21 GEOLOGICA BALCANICA, 41. 1–3, Sofia, Dec. 2012, p. 21–37. Integrated biostratigraphy of the Upper Cretaceous Abderaz Formation of the East Kopet Dagh Basin (NE Iran) Meysam Shafiee Ardestani 1 , Mohammad Vahidinia 1 , Abbas Sadeghi 2 , José Antonio Arz 3 , Docho Dochev 4 1 Faculty of Science, Department of Geology, Ferdowsi University of Mashhad, Mashhad, Iran; e-mail: [email protected]2 Department of Geology, Faculty of Science, University of Shahid Beheshti, Tehran, Iran 3 Departamento de Ciencias de la Tierra (Paleontología) e Instituto de Investigación en Ciencias Ambientales (IUCA), Universidad de Zaragoza, C/ Pedro Cerbuna 12, E-50009 Zaragoza, Spain 4 Department of Geology, Paleontology and Fossil Fuels, Faculty of Geology and Geography, Sofia University “St. Kliment Ohridski”, Sofia, Bulgaria (Accepted in revised form: November2012) Abstract. Based on planktonic foraminifera, inoceramids and echinoids, we present a detailed biostratigraphic analysis of the Abderaz Formation at the 606 m thick Padeha section, NE Iran. This sequence consists mainly of gray shales and marls with four levels of chalky limestones intercalated. The lower boundary of the Abderaz Formation with the Aitamir Formation is a paraconformity, while the upper boundary with the Abtalkh Formation represents a gradual transition. Fifty four species of planktonic foraminifera from 15 genera were identified, and five zones were recognized, namely: Whiteinella archaeocretacea (Bolli) Partial-Range- Zone; Helvetoglobotruncana helvetica (Sigal) Total-Range-Zone; Marginotruncana schneegansi (Dalbiez) Interval-Range-Zone; Dicarinella concavata (Brotzen) Interval-Range-Zone; and Dicarinella asymetrica (Sigal) Total-Range-Zone. Based on these data, the age of the Abderaz Formation is determined as earli- est Turonian to earliest Campanian. Inoceramid bivalves Cremnoceramus walterdorfensis walterdorfensis (Andert) and Cremnoceramus deformis deformis (Meek) were identified in the uppermost Turonian and in the middle part of the early Coniacian, respectively. Echinocorys ex. gr. scutata and Cordiceramus sp. were recorded near the Coniacian/Santonian boundary. Shafiee Ardestani, M., Vahidinia, M., Sadeghi, A., Arz, J.A., Dochev, D. 2012. Integrated biostratigraphy of the Upper Cretaceous Abderaz Formation of the East Kopet Dagh Basin (NE Iran). Geologica Balcanica 40(1–3), 21–37. Key words: Upper Cretaceous, Abderaz Formation, Biostratigraphy, Planktonic foraminiferal zonation, Inoceramids, NE Iran. INTRODUCTION The Kopet Dagh sedimentary basin was formed after the Middle Triassic orogeny in northeast Iran (Afshar-Harb, 1969). The basin was framed due to the faulting activ- ity with approximately NW–SE trend. Four of these ma- jor active basement faults have been recognized in the central and western part of the basin. Sedimentation was more or less continuous throughout the Middle Jurassic to Oligocene interval in the eastern part of the basin, where five major transgressive-regressive sequences have been identified. Moussavi-Harami and Brenner (1992) concluded that subsidence in the eastern part of the basin was predominantly a result of sediment load- ing. There is no evidence of major tectonic activity in this region; all formations are conformable, except for a few disconformities within the Cretaceous succession. These can be related to epeirogenic movements in the basin. All formations thin out from north-west to south-east. They were folded during the Late Alpine Orogeny and formed the structural traps of the Khangiran and Gonbadli gas fields (Raisossadat and Moussavi-Harami, 2000). The Cretaceous sediments in the Kopet Dagh Basin are di- vided into nine different formations, composed mainly of sandstones, conglomerates, mudstones, limestones and dolostones with subordinate evaporites. The thick- ness of these sediments is normally more than 4000 m, showing a prominent decrease to 2500 m in the eastern part of the basin (Shafiee Ardestani et al., 2011). The Abderaz Formation is the major lithostratigraphic unit of
Transcript
21
GEOLOGICA BALCANICA, 41. 1–3, Sofia, Dec. 2012, p. 21–37.
Integrated biostratigraphy of the Upper Cretaceous Abderaz Formation of the East Kopet Dagh Basin (NE Iran)
Meysam Shafiee Ardestani1, Mohammad Vahidinia1, Abbas Sadeghi2, José Antonio Arz3, Docho Dochev4
1 Faculty of Science, Department of Geology, Ferdowsi University of Mashhad, Mashhad, Iran; e-mail: [email protected] 2 Department of Geology, Faculty of Science, University of Shahid Beheshti, Tehran, Iran 3 Departamento de Ciencias de la Tierra (Paleontología) e Instituto de Investigación en Ciencias Ambientales (IUCA), Universidad de Zaragoza, C/ Pedro Cerbuna 12, E-50009 Zaragoza, Spain 4 Department of Geology, Paleontology and Fossil Fuels, Faculty of Geology and Geography, Sofia University “St. Kliment Ohridski”, Sofia, Bulgaria(Accepted in revised form: November2012)
Abstract. Based on planktonic foraminifera, inoceramids and echinoids, we present a detailed biostratigraphic analysis of the Abderaz Formation at the 606 m thick Padeha section, NE Iran. This sequence consists mainly of gray shales and marls with four levels of chalky limestones intercalated. The lower boundary of the Abderaz Formation with the Aitamir Formation is a paraconformity, while the upper boundary with the Abtalkh Formation represents a gradual transition. Fifty four species of planktonic foraminifera from 15 genera were identified, and five zones were recognized, namely: Whiteinella archaeocretacea (Bolli) Partial-Range-Zone; Helvetoglobotruncana helvetica (Sigal) Total-Range-Zone; Marginotruncana schneegansi (Dalbiez) Interval-Range-Zone; Dicarinella concavata (Brotzen) Interval-Range-Zone; and Dicarinella asymetrica (Sigal) Total-Range-Zone. Based on these data, the age of the Abderaz Formation is determined as earli-est Turonian to earliest Campanian. Inoceramid bivalves Cremnoceramus walterdorfensis walterdorfensis (Andert) and Cremnoceramus deformis deformis (Meek) were identified in the uppermost Turonian and in the middle part of the early Coniacian, respectively. Echinocorys ex. gr. scutata and Cordiceramus sp. were recorded near the Coniacian/Santonian boundary.
Shafiee Ardestani, M., Vahidinia, M., Sadeghi, A., Arz, J.A., Dochev, D. 2012. Integrated biostratigraphy of the Upper Cretaceous Abderaz Formation of the East Kopet Dagh Basin (NE Iran). Geologica Balcanica 40(1–3), 21–37.
The Kopet Dagh sedimentary basin was formed after the Middle Triassic orogeny in northeast Iran (Afshar-Harb, 1969). The basin was framed due to the faulting activ-ity with approximately NW–SE trend. Four of these ma-jor active basement faults have been recognized in the central and western part of the basin. Sedimentation was more or less continuous throughout the Middle Jurassic to Oligocene interval in the eastern part of the basin, where five major transgressive-regressive sequences have been identified. Moussavi-Harami and Brenner (1992) concluded that subsidence in the eastern part of the basin was predominantly a result of sediment load-ing. There is no evidence of major tectonic activity in this
region; all formations are conformable, except for a few disconformities within the Cretaceous succession. These can be related to epeirogenic movements in the basin. All formations thin out from north-west to south-east. They were folded during the Late Alpine Orogeny and formed the structural traps of the Khangiran and Gonbadli gas fields (Raisossadat and Moussavi-Harami, 2000). The Cretaceous sediments in the Kopet Dagh Basin are di-vided into nine different formations, composed mainly of sandstones, conglomerates, mudstones, limestones and dolostones with subordinate evaporites. The thick-ness of these sediments is normally more than 4000 m, showing a prominent decrease to 2500 m in the eastern part of the basin (Shafiee Ardestani et al., 2011). The Abderaz Formation is the major lithostratigraphic unit of
22
Late Cretaceous age in the Kopet Dagh Basin. It includes highly diverse and abundant micro- and macrofossils.
