This article was originally published in the The Desk Encyclopedia of Microbiology published by Elsevier, and the attached copy is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who you know, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at: http://www.elsevier.com/locate/permissionusematerial B Jagannathan and J H Golbeck. Photosynthesis: Microbial. The Desk Encyclopedia of Microbiology. (Moselio Schaechter, Editor), pp. 844-860, Oxford: Elsevier.
Transcript
This article was originally published in the The Desk Encyclopedia of Microbiology published by Elsevier, and the attached copy is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who
you know, and providing a copy to your institution’s administrator.
All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or
institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at:
B Jagannathan and J H Golbeck. Photosynthesis: Microbial. The Desk Encyclopedia of
Microbiology. (Moselio Schaechter, Editor), pp. 844-860, Oxford: Elsevier.
Author's personal copy
Photosynthesis: MicrobialB Jagannathan and J H Golbeck, The Pennsylvania State University, University Park, PA, USA
ª 2009 Elsevier Inc. All rights reserved.
Defining Statement
Introduction
Historical Perspective
Classification of Photosynthetic Organisms
The Constituent Processes of Photosynthesis
Absorption and Transfer of Light Energy
The Water-Splitting Complex
844
Transmembrane Electron Transfer
Calvin–Benson–Bassham Cycle
Anoxygenic Photosynthesis
The Evolution of Photosynthesis
Future Research Directions
Further Reading
Glossaryanoxygenic photosynthesis A type of photosynthesis
performed by bacteria that do not use water as the
electron donor for carbon fixation, and hence do not
liberate oxygen.
antenna A network of closely spaced pigment
molecules that captures light energy and transfers it
efficiently to the reaction center.
chlorosome An extensive antenna system consisting
of bacteriochlorophyll c, d, and e found in green sulfur
bacteria.
iron–sulfur cluster A redox cofactor consisting of
covalently linked iron and sulfur atoms that is involved in
one-electron transfers.
reaction center A membrane-embedded pigment–
protein complex responsible for transferring electrons
across the photosynthetic membrane.
thylakoid A membranous structure wherein all the
light-dependent processes of photosynthesis occur.
water-splitting complex A cluster of four manganese
atoms and one calcium atom that oxidatively splits
water, producing electrons for carbon fixation.
Abbreviations
3PG 3-phosphoglycerate
CBB
Calvin–Benson–Bassham
Chl
Chlorophyll
DHAP
dihydroxyacetone phosphate
EXAFS
extended x-ray absorption fine structure
F6P
fructose-6-phosphate
FBP
fructose-1,6-biphosphate
FNR
ferredoxin-NADPþ oxidoreductase
G1P
glucose-1-phosphate
G3P
glyceraldehyde-3-phosphate
NADPH
nicotinamide adenine dinucleotide
phosphate
PS
Photosystem
RuBP
ribulose-1,5-biphosphate
Defining Statement
The aim of this article is to move beyond the textbookequation of photosynthesis and describe the design prin-ciples behind photosynthetic electron transfer. Theevents that constitute a photosynthetic cycle aredescribed in the exact order they occur, using the cyano-bacterial system as a model. Photosynthesis in lesser-known phototrophs is also discussed.
Introduction
Photosynthesis is the biochemical process carried out bycertain bacteria, algae, and higher plants in which light is
converted into chemical bond energy. The process iscrucial, since nearly all life on earth depends on sunlighteither directly or indirectly for energy, food, and O2. Theadvent of photosynthetic prokaryotes with the ability toconsume CO2 and produce O2 from H2O resulted in ahospitable environment on earth for advanced forms oflife. Fossil records indicate that the first oxygenic photo-synthetic bacteria appeared around 3.5� 109 years ago.Earlier, organisms survived by anaerobic metabolism, aprocess that generates only a fraction of the energy pro-duced by aerobic metabolism. It is likely that in theabsence of oxygenic photosynthesis, advanced forms oflife would not have emerged and only microorganismswould now exist. Today, as the primary means of carbonfixation, oxygenic photosynthesis forms one half of the
Author's personal copyPhotosynthesis: Microbial 845
energy-carbon cycle. Phototrophic organisms reduceCO2 to carbohydrates, which are oxidized back to CO2
by heterotrophic (as well as phototrophic) organisms. Theenergy released during the oxidation reaction is stored inthe form of NADH and ATP, which are subsequentlyused for growth, metabolism, and reproduction. In addi-tion, prehistoric plants and algae were largely responsiblefor the generation of the vast reserves of fossil fuels thatare now being mined for their energy value. They pro-vided a large portion of the initial biomass, which wasconverted into oil and coal over millions of years throughpressure, heat, and microbial action.
The general process of photosynthesis is described byVan Niel’s equation:
2H2Aþ CO2 ! 2Aþ CH2OþH2O ½1�
where H2A is the reductant and A is the oxidized product.Van Niel’s equation can be applied to oxygenic photo-
synthesis as:
6CO2 þ 6H2Oþ light! C6H12O6 þ 6O2 ½2�
Although complete, this equation belies the overwhelm-ing complexity of the process. For example, thegeneration of the light-induced charge-separated stateand its subsequent stabilization over time requires alarge number of pigments and cofactors arranged in aspecific protein environment. The splitting of H2O intoO2 is extremely difficult to replicate in the laboratory, yetplants and cyanobacteria perform the task repeatedly withseeming ease. The conversion of CO2 into sugars isanother intricate process that requires an extensive setof physical and chemical reactions to occur in a highlycoordinated fashion.
In this article, we will expand on this simple equation.In addition to describing the general design principlesbehind the sophisticated biomachinery involved in photo-synthesis, we will provide structural and functionaldetails, placing special emphasis on light-induced elec-tron transfer in aerobic and anaerobic organisms.
Historical Perspective
The first experiments on photosynthetic organisms wereperformed in the 1770s when Joseph Priestley showed thatplants were capable of generating a gas that could supportcombustion. Building on his work, Jan Ingenhousz estab-lished that sunlight was required, and Jean Senebier andNicolas Theodore de Saussure demonstrated the indispen-sability of CO2 and H2O. In 1845, Julius Robert von Meyerpostulated that plants convert light into chemical energyduring photosynthesis. Early scientists believed that the O2
was produced from the splitting of CO2, and it was notuntil the 1930s that Cornelius van Niel proposed, correctly,
that H2O was the source of O2. It is interesting that 75 yearslater the exact biochemical mechanism of H2O splittingremains to be elucidated.
Photosynthesis research has had its share of Nobellaureates. Melvin Calvin won the chemistry prize in1961 for identifying most of the intermediates in theconversion of CO2 into carbohydrates. Peter Mitchellwas the sole recipient of the chemistry award in 1978for his work on the chemi-osmotic theory of protontranslocation. Johann Deisenhofer, Robert Huber, andHartmut Michel won the chemistry prize in 1988 forsolving the first crystal structure of a photosyntheticreaction center. Rudolph Marcus’s investigation of thefactors guiding electron transfer in chemical systemsremains the paradigm for theoretical calculations of elec-tron transfer in photosynthetic reaction centers. He wasawarded the Nobel Prize in chemistry in 1992. Morerecently, Paul Boyer and John Walker were awarded theprize in chemistry in 1997 for elucidating the enzymaticmechanism underlying the synthesis of ATP.
Artificial photosynthesis has seen a recent spurt ofactivity, largely due to an increased awareness of thedepletion of fossil fuel reserves and the effect of theircombustion products on the earth’s climate. The goal isto synthesize inexpensive and long-lasting organic andinorganic molecules that convert light into chemicalenergy, thereby mimicking the basic process of photo-synthesis. This has brought new disciplines such asmaterial science and bioengineering into photosynthesis,making the field truly interdisciplinary.
Classification of PhotosyntheticOrganisms
There exist five bacterial phyla with members capable ofchlorophyll-based phototrophy: Firmicutes, Chloroflexi,Chlorobi, Proteobacteria, and Cyanobacteria. With therecent discovery of Chloracidobacterium thermophilum,Acidobacteria have become the sixth known phylum tocarry out the process of photosynthesis.
All photosynthetic organisms can be classified as eitheroxygenic or anoxygenic. Oxygenic phototrophs employH2O as the source of electrons and liberate O2 as the by-product. Anoxygenic phototrophs derive their electronsfrom organic or inorganic molecules, and hence they donot evolve O2. Of the five well-established phototrophicbacterial phyla, only the Cyanobacteria are capable ofperforming oxygenic photosynthesis. In addition, alleukaryotic phototrophs such as higher plants and algae,which evolved later than cyanobacteria, produce O2 dur-ing photosynthesis.
The remaining four phyla include anaerobes such asthe purple nonsulfur bacteria, purple sulfur bacteria,green sulfur bacteria, and heliobacteria, which survive
Author's personal copy846 Photosynthesis: Microbial
only under low concentrations of O2. The recently dis-covered Acidobacteria have been reported to live underoxic conditions, although a detailed physiological char-acterization of this organism remains to be carried out.
