Original article Nannoplankton biostratigraphic calibration of the evaporitic events in the Neogene Fortuna Basin (SE Spain) § Calibration biostratigraphique des événements évaporitiques du bassin néogène de Fortuna fondée sur les nannofossiles calcaires (SE Espagne) Carlos Lancis a , José-Enrique Tent-Manclús a, * , Jesús-Miguel Soria a , Jesús-Esteban Caracuel a,1 , Hugo Corbí a , Jaume Dinarès-Turell b , Antonio Estévez a , Alfonso Yébenes a a Universidad de Alicante, Departamento de Ciencias de la Tierra y del Medio Ambiente, Apto. 99, 03080 San Vicente del Raspeig, Alicante, Spain b Istituto Nazionale di Geofisica e Vulcanologia, Via di Vigna Murata, 605, 00143 Roma, Italy Received 20 October 2008; accepted 14 September 2009 Available online 1 February 2010 Abstract The Fortuna Basin is an example of a marginal Mediterranean basin with evaporitic sedimentation during the Late Tortonian and Messinian. This basin shows an early restriction event before the main Messinian Salinity Crisis (MSC) that allows the Tortonian Salinity Crisis (TSC) to be proposed as a tectonic uplift event isolating the eastern Betic basins. Four evaporitic events are present in the central part of the Fortuna Basin, from bottom to top: Los Baños Marls Formation (composed by Fenazar Conglomerate Bed, Lower Gypsum Member [Mb] and Sanel Mb), Tale Gypsum Formation (Fm), Chicamo Diatomites and Gypsum Cycles Fm, and Rambla Salada Gypsum Fm. The present work documents the first biostratigraphic dating based on calcareous nannoplankton of these events. The lowest occurrence (LO) of Amaurolithus primus is registered at the upper part of the Sanel Mb, below the Tale Gypsum Fm. The LOs of Amaurolithus delicatus and Reticulofenestra rotaria, which mark the base of the Messinian, occur in the lower part of the Chicamo Cycles Fm, above the Tale Gypsum Fm, the Triquetrorhabdulus rugosus-Nicklithus amplificus integrate form and the LO of Nicklithus cf. amplificus in the upper part of the Chicamo Cycles Fm. Taking into account these results, a new calibration of the available magnetostratigraphic data is presented: the Chicamo Cycles Fm were formed during the reverse chron C3Ar and the Tortonian-Messinian boundary should be found within the Tale Gypsum Fm or near the top of the Sanel Mb. The onset of the TSC, the first restriction phase of the Fortuna Basin, is represented by the Fenazar Conglomerate Bed, bottom of the Los Baños Fm, and not by the Tale Gypsum Fm, as previously considered. # 2010 Elsevier Masson SAS. All rights reserved. Keywords: Betic Cordillera; Tortonian Salinity Crisis; Calcareous nannoplankton; Messinian; Fortuna Basin Résumé Le bassin de Fortuna est un exemple de bassin méditerranéen marginal avec sédimentation évaporitique pendant le Tortonien supérieur et le Messinien. Il présente l’enregistrement d’événements de restriction avant la Crise de Salinité Messinienne (CSM), ce qui a permis de mettre en évidence une Crise de Salinité Tortonienne (CST) induite par un soulèvement tectonique qui a isolé le bassin. Dans la partie centrale du bassin, on peut reconnaître quatre ensembles évaporitiques, avec de bas en haut, la Formation Los Baños (Couche Conglomératique de Fenazar, Membre [Mb] du Gypse Inférieur, Mb des Marnes de Sanel), la Formation (Fm) Gypse de Tale, la Fm Cycles de Diatomites et Gypse de Chicamo et la Fm Gypse de Rambla Salada. Ici, pour la première fois, nous datons ces ensembles sur la base d’une analyse biostratigraphique du nannoplancton calcaire. La première occurrence d’Amaurolithus primus est enregistrée dans la partie supérieure du Mb Sanel, au-dessous de la Fm Gypse de Tale. Les premières occurrences d’Amaurolithus delicatus et de Reticulofenestra rotaria, qui marquent la base du Messinien, sont localisées dans la partie inférieure de la Fm Cycles de Chicamo, au-dessus de la Fm Gypse de Tale. Finalement, la transition Triquetrorhabdulus rugosus-Nicklithus amplificus et la première occurrence de Nicklithus cf. amplificus sont enregistrées dans la partie supérieure de la Fm Cycles de Chicamo. Sur la base Geobios 43 (2010) 201–217 § Corresponding editor: Frédéric Quillévéré. * Corresponding author. E-mail address: [email protected](J.-E. Tent-Manclús). 1 Professor Jesús Caracuel passed away in the course of this work. We dedicate it to his memory. 0016-6995/$ – see front matter # 2010 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.geobios.2009.09.007
Transcript
Original article
Nannoplankton biostratigraphic calibration of the evaporitic eventsin the Neogene Fortuna Basin (SE Spain)§
Calibration biostratigraphique des événements évaporitiques du bassin néogènede Fortuna fondée sur les nannofossiles calcaires (SE Espagne)
Carlos Lancis a, José-Enrique Tent-Manclús a,*, Jesús-Miguel Soria a, Jesús-Esteban Caracuel a,1,Hugo Corbí a, Jaume Dinarès-Turell b, Antonio Estévez a, Alfonso Yébenes a
a Universidad de Alicante, Departamento de Ciencias de la Tierra y del Medio Ambiente, Apto. 99, 03080 San Vicente del Raspeig, Alicante, Spainb Istituto Nazionale di Geofisica e Vulcanologia, Via di Vigna Murata, 605, