Page 1
This article was downloaded by: [117.169.1.120]On: 02 July 2015, At: 08:51Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: 5 HowickPlace, London, SW1P 1WG
Scandinavian Journal of Forest ResearchPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/sfor20
Heart-rot and associated fungi in Alnus glutinosastands in LatviaNatalija Arhipova a b , Talis Gaitnieks b , Janis Donis b , Jan Stenlid a & Rimvydas Vasaitisa
a Department of Forest Mycology and Pathology , Swedish University of AgriculturalSciences , SE-75007 , Uppsala , Swedenb Latvian State Forest Research Institute “Silava” , LV2169 , Salaspils , LatviaAccepted author version posted online: 01 Mar 2012.Published online: 26 Mar 2012.
To cite this article: Natalija Arhipova , Talis Gaitnieks , Janis Donis , Jan Stenlid & Rimvydas Vasaitis (2012) Heart-rotand associated fungi in Alnus glutinosa stands in Latvia, Scandinavian Journal of Forest Research, 27:4, 327-336, DOI:10.1080/02827581.2012.670727
To link to this article: http://dx.doi.org/10.1080/02827581.2012.670727
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose ofthe Content. Any opinions and views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be reliedupon and should be independently verified with primary sources of information. Taylor and Francis shallnot be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and otherliabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Page 2
ORIGINAL ARTICLE
Heart-rot and associated fungi in Alnus glutinosa stands in Latvia
NATALIJA ARHIPOVA1,2, TALIS GAITNIEKS2, JANIS DONIS2, JAN STENLID1 &
RIMVYDAS VASAITIS1
1Department of Forest Mycology and Pathology, Swedish University of Agricultural Sciences, SE-75007 Uppsala, Sweden,
and 2Latvian State Forest Research Institute ‘‘Silava’’, LV2169 Salaspils, Latvia
AbstractThe interest in Alnus glutinosa (L.) Gaertn. as plantation species has increased during last years, but its prospects should beevaluated from the perspective of forest health. The aims of the present study were to: (1) estimate the incidence of stemdecay in Latvian A. glutinosa stands, (2) measure the extent of decay within individual stems and on a stand level and (3)identify decay-causing fungi. In four A. glutinosa stands, 450 trees were randomly sampled with an increment borer and thepresence/absence of decay was recorded. As a result, 112 sound-looking and 338 decayed trees were detected, and acorresponding number of wood samples were collected for fungal isolations. A total of 34 stems with decay symptoms werecut to measure the extent of internal decay. The incidence of decayed stems in studied stands was 75.1% on average. Thelength of the decay column was 7.795.4 m on average, and that of spongy rot was 4.294.5 m on average, implying thatyield losses for fully stocked 80-years-old A. glutinosa stand would comprise 49.2% of the total stand volume, and the lossesfrom spongy rot alone � 30.5%. In total, 1134 isolates representing 68 fungal taxa were obtained. The most common decay-causing fungi were Inonotus radiatus and Armillaria sp.
Keywords: Black alder, Inonotus radiatus, Armillaria, stem decay, wood-inhabiting fungi, yield losses.
Introduction
Currently, stands of black alder (Alnus glutinosa (L.)
Gaertn.) comprise 5.1% (161,200 ha) of the total
forest area of Latvia (Central Statistical Bureau of
Latvia, 2008�2010). In Latvia, A. glutinosa typically
grows on wet peatlands, usually comprising pure
stands, or stands mixed with Alnus incana (L.)
Moench., Betula spp., Populus tremula L. and Picea
abies (L.) Karst. (Kundzins, 1969; Prieditis, 1993).
Specific characteristics of this tree species are frost
and waterlogging tolerance, a strong root system that
penetrates both vertically and horizontally, adapta-
tion to various soil conditions, and ability to fix
nitrogen (McVean, 1953; Wheeler et al., 1986).
Leaves are also nitrogen-rich and, after being shed,
increase nitrogen concentration in soil (Cote &
Camire, 1985; Dawson & Funk, 1981;
Perez-Corona et al., 2006). A. glutinosa grows well
on marshlands, riverbanks and other kinds of wet
sites, and is an excellent pioneer species (Claessens
et al., 2010; Fremstad, 1983; Obidzinski, 2004).
Under suitable conditions it can be as productive as
Fraxinus or Acer, yielding wood of high quality
(Claessens et al., 2010), usable for wide variety of
purposes, e.g. sawn timber, pulp and others
(Claessens et al., 2010; Fennessy, 2004; McVean,
1953).
The combination of rapid early growth with a
coppicing ability makes A. glutinosa suitable for a
short rotation forestry (Wittwer & Immel, 1978;
Wittwer & Stringer, 1985), while the capacity for
pioneering and nitrogen fixation makes this species
suitable for afforestation of former agricultural land
and non-productive sites, as, e.g. reclaimed mining
areas (Chodak & Niklinska, 2010; Kuznetsova et al.,
2010; Pregent & Camire, 1985; Torbert et al., 1985;
Vares et al., 2004; Wittwer & Immel, 1978). The
species has also been used in mixed forest planta-
tions to increase overall wood production (Bohanek
& Groninger, 2005; Chodak & Niklinska, 2010), as
interplanting with A. glutinosa was shown to have
positive effects on growth of adjacent trees of other
Correspondence: Natalija Arhipova, Department of Forest Mycology and Pathology, Uppsala BioCenter, Swedish University of Agricultural Sciences, PO Box
7026, SE-75007 Uppsala, Sweden. E-mail: [email protected]
Scandinavian Journal of Forest Research, 2012; 27: 327�336
(Received 29 August 2011; accepted 22 February 2012)
ISSN 0282-7581 print/ISSN 1651-1891 online # 2012 Taylor & Francis
http://dx.doi.org/10.1080/02827581.2012.670727
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 3
species (Cote & Camire, 1984, 1987; Hansen &
Dawson, 1982; Paschke et al., 1989; Plass, 1977).
Consequently, nowadays A. glutinosa is becoming
increasingly important as a plantation species. Aside
from commercial forestry, this tree species is im-
portant for riparian ecosystems and their biodiver-
sity, providing habitats for specific wetland flora and
fauna and stabilising riverbanks (Brown et al., 1997;
Claessens et al., 2010; Popovska et al., 2008;
Prieditis, 1997).
The potential of different tree species considered
for afforestation needs to be evaluated from many
points of view, and the aspect of forest health is an
important consideration. As in numerous countries
with intense management of boreal � temperate
forests, heart-rot of standing trees is a considerable
problem also in Latvian forestry. Based on observed
average heart-rot incidence, spread of the decay
inside a stem, and applying stand growth models it
was estimated that in fully stocked stands of P. abies
volumes of wood, degraded by the heart-rot
comprise about 20 m3 ha�1 at the age of 40 years,
55�60 m3 ha�1 at the age of 60�100 years and about
90 m3 ha�1 at the age of 120 years, corresponding to
6�16% of a total standing volume (Arhipova et al.,
2011a). Similar situation was also observed in fully
stocked pure stands of A. incana, where volumes of
decayed wood comprised 32 m3 ha�1 at the age
of 45�50 years and 60 m3 ha�1 at the age of
60�65 years, corresponding to about 10% and
20% of all standing volume (Arhipova et al.,
2011b). To date, however, no published data in
this respect are available for A. glutinosa.
