thesmall intestine, which cluster separately by both geneand protein expression assays [11,37,38] and seem tohave a worse prognosis [39]. However, our correla-tions in this regard did not reach statistical signifi-cance, possibly as a result of the limited number ofcases.
Although more cases need to be examined, FASNexpression appears to be associated with advancedstage, since most metastases expressed high levels ofthe enzyme.
FASN knock-down led to decreased expressionof cyclin A. Cyclin A is a S-phase-specific cyclinrequired for the initiation of DNA replication and is aninformative marker of proliferation in GIST [40]. Inaddition, FASN expression correlated with high pro-liferative rates in both primary and metastatic GISTs,as was previously demonstrated in soft tissue sarco-mas [7]. FASN inhibition has been shown to decreaseDNA synthesis and prevent cells from progressing intoS-phase [3]. It has been proposed that the link betweenFASN and DNA replication may result from de novosynthesis of fatty acids required for the biogenesis ofcellular membranes and second messengers critical inintracellular signal transduction during cell prolifer-ation [41]. More recently, FASN has been found tointerfere with ubiquitin-dependent proteolysis of cell-cycle proteins. In particular, FASN blockade seems tocause dramatic down-regulation of Skp2, a componentof E3 ubiquitin ligase that tags p27 for degradation bythe proteosome in mammary epithelial cell lines [42].In our study, knocking down FASN did not affect p27levels, but did result in decreased proliferation. Thismay be attributed, in part, to Skp2 inhibition. In addi-tion, FASN knock-down did not result in increasedlevels of cleaved PARP, suggesting that FASN doesnot affect apoptosis in GIST, in contrast with whathas been shown in prostate cancer cell lines [1,4,5].
A recent breakthrough in medical oncology has beenthe advent of targeted molecular therapy with thetyrosine kinase inhibitor imatinib. Partial responsesare observed in most patients with advanced GIST,offering a valid therapeutic strategy for a formerlyorphan tumour [23]. However, not all GISTs aresensitive to imatinib: GISTs with KIT or PDGFR-A mutations in the kinase activation loop domain,and GISTs lacking KIT or PDGFR-A mutations, areoften resistant to imatinib [22,25]. Moreover, acquiredresistance, dependent on mis-sense mutations in the
KIT kinase domain, may occur in initially sensitiveGISTs [24,43,44]. In this study, FASN expressionwas demonstrated in all metastatic tumours, includingone from a patient who did not show a partialresponse but was stable on imatinib. In addition, FASNexpression was found in each of five GIST cell lines,including four cell lines that are imatinib-resistant dueto secondary kinase domain mutations or loss of KITexpression. These data raise the intriguing possibilitythat FASN may represent a therapeutic target inGIST patients with primary or acquired resistance toimatinib. FASN inhibitors, such as Cerulenin [1] and,more recently, Orlistat [45], have been demonstratedto halt tumour cell proliferation, induce apoptosis, andinhibit the growth of FASN-positive tumours in nudemice.
In conclusion, we have demonstrated that FASN isexpressed in primary and metastatic GISTs as wellas in GIST cell lines, with preferential expression incases with features of aggressive biological behaviour.FASN gene silencing results in decreased proliferationin the GIST882 cell line. Our data suggest that FASNmay represent a novel biomarker and a potentialtherapeutic target in GISTs that show primary oracquired resistance to imatinib.
Acknowledgements
We thank Mrs Serena Chinellato for her excellent technicalsupport. This work was supported by NIH grants (SPOREP50CA90381, PO1-CA89021) as well as a Prostate CancerFoundation award to ML. ML is a Dana Farber-NovartisInvestigator.
References
1. Kuhajda FP. Fatty-acid synthase and human cancer: new perspec-tives on its role in tumor biology. Nutrition 2000;16:202–208.
2. Zhang L, Joshi AK, Smith S. Cloning, expression, characteriza-tion, and interaction of two components of a human mitochondrialfatty acid synthase. Malonyltransferase and acyl carrier protein. JBiol Chem 2003;278:40 067–40 074.
3. Pizer ES, Chrest FJ DiGiuseppe JA, Han WF. Pharmacologicalinhibitors of mammalian fatty acid synthase suppress DNAreplication and induce apoptosis in tumor cell lines. Cancer Res1998;58:4611–4615.
4. De Schrijver E, Brusselmans K, Heyns W, Verhoeven G, Swin-nen JV. RNA interference-mediated silencing of the fatty acid syn-thase gene attenuates growth and induces morphological changesand apoptosis of LNCaP prostate cancer cells. Cancer Res2003;63:3799–3804.
5. Heiligtag SJ, Bredehorst R, David KA. Key role of mito-chondria in cerulenin-mediated apoptosis. Cell Death Differ2002;9:1017–1025.
6. Agostini M, Silva SD, Zecchin KG, Coletta RD, Jorge J, Loda M,et al. Fatty acid synthase is required for the proliferation of humanoral squamous carcinoma cells. Oral Oncol 2004;40:728–735.