Planktonic foraminiferal biostratigraphic analyses, together with the identification of invertebrate (mainly inoceramid) bio-events were conducted in order to pre-cise the age of the Abderaz Formation. It is based on the study of the Padeha section (Mashhad Province, NE Iran), since this section was found to be well exposed and containing enough micro- and macrofossils of high bios-tratigraphic value to give a reliable age determination.
LIThOsTRaTIgRaphy
The Padeha section is located southeast of Mashhad. It crops out along the Mashhad-Sarakhs road, 25 kilo-meters east of type locality of the Abderaz Formation, and 2 kilometers from the Padeha rural road, near the Rakhshani region. The geographical position of the sec-tion is 60°44'39.6" E and 36°06'23.7" N (Fig. 1).
The Abderaz Formation at Padeha section consists of 11 lithological packets from top to bottom, namely (Fig. 2):
11) Thin bedded light gray shales, 28 m thick.10) White chalk limestones, 18 m thick.9) Thin bedded light gray marls, 300 m thick.8) Thin bedded gray marls, 10 m thick.7) Thin bedded light gray shales, 40 m thick.
6) Thick bedded white chalky limestones, 20 m thick.5) Thin bedded light gray shales, 44 m thick.4) White chalky limestones, 7 m thick.3) Thin bedded light gray shales, 79 m thick.2) White chalky limestones, 20 m thick.1) Thin bedded yellowish to gray marls, 40 m thick.The lower boundary of the Abderaz Formation with
the Aitamir Formation is a paraconformity. Upwards, the Abderaz Formation passes gradually into the Abtalkh Formation. The rocks contain common inoceramids, fre-quent echinoids, and some ammonites. Abundant plank-tonic foraminifers occur, especially in the chalky lime-stones. The total thickness of the section is 606 m.
METhODOLOgy
The sampling density at the Padeha section was 3 me-ters, thus a total of 200 samples was collected. Samples were soaked in water with diluted hydrogen peroxide, and washed through 63 μm, 150 μm and 250 μm sieves. About 200–300 foraminiferal specimenss were picked up from each sample in two size fractions (63–150 μm and >150 μm), determined, and mounted on microslides for permanent record and future revisions.
The most representative and the best preserved fora-miniferal specimens were selected for scanning electron
Fig. 1. Geological and geographical sketch-maps of the area of Padeha section (NE Iran)
23
of Ferdowsi University of Mashhad. Taxonomy is based on Bolli (1957), Postuma (1971), Robaszynski and Caron (1979, 1995), Caron (1985), Loeblich and Tappan (1988), Nederbragt (1990), Ellis and Messina (1999), and Premoli Silva and Verga (2004).
FORaMINIFERaL BIOsTRaTIgRaphy
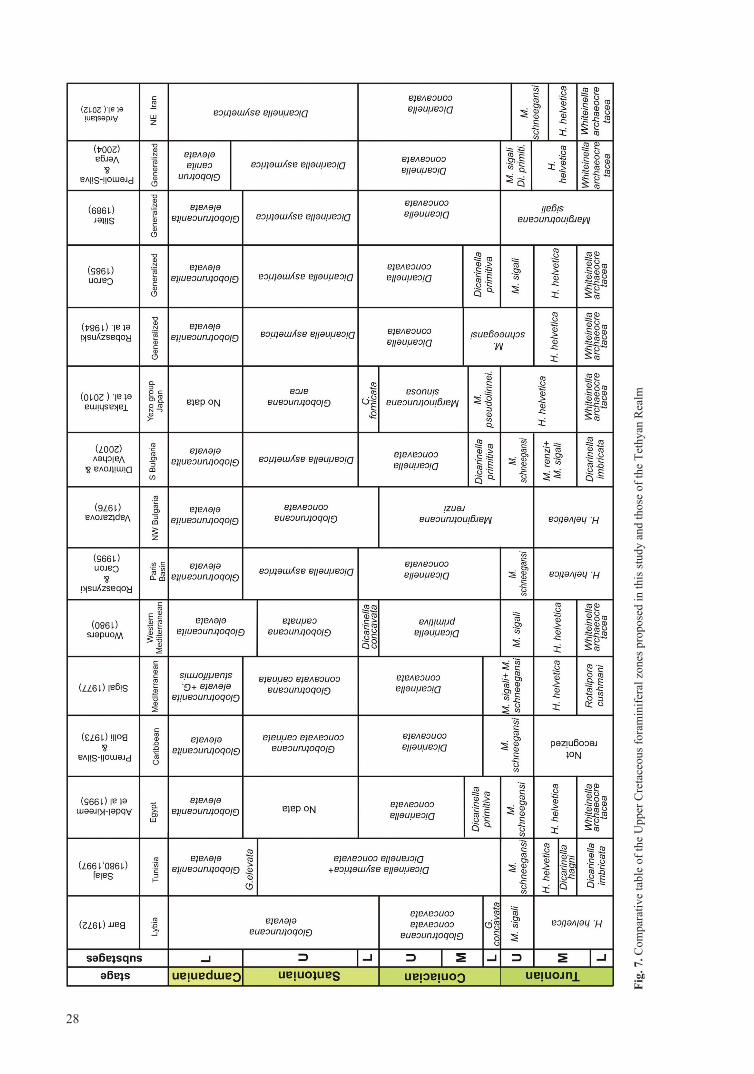
The planktonic foraminiferal zonation here used is com-pared with other Tethyan biozonations (Fig. 7) (Barr, 1972; Premoli-Silva and Bolli, 1973; Vaptzarova, 1976; Sigal, 1977; Wonders 1980; Salaj, 1980, 1997; Robaszynski et al. 1984; Caron, 1985; Sliter, 1989; Abdel-Kireem et al., 1995; Robaszynski and Caron 1995; Premoli-Silva and Verga, 2004; Dimitrova and Valchev, 2007; Takashima et al., 2010). This zonation is similar to that of Robaszynski and Caron (1995).
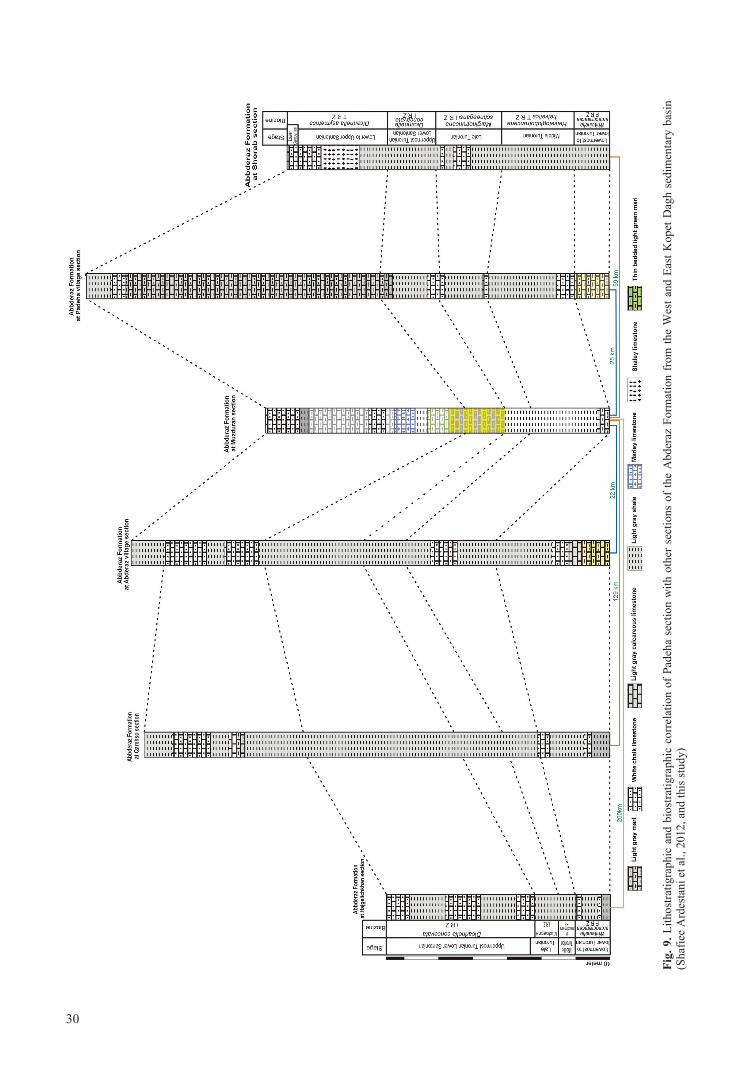
In this study, 54 species belonging to 15 genera of planktonic foraminifera, and 11 invertebrate species (8 inoceramids, 2 echinoids and 1 ammonite), were recognized in the Abderaz Formation (Fig. 8). The Padeha section has been correlated with other sec-tions of the Abderaz Formation from West and East Kopet-Dagh Basin, which were previously studied by Shafiee Ardestani et al. (2012) (Fig. 9). These are the Hajgelichkhan section (Lowermost Turonian-Lowermost Santonian), Qarehso section (Lowermost Turonian-Late Santonian), Abderaz section (Early Turonian-Lowermost Cam panian), Muz duran type section (Middle Turonian-Lowermost Campanian), and Shorab section (Lowermost Turonian-Lowermost Campanian) (Fig 9). Di. asymetrica Zone is of greater thickness than the others (Fig. 9). Rates of sedimenta-tion in all studied sections are different.
