Identification and Expression of Multidrug Transporters Responsiblefor Fluconazole Resistance in Candida dubliniensis
GARY P. MORAN,1 DOMINIQUE SANGLARD,2 SAMANTHA M. DONNELLY,1
DIARMUID B. SHANLEY,1 DEREK J. SULLIVAN,1
AND DAVID C. COLEMAN1*
Department of Oral Surgery, Oral Medicine and Pathology, School of Dental Science and DublinDental Hospital, Trinity College, University of Dublin, Dublin 2, Republic of Ireland,1
and Institut de Microbiologie, Centre Hospitalier Universitaire Vaudois,1011 Lausanne, Switzerland2
Received 18 March 1998/Returned for modification 10 April 1998/Accepted 4 May 1998
Candida dubliniensis is a recently described Candida species associated with oral candidosis in human im-munodeficiency virus (HIV)-infected and AIDS patients, from whom fluconazole-resistant clinical isolates havebeen previously recovered. Furthermore, derivatives exhibiting a stable fluconazole-resistant phenotype havebeen readily generated in vitro from fluconazole-susceptible isolates following exposure to the drug. In thisstudy, fluconazole-resistant isolates accumulated up to 80% less [3H]fluconazole than susceptible isolates andalso exhibited reduced susceptibility to the metabolic inhibitors 4-nitroquinoline-N-oxide and methotrexate.These findings suggested that C. dubliniensis may encode multidrug transporters similar to those encoded bythe C. albicans MDR1, CDR1, and CDR2 genes (CaMDR1, CaCDR1, and CaCDR2, respectively). A C. dubliniensishomolog of CaMDR1, termed CdMDR1, was cloned; its nucleotide sequence was found to be 92% identical tothe corresponding CaMDR1 sequence, while the predicted CdMDR1 protein was found to be 96% identical tothe corresponding CaMDR1 protein. By PCR, C. dubliniensis was also found to encode homologs of CDR1 andCDR2, termed CdCDR1 and CdCDR2, respectively. Expression of CdMDR1 in a fluconazole-susceptible Dpdr5null mutant of Saccharomyces cerevisiae conferred a fluconazole-resistant phenotype and resulted in a 75%decrease in accumulation of [3H]fluconazole. Northern analysis of fluconazole-susceptible and -resistant iso-lates of C. dubliniensis revealed that fluconazole resistance was associated with increased expression of CdMDR1mRNA. In contrast, most studies showed that overexpression of CaCDR1 was associated with fluconazole re-sistance in C. albicans. Increased levels of the CdMdr1p protein were also detected in fluconazole-resistantisolates. Similar results were obtained with fluconazole-resistant derivatives of C. dubliniensis generated invitro, some of which also exhibited increased levels of CdCDR1 mRNA and CdCdr1p protein. These results dem-onstrate that C. dubliniensis encodes multidrug transporters which mediate fluconazole resistance in clinical isolatesand which can be rapidly mobilized, at least in vitro, on exposure to fluconazole.
The triazole antifungal drug fluconazole is commonly usedto treat oral candidosis and since its introduction has provedeffective in the treatment of oral yeast infections. However,recent studies have reported an increasing incidence of resis-tance to this compound among clinical isolates of Candidaalbicans from human immunodeficiency virus (HIV)-infectedand AIDS patients (14, 15, 25, 31). Furthermore, some evi-dence suggests that since the introduction of fluconazole, theincidence of infections caused by non-C. albicans species ofCandida, including C. glabrata and C. krusei, which are inher-ently less susceptible to fluconazole, has increased (14, 20, 22,38, 42).
Candida dubliniensis is a recently described Candida speciesassociated with oral candidosis in HIV-infected and AIDSpatients, especially in those with recurrent infections. C. dub-liniensis is phylogenetically closely related to C. albicans andhas recently been shown to have a worldwide distribution (4,18, 34–36). In a recent study of Irish subjects, C. dubliniensiswas recovered from the oral cavities of 27% of HIV-infectedindividuals and 32% of AIDS patients presenting with symp-toms of oral candidosis (6). The majority of C. dubliniensis
clinical isolates tested to date are susceptible to fluconazole(MIC range, 0.125 to 1.0 mg/ml) and to other commonly usedantifungal drugs, including ketoconazole, itraconazole, andamphotericin B (9, 17). Based on a limited study, Moran et al.(19) reported the occurrence of fluconazole resistance in 20%of oral isolates (MIC range, 8 to 32 mg/ml) of C. dubliniensisrecovered from AIDS patients who had been treated previ-ously with fluconazole. Furthermore, sequential exposure offluconazole-susceptible clinical isolates of C. dubliniensis toincreasing concentrations of fluconazole in agar medium re-sulted in the recovery of derivatives which expressed a stablefluconazole-resistant phenotype (MIC range, 16 to 64 mg/ml).It has been suggested that the ability of C. dubliniensis torapidly develop resistance to fluconazole may contribute to itsability to successfully colonize the oral cavities of HIV-infectedindividuals who are receiving long-term therapy with this com-pound (19). Furthermore, this may, at least in part, explain theapparent recent emergence of this organism.
Several studies have demonstrated the importance of spe-cific multidrug transporters in the development of fluconazoleresistance in C. albicans. Sanglard et al. (30, 31) have shownthat three C. albicans proteins, namely the ATP-binding cas-sette (ABC) transporters Cdr1p and Cdr2p, encoded by theCDR1 and CDR2 genes, respectively, and the major facilitatorprotein Mdr1p (also known as Benp), encoded by the MDR1gene, play important roles in reducing the intracellular flucon-
* Corresponding author. Mailing address: Dental School Office,School of Dental Science, Trinity College, University of Dublin, Dub-lin 2, Republic of Ireland. Phone: 353 1 6081814. Fax: 353 1 6799294.E-mail: [email protected].
azole content of fluconazole-resistant C. albicans isolates by aprocess of active drug efflux. White (39) has also demonstratedthe importance of these proteins in fluconazole-resistant C. al-bicans. In addition, Loffler et al. (17), Sanglard et al. (29), andWhite (40) have characterized mutations in the cytochromeP-450 lanosterol 14a-demethylase enzyme, the intracellulartarget of fluconazole. Some of these mutations have beenshown to reduce this protein’s affinity for fluconazole in resis-tant clinical isolates of C. albicans (29, 40).
The objectives of the present study were to investigate themechanism(s) of fluconazole resistance in C. dubliniensis clin-ical isolates and in in vitro-generated fluconazole-resistant de-rivatives. Homologs of the C. albicans CDR1, CDR2, and MDR1genes were identified in C. dubliniensis. (For maximum clarity,the C. albicans and C. dubliniensis genes, as well as their prod-ucts, will be given the prefixes Ca and Cd, respectively.) Ex-pression of these genes was examined in fluconazole-suscepti-ble and -resistant clinical isolates of C. dubliniensis in order tocharacterize their role in fluconazole resistance in this organ-ism.
MATERIALS AND METHODS
C. dubliniensis clinical isolates, in vitro-generated derivatives, and cultureconditions. C. dubliniensis isolates were routinely cultured on potato dextroseagar (Oxoid) medium, pH 5.6, at 37°C. For liquid culture, isolates were grown inyeast extract-peptone-dextrose (YPD) broth at 37°C in an orbital incubator(Gallencamp, Leicester, United Kingdom) at 200 rpm. Many of the C. dublini-ensis clinical isolates and in vitro-generated fluconazole-resistant derivatives usedin this study (Table 1) were previously described by Moran et al. (19). Also
included in the study was the oral isolate CD72 and a series of nine in vitro-generated derivatives of the clinical C. dubliniensis isolate CD57, termed CD57C,CD57D, CD57E, CD57F, CD57G, CD57H, CD57I, CD57J, and CD57K (Table1). C. dubliniensis CD72 was isolated from an oral swab specimen taken from themid-dorsum of the tongue of an Irish AIDS patient without clinical symptomsindicative of oral candidosis who had previously been treated with fluconazolefor oral candidosis. This specimen yielded 61 yeast colonies on potato dextroseagar, consisting of 52 colonies of C. dubliniensis and 9 colonies of Saccharomycescerevisiae, which were identified as previously described (19). The derivativeseries CD57C to CD57K was generated in vitro as described by Moran et al. (19)and traces the origin of the fluconazole-resistant derivative CD57K, which wasisolated from a plate containing 50 mg of fluconazole/ml. Initially, a fluconazole-susceptible colony of the C. dubliniensis clinical isolate CD57 (MIC, 0.5 mg/ml)was cultured on YPD agar medium containing 0.5 mg of fluconazole/ml for 48 hat 37°C. Subsequently, this colony was further subcultured on fresh YPD agarmedium containing 0.5 mg of fluconazole/ml under the same conditions. Thisorganism was then successively subcultured as described above, twice in eachcase (first [1°] and second [2°] subcultures), on YPD agar medium containing 1,5, 10, or 25 mg of fluconazole/ml. The organism was finally subcultured on YPDmedium containing 50 mg of fluconazole/ml, from which the derivative termedCD57K was recovered. Colonies were selected from the plates containing flu-conazole (at the indicated concentrations) following incubation to obtain theseries of derivatives termed CD57C to CD57K (Table 1) as follows: CD57C (1°;0.5 mg/ml), CD57D (1°; 1.0 mg/ml), CD57E (1°; 5.0 mg/ml), CD57F (2°; 5.0mg/ml), CD57G (1°; 10.0 mg/ml), CD57H (2°; 10.0 mg/ml), CD57I (1°; 25.0mg/ml), CD57J (2°; 25.0 mg/ml), and CD57K (1°; 50.0 mg/ml). Series derivativeswhich exhibited an increase in the fluconazole MIC relative to the that forparental isolate CD57 (i.e., derivatives CD57F to CD57K), as determined bybroth microdilution in RPMI–2% (wt/vol) glucose, were subcultured at least 10times on fluconazole-free medium at 37°C for 48 h to determine if the flucon-azole resistance phenotype was stable.