7. Takahiro T, Shinichi K, Toshimitsu S. Expression of fatty acidsynthase as a prognostic indicator in soft tissue sarcomas. ClinCancer Res 2003;9:2204–2212.
8. Blay JY, Bonvalot S, Casali P, Choi H, Debiec-Richter M, DeiTos AP, et al. Consensus meeting for the management ofgastrointestinal stromal tumors. Report of the GIST ConsensusConference of 20–21 March 2004, under the auspices of ESMO.Ann Oncol 2005;16:566–578.
J Pathol 2006; 209: 369–375
FASN in GIST 375
9. Cunningham RE, Abbondanzo SL, Chu WS, Emory TS,Sobin LH, O’Leary TJ. Apoptosis, bcl-2 expression, and p53expression in gastrointestinal stromal/smooth muscle tumors. ApplImmunohistochem Mol Morphol 2001;9:19–23.
10. Panizo-Santos A, Sola I, Vega F, de Alava E, Lozano MD,Idoate MA, et al. Predicting metastatic risk of gastrointestinalstromal tumors: role of cell proliferation and cell cycle regulatoryproteins. Int J Surg Pathol 2000;8:133–144.
11. Haller F, Gunawan B, von Heydebreck A, Schwager S, Schul-ten HJ, Wolf-Salgo J, et al. Prognostic role of E2F1 and membersof CDKN2A network in gastrointestinal stromal tumors. Clin Can-cer Res 2005;11:6589–6597.
12. Sakurai S, Fukayama M, Kaizaki Y, Saito K, Kanazawa K,Kitamura M, et al. Telomerase activity in gastrointestinal stromaltumors. Cancer 1998;83:2060–2066.
13. Gelen T, Elpek GO, Aksoy NH, Ogus M, Keles N. p27 labelingindex and proliferation in gastrointestinal stromal tumors:correlations with clinicopathologic factors and recurrence. Jpn JClin Oncol 2003;33:346–352.
14. Pruneri G, Mazzarol G, Fabris S, Del Curto B, Bertolini F,Neri A, et al. Cyclin D3 immunoreactivity in gastrointestinalstromal tumors is independent of cyclin D3 gene amplificationand is associated with nuclear p27 accumulation. Mod Pathol2003;16:886–892.
15. Schneider-Stock R, Boltze C, Lasota J, Miettinen M, Peters B,Pross M, et al. High prognostic value of p16INK4 alterations ingastrointestinal stromal tumors. J Clin Oncol 2003;21:1688–1697.
16. Schneider-Stock R, Boltze C, Lasota J, Peters B, Corless CL,Ruemmele P, et al. Loss of p16 protein defines high-risk patientswith gastrointestinal stromal tumors: a tissue microarray study.Clin Cancer Res 2005;11:638–645.
17. Montgomery E, Abraham SC, Fisher C, Deasel MR, Amr SS,Sheikh SS, et al. CD44 loss in gastric stromal tumors as aprognostic marker. Am J Surg Pathol 2004;28:168–177.
18. Hirota S, Isozaki K, Moriyama Y, Hashimoto K, Nishida T,Ishiguro S, et al. Gain-of-function mutations of c-kit in humangastrointestinal stromal tumors. Science 1998;279:577–580.
20. Kim TW, Lee H, Kang YK, Choe MS, Ryu MH, ChangHM, et al. Prognostic significance of c-kit mutation inlocalized gastrointestinal stromal tumors. Clin Cancer Res2004;10:3076–3081.
21. Kinoshita K, Hirota S, Isozaki K, Ohashi A, Nishida T, Kita-mura Y, et al. Absence of c-kit gene mutations in gastrointestinalstromal tumours from neurofibromatosis type 1 patients. J Pathol2004;202:80–85.
22. Heinrich MC, Corless CL, Duensing A, Blanke CD, vonMehren M, Joensuu H, et al. PDGFRA activating mutations ingastrointestinal stromal tumors. Science 2003;299:708–710.
23. Verweij J, Casali PG, Zalcberg J, LeCesne A, Reichardt P,Blay JY, et al. Progression-free survival in gastrointestinal stromaltumours with high-dose imatinib: randomised trial. Lancet2004;364:1127–1134.
24. Antonescu CR, Besmer P, Guo T, Arkun K, Hom G, Kory-otowski B, et al. Acquired resistance to imatinib in gastrointestinalstromal tumor occurs through secondary gene mutation. Clin Can-cer Res 2005;11:4182–4190.
25. Debiec-Rychter M, Dumez H, Judson I, Wasag B, Verweij J,Brown M, et al. Use of c-KIT/PDGFRA mutational analysis topredict the clinical response to imatinib in patients with advancedgastrointestinal stromal tumours entered on phase I and II studiesof the EORTC Soft Tissue and Bone Sarcoma Group. Eur J Cancer2004;40:689–695.