Planktonic foraminiferal biostratigraphy indicates early Turonian to earliest Campanian age of the Abderaz Formation. Five successive zones are described below.
Definition. Interval with Whiteinella archaeocretacea between the last occurrence data (LOD) of Rotalipora cushmani and the first occurrence data (FOD) of Helvetoglobotruncana helvetica.planktonic foraminifera assemblage: Dicarinella cana liculata (Reuss, 1854), Di. hagni (Scheibnerova, 1962), Heterohelix globulosa (Ehrenberg, 1840), H. more mani (Cushman, 1938), H. reussi (Cushman, 1938), Macroglobigerinelloides bollii (Pessagno, 1967), M. ca seyi (Bolli, Loeblich and Tappan, 1957), M. ultra-micra (Subbotina, 1949), Muricohedbergella delrioensis (Carsey, 1926), M. simplex (Morrow, 1934), Whiteinella aprica (Loeblich and Tappan, 1961), W. baltica Douglas and Rankin, 1969, W. brittonensis (Loeblich and Tappan, 1961), W. paradubia (Sigal, 1952), and W. praehelvetica (Trujillo, 1960).Remarks. The base of this zone coincides with the bound-ary between the Aitamir and the Abderaz Formation, indicating an Early Turonian age for the lower part of the Abderaz Formation. The planktonic foraminifers
Fig. 2. Lithological column of the Abderaz Formation at Padeha section
microscopy (SEM), using a JEOL JSM 6400 SEM at the Microscopy Service of the Universidad de Zaragoza (Spain). They are illustrated in Figures 3–6. We also studied planktonic foraminiferal collection from the Abderaz Formation deposited at the National Museum
24
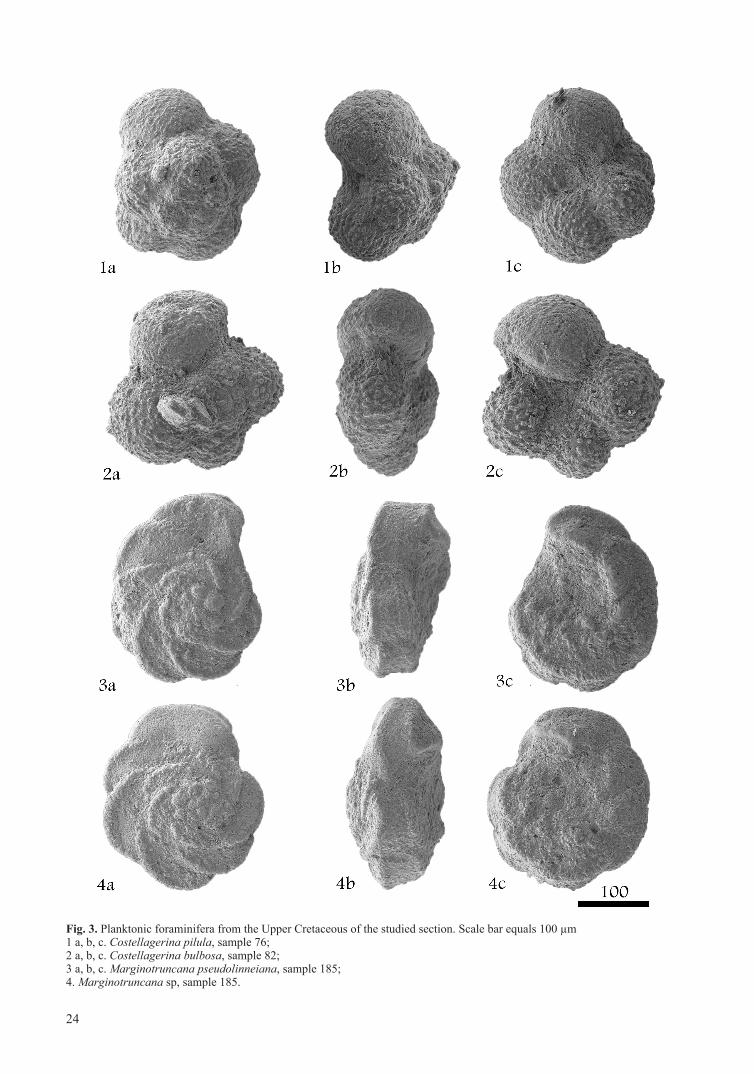
Fig. 3. Planktonic foraminifera from the Upper Cretaceous of the studied section. Scale bar equals 100 µm1 a, b, c. Costellagerina pilula, sample 76;2 a, b, c. Costellagerina bulbosa, sample 82;3 a, b, c. Marginotruncana pseudolinneiana, sample 185; 4. Marginotruncana sp, sample 185.
25
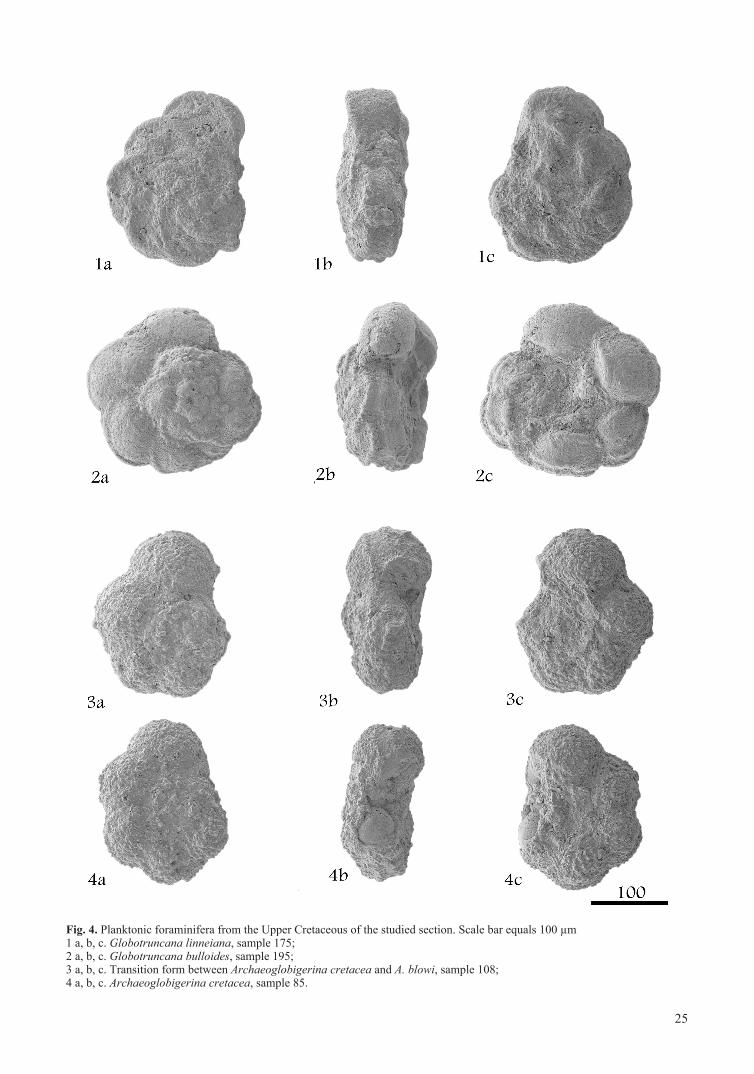
Fig. 4. Planktonic foraminifera from the Upper Cretaceous of the studied section. Scale bar equals 100 µm1 a, b, c. Globotruncana linneiana, sample 175; 2 a, b, c. Globotruncana bulloides, sample 195; 3 a, b, c. Transition form between Archaeoglobigerina cretacea and A. blowi, sample 108; 4 a, b, c. Archaeoglobigerina cretacea, sample 85.