Escherichia coli and S. cerevisiae strains, culture media, and growth conditions.E. coli DH5a was used as the host strain for the phagemid pBluescript II KS(2)(Stratagene, La Jolla, Calif.) and its recombinant derivatives and was maintainedon Luria-Bertani (LB) agar (27) containing ampicillin at 100 mg/ml. For liquid
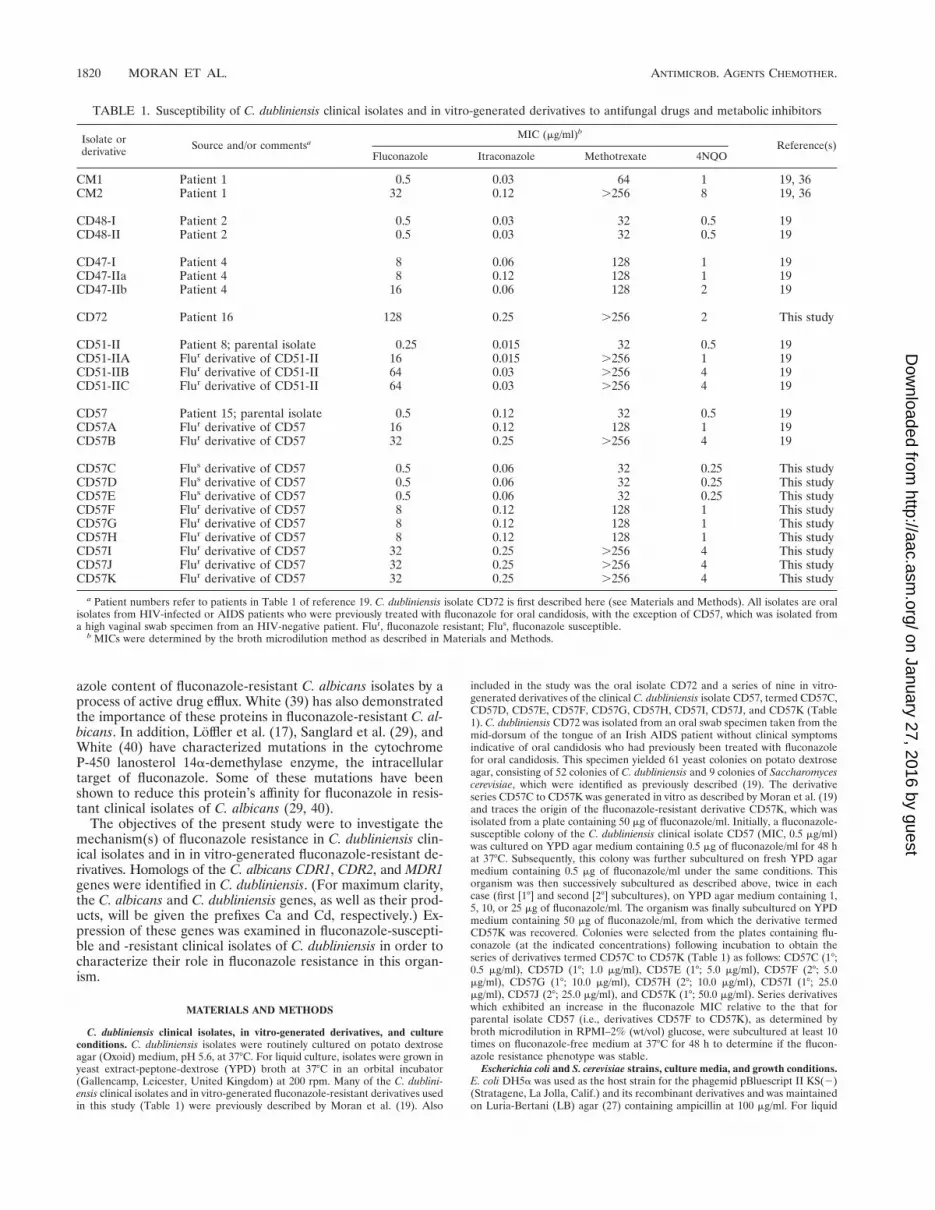
TABLE 1. Susceptibility of C. dubliniensis clinical isolates and in vitro-generated derivatives to antifungal drugs and metabolic inhibitors
CD57C Flus derivative of CD57 0.5 0.06 32 0.25 This studyCD57D Flus derivative of CD57 0.5 0.06 32 0.25 This studyCD57E Flus derivative of CD57 0.5 0.06 32 0.25 This studyCD57F Flur derivative of CD57 8 0.12 128 1 This studyCD57G Flur derivative of CD57 8 0.12 128 1 This studyCD57H Flur derivative of CD57 8 0.12 128 1 This studyCD57I Flur derivative of CD57 32 0.25 .256 4 This studyCD57J Flur derivative of CD57 32 0.25 .256 4 This studyCD57K Flur derivative of CD57 32 0.25 .256 4 This study
a Patient numbers refer to patients in Table 1 of reference 19. C. dubliniensis isolate CD72 is first described here (see Materials and Methods). All isolates are oralisolates from HIV-infected or AIDS patients who were previously treated with fluconazole for oral candidosis, with the exception of CD57, which was isolated froma high vaginal swab specimen from an HIV-negative patient. Flur, fluconazole resistant; Flus, fluconazole susceptible.
b MICs were determined by the broth microdilution method as described in Materials and Methods.
culture, E. coli DH5a harboring recombinant plasmids was routinely grown inLB broth containing 100 mg of ampicillin/ml in an orbital incubator (Gallen-camp) at 37°C and 200 rpm. Transformation of E. coli DH5a and the identifi-cation of transformant derivatives harboring recombinant plasmids were carriedout by standard protocols (27). E. coli LE392 and its lysogenic derivativeLE392-P2 were used for the propagation of the bacteriophage lambda cloningvector EMBL3 and its recombinant derivatives, respectively, on LB mediumsupplemented with 10 mM MgSO4 and 0.2% (wt/vol) maltose as described bySambrook et al. (27).
S. cerevisiae YKKB-13 (MATa ura3-52 lys2-801amber ade2-101ochre trp1-D63his3-D200 leu2-1 Dpdr5::TRP1) (31) was routinely cultured on yeast nitrogen base(YNB; Difco) medium supplemented with uracil, lysine, adenine, tryptophan,and histidine (each at 50 mg/ml) and containing 2% (wt/vol) glucose (YNB–2%glucose) at 30°C. YKKB-13 is defective for the ABC transporter Sts1p (alsoknown as Pdr5p) and is hypersusceptible to azole drugs (31).
Chemicals, enzymes, radioisotopes, and oligonucleotides. Analytical-grade ormolecular biology-grade chemicals were purchased from Sigma, BDH (Poole,Dorset, United Kingdom), or Boehringer Mannheim (Lewes, East Sussex,United Kingdom). Enzymes were purchased from the Promega Corporation(Madison, Wis.) or Boehringer Mannheim and used according to the manufac-turer’s instructions. [a-32P]dATP (3,000 Ci mmol21; 110 TBq mmol21) waspurchased from Amersham International Plc. (Little Chalfont, Buckingham-shire, United Kingdom). Fluconazole powder was a gift from Pfizer Pharmaceu-ticals (Sandwich, Kent, United Kingdom), itraconazole powder was a gift fromJanssen Pharmaceuticals (Cork, Republic of Ireland), and amphotericin B was agift from E. R. Squibb (Swords, Republic of Ireland). All other metabolicinhibitors used for the susceptibility testing of C. dubliniensis isolates and deriv-atives and S. cerevisiae YKKB-13 and its derivatives were purchased from Sigma,with the exception of terbinafine (Sandoz Pharma, Surrey, United Kingdom) andamorolfine chloride (Hoffman La Roche, Basel, Switzerland). Custom-synthe-sized oligonucleotides were purchased from Genosys Biotechnologies (Europe)Ltd. (Pampisford, Cambridgeshire, United Kingdom.)
Susceptibility testing procedures. Fluconazole and itraconazole susceptibilitytesting of C. dubliniensis clinical isolates and their derivatives was carried out in96-well microdilution plates (Corning-Costar) in RPMI 1640 medium (Sigma)supplemented with 2% (wt/vol) glucose (RPMI–2% glucose [26]) as described byMoran et al. (19). Amphotericin B susceptibility tests were carried out by themethod of Rex et al. (24). Some C. dubliniensis isolates and derivatives weretested for susceptibility to metabolic inhibitors (concentration ranges are shownin parentheses) as follows: benomyl (0.5 to 256 mg/ml), 4-nitroquinoline-N-oxide(4NQO; 0.03 to 16 mg/ml), methotrexate (0.5 to 256 mg/ml), and cycloheximide(0.5 to 256 mg/ml) were tested in 96-well plates, with RPMI–2% glucose as thegrowth medium, essentially as described for the fluconazole susceptibility testing.Azole drug susceptibility testing of S. cerevisiae YKKB-13 and its derivatives wascarried out by the broth microdilution method at 30°C in 96-well plates withYNB–2% glucose as the growth medium. The susceptibility of S. cerevisiae toother metabolic inhibitors was assayed by the method of Sanglard et al. on solidmedia by incorporating the inhibitors into 15 ml of YPD agar medium containedin 90-mm-diameter petri dishes to achieve the desired drug concentration (30).Yeast inocula for inhibitor susceptibility testing were prepared by harvesting cellsby centrifugation from cultures grown for 18 h with shaking (180 rpm) at 30°C inYNB–2% glucose medium and resuspending them in 0.9% (wt/vol) NaCl at a celldensity of 2 3 107 CFU/ml. Serial dilutions (1/10) of the suspensions were madein 0.9% (wt/vol) NaCl, and 5-ml volumes of each dilution were spotted ontoinhibitor-containing agar plates which were subsequently incubated at 30°C for24 h. The susceptibility of recombinant-plasmid-harboring S. cerevisiae YKKB-13to each inhibitor was determined based on the highest dilution of the culturewhich could grow in the presence of the inhibitor as described previously bySanglard et al. (30).
Accumulation of [3H]fluconazole in C. dubliniensis isolates. Accumulation of[3H]fluconazole (Amersham) in C. dubliniensis and S. cerevisiae was assessed bythe method of Sanglard et al. (31). All experiments were repeated on twoseparate occasions.
DNA isolation and Southern hybridization analysis. Total genomic DNA ofC. dubliniensis clinical isolates and derivatives was prepared from cells grown for18 h in YPD broth cultures, as described by Gallagher et al. (8). Large-scale andsmall-scale E. coli plasmid DNA preparations were as described by Sambrook etal. (27). Restriction endonuclease-digested DNA was transferred to Magna-Graph nylon membranes (MSI, Westboro, Mass.) as described by Sullivan et al.(33), and hybridization reactions were carried out under high-stringency condi-tions with DNA probes labelled with [a-32P]dATP by random primer labelling ina rotary hybridization oven (Hybaid, Middlesex, United Kingdom) as describedby Sullivan et al. (33).
Yeast chromosomes were prepared as described by Vazquez et al. (37) andseparated in 1.3% (wt/vol) agarose gels by using the CHEF-Mapper system(Bio-Rad, Hercules, Calif.) as described by Sullivan et al. (36). Chromosome-sized DNA was transferred to nylon membranes for hybridization analysis bystandard Southern blotting techniques (33).
Construction of a C. dubliniensis genomic DNA library. High-molecular-masstotal cellular DNA from C. dubliniensis CD36 was isolated as described byBennett et al. (3) and was used for the construction of a lambda EMBL3 library.Sau3A-generated partial-digest products of C. dubliniensis CD36 DNA of greaterthan 10 kb in size were ligated with BamHI-generated prepared lambda bacte-riophage replacement vector EMBL3 arms (Promega) and then packaged invitro, using preprepared phage heads and tails (Promega), according to themanufacturer’s instructions. Following packaging, recombinant phage particleswere propagated on the E. coli P2 lysogenic strain LE392-P2 as described bySambrook et al. (27). A recombinant phage library containing 2 3 105 PFU wasobtained.
Recombinant phages were propagated on E. coli LE392 to yield ;600 to 700PFU per plate for 10 90-mm-diameter petri dishes and were transferred from theplaques onto nitrocellulose membrane filters (Schleicher and Schuell, Dassel,Germany) by overlaying the plaques with the filters. These were then screened byplaque hybridization (27), using as a probe the a-32P-labelled C. albicans MDR1gene contained on a 2.9-kb HindIII-BamHI fragment excised from plasmidp2002 (7). The genomic DNA of a recombinant EMBL3 phage, termed fCD1,which hybridized strongly with the probe was purified as described by Sambrooket al. (27). The cloned DNA insert of fCD1 was mapped with restriction endo-nucleases, and specific fragments were subcloned into pBluescript by conven-tional methods (27).
DNA sequencing. DNA sequencing was performed by the dideoxy chain ter-mination method of Sanger et al. (28), using an Applied Biosystems model 370Aautomated DNA sequencer. Sequencing reactions were carried out with anApplied Biosystems Prism dye terminator cycle sequencing reaction kit. Searchesof the EMBL and GenBank databases for nucleotide and amino acid sequencesimilarities were performed with the BLAST family of computer programs (2).