Generally, A. glutinosa is regarded a short-living
tree species, starting to die out naturally at about
60 years of age under Central European conditions
(Vyhlıdkova et al., 2005). However, depending on
the region and growth conditions, the species might
reach age of 100�160 years (Claessens et al., 2010).
Some authors noted that A. glutinosa is usually
attacked by a stem rot at the age of 50�70 years,
especially on wet sites (Claessens, 2005; Claessens
et al., 2010; Immler, 2004; Kotar, 2000). Moreover,
there are observations that a large proportion of trees
with a diameter at breast height over 30 cm are
attacked by decay fungi and become susceptible to
stem-breakage (Ilisson et al., 2004), or have stem
cavities (Remm et al., 2006). In his review McVean
(1953) named polypore Inonotus radiatus (Sowerby)
Karst. as the most important cause of heart-rot in
A. glutinosa, which was later supported by the
observations from Central Europe (Schumacher
et al., 2001, Vyhlıdkova et al., 2005). To date,
information on fungal communities in living stems of
A. glutinosa is scarce, comprising only a couple of
records of endophytic fungi (Fisher & Petrini, 1990;
Moricca, 2002). In this context, the aims of the
present study were to: (1) estimate the incidence of
stem decay in Latvian A. glutinosa stands, (2)
measure the extent of decay within individual stems
and on a stand level and (3) identify decay-causing
and other fungi that inhabit living A. glutinosa stems.
Materials and methods
Field work
The fieldwork included: (1) sampling living stems of
A. glutinosa for estimating the frequency of decay
and discoloration and for subsequent fungal isola-
tion, (2) felling and dissection of decayed stems
containing decay and measuring the extent of
decay. Five 51- to 84-year-old A. glutinosa stands
of 0.5�3.6 ha in size were investigated (Table I). The
stands were located in central-eastern Latvia,
Kalsnava forest district (56.6838 N, 25.9678 E). A
total of 450 trees have been sampled at 10�20 cm
height using an increment borer (Table I) and the
presence/absence of stem decay was recorded follow-
ing visual examination of each core. Each tree was
sampled once, extracting 10- to 20-cm-long bore
cores. All 450 wood samples were placed into sterile
plastic tubes and transported to the laboratory, and
full length of the cores was subsequently used for
fungal isolations. In each stand, trees were sampled
by random, always choosing the most adjacent tree
to the one previously sampled. From those sampled
trees, 34 decayed stems (as determined by presence
of decay in the extracted cores) were felled and
dissected. Age, height, stem diameter at breast
Table I. Characteristics of investigated A. glutinosa stands and number of trees cut for heart-rot examination.
Age, years Species composition (%)* Forest site type Sampled trees, no. Decayed stems (%) Cut trees, no.
51 100 A.g. Mercurialiosa** 100 98 �74 80 A.g.�20 A.i. Filipendulosa 100 82 14
76 60 A.i.�30 A.g.�10 P.a. Oxalidosa turf.** 150 53 20
84 80 A. g.�20 A. i. Oxalidosa turf.** 100 78 �Average 75
*A.g. � Alnus glutinosa; A.i. � Alnus incana; P.a. � Picea abies.**Drained (ameliorated).
328 N. Arhipova et al.
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 4
height (d.b.h.) and stump diameter were assessed for
every tree. Age of the felled trees ranged from 74 to
81 years (7793 years), stump diameter � from 15.3
to 43.1 cm (28.297.5 cm), d.b.h. � from 14.1 to
36.3 cm (23.296.1 cm) and height � from 13.2 to
27.3 m (21.893.5 m). Two types of decay were
distinguished: (1) discoloured wood without or with
slight changes in mechanical properties, throughout
the paper referred as ‘‘decay’’ and (2) decomposed
wood squeezable with fingers, throughout the paper
classed as ‘‘spongy rot’’. Columns of the spongy rot
were always situated inside decay columns, being
shorter in length and smaller in diameter. Diameters
at stump level and the total column lengths were
separately measured both for decay and, when
present, for spongy rot.
Isolation and identification of fungi
In the laboratory, the procedure of fungal isolation
from wood samples was performed as in our earlier
studies (Vasiliauskas & Stenlid, 1998; Vasiliauskas
et al., 1996). Briefly, the isolation was done next day
after the sampling. Overnight samples were stored in
refrigerator at 48C. All samples were flame-sterilised,
placed on Hagem agar (Modess, 1941) media and
incubated at 198C in the dark for 1�2 weeks. To
obtain pure cultures, the emerging mycelia were
subcultured in separate Petri dishes, containing
Hagem agar media. After 2�3 weeks of incubation,
all pure cultures were examined under the light
microscope (Leica DM400B) and grouped accord-
ing to morphological features of the fungal myce-
lium. From those, several species and genera
(possessing distinct mycelial morphotypes and re-
peatedly sequenced in our previous studies) were
identified microscopically (Table III).
One to three representatives from the morpho-
types that have not been identified microscopically
were subjected to a molecular identification
(Vasiliauskas et al., 2004, 2005). DNA extraction
and PCR amplification followed established proto-
cols (Karen et al., 1997). After the amplification,
PCR products were purified using Calf Intestine
Alkaline Phosphatase (CIAP) (Fermentas GmbH,
St. Leon-Rot, Germany) and Exonuclease I (Exo I)
(Fermentas GmbH, St. Leon-Rot, Germany) and
sent for Sanger sequencing (Sanger et al., 1977) to
Macrogen Inc. (Seoul, Republic of Korea). For the
ITS region sequencing, primer ITS4 was used for
every specimen. All sequences were aligned and
manually edited using Lasergene software package
SeqMan (version 5.07, DNASTAR, Madison, WI,
USA). BLAST (Basic Local Alignment Search Tool)
searches (Altschul et al., 1997) were performed
using two reference databases � one at the Depart-
ment of Forest Mycology and Pathology, Swedish
University of Agricultural Sciences, and one of the
GenBank (http://www.ncbi.nlm.nih.gov/blast). The
ITS sequence homology was set at 98�100% for
species level and 94�97% for genus level as in our
previous studies (Arhipova et al., 2011a, 2011b;
Bakys et al., 2009a, 2009b, 2011; Vasiliauskas et al.,
2004, 2005). ITS sequence information for selected
isolates was deposited in the GenBank (Table III).
