Manganese nutrition of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit disorder incidence A. Silber a, *, A. Bar-Tal a , I. Levkovitch a , M. Bruner b , H. Yehezkel b , D. Shmuel b , S. Cohen b , E. Matan b , L. Karni c , H. Aktas c,1 , E. Turhan c,2 , B. Aloni c a Institute of Soils, Water and Environmental Sciences, Agricultural Research Organization, the Volcani Center, P.O. Box 6, Bet Dagan 50250, Israel b Besor Experimental Station, Israel c Institute of Plant Sciences, Agricultural Research Organization, the Volcani Center, P.O. Box 6, Bet Dagan 50250, Israel 1. Introduction Manganese (Mn) is an essential plant micronutrient that is involved in the activation of many enzymes in plant systems, mostly in oxidation–reduction, decarboxylation and hydrolytic reactions (Marschner, 1995). Rapid (on a scale of seconds or minutes) decrease of Mn to deficiency levels after addition of Mn(II) fertiliser to plant-growth substrate has been reported (Sonneveld and Voogt, 1997; Savvas et al., 2003; Silber et al., 2005). This might be attributed to fast adsorption reactions on negatively charged surfaces, in accordance with analogous finding in soils (Norvell and Lindsay, 1969; Benhammou et al., 2005) or soil constituents (Davies and Morgan, 1989). Recently, Silber et al. (2008) reported that Mn(II) solubility was controlled, on a time scale of seconds to a few hours after application to perlite media, by adsorption reactions, mainly onto the organic constituents accumulated from root excretions or decomposition and rhizo- sphere biota. With advancing time, biotic oxidation became the predominant mechanism of Mn(II) removal (Silber et al., 2008). Stressful conditions, such as high temperature, high light intensity, water deficit and high salinity are known to elicit oxidative stress in plants. In addition, many studies have demon- strated that reactive oxygen species (ROS) are involved in stress- induced damage to plant tissues (Hernandez et al., 1995; Burdon et al., 1996; Shalata and Tal, 1998; Shalata and Neumann, 2001). Plant tissues have the capacity to alleviate oxidative stress by Scientia Horticulturae xxx (2009) xxx–xxx ARTICLE INFO Article history: Received 18 May 2009 Received in revised form 5 July 2009 Accepted 10 August 2009 Keywords: Colour spots Heat damage Mn deficiency Nitrite toxicity ABSTRACT This research included two independent yet build up objectives: (i) to examine the effects of Mn concentration in the irrigation water on the yield and fruit quality of greenhouse-grown bell pepper exposed to heat stress; and (ii) to investigate in a laboratory experiment the role of Mn addition on oxidative and anti-oxidative components of fruit apoplast. The field experiment included four concentrations of Mn in the irrigation water (0, 0.2, 0.6 and 1.0 mg L 1 ). The effect of Mn application rate on vegetative growth and total fruit yield was insignificant and despite low Mn concentrations in zero- Mn-fed plants, no visible foliar Mn symptoms were observed. A quadratic regression was obtained between high-quality fruit yield and Mn concentration in fruit. Based on the quadratic equation, maximum high-quality yield was obtained when fruit-Mn concentration approached 44 mg kg 1 DW. The correlation also indicates that variations in high-quality yield could be explained by differences in fruit-Mn concentration. The combination of low-Mn nutrition and high air temperature which prevailed during the experiment (summer season) induced severe pale spots and blemished fruits; approximately 50% of fruits were infected in zero-Mn-fed plants. In a laboratory experiment with detached-fruit under high temperature the H 2 O 2 concentration increased whereas ascorbic acid concentration decreased in the apoplast. In Mn-fed fruit the H 2 O 2 concentration was attenuated and the activity of ascorbate oxidase was suppressed, with the result that apoplastic ascorbic acid concentration increased. In light of the known crucial role of Mn in enzyme activities and in detoxification of oxygen free-radicals, the pale spots might be related to Mn effect on oxidative stress-related apoplastic activities. The relationships between incidence of pale spots and plant-Mn concentration may support this hypothesis, but further research is needed to validate it. ß 2009 Elsevier B.V. All rights reserved. * Corresponding author. E-mail address: [email protected](A. Silber). 1 Present address: Harran University, Faculty of Agriculture, Department of Horticulture, 63040 Sanlıurfa, Turkey. 2 Present address: Canakkale, Onsekiz Mart University, Boyramic Vocational College, 17700 Boyramic/Canakkale, Turkey. G Model HORTI-3388; No of Pages 7 Please cite this article in press as: Silber, A., et al., Manganese nutrition of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit disorder incidence. Sci. Hortic. (2009), doi:10.1016/j.scienta.2009.08.005 Contents lists available at ScienceDirect Scientia Horticulturae journal homepage: www.elsevier.com/locate/scihorti 0304-4238/$ – see front matter ß 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.scienta.2009.08.005
A. Silber a,*, A. Bar-Tal a, I. Levkovitch a, M. Bruner b, H. Yehezkel b, D. Shmuel b, S. Cohen b,E. Matan b, L. Karni c, H. Aktas c,1, E. Turhan c,2, B. Aloni c
a Institute of Soils, Water and Environmental Sciences, Agricultural Research Organization, the Volcani Center, P.O. Box 6, Bet Dagan 50250, Israelb Besor Experimental Station, Israelc Institute of Plant Sciences, Agricultural Research Organization, the Volcani Center, P.O. Box 6, Bet Dagan 50250, Israel
A R T I C L E I N F O
Article history:
Received 18 May 2009
Received in revised form 5 July 2009
Accepted 10 August 2009
Keywords:
Colour spots
Heat damage
Mn deficiency
Nitrite toxicity
A B S T R A C T
This research included two independent yet build up objectives: (i) to examine the effects of Mn
concentration in the irrigation water on the yield and fruit quality of greenhouse-grown bell pepper
exposed to heat stress; and (ii) to investigate in a laboratory experiment the role of Mn addition on
oxidative and anti-oxidative components of fruit apoplast. The field experiment included four
concentrations of Mn in the irrigation water (0, 0.2, 0.6 and 1.0 mg L�1). The effect of Mn application rate
on vegetative growth and total fruit yield was insignificant and despite low Mn concentrations in zero-
Mn-fed plants, no visible foliar Mn symptoms were observed. A quadratic regression was obtained
between high-quality fruit yield and Mn concentration in fruit. Based on the quadratic equation,
maximum high-quality yield was obtained when fruit-Mn concentration approached 44 mg kg�1 DW.
The correlation also indicates that variations in high-quality yield could be explained by differences in
fruit-Mn concentration. The combination of low-Mn nutrition and high air temperature which prevailed
during the experiment (summer season) induced severe pale spots and blemished fruits; approximately
50% of fruits were infected in zero-Mn-fed plants. In a laboratory experiment with detached-fruit under
high temperature the H2O2 concentration increased whereas ascorbic acid concentration decreased in
the apoplast. In Mn-fed fruit the H2O2 concentration was attenuated and the activity of ascorbate oxidase
was suppressed, with the result that apoplastic ascorbic acid concentration increased. In light of the
known crucial role of Mn in enzyme activities and in detoxification of oxygen free-radicals, the pale spots
might be related to Mn effect on oxidative stress-related apoplastic activities. The relationships between
incidence of pale spots and plant-Mn concentration may support this hypothesis, but further research is
needed to validate it.