We will discuss oxygenic photosynthesis first, usingcyanobacteria as the model organism. Cyanobacteria arephotosynthetic prokaryotes that are found in every con-ceivable habitat from oceans to fresh water to soil. TheseGram-negative bacteria are responsible for generating themajority of the O2 in the earth’s atmosphere. The mostwidely used cyanobacterial strains for current experimen-tal research are Synechocystis sp. PCC 6803, Synechococcus
sp. PCC 7002, and Thermosynechococcus elongatus.In cyanobacteria, photosynthesis is associated with a
well-organized system of internal membranes in the cyto-plasm. These are called thylakoids, from the Greek wordthylakos meaning sac. These membranes are highly folded,allowing the cell to pack a large amount of surface areainto a small space. The interior space enclosed by thethylakoid membrane is termed the lumen and the matrixsurrounding the thylakoids is termed the stroma. Thethylakoids are home to the integral membrane proteincomplexes that are involved in the light reactions ofphotosynthesis.
Eukaryotic organisms such as higher plants and algaeconduct photosynthesis in membrane-bound organellescalled chloroplasts. They consist of an outer, freely perme-able membrane and a selectively permeable innermembrane that encloses the stroma. The sac-like thyla-koids immersed in the stroma are similar in organization tothe comparable membranes in cyanobacteria. Chloroplastthylakoids, however, tend to form well-defined stackscalled grana, which are connected to other stacks byintergrana thylakoids called lamellae. It is widely thoughtthat chloroplasts evolved from an endosymbiotic relation-ship of a heterotrophic prokaryote with a cyanobacterium.
The Constituent Processes ofPhotosynthesis
Eqn [1] is the end product of a large number of events thatoccur during a typical photosynthetic cycle. The basicprocesses that constitute oxygenic photosynthesis are:
• Absorption of light by pigment molecules and transferof the excitation energy to two reaction centers,Photosystem II (PS II) and Photosystem I (PS I).
• Light-induced transfer of an electron across the photo-synthetic membrane and splitting of H2O into O2 byPS II.
• Light-induced excitation and transfer of an electronacross the photosynthetic membrane, generating redu-cing equivalents in the form of nicotinamide adeninedinucleotide phosphate (NADPH) by PS I.
• Production of ATP using the proton gradient gener-ated across the membrane from both H2O splitting andelectron transfer through the cytochrome b6f complex.
• Conversion of CO2 into carbohydrates using ATP andthe reducing power of NADPH.
The division of photosynthetic labor is relatively straight-forward. All the light reactions occur within or on thethylakoid membrane. The ATP and NADPH producedby the light reactions are released into the stroma wherethe dark reactions of CO2 fixation are carried out. Wefocus first on the overall design philosophy of the processof converting light to stable chemical energy.
Absorption and Transfer of Light Energy
The Light-Absorbing Chromophores
Photosynthesis in cyanobacteria and plants is driven bylight in the visible (380–750 nm) region of the electromag-netic spectrum. Phototrophic organisms such as purplebacteria, green sulfur bacteria, and heliobacteria extendthis region to the near-infrared so as to exploit uniqueecological niches. All of this makes evolutionary sense asthe majority of the sun’s energy that reaches the earth’ssurface lies in this range. Ultraviolet radiation and far-infrared radiation are both limited in amount; also, theformer is too energetic and is capable of breaking chemicalbonds, while the latter contains insufficient energy to beuseful for most photochemical processes.
Primary chromophores
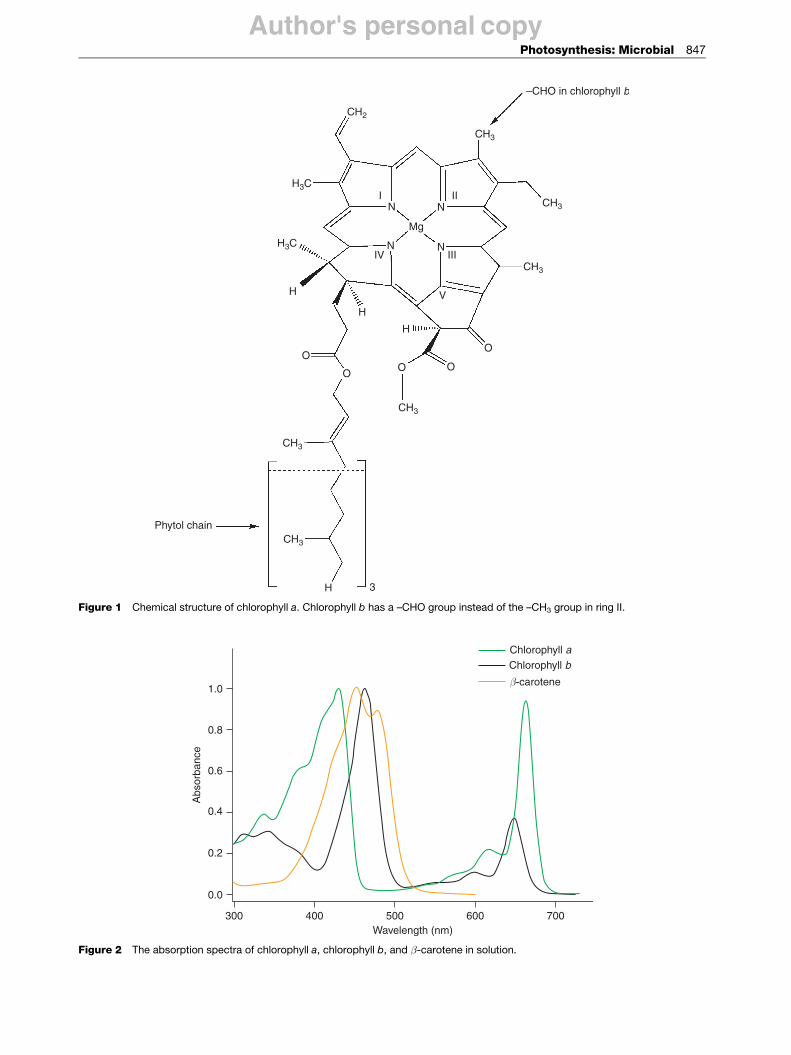
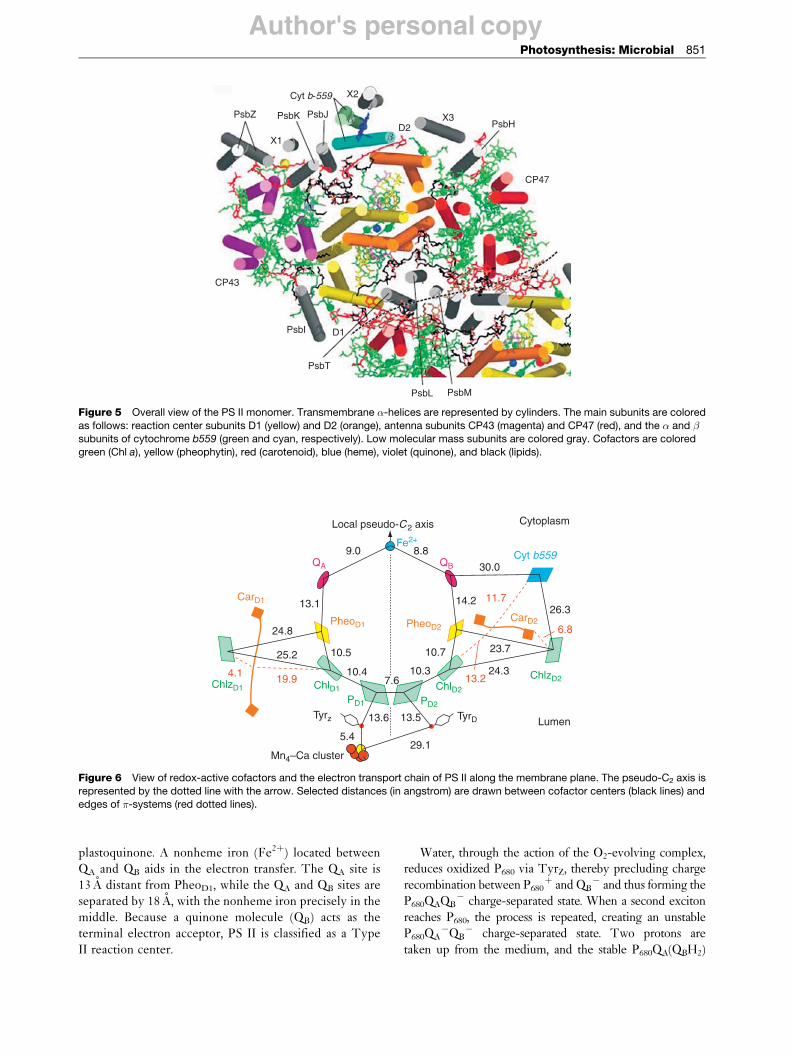
Photosynthetic organisms use a range of chromophores toefficiently capture photons in the visible and near-IRregions. The most abundant chromophore involved inphotosynthesis is chlorophyll, a molecule structurallysimilar to, and produced by, the same metabolic pathwayas porphyrin pigments such as heme. The basic structureof the chlorophyll molecule is a chlorin ring coordinatedto a central magnesium atom (Figure 1). The addition ofa long phytol tail makes chlorophyll insoluble in water.There are four common types of chlorophyll molecules inphotosynthetic organisms, named chlorophyll a, b, c, andd. Their overall structure is similar, with minor changes inthe side-chain groups that result in slightly differentabsorption spectra (Figure 2). Cyanobacteria employchlorophyll a, while plants utilize both chlorophyll a
and b. Some species of algae contain chlorophyll c, and afew species of cyanobacteria contain chlorophyll d.Chlorophylls absorb primarily in the blue and red regionsof the visible spectrum and have a high molar extinctioncoefficient. They have an inherently high fluorescenceyield, which guarantees a long-lived excited singlet state,making them the ideal chromophore.