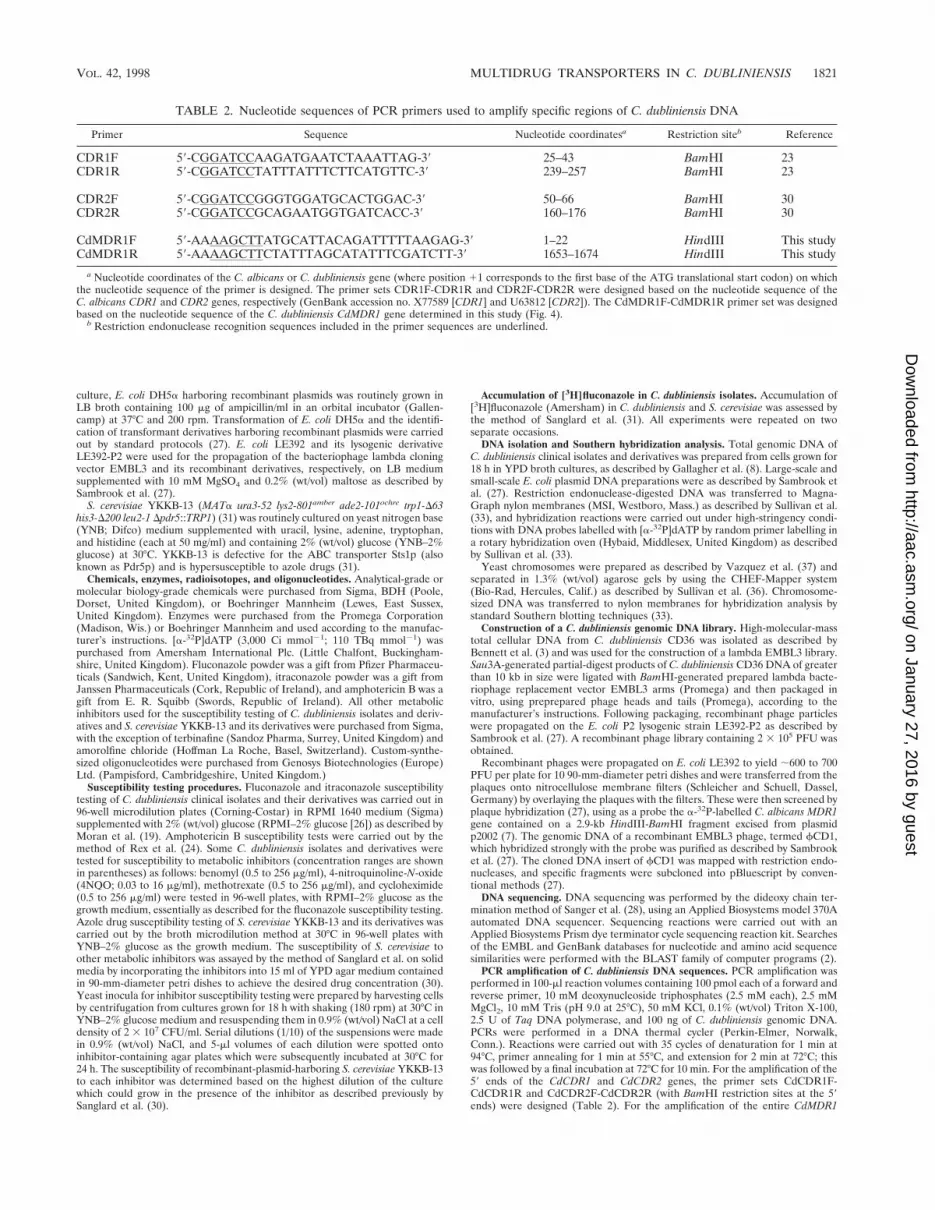
PCR amplification of C. dubliniensis DNA sequences. PCR amplification wasperformed in 100-ml reaction volumes containing 100 pmol each of a forward andreverse primer, 10 mM deoxynucleoside triphosphates (2.5 mM each), 2.5 mMMgCl2, 10 mM Tris (pH 9.0 at 25°C), 50 mM KCl, 0.1% (wt/vol) Triton X-100,2.5 U of Taq DNA polymerase, and 100 ng of C. dubliniensis genomic DNA.PCRs were performed in a DNA thermal cycler (Perkin-Elmer, Norwalk,Conn.). Reactions were carried out with 35 cycles of denaturation for 1 min at94°C, primer annealing for 1 min at 55°C, and extension for 2 min at 72°C; thiswas followed by a final incubation at 72°C for 10 min. For the amplification of the59 ends of the CdCDR1 and CdCDR2 genes, the primer sets CdCDR1F-CdCDR1R and CdCDR2F-CdCDR2R (with BamHI restriction sites at the 59ends) were designed (Table 2). For the amplification of the entire CdMDR1
TABLE 2. Nucleotide sequences of PCR primers used to amplify specific regions of C. dubliniensis DNA
Primer Sequence Nucleotide coordinatesa Restriction siteb Reference
CdMDR1F 59-AAAAGCTTATGCATTACAGATTTTTAAGAG-39 1–22 HindIII This studyCdMDR1R 59-AAAAGCTTCTATTTAGCATATTTCGATCTT-39 1653–1674 HindIII This study
a Nucleotide coordinates of the C. albicans or C. dubliniensis gene (where position 11 corresponds to the first base of the ATG translational start codon) on whichthe nucleotide sequence of the primer is designed. The primer sets CDR1F-CDR1R and CDR2F-CDR2R were designed based on the nucleotide sequence of theC. albicans CDR1 and CDR2 genes, respectively (GenBank accession no. X77589 [CDR1] and U63812 [CDR2]). The CdMDR1F-CdMDR1R primer set was designedbased on the nucleotide sequence of the C. dubliniensis CdMDR1 gene determined in this study (Fig. 4).
b Restriction endonuclease recognition sequences included in the primer sequences are underlined.
VOL. 42, 1998 MULTIDRUG TRANSPORTERS IN C. DUBLINIENSIS 1821
gene, the primer set CdMDR1F-CdMDR1R (with HindIII restriction sites at the59 ends) (Table 2) was designed based on the nucleotide sequence of the C. dub-liniensis CD36 gene determined in this study and the amplification reaction wascarried out with a high-fidelity thermostable DNA polymerase (VentR; NewEngland BioLabs, Beverly, Mass.). PCR products were cloned into pBluescript IIKS(2) by conventional methods (27). Following digestion with HindIII, theCdMDR1 amplimer was cloned into the HindIII-cleaved S. cerevisiae expressionvector plasmid pAAH5 to create recombinant plasmid pGM3. Plasmid pAAH5contains a unique HindIII restriction site downstream of the S. cerevisiae ADC1promoter, which allows constitutive expression of cloned sequences from thispromoter region in S. cerevisiae (31). Both pAAH5 and pGM3 were used totransform S. cerevisiae YKKB-13 by standard protocols (31).
RNA extraction and Northern analysis. RNA was extracted from C. dublini-ensis cultures grown to mid-exponential phase (optical density at 600 nm, 0.6) in50-ml volumes of YPD broth at 37°C with shaking at 200 rpm in an orbitalincubator (Gallenkamp). Extractions were carried out by the glass bead disrup-tion method described by Hube et al. (13). To remove contaminating DNA, 2volumes of 6 M LiCl was added to each RNA sample, and after incubation of thesolutions at 220°C for at least 2 h, they were centrifuged at 11,600 3 g. PelletedRNA was resuspended in diethyl pyrocarbonate-treated water (;150 ml), and20-mg quantities, in 5- to 10-ml volumes, were used for electrophoresis in 1.2%(wt/vol) agarose gels containing 6% (vol/vol) formaldehyde as described by Hubeet al. (13). RNA was transferred onto MagnaGraph nylon membranes by capil-lary transfer in 203 SSC buffer (3 M NaCl, 0.3 M trisodium citrate). The RNAwas fixed by baking the membranes for 30 min at 80°C followed by UV cross-linking in a Bio-Rad UV cross-linker. Hybridization reactions were performedwith a dextran sulfate-containing hybridization solution at 42°C by the method ofSanglard et al. (30). The membranes were then exposed to BioMax MS film(Eastman Kodak Company, Rochester, N.Y.) for 24 to 72 h. All membranes werehybridized with a probe homologous to the C. albicans TEF3 gene, consisting ofa 0.7-kb EcoRI-PstI fragment from plasmid pDC1, as described by Hube et al.(13). Relative levels of mRNA expression were measured by using an imagingdensitometer (Bio-Rad model GS-670) to scan the hybridization signal intensityon autoradiograms, with the signal intensity of TEF3 mRNA being employed asa loading control.
Extraction and Western blotting of proteins from C. dubliniensis. Crude pro-tein extracts were prepared from C. dubliniensis isolates and derivatives grown inYNB broth containing 2% (wt/vol) glucose to mid-exponential phase. A 2-mlvolume of each culture was harvested by centrifugation at 5,000 3 g for 5 min,and each portion of cells was resuspended in 1 ml of sterile distilled H2O. Cellswere lysed by the addition of 150 ml of 1.85 M NaOH–7.5% (vol/vol) b-mercap-toethanol and then incubated on ice for 10 min. Protein was precipitated by theaddition of 150 ml of ice-cold 50% (vol/vol) trichloroacetic acid and incubationon ice for 10 min; this was followed by centrifugation at 10,000 3 g for 5 min at4°C. Each sample was resuspended in 100 ml of sample buffer (40 mM Tris-HCl,8 M urea, 5% [wt/vol] sodium dodecyl sulfate, 0.1 mM EDTA, 1% [vol/vol]b-mercaptoethanol, and 0.1 mg of bromophenol blue per ml), incubated for 15min at 37°C, and then centrifuged as described above to remove cell debris. Forelectrophoresis, 10-ml volumes (each containing approximately 20 mg of protein)of each sample were loaded on sodium dodecyl sulfate–10% (wt/vol) polyacryl-amide gels and electrophoresed in a Mini-PROTEAN II electrophoresis cell(Bio-Rad). Following electrophoresis, proteins were transferred to nitrocellulosemembranes by Western blotting, using the Bio-Rad Mini Trans-blot electro-phoretic transfer cell in accordance with the manufacturer’s instructions. Immu-nodetection of proteins was carried out with polyclonal rabbit sera raised againstpurified glutathione S-transferase-fused N-terminal regions of the C. albicansMdr1p, Cdr1p, and Cdr2p proteins (28a). Antibody-protein complexes weredetected with horseradish peroxidase-conjugated anti-rabbit sera (Sigma). Sig-nals were developed by using the Supersignal chemiluminescent substrate(Pierce). Membranes were exposed to X-ray film (Fuji, Tokyo, Japan) for doc-umentation.
Nucleotide sequence accession number. The sequence of CdMDR1 has beendeposited in the EMBL nucleotide sequence database under accession no.AJ227752.
RESULTS
Susceptibility testing of C. dubliniensis isolates. All of theC. dubliniensis clinical isolates listed in Table 1, except CD72,were tested previously for their susceptibilities to the azoleantifungal drugs fluconazole and itraconazole and the polyeneantifungal drug amphotericin B (19) (Table 1). The clinicalisolate CM2 from patient no. 1 and the CD47 series of isolatesfrom patient no. 4 all displayed reduced susceptibility to flu-conazole (MICs, 8 to 32 mg/ml) (Table 1). No cross-resistanceto itraconazole or amphotericin B was observed for these iso-lates. The C. dubliniensis clinical isolate CD72, first describedin this study, was found to display the highest level of flucon-azole resistance (MIC, 128 mg/ml) of all the clinical isolates
tested here or previously but was not cross resistant to itracon-azole or amphotericin B (Table 1). The fluconazole-resistantC. dubliniensis derivatives previously described by Moran et al.(19) (Table 1) also showed reduced susceptibility to flucon-azole compared to their respective parental isolates (MICs, 16to 64 mg/ml) (Table 1). These derivatives were originally gen-erated by culturing their respective fluconazole-susceptible pa-rental isolates successively on agar media containing increasingconcentrations of fluconazole. To investigate the developmentof fluconazole resistance in C. dubliniensis more closely, thefluconazole-susceptible clinical isolate CD57 was cultured onYPD agar containing progressively increasing concentrationsof fluconazole (0.5 to 50 mg/ml). The derivative series gener-ated, CD57C to CD57K (Table 1), follows the development ofaltered levels of fluconazole susceptibility, from the susceptibleparental isolate (CD57) through the final fluconazole-resistantderivative, CD57K. The derivatives CD57C to CD57E werefound to exhibit the same fluconazole susceptibility as theirparental isolate, CD57 (MIC, 0.5 mg/ml); derivatives CD57F toCD57H each exhibited a fluconazole MIC of 8 mg/ml, andderivatives CD57I-K exhibited a fluconazole MIC of 32 mg/ml.No cross-resistance to itraconazole was exhibited by any of thederivative series CD57C to CD57K (Table 1).