Volume calculations and statistical analyses
Stand characteristics were obtained from the stand
inventory data in the Forest State Register
(Mr. Indulis Brauners, Latvian State Forests Inc.,
personal communication). Calculations of decayed
log volume were based on actual lengths of decay
and spongy rot in the analysed trees (Arhipova et al.,
2011a, 2011b; Ozolins, 2000). Volume calculations
of decayed logs at the stand level were made using
individual stem data in the context of regional forest
yield models and tables (Kuliesis, 1993). The
calculations were accomplished closely following
the procedure used in our previous studies (Arhipova
et al., 2011a, 2011b). Correlation analyses were
made using Microsoft Excel and significance levels
were evaluated according to Liepa (1974). Analyses
of similarity between fungal communities detected in
sound-looking and decayed stems of A. glutinosa, as
well as between the communities inhabiting
Table II. Correlations between tree and decay parameters in 34 dissected stems of A. glutinosa.
Decay column parametersTree parameters
Stump diameter d.b.h. Height
Decay length inside stem 0.671 0.658 0.676
Length of spongy rot within decay column/stem 0.498 0.489 0.482
Decay diameter at stump level 0.652 0.678 0.436*
Diameter of spongy rot within decay column at stump level 0.542 0.569 0.339**
Correlation coefficients not labelled with stars are significant at pB0.01.*Significant at pB0.05**Not significant; p�0.05
Heart-rot of Alnus glutinosa 329
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 5
Table III. Percentages of A. glutinosa stems from which each fungal taxa were isolated.
Living stems
Fungal taxa GenBank accession no.* Sound (n�112) Decayed (n�338)
Basidiomycetes
Armillaria sp. � � 5.9
Bjerkandera adusta (Willd.) Karst. JF340266 0.9 4.1
Chondrostereum purpureum (Pers.) Pouzar JF340265 � 1.2
Cinereomyces lindbladii (Berk.) Julich JF340289 � 0.3
Daedaleopsis confragosa (Bolton) Schrot. JF340288 � 0.3
Flammula alnicola (Fr.) P. Kumm. JF340270 � 2.1
Fomes fomentarius (L.) Fr. JF340284 � 0.9
Fomitopsis pinicola (Sw.) Karst. JF340282 0.9 �Gymnopilus penetrans (Fr.) Murrill JF340274 � 0.3
Inonotus radiatus (Sowerby) Karst. JF340239 0.9 42.6
Mycena galericulata (Scop.) Gray JF340273 � 1.2
Peniophora cinerea (Pers.) Cooke JF340279 0.9 0.6
Phanerochaete sordida (Karst.) Erikss. & Ryvarden JF340276 0.9 �Pholiota aurivella (Batsch) P. Kumm. JF340271 � 1.8
Polyporus sp. M57 JF340264 � 0.3
Unidentified sp. M85 JF340277 � 1.5
All basidiomycetes 4.5 55.3
Ascomycetes/anamorphic fungi
Alternaria alternata (Fr.) Keissl. JF340247 2.7 3.8
Ascocoryne cylichnium (Tul.) Korf JF340286 4.5 28.7
Aspergillus sp. � 2.7 2.4
Aureobasidium pullulans (de Bary) G. Arnaud JF340246 0.9 1.2
Bionectria sp. M75 JF340272 1.8 2.4
Botryotinia fuckeliana (de Bary) Whetzel � � 0.3
Cadophora malorum (Kidd & Beaumont) Gams JF340236 4.5 11.8
Chaetosphaeria myriocarpa (Fr.) Booth JF340253 � 0.3
Chaunopycnis pustulata Bills, Polishook & White JF340250 � 0.6
Cladosporium cladosporioides (Fresen.) de Vries JF340280 1.8 1.2
Coniothyrium fuckelii Sacc. JF340287 0.9 �Cosmospora vilior (Starback) Rossman & Samuels JF340292 0.9 0.3
Cryptosporiopsis diversispora Robak JF340249 � 4.1
Cylindrocarpon sp. M46 JF340259 � 0.3
Daldinia loculata (Lev.) Sacc. JF340281 � 0.3
Diaporthe sp. M116 JF340291 � 0.2
Epicoccum nigrum Link JF340252 1.8 0.3
Eutypella cerviculata (Fr.) Sacc. JF340269 � 0.6
Fimetariella rabenhorstii (Niessl) Lundq. JF340278 2.7 �Gibberella avenacea Cook JF340258 3.6 0.6
Helotiales sp. M17 JF340244 � 0.6
Leptodontium elatius (Mangenot) de Hoog JF340290 0.9 1.2
Leucostoma persoonii (Nitschke) Hohn. JF340255 2.7 0.6
Lewia infectoria (Fuckel) Barr & Simmons JF340283 0.9 0.3
Mariannaea elegans (Corda) Samson JF340240 � 0.6
Melanconis stilbostoma (Fr.) Tul. & Tul. JF340241 2.7 4.1
Nectria cinnabarina (Tode) Fr. JF340285 0.9 �Paraconiothyrium sporulosum (Gams & Domsch) Verkley JF340257 � 0.3
Paraphaeosphaeria michotii (Westend.) Erikss. JF340243 � 0.3
Penicillium sp. � 17.9 34.3
Penicillium spinulosum Thom JF340242 2.7 2.7
Pestalotiopsis besseyi (Guba) Nag Raj JF340248 0.9 �Phialocephala sp. M36 JF340237 1.8 6.2
Phialocephala sp. M49 JF340261 1.8 8.2
Phoma herbarum Westend. JF340263 � 1.5
Phoma sp. M37 JF340254 � 0.9
Plectosphaerella sp. M32 JF340251 0.9 �Sclerotinia sp. M40 JF340256 � 0.3
Scytalidium lignicola Pesante JF340238 0.9 4.1
Spadicoides bina (Corda) Hughes JF340260 � 0.3
Thysanophora penicillioides (Roum.) Kendr. JF340235 2.7 0.3
Trichoderma sp. � 8.0 15.4
330 N. Arhipova et al.
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 6
A. glutinosa and A. incana stems (Arhipova et al.
2011b) were performed by calculating qualitative
Sorensen similarity indices (Magurran, 1988).