� 2009 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Scientia Horticulturae
journal homepage: www.e lsev ier .com/ locate /sc ihor t i
1. Introduction
Manganese (Mn) is an essential plant micronutrient that isinvolved in the activation of many enzymes in plant systems,mostly in oxidation–reduction, decarboxylation and hydrolyticreactions (Marschner, 1995). Rapid (on a scale of seconds orminutes) decrease of Mn to deficiency levels after addition ofMn(II) fertiliser to plant-growth substrate has been reported(Sonneveld and Voogt, 1997; Savvas et al., 2003; Silber et al., 2005).
* Corresponding author.
E-mail address: [email protected] (A. Silber).1 Present address: Harran University, Faculty of Agriculture, Department of
Please cite this article in press as: Silber, A., et al., Manganese nutritiodisorder incidence. Sci. Hortic. (2009), doi:10.1016/j.scienta.2009.08
0304-4238/$ – see front matter � 2009 Elsevier B.V. All rights reserved.
doi:10.1016/j.scienta.2009.08.005
This might be attributed to fast adsorption reactions on negativelycharged surfaces, in accordance with analogous finding in soils(Norvell and Lindsay, 1969; Benhammou et al., 2005) or soilconstituents (Davies and Morgan, 1989). Recently, Silber et al.(2008) reported that Mn(II) solubility was controlled, on a timescale of seconds to a few hours after application to perlite media, byadsorption reactions, mainly onto the organic constituentsaccumulated from root excretions or decomposition and rhizo-sphere biota. With advancing time, biotic oxidation became thepredominant mechanism of Mn(II) removal (Silber et al., 2008).
Stressful conditions, such as high temperature, high lightintensity, water deficit and high salinity are known to elicitoxidative stress in plants. In addition, many studies have demon-strated that reactive oxygen species (ROS) are involved in stress-induced damage to plant tissues (Hernandez et al., 1995; Burdonet al., 1996; Shalata and Tal, 1998; Shalata and Neumann, 2001).Plant tissues have the capacity to alleviate oxidative stress by
n of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit.005
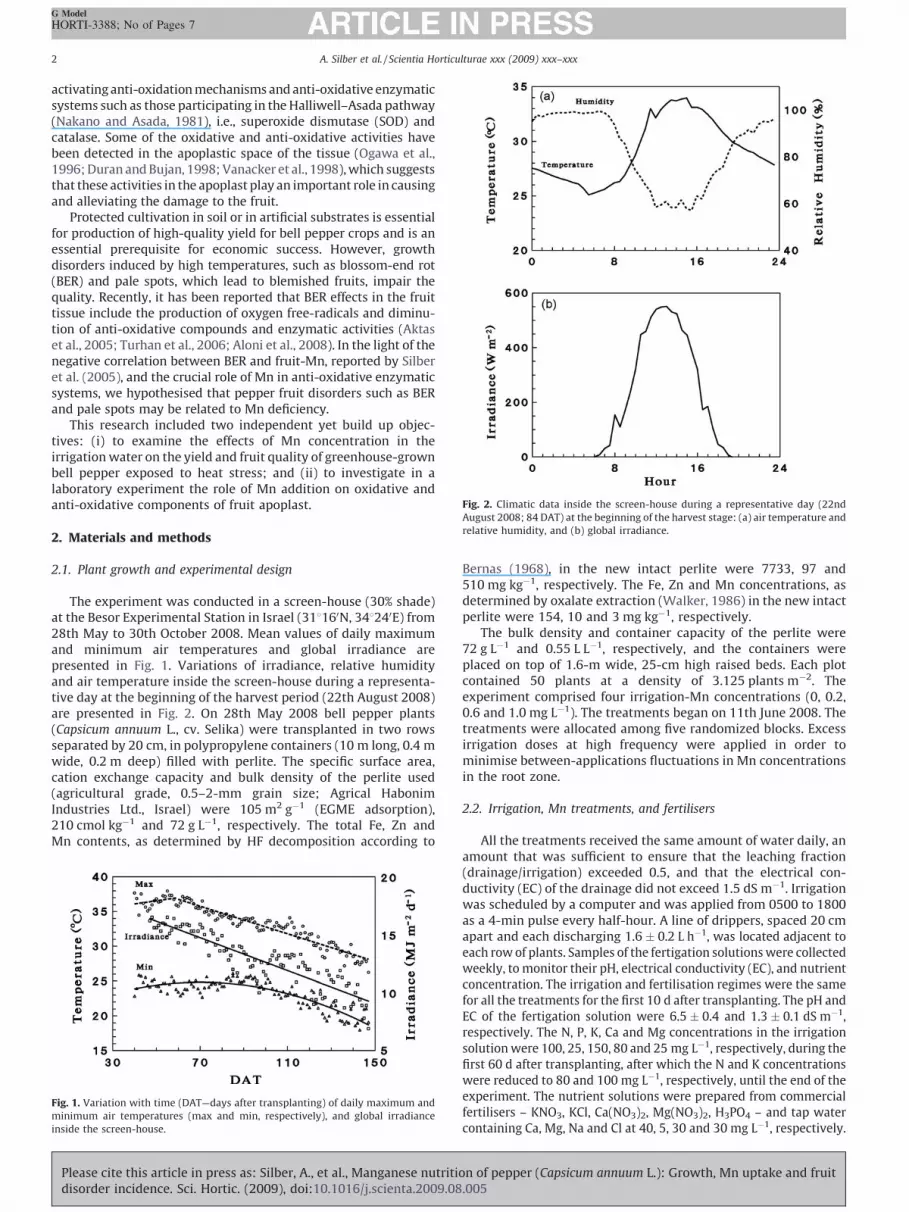
Fig. 2. Climatic data inside the screen-house during a representative day (22nd
August 2008; 84 DAT) at the beginning of the harvest stage: (a) air temperature and
relative humidity, and (b) global irradiance.
A. Silber et al. / Scientia Horticulturae xxx (2009) xxx–xxx2
G Model
HORTI-3388; No of Pages 7
activating anti-oxidation mechanisms and anti-oxidative enzymaticsystems such as those participating in the Halliwell–Asada pathway(Nakano and Asada, 1981), i.e., superoxide dismutase (SOD) andcatalase. Some of the oxidative and anti-oxidative activities havebeen detected in the apoplastic space of the tissue (Ogawa et al.,1996; Duran and Bujan, 1998; Vanacker et al., 1998), which suggeststhat these activities in the apoplast play an important role in causingand alleviating the damage to the fruit.
Protected cultivation in soil or in artificial substrates is essentialfor production of high-quality yield for bell pepper crops and is anessential prerequisite for economic success. However, growthdisorders induced by high temperatures, such as blossom-end rot(BER) and pale spots, which lead to blemished fruits, impair thequality. Recently, it has been reported that BER effects in the fruittissue include the production of oxygen free-radicals and diminu-tion of anti-oxidative compounds and enzymatic activities (Aktaset al., 2005; Turhan et al., 2006; Aloni et al., 2008). In the light of thenegative correlation between BER and fruit-Mn, reported by Silberet al. (2005), and the crucial role of Mn in anti-oxidative enzymaticsystems, we hypothesised that pepper fruit disorders such as BERand pale spots may be related to Mn deficiency.