26
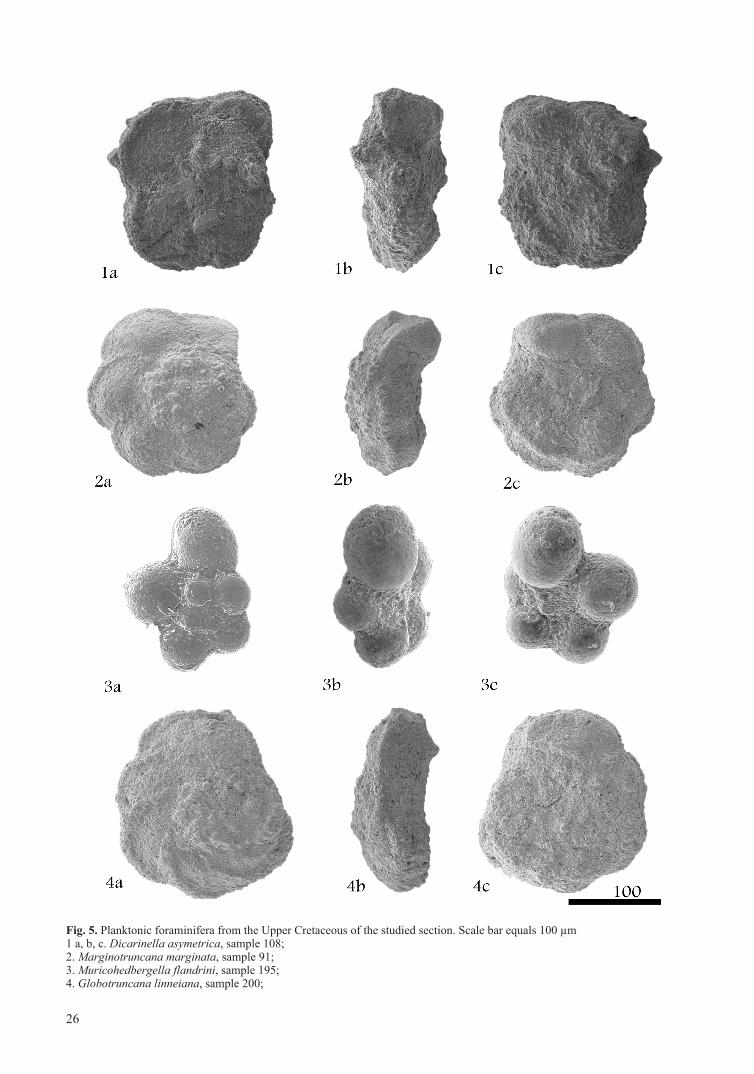
Fig. 5. Planktonic foraminifera from the Upper Cretaceous of the studied section. Scale bar equals 100 µm1 a, b, c. Dicarinella asymetrica, sample 108; 2. Marginotruncana marginata, sample 91; 3. Muricohedbergella flandrini, sample 195; 4. Globotruncana linneiana, sample 200;
27
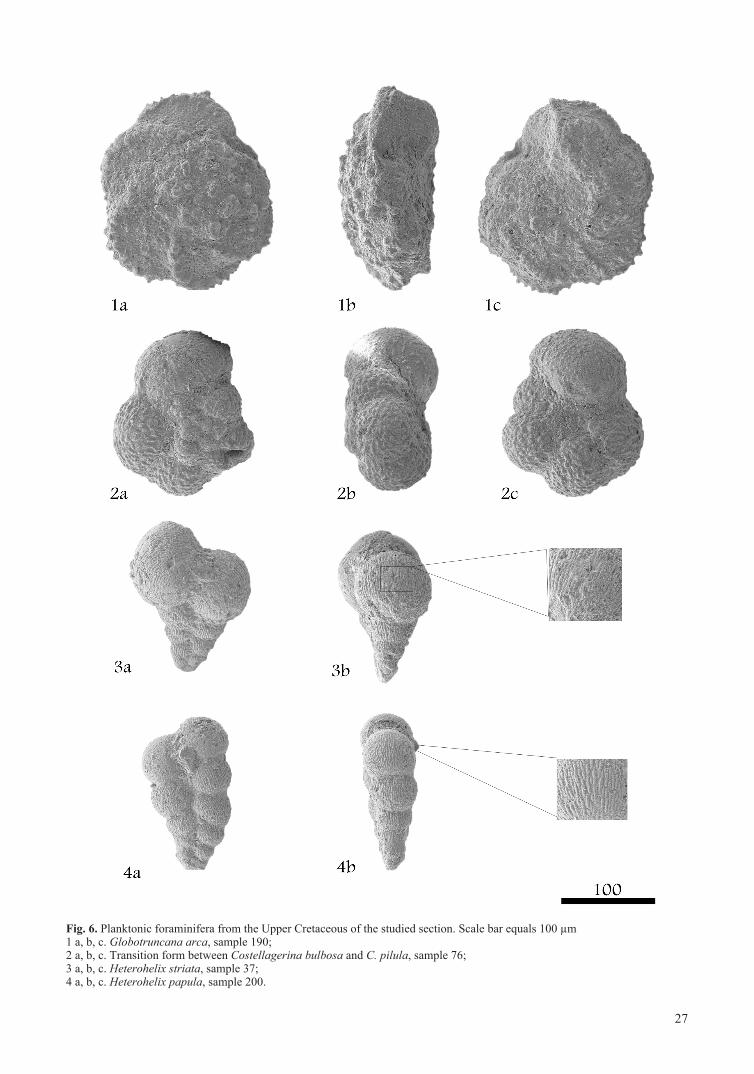
Fig. 6. Planktonic foraminifera from the Upper Cretaceous of the studied section. Scale bar equals 100 µm1 a, b, c. Globotruncana arca, sample 190; 2 a, b, c. Transition form between Costellagerina bulbosa and C. pilula, sample 76;3 a, b, c. Heterohelix striata, sample 37;4 a, b, c. Heterohelix papula, sample 200.
28
Fig.
7. C
ompa
rativ
e ta
ble
of th
e U
pper
Cre
tace
ous f
oram
inife
ral z
ones
pro
pose
d in
this
stud
y an
d th
ose
of th
e Te
thya
n R
ealm
29
Fig.
8. B
iost
ratig
raph
ic ra
nge-
char
t of p
lank
toni
c fo
ram
inife
rs a
nd m
acro
foss
il in
verte
brat
es re
cord
ed fr
om th
e A
bder
az F
orm
atio
n at
Pad
eha
sect
ion
30
Fig.
9. L
ithos
tratig
raph
ic a
nd b
iost
ratig
raph
ic c
orre
latio
n of
Pad
eha
sect
ion
with
oth
er s
ectio
ns o
f th
e A
bder
az F
orm
atio
n fr
om th
e W
est a
nd E
ast K
opet
Dag
h se
dim
enta
ry b
asin
(S
hafie
e A
rdes
tani
et a
l., 2
012,
and
this
stud
y)
31
are poorly preserved. This zone consists of yellowish to gray marls 39 m thick (see Fig. 8). Inoceramus sp. and Mytiloides sp. were also determined (Fig. 10).