26. Medeiros F, Corless CL, Duensing A, Hornick JL, Oliveira AM,Heinrich MC, et al. KIT-negative gastrointestinal stromal tumors:proof of concept and therapeutic implications. Am J Surg Pathol2004;28:8889–8894.
27. Nilsson B, Bumming P, Meis-Kindblom JM, Oden A, Dortok A,Gustavsson B, et al. Gastrointestinal stromal tumors: the inci-dence, prevalence, clinical course and prognostication in thepreimatinib mesylate era — a population-based study in westernSweden. Cancer 2005;103:821–829.
28. Tuveson DA, Willis NA, Jacks T, Griffin JD, Singer S, FletcherCD, et al. STI571 inactivation of the gastrointestinal stromal tumorc-KIT oncoprotein: biological and clinical implications. Oncogene2001;20:5054–5058.
29. Waltregny D, Leav I, Signoretti S, Soung P, Lin D, Merk F,et al. Androgen-driven prostate epithelial cell proliferation anddifferentiation in vivo involve the regulation of p27. MolEndocrinol 2001;15:765–782.
30. Tewari M, Quan LT, O’Rourke K, Dixit V, Snipas SJ, Poirier GG,et al. Yama/CPP32 beta, a mammalian homolog of CED-3, isa CrmA-inhibitable protease that cleaves the death substratepoly(ADP-ribose) polymerase. Cell 1995;81:801–809.
31. Lazebnik YA, Kaufmann SH, Desnoyers S, Poirier GG, Earn-shaw WC. Cleavage of poly(ADP-ribose) polymerase by a pro-teinase with properties like ICE. Nature 1994;371:346–347.
32. Gansler TS, Hardman W 3rd, Hunt DA, Schaffel S, Hennigar RA.Increased expression of fatty acid synthase (OA-519) inovarian neoplasms predicts shorter survival. Hum Pathol1997;28:686–692.
33. Swinnen JV, Roskams T, Joniau S, Van Poppel H, Oyen R,Baert L, et al. Overexpression of fatty acid synthase is an earlyand common event in the development of prostate cancer. Int JCancer 2002;98:19–22.
34. Pizer ES, Lax SF, Kuhajda FP, Pasternack GR, Kurman RJ.Fatty acid synthase expression in endometrial carcinoma:correlation with cell proliferation and hormone receptors. Cancer1998;83:528–537.
35. Epstein JI, Carmichael M, Partin AW. OA-519 (fatty acidsynthase) as an independent predictor of pathologic state inadenocarcinoma of the prostate. Urology 1995;45:81–86.
36. Shurbaji MS, Kalbfleisch JH, Thurmond TS. Immunohistochemi-cal detection of a fatty acid synthase (OA-519) as a predictor ofprogression of prostate cancer. Hum Pathol 1996;27:917–921.
37. Antonescu CR, Viale A, Sarran L, Tschernyavsky SJ, Gonen M,Segal NH, et al. Gene expression in gastrointestinal stromaltumors is distinguished by KIT genotype and anatomic site. ClinCancer Res 2004;10:3282–3290.
38. Nishitani A, Hirota S, Nishida T, Isozaki K, Hashimoto K,Nakagomi N, et al. Differential expression of connexin 43 ingastrointestinal stromal tumours of gastric and small intestinalorigin. J Pathol 2005;206:377–382.
39. Miettinen M, El Rifai W, Sobin HL, Lasota J. Evaluation ofmalignancy and prognosis of gastrointestinal stromal tumors: areview. Hum Pathol 2002;33:478–483.
40. Duensing A, Medeiros F, McConarty B, Joseph NE, Panigray D,Singer S, et al. Mechanisms of oncogenic KIT signal transductionin primary gastrointestinal stromal tumors (GISTs). Oncogene2004;23:3999–4006.
41. Baron A, Migita T, Tang D, Loda M. Fatty acid synthase:a metabolic oncogene in prostate cancer? J Cell Biochem2004;91:47–53.
42. Knowles LM, Axelrod F, Browne CD, Smith JW. A fatty acidsynthase blockade induces tumor cell-cycle arrest by down-regulating Skp2. J Biol Chem 2004;279:30 540–30 545.
43. Tamborini E, Bonadiman L, Greco A, Albertini V, Negri T,Gronchi A, et al. A new mutation in the KIT ATP pocket causesacquired resistance to imatinib in a gastrointestinal stromal tumorpatient. Gastroenterology 2004;127:294–299.
44. Chen LL, Trent JC, Wu EF, Fuller GN, Ramdas L, Zhang W,et al. A missense mutation in KIT kinase domain 1 correlates withimatinib resistance in gastrointestinal stromal tumors. Cancer Res2004;64:5913–5919.
45. Kridel SJ, Axelrod F, Rozenkrantz N, Smith JW. Orlistat is anovel inhibitor of fatty acid synthase with antitumor activity.Cancer Res 2004;64:2070–2075.