Author's personal copy
CH2
CH3
–CHO in chlorophyll b
CH3
CH3
CH3
CH3
CH3
Phytol chain
H3C
N
Mg
H
H
H 3
H
OO
OO
O
N N
NI II
IIIIV
V
H3C
Figure 1 Chemical structure of chlorophyll a. Chlorophyll b has a –CHO group instead of the –CH3 group in ring II.
1.0
0.8
0.6
0.4
0.2
0.0
400300 500Wavelength (nm)
Abs
orba
nce
600 700
β-carotene
Chlorophyll bChlorophyll a
Figure 2 The absorption spectra of chlorophyll a, chlorophyll b, and �-carotene in solution.
Photosynthesis: Microbial 847
Author's personal copy848 Photosynthesis: Microbial
Accessory chromophoresBesides containing chlorophylls, photosynthetic organ-isms contain accessory pigments that extend the rangeof absorbed wavelengths. Carotenoids are the main acces-sory pigment found in cyanobacteria, algae, and higherplants. They belong to the tetraterpenoid family, that is,contain 40 carbon atoms, and absorb light in the400–500 nm region. Structurally, these compounds arecomposed of two small six-carbon rings connected by apolyene chain of carbon atoms. They are insoluble inwater and are normally attached to proteins that areattached to the membrane. There are over 600 types ofcarotenoids, which are classified as either carotenes orxanthophylls. Carotenes consist exclusively of carbonand hydrogen, while xanthophylls also contain oxygen.The most abundant carotenoid in cyanobacteria is �-carotene, which is the same pigment that gives carrotsits distinctive color (Figure 3). In addition to functioningas an accessory pigment, carotenoids play a vital role indissipating excess light energy, which would otherwiselead to the generation of superoxide radicals. These radi-cals are highly reactive to chemical bonds and could bepotentially lethal to the cell if left unchecked.
Cyanobacteria and certain types of algae contain addi-tional pigments called phycobilins, which absorb lightbetween 500 and 650 nm. Phycobilins consist of an openchain of four pyrrole rings and are water-soluble. Theyare attached to proteins termed phycobiliproteins andthey pass on the absorbed light energy to nearby antennachlorophyll molecules.
Plants and cyanobacteria therefore use a combinationof chlorophylls and accessory pigments to effectivelyblanket a large majority of the visible spectrum. Bothappear dark green or blue-green because the few photonsthat are not absorbed lie between the blue and red regionsof the spectrum.
The Light-Gathering Structures and ResonanceEnergy Transfer
The task of the photosynthetic reaction center is to con-vert the energy stored in the excited singlet state ofchlorophyll to a form useful for work. In photosynthesis,work refers to the creation of a charge-separated stateconsisting of a donor, Dþ, and an acceptor, A�, pair. At
CH3 CH3CH3
CH3
CH3
Figure 3 Chemical structure of �-carotene.
one extreme of time, the creation of the singlet excitedstate occurs within 10�15 s of absorbing a photon. At theother extreme, the captured light energy must be utilizedwithin 10�8 s, otherwise the energy will be lost as heat orfluorescence as the excited state decays. The generationof the charge-separated state must occur within this win-dow of time.
A network of closely spaced chlorophyll molecules,termed the antenna system, absorbs the photon and theresulting excited state migrates to a neighboring antennachlorophyll by a process known as resonance energytransfer. This occurs on a timescale of 10�12 s and is anonradiative process. The excited state, known as anexciton, randomly wanders about the antenna systemuntil it chances upon the specialized reaction centerchlorophylls associated with PS I and PS II. The energylevels of these specialized chlorophylls are slightly lowerthan the antenna chlorophylls because they are in a dif-ferent protein environment. This allows these specializedchlorophylls to trap the exciton and use it to create acharge-separated state. In most photosynthetic reactioncenters, this state is generated within 10�10 s followingphoton absorption. Accessory pigments also transmit theabsorbed energy to antenna chlorophylls by a similarprocess of resonance energy transfer.
The Water-Splitting Complex
We now turn our attention to the source of electrons inoxygenic photosynthesis. The catalytic redox center thatcarries out H2O splitting is termed the O2-evolving com-plex, and is an integral component of PS II. The water-splitting reaction can be summarized by the followingequation:
2H2O! O2 þ 4e – þ 4Hþ ½3�
The 3.0 A X-ray crystal structure of PS II from T. elongatus
(PDB ID 2AXT), as well as extended X-ray absorptionfine structure (EXAFS) studies on PS II crystals, has led toa structural model of the O2-evolving complex(Figure 4). This structure is the starting point for discus-sion on the mechanism of O2 evolution.
A cluster of four manganese atoms and a calcium atomis responsible for stripping four electrons from two H2O
CH3 CH3CH3H3C
H3C
Author's personal copy
Tyr 161
Asp 170
Glu 333
His 332
His 337Glu 354
Asp 342
Ala 344
Gln 165
His 190
Glu 189
CP43
MnA
MnC
MnBMnD Ca
Figure 4 Proposed structure of the water-splitting complex ofPS II, based on the 3.0 A resolution X-ray crystal structure and on
extended X-ray absorption fine structure (EXAFS) data. The
spheres represent manganese (red), calcium (green), and the
bridging oxygen ligand atoms (gray).
Photosynthesis: Microbial 849
molecules, liberating O2 in the process. The protons arereleased into the thylakoid lumen, thereby generating aportion of the pH gradient that is used to synthesize ATP.
The structural model of PS II shows the metal atomsarranged in an extended cubane structure, with three man-ganese and one calcium at the corners, and the fourthmanganese located immediately to one side. The metalatoms are connected to each other by mono-�-oxo, di-�-oxo, and/or hydroxo bridges, but the amino acids that con-tribute the ligands are not known with complete certainty.
Since 1970, the paradigm for understanding O2 evolu-tion has been the S-state cycle proposed by Bessel Kok.This model includes five oxidation states (S-states) for the4Mn–Ca cluster. The cluster is oxidized in one-electronsteps from S0 (most reduced) to S4 (most oxidized) by asuccessively photooxidized reaction center chlorophyll inPS II. The 4Mn–Ca cluster thus accumulates four equiva-lents of oxidizing power and uses it to split two H2Omolecules. An O2 molecule is released after the S4 state,returning the 4Mn–Ca cluster to the S0 state.
A large number of questions remain unsolved concern-ing the catalytic mechanism of water splitting. Mostimportantly, the S4 state of the cycle is fleeting and hasnot been observed spectroscopically. This state is criticalbecause it may be the starting point for O–O bond forma-tion. Without knowledge of its chemistry, the precisecatalytic mechanism of the water-splitting complex is dif-ficult to formulate. The binding site of the two H2Omolecules represents another uncertainty, although arecent proposal suggests that the calcium coordinates oneof the H2O molecules, with a manganese binding the other.
The electrons from the water-splitting complex aredonated to an oxidized tyrosine residue termed TyrZ.
According to the X-ray crystal structure, the calciumatom is positioned between the 4Mn–Ca cluster andTyrZ. A chloride ion is bound near the water-splittingcomplex in the vicinity of TyrZ. It is believed that its rolein O2 evolution is to neutralize accumulated charge.
Transmembrane Electron Transfer
Stabilization of the Charge-Separated State
The electrons obtained from the splitting of H2O are ulti-mately used to reduce CO2. To achieve this goal, theelectrons must be transferred against a highly unfavorablethermodynamic gradient from the lumenal side of the mem-brane to the stromal side, where the dark process of carbonfixation occurs. The electrons traverse a distance equal to thethickness of the membrane, which is around 40 A. Thisdistance appears short, but it is significant on the scale of anelectron.
Consider a hypothetical donor–acceptor pair, D–A,where D becomes excited to D� and donates an electronto A, thereby creating a charge-separated state Dþ–A�:
D–A! D�–A! Dþ–A –
Due to their close proximity, the Dþ–A� charge-separated state is unstable and short-lived. Adding a sec-ond closely spaced acceptor molecule, A1, can extend thelifetime of the charge-separated state:
D–A–A1 ! D�–A–A1 ! Dþ–A – –A1 ! Dþ–A–A –1
The driving force for electron transfer is a drop in Gibbs freeenergy between A� and A1, thus altering the equilibriumconstant in favor of A1. This results in the expenditure ofsome of the original energy of the photon, but it is anecessary trade-off to extend the lifetime of the charge-separated state.
Adding a second closely spaced donor molecule, D1,has a similar effect:
D1–D–A! D1–D�–A! D1–Dþ–A –!D þ1 –D–A –
The positive charge will migrate from Dþ to D1 due to adrop in Gibbs free energy between D1 and D, thus alter-ing the equilibrium constant in favor of D.