Results
As a result of sampling, 112 visually healthy and
338 decayed bore cores were collected. The inci-
dence of decay-infected stems in the investigated
A. glutinosa stands varied between 53 and 98% (75%
on average, Table I). Decay was present in all
34 dissected stems as presumed during the core
sampling, and spongy rot was detected in 29 of the
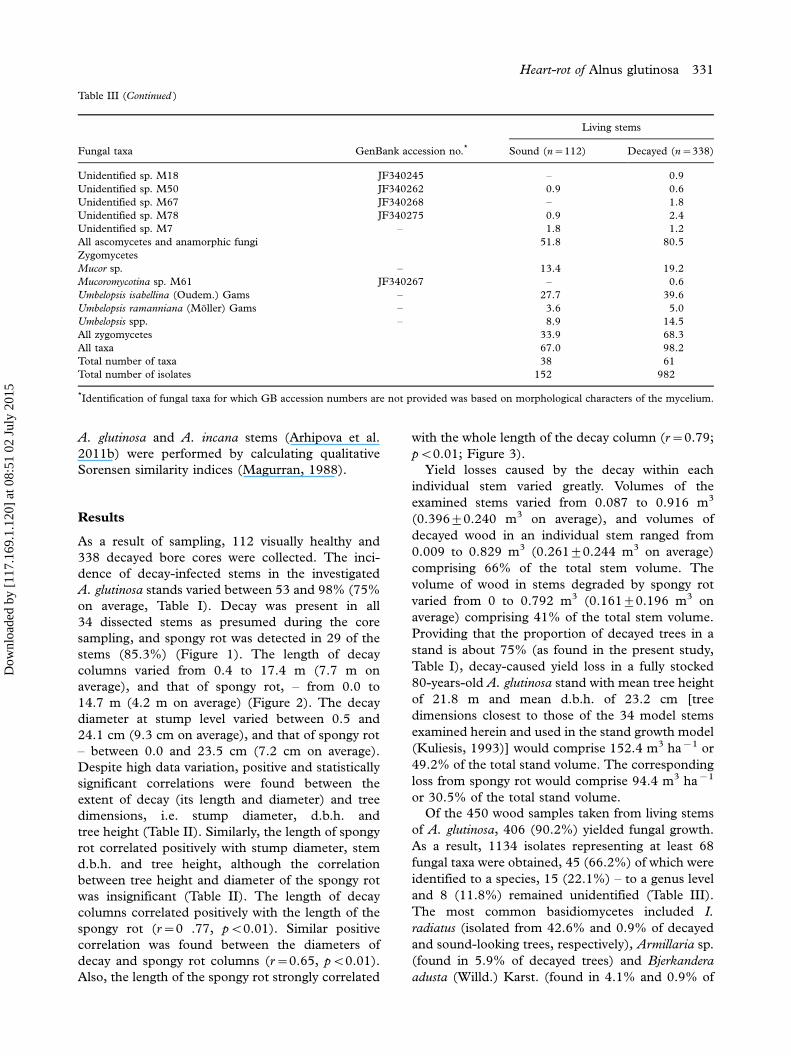
stems (85.3%) (Figure 1). The length of decay
columns varied from 0.4 to 17.4 m (7.7 m on
average), and that of spongy rot, � from 0.0 to
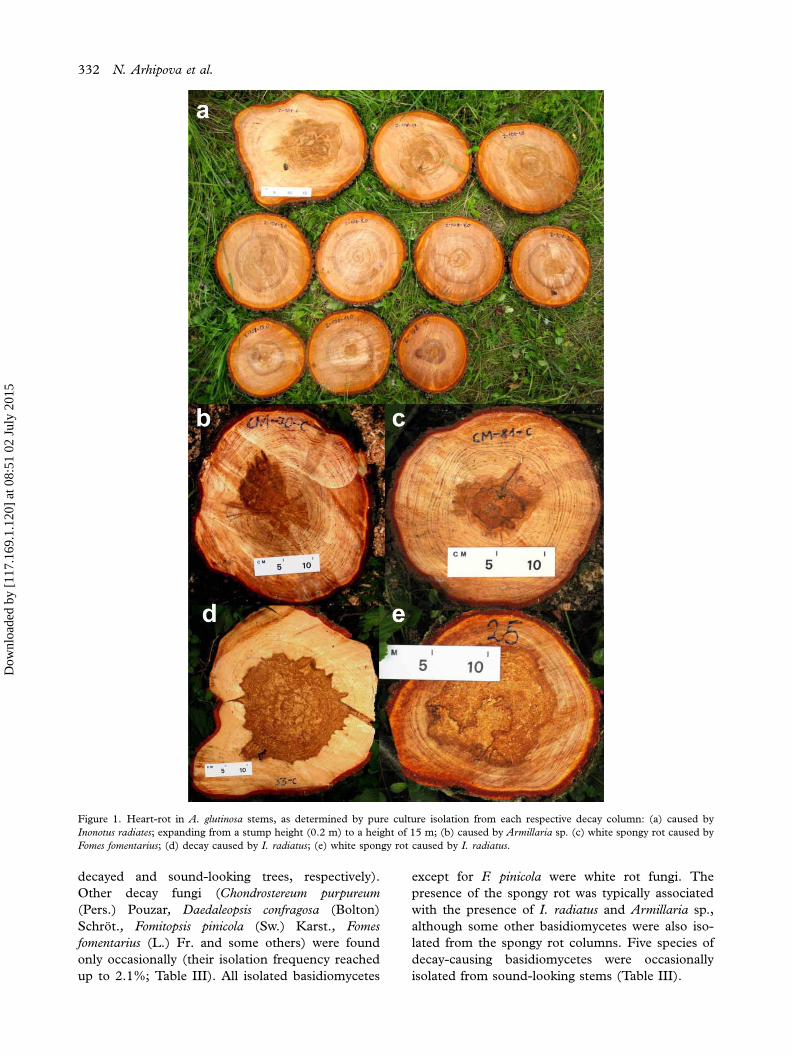
14.7 m (4.2 m on average) (Figure 2). The decay
diameter at stump level varied between 0.5 and
24.1 cm (9.3 cm on average), and that of spongy rot
� between 0.0 and 23.5 cm (7.2 cm on average).
Despite high data variation, positive and statistically
significant correlations were found between the
extent of decay (its length and diameter) and tree
dimensions, i.e. stump diameter, d.b.h. and
tree height (Table II). Similarly, the length of spongy
rot correlated positively with stump diameter, stem
d.b.h. and tree height, although the correlation
between tree height and diameter of the spongy rot
was insignificant (Table II). The length of decay
columns correlated positively with the length of the
spongy rot (r�0 .77, pB0.01). Similar positive
correlation was found between the diameters of
decay and spongy rot columns (r�0.65, pB0.01).