This zone was previously identified in the Lower Turonian of the western Tethys (Wonders, 1980). According to Caron (1985) and Wonders (1980), the W. archaeocretacea Zone is Early Turonian in age, but they suggested the FOD of W. archaeocretacea is uppermost Cenomanian, too. However, Pessagno (1967) consid-ered this zone as corresponding to the Upper Turonian. Postuma (1971) used Helvetoglobotruncana helvetica as the index species in defining a Total-Range-Zone of Turonian age. Sigal (1977) used the Rotalipora cush-mani Total-Range-Zone for marking the Cenomanian-Turonian transition. In low latitudes, Premoli-Silva and Verga (2004) indicated that the W. archaeocretacea Zone is of Cenomanian-Turonian age, but Walaszczyk et al. (2004) considered this zone being of Turonian age in the Bryansk Region.Chronostratigraphy. Uppermost Cenomanian to Lower Turonian.
Definition. Interval of the total range of Helvetoglobo-truncana helvetica.planktonic foraminifera assemblage: Dicarinella al-geriana (Caron, 1966), Di. hagni (Scheibnerova, 1962), Di. imbricata (Monrod, 1950), Macroglobigerinelloides ultramicra (Subbotina, 1949), Heterohelix globulosa (Ehrenberg, 1840), H. moremani (Cushman, 1938), Muricohedbergella delrioensis (Carsey, 1926), M. plan-ispira (Tappan, 1940), Marginotruncana renzi (Gandolfi, 1942), M. sinuosa Porthault, 1970, M. sigali (Reichel, 1950), Whiteinella aprica (Loeblich and Tappan, 1961), W. baltica Douglas and Rankin 1969, W. praehelvetica Trujillo, 1960, and W. paradubia (Sigal, 1952).Remarks. This zone is 102 m thick (samples 13-47) and includes chalky limestones with shales intercalated (Figs 2, 8). Inoceramus sp. is frequent. The zone was previ-ously identified from the Middle Turonian in the Atlantic (McNulty, 1976; Premoli-Silva and Sliter 1981), western Tethys (Wonders, 1980), central Tethys (Fleury, 1980; Sigal, 1977) and Pacific (Gradstein et al., 1978; Pessagno and Longoria, 1973). Keller et al. (2004) and Keller and Pardo (2004) considered that H. helvetica range is dia-chronous in different localities.Chronostratigraphy. Lower and Middle Turonian.
Definition. The interval between the FOD of Margino-truncana schneegansi and the FOD of Dicarinella primitiva.planktonic foraminifera assemblage: Archaeoglobige-rina bosquensis Pessagno, 1967, Dicarinella canaliculata (Reuss, 1854), Heterohelix globulosa (Ehrenberg, 1840), Macroglobigerinelloides prairiehillensis (Pessagno, 1967), M. caseyi (Bolli et al., 1957), Marginotruncana paraconcavata Porthault, 1970, M. pseudolinneiana
Pessagno, 1967, M. renzi (Gandolfi, 1942), M. sigali (Reichel, 1950), Muricohedbergella delrioensis (Carsey, 1926), M. planispira (Tappan, 1940), Whiteinella ar-chaeocretacea Pessagno, 1967, W. baltica Douglas and Rankin, 1969, and W. paradubia (Sigal, 1952).Remarks. This zone is 42 m thick (samples 47 to 61), and consists of light gray shales with yellowish to white chalky limestones (Figs 2, 8). Inoceramus sp., Mytiloides sp., Cremnoceramus walterdorfensis walterdorfensis and Mytiloides herbichi were identified (Fig. 10).
This interval is also known as the Middle to Upper Turonian Marginotruncana sigali-Dicarinella primitiva Zone (Premoli Silva and Sliter, 1995). It was identified from the Upper Turonian to Lower Coniacian in the Atlantic (McNulty, 1976; Premoli Silva and Sliter, 1981, 1995), western Tethys (Wonders, 1980), central Tethys (Fleury, 1980, Sigal, 1977), eastern Tethys (Gorbachik, 1971a, b; Maslakova, 1971) and Pacific (Gradstein et al., 1978; Pessagno and Longoria, 1973). Robaszynski and Caron (1995) identified this biozone in the whole Tethyan Realm.Chronostratigraphy Upper Turonian to Lower Coniacian.
Dicarinella concavata Interval Zone sigal, 1955
Definition. The interval between the FOD of Dicari nella concavata and the FOD of Dicarinella asymetrica.planktonic foraminifera assemblage: Archaeoglo-bigerina bosquensis Pessagno, 1967, A. cretacea (d’Or-bigny, 1840), Costellagerina pilula (Belford, 1960), C. bulbosa (Belford, 1960), Dicarinella canaliculata (Reuss, 1854), Di. hagni (Scheibnerova, 1962), Di. pri mitiva (Dalbiez, 1955), Macroglobigerinelloides caseyi (Bolli et al., 1957), Muricohedbergella delri-oensis (Carsey, 1926), M. flandrini (Porthault, 1970), M. planispira (Tappan, 1940), M. simplex (Morrow, 1934), Heterohelix globulosa (Ehrenberg, 1840), Mar-ginotruncana marginata (Reuss, 1845), M. paraconca-vata Porthault, 1970, M. renzi (Gandolfi, 1942), M. si-gali (Reichel, 1950), M. schneegansi (Sigal, 1952), M. sinuosa Porthault, 1970, Whiteinella archeocretacea Pessagno, 1967, W. baltica Douglas and Rankin, 1969, W. brittonensis (Loeblich and Tappan, 1961), W. au-malensis (Sigal, 1952), and W. paradubia (Sigal, 1952).Remarks. The thickness is about 78 m (samples 62 to 88), and the zone consists of shales, chalky limestones and light gray marls (Figs 2, 8)
This zone was defined by Sigal (1955) who used FOD of Di. primitiva as the lower boundary indicator. FODs of Di. primitiva and Di. concavata have been documented almost at the same level in the Padeha sec-tion, thus we define the base of Di. concavata Zone at the FOD of the index species. Sigal (1955) suggested a Late Coniacian to Early Santonian age for this zone, but later it was dated as Coniacian to early Santonian by Premoli Silva and Verga (2004). Walaszczyk et al. (2012) documented that the FODs of Di. concavata and Di. primiriva are coeval in the Upper Turonian Mytiloides scupini inoceramid zone.
The zone was identified in the Coniacian to Lower Santonian from the Atlantic Realm (McNulty, 1976;
32
33
Premoli-Silva and Sliter 1981), western Tethys (Wonders, 1980), central Tethys (Fleury, 1980; Sigal, 1977) and the Pacific Realm (Gradstein et al. 1978, 1994; Pessagno and Longoria, 1973). The following invertebrate macrofossils have also been found: Inoceramus sp. (Upper Turonian), Mytiloides sp. (Upper Turonian), Cremnoceramus wal-terdorfensis walterdorfensis (Andert, 1911) (Uppermost Turonian), Mytiloides herbichi (Atabekian, 1968) (Upper Turonian), Cremnoceramus cf. deformis deformis (Meek, 1871), Inoceramus cf. vistulensis (Walaszczyk, 1992), and Echinocorys gr. scutata (base of Santonian).Chronostratigraphy. Uppermost Turonian, Cenomanian and Lowermost Santonian.