This research included two independent yet build up objec-tives: (i) to examine the effects of Mn concentration in theirrigation water on the yield and fruit quality of greenhouse-grownbell pepper exposed to heat stress; and (ii) to investigate in alaboratory experiment the role of Mn addition on oxidative andanti-oxidative components of fruit apoplast.
2. Materials and methods
2.1. Plant growth and experimental design
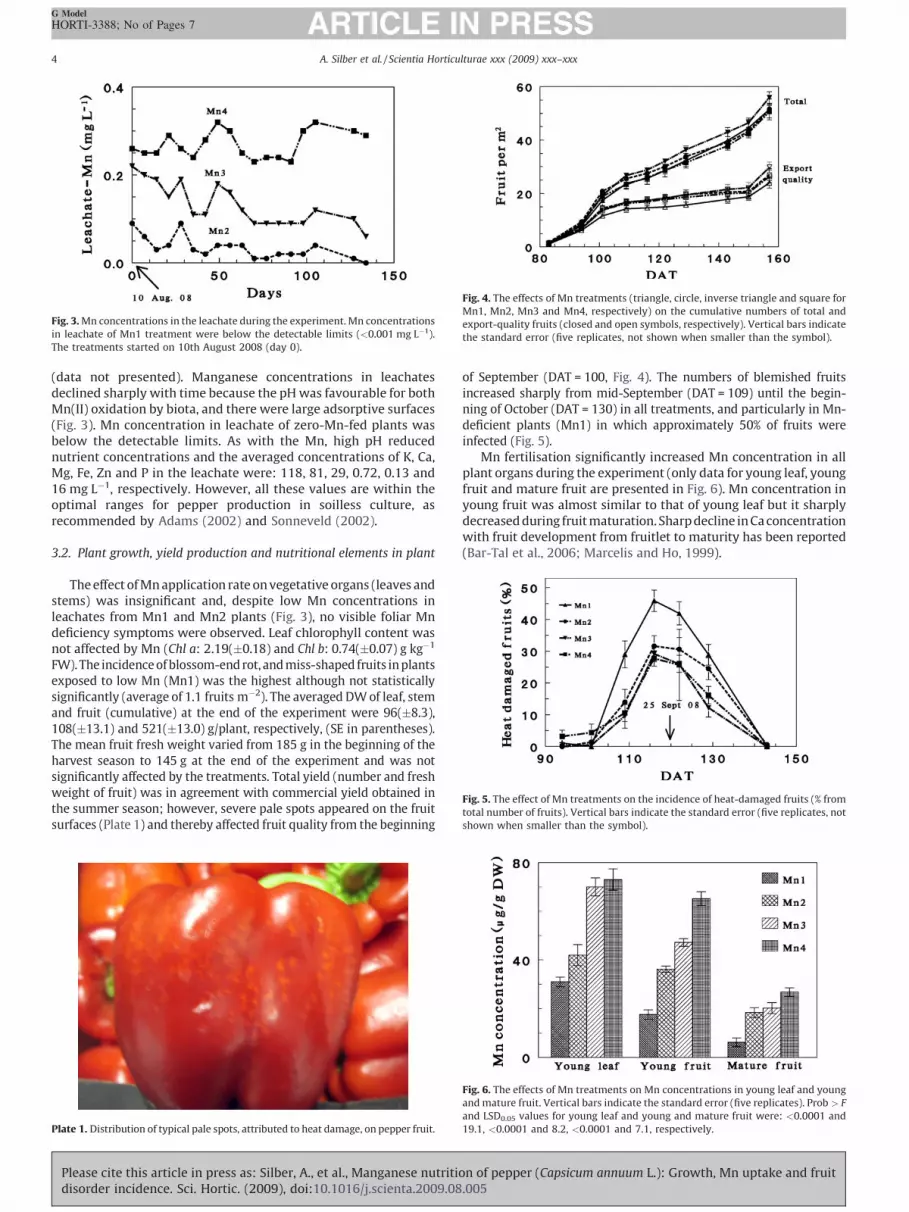
The experiment was conducted in a screen-house (30% shade)at the Besor Experimental Station in Israel (318160N, 348240E) from28th May to 30th October 2008. Mean values of daily maximumand minimum air temperatures and global irradiance arepresented in Fig. 1. Variations of irradiance, relative humidityand air temperature inside the screen-house during a representa-tive day at the beginning of the harvest period (22th August 2008)are presented in Fig. 2. On 28th May 2008 bell pepper plants(Capsicum annuum L., cv. Selika) were transplanted in two rowsseparated by 20 cm, in polypropylene containers (10 m long, 0.4 mwide, 0.2 m deep) filled with perlite. The specific surface area,cation exchange capacity and bulk density of the perlite used(agricultural grade, 0.5–2-mm grain size; Agrical HabonimIndustries Ltd., Israel) were 105 m2 g�1 (EGME adsorption),210 cmol kg�1 and 72 g L�1, respectively. The total Fe, Zn andMn contents, as determined by HF decomposition according to
Fig. 1. Variation with time (DAT—days after transplanting) of daily maximum and
minimum air temperatures (max and min, respectively), and global irradiance
inside the screen-house.
Please cite this article in press as: Silber, A., et al., Manganese nutritiodisorder incidence. Sci. Hortic. (2009), doi:10.1016/j.scienta.2009.08
Bernas (1968), in the new intact perlite were 7733, 97 and510 mg kg�1, respectively. The Fe, Zn and Mn concentrations, asdetermined by oxalate extraction (Walker, 1986) in the new intactperlite were 154, 10 and 3 mg kg�1, respectively.
The bulk density and container capacity of the perlite were72 g L�1 and 0.55 L L�1, respectively, and the containers wereplaced on top of 1.6-m wide, 25-cm high raised beds. Each plotcontained 50 plants at a density of 3.125 plants m�2. Theexperiment comprised four irrigation-Mn concentrations (0, 0.2,0.6 and 1.0 mg L�1). The treatments began on 11th June 2008. Thetreatments were allocated among five randomized blocks. Excessirrigation doses at high frequency were applied in order tominimise between-applications fluctuations in Mn concentrationsin the root zone.
2.2. Irrigation, Mn treatments, and fertilisers
All the treatments received the same amount of water daily, anamount that was sufficient to ensure that the leaching fraction(drainage/irrigation) exceeded 0.5, and that the electrical con-ductivity (EC) of the drainage did not exceed 1.5 dS m�1. Irrigationwas scheduled by a computer and was applied from 0500 to 1800as a 4-min pulse every half-hour. A line of drippers, spaced 20 cmapart and each discharging 1.6 � 0.2 L h�1, was located adjacent toeach row of plants. Samples of the fertigation solutions were collectedweekly, to monitor their pH, electrical conductivity (EC), and nutrientconcentration. The irrigation and fertilisation regimes were the samefor all the treatments for the first 10 d after transplanting. The pH andEC of the fertigation solution were 6.5 � 0.4 and 1.3 � 0.1 dS m�1,respectively. The N, P, K, Ca and Mg concentrations in the irrigationsolution were 100, 25, 150, 80 and 25 mg L�1, respectively, during thefirst 60 d after transplanting, after which the N and K concentrationswere reduced to 80 and 100 mg L�1, respectively, until the end of theexperiment. The nutrient solutions were prepared from commercialfertilisers – KNO3, KCl, Ca(NO3)2, Mg(NO3)2, H3PO4 – and tap watercontaining Ca, Mg, Na and Cl at 40, 5, 30 and 30 mg L�1, respectively.
n of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit.005
A. Silber et al. / Scientia Horticulturae xxx (2009) xxx–xxx 3
G Model
HORTI-3388; No of Pages 7
The concentration of micronutrients (Fe, Zn, Mn and Cu) in the tapwater was below the detectable limits. Micronutrients Zn, Fe, Cu, B,and Mo, all EDTA based, were applied at concentrations of 0.25, 1.0,0.02, 0.2, and 0.02 mg L�1, respectively. Leachates from eachtreatment were collected and the volumes monitored daily. TheEC, pH values, and element concentrations in the leachates wereanalyzed once a week. Element concentrations were determined asfollows: NO3-N, and P with an injector Lachat Autoanalyzer (LachatInstruments, Milwaukee, WI, USA); K with a flame photometer(M410, Sherwood Sci. Ltd., Cambridge, England); Ca, Mg, Fe, Zn andMn with an Analyst 800 atomic absorption spectrophotometer(PerkinElmer, Shelton, CT, USA).