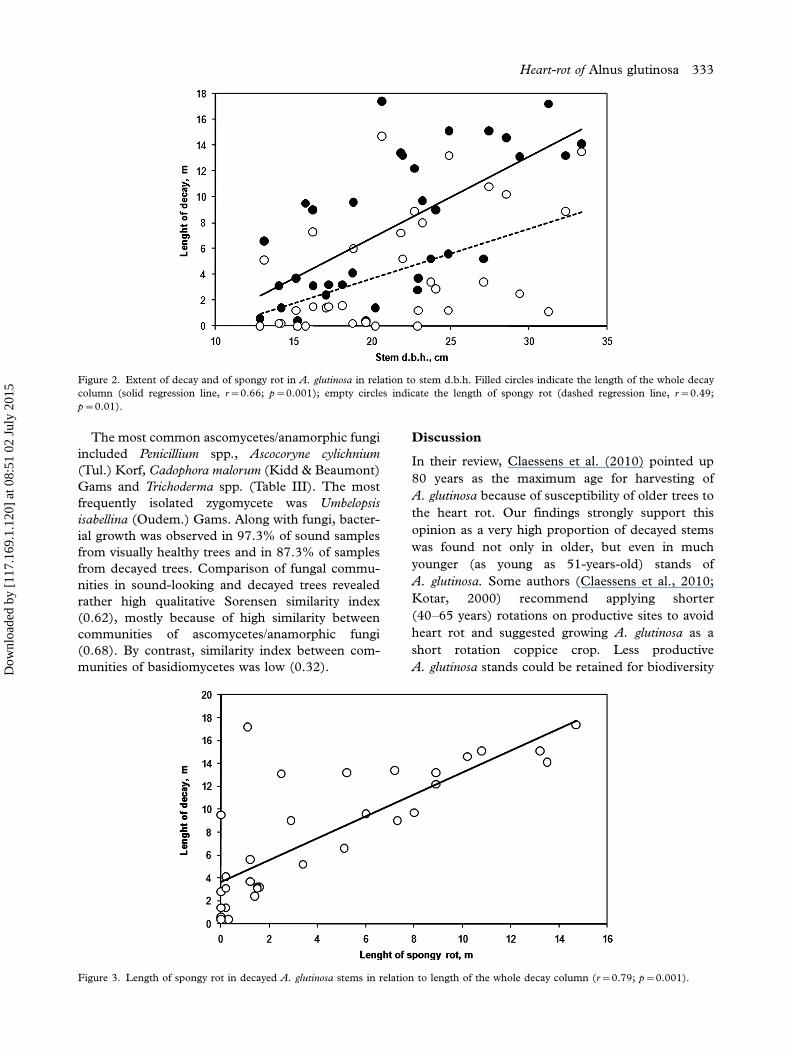
Also, the length of the spongy rot strongly correlated
with the whole length of the decay column (r�0.79;
pB0.01; Figure 3).
Yield losses caused by the decay within each
individual stem varied greatly. Volumes of the
examined stems varied from 0.087 to 0.916 m3
(0.39690.240 m3 on average), and volumes of
decayed wood in an individual stem ranged from
0.009 to 0.829 m3 (0.26190.244 m3 on average)
comprising 66% of the total stem volume. The
volume of wood in stems degraded by spongy rot
varied from 0 to 0.792 m3 (0.16190.196 m3 on
average) comprising 41% of the total stem volume.
Providing that the proportion of decayed trees in a
stand is about 75% (as found in the present study,
Table I), decay-caused yield loss in a fully stocked
80-years-old A. glutinosa stand with mean tree height
of 21.8 m and mean d.b.h. of 23.2 cm [tree
dimensions closest to those of the 34 model stems
examined herein and used in the stand growth model
(Kuliesis, 1993)] would comprise 152.4 m3 ha�1 or
49.2% of the total stand volume. The corresponding
loss from spongy rot would comprise 94.4 m3 ha�1
or 30.5% of the total stand volume.
Of the 450 wood samples taken from living stems
of A. glutinosa, 406 (90.2%) yielded fungal growth.
As a result, 1134 isolates representing at least 68
fungal taxa were obtained, 45 (66.2%) of which were
identified to a species, 15 (22.1%) � to a genus level
and 8 (11.8%) remained unidentified (Table III).
The most common basidiomycetes included I.
radiatus (isolated from 42.6% and 0.9% of decayed
and sound-looking trees, respectively), Armillaria sp.
(found in 5.9% of decayed trees) and Bjerkandera
adusta (Willd.) Karst. (found in 4.1% and 0.9% of
Table III (Continued )
Living stems
Fungal taxa GenBank accession no.* Sound (n�112) Decayed (n�338)
Unidentified sp. M18 JF340245 � 0.9
Unidentified sp. M50 JF340262 0.9 0.6
Unidentified sp. M67 JF340268 � 1.8
Unidentified sp. M78 JF340275 0.9 2.4
Unidentified sp. M7 � 1.8 1.2
All ascomycetes and anamorphic fungi 51.8 80.5
Zygomycetes
Mucor sp. � 13.4 19.2
Mucoromycotina sp. M61 JF340267 � 0.6
Umbelopsis isabellina (Oudem.) Gams � 27.7 39.6
Umbelopsis ramanniana (Moller) Gams � 3.6 5.0
Umbelopsis spp. � 8.9 14.5
All zygomycetes 33.9 68.3
All taxa 67.0 98.2
Total number of taxa 38 61
Total number of isolates 152 982
*Identification of fungal taxa for which GB accession numbers are not provided was based on morphological characters of the mycelium.
Heart-rot of Alnus glutinosa 331
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 7
decayed and sound-looking trees, respectively).
Other decay fungi (Chondrostereum purpureum
(Pers.) Pouzar, Daedaleopsis confragosa (Bolton)
Schrot., Fomitopsis pinicola (Sw.) Karst., Fomes
fomentarius (L.) Fr. and some others) were found
only occasionally (their isolation frequency reached
up to 2.1%; Table III). All isolated basidiomycetes
except for F. pinicola were white rot fungi. The
presence of the spongy rot was typically associated
with the presence of I. radiatus and Armillaria sp.,
although some other basidiomycetes were also iso-
lated from the spongy rot columns. Five species of
decay-causing basidiomycetes were occasionally
isolated from sound-looking stems (Table III).
Figure 1. Heart-rot in A. glutinosa stems, as determined by pure culture isolation from each respective decay column: (a) caused by
Inonotus radiates; expanding from a stump height (0.2 m) to a height of 15 m; (b) caused by Armillaria sp. (c) white spongy rot caused by
Fomes fomentarius; (d) decay caused by I. radiatus; (e) white spongy rot caused by I. radiatus.
332 N. Arhipova et al.
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 8
The most common ascomycetes/anamorphic fungi
included Penicillium spp., Ascocoryne cylichnium
(Tul.) Korf, Cadophora malorum (Kidd & Beaumont)
Gams and Trichoderma spp. (Table III). The most
frequently isolated zygomycete was Umbelopsis
isabellina (Oudem.) Gams. Along with fungi, bacter-
ial growth was observed in 97.3% of sound samples
from visually healthy trees and in 87.3% of samples
from decayed trees. Comparison of fungal commu-
nities in sound-looking and decayed trees revealed
rather high qualitative Sorensen similarity index
(0.62), mostly because of high similarity between
communities of ascomycetes/anamorphic fungi
(0.68). By contrast, similarity index between com-
munities of basidiomycetes was low (0.32).