Further susceptibility tests were carried out with methotrex-ate, 4NQO, cycloheximide, and benomyl. Fluconazole-suscep-tible isolates and derivatives had methotrexate MICs of 32 to64 mg/ml, whereas isolates with reduced susceptibility to flu-conazole (MICs, $8 mg/ml) had MICs which were up to four-fold higher (128 to .256 mg/ml) (Table 1). Similarly, resistanceto fluconazole was also associated with reduced susceptibilityto 4NQO. Isolates with fluconazole MICs of $8 mg/ml had4NQO MICs which were four- to eightfold higher than thosefor fluconazole-susceptible isolates and derivatives (Table 1).The MICs of cycloheximide were .256 mg/ml for all of theclinical isolates and derivatives tested, with the exception of theclinical isolates CD48-I and CD48-II, each of which had acycloheximide MIC of 128 mg/ml, and the fluconazole-suscep-tible parental isolate CD51-II, which had a cycloheximide MICof 64 mg/ml. Benomyl MICs were found to range between 16 to32 mg/ml for all the isolates and derivatives tested. No corre-lation between fluconazole resistance and increased benomylor cycloheximide MICs was found. These data suggested thatfluconazole resistance in C. dubliniensis is associated withcross-resistance to the structurally unrelated compounds meth-otrexate and 4NQO and therefore would indicate a multidrug-resistant phenotype.
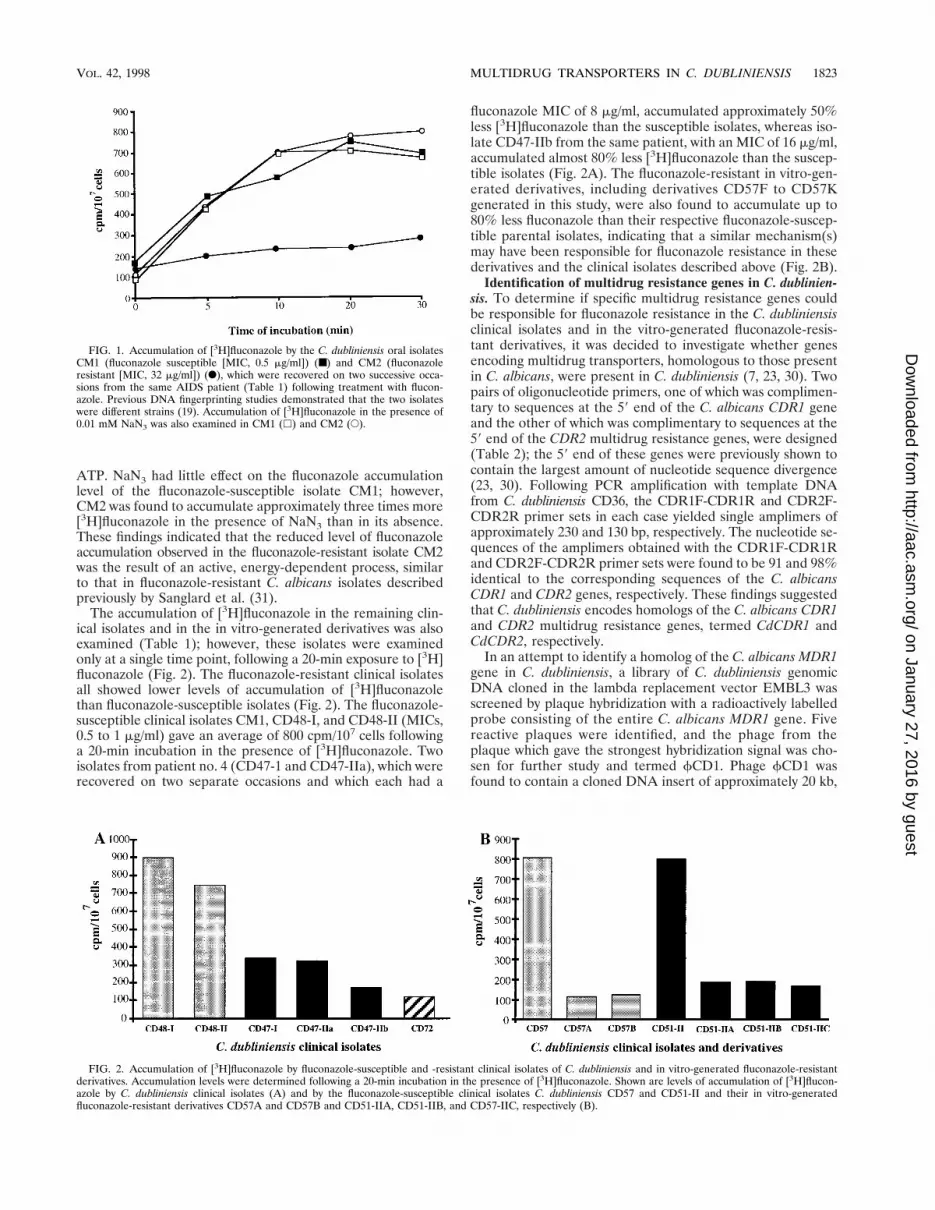
Accumulation of [3H]fluconazole in fluconazole-susceptibleand -resistant cells. To determine if alterations in cellularpermeability to fluconazole could be responsible for flucon-azole resistance in C. dubliniensis isolates and derivatives, cellswere incubated in the presence of [3H]fluconazole and theintracellular contents of this compound in fluconazole-suscep-tible and -resistant cells were determined. The clinical isolatesCM1 (fluconazole MIC, 0.5 mg/ml) and CM2 (fluconazoleMIC, 32 mg/ml) were recovered from the same AIDS patienton two separate occasions, and by using DNA fingerprinting itwas shown previously that the two isolates were differentstrains (Table 1) (19). These isolates were exposed to [3H]flu-conazole and examined at different time intervals (Fig. 1). Thetwo isolates were found to differ with regard to fluconazole ac-cumulation. The fluconazole-resistant isolate CM2 was foundto accumulate 75% less [3H]fluconazole than the susceptibleisolate CM1 following a 20-min incubation (Fig. 1). To deter-mine if this was an active, energy-dependent process, bothisolates were exposed to a subinhibitory concentration of so-dium azide (NaN3; 0.01 mM), which affects the generation of
ATP. NaN3 had little effect on the fluconazole accumulationlevel of the fluconazole-susceptible isolate CM1; however,CM2 was found to accumulate approximately three times more[3H]fluconazole in the presence of NaN3 than in its absence.These findings indicated that the reduced level of fluconazoleaccumulation observed in the fluconazole-resistant isolate CM2was the result of an active, energy-dependent process, similarto that in fluconazole-resistant C. albicans isolates describedpreviously by Sanglard et al. (31).
The accumulation of [3H]fluconazole in the remaining clin-ical isolates and in the in vitro-generated derivatives was alsoexamined (Table 1); however, these isolates were examinedonly at a single time point, following a 20-min exposure to [3H]fluconazole (Fig. 2). The fluconazole-resistant clinical isolatesall showed lower levels of accumulation of [3H]fluconazolethan fluconazole-susceptible isolates (Fig. 2). The fluconazole-susceptible clinical isolates CM1, CD48-I, and CD48-II (MICs,0.5 to 1 mg/ml) gave an average of 800 cpm/107 cells followinga 20-min incubation in the presence of [3H]fluconazole. Twoisolates from patient no. 4 (CD47-1 and CD47-IIa), which wererecovered on two separate occasions and which each had a
fluconazole MIC of 8 mg/ml, accumulated approximately 50%less [3H]fluconazole than the susceptible isolates, whereas iso-late CD47-IIb from the same patient, with an MIC of 16 mg/ml,accumulated almost 80% less [3H]fluconazole than the suscep-tible isolates (Fig. 2A). The fluconazole-resistant in vitro-gen-erated derivatives, including derivatives CD57F to CD57Kgenerated in this study, were also found to accumulate up to80% less fluconazole than their respective fluconazole-suscep-tible parental isolates, indicating that a similar mechanism(s)may have been responsible for fluconazole resistance in thesederivatives and the clinical isolates described above (Fig. 2B).
Identification of multidrug resistance genes in C. dublinien-sis. To determine if specific multidrug resistance genes couldbe responsible for fluconazole resistance in the C. dubliniensisclinical isolates and in the vitro-generated fluconazole-resis-tant derivatives, it was decided to investigate whether genesencoding multidrug transporters, homologous to those presentin C. albicans, were present in C. dubliniensis (7, 23, 30). Twopairs of oligonucleotide primers, one of which was complimen-tary to sequences at the 59 end of the C. albicans CDR1 geneand the other of which was complimentary to sequences at the59 end of the CDR2 multidrug resistance genes, were designed(Table 2); the 59 end of these genes were previously shown tocontain the largest amount of nucleotide sequence divergence(23, 30). Following PCR amplification with template DNAfrom C. dubliniensis CD36, the CDR1F-CDR1R and CDR2F-CDR2R primer sets in each case yielded single amplimers ofapproximately 230 and 130 bp, respectively. The nucleotide se-quences of the amplimers obtained with the CDR1F-CDR1Rand CDR2F-CDR2R primer sets were found to be 91 and 98%identical to the corresponding sequences of the C. albicansCDR1 and CDR2 genes, respectively. These findings suggestedthat C. dubliniensis encodes homologs of the C. albicans CDR1and CDR2 multidrug resistance genes, termed CdCDR1 andCdCDR2, respectively.
In an attempt to identify a homolog of the C. albicans MDR1gene in C. dubliniensis, a library of C. dubliniensis genomicDNA cloned in the lambda replacement vector EMBL3 wasscreened by plaque hybridization with a radioactively labelledprobe consisting of the entire C. albicans MDR1 gene. Fivereactive plaques were identified, and the phage from theplaque which gave the strongest hybridization signal was cho-sen for further study and termed fCD1. Phage fCD1 wasfound to contain a cloned DNA insert of approximately 20 kb,
FIG. 1. Accumulation of [3H]fluconazole by the C. dubliniensis oral isolatesCM1 (fluconazole susceptible [MIC, 0.5 mg/ml]) (■) and CM2 (fluconazoleresistant [MIC, 32 mg/ml]) (F), which were recovered on two successive occa-sions from the same AIDS patient (Table 1) following treatment with flucon-azole. Previous DNA fingerprinting studies demonstrated that the two isolateswere different strains (19). Accumulation of [3H]fluconazole in the presence of0.01 mM NaN3 was also examined in CM1 (h) and CM2 (E).
FIG. 2. Accumulation of [3H]fluconazole by fluconazole-susceptible and -resistant clinical isolates of C. dubliniensis and in vitro-generated fluconazole-resistantderivatives. Accumulation levels were determined following a 20-min incubation in the presence of [3H]fluconazole. Shown are levels of accumulation of [3H]flucon-azole by C. dubliniensis clinical isolates (A) and by the fluconazole-susceptible clinical isolates C. dubliniensis CD57 and CD51-II and their in vitro-generatedfluconazole-resistant derivatives CD57A and CD57B and CD51-IIA, CD51-IIB, and CD57-IIC, respectively (B).