Dicarinella asymetrica Total Range Zone postuma, 1971
Definition. Total range zone of Dicarinella asymetrica.planktonic foraminifera assemblage: Archaeoglobi-gerina bosquensis Pessagno, 1967, A. cretacea (d’Or-bigny, 1840), Contusotruncana fornicata (Plummer, 1931), C. pateliformis (Gandolfi, 1955), Dicarinella ca-naliculata (Reuss, 1854), Di. concavata (Brotzen, 1934), Di. hagni (Scheibnerova, 1962), Globotruncana arca (Cushman, 1926), G. bulloides Vogler, 1941, G. linneia-na (d’Orbigny, 1839), Heterohelix globulosa (Ehrenberg, 1840), H. papula (Belford, 1960), Laeviheterohelix pul-chra (Brotzen, 1936), Macroglobigerinelloides alvarezi (Eternod Olvera, 1959), M. bollii (Pessagno, 1967), M. escheri Kaufmann, 1865, M. prairiehillensis (Pessagno, 1967), M. sp., Marginotruncana coronata (Bolli, 1945), M. marginata (Reuss, 1845), M. pseudolinneiana Pessag-no, 1967, M. paraconcavata Porthault, 1970, M. ren-zi (Gandolfi, 1942), M. schneegnasi (Sigal, 1952), M. sigali (Reichel, 1950), M. sinuosa Porthault, 1970, M. tarfayensis (Lehmann, 1963), Muricohedbergella flandrini (Porthault, 1970), M. holmdelensis (Olsson, 1964), M. planispira (Tappan, 1940), M. simplex (Morrow, 1934), Pseudotextularia nuttalli (Voorwijk, 1937), Pseudoguembelina costellifera Masters, 1976, Rugoglobigerina pennyi Brönnimann, 1952, R. rugo- R. rugo-R. rugo-sa (Plummer, 1926), Ventillabrella austiana Cushman, 1938, Whiteinella aumalensis (Sigal, 1952), and W. ar-chaeocretacea Pessagno, 1967.Remarks. This biozone is 333 m thick (samples 89 to 200) (Fig. 8). The Santonian/Campanian boundary is
placed within this zone. As far as Globotruncanita el-evata was not found, therefore we used the LOD of mar-ginotruncanids and the LOD of Ventilabrella austiniana to distinguish the base of Campanian. The bulk of the macrofossils identified from the Padeha section (Fig. 10) came from this zone: Echinocorys ex gr. scutata (low-ermost Santonian), Platyceramus cf. ahsenensis (lower Santonian), Texanites hispanicus (Upper Santonian), and Echinocorys sp. (lowermost Campanian).Chronostratigraphy. Lower Santonian – lowermost Campanian
INTEgRaTED FORaMINIFERaL aND INOCERaMID BIOsTRaTIgRaphy
Base of Coniacian
Caron (1979), Robaszynski and Caron (1979), Birkelund et al. (1984), and Marks and Stan (1983) considered that the FOD of Dicarinella primitiva is located at the basal part of the Coniacian. However, this biohorizon was later placed at the Turonian/Coniacian bound-ary (Robaszynski et al., 1990; Robaszynski and Caron, 1995). The latter authors indicated that the FODs of Di. primitiva and Di. concavata coincide in the late Turonian and are well-correlated with the FOD of the ammonite Subprionocyclus neptuni. This event was recorded close to the Middle-Upper Turonian transition (Robaszynski et al. 1990; Robaszynski and Caron, 1995). Gebhardt (2001) considered the FOD of the ammonite Prionocycloceras milticostatum as indicative for the Turonian/Coniacian boundary.
Although most planktonic foraminiferal special-ists considered the Turonian/Coniacian boundary as not traceable precisely upon planktonic foraminifera, others suggested that the FOD of Marginotruncana sinuosa can be used to define the base of Coniacian (Birkelund et al. 1984; Marks and Stan, 1983; Kauffman et al., 1996, Walaszczyk et al., 2010). Gebhardt (2001 a, b) used Muricohedbergella flandrini to recognize the Late Turonian. The LODs of Muricohedbergella simplex and Dicarinella primitiva precede the FOD of Dicarinella concavata (Koutsoukos and Bengtson, 1993).
The FOD of Cremnoceramus deformis erectus (Meek, 1811) was used to place the base of Coniacian (Wood et
al., 1984, 2004; Kauffman et al., 1996; Walaszczyk and Wood, 1999, 2000, 2008; Walaszczyk and Coban 2000; Ogg et al., 2004; Walaszczyk et al., 2010, 2012) (Fig. 11). The FOD of Cremnoceramus rotundatus (sensu Troger) (= Cremnoceramus deformis erectus), was recommend-ed for defining the Turonian/Coniacian boundary by the Coniacian Working Group (CWG) of the Cretaceous Subcomission on Stratigraphy (Kauffman et al., 1996; Walaszczyk et al., 2010). In the studied area, C. deformis erectus has not been found, but the FODs of Di. primi-tiva and Di. concavata have been recognized below the FOD of Cremnoceramus deformis deformis in the Early Coniacian, thus confirming the foraminiferal FODs latest Turonian age. Lamolda and Paul (2007) used the FOD of Costellagerina pilula as the main key planktonic fo-raminiferal horizon for the Coniacian/Santonian bound-ary in Spain. However, the present study recorded the
FOD of C. pilula to be lower than the FOD of C. de-formis deformis. Therefore, Marginotruncana paracon-cavata and Costellagerina pilula are latest Turonian in age, thus representing reliable indicators for recognizing the Turonian/Coniacian boundary in Iran.
Base of santonian
Caron (1985) and Robaszynski and Caron (1979) reported Dicarinella concavata from the Late Coniacian to Early Santonian, whereas other authors (Wonders 1980; Van Hinte, 1976; Sigal, 1977; Postuma, 1971; Pessango, 1967; Bolli, 1966) considered the Di. concavata Zone as corresponding to the Coniacian only. Consequently, the Turonian/Coniacian and Coniacian/Santonian boundaries have been placed within the D. concavata Zone of large stratigraphic volume in the studied area. The FOD of the
Fig. 11. Correlation of inoceramid zonations
35
inoceramid Cladoceramus undulatoplicatus (Roemer) was used as a global marker of the Coniacian/Santonian boundary (Gallemí et al., 2007a, b; Hampton et al., 2007; Ion et al., 1994; Kopaevich et al., 2007; Lamolda and Hancock, 1996; Lamolda and Paul, 2007; Melinte et al., 2007; Peryt and Lamolda, 2002, 2007). We have not recognized Cladoceramus undulatoplicatus. For this reason we have used the FOD of the echinoid Echinocurys ex gr. scutata as a secondary marker of the base of Santonian as proposed by Gallemí et al. (2007a, b). Using the FOD of E. ex gr. scutata, the Coniacian/Santonian boundary has been placed at the position of the sample 85 in the Padeha section. The FOD of Dicarinella asymetrica has been documented in sample 88, i.e. 9 m above the Coniacian/Santonian boundary (Fig. 8). It is evident, therefore, that the FOD of Di. asymetrica is not a reliable horizon to place the Coniacian/Santonian boundary in the studied section (Figs 1a–c and 5).
CONCLUsIONs
Planktonic foraminiferal biostratigraphic study of the Abderaz Formation in the Padeha section has been based on distribution of 54 species belonging to 15 genera. Five successive foraminiferal zones have been recognized, from the lower Turonian to the lowermost Campanian. In ascending order these are:
Whiteinella archaeocretacea (Bolli) Partial Range Zone;
Helvetoglobotruncana helvetica (Sigal) Total Range Zone;
Marginotruncana schneegansi (Dalbiez) Interval Range Zone;
Dicarinella concavata (Sigal) Interval Range Zone;
Dicarinella asymetrica (Postuma) Total Range Zone. These zones were correlated with previously docu-
mented occurrences in four sections in West and East Kopet-Dagh Basin: Hajgelichkhan, Qarehso, Abderaz, Muzduran, and Shorab, (Shafiee Ardestani et al., 2012).
Biostratigraphic data indicate that the Abderaz Formation of Padeha section is continuous and allows the recognition of three stage boundaries of the Upper Cretaceous: the Turonian/Coniacian, Coniacian/Santonian and Santonian/Campanian. Parallel to the biostratigraphy of the planktonic foraminifers, the stratigraphic distribu-tion of several macrofossil taxa has also been identified: eight inoceramid species, 2 echinoids and 1 ammonite. The FOD of the inoceramid Cremnoceramus walterdor-fensis walterdorfensis has been used to recognize the uppermost Turonian, and the FOD of Cremnoceramus deformis deformis to identify the middle part of Lower Coniacian in the Abderaz Formation. The base of the Santonian has also been recognized using the FOD of the echinoid Echinocorys ex gr. scutata. Finally, the LOD of the foramininfers of the Marginotruncanid group in addi-tion to the LOD of Ventilabrella austiniana were used to identify the base of Campanian.
Acknowledgments
This research was funded by Ferdowsi University of Mashhad, the Spanish Ministerio de Ciencia e Innovación project CGL2011-22912, co-financed by the ERDF (European Regional Development Fund), and the Aragonian Departamento de Educación y Ciencia (DGA group E05). The authors would like to express their deep gratitude to Prof. Robaszynski for his invaluable help during the study. The paper is proudly dedicated to the living memory of Prof. Hesabi.