2.3. Harvest and plant analyses
Red fruits reaching 80% red were selectively harvested weekly,from 83 d after transplanting (DAT) until the end of the experimentat 157 DAT, and total fruit numbers and weights, numbers of fruitswith physiological disorders (blossom-end rot, virus infections,deformities, flat and heat-damaged fruits) and fruit quality weredetermined. Fruit quality was classified according to a three-gradescale: (a) export (fruits with perfect shape and weighing over140 g); (b) local market (fruits with perfect shape and weighingunder 140 g); and (c) unmarketable low quality (damaged fruits orthose with physiological disorders). Top leaves from each replicatewere sampled on 9th July 2008, washed with distilled water, driedfor 1 week in a ventilated oven at 60 8C, and stored pendingchemical analysis. Two plants from each replicate were collectedon 6th August 2008 and at the end of the experiment on 30thOctober 2008; they were divided into leaves, stems and fruits,weighed immediately, washed with distilled water, dried for 1week in a ventilated oven at 60 8C, and stored pending chemicalanalysis. The dry tissue (DW) was ground to pass a 20-mesh sieve,and 100-mg samples were wet ashed with H2SO4
�–H2O2 andanalyzed for Na, K, organic-N, and P. Ashing in HClO4–HNO3 wasused to analyze for Ca, Mg, and micronutrients. Elementconcentrations were determined as mentioned above. Uptake ofMn by the vegetative organs (leaves and stems) was calculatedfrom the Mn concentration and dry weight data collected on 30thOctober 2008, and that by the fruits from the total dry weightaccumulated throughout the harvest period and the correspondinganalyses of fruit-Mn concentrations.
The chlorophyll of the sun-exposed fully expanded leaves wasextracted with N,N-dimethylformamide and determined at wavelengths of 647 and 664.5 nm with a model DMS 100 UV-visiblespectrophotometer (UVICON 933; double beam) fitted with acuvette having a path length of 1 cm. The concentrations ofchlorophyll a and b and total chlorophyll were calculated asdescribed by Inskeep and Bloom (1985).
2.4. Mn extraction from growth medium by ammonium oxalate
Samples of the top and bottom 10-cm layers of the growthmedium were collected at the end of the growth period andextracted by the ammonium oxalate (NH4Ox) method (Walker,1986) as follows: 1 g of substrate was added to a centrifuge tube,together with 40 mL of 0.2 M (NH4)2C2O4, acidified to pH 3.15 withoxalic acid. After shaking in the dark for 4 h, the supernatant wascollected, centrifuged at 10,000 rpm and filtered through a 45-mmmillipore filter.
2.5. Fruit feeding with Mn solutions
Young pepper fruits at the expansion stage, approximately 3 cmin length, were harvested with their attached peduncle. Each fruitwas fed by embedding its peduncle into a 20-mL vial filled with
Please cite this article in press as: Silber, A., et al., Manganese nutritiodisorder incidence. Sci. Hortic. (2009), doi:10.1016/j.scienta.2009.08
20 mL of either distilled water (control) or 1 mM MnSO4. Thesefruits were incubated in incubators at either 25 or 35 8C for 72 h.Humidity was approximately 90% in each incubator. Each day thevials were supplemented with the corresponding solution.Following incubation, the fruits were subjected to intercellularwashing fluid—IWF (apoplast).
2.6. Preparation of intercellular washing fluid (IWF)
Apoplastic IWF was extracted according to Fecht-Christofferset al. (2003). The pericarp tissue of the treated fruits was cut intosmall pieces, approximately 5 mm in width, and rinsed for 2 minwith tap water. Fruit pericarp samples of approximately 10 g wereimmersed in distilled water for 1 min, then vacuum infiltrated for5 min. The infiltrated tissue was gently surface dried andcentrifuged at room temperature for 1 min at 1200 � g. Theresulting wash water was collected in a plastic tube, placed inliquid N2 and kept at �80 8C pending analysis.
2.7. Determination of H2O2 and SOD activity
Determination of H2O2 was according to Ngo and Lenhoff(1980). The assay solution consisted of 1 mL of 0.1 M sodiumphosphate pH 6.5, containing 3.3 mmoles of 3-dimethylamino-benzoic acid (DMAB), 0.07 mmoles 3-methyl-2-benzothiazolinonehydrazone (MBTH) and 10 mg of horseradish peroxidase. Thereaction was initiated by adding up to 1000 mL of the IWF sample,and the absorbance was recorded as optical density (O.D.) at590 nm and compared with that of H2O2 standards. Total SODactivity in the IWF was determined by the xanthine oxidase—cytochrome C method (McCord and Fridovich, 1969).
2.8. Ascorbic acid extraction and determination
Ascorbic acid concentration in the IWF was determinedaccording to Schoner and Krause (1990). Up to 500 mL of IWFwas mixed with 500 mL of 10% metaphosphoric acid, 1 mL of Na-citrate buffer pH 2.6, and 1 mL of dichlorophenol-indophenol(DCPIP). Following incubation for 30 s, the O.D. of the solution wasdetermined at 524 nm and compared with that of standardsolutions of ascorbic acid.
2.9. Statistical analysis and calculations
Statistical analyses were carried out with the JMP software(SAS, 2006). The measured values of dry weight, yield parameters,and nutrient concentrations in leaves and fruits during theproductive stage were analyzed for treatment effects by meansof the General Linear Model Procedure of SAS. Differences amongmeans were subjected to the standard least squares mode ofANOVA, followed by Tukey’s pair-wise comparison of the means.Differences with a probability larger than 95% were taken to besignificant. Linear regression and the stepwise procedure of JMPsoftware were used to determine the relationship between dryweight production and nutrient concentrations in the leaves. Ionspeciation in leachate solutions was calculated with the MINEQL+software (Schecher and McAvoy, 2002).