VOL. 42, 1998 MULTIDRUG TRANSPORTERS IN C. DUBLINIENSIS 1823
and Southern hybridization analysis of restriction endonucle-ase-generated fragments of fCD1 DNA with the C. albicansMDR1 gene as a probe identified a strongly hybridizing XbaI-EcoRI insert DNA fragment of 5 kb. This fragment was clonedinto pBluescript, and the resulting plasmid was termed pGM1(Fig. 3). Further restriction endonuclease mapping studies andSouthern hybridization analysis with the C. albicans MDR1gene identified a 2.6-kb ClaI-SpeI fragment within the clonedDNA of pGM1; this was also subcloned in pBluescript to yieldplasmid pGM2 (Fig. 3). To identify an open reading frame(ORF), approximately 2 kb of the ClaI-SpeI fragment of pGM2was sequenced on both strands, corresponding to the regionbetween the ClaI site and the RsaI site, as shown in Fig. 3.Computer analysis of the 1,967-bp ClaI-RsaI fragment ofpGM2 revealed the presence of one significant ORF of 1,815bp with two potential ATG start codons at nucleotide positions2141 and 11 (numbering the sequence in the 59-to-39 direc-tion from the first base [11] of the proposed translation startcodon [Fig. 4]). The size of the protein encoded by CdMDR1,as determined in Western blotting experiments, and compari-son with the corresponding sequence of CaMDR1 suggestedthat the actual coding sequence starts at position 11, as shownin Fig. 4. This proposed start codon is preceded by a putativepromoter region in the 59 flanking sequence, including a CTblock at nucleotide positions 2104 to 286 and an adenineresidue at position 23. Although a number of TA-rich regionswere present, none matched the transcription initiation con-sensus TATAA (Fig. 4).
This ORF, termed CdMDR1, has the capacity to encode aprotein of 557 amino acids with a predicted molecular weightof 62.2 kDa and a pI of 6.4 (Fig. 4). A hydropathy plot gener-ated by the method of Kyte and Doolittle (16) indicates thatthe structure of the predicted protein encoded by CdMDR1,termed CdMdr1p, is very similar to that of the correspondingC. albicans protein, CaMdr1p, consisting of two halves, eachwith six putative transmembrane hydrophobic domains, typicalof the 12-transmembrane segment (12-TMS) family of drugexport proteins within the major facilitator superfamily (MFS)of transporters (11, 21). Also in common with the correspond-ing C. albicans Mdr1p protein is the presence of a hydrophilicstretch of amino acids near the N terminus.
The C. dubliniensis and C. albicans MDR1 genes are highly
FIG. 3. Restriction map of CdMDR1-encoding DNA from C. dubliniensisCD36. The black rectangular boxes represent C. dubliniensis genomic DNA. Theupper part of the figure shows the 5-kb CdMDR1-encoding EcoRI-XbaI fragmentsubcloned from recombinant phage fCD1 into vector plasmid pBluescript, yield-ing recombinant plasmid pGM1. The lower part of the figure shows the 2.6-kbCdMDR1-encoding SpeI-ClaI fragment of pGM1 subcloned into pBluescript,yielding recombinant plasmid pGM2. The thin double-arrowed line representsthe 2-kb fragment of pGM2 insert DNA which was sequenced. The single-arrowed line, indicating the position of and showing the direction of transcriptionof the 1,674-bp ORF encoding the CdMDR1 gene, represents the region whichwas PCR amplified from pGM2 insert DNA, using a high-fidelity proofreadingpolymerase, and subcloned into the S. cerevisiae expression vector plasmidpAAH5, yielding recombinant plasmid pGM3. Restriction endonuclease cleav-age sites are abbreviated as follows: A, AccI; B, BstxI; C, ClaI; E, EcoRI; K, KpnI;R, RsaI; S, SpeI; and X, XbaI.
FIG. 4. Nucleotide sequence and deduced amino acid sequence of theC. dubliniensis CdMDR1 gene. Nucleotide sequences are numbered in the 59-to-39 direction from the first base (11) of the ATG translation start codon.Amino acid sequences are numbered from the initial methionine. A putative CTblock is shown in boldface at nucleotide positions 2104 to 286. Amino acidresidues which are underlined show the positions of motifs typical of proteinswithin the MFS of transporter proteins, and residues shown in boldface are thosewhich match the consensus motif as described by Paulsen et al. (21). Thesecorrespond to motif D2 (residues 124 to 134), motif A (residues 168 to 180),motif B (residues 203 to 215), motif C (residues 248 to 259), and motif G(residues 499 to 505). Also underlined are the WRW and PET motifs, at residues265 to 267 and residues 288 to 290, respectively. The WRW and PET motifscorrespond to highly conserved regions in related MFS proteins from S. cerevi-siae, although their functions are unknown (11).
homologous, being 92% identical at the nucleotide sequencelevel, as determined with the CLUSTAL sequence alignmentcomputer program (12). CdMdr1p, at 557 amino acids inlength, is 7 amino acids shorter than CaMdr1p (7). Alignmentof the amino acid sequences of the two proteins shows thatthey are highly homologous, being 96.2% identical. Much ofthe divergence occurs within the hydrophilic N terminus.CaMdr1p contains an asparagine-rich region from amino acidresidues 81 to 87 which is partly absent in CdMdr1p (Fig. 5).Most of the remaining amino acid substitutions in CdMdr1pare conservative in nature, the two proteins being 98.7% sim-ilar. Interestingly, CdMDR1, like CaMDR1, does not encode aCUG codon.
A number of motifs, described by Paulsen et al. (21), whichare conserved within the 12-TMS family of drug export pro-teins can be identified in the CdMdr1p and CaMdr1p aminoacid sequences. Motifs A (CdMdr1p amino acid residues 168to 180, G x L a D r x G r K x x l, where residues shown inuppercase type are present in at least 70% of aligned se-quences analyzed by Paulsen et al. [21] and those shown inlower case are present in approximately 50% of the analyzedsequences) and B (CdMdr1p amino acid residues 203 to 215, lx x x R x x q G g a s) are common throughout the MFS and arebelieved to play a critical structural role (Fig. 4). Motif A ispoorly conserved; however, motif B can be clearly identified inboth proteins. Motif C (CdMdr1p amino acid residues 248 to259, g x x x G P x x G G x l), which is specific for drugtransporters within the MFS, is well conserved and may play arole in drug binding or transport. Motifs D2 (CdMdr1p aminoacid residues 124 to 134, l g x x x x x P v x P) and G (CdMdr1pamino acid residues 248 to 259; G x x x G P L) are specific tothe 12-TMS transporters and are partly conserved. Also pres-ent are the WRW and PET motifs at amino acid residues 265to 267 and 288 to 290, respectively, as described by Goffeau etal. (11). These motifs, found preceding and just after the sixthtransmembrane span, respectively, were identified as highlyconserved regions in related MFS proteins from S. cerevisiae,although their function is unknown (11).
Southern hybridization analysis of EcoRI-XbaI restrictionendonuclease-digested C. dubliniensis CD36 genomic DNA,with the CdMDR1 gene, localized the CdMDR1 gene to a sin-gle 5-kb fragment, identical in size to the fragment isolatedfrom the recombinant phage fCD1 (data not shown). Furtheranalysis of ClaI-KpnI-digested CD36 genomic DNA identifieda band of approximately 1.4 kb, similar in size to the ClaI-KpnIregion of pGM2 as shown in Fig. 3, and a band of 5 kb corre-sponding to the 39 end of the gene and its flanking sequences.Southern hybridization analysis of chromosome-sized DNAmolecules from CD36 separated by pulsed-field gel electro-phoresis with the CdMDR1 gene as a probe showed that theCdMDR1 gene was located on a chromosome of approximately1.3 Mb in size, which is similar in size to C. albicans chromo-some no. 6, which has been reported as the chromosomal lo-cation of CaMDR1 (7).
Expression of the C. dubliniensis CdMDR1 gene in S. cerevi-siae. Sanglard et al. (31) demonstrated that expression of theC. albicans MDR1 gene in an azole-susceptible S. cerevisiaestrain led to the expression of a fluconazole-resistant pheno-type. In the present study, similar experiments were carried outwith the CdMDR1 structural gene, using the S. cerevisiae Dpdr5mutant strain YKKB-13. The S. cerevisiae PDR5 gene is a func-tional homolog of the C. albicans CDR1 gene, and in S. cer-evisiae YKKB-13 the PDR5 deletion renders the organism hy-persusceptible to fluconazole. The entire CdMDR1 gene wasamplified from CD36, using a high-fidelity thermostable DNApolymerase, with the primer set CdMDR1F-CdMDR1R (Ta-ble 2). A single amplification product was obtained, which wascloned into the S. cerevisiae expression vector plasmid pAAH5via the HindIII restriction endonuclease cleavage sites withinthe primer sequences, yielding the plasmid pGM3 (Fig. 3). Theplasmid pAAH5 contains the promoter for the S. cerevisiaeADC1 gene, which allows for constitutive expression of geneswhen cloned into this vector in S. cerevisiae. A representativetransformant of YKKB-13 harboring pGM3, termed YGM3,was tested for susceptibility to fluconazole and was found tohave a fluconazole MIC of 128 mg/ml, whereas the fluconazoleMIC for a transformant of YKKB-13 bearing only the vectorplasmid pAAH5 (termed YP5) was only 2 mg/ml. No differ-ences in the MICs of itraconazole and ketoconazole werefound for YGM3 and YP5. Examination of the fluconazoleaccumulation levels in YGM3 and YP5 showed that YGM3accumulated approximately 75% less [3H]fluconazole than YP5.These findings illustrate that CdMDR1 mediated expression ofa fluconazole resistance phenotype in S. cerevisiae.
To assess whether CdMDR1 could confer a multidrug resis-tance phenotype on YGM3, susceptibility to a number of un-related compounds which are known multidrug transportersubstrates was tested. Susceptibility tests were carried out onYPD agar medium as described in Materials and Methods.Transformant YGM3 was found to be less susceptible thanYP5 to benomyl, brefeldin A, cerulenin, cycloheximide, flu-phenazine, 4-NQO, 1,10-phenanthroline, sulfometuron meth-yl, and terbinafine (Table 3). This range of substrates is similarto that which has been described for the C. albicans Mdr1ptransporter. Reduced susceptibility to amorolfine was also not-
FIG. 5. Alignment of the N-terminal amino acid sequence of the Mdr1pprotein encoded by the C. albicans MDR1 gene and the corresponding aminoacid sequence of the CdMdr1p protein, encoded by the C. dubliniensis CdMDR1gene, generated with the CLUSTAL sequence alignment program (12). Asterisksindicate identical residues, dots represent similar residues, and colons representdissimilar residues. Dashes indicate gaps created to obtain alignment.
TABLE 3. Susceptibility of the S. cerevisiae transformants YP5,harboring vector plasmid pAAH5, and YGM3,
a Parenthetical values are concentrations of inhibitor (in micrograms per mil-liliter) incorporated into YPD agar medium.
b Values refer to the growth on YPD agar of colonies from inocula preparedat various dilutions as described in Materials and Methods. 4, growth at 1024
dilution; 3, growth at 1023 dilution; 2, growth at 1022 dilution; 1, growth at 1021
dilution; 0, growth at 1020 (undiluted culture); —, no growth at 1020, as deter-mined by the method of Sanglard et al. (30).
VOL. 42, 1998 MULTIDRUG TRANSPORTERS IN C. DUBLINIENSIS 1825
ed in YGM3, which has not been described previously as asubstrate for the C. albicans Mdr1p transporter (7, 30).