Photosynthetic complexes adopt both strategies andtrade off some of the energy of the original photon toextend the lifetime of the otherwise short-lived charge-separated state.
Factors Affecting the Rate of ElectronTransfer – Marcus Theory
To further understand this process, we must delve furtherinto the factors that govern the rate of electron transfer inproteins. Marcus theory states that the rate of electron
Author's personal copy850 Photosynthesis: Microbial
transfer between a donor and an acceptor pair depends ontwo factors. The first is the Frank Condon factor, whichincludes the change in the Gibbs free energy, the reorga-nization energy, and the temperature. Mathematically, itis expressed as:
ket _ exp–�G0 þ �ð Þ
4�kBT
2
½4�
where ket is the rate of electron transfer, �G0 is the differencein free energy between the product and the reactant, � is thereorganization energy, kB is the Boltzmann’s constant, and T
is the absolute temperature. This is the equation of a para-bola; as �G increases, the rate first increases, then attains amaximum, and finally decreases. The difference in Gibbsfree energy between the product (Dþ–A�) and the reactant(D–A) translates into the thermodynamic driving force forthe reaction. The reorganization energy corresponds to theamount of energy required to alter the microenvironment ofthe reactants before electron transfer so that it resembles theequilibrium microenvironment of the products after electrontransfer. This term reflects small changes in bond lengths andreorientation of dipoles around the redox centers to reflectthe new pattern of electric fields before and after the electrontransfer event. It is difficult to measure experimentally and isusually assumed to be a constant 0.7 eV in proteins.
The second factor that influences the rate of electrontransfer is the matrix coupling element. It relates electrontransfer rate to the distance between the donor and theacceptor pairs. The following equation describes therelationship:
HABj j2 _ exp – �Rð Þ ½5�
where HABj j is the coupling probability between the donorand acceptor wave functions, � is a constant whose value is
1.4 A�1 in proteins, and R is the edge-to-edge distancebetween the donor and the acceptor molecules.
In proteins, this logarithmic relationship translates to atenfold change in the rate of electron transfer for every 1.7 Achange in distance. Accordingly, an electron would takenearly a century to traverse a distance of 45 A. In bioener-getic membranes, large distances are traversed byintroducing several cofactors into the membrane so as toshorten the distance between any two redox pairs. In photo-synthetic systems, an electron traverses the width of thethylakoid membrane in less than 1ms due to the presenceof multiple electron transfer cofactors. With detailed knowl-edge of the factors that affect electron transfer in proteins, wenow turn to how nature has incorporated these concepts intothe design of a photosynthetic reaction center.
The Photosynthetic Reaction Center
The pigment–protein complex that is responsible fortranslocating the electron across the photosynthetic mem-brane is termed the photosynthetic reaction center. Itcomprises the antenna pigments, the organic and inor-ganic molecules that function as electron transfercofactors, and the proteins that provide the scaffold forthese components.
Oxygenic phototrophs employ two photosyntheticreaction centers in series for achieving transmembraneelectron transfer.
Photosystem II
The net reaction performed by PS II can be summarizedas:
Plastoquinone
2H2O + 2
Plastoquinol
2
n = 6–10OH
OH
CH3
O
O
CH3
CH3 CH3
CH3
(CH2–CH) C–CH2)n–H + O2(CH2–CH) C–CH2)n–H [6]
CH3
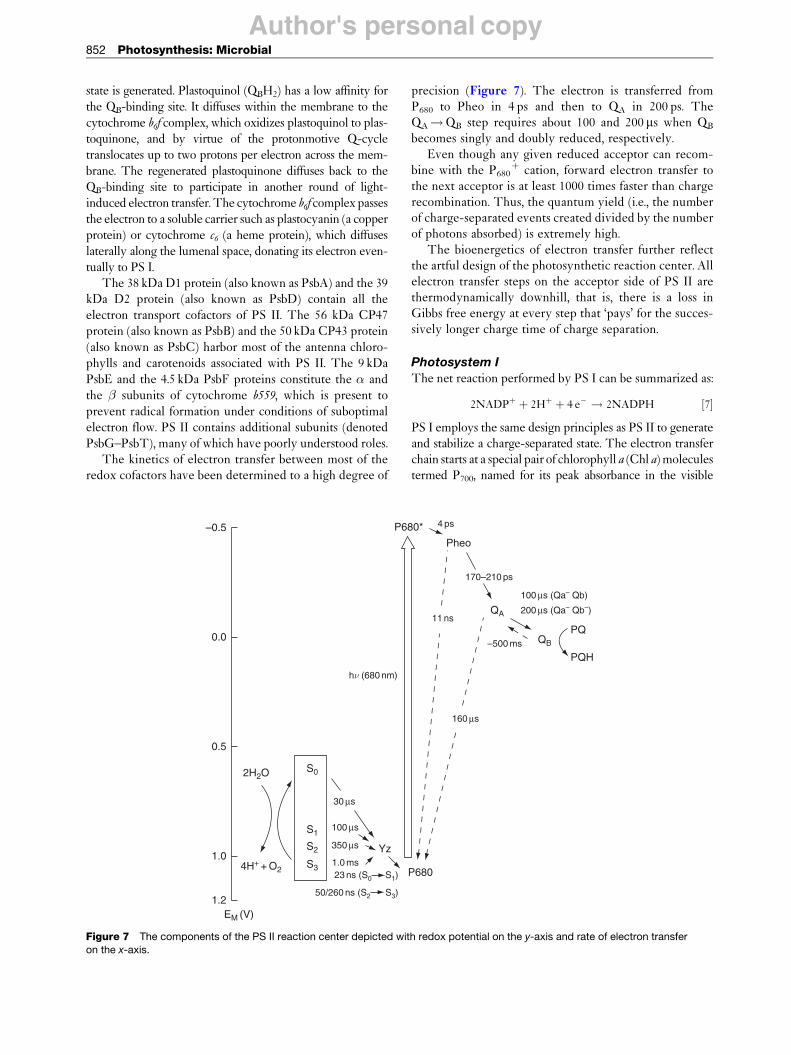
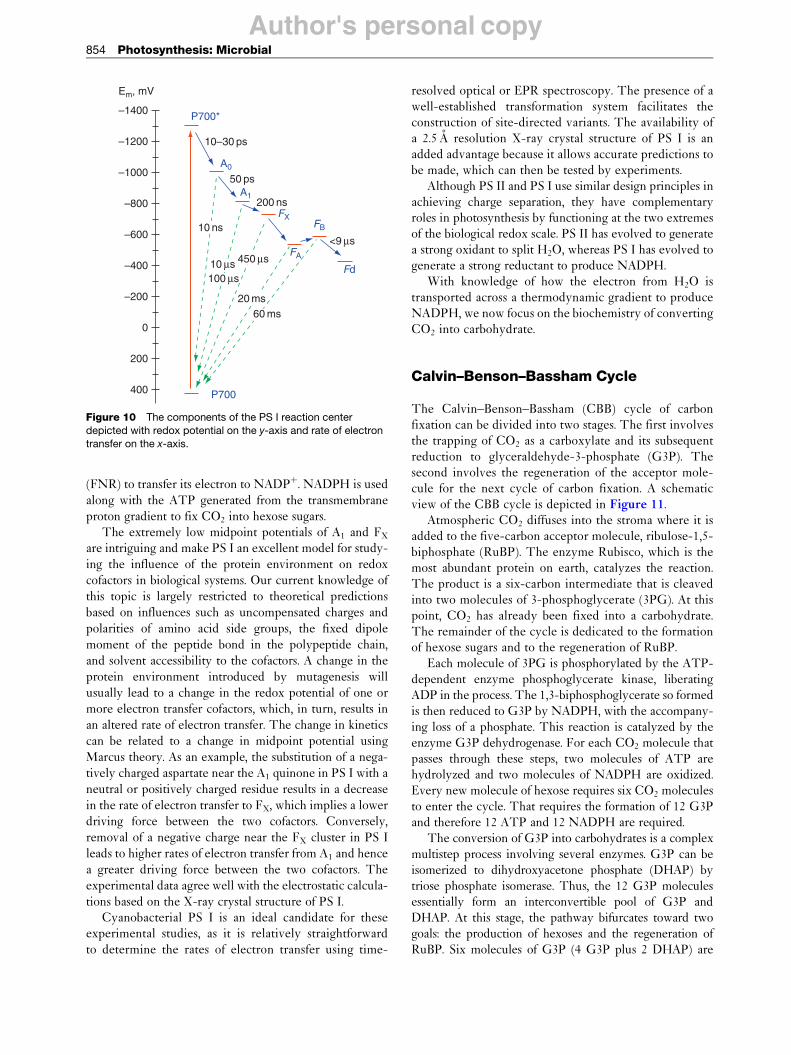
The structural model of cyanobacterial PS II (Figure 5)shows that the redox cofactors bound to the D1 and D2proteins form two branches that are arranged symmetri-cally along a pseudo-C2 axis of symmetry (Figure 6).