Discussion
In their review, Claessens et al. (2010) pointed up
80 years as the maximum age for harvesting of
A. glutinosa because of susceptibility of older trees to
the heart rot. Our findings strongly support this
opinion as a very high proportion of decayed stems
was found not only in older, but even in much
younger (as young as 51-years-old) stands of
A. glutinosa. Some authors (Claessens et al., 2010;
Kotar, 2000) recommend applying shorter
(40�65 years) rotations on productive sites to avoid
heart rot and suggested growing A. glutinosa as a
short rotation coppice crop. Less productive
A. glutinosa stands could be retained for biodiversity
Figure 3. Length of spongy rot in decayed A. glutinosa stems in relation to length of the whole decay column (r�0.79; p�0.001).
Figure 2. Extent of decay and of spongy rot in A. glutinosa in relation to stem d.b.h. Filled circles indicate the length of the whole decay
column (solid regression line, r�0.66; p�0.001); empty circles indicate the length of spongy rot (dashed regression line, r�0.49;
p�0.01).
Heart-rot of Alnus glutinosa 333
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 9
conservation (Claessens et al., 2010; Hall & Bunce,
2011).
The present study shows that a large proportion
(53�98%) of trees in the investigated mature and
over-mature (51- to 84-years-old) A. glutinosa stands
are damaged by heart rot. As the recommended age
of the final felling of A. glutinosa stands is 71 years
(corresponding clause nr. 9, Forest Law of Latvia),
our results suggest that this late felling may un-
avoidably lead to serious yield losses due to stem
decay. Generally lower heart-rot incidence, compris-
ing 21.6% of stems on average, was observed in
Latvian 53- to 65-year-old stands of A. incana,
although rot incidence increased with stand age,
and in some stands the proportion of decayed
A. incana trees was over 50% (Arhipova et al.,
2011b), indicating that susceptibility to heart-rot of
both tree species is rather similar. As in the present
study, the size of decay column in A. incana trees was
larger in larger stems, which might be determined by
the higher volume of heartwood in larger trees that is
favoured by the heart rot-causing fungi. Moreover,
the extent of spongy rot in relation to whole decay
column observed in A. glutinosa (r�0.79; Figure 3)
was higher than that in A. incana (r�0.48; Arhipova
et al., 2011b), what might also be explained by the
fact that older A. glutinosa stems were examined. It is
also worth noticing that the heart-rot frequency
was related to stand species composition: pure
A. glutinosa stand had the highest heart-rot incidence
(98%), whereas stand composed of three species
showed the lowest incidence (53%), and the rest two
of the investigated stands that were composed of two
species were in-between (78�82%) (Table I). How-
ever, the present study included too few (4) stands to
draw any definite conclusion on this observation,
and more comprehensive investigations are needed
to clarify eventual relationships between stand spe-
cies composition and heart-rot incidence in
A. glutinosa.
The most common decay fungi in living stems of
A. glutinosa were I. radiatus and Armillaria sp., while
other basidiomycetes were only occasionally isolated.
As those fungi were mostly found in columns of the
spongy rot, both species, and in particular I. radiatus,
should be regarded as the most important heart-rot
agents of A. glutinosa. Results of this work therefore
fully support findings by Schumacher et al. (2001)
and Vyhlıdkova et al. (2005) which investigated
heart rot of black alder in Germany and Czech
Republic. Moreover, I. radiatus was pointed out as
the most important heart-rot agent of A. glutinosa
already in early British review (McVean, 1953). In
nature, I. radiatus is most commonly observed on
dead Alnus wood: following tree death it grows as a
saprotroph and continues to decompose snags, logs
and stumps, producing on them abundant sporo-
carps (Ryvarden & Gilbertson, 1993). The present
study for the first time provided the information on
wood yield loss caused by I. radiatus in stands of
A. glutinosa, showing the fungus to be an important
forest pathogen capable to degrade up to 50% of a
total wood harvest. Nevertheless, little is known
on infection biology and infection pathways of
I. radiatus during attacks to living trees, thus the
subject deserves further investigations.
To date, the present work represents the most
comprehensive study of mycobiota inhabiting living
A. glutinosa stems so far (Table III). Interestingly,
species richness in living (sound-looking and de-
cayed) stems of A. glutinosa (0.15 taxa per tree) was
much lower than that in A. incana (0.75 taxa per
tree), detected during our previous study (Arhipova
et al., 2011b), implying that ecological conditions for
fungal colonisation of and growth in living stems of
those two related tree species markedly differ. This
hypothesis is furthermore supported by the fact that
qualitatively different fungal communities were
found in stems of A. glutinosa and A. incana
(Sorensen similarity index 0.39; datasets: Table III
vs. Arhipova et al., 2011b), despite trees being
equally exposed to fungal infections while growing
in mixed stands (Table I). The dominant decay fungi
were also different: C. purpureum and Phlebia
tremellosa (Schrad.) Nakasone & Burds. were domi-
nant in stems of A. incana (Arhipova et al., 2011b),
while in A. glutinosa those were I. radiatus and
Armillaria sp. In sound-looking stems of living
A. glutinosa, it was also surprising to find a number
of basidiomycetes capable of causing active wood
decay, namely F. pinicola, B. adusta, I. radiatus,
Peniophora cinerea (Pers.) Cooke and Phanerochaete
sordida (Karst.) Erikss. & Ryvarden. This was also
the case in our previous study (Arhipova et al.,
2011b) where 12 taxa of decay-causing basidiomy-
cetes (including B. adusta and P. cinerea) have been
isolated from visually healthy stems of A. incana.
Some earlier studies had also reported an occasional
occurrence of decay fungi in healthy xylem of beech
and aspen stems (Baum et al., 2003; Chapela,
1989), but more recent investigations employing
molecular methods had demonstrated numerous
wood decay fungi latently present in sapwood of
wide range of angiosperm trees (Parfitt et al., 2010).
Pathways of infection, ecological roles and factors
triggering pathogenicity of those fungi in healthy tree
stems are not clear and are subject of current and
future research.
In conclusion, results of the present study show
that prevailing majority of stems in mature and over-
mature A. glutinosa stands of Latvia are invaded by
extensive heart-rot, and that wood harvest loss due
334 N. Arhipova et al.
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 10
to decay might comprise over 50% of a total stand
volume, degrading its most valuable part (butt log).
The pathogen, mainly responsible for the loss is
wood-decay fungus I. radiatus, biology, ecology and
infection pathways of which are to date poorly
studied, particularly in relation to living trees. Yet,
as only relatively old stands were investigated,
information on susceptibility of younger stands to
the heart-rot is still lacking. Taking into account that
in the region A. glutinosa stands may achieve
productivity of up to 150�190 m3 ha�1 already at
the age of 20 years (Vares et al., 2004), such
knowledge could provide important implications
for management of this tree species, including
recommendations for rotation length in, e.g. energy
plantations.