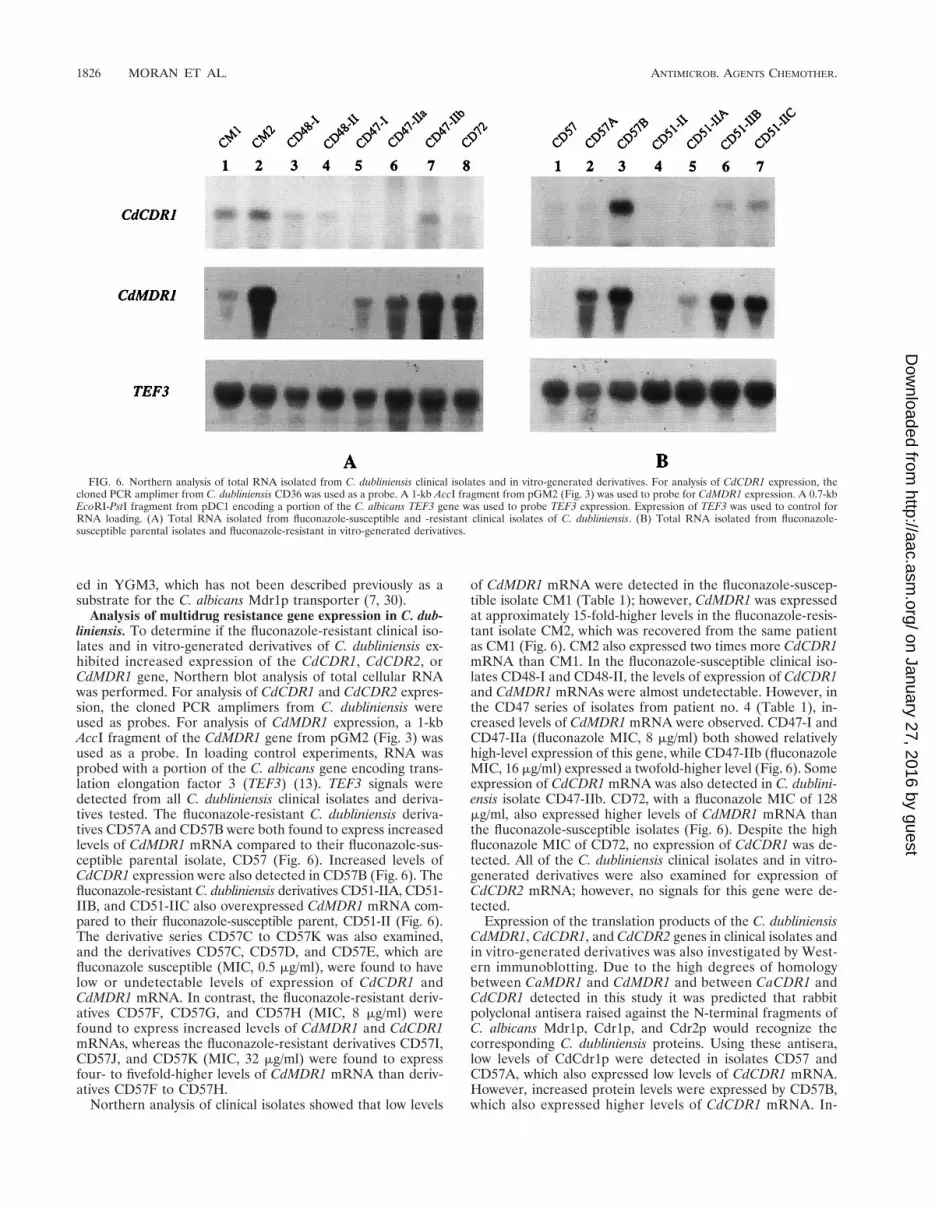
Analysis of multidrug resistance gene expression in C. dub-liniensis. To determine if the fluconazole-resistant clinical iso-lates and in vitro-generated derivatives of C. dubliniensis ex-hibited increased expression of the CdCDR1, CdCDR2, orCdMDR1 gene, Northern blot analysis of total cellular RNAwas performed. For analysis of CdCDR1 and CdCDR2 expres-sion, the cloned PCR amplimers from C. dubliniensis wereused as probes. For analysis of CdMDR1 expression, a 1-kbAccI fragment of the CdMDR1 gene from pGM2 (Fig. 3) wasused as a probe. In loading control experiments, RNA wasprobed with a portion of the C. albicans gene encoding trans-lation elongation factor 3 (TEF3) (13). TEF3 signals weredetected from all C. dubliniensis clinical isolates and deriva-tives tested. The fluconazole-resistant C. dubliniensis deriva-tives CD57A and CD57B were both found to express increasedlevels of CdMDR1 mRNA compared to their fluconazole-sus-ceptible parental isolate, CD57 (Fig. 6). Increased levels ofCdCDR1 expression were also detected in CD57B (Fig. 6). Thefluconazole-resistant C. dubliniensis derivatives CD51-IIA, CD51-IIB, and CD51-IIC also overexpressed CdMDR1 mRNA com-pared to their fluconazole-susceptible parent, CD51-II (Fig. 6).The derivative series CD57C to CD57K was also examined,and the derivatives CD57C, CD57D, and CD57E, which arefluconazole susceptible (MIC, 0.5 mg/ml), were found to havelow or undetectable levels of expression of CdCDR1 andCdMDR1 mRNA. In contrast, the fluconazole-resistant deriv-atives CD57F, CD57G, and CD57H (MIC, 8 mg/ml) werefound to express increased levels of CdMDR1 and CdCDR1mRNAs, whereas the fluconazole-resistant derivatives CD57I,CD57J, and CD57K (MIC, 32 mg/ml) were found to expressfour- to fivefold-higher levels of CdMDR1 mRNA than deriv-atives CD57F to CD57H.
Northern analysis of clinical isolates showed that low levels
of CdMDR1 mRNA were detected in the fluconazole-suscep-tible isolate CM1 (Table 1); however, CdMDR1 was expressedat approximately 15-fold-higher levels in the fluconazole-resis-tant isolate CM2, which was recovered from the same patientas CM1 (Fig. 6). CM2 also expressed two times more CdCDR1mRNA than CM1. In the fluconazole-susceptible clinical iso-lates CD48-I and CD48-II, the levels of expression of CdCDR1and CdMDR1 mRNAs were almost undetectable. However, inthe CD47 series of isolates from patient no. 4 (Table 1), in-creased levels of CdMDR1 mRNA were observed. CD47-I andCD47-IIa (fluconazole MIC, 8 mg/ml) both showed relativelyhigh-level expression of this gene, while CD47-IIb (fluconazoleMIC, 16 mg/ml) expressed a twofold-higher level (Fig. 6). Someexpression of CdCDR1 mRNA was also detected in C. dublini-ensis isolate CD47-IIb. CD72, with a fluconazole MIC of 128mg/ml, also expressed higher levels of CdMDR1 mRNA thanthe fluconazole-susceptible isolates (Fig. 6). Despite the highfluconazole MIC of CD72, no expression of CdCDR1 was de-tected. All of the C. dubliniensis clinical isolates and in vitro-generated derivatives were also examined for expression ofCdCDR2 mRNA; however, no signals for this gene were de-tected.
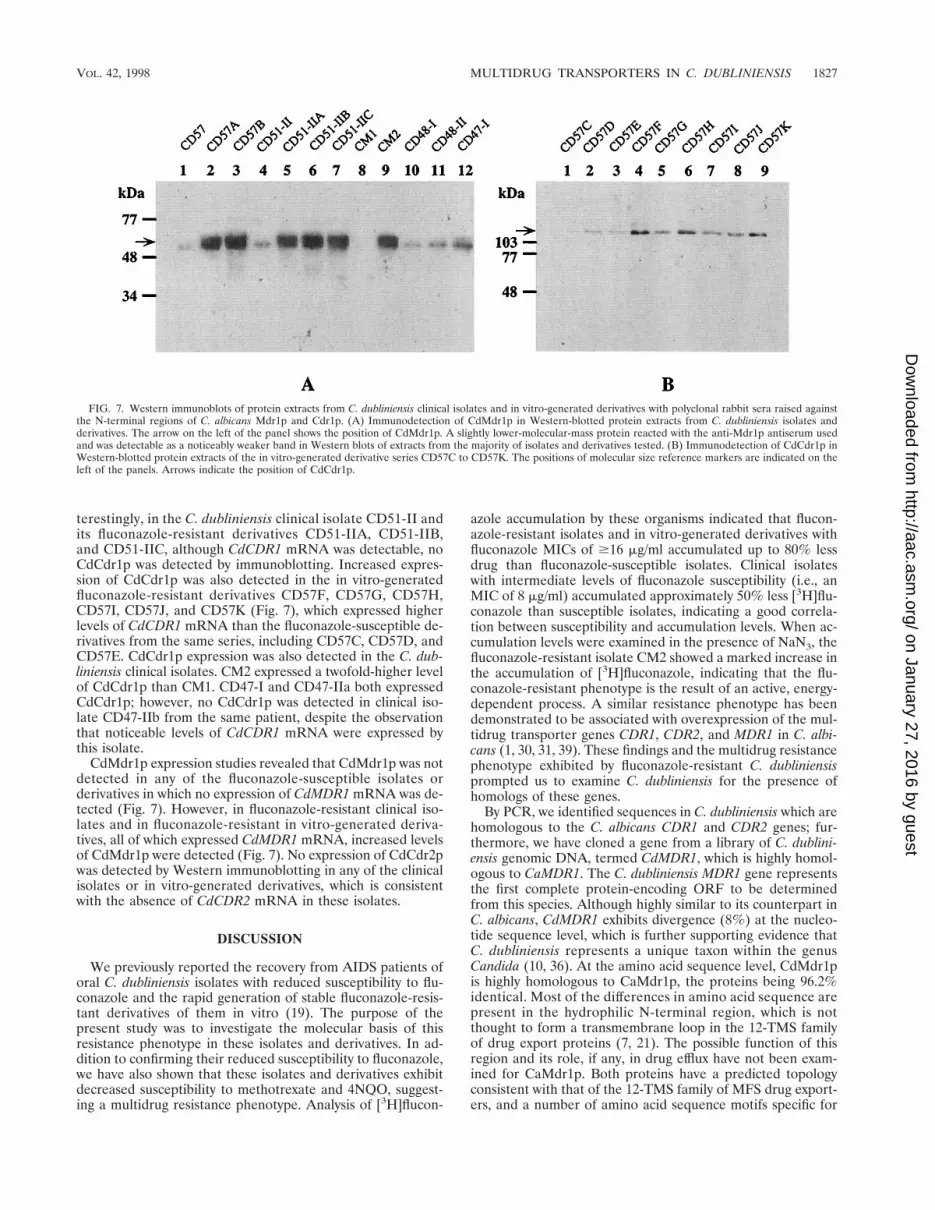
Expression of the translation products of the C. dubliniensisCdMDR1, CdCDR1, and CdCDR2 genes in clinical isolates andin vitro-generated derivatives was also investigated by West-ern immunoblotting. Due to the high degrees of homologybetween CaMDR1 and CdMDR1 and between CaCDR1 andCdCDR1 detected in this study it was predicted that rabbitpolyclonal antisera raised against the N-terminal fragments ofC. albicans Mdr1p, Cdr1p, and Cdr2p would recognize thecorresponding C. dubliniensis proteins. Using these antisera,low levels of CdCdr1p were detected in isolates CD57 andCD57A, which also expressed low levels of CdCDR1 mRNA.However, increased protein levels were expressed by CD57B,which also expressed higher levels of CdCDR1 mRNA. In-
FIG. 6. Northern analysis of total RNA isolated from C. dubliniensis clinical isolates and in vitro-generated derivatives. For analysis of CdCDR1 expression, thecloned PCR amplimer from C. dubliniensis CD36 was used as a probe. A 1-kb AccI fragment from pGM2 (Fig. 3) was used to probe for CdMDR1 expression. A 0.7-kbEcoRI-PstI fragment from pDC1 encoding a portion of the C. albicans TEF3 gene was used to probe TEF3 expression. Expression of TEF3 was used to control forRNA loading. (A) Total RNA isolated from fluconazole-susceptible and -resistant clinical isolates of C. dubliniensis. (B) Total RNA isolated from fluconazole-susceptible parental isolates and fluconazole-resistant in vitro-generated derivatives.
terestingly, in the C. dubliniensis clinical isolate CD51-II andits fluconazole-resistant derivatives CD51-IIA, CD51-IIB,and CD51-IIC, although CdCDR1 mRNA was detectable, noCdCdr1p was detected by immunoblotting. Increased expres-sion of CdCdr1p was also detected in the in vitro-generatedfluconazole-resistant derivatives CD57F, CD57G, CD57H,CD57I, CD57J, and CD57K (Fig. 7), which expressed higherlevels of CdCDR1 mRNA than the fluconazole-susceptible de-rivatives from the same series, including CD57C, CD57D, andCD57E. CdCdr1p expression was also detected in the C. dub-liniensis clinical isolates. CM2 expressed a twofold-higher levelof CdCdr1p than CM1. CD47-I and CD47-IIa both expressedCdCdr1p; however, no CdCdr1p was detected in clinical iso-late CD47-IIb from the same patient, despite the observationthat noticeable levels of CdCDR1 mRNA were expressed bythis isolate.