The electron transfer chain in PS II starts at a specialpair of chlorophyll a (Chl a) molecules termed P680,named for its peak absorbance in the visible region. Themagnesium atoms between the two Chl a molecules(PD1 and PD2) are separated by a distance of only 7.6 A.When the exciton reaches P680 via the antenna system, itbecomes excited to the singlet state and the electron is
transferred to the primary acceptor, a pheophytin mole-cule. A bridging Chl a molecule acts as an intermediatebetween P680 and the pheophytin. The bridging Chl a
molecules on either side of the pseudo-C2 axis (ChlD1 andChlD2) are located 10.4 and 10.3 A, respectively, from PD1
and PD2. The two pheophytin molecules (PheoD1 andPheoD2) are located at a distance of 10.5 and 10.7 A,respectively, from ChlD1 and ChlD2.
The charge-separated state between P680þ and Pheo�
is stabilized by the transfer of the electron from Pheo� toQA and then to QB, both of which are molecules of
Author's personal copy
PsbZ PsbK PsbJPsbH
CP47
D1PsbI
CP43
PsbT
PsbL PsbM
D2α
β
Cyt b-559 X2
X1
X3
Figure 5 Overall view of the PS II monomer. Transmembrane �-helices are represented by cylinders. The main subunits are colored
as follows: reaction center subunits D1 (yellow) and D2 (orange), antenna subunits CP43 (magenta) and CP47 (red), and the � and �subunits of cytochrome b559 (green and cyan, respectively). Low molecular mass subunits are colored gray. Cofactors are colored
green (Chl a), yellow (pheophytin), red (carotenoid), blue (heme), violet (quinone), and black (lipids).
Local pseudo-C2 axis Cytoplasm
Lumen
9.0QA QB
8.8
30.0
14.226.3
23.7
24.3
10.7
10.37.6
10.4
Tyrz
Mn4–Ca cluster
13.6 13.5
5.429.1
TyrD
10.525.2
19.9 13.2
6.8
11.7
4.1ChlzD1 ChlD1
PD1 PD2
ChlD2
ChlzD2
24.8
13.1
Fe2+
CarD1
CarD2PheoD1 PheoD2
Cyt b559
Figure 6 View of redox-active cofactors and the electron transport chain of PS II along the membrane plane. The pseudo-C2 axis is
represented by the dotted line with the arrow. Selected distances (in angstrom) are drawn between cofactor centers (black lines) andedges of �-systems (red dotted lines).
Photosynthesis: Microbial 851
plastoquinone. A nonheme iron (Fe2þ) located betweenQA and QB aids in the electron transfer. The QA site is13 A distant from PheoD1, while the QA and QB sites areseparated by 18 A, with the nonheme iron precisely in themiddle. Because a quinone molecule (QB) acts as theterminal electron acceptor, PS II is classified as a TypeII reaction center.
Water, through the action of the O2-evolving complex,reduces oxidized P680 via TyrZ, thereby precluding charge
recombination between P680þ and QB
� and thus forming theP680QAQB
� charge-separated state. When a second exciton
reaches P680, the process is repeated, creating an unstableP680QA
�QB� charge-separated state. Two protons are
taken up from the medium, and the stable P680QA(QBH2)
Author's personal copy852 Photosynthesis: Microbial
state is generated. Plastoquinol (QBH2) has a low affinity forthe QB-binding site. It diffuses within the membrane to thecytochrome b6f complex, which oxidizes plastoquinol to plas-toquinone, and by virtue of the protonmotive Q-cycletranslocates up to two protons per electron across the mem-brane. The regenerated plastoquinone diffuses back to theQB-binding site to participate in another round of light-induced electron transfer. The cytochrome b6f complex passesthe electron to a soluble carrier such as plastocyanin (a copperprotein) or cytochrome c6 (a heme protein), which diffuseslaterally along the lumenal space, donating its electron even-tually to PS I.
The 38 kDa D1 protein (also known as PsbA) and the 39kDa D2 protein (also known as PsbD) contain all theelectron transport cofactors of PS II. The 56 kDa CP47protein (also known as PsbB) and the 50 kDa CP43 protein(also known as PsbC) harbor most of the antenna chloro-phylls and carotenoids associated with PS II. The 9 kDaPsbE and the 4.5 kDa PsbF proteins constitute the � andthe � subunits of cytochrome b559, which is present toprevent radical formation under conditions of suboptimalelectron flow. PS II contains additional subunits (denotedPsbG–PsbT), many of which have poorly understood roles.
The kinetics of electron transfer between most of theredox cofactors have been determined to a high degree of
–0.5
0.0
0.5
1.04H+
+ O2
2H2O S0
30 µs
100 µs
350 µs
1.0 ms
hν (680 nm)
P6
Yz
S1
S2
S3
EM (V)1.2
23 ns (S0 S1)
50/260 ns (S2 S3)
Figure 7 The components of the PS II reaction center depicted witon the x-axis.
precision (Figure 7). The electron is transferred fromP680 to Pheo in 4 ps and then to QA in 200 ps. TheQA!QB step requires about 100 and 200 ms when QB
becomes singly and doubly reduced, respectively.Even though any given reduced acceptor can recom-
bine with the P680þ cation, forward electron transfer to
the next acceptor is at least 1000 times faster than chargerecombination. Thus, the quantum yield (i.e., the numberof charge-separated events created divided by the numberof photons absorbed) is extremely high.
The bioenergetics of electron transfer further reflectthe artful design of the photosynthetic reaction center. Allelectron transfer steps on the acceptor side of PS II arethermodynamically downhill, that is, there is a loss inGibbs free energy at every step that ‘pays’ for the succes-sively longer charge time of charge separation.
Photosystem I
The net reaction performed by PS I can be summarized as:
2NADPþ þ 2Hþ þ 4 e – ! 2NADPH ½7�
PS I employs the same design principles as PS II to generateand stabilize a charge-separated state. The electron transferchain starts at a special pair of chlorophyll a (Chl a) moleculestermed P700, named for its peak absorbance in the visible
11 ns
170–210 ps
100 µs (Qa– Qb)
200 µs (Qa– Qb–)
–500 ms
160 µs
4 ps80*
Pheo
QA
QB
PQ
PQH
P680
h redox potential on the y-axis and rate of electron transfer
Author's personal copy
FBFB
FA
FX FX
QK-A QK-B
eC-A3 eC-B3
eC-A2
eC-B1
eC-B2
eC-A1
22.0
12.3
14.9
14.1
8.6
8.2
12.0
14.2
8.6
8.8
11.7
Figure 9 The arrangement of cofactors in PS I with center-to-
center distances (in angstrom). The Chl a molecules, eC-A1 and
eC-B1, constitute the special pair of chlorophylls P700. Thechlorophylls eC-A3 and eC-B3 represent A0 on the PsaA and
PsaB branches, respectively. QK-A and QK-B represent
phylloquinone (A1) on the PsaA and PsaB branches, respectively.
The three [4Fe–4S] clusters are FX, FA, and FB.
Photosynthesis: Microbial 853
region. When P700 becomes excited to the singlet state, itselectron is transferred to the primary acceptor, a Chl a
monomer. An additional molecule of Chl a located betweenP700 and A0 acts as an electron transfer bridge. The charge-separated state is stabilized by rapid transfer of the electron toa molecule of bound phylloquinone (A1) and then to a seriesof [4Fe–4S] clusters termed FX, FA, and FB, which function asan electron transfer wire. Because the terminal acceptors areiron–sulfur clusters, PS I is a Type I reaction center.
A model based on the 2.5 A resolution crystal structureof the trimeric PS I reaction center from the cyanobacter-
ium Synechoccoccus elongatus (PDB ID 1JB0) has provided
detailed structural information on the arrangement of
proteins and cofactors (Figure 8). Each monomer of the
PS I heterodimer contains 96 Chl a molecules, 22 carote-
noids, and 4 lipids. All of the organic cofactors in the
electron transport chain are arranged in two pseudo-C2
symmetric branches along the PsaA–PsaB heterodimer
(Figure 9). However, unlike PS II, in which electron
transfer occurs only along one branch, both the branches
in PS I participate to varying degrees. In higher plants and
algae, electron transfer along the PsaB branch is nearly as
active as along the PsaA branch, whereas in cyanobacteria
electron transfer is more asymmetrical, with only a small
minority of electrons transported along the PsaB branch.However, which branch is most active has little prac-
tical consequence, as the two branches converge at Fx, a
unique interpolypeptide [4Fe-4S] cluster ligated by two
cysteine residues from PsaA and two cysteine residues
from PsaB. Fx is possibly the most reducing iron–sulfur
cluster in biology, with a measured midpoint potential
of –710 to –730 mV. The terminal [4Fe–4S] clusters, FA
PsaE
PsaCPsaD Stroma
Lumen
Figure 8 Side view of the arrangement of all proteins in one
monomer of PS I. The main subunits are colored as follows: PsaA
(blue), PsaB (red), PsaC (pink), PsaD (turquoise), PsaE (green),and PsaF (yellow). The iron–sulfur clusters in PsaC are colored
yellow. The antenna pigments have been omitted for clarity. The
vertical line (right) depicts the crystallographic C3 axis.
and FB, are located in the PsaC subunit, which is located
on the stromal side of the membrane. To our knowledge,
PsaC is the only membrane-associated protein whose
three-dimensional structure has been solved in both the
bound and unbound forms. PsaD and PsaE are two addi-
tional stromal proteins that flank PsaC, stabilizing
its association with the heterodimeric core. These
stromal proteins are vital for the docking of soluble
electron acceptors such as ferredoxin and flavodoxin.