REFERENCEs
Abdel-Kireem, M.R., Samir, A.M., Ibrahim, M.I.A. 1995. Upper Cretaceous planktonic foraminiferal zonation and correla-tion in the northern part of Western Desert, Egypt. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 198 (3), 329–361.
Afshar-Harb, A. 1969. A brief history of geological explora-A brief history of geological explora-tion and geology of the Sarakhs area and the Khangiran gas field. Bulletin of the Iranian Petroleum Institute 37, 86–96 (in Persian).
Barr, F.T. 1972. Cretaceous biostratigraphy and planktonic fo-raminifera of Lybia. Micropaleontology 18(1), 1–46.
Birkelund, T., Hancock, J.M., Hart, M.B., Rawson, P.F., Remane, J., Robaszynski, F., Schmid, F., Surlyk, F. 1984. Cretaceous stage boundaries-proposals. Bulletin of the Geological Society of Denmark 33, 3–20.
Bolli, H.M.1957. The genera Praeglobotruncana, Globotruncana, Rotalipora, Abathomphalus in the Upper Cretaceous in Trinidad. National Museum of America Bulletin 215, 55–61.
Bolli, H,M.1966. Zonation of Cretaceous to Pliocene ma-rine sediments based on planktonic foraminifera. Boletin
Informativo Asociacion Venezolana de Geologia, Minería y Petróleo 9, 3–32.
Caron, M. 1979. Cretaceous planktonic foraminifers from DSDP Leg 40, Southeastern Atlantic Ocean. Initial. Reports. Deep Sea drilling. Project 40, 651–678.
Caron, M. 1985. Cretaceous planktic foraminifera. In: Bolli, H.M., Saunders, J.B., Perch Nielsen, K. (Eds). Plankton Stratigraphy. Cambridge University Press, 17–86.
Dalbiez, F. 1955. The genus Globotruncana in Tunisia. Micropaleontology 1, 161–171.
Dimitrova, E., Valchev, B. 2007. Attempt for Upper Cretaceous planktic foraminiferal zonation of the Srednogorie and Eastern Balkan Zones (Bulgaria). Geologica Balcanica 36(1-2), 55–63.
Dochev, D. 2006. New biostratigraphic data based on inocera-mid bivalves from the interval Upper Turonian-Coniacian in Breznik, West Srednogorie Mountains, Bulgaria. International Conference “Geosciences 2006”, Bulgarian Geological Society, 61–64.
Ellis, B.F., Messina, A.R. 1999. Catalogue of foraminifera on CD ROM. American Museum of Natural History.
36
Fleury J.J. 1980. Les zones de Gavrovo-Tripolitza et du Pinde Olonos (Gréce continentale et Peloponnese du Nord). Evolution d’une plate-forme et d’un bassin dans leur cadre alpin. Societe Geologique du Nord 4, 648 pp.
Gallemí, J., López, G., Martínez, R., Pons, J.M., 2007a. Macrofauna of the Cantera de Margas section, Olazagutia: Coniacian/Santonian boundary, Navarro-Cantabrian Basin, northern Spain. Cretaceous Research 28, 5–17.
Gallemí, J., López, G., Martínez, R., Pons, J.M. 2007b. Macrofauna of the Villamartín Section: Coniacian/Santonian boundary, North Castilian Platform, Burgos, Spain. Cretaceous Research 28, 93–107.
Gebhardt, H., 2001. Inoceramids, Didymotis and ammonites from the Nkalagu Formation type locality (late Turonian to Coniacian, southern Nigeria): biostratigraphy and palae-oecologic implications. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 193–212.
Gorbachik, T.N. 1971a. On the Cretaceous foraminifera of the Crimea. Voprosy Micropaleontologii, 125–211.
Gorbachik, T.N. 1971b. A brief characteristic of Cretaceous and Paleogene deposits on the Mountain Crimea. XII European Micropaleontological Colloquium, 13–28.
Gradstein, F.M., Agterberg, F.P., Ogg, J.G., Hardenbol, J., Huang, Z. 1994. A Mezozoic Time Scale. Journal of Geophysical Research 99, p. 24.
Grandstein, F.M., Bukry, D., Hbib, D., Renz, O., Roth, P.H. 1978. Summary of DSDP Leg 44: Western North Atlantic Ocean. Initial Report Deep Sea Drill Project 44, 567–562.
Hampton, M.J., Bailey, H.W., Gallagher, L.T., Mortimore, R.N., Wood, C.J. 2007. The biostratigraphy of Seaford Head, Sussex, southern England. An international reference section for the basal boundaries for the Santonian and Campanian Stages in chalk facies. Cretaceous Research 28, 46–60.
Ion, J., Szasz, L. 1994. Biostratigraphy of the Upper Cretaceous of Romania. Cretaceous Research 15, 59– 87.
Kauffman, E.G., Kennedy, W.J. Wood, C.J. 1996. The Coniacian stage and substage boundaries. Bulletin de l’Institut Royal des Sciences naturelles de Belgique, Sciences de la Terre 66 (Supplement), 81–94.
Keller, G., Pardo, A. 2004. Paleoecology of the Cenomanian – Turonian Stratotype Section (GSSP) at Pueblo, Colorado. Marine Micropleontology 51, 95–128.
Keller, G., Stueben, D., Zsolt, B., Adatte, T. 2004. Cenomanian – Turonian sea level and salinity variations at Pueblo, Colorado. Palaeogeography, Palaeoclimatology, Palaeoecology 211, 19–43
Kopaevich, L.F., Beniamovski, V.N, Sadekov, A.Yu. 2007. Middle Coniacian – Santonian foraminiferal bioevents around the Mangyshlak Peninsula and Russian Platform. Cretaceous Research 28, 108–118.
Koutsoukos, E.A.M., Bengtson, P. 1993. Towards an integrat-ed biostratigraphy of the upper Aptian – Maastrichtian of the Sergipe Basin, Brazil. Documents de Laboratoire de Géologie de Lyon 125, 241–262.
Lamolda, M.A., Hancock, J.M. 1996. The Santonian stage and substages. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 66 (Sup-plement), 95–102.
Lamolda, M.A., Paul, C.R.C. 2007. Carbon and oxygen sta-Carbon and oxygen sta-ble isotopes across the Coniacian/Santonian boundary at Olazagutia, northern Spain. Cretaceous Research 28, 37–45.
Loeblich, A.R.Jr, Tappan, H. 1988. Foraminiferal genera and their classification. Van Nostrand Reinchold Company, New York, pp. 970.
Marks, P., Stan, N. 1983. Proposal for the recognition of bound-aries between Cretaceous stages by means of planktonic foraminiferal biostratigraphy, Bullten of the Geological
Society of Denmark 33, 163–169.Maslakova, N. I. 1971. Contribution to the systematic and phyl-
ogeny of the Globotruncanids. Voprosy Micropaleontologii 8, 102–117.
McNulty, C. L. 1976. Cretaceous foraminiferal stratigraphy, DSDP Leg 33, Holes 315A-317a. Initial Report Deep Sea Drill Project 33, 369–81.
Melinte, M. C., Lamolda, M. A. 2007. Calcareous nannofos-Calcareous nannofos-sil biostratigraphy of the Coniacian/Santonian boundary interval in Romania and comparison with other European regions. Cretaceous Research 28(1), 119–127.
Moussavi-Harami, R. Brenner, R. 1992. Geohistory analysis and petroleum reservoir characteristics of Lower Cretaceous (Neocomian) sandstones, eastern Kopet Dagh Basin, northeastern Iran. Bulletin of the American Association of Petroleum Geologists 76, 1200–1208.
Nederbragt, A.J. 1990. Biostratigraphy and paleoceano-graphic potential of the Cretaceous planktic foraminifera Heterohelicidae. PhD Thesis, Centrale Huisdrukkerij Vrije Universiteit, Amsterdam, 203 pp.
Ogg, J., Agterberg, F.P., Gradstein, F.M. 2004. The Cretaceous Period. In: Gradstein, F.M., Ogg, J., Smith, A. (Eds), Geological time scale. Cambridge University press, Cambridge, 344–383.
Peryt, D., Lamolda, M.A. 2002. Benthic foraminifers from the Coniacian/Santonian boundary interval at Olazagutia, Spain. In: Lamolda, M.A. (Comp.), Meeting on the Coniacian-Santonian Boundary, Bilbao, September 14–16, 2002. Abstracts and Field Guide Book, p. 19.