CdMdr1p expression studies revealed that CdMdr1p was notdetected in any of the fluconazole-susceptible isolates orderivatives in which no expression of CdMDR1 mRNA was de-tected (Fig. 7). However, in fluconazole-resistant clinical iso-lates and in fluconazole-resistant in vitro-generated deriva-tives, all of which expressed CdMDR1 mRNA, increased levelsof CdMdr1p were detected (Fig. 7). No expression of CdCdr2pwas detected by Western immunoblotting in any of the clinicalisolates or in vitro-generated derivatives, which is consistentwith the absence of CdCDR2 mRNA in these isolates.
DISCUSSION
We previously reported the recovery from AIDS patients oforal C. dubliniensis isolates with reduced susceptibility to flu-conazole and the rapid generation of stable fluconazole-resis-tant derivatives of them in vitro (19). The purpose of thepresent study was to investigate the molecular basis of thisresistance phenotype in these isolates and derivatives. In ad-dition to confirming their reduced susceptibility to fluconazole,we have also shown that these isolates and derivatives exhibitdecreased susceptibility to methotrexate and 4NQO, suggest-ing a multidrug resistance phenotype. Analysis of [3H]flucon-
azole accumulation by these organisms indicated that flucon-azole-resistant isolates and in vitro-generated derivatives withfluconazole MICs of $16 mg/ml accumulated up to 80% lessdrug than fluconazole-susceptible isolates. Clinical isolateswith intermediate levels of fluconazole susceptibility (i.e., anMIC of 8 mg/ml) accumulated approximately 50% less [3H]flu-conazole than susceptible isolates, indicating a good correla-tion between susceptibility and accumulation levels. When ac-cumulation levels were examined in the presence of NaN3, thefluconazole-resistant isolate CM2 showed a marked increase inthe accumulation of [3H]fluconazole, indicating that the flu-conazole-resistant phenotype is the result of an active, energy-dependent process. A similar resistance phenotype has beendemonstrated to be associated with overexpression of the mul-tidrug transporter genes CDR1, CDR2, and MDR1 in C. albi-cans (1, 30, 31, 39). These findings and the multidrug resistancephenotype exhibited by fluconazole-resistant C. dubliniensisprompted us to examine C. dubliniensis for the presence ofhomologs of these genes.
By PCR, we identified sequences in C. dubliniensis which arehomologous to the C. albicans CDR1 and CDR2 genes; fur-thermore, we have cloned a gene from a library of C. dublini-ensis genomic DNA, termed CdMDR1, which is highly homol-ogous to CaMDR1. The C. dubliniensis MDR1 gene representsthe first complete protein-encoding ORF to be determinedfrom this species. Although highly similar to its counterpart inC. albicans, CdMDR1 exhibits divergence (8%) at the nucleo-tide sequence level, which is further supporting evidence thatC. dubliniensis represents a unique taxon within the genusCandida (10, 36). At the amino acid sequence level, CdMdr1pis highly homologous to CaMdr1p, the proteins being 96.2%identical. Most of the differences in amino acid sequence arepresent in the hydrophilic N-terminal region, which is notthought to form a transmembrane loop in the 12-TMS familyof drug export proteins (7, 21). The possible function of thisregion and its role, if any, in drug efflux have not been exam-ined for CaMdr1p. Both proteins have a predicted topologyconsistent with that of the 12-TMS family of MFS drug export-ers, and a number of amino acid sequence motifs specific for
FIG. 7. Western immunoblots of protein extracts from C. dubliniensis clinical isolates and in vitro-generated derivatives with polyclonal rabbit sera raised againstthe N-terminal regions of C. albicans Mdr1p and Cdr1p. (A) Immunodetection of CdMdr1p in Western-blotted protein extracts from C. dubliniensis isolates andderivatives. The arrow on the left of the panel shows the position of CdMdr1p. A slightly lower-molecular-mass protein reacted with the anti-Mdr1p antiserum usedand was detectable as a noticeably weaker band in Western blots of extracts from the majority of isolates and derivatives tested. (B) Immunodetection of CdCdr1p inWestern-blotted protein extracts of the in vitro-generated derivative series CD57C to CD57K. The positions of molecular size reference markers are indicated on theleft of the panels. Arrows indicate the position of CdCdr1p.
VOL. 42, 1998 MULTIDRUG TRANSPORTERS IN C. DUBLINIENSIS 1827
this family of proteins are present within the two proteins (Fig.4), providing evidence that they are involved in drug efflux.
To demonstrate that CdMDR1 can confer resistance to flu-conazole, the gene was cloned into the S. cerevisiae expressionvector pAAH5, yielding plasmid pGM3 (Fig. 3). pGM3 wastransformed into S. cerevisiae YKKB-13, which is fluconazolesusceptible due to the deletion of the gene encoding the ABCtransporter Pdr5p. Transformants harboring pGM3 exhibited afluconazole MIC of 128 mg/ml and accumulated 75% less[3H]fluconazole than YKKB-13 transformants harboring onlythe vector (MIC, 2.0 mg/ml). No difference in itraconazole orketoconazole susceptibility was observed for the two types oftransformants. S. cerevisiae harboring pGM3 also exhibited re-duced susceptibility to a number of other compounds, includ-ing 4NQO, which suggests that CdMDR1 may be responsiblefor the multidrug-resistant phenotype of fluconazole-resistantC. dubliniensis isolates and their derivatives.
Involvement of the C. dubliniensis MDR1, CDR1, and CDR2genes in the fluconazole resistance of clinical isolates and invitro-generated derivatives was examined by probing for theirexpression in Northern and Western blots with polyclonal seraraised against the N-terminal regions of CaMdr1p, CaCdr1p,and CaCdr2p. All of the C. dubliniensis clinical isolates withreduced susceptibility to fluconazole were found to express in-creased levels of CdMDR1 mRNA and CdMdr1p, whereasin fluconazole-susceptible isolates, the levels of expressionof CdMDR1 mRNA and CdMdr1p were low or absent (Table4). These findings directly reflect previous findings with theC. albicans MDR1 gene, which was shown to confer a flucon-azole-specific azole resistance phenotype when expressed inS. cerevisiae (31). Interestingly, CdCdr1p was not detected inCD47-IIb, which showed significant levels of CdCDR1 mRNA.
Isolate CD72 was found to have a fluconazole MIC of 128 mg/ml; however, it expressed CdMDR1 mRNA at levels similar tothose of clinical isolates with fluconazole MICs of 16 to 32 mg/ml. In addition, CD72 expressed very low levels of CdCDR1mRNA. White (40), Loffler et al. (17), and Sanglard et al. (29)have all reported that mutations in the cytochrome P-450lanosterol 14a-demethylase enzyme can be associated with flu-conazole resistance. We have not yet examined the possibilitythat similar mutations are involved in fluconazole resistance inthe C. dubliniensis clinical isolates studied here, but it is pos-sible that a mutation(s) in the cytochrome P-450 lanosterol14a-demethylase enzyme could contribute to the fluconazole-resistant phenotype, particularly in CD72. It is also possiblethat an additional multidrug transporter is involved.
Although only a relatively small number of fluconazole-re-sistant C. dubliniensis and C. albicans isolates have been exam-ined so far, the patterns of expression of the CaMDR1 andCaCDR1 genes and their homologs in C. dubliniensis appear todiffer in the two species. Two recent studies reported thatincreased expression of CDR1 was the primary mechanism offluconazole resistance in C. albicans clinical isolates (1, 31),while it appears that increased expression of CdMDR1 is themain mechanism involved in C. dubliniensis clinical isolates.Expression of CDR2 has also been reported in some flucon-azole-resistant C. albicans isolates; however, no expression ofCdCDR2 mRNA or CdCdr2p was detected in the C. dublini-ensis isolates and derivatives described here.
Examination of the CdMDR1 mRNA expression levels in thein vitro-generated fluconazole-resistant derivatives recoveredfrom the fluconazole-susceptible isolates CD51-II and CD57yielded results similar to those obtained for the fluconazole-resistant clinical isolates (Table 4). All of the fluconazole-re-sistant derivatives were found to overexpress the CdMDR1 geneand CdMdr1p, whereas very low levels of expression were ob-served with the isogenic fluconazole-susceptible parental iso-lates. Three separate events could be identified during the de-velopment of fluconazole resistance in this series of derivatives(CD57C to CD57K), the first two being increases in expressionof CdMDR1 and CdCDR1 mRNAs, correlating with an increasein the fluconazole MIC to 8 mg/ml, and the second, occurringsubsequently, being further overexpression of CdMDR1, asso-ciated with a fluconazole MIC of 32 mg/ml (Table 4). White(39) reported a similar series of events, in a series of clinicalisolates of C. albicans from a single patient, in which a numberof separate events, including overexpression of the CDR1 andCaMDR1 genes, led to the development of fluconazole resis-tance.
By passage of a susceptible isolate in fluconazole-containingbroth cultures, Albertson et al. (1) isolated a fluconazole-re-sistant mutant of C. albicans which was found to express in-creased levels of the CaMDR1 gene. Calvet et al. (5) also iso-lated unstable fluconazole-resistant mutants of C. albicans bysimilar means, although no involvement of the MDR1 or CDR1gene was observed. In our experience, exposure of C. albicansto fluconazole in agar medium did not lead to a stable changein fluconazole susceptibility, and others have also reporteddifficulties in using solid media for this purpose (5). However,fluconazole-resistant derivatives of susceptible C. dublinien-sis isolates which overexpress CaMDR1 and, in some cases,CdCDR1 mRNA can be readily generated on fluconazole-containing agar medium. The fluconazole-resistant phenotypeof these derivatives is stable in the absence of fluconazole; itappears to be due to a heritable genetic change(s) rather thanto transient stress-activated transcription of multidrug trans-porter genes. The genomic DNA fingerprinting and karyotypeprofiles of some in vitro-generated fluconazole-resistant deriv-
TABLE 4. Summary of resistance mechanisms observedin fluconazole-resistant C. dubliniensis
atives of C. dubliniensis have been found to differ from those oftheir fluconazole-susceptible parental isolates (19). It is possi-ble that alterations of sequences flanking drug transportergenes or trans-acting factors influence their rates or regulationof transcription (41).