Cyanobacterial PS I contains seven additional subunits
(PsaF, PsaI, PsaJ, PsaK, PsaL, PsaM, and PsaX) whose
roles are not well established. Some function as additional
antenna proteins by binding one or two chlorophyll a
molecules. Others, such as PsaL, along with PsaI and
PsaM, contribute to the trimerization of cyanobacterial
PS I. It is interesting that the absence of a portion of the
C-terminus in the PsaL subunit leads to a completely
monomeric PS I in higher plants and algae.The electron transfer kinetics among the bound cofac-
tors in PS I are roughly comparable to those in PS II
(Figure 10). The primary electron donor, P700, becomes
oxidized in �20 ps, and the electron is transferred from A0
to A1 in 50 ps. The electron transfer step from A1 to FX is
biphasic with lifetimes of�200 ns and 20 ns. It is likely that
the slower kinetic phase represents electron transfer along
the PsaA branch of cofactors, while the faster phase repre-
sents electron transfer along the PsaB branch. The rates of
electron transfer between FX, FA, and FB are not known
with certainty; however, soluble ferredoxin is known to be
reduced within 500 ns. The reduced ferredoxin interacts
with the enzyme ferredoxin-NADPþ oxidoreductase
Author's personal copy
–1400
Em, mV
–1200 10–30 ps
P700*
P700
A0
A1
FXFB
FA
Fd
50 ps
200 ns
20 ms60 ms
10 ns<9 µs
450 µs10 µs100 µs
–1000
–800
–600
–400
–200
200
400
0
Figure 10 The components of the PS I reaction center
depicted with redox potential on the y-axis and rate of electron
transfer on the x-axis.
854 Photosynthesis: Microbial
(FNR) to transfer its electron to NADPþ. NADPH is usedalong with the ATP generated from the transmembraneproton gradient to fix CO2 into hexose sugars.
The extremely low midpoint potentials of A1 and FX
are intriguing and make PS I an excellent model for study-ing the influence of the protein environment on redoxcofactors in biological systems. Our current knowledge ofthis topic is largely restricted to theoretical predictionsbased on influences such as uncompensated charges andpolarities of amino acid side groups, the fixed dipolemoment of the peptide bond in the polypeptide chain,and solvent accessibility to the cofactors. A change in theprotein environment introduced by mutagenesis willusually lead to a change in the redox potential of one ormore electron transfer cofactors, which, in turn, results inan altered rate of electron transfer. The change in kineticscan be related to a change in midpoint potential usingMarcus theory. As an example, the substitution of a nega-tively charged aspartate near the A1 quinone in PS I with aneutral or positively charged residue results in a decreasein the rate of electron transfer to FX, which implies a lowerdriving force between the two cofactors. Conversely,removal of a negative charge near the FX cluster in PS Ileads to higher rates of electron transfer from A1 and hencea greater driving force between the two cofactors. Theexperimental data agree well with the electrostatic calcula-tions based on the X-ray crystal structure of PS I.
Cyanobacterial PS I is an ideal candidate for theseexperimental studies, as it is relatively straightforwardto determine the rates of electron transfer using time-
resolved optical or EPR spectroscopy. The presence of awell-established transformation system facilitates theconstruction of site-directed variants. The availability ofa 2.5 A resolution X-ray crystal structure of PS I is anadded advantage because it allows accurate predictions tobe made, which can then be tested by experiments.
Although PS II and PS I use similar design principles inachieving charge separation, they have complementaryroles in photosynthesis by functioning at the two extremesof the biological redox scale. PS II has evolved to generatea strong oxidant to split H2O, whereas PS I has evolved togenerate a strong reductant to produce NADPH.
With knowledge of how the electron from H2O istransported across a thermodynamic gradient to produceNADPH, we now focus on the biochemistry of convertingCO2 into carbohydrate.
Calvin–Benson–Bassham Cycle
The Calvin–Benson–Bassham (CBB) cycle of carbonfixation can be divided into two stages. The first involvesthe trapping of CO2 as a carboxylate and its subsequentreduction to glyceraldehyde-3-phosphate (G3P). Thesecond involves the regeneration of the acceptor mole-cule for the next cycle of carbon fixation. A schematicview of the CBB cycle is depicted in Figure 11.
Atmospheric CO2 diffuses into the stroma where it isadded to the five-carbon acceptor molecule, ribulose-1,5-biphosphate (RuBP). The enzyme Rubisco, which is themost abundant protein on earth, catalyzes the reaction.The product is a six-carbon intermediate that is cleavedinto two molecules of 3-phosphoglycerate (3PG). At thispoint, CO2 has already been fixed into a carbohydrate.The remainder of the cycle is dedicated to the formationof hexose sugars and to the regeneration of RuBP.
Each molecule of 3PG is phosphorylated by the ATP-dependent enzyme phosphoglycerate kinase, liberatingADP in the process. The 1,3-biphosphoglycerate so formedis then reduced to G3P by NADPH, with the accompany-ing loss of a phosphate. This reaction is catalyzed by theenzyme G3P dehydrogenase. For each CO2 molecule thatpasses through these steps, two molecules of ATP arehydrolyzed and two molecules of NADPH are oxidized.Every new molecule of hexose requires six CO2 moleculesto enter the cycle. That requires the formation of 12 G3Pand therefore 12 ATP and 12 NADPH are required.
The conversion of G3P into carbohydrates is a complexmultistep process involving several enzymes. G3P can beisomerized to dihydroxyacetone phosphate (DHAP) bytriose phosphate isomerase. Thus, the 12 G3P moleculesessentially form an interconvertible pool of G3P andDHAP. At this stage, the pathway bifurcates toward twogoals: the production of hexoses and the regeneration ofRuBP. Six molecules of G3P (4 G3P plus 2 DHAP) are
Author's personal copy
3 6
6
6
3-Phosphoglycerate
1,3-Diphosphoglycerate
3C
3C
5C
1C3 CO2
Ribulose-1,5-biphosphate
3
5
5C
3C
1
3C
6Pi2Pi
6 ATP
6 ADP3 ATP3 ADP
6 NADPH
6 NADP+
3C
Ribulose-5-phosphate
Glyceraldehyde-3-phosphate
Glyceraldehyde-3-phosphate
Glyceraldehyde-3-phosphate
Carbohydrates
Figure 11 Schematic view of the Calvin–Benson–Bassham (CCB) cycle of carbon fixation. The net uptake of ATP and NADPH at each
step is indicated.
Photosynthesis: Microbial 855
diverted to the regeneration pathway, while the remainingsix molecules (3 G3P plus 3 DHAP) are used for carbohy-drate synthesis.
Three molecules of G3P combine with three moleculesof DHAP via the enzyme fructose biphosphate aldolase toyield three molecules of fructose-1,6-biphosphate (FBP).The FBP is then dephosphorylated to provide three mole-cules of fructose-6-phosphate (F6P).
Of these, two molecules will be used in the regenera-tion pathway, leaving one as the net product of the CCBcycle. F6P is isomerized to glucose-6-phosphate (G6P)and finally to glucose-1-phosphate (G1P), which is theprecursor for oligosaccharide and polysaccharide forma-tion. G1P can be hydrolyzed to form glucose, or it canbe converted to amylose and sucrose via separatepathways.
The input molecules for the regeneration cycle are 4G3P, 2 DHAP, and 2 F6P molecules. Enzymes termedtrans-ketolases and aldolases perform molecular rearran-gements that are necessary to form five-carbon moleculesfrom six-carbon and three-carbon molecules. The finalstep in the regeneration of RuBP is a phosphorylationreaction catalyzed by the ATP-dependent enzyme ribu-lose-5-phosphate kinase. An additional six ATP arerequired for six rounds of this step. The overall CCBcycle can thus be summarized as:
As the name suggests, anoxygenic photosynthetic bacteriado not evolve O2 as a by-product of photosynthesis. Thesedescendants of ancient microbes contain only one type ofreaction center and hence the electrons used to reduce CO2
are taken from highly reduced molecules such as succinateand sulfide. Although most photosynthetic bacteria use theCCB cycle to fix carbon, some are able to fix atmosphericCO2 by other biochemical pathways. Most anaerobic photo-trophs can survive only under very low concentrations of O2.