Acknowledgements
The study was financially supported by the Latvian
State Research Program ‘‘Substantiation of decid-
uous tree cultivation and rational utilization: new
products and technologies 2005�2009’’, the Latvian
State Forest Research Institute ‘‘Silava’’, the
Swedish Energy Agency (STEM) and the Faculty
of Natural Resources and Agricultural Sciences
(research program TEMA) and the Swedish
University of Agricultural Sciences. We thank
Dr. Anna Hopkins for language revision.
References
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J.,
Zhang, Z., Miller, W. et al. (1997). Gapped BLAST and
PSI-BLAST: A new generation of protein database search
programs. Nucleic Acids Research, 25, 3389�3402.
Arhipova, N., Gaitnieks, T., Donis, J., Stenlid, J. & Vasaitis, R.
(2011a). Butt rot incidence, causal fungi, and related yield
loss in Picea abies stands of Latvia. Canadian Journal of Forest
Research, 41, 2337�2345.
Arhipova, N., Gaitnieks, T., Donis, J., Stenlid, J. & Vasaitis, R.
(2011b). Decay, yield loss and associated fungi in stands of
grey alder (Alnus incana) in Latvia. Forestry, 84, 337�348.
Bakys, R., Vasaitis, R., Barklund, P., Thomsen, I. M. & Stenlid, J.
(2009a). Occurrence and pathogenicity of fungi in necrotic
and non-symptomatic shoots of declining common ash
(Fraxinus excelsior) in Sweden. European Journal of Forest
Research, 128, 51�60.
Bakys, R., Vasaitis, R., Barklund, P., Thomsen, I. M. & Stenlid, J.
(2009b). Investigations concerning the role of Chalara
fraxinea in declining Fraxinus excelsior. Plant Pathology, 58,
284�292.
Bakys, R., Vasiliauskas, A., Ihrmark, K., Stenlid, J., Menkis, A. &
Vasaitis, R. (2011). Root rot, associated fungi and their
impact on health condition of declining Fraxinus excelsior
stands in Lithuania. Scandinavian Journal of Forest Research,
26, 128�135.
Baum, S., Sieber, T. N., Schwarze, F. W. M. R. & Fink, S. (2003).
Latent infection of Fomes fomentarius in the xylem of
European beech (Fagus sylvatica). Mycological Progress, 2,
141�148.
Bohanek, J. R. & Groninger, J. W. (2005). Productivity of
European black alder (Alnus glutinosa) interplanted with
black walnut (Juglans nigra) in Illinois, USA. Agroforestry
Systems, 64, 99�106.
Brown, A. G., Harper, D. & Peterken, G. F. (1997). European
floodplain forests: Structure, functioning and management.
Global Ecology and Biogeography Letters, 6, 169�178.
Central statistical Bureau of Latvia (2008�2010). Tree species
in forests of Latvia. Retrieved, from http://www.csb.gov.lv/
en/statistikas-temas/natural-resources-key-indicators-30500.
html (cited 13.06.2011).
Chapela, I. H. (1989). Fungi in healthy stems and branches of
American beech and aspen: A comparative study. New
Phytologist, 113, 65�75.
Chodak, M. & Niklinska, M. (2010). The effect of different tree
species on the chemical and microbial properties of re-
claimed mine soils. Biology and Fertility of Soils, 46, 555�566.
Claessens, H. (2005). L’aulne glutineux. Ses stations et sa sylvicul-
ture. [The Black Alder it’s stations and forestry]. Gembloux,
Belgique: Foret Wallonne asbl, 189 pp. (In French).
Claessens, H., Oosterbaan, A., Savill, P. & Rondeux, J. (2010). A
review of the characteristics of black alder (Alnus glutinosa
(L.) Gaertn.) and their implications for silvicultural
practices. Forestry, 83, 163�175.
Cote, B. & Camire, C. (1984). Growth, nitrogen accumulation,
and symbiotic dinitrogen fixation in pure and mixed plant-
ings of hybrid poplar and black alder. Plant and Soil, 78,
209�220.
Cote, B. & Camire, C. (1985). Nitrogen cycling in dense
plantings of hybrid poplar and black alder. Plant and Soil,
87, 195�208.
Cote, B. & Camire, C. (1987). Tree growth and nutrient cycling
in dense plantings of hybrid poplar and black alder.
Canadian Journal of Forest Research, 17, 516�523.
Dawson, J. O. & Funk, D. T. (1981). Notes: Seasonal change in
foliar nitrogen concentration of Alnus glutinosa. Forest
Science, 27, 239�243.
Fennessy, J. (2004). Common alder (Alnus glutinosa) as a forest tree
in Ireland. Dublin, Ireland: Coford Connects, Reproductive
Material No. 8, 4 pp.
Fisher, P. J. & Petrini, O. (1990). A comparative study of fungal
endophytes in xylem and bark of Alnus species in England
and Switzerland. Mycological Research, 94, 313�319.
Fremstad, E. (1983). Role of black alder (Alnus glutinosa) in
vegetation dynamics in West Norway. Nordic Journal of
Botany, 3, 393�410.
Hall, S. & Bunce, B. (2011). Mature trees as keystone structures
in Holarctic ecosystems. A quantitative species comparison
in a northern English park. Plant Ecology & Diversity, 4,
243�250.
Hansen, E. A. & Dawson, J. O. (1982). Effect of Alnus glutinosa on
hybrid Populus height growth in a short-rotation intensively
cultured plantation. Forest Science, 28, 49�59.
Ilisson, T., Metslaid, M. & Jogiste, K. (2004). Storm as a
disturbance factor in forest ecosystems: A case study of
Tudu and Halliku. Translation of the Faculty of Forestry,
Estonian Agricultural University, 37, 104�107.
Immler, T. (2004). Waldbauliches Konzept zur Pflege der
Schwarzerle. [Forest planning concept for the management
of the black alder]. In: Bayerische Landesanstalt fur Wald
und Forstwirtschaft, Beitrage zur Schartzerle. LWF Wissen,
42, 27�30.
Karen, O., Hogberg, N., Dahlberg, A., Jonsson, L. &
Nylund, J. E. (1997). Inter- and intraspecific variation in
the ITS region of rDNA of ectomycorrhizal fungi in
Fennoscandia as detected by endonuclease analysis. New
Phytologist, 136, 313�325.
Heart-rot of Alnus glutinosa 335
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015
Page 11
Kotar, M. (2000). Influence of age and stem diameter on yield in
a forest. In: I. Potocnik (Ed.), Nova znanja v gozdarstvu �prispevek visokego solstva: zbornik referatov studijskih dni,
pp. 169�190, Slovenia: Kranjska Gora [In Slovenian with
English summary].
Kuliesis, A. (1993). Forest yield models and tables in Lithuania, 383
pp. Kaunas, Lithuania: LMS.
Kundzins, A. (1969). Investigations on genus Alnus Gaertn. in
Latvian SSR, Doctoral Thesis, Latvian Academy of Agri-
culture, Jelgava, Latvia, 50 p (in Russian).
Kuznetsova, T., Lukjanova, A., Mandre, M. & Lohmus, K.