3. Results
3.1. Nutrient concentration in the leachates
The pH in leachates increased immediately after the treatmenthad begun (10th August 2008) and reached an average value of 7.4as the experiment proceeded. There was no effect of Mnsupplements on the leachates pH throughout the experiment
n of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit.005
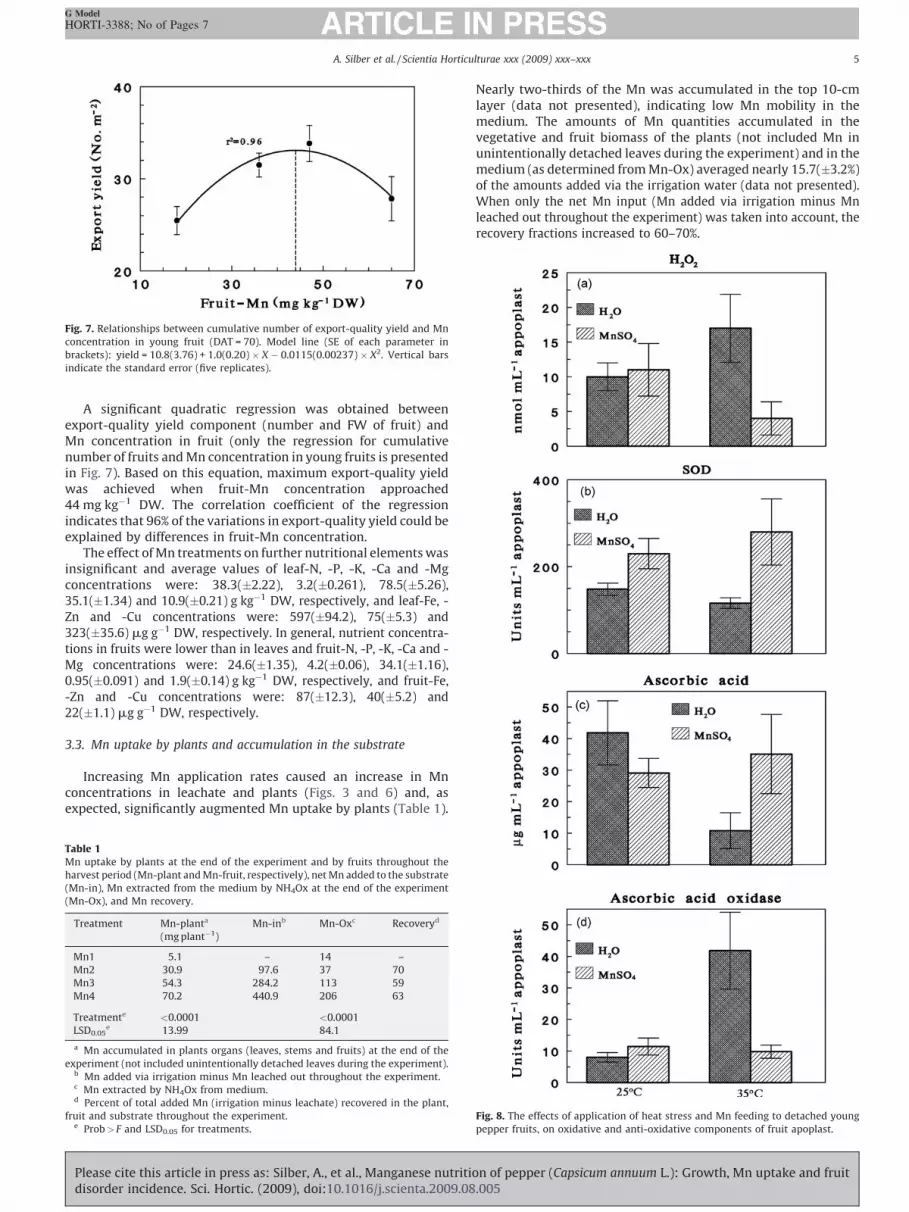
Fig. 3. Mn concentrations in the leachate during the experiment. Mn concentrations
in leachate of Mn1 treatment were below the detectable limits (<0.001 mg L�1).
The treatments started on 10th August 2008 (day 0).
Fig. 4. The effects of Mn treatments (triangle, circle, inverse triangle and square for
Mn1, Mn2, Mn3 and Mn4, respectively) on the cumulative numbers of total and
export-quality fruits (closed and open symbols, respectively). Vertical bars indicate
the standard error (five replicates, not shown when smaller than the symbol).
Fig. 5. The effect of Mn treatments on the incidence of heat-damaged fruits (% from
total number of fruits). Vertical bars indicate the standard error (five replicates, not
shown when smaller than the symbol).
A. Silber et al. / Scientia Horticulturae xxx (2009) xxx–xxx4
G Model
HORTI-3388; No of Pages 7
(data not presented). Manganese concentrations in leachatesdeclined sharply with time because the pH was favourable for bothMn(II) oxidation by biota, and there were large adsorptive surfaces(Fig. 3). Mn concentration in leachate of zero-Mn-fed plants wasbelow the detectable limits. As with the Mn, high pH reducednutrient concentrations and the averaged concentrations of K, Ca,Mg, Fe, Zn and P in the leachate were: 118, 81, 29, 0.72, 0.13 and16 mg L�1, respectively. However, all these values are within theoptimal ranges for pepper production in soilless culture, asrecommended by Adams (2002) and Sonneveld (2002).
3.2. Plant growth, yield production and nutritional elements in plant
The effect of Mn application rate on vegetative organs (leaves andstems) was insignificant and, despite low Mn concentrations inleachates from Mn1 and Mn2 plants (Fig. 3), no visible foliar Mndeficiency symptoms were observed. Leaf chlorophyll content wasnot affected by Mn (Chl a: 2.19(�0.18) and Chl b: 0.74(�0.07) g kg�1
FW). The incidence of blossom-end rot, and miss-shaped fruits in plantsexposed to low Mn (Mn1) was the highest although not statisticallysignificantly (average of 1.1 fruits m�2). The averaged DW of leaf, stemand fruit (cumulative) at the end of the experiment were 96(�8.3),108(�13.1) and 521(�13.0) g/plant, respectively, (SE in parentheses).The mean fruit fresh weight varied from 185 g in the beginning of theharvest season to 145 g at the end of the experiment and was notsignificantly affected by the treatments. Total yield (number and freshweight of fruit) was in agreement with commercial yield obtained inthe summer season; however, severe pale spots appeared on the fruitsurfaces (Plate 1) and thereby affected fruit quality from the beginning
Plate 1. Distribution of typical pale spots, attributed to heat damage, on pepper fruit.
Please cite this article in press as: Silber, A., et al., Manganese nutritiodisorder incidence. Sci. Hortic. (2009), doi:10.1016/j.scienta.2009.08
of September (DAT = 100, Fig. 4). The numbers of blemished fruitsincreased sharply from mid-September (DAT = 109) until the begin-ning of October (DAT = 130) in all treatments, and particularly in Mn-deficient plants (Mn1) in which approximately 50% of fruits wereinfected (Fig. 5).
Mn fertilisation significantly increased Mn concentration in allplant organs during the experiment (only data for young leaf, youngfruit and mature fruit are presented in Fig. 6). Mn concentration inyoung fruit was almost similar to that of young leaf but it sharplydecreased during fruit maturation. Sharp decline in Ca concentrationwith fruit development from fruitlet to maturity has been reported(Bar-Tal et al., 2006; Marcelis and Ho, 1999).