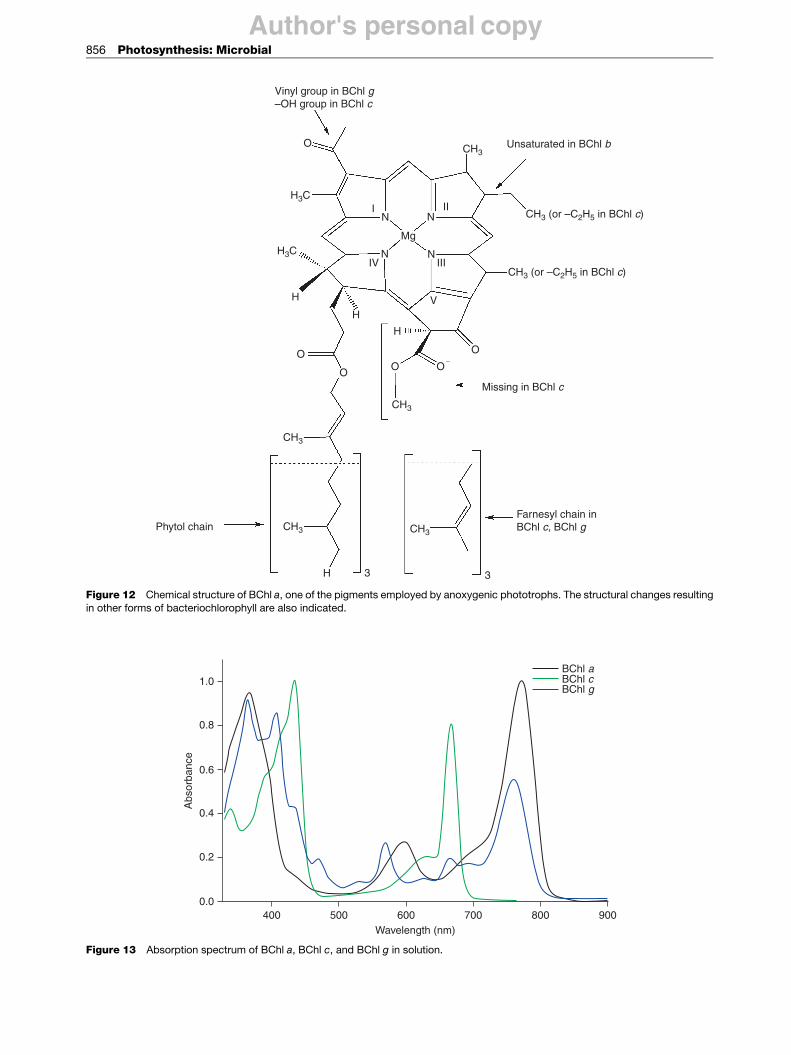
Despite these differences, the general principles ofenergy transduction in anoxygenic photosynthesis aresimilar to those in oxygenic photosynthesis. The primarychromophore belongs to a family of molecules calledbacteriochlorophylls. There are six types of bacterio-chlorophylls, denoted bacteriochlorophyll (BChl) a, b, c,d, e, and g (Figure 12). They are similar to chlorophylls,but absorb light in the near-infrared region (Figure 13).As in aerobic photosynthesis, electron transfer is coupledto the generation of a proton gradient that is used tosynthesize ATP. The energy required to reduce CO2 isprovided by ATP and NADH, a molecule similar toNADPH but lacking the phosphate.
Purple Bacteria
Purple photosynthetic bacteria are a versatile group ofproteobacteria that can be classified further into purple
Author's personal copy
1.0BChl aBChl cBChl g
0.8
0.6
0.4
0.2
0.0400 500 600
Wavelength (nm)
Abs
orba
nce
700 800 900
Figure 13 Absorption spectrum of BChl a, BChl c, and BChl g in solution.
O
H3C
H3C
CH3
CH3
CH3 CH3
Farnesyl chain inBChl c, BChl gPhytol chain
H 3 3
H
H
H
O O
O
O
O
I II
IIIIVN
N N
N
Mg
V
Vinyl group in BChl g–OH group in BChl c
CH3
CH3 (or –C2H5 in BChl c)
CH3 (or –C2H5 in BChl c)
Missing in BChl c
Unsaturated in BChl b
Figure 12 Chemical structure of BChl a, one of the pigments employed by anoxygenic phototrophs. The structural changes resultingin other forms of bacteriochlorophyll are also indicated.
856 Photosynthesis: Microbial
Author's personal copyPhotosynthesis: Microbial 857
nonsulfur bacteria and purple sulfur bacteria. All purplebacteria use a Type II reaction center to generate a protongradient for ATP synthesis, that is, there is no formationof NADPH. The reductant for carbon fixation is derivedfrom organic compounds such as succinate and malate(nonsulfur bacteria) or from inorganic sulfide (sulfur bac-teria). Light-driven electron transfer in purple bacteria iscyclic and hence no net oxidation or reduction occurs.
Purple nonsulfur bacteria are found in ponds, mud,and sewage. Purple sulfur bacteria are obligate anaerobesand are found in illuminated anoxic zones of lakes whereH2S accumulates and also in geothermal sulfur springs.Both fix carbon via the CBB cycle.
All purple bacteria have a very efficient antenna systemconsisting of BChl a, BChl b, and carotenoids. The presenceof purple carotenoids such as spirilloxanthin gives thesebacteria their distinct color. The first three-dimensionalX-ray crystal structures of a photosynthetic reaction centerwere from purple nonsulfur bacteria (Rhodopseudomonas vir-
idis and Rhodobacter sphaeroides). The basic composition oftheir reaction centers is similar to that of PS II. The primarydonor is a special pair of BChl a molecules, which, afterexcitation by light, transfer the electron to bacteriopheo-phytin, the primary electron acceptor. The charge-separated state is stabilized by successive electron transferto two ubiquinone molecules, QA and QB. After two cyclesof reduction, two protons are taken up from inside themembrane to form the doubly reduced dihydroubiquinolin the QB site. Dihydroubiquinol diffuses to the cytochromebc1 complex, where it becomes oxidized, regenerating ubi-quinone. The cytochrome bc1 complex employs theprotonmotive Q-cycle and translocates up to two protonsper electron across the membrane. The energy stored in theresulting electrochemical proton gradient is used to synthe-size ATP via the membrane-bound ATP synthase complex.The cytochrome bc1 complex completes the cycle by trans-ferring the electron back to the primary donor via thesoluble carrier protein cytochrome c.
Green Sulfur Bacteria
Green sulfur bacteria such as Chlorobium tepidum andChlorobium vibrioforme belong to the phyla Chlorobi andare strictly anaerobic photoautotrophs. They use reducedsulfur compounds as their electron donors and fix carbonusing the reverse TCA cycle. Unlike purple bacteria,light-induced electron transfer is noncyclic in green sul-fur bacteria; hence NADPH is generated. These bacterialive in sulfur-rich environments that have characteristi-cally low light intensities. They employ a unique antennacomplex termed the chlorosome, which comprises BChl c,BChl d, and BChl e. It is the largest known antennastructure in biology, with each chlorosome containing�200 000 BChl molecules. The habitats of green sulfurbacteria necessitate such an extensive antenna system,
requiring a very large optical cross section to capture
the few available photons. The light energy is transferred
to a homodimeric Type I reaction center via the BChl a
containing the Fenna–Matthews–Olsen (FMO) protein.
The FMO protein is soluble in water, and was the
first chlorophyll-containing protein to have its three-
dimensional structure solved. The reaction center core
is a homodimer of PscA, and it contains most of the redox
cofactors. Electron transfer begins at P840, a special pair
of BChl a molecules, and proceeds through the primary
acceptor, a Chl a molecule monomer, and three [4Fe–4S]
clusters FX, FA, and FB. It is uncertain whether a quinone
functions as an intermediate electron transfer cofactor
between A0 and FX. FA and FB are bound to a protein
named PscB, which has an unusually long N-terminal
segment of proline, lysine, and arginine residues. A pro-
tein named PscD is thought to be involved in the docking
of soluble ferredoxin and in the stabilization of the FMO
protein. Another protein, PscC, is a tightly bound cyto-
chrome, c551, that donates electrons to P840.
Heliobacteria
Heliobacteria (e.g., Heliobacterium mobilis and Heliobacterium
modesticaldum) are members of the phylum Firmicutes and
are the only known Gram-positive photosynthetic organ-
isms. They were discovered 25 years ago in soil on
the campus of Indiana University, Bloomington.
Heliobacteria are anaerobic photoheterotrophs that fix nitro-
gen and are commonly found in rice fields. They can grow on
selected organic substrates like pyruvate, lactate, and buty-
rate. Heliobacteria do not contain ribulose-1,5-bisphosphate
or ATP-citrate lyase, the two enzymes commonly used in
carbon fixation, but rather incorporate carbon via an incom-
plete reductive carboxylic acid pathway. These bacteria use
BChl g as their primary pigment and employ a simple homo-
dimeric Type I reaction center to perform noncyclic electron
transfer. The components of the electron transfer chain are
similar to green sulfur bacteria except that the pigment used
as the special pair (P798) is BChl g. The reaction center core is
a homodimer of PshA, and it contains the primary donor and
acceptor chlorophylls and the FX iron–sulfur cluster. The FA
and FB iron–sulfur clusters are harbored on a low molecular
mass polypeptide termed PshB. Similar to the reaction cen-
ters in the phylum Chlorobi, the participation of a quinone as
an electron transfer cofactor between A0 and FX is still under
debate.Little or no structural information is available on any
homodimeric Type I reaction center. Based on analogy
with PS I, it is believed that a bifurcating electron transfer
chain with two equivalent branches of cofactors exists in
these reaction centers, but there is no spectroscopic evi-
dence yet to support this proposal.
Author's personal copy858 Photosynthesis: Microbial
Other Photosynthetic Bacteria
Some species of photosynthetic bacteria do not fall underany of the previously discussed categories. The greengliding bacteria (Chloroflexi), also known as green filamen-tous bacteria, can grow photosynthetically under anaerobicconditions or in the dark by respiration under aerobicconditions. Like green sulfur bacteria, they harvest lightby using chlorosomes, but like purple bacteria, theyemploy a Type II reaction center. These poorly studiedorganisms fix CO2 via the 3-hydroxypropionate pathway.