(2010). Aboveground biomass and nutrient accumulation
dynamics in young black alder, silver birch and Scots pine
plantations on reclaimed oil shale mining areas in Estonia.
Forest Ecology and Management, 262, 56�64.
Liepa, I. (1974). Biometrija. [Biometrics], 332 pp. Riga, Latvia:
Zvaigzne (In Latvian).
Magurran, A. E. (1988). Ecological diversity and its measurement,
179 pp. Princeton, NJ: Princeton University Press.
McVean, D. N. (1953). Alnus glutinosa (L.) Gaertn. Journal of
Ecology, 41, 447�466.
Modess, O. (1941). Zur Kenntnis der Mykorrhizabildner von
Kiefer und Fichte [Notes on mycorrhization of pine
and spruce]. Ph.D. Dissertation, Almqvist & Wiksells
Boktryckeri A.B, Uppsala, Sweden.
Moricca, S. (2002). Phomopsis alnea, the cause of dieback of black
alder in Italy. Plant Pathology, 51, 755�764.
Obidzinski, A. (2004). Black alder (Alnus glutinosa Gaertn.) as a
pioneer species in regeneration of fresh oak-linden-
hornbeam forest (Tilio-Carpinetum Typicum Traczyk 1962)
in Bialowieza forest (East Poland). Polish Journal of Ecology,
52, 533�551.
Ozolins, R. (2000). Forest stand assortment structure analysis
using mathematical modelling. Metsanduslikud uurimused.
(Forestry studies, Tartu), 37, 33�47.
Parfitt, D., Hunt, J., Dockrell, D., Rogers, H. J. & Boddy, L.
(2010). Do all trees carry the seeds of their own destruction?
PCR reveals numerous wood decay fungi latently present in
sapwood of wide range of angiosperm trees. Fungal Ecology,
3, 338�346.
Paschke, M. W., Dawson, J. O. & David, M. B. (1989). Soil
nitrogen mineralization of Juglans nigra interplanted with
actinorhyzal Eleagnus umbellata or Alnus glutinosa. Plant and
Soil, 118, 33�42.
Perez-Corona, E. M., Perez Hernandez, C. M. & Bermudez de
Castro, F. (2006). Decomposition of alder, ash and poplar
litter in a Mediterranean riverine area. Communications in
Plant Sciences and Plant Analysis, 37, 1111�1125.
Plass, W. T. (1977). Growth and survival of hardwoods and pine
interplanted with European alder. Forest Service, U.S. Depart-
ment of Agriculture Northeastern Forest Experiment Sta-
tion, Broomall, PA, USDA Forest Service Research Paper
NE-376, 10 p.
Popovska, C., Ivanovski, D., & Jovanovski, M. (2008). Disturbed
river corridors and protection measures. Balwois 27, 31 May,
Ohrid, Republic of Macedonia, pp. 1�7.
Pregent, G. & Camire, C. (1985). Biomass production by alders
on four abandoned agricultural soils in Quebec. Plant and
Soil, 87, 185�193.
Prieditis, N. (1993). Black alder swamps on forested peatlands in
Latvia. Folia Geobotanica & Phytotaxonomica, 28, 261�277.
Prieditis, N. (1997). Alnus glutinosa � Dominated wetland of the
Baltic Region: Community structure, syntaxonomy and
conservation. Plant Ecology, 129, 49�94.
Remm, J., Lohmus, A. & Remm, K. (2006). Tree cavities in
riverine forests: What determines their occurrence and use
by hole-nesting passerines? Forest Ecology and Management,
221, 167�277.
Ryvarden, L. & Gilbertson, R. L. (1993). European polypores 1.
Synopsis fungorum, 6, 1�387.
Sanger, F., Nicklen, S. & Coulson, A. R. (1977). DNA sequen-
cing with chain-terminating inhibitors. Proceedings of the
National Academy of Sciences of the United States of America,
74, 5463�5467.
Schumacher, J., Heydeck, P. & Roloff, A. (2001). Lignicole
Pilze an Schwarz-Erle (Alnus glutinosa [L.] Gaertn.) �welche Arten sind bedeutsame Faeuleerreger? [Lignicolous
fungi on common alder (Alnus glutinosa [L.] Gaertn.) �Which of the species of wood-decay fungi are crucial?]
Forstwissenschaftliches Centralblatt � Tharandter Forstliches
Jahrbuch, 120, 8�17.
Torbert, J. L., Burger, J. A., Lien, J. N. & Schoenholtz, S. H.
(1985). Results of a tree species trial on a recontoured
surface mine in Southwestern Virginia. Southern Journal of
Applied Forestry, 9, 150�153.
Vares, A., Lohmus, K., Truu, M., Truu, J., Tullus, H. & Kanal, A.
(2004). Productivity of black alder (Alnus glutinosa (L.)
Gaertn.) plantations on reclaimed oil-shale mining detritus
and mineral soils in relation to rhizosphaere conditions. Oil
Shale, 21, 43�58.
Vasiliauskas, R. & Stenlid, J. (1998). Discoloration following bark
stripping wounds on Fraxinus excelsior. European Journal of
Forest Pathology, 28, 383�390.
Vasiliauskas, R., Stenlid, J. & Johansson, M. (1996). Fungi in bark
peeling wounds of Picea abies in central Sweden. European
Journal of Forest Pathology, 26, 285�296.
Vasiliauskas, R., Lygis, V., Thor, M. & Stenlid, J. (2004). Impact
of biological (Rotstop) and chemical (urea) treatments on
fungal community structure in freshly cut Picea abies stumps.
Biological Control, 31, 405�413.
Vasiliauskas, R., Lygis, V., Larsson, K.-H. & Stenlid, J. (2005).
Airborne fungal colonisation of coarse woody debris in
north-temperate Picea abies forest: Impact of season and
local spatial scale. Mycological Research, 109, 487�496.
Vyhlıdkova, I., Palovcıkova, D., Rybnıcek, M., Cermak, P. &
Jankovsky, L. (2005). Some aspects of alder decline along
the Luznice River. Journal of Forest Science, 51, 381�391.
Wheeler, C. T., Hooker, J. E., Crowe, A. & Berrie, A. M. M.
(1986). The improvement and utilization in forestry of
nitrogen fixation by actinorhizal plants with special reference
to Alnus in Scotland. Plant and Soil, 90, 393�406.
Wittwer, R. F. & Immel, M. J. (1978). A comparison of five tree
species for intensive fiber production. Forest Ecology and
Management, 1, 249�254.
Wittwer, R. F. & Stringer, J. W. (1985). Biomass production and
nutrient accumulation in seedling and coppice hardwood
plantations. Forest Ecology and Management, 13, 223�233.
336 N. Arhipova et al.
Dow
nloa
ded
by [
117.
169.
1.12
0] a
t 08:
51 0
2 Ju
ly 2
015