Fig. 6. The effects of Mn treatments on Mn concentrations in young leaf and young
and mature fruit. Vertical bars indicate the standard error (five replicates). Prob > F
and LSD0.05 values for young leaf and young and mature fruit were: <0.0001 and
19.1, <0.0001 and 8.2, <0.0001 and 7.1, respectively.
n of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit.005
A. Silber et al. / Scientia Horticulturae xxx (2009) xxx–xxx 5
G Model
HORTI-3388; No of Pages 7
A significant quadratic regression was obtained betweenexport-quality yield component (number and FW of fruit) andMn concentration in fruit (only the regression for cumulativenumber of fruits and Mn concentration in young fruits is presentedin Fig. 7). Based on this equation, maximum export-quality yieldwas achieved when fruit-Mn concentration approached44 mg kg�1 DW. The correlation coefficient of the regressionindicates that 96% of the variations in export-quality yield could beexplained by differences in fruit-Mn concentration.
The effect of Mn treatments on further nutritional elements wasinsignificant and average values of leaf-N, -P, -K, -Ca and -Mgconcentrations were: 38.3(�2.22), 3.2(�0.261), 78.5(�5.26),35.1(�1.34) and 10.9(�0.21) g kg�1 DW, respectively, and leaf-Fe, -Zn and -Cu concentrations were: 597(�94.2), 75(�5.3) and323(�35.6) mg g�1 DW, respectively. In general, nutrient concentra-tions in fruits were lower than in leaves and fruit-N, -P, -K, -Ca and -Mg concentrations were: 24.6(�1.35), 4.2(�0.06), 34.1(�1.16),0.95(�0.091) and 1.9(�0.14) g kg�1 DW, respectively, and fruit-Fe,-Zn and -Cu concentrations were: 87(�12.3), 40(�5.2) and22(�1.1) mg g�1 DW, respectively.
3.3. Mn uptake by plants and accumulation in the substrate
Increasing Mn application rates caused an increase in Mnconcentrations in leachate and plants (Figs. 3 and 6) and, asexpected, significantly augmented Mn uptake by plants (Table 1).
Table 1Mn uptake by plants at the end of the experiment and by fruits throughout the
harvest period (Mn-plant and Mn-fruit, respectively), net Mn added to the substrate
(Mn-in), Mn extracted from the medium by NH4Ox at the end of the experiment
(Mn-Ox), and Mn recovery.
Treatment Mn-planta
(mg plant�1)
Mn-inb Mn-Oxc Recoveryd
Mn1 5.1 – 14 –
Mn2 30.9 97.6 37 70
Mn3 54.3 284.2 113 59
Mn4 70.2 440.9 206 63
Treatmente <0.0001 <0.0001
LSD0.05e 13.99 84.1
a Mn accumulated in plants organs (leaves, stems and fruits) at the end of the
experiment (not included unintentionally detached leaves during the experiment).b Mn added via irrigation minus Mn leached out throughout the experiment.c Mn extracted by NH4Ox from medium.d Percent of total added Mn (irrigation minus leachate) recovered in the plant,
fruit and substrate throughout the experiment.e Prob> F and LSD0.05 for treatments.
Please cite this article in press as: Silber, A., et al., Manganese nutritiodisorder incidence. Sci. Hortic. (2009), doi:10.1016/j.scienta.2009.08
Nearly two-thirds of the Mn was accumulated in the top 10-cmlayer (data not presented), indicating low Mn mobility in themedium. The amounts of Mn quantities accumulated in thevegetative and fruit biomass of the plants (not included Mn inunintentionally detached leaves during the experiment) and in themedium (as determined from Mn-Ox) averaged nearly 15.7(�3.2%)of the amounts added via the irrigation water (data not presented).When only the net Mn input (Mn added via irrigation minus Mnleached out throughout the experiment) was taken into account, therecovery fractions increased to 60–70%.
Fig. 8. The effects of application of heat stress and Mn feeding to detached young
pepper fruits, on oxidative and anti-oxidative components of fruit apoplast.
n of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit.005
A. Silber et al. / Scientia Horticulturae xxx (2009) xxx–xxx6
G Model
HORTI-3388; No of Pages 7
3.4. Effect of Mn on oxidative stress-related activities
In order further study effects of Mn associated with theoccurrence of the blemish spots on pepper fruits, we exposeddetached fruits to normal or high temperatures, with or withoutfeeding with Mn solutions. Elevating the temperature in theabsence of Mn increased the H2O2 concentration in the apoplast(IWF) of the fruit, whereas it diminished the activity of SOD and theconcentration of ascorbic acid (Fig. 8a–c, respectively). Concomi-tantly, the activity of ascorbate oxidase in the apoplast wasenhanced approximately fivefold (Fig. 8d). In contrast, elevatingthe temperature of Mn-fed fruits lowered the apoplastic H2O2 leveland increased the activity of SOD (Fig. 8a and b). The apoplasticconcentrations of ascorbic acid were increased in Mn-fed, high-temperature-exposed fruits (Fig. 8c). In accordance with thechange in ascorbic acid content, the activity of apoplastic ascorbateoxidase was decreased in these fruits (Fig. 8d).
4. Discussion
4.1. Sources of Mn supply
Accumulation of Mn in the plants and in the growth media inthe zero-added Mn treatment (Mn1) reached 19.1 mg/plant(Table 1). The total and soluble Mn contents in fresh intact perlite,as determined by HF decomposition and oxalate extraction,respectively, were 510 and 3 mg kg�1, respectively (Silber et al.,2008). Considering the fact that nearly 1 kg of perlite per plant wasused in this experiment, the quantities of Mn accumulated(plants + perlite substrate) throughout this experiment were farabove the quantities of soluble Mn. Part of the accumulated Mnwas probably incorporated with the seedlings and their associatedmedium, and more might have been added as undetectableamounts associated with the fertiliser compounds. Extremelyhighly efficient mechanisms for plant Mn acquisition would berequired if these undetectable amounts had been the only source ofMn nutrition in this treatment. Alternatively, the possiblecapability of pepper roots to alter the chemical and biologicalproperties of the rhizosphere by excretion of organic compounds inorder to enhance Mn uptake should not be ignored, and meritsfurther research.
4.2. Relationships between yellow spots, air temperature and Mn
concentration
The most pronounced effect of growing peppers under thepresent experimental conditions was the appearance of yellowspots on the fruit surface (Plate 1). Occurrence of colour spots,flecks, specks and scrap on pepper fruits, with consequentdeterioration of fruit quality have been reported (Aloni et al.,1994; Jovicich et al., 2007; Savvas et al., 2008). The question ofwhether the pale spots shown in Plate 1, as well as other colourdisorders recently reviewed by Savvas et al. (2008) actuallyrepresent the same physiological disorder is of interest. Aloni et al.(1994) described colour spots in bell pepper fruit promoted byhigh-N nutrition and shading; however, that colouring looksdifferent from the pale spot in Plate 1. Jovicich et al. (2007)reported that under a low-nutrition regime incidence of yellowspots in pepper fruits increased as irrigation frequency increased.The yellow spots appeared solely in the hot season, which mayindicate a growth disorder induced by high temperature togetherwith a certain nutrient deficiency, as in the present study. The palespots that lead to blemished fruits, as presented in Plate 1, areabundant on fruits exposed to the sun during the summer and areusually attributed by growers to heat damage. However, the causeof this physiological disorder is not clear. Usually, these spots are
Please cite this article in press as: Silber, A., et al., Manganese nutritiodisorder incidence. Sci. Hortic. (2009), doi:10.1016/j.scienta.2009.08
noticeable approximately 10 d before harvest, as the fruit colouralters from green to red. In the present experiment the blemishedfruits were first noted at the beginning of September (DAT = 100;Fig. 5), so that the initiation and development of the damage mayhave occurred between 50 and 90 DAT. During this period themaximum air temperature inside the screen-house fluctuatedbetween 34 and 38 8C (Figs. 1 and 2). Temperatures of fruits locatedat the top of the plant were found to be 7–10 8C higher than theaverage air temperature (Bar-Tal et al., 2006), so the maximumtemperature of the fruits could have reached 408 or more forseveral hours per day during this period. Such extreme tempera-ture may lead to high oxidative stress (Levine, 1999), and that this,in turn, could lead to this physiological disorder.