The most recent addition to the list of photosyntheticmicrobes is an acidobacterium, C. thermophilum, whichreportedly synthesizes BChl a and BChl c in aerobicenvironments. This organism was isolated from microbialmats at an alkaline hot spring and is thought to containchlorosomes and a homodimeric Type I reaction center.Further studies are needed to determine whether thephotosynthetic apparatus has new and interesting fea-tures, or whether it falls into a typical Type I class.
The Evolution of Photosynthesis
The origin of photosynthesis is such an ancient event thatthe details of how this biological process developed may beirretrievably lost. Nevertheless, an extensive interdisciplin-ary effort involving researchers in biochemistry, genetics,biophysics, geology, biogeochemistry, and bioinformaticshas led to new and important insights into the evolutionaryhistory of photosynthesis.
There is widespread agreement that the first photosyn-thetic organisms were anoxygenic prokaryotes and that the
Primordial homo
Primitive type II RC
Purple bacterial RC Photosystem II
Cyanobac
Figure 14 Schematic representation of the homodimeric ‘primordi
cyanobacteria, with their much more complicated system of
two linked photosystems, were a later development.
However, the origin of the most primitive photosynthetic
machinery remains shrouded in mystery.According to one school of thought, a common single
polypeptide could have been the ancestor to both Type II
and Type I reaction centers. This primordial reaction cen-
ter would have diverged into a homodimeric reaction
center and subsequent development would have led to the
present reaction centers found in anoxygenic phototrophs.
Alternately, the formation of a homodimeric reaction center
might have been the initial photosynthetic event
(Figure 14). From a geometrical perspective, a three-
dimensional surface is easiest to construct by abutting two
two-dimensional surfaces. A highly protected environment
at the interface of the two identical proteins would neatly
solve the problem of shielding the electron transfer cofac-
tors from water without requiring a highly folded three-
dimensional structure such as the interior of a protein.
Inherent in both ideas, the formation of a heterodimeric
core would have been a result of gene divergence from a
homodimeric core. The similar motifs of charge separation
and charge stabilization in both Type I and Type II reac-
tion centers are certainly compatible with a common origin.
Genetic fusion between two organisms, one having a Type
II and the other a Type I reaction center, would have led to
the colocation of both in the same organism. This fusion
organism likely evolved into the present-day cyanobacteria.The terminal electron acceptor of the primordial reac-
tion center in either case could have been a quinone or an
iron–sulfur cluster. The recruitment of a bacterial ferre-
doxin later in evolution would account for the presence of
dimeric RC
Green sulfur bacterial/heliobacterial RC
Photosystem I
Primitive type I RC
teria
al reaction center’ model of photosynthetic evolution.
Author's personal copyPhotosynthesis: Microbial 859
FA and FB in heterodimeric as well as homodimeric TypeI reaction centers. It is also possible that a completelydifferent moiety could have functioned as the primordialterminal acceptor, with subsequent modifications that ledto the emergence of Type II and Type I reaction centers.
In Type II reaction centers, the transition from homo-dimeric to heterodimeric state might have occurred tospecialize the function of the QA and QB quinones as one-electron and two-electron gates, respectively. The reasonfor the transition is less clear in Type I reaction centers,which do not require a division of labor between the twoquinones. It is nevertheless interesting that heterodimericType I reaction centers are exclusively associated withcyanobacteria, algae and plants, organisms that employaccessory antenna chlorophyll proteins. They might haveevolved from a homodimeric to a heterodimeric state toprovide specialized binding sites for these additionalstructures. The two branches of redox cofactors are highlysymmetrical in heterodimeric Type I and Type II reac-tion centers, suggesting that this transition has notresulted in a significant alteration of the protein environ-ment surrounding the electron transport cofactors.
Although analysis of protein similarity is a highlyeffective method to predict evolutionary events, it doesnot help in understanding the development of complexpigment molecules such as chlorophyll. The originalGranick hypothesis holds that biosynthetic pathways forthe formation of chlorophyll recapitulate their evolution.It proposes that the pathway is built forward, with eachstep fulfilling a function, eventually being replaced by thenext step selected for improved utility. The problem withthis proposal is that the synthesis of bacteriochlorophyllproceeds through a chlorophyll-like intermediate step. Astrict interpretation of the Granick hypothesis wouldtherefore imply that bacteriochlorophyll-containingorganisms (anoxygenic) evolved later than chlorophyll-containing organisms (oxygenic). Because the opposite ismost likely the case, the reaction centers and chlorophyllsmay have followed a dissimilar history in evolution.
The origin of the water-splitting complex is also uncer-tain, although some groups have postulated that it evolvedfrom a manganese-containing catalase. According to thisidea, weak electron donors such as H2O2 and Fe(OH)þ
once provided the electrons to PS II. The incorporation ofthe water-splitting complex might have been driven by thenecessity to replace these rare electron donors with themost abundant electron donor on earth, H2O.
Future Research Directions
Although extensive research in the last four decades hasled to a good understanding of the overall design philo-sophy of photosynthetic electron transfer, there remain
many unanswered questions. The mechanism behindthe splitting of H2O is still unknown, primarily becausethe S4 state of the manganese cluster, which is the startingpoint for O–O bond formation, has not been observed.High-resolution crystal structures are invaluable, butthey only provide a static ground-state depiction ofthe proteins and cofactors. Sophisticated spectroscopictechniques, particularly electron paramagnetic resonance,are required to probe the excited states, providing precisedetails of the critical steps in oxygenic photosynthesis.
Homodimeric Type I reaction centers remain poorlyunderstood. The presence of two branches of electrontransfer cofactors in identical environments begs thequestion of how charge separation is initiated and howthe electron ‘chooses’ which of two equivalent pathwaysto take. The main hindrance to progress lies in the inabil-ity to spectroscopically distinguish electron transfer oneither branch.
Photosynthetic reaction centers are undoubtedly thebest model system for studying the influence of the proteinenvironment on the redox potentials of organic and inor-ganic cofactors. This is best exemplified by comparing thequinones in Type II and Type I reaction centers, whichare structurally similar but differ in redox potential byhundreds of millivolts. QA and QB, which are both plasto-quinones, have midpoint potentials of –150 mV andþ100 mV, respectively, when bound to PS II. An evenmore dramatic case is provided by A1, the phylloquinonebound to PS I. Its redox potential in organic solvent issimilar to that of plastoquinone, but when bound to PS I, ithas a midpoint of –800 mV. Thus, the redox cofactors aretuned by the protein to provide an appropriatemidpoint potential for a given electron transfer step.Detailed knowledge of how the tuning occurs will beexceedingly useful in the designing of artificial photosyn-thetic systems.
Indeed, the next decade will probably see a majoradvance in artificial photosynthesis, mainly due to theconcern over the need to develop new sources of energyafter the depletion of fossil fuels. The basic outline ofsolar energy conversion is now known from the knowl-edge of natural photosynthesis, and these principles canbe used to design artificial organic and inorganic systemsof high efficiency. A solar cell consisting of photosyn-thetic reaction centers from spinach and from purplebacteria layered on a silver electrode has already beenshown to generate an electric current. Efforts to developsynthetic counterparts of the reaction center and theoxygen-evolving complex have had limited success dueto the inability to replicate the complex environment of aliving cell. However, the coming years will undoubtedlysee new developments in this field.
Several research groups are also trying to use photo-synthetic organisms to generate H2 that can be used as analternate source of stored solar energy. Several promising
Author's personal copy860 Photosynthesis: Microbial
ideas include the manipulation of cyanobacterial genes toenhance H2 production in whole cells and the synthesis ofa PS I–hydrogenase hybrid complex.
It is somewhat ironic that after depleting the fossilfuels that were produced by photosynthetic organisms inthe first place we are again looking to photosynthesis forour long-term energy needs. Photosynthesis truly hascome full circle.
Acknowledgments
Research in this laboratory is funded by grants from theNational Science Foundation (MCB-0519743) and theUnited States Department of Energy (DE-FG-02-98-ER20314).
Further Reading
Blankenship RE (1992) Origin and early evolution of photosynthesis.Photosynthesis Research 33: 91–111.
Blankenship RE, Madigan MT, and Bauer CE (eds.) (1995) AnoxygenicPhotosynthetic Bacteria. Dordrecht (The Netherlands): KluwerAcademic Publishers.
Golbeck JH (2002) Photosynthetic reaction centers: So little time, somuch to do. Biophysics Textbook Online.http://www.biophysics.org/education/golbeck.pdf
Golbeck JH (ed.) (2007) Photosystem I: The Light Driven Plastocyanin:Ferredoxin Oxidoreductase. Dordrecht (The Netherlands): Springer.
Heinnickel M and Golbeck JH (2007) Heliobacterial photosynthesis.Photosynthesis Research 92: 35–53.
Wydrzynski TJ and Satoh K (2005) Photosystem II: The Light-DrivenWater: Plastoquinone Oxidoreductase. Dordrecht (The Netherlands):Springer.