Peryt, D., Lamolda, M.A. 2007. Neoflabellinids (benthic fo-raminifers) from the Upper Coniacian and Lower Santonian at Olazagutia, Navara Province, Spain; taxonomy and cor-relation potential. Cretaceous Research 28, 30–36.
Pessagno, E.A.Jr. 1967. Upper Cretaceous planktonic foraminif-era from the Western Gulf Coastal Plain. Palaeontographica Americana 5, 259–441.
Pessagno, E.A.Jr, Longoria J.F. 1973. Shore laboratory re-port on Mesozoic planktonic foraminifera. Deep Sea Drill Project Leg 16, 16, 891–894
Postuma, J. 1971. Manual of Planktonic Foraminifera. Elsevier Publishing Company. Amsterdam, pp 420.
Premoli Silva, I., Bolli, H.M. 1973. Late Cretaceous to Eocene planktonic foraminifera and stratigraphy of Leg 15 sites in the Caribbean Sea. Initial Reports Deep Sea Drill Project 15, 499–547.
Premoli Silva, I., Sliter, W.V. 1981. Cretaceous planktonic foraminifers from the Nauru Basin, Leg 61, Site 462 Western equatorial Pacific. Initial Report Deep Sea Drill Project 61, 423–437.
Premoli Silva, I., Sliter, W.V. 1995. Cretaceous planktonic fo-raminiferal biostratigraphy and evolutionary trends from the Bottaccione section, Gubbio, Italy. Palaeontographia Italica 82, 1–89.
Premoli Silva, I., Verga, D. 2004. Practical Manual of Cretaceous Planktonic Foraminifera. In: Verga, D., Rettori, R., (Eds). International school on Planktonic Foraminifera, Universities of Perugia and Milano, Tipografia Pontefelcino, Perugia, 283 pp.
Raisossadat, N., Moussavi-Harami, R. 2000. Lithostratigraphic and facies analyses of the Sarcheshmeh Formation (Lower Cretaceous) in the eastern Kopet Dagh Basin, NE Iran. Cretaceous Research 21(4), 507–516.
Robaszynski, F., Caron, M. 1979. Atlas de foraminifères planctoniques du Crétacé moyen (Mer Boreale et Tethys), première partie. Cahiers de Micropaléontologie 1, 185 pp.
Robaszynski, F., Caron, M. 1995. Foraminifères planktoni-ques du Crétacé. Bulletin de la Societé Géologique de France 166, 681–698.
37
Robaszynski, F., Caron, M., Dupuis, C., Amedro, F., Gonzalez Donoso, J.M., Linares, D., Hardenbol, J., Gartner, S., Calandra, F., Deloffre, R. 1990. A tentative integrated stratigraphy in the Turonian of Central Tunisia: Formations, zones and sequential stratigraphy in the Kalaat Senan area. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine 14, 213–384.
Robaszynski, F., Caron, M., Gonzalez Donoso, J. M., Wonders, A. A. H. 1984. Atlas of Late Cretaceous Globotruncanids. Revue de Micropaléontologie 26, 145-305.
Salaj, J., 1980. Microbiostratigraphie du Crétacé et du Pa-léo gène de la Tunisie Septentrionale et Orientale (Hypo-stratotypes Tunisiens). Institut Géologique de Dionýz Štúr, Bratislava, 238 pp.
Salaj, J. 1997. Microbiostratigraphical (Foraminifera) division of the Turonian to Santonian in Tunisia (El-Kef and Dj. Fgnira Salah Area). Geologica Carpathica 48(3), 171–178.
Shafiee Ardestani, M., Vahidinia, M., Youssef A.M. 2011. Biostratigraphy and foraminiferal bioevents of the Abderaz Formation (middle Turonian-lower Campanian) in Kopet-Dagh sedimentary basin, northeastern Iran. Egyptian Journal of Paleontology 11, 1-16.
Shafie Ardestani, M., 2012. Biostratigraphy, biocorrelation and bio-events of the base the Coniacian and Santonian stages in Abderaz Formation at east and west of Kopet-Dagh basin, NE Iran. PhD Thesis, 569 pp. (in Persian).
Sigal, J. 1955. Notes micropaléontologiques nord-africaines. I. Du Cénomanian au Santonien: zones et limites en faciès pélagique. Comptes rendus sommaires des siences de la Société Géologique de France 8, 157–160.
Sigal, J. 1977. Essai du zonation du Crétacé méditerranéen-ne a l’aide des foraminifères planctoniques. Géologie Méditerranéenne 4, 99–108.
Sliter, W.V. 1989. Biostratigraphic zonation for Cretaceous planktonic foraminifera examined in thin section: Journal of Foraminiferal Research 19(1), 1–19.
Takashima, R., Nishi, H., Yamanaka, T., Hayashi, K., Waseda, A., Obuse, A., Tomosugi, T., Deguchi, N., Mochizuki, S. 2010. High-resolution terrestrial carbon isotope and plank-tic foraminiferal records of the Upper Cenomanian to the Lower Campanian in the Northwest Pacific. Earth and Planetary Science Letters 289, 570–582.
Van Hinte, J. E. 1976. A Cretaceous time scale. Petroleum Geology Society of America Bulletin 60, 498–516.
Vaptzarova, Y. 1976. Zonation du Crétacé supérieur du type Carpatique en Bulgarie du Nord-Ouest d’apres les Foraminifères planctoniques. Geologica Balcanica 6(3), 93–109.
Walaszczyk, I., Coban, W.A. 2004. Inoceramid fauna and bios-tratigraphy of the Middle-Upper Coniacian and Santonian of the US Western Interior. Cretaceous Research 49, 53–60.
Walaszczyk, I. Kopaevich, L F., Olferiev A. G. 2004. Inoceramid foraminiferal succession of the Turonian and Coniacian (Upper Cretaceous) of the Briasnik region (Central European Russia). Acta Geologica Polonica 54(4), 597–609.
Walaszczyk, I., Lees, J.A., Peryt, D., Cobban, W.A.,Wood, C.J. 2012. Testing the congruence of the macrofossil versus mi-crofossil record in the Turonian-Coniacian boundary suc-cession of the Wagon Mound-Springer composite section (NE New Mexico, USA). Acta Geologica Polonica 62(4), 581–594.
Walaszczyk, I., Wood, C. J. 1999. Inoceramids and biostratig-raphy at the Turonian/Coniacian boundary; based on the Salzgitter-Salder Quarry, Lower Saxony, Germany and the Słupia Nadbrzeżna section, Central Poland. Acta Geologica Polonica 48, 395–434.
Walaszczyk, I., Wood, C.J. 2000. Inoceramid bivalves at the Turonian/Coniacian boundary: biostratigraphy, events and diversity trends. Acta Geologica Polonica 50(4), 421–430.
Walaszczyk, I., Wood, C, J. 2008. Turonian-Coniacian bound-ary; definition, recognition and stratotype problems. International geological Congress, Oslo 2008, August, 6–14.
Walaszczyk, I., Wood, C. J., Lees, J. A., Peryt, D., Voigt, S., Wiese, F. 2010. The Salzgitter-Salder Quarry (Lower Saxony, Germany) and Słupia Nadbrezna river cliff section (central Poland) a proposed candidate composite Global Boundary Stratotype Section and Point for the Coniacian Stage (Upper Cretaceous). Acta Geologica Polonica 60(4), 445–477.
Wonders, A. A. 1980. Middle and late Cretaceous planktonic Foraminifera of the western Mediterranean area. Utrecht Micropaleontology Bulletin 24, 1–158.
Wood, C. J., Ernst, G., Rasemann, G. 1984. The Turonian-Coniacian stage boundary in Lower Saxony (Germany) and adjacent areas: the Salzgitter-Salder Quarry as a proposed international standard section. Bulletin of the Geological Society of Denmark 33(1–2), 225–238.
Wood, C. J., Walaszczyk, I., Mortimore, R.N., Woods, M.A. 2004. New observations on the inoceramid biostratigraphy of the higher part of the Upper Turonian and the Turonian – Coniacian boundary transition in Poland, Germany and the UK. Acta Geologica Polonica 54, 541–549.