In the light of the fundamental role of Mn in anti-oxidativesystems, the significant increases in numbers of heat-damagedfruits in Mn-deficient plants (Fig. 5) might have been caused byaccumulation of reactive oxygen species (ROSs) in the fruit.
Enhancement of ROS production by salinity stress and ROSscavenging by Mn were recently reported (Kawano et al., 2002) intobacco cell suspensions. Tachibana and Inden (1976) showed thathigh salinity repressed Mn2+ absorption in the root by competingwith Mn2+ for binding sites, and that the main competing ionscould be K+ and Ca2+. Ragab et al. (2001) demonstrated that thequality of tomato fruits grown under saline conditions wasimproved by a manganese spray. Aloni and Karni (2001) andAktas et al. (2005) suggested that occurrence of BER in peppersunder salinity might result from oxidative stress in the fruits. It is,therefore, to be expected that by reducing oxidative stress it wouldbe possible to attenuate stress-related disorders in pepper fruits. Ithas been reported that manganese elicits anti-oxidative effects inplant tissues (Archibald and Fridovich, 1982; Coassin et al., 1992;Tanaka et al., 1999). Manganese may be essential for the anti-oxidative activity of Mn-SOD, but may also serve as an anti-oxidantper se by being oxidized from Mn2+ to Mn3+ (Coassin et al., 1992).To prevent ROS-related injuries, plant cells contain anti-oxidativesystems consisting of enzymes, such as catalase, peroxidase,superoxide dismutase (SOD), and low-molecular-weight anti-oxidants such as ascorbic acid, glutathione and a-tocopherol. Someof these components are present in the apoplastic fluid (Takahamaet al., 1999; Hernandez et al., 2001), where they may play asignificant role in protection mechanisms. In the present study weshowed that apoplast-associated ascorbic acid, SOD and H2O2 mayplay important roles in controlling high temperature-relateddamage to pepper fruit. The H2O2 concentration increased underhigh temperature in the detached-fruit apoplast, and its concen-tration was attenuated by feeding with Mn. Such an Mn effect maybe due either to its own anti-oxidative properties or to itsinhibitory effect on plasma membrane-bound NADPH oxidaseactivity (Aktas et al., 2005). The other apoplastic anti-oxidantcomponent which decreased in heat-treated fruits is ascorbic acid,and this effect may be due to the enhancement of apoplasticascorbate oxidase under these conditions. In Mn-fed fruit theactivity of this enzyme was suppressed, with the result thatapoplastic ascorbic acid concentration increased (Fig. 8c). Theincrease in the apoplastic concentration of H2O2 and probably ofadditional oxygen radicals at high temperature may cause thecharacteristic pale spot. Manganese feeding might alleviate thesesymptoms by reducing oxidative stress. However, the causalrelationship between oxygen radicals, the development of fruit-blemish spots and the protective effect of Mn still requires furtherstudy.
Acknowledgments
The authors are grateful to the anonymous reviewer for criticalreadings of the article and the constructive comments. This paper
n of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit.005
A. Silber et al. / Scientia Horticulturae xxx (2009) xxx–xxx 7
G Model
HORTI-3388; No of Pages 7
is contribution no. 608/09 of the Agricultural Research Organiza-tion, the Volcani Center, Israel.
References
Adams, P., 2002. Nutritional control in hydroponics. In: Savvas, D., Passam, H.C.(Eds.), Hydroponic Production of Vegetables and Ornamentals. Embryo Pub-lications, Athens, pp. 211–262.
Aktas, H., Karni, L., Chang, D.C., Turhan, E., Bar-Tal, A., Aloni, B., 2005. The suppres-sion of salinity-associated oxygen radicals production, in pepper (Capsicumannuum) fruit, by manganese, zinc and calcium in relation to its sensitivity toblossom-end rot. Physiol. Plant. 123, 67–74.
Aloni, B., Karni, L., 2001. Biochemical changes in pepper (Capsicum annuum L.) fruitassociated with blossom-end rot. HortScience 36, 485.
Aloni, B., Karni, L., Rylski, I., Zaidman, Z., 1994. The effect of nitrogen fertilization andshading on the incidence of ‘‘colour spots’’ in sweet pepper (Capsicum annuum)fruit. J. Hortic. Sci. 69, 763–773.
Aloni, B., Karni, L., Deventurero, G., Turhan, E., Aktas, H., 2008. Changes in ascorbicacid concentration, ascorbate oxidase activity, and apoplastic pH in relation tofruit development in pepper (Capsicum annuum L.) and the occurrence ofblossom-end rot. J. Hortic. Sci. Biotechnol. 83, 100–105.
Archibald, F.A., Fridovich, I., 1982. The scavenging of superoxide radicals by man-ganous complexes: In Vitro. Arch. Biochem. Biophys. 214, 452–463.
Bar-Tal, A., Aloni, B., Arbel, A., Barak, M., Karni, L., Oserovitz, J., Hazan, A., Gantz, S.,Avidan, A., Posalski, I., Keinan, M., 2006. Effects of an evaporative cooling systemon greenhouse climate, fruit disorders and yield in bell pepper (Capsicumannuum L.). J. Hortic. Sci. Biotechnol. 81, 599–606.
Benhammou, A., Yaacoubi, A., Nibou, L., Tanouti, B., 2005. Adsorption of metal ionsonto Moroccan stevensite: kinetic and isotherm studies. J. Colloid Interface Sci.282, 320–326.
Bernas, B., 1968. A new method for decomposition and comprehensive analysis ofsilicates by atomic absorption spectrometry. Analytical Chemistry 40, 1682–1686.
Burdon, R.H., O’Kane, D., Fadzillah, N., Gill, V., Boyd, P.A., Finch, R.P., 1996. Oxidativestress and responses in Arabidopsis thaliana and Oryza sativa subjected tochilling and salinity stress. Biochem. Soc. Trans. 24, 468–472.
Coassin, M., Ursini, F., Bindoli, A., 1992. Antioxidant effect of manganese. Arch.Biochem. Biophys. 299, 330–333.
Davies, S.H.R., Morgan, J.J., 1989. Manganese (II) oxidation kinetics on metal oxidesurfaces. J. Coll. Interface Sci. 129, 63–77.
Duran, C.M.V., Bujan, C.R., 1998. Antioxidant system in the leaf apoplast compart-ment of Pinus pinaster Ait. and Pin plants exposed to SO2. Ann. Appl. Biol. 133,455–466.
Fecht-Christoffers, M.M., Maier, P., Horst, W.J., 2003. Apoplastic peroxidase andascorbate are involved in manganese toxicity and tolerance of Vigna unguicu-lata. Physiol. Plant. 117, 237–244.