The ability of C. dubliniensis to rapidly develop fluconazoleresistance in vitro may have implications for antifungal resis-tance in vivo. If the development of fluconazole resistance inC. dubliniensis in vitro correlates with the development offluconazole resistance in vivo, it may prove to be a usefulmodel system for studying the mechanisms involved in thedevelopment of fluconazole resistance in a clinical context. It isstill unknown why C. dubliniensis has emerged, apparently,only in recent years. However, the appearance of C. dublini-ensis shortly after the widespread introduction of fluconazolefor the treatment of oral candidosis in HIV-infected and AIDSpatients, particularly in patients with recurrent infection, maybe correlated. Perhaps the ability of C. dubliniensis to rapidlyswitch on expression of the CdMDR1 gene enables this organ-ism to persist in the oral cavities of patients undergoing flu-conazole therapy. However, it is important to note that not allHIV-infected and AIDS patients undergoing fluconazole ther-apy who are colonized with C. dubliniensis yield fluconazole-resistant isolates, a situation similar to that observed with C. al-bicans (15, 24). The development of fluconazole resistance maydepend on the dosage of drug administered, the duration oftherapy, or the immune status of the patient, as has beenobserved in the case of C. albicans infection (15, 25). In addi-tion, few investigators have examined how Candida speciesrespond to fluconazole exposure in vivo, and although we ob-served stable changes in C. dubliniensis multidrug resistancegene expression in vitro, it is not known whether exposure tofluconazole in vivo could lead to changes in multidrug resis-tance gene expression. Schoofs et al. (32) demonstrated that invivo populations of C. albicans can consist of a large number ofsubtypes which differ in their relative susceptibility to antifun-gal agents, and the authors suggested that exposure to flucon-azole in vivo could lead to selection of such fluconazole-resistant subtypes. Clearly, in vivo Candida populations,including those of C. dubliniensis, may have the ability to re-spond in a dynamic fashion to antifungal therapy, a responsewhich may involve changes in gene expression or the relativeabundance of yeast species in the population. To determinethe extent and nature of fluconazole resistance in populationsof C. dubliniensis, epidemiological studies are currently con-tinuing to follow the progress of C. dubliniensis-colonizedpatients who are undergoing fluconazole therapy.
ACKNOWLEDGMENTS
Work performed in the laboratory of D.C.C. was supported by agrant from the Wellcome Trust (no. 047204). G.P.M. was supported bythe School of Dental Science and Dublin Dental Hospital, TrinityCollege Dublin, and by a short-term EMBO fellowship (no. ASTF8887). D.S. was supported by a grant from the Swiss Research NationalFoundation.
We thank B. B. Magee for the gift of plasmid p2002 and B. Hube forplasmid pDC1.
REFERENCES
1. Albertson, G. D., M. Niimi, R. D. Cannon, and H. F. Jenkinson. 1996.Multiple efflux mechanisms are involved in Candida albicans fluconazoleresistance. Antimicrob. Agents Chemother. 40:2835–2841.
2. Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990.Basic local alignment search tool. J. Mol. Biol. 215:403–410.
3. Bennett, D. E., C. E. McCreary, and D. C. Coleman. 1998. Genetic charac-terisation of a phospholipase C gene from Candida albicans: presence ofhomologous sequences in Candida species other than C. albicans. Microbi-ology 144:55–72.
4. Boerlin, P., F. Boerlin-Petzold, C. Durussel, M. Addo, J.-L. Pagani, J.-P.Chave, and J. Bille. 1995. Cluster of oral atypical Candida albicans isolatesin a group of human immunodeficiency virus-positive drug users. J. Clin.Microbiol. 33:1129–1135.
5. Calvet, H. M., M. R. Yeaman, and S. G. Filler. 1997. Reversible fluconazoleresistance in Candida albicans: a potential in vitro model. Antimicrob.Agents Chemother. 41:535–539.
6. Coleman, D. C., D. J. Sullivan, D. E. Bennett, M. C. Henman, G. P. Moran,H. J. Barry, and D. B. Shanley. 1997. The emergence of Candida dubliniensis:a novel Candida species associated with oral candidiasis in individuals in-fected with HIV and with AIDS. AIDS 11:557–567.
7. Fling, M. E., J. Kopf, A. Tamarkin, J. A. Gorman, H. A. Smith, and Y. Koltin.1991. Analysis of a Candida albicans gene that encodes a novel mechanismfor resistance to benomyl and methotrexate. Mol. Gen. Genet. 227:318–329.
8. Gallagher, P. J., D. E. Bennett, M. C. Henman, R. J. Russell, S. R. Flint,D. B. Shanley, and D. C. Coleman. 1992. Reduced azole susceptibility ofCandida albicans from HIV-positive patients and a derivative exhibitingcolony morphology variation. J. Gen. Microbiol. 138:1901–1911.
9. Genetics Computer Group. 1994. Program manual for the GCG package,version 8.0, September 1994. Genetics Computer Group, Madison, Wis.
10. Gilfillan, G. D., D. J. Sullivan, K. Haynes, T. Parkinson, D. C. Coleman, andN. A. R. Gow. 1998. Candida dubliniensis: phylogeny and putative virulencefactors. Microbiology 144:829–838.
11. Goffeau, A., J. Park, I. T. Paulsen, J.-L. Jonniaux, T. Dinh, P. Mordant, andM. H. Saier, Jr. 1997. Multidrug-resistant transport proteins in yeast: com-plete inventory and phylogenetic characterisation of yeast open readingframes within the major facilitator superfamily. Yeast 13:43–54.
12. Higgins, D. G., and P. M. Sharp. 1998. CLUSTAL: a package for performingmultiple sequence alignment on a microcomputer. Gene 73:237–244.
13. Hube, B., M. Monod, D. A. Schofield, A. J. P. Brown, and N. A. R. Gow. 1994.Expression of seven members of the gene family encoding secretory aspartylproteinases in Candida albicans. Mol. Microbiol. 14:87–99.
14. Johnson, E. M., and D. W. Warnock. 1995. Azole drug resistance in yeasts.J. Antimicrob. Chemother. 36:751–755.
15. Klepser, M. E., E. J. Ernst, and M. A. Pfaller. 1997. Update on antifungalresistance. Trends Microbiol. 5:372–375.
16. Kyte, J., and R. Doolittle. 1982. A simple method for displaying the hydro-pathic character of a protein. J. Mol. Biol. 157:105–132.
17. Loffler, J., S. L. Kelly, H. Hebart, U. Schumacher, C. Lass-Florl, and H.Einsele. 1997. Molecular analysis of cyp51 from fluconazole resistant Can-dida albicans isolates. FEMS Microbiol. Lett. 151:263–268.
18. McCullough, M., B. Ross, and P. Reade. 1995. Characterization of geneti-cally distinct subgroup of Candida albicans strains isolated from oral cavitiesof patients infected with human immunodeficiency virus. J. Clin. Microbiol.33:696–700.
19. Moran, G. P., D. J. Sullivan, M. C. Henman, C. E. McCreary, B. J. Har-rington, D. B. Shanley, and D. C. Coleman. 1997. Antifungal drug suscep-tibilities of oral Candida dubliniensis isolates from human immunodeficiencyvirus (HIV)-infected and non-HIV-infected subjects and generation of stablefluconazole-resistant derivatives in vitro. Antimicrob. Agents Chemother. 41:617–623.
20. Nguyen, M. H., J. E. Peacock, Jr., A. J. Morris, D. C. Tanner, M. L. Nguyen,D. R. Snydman, M. M. Wagener, M. G. Rinaldi, and V. L. Yu. 1996. Thechanging face of candidemia: emergence of non-Candida albicans speciesand antifungal resistance. Am. J. Med. 100:617–623.
21. Paulsen, I. T., M. H. Brown, and R. A. Skurray. 1996. Proton-dependentmultidrug efflux systems. Microbiol. Rev. 60:575–608.
22. Pfaller, M. A. 1996. Nosocomial candidiasis: emerging species, reservoirs,and modes of transmission. Clin. Infect. Dis. 22(Suppl. 2):S89–S94.
23. Prasad, R., P. De Wergifosse, A. Goffeau, and E. Balzi. 1995. Molecularcloning and characterisation of a novel gene of Candida albicans, CDR1,conferring multiple resistance to drugs and antifungals. Curr. Genet. 27:320–329.
24. Rex, J. H., C. R. Cooper, Jr., W. G. Merz, J. N. Galgiani, and E. J. Anaissie.1995. Detection of amphotericin B-resistant Candida isolates in a broth-based system. Antimicrob. Agents Chemother. 39:906–909.
25. Rex, J. H., M. G. Rinaldi, and M. A. Pfaller. 1995. Resistance of Candidaspecies to fluconazole. Antimicrob. Agents Chemother. 39:1–8.
26. Rodriguez-Tudela, J. L., and J. V. Martinez-Suarez. 1995. Defining condi-tions for microbroth antifungal susceptibility tests: influence of RPMI andRPMI-2% glucose on the selection of endpoint criteria. J. Antimicrob. Che-mother. 35:739–749.
27. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
28. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequencing withchain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463–5467.
28a.Sanglard, D. Unpublished data.29. Sanglard, D., F. Ischer, L. Kogmans, and J. Bille. 1998. Amino acid substi-
tutions in the cytochrome P-450 lanosterol 14a-demethylase (CYP51A1)from azole-resistant Candida albicans clinical isolates contribute to resis-
VOL. 42, 1998 MULTIDRUG TRANSPORTERS IN C. DUBLINIENSIS 1829
tance to azole antifungal agents. Antimicrob. Agents Chemother. 42:241–253.
30. Sanglard, D., F. Ischer, M. Monod, and J. Bille. 1997. Cloning of Candidaalbicans genes conferring resistance to azole antifungal agents: characteri-sation of CDR2, a new multidrug ABC transporter gene. Microbiology 143:405–416.
31. Sanglard, D., K. Kuchler, F. Ischer, J.-L. Pagani, M. Monod, and J. Bille.1995. Mechanisms of resistance to azole antifungal agents in Candida albi-cans isolates from AIDS patients involve specific multidrug transporters.Antimicrob. Agents Chemother. 39:2378–2386.
32. Schoofs, A., F. C. Odds, R. Colebunders, M. Ieven, L. Wouters, and H.Goosens. 1997. Isolation of Candida species on media with and withoutadded fluconazole reveals high variability in relative growth susceptibilityphenotypes. Antimicrob. Agents Chemother. 41:1625–1635.
33. Sullivan, D., D. Bennett, M. Henman, P. Harwood, S. Flint, F. Mulcahy, D.Shanley, and D. Coleman. 1993. Oligonucleotide fingerprinting of isolates ofCandida species other than C. albicans and of atypical Candida species fromhuman immunodeficiency virus-positive and AIDS patients. J. Clin. Micro-biol. 31:2124–2133.
34. Sullivan, D., K. Haynes, J. Bille, P. Boerlin, L. Rodero, S. Lloyd, M. Hen-man, and D. Coleman. 1997. Widespread geographic distribution of oralCandida dubliniensis strains in human immunodeficiency virus-infected indi-viduals. J. Clin. Microbiol. 35:960–964.
35. Sullivan, D. J., and D. C. Coleman. 1998. Candida dubliniensis: characteris-
tics and identification. J. Clin. Microbiol. 36:329–334.36. Sullivan, D. J., T. J. Westerneng, K. A. Haynes, D. E. Bennett, and
D. C. Coleman. 1995. Candida dubliniensis sp. nov.: phenotypic and molec-ular characterisation of a novel species associated with oral candidosis inHIV-infected individuals. Microbiology 141:1507–1521.
37. Vazquez, J. A., A. Beckley, J. D. Sobel, and M. J. Zervos. 1991. Comparisonof restriction enzyme analysis and pulsed-field gradient gel electrophoresis astyping systems for Candida albicans. J. Clin. Microbiol. 29:962–967.
38. Warnock, D. W. 1992. Azole drug resistance in Candida species. J. Med.Microbiol. 37:225–226.
39. White, T. C. 1997. Increased mRNA levels of ERG16, CDR, and MDR1correlate with increases in azole resistance in Candida albicans isolates froma patient infected with human immunodeficiency virus. Antimicrob. AgentsChemother. 41:1482–1487.
40. White, T. C. 1997. Presence of an R467K amino acid substitution and loss ofallelic variation correlate with an azole-resistant lanosterol 14a demethylasein Candida albicans. Antimicrob. Agents Chemother. 41:1488–1494.
41. Wickes, B. L., and R. Petter. 1996. Genomic variation in C. albicans. Curr.Top. Med. Mycol. 7:71–86.
42. Wingard, J. R., W. G. Merz, M. G. Rinaldi, T. R. Johnson, J. E. Karp, andR. Saral. 1991. Increase in Candida krusei infection among patients withbone marrow transplantation and neutropenia treated prophylactically withfluconazole. N. Engl. J. Med. 325:1274–1277.