Hernandez, J.A., Olmos, E., Corpas, F.J., Sevilla, F., del Rio, L.A., 1995. Salt inducedoxidative stress in chloroplasts of pea plants. Plant Sci. 105, 151–167.
Hernandez, J.A., Ferrer, M.A., Jimenez, A., Barcelo, A.R., Sevilla, F., 2001. Antioxidantsystem and O2-/H2O2 production in the apoplast of pea leaves. Its relation withsalt-induced necrotic lesions in minor veins. Plant Physiol. 127, 817–831.
Inskeep, W.P., Bloom, P.R., 1985. Extinction coefficients of chlorophyll a and b inN,N-dimethylformamide and 80% acetone. Plant Physiol. 77, 483–485.
Jovicich, E., Cantliffe, D.J., Stoffella, P.J., Haman, D.Z., 2007. Bell pepper fruit yield andquality as influenced by solar radiation-based irrigation and container media ina passively ventilated greenhouse. HortScience 42, 642–652.
Kawano, T., Kawano, N., Muto, S., Lapeyyrie, F., 2002. Retardation and inhibition ofthe cation-induced superoxide generation in BY-2 tobacco cell suspensionculture by Zn2+ and Mn2+. Physiol. Plant. 114, 395–404.
Levine, A., 1999. Oxidative stress as a regular of environmental responses in plants.In: Lerner, H.R. (Ed.), Plant Responses to Environmental Stresses, from Phyto-hormones to Genome Reorganization. CRC Press, New York, pp. 247–264.
Marcelis, L.F.M., Ho, L.C., 1999. Blossom-end rot in relation to growth rate andcalcium content in fruits of sweet pepper (Capsicum annuum). J. Exp. Bot. 50,357–363.
Please cite this article in press as: Silber, A., et al., Manganese nutritiodisorder incidence. Sci. Hortic. (2009), doi:10.1016/j.scienta.2009.08
Marschner, H., 1995. Mineral Nutrition of Higher Plants, 2nd ed. Academic Press,New York.
McCord, J.M., Fridovich, I., 1969. Superoxide dismutase. J. Biol. Chem. 244, 6049–6055.
Nakano, Y., Asada, K., 1981. Hydrogen peroxide is scavenged by ascorbate-specificperoxidase in spinach chloroplasts. Plant Cell Physiol. 22, 867–880.
Ngo, T.T., Lenhoff, H.M., 1980. A sensitive and versatile chromogenic assay forperoxidase and peroxidase-coupled reactions. Anal. Biochem. 105, 389–397.
Norvell, W.A., Lindsay, W.L., 1969. Reactions of EDTA complexes of Fe, Zn, Mn, andCu with soils. Soil Sci. Soc. Am. J. 33, 86–91.
Ogawa, K., Kanematsu, S., Asada, K., 1996. Intra- and extra-cellular localization of‘‘cytosolic’’ CuZn-superoxide dismutase in spinach leaf and hypocotyl. Plant CellPhysiol. 37, 790–799.
Ragab, M.E., Helal, R.M., Khalaf, S.M., Hafez, M.R., 2001. Improving productivity oftomato under saline conditions by proline or manganese foliar spray. Ann.Agric. Sci. 46, 773–784.
SAS Institute, 2006. JMP User’s Guide, Version 6.0. SAS Institute Inc., Cary, NC.Savvas, D., Karagianni, V., Kotsiras, A., Demopoulus, V., Karkamisi, I., Pakou, I., 2003.
Interactions between ammonium and pH of the nutrient solution supplied togerbera (Gerbera jamesonii) grown in pumice. Plant Soil 254, 393–402.
Savvas, D., Ntatsi, G., Passam, H., 2008. Plant nutrition and physiological disorders ingreenhouse grown tomato, pepper and eggplant. Eur. J. Plant Sci. Biotechnol. 2,46–61.
Schecher, W., McAvoy, D.C., 2002. MINEQL+: A Chemical Equilibrium ModelingSystem, Version 4.5 for Windows. Environmental Research Software, Hallowell,ME.
Schoner, S., Krause, G.H., 1990. Protective systems against active oxygen species inspinach: response to cold acclimation in excess light. Planta 180, 383–389.
Shalata, A., Neumann, P.M., 2001. Exogenous ascorbic acid (vitamin C) increasesresistance to salt stress and reduces lipid peroxidation. J. Exp. Bot. 52, 2207–2211.
Shalata, A., Tal, M., 1998. The effect of salt stress on lipid peroxidation andantioxidants in the leaf of the cultivated tomato and its wild salt-tolerantrelative Lycopersicun pennellii. Physiol. Plant. 104, 169–174.
Silber, A., Bruner, M., Kenig, E., et al., 2005. High fertigation frequency andphosphorus level: effects on summer-grown bell pepper growth and blos-som-end rot incidence. Plant Soil 270, 135–146.
Silber, A., Bar-Yosef, B., Levkovitch, I., Kautzky, L., Minz, D., 2008. Kinetics andmechanisms of pH-dependent Mn(II) reactions in plant-growth medium. SoilBiol. Biochem. 40, 2787–2795.
Sonneveld, C., 2002. Composition of nutrient solutions. In: Savvas, D., Passam, H.C.(Eds.), Hydroponic Production of Vegetables and Ornamentals. Embryo Pub-lications, Athens, pp. 179–210.
Sonneveld, C., Voogt, W., 1997. Effects of pH value and Mn application on yieldand nutrient absorption with rockwool grown gerbera. Acta Hortic. 450, 139–147.
Tachibana, S., Inden, T., 1976. Studies on the fertilization and cultivation of horti-cultural crops under covering conditions. IV. Effect of the concentration ofexternal solutions on the absorption and translocation of manganese by tomatoplants. J. Jpn. Soc. Hortic. Sci. 45, 153–159.
Takahama, U., Hirotsu, M., Oniki, T., 1999. Age-dependent changes in levels ofascorbic acid and chlorogenic acid, and activities of peroxidase and superoxidedismutase in the apoplast of tobacco leaves: mechanism of the oxidation ofchlorogenic acid in the apoplast. Plant Cell Physiol. 40, 716–724.
Tanaka, Y., Hibino, T., Hayashi, Y., Tanaka, A., Kishitani, S., Takabe, T., Yokota, T.,Yokata, S., Takabe, T., 1999. Salt tolerance of transgenic rice overexpressingyeast mitochondrial Mn-SOD in chloroplasts. Plant Sci. 148, 131–138.
Turhan, E., Karni, L., Aktas, H., Deventurero, G., Chang, D.C., Bar-Tal, A., Aloni, B.,2006. Apoplastic anti-oxidants in pepper (Capsicum annuum L.) fruit and theirrelationship to blossom-end rot. J. Hortic. Sci. Biotechnol. 81, 661–667.
Vanacker, H., Harbinson, J., Ruisch, J., Carver, T.L.W., Foyer, C.H., Asard, H., Navas, P.,1998. Plasma membrane redox systems and their role in biological stress anddisease. Protoplasma 205, 129–140.
Walker, A., 1986. Oxalate-extractable iron and aluminium. In: Singer, M.J., Janitzky,P. (Eds.), Field and Laboratory Procedures Used in a Soil Chronosequence Study,vol. 1648. US Geological Survey, Bull., pp. 41–42.
n of pepper (Capsicum annuum L.): Growth, Mn uptake and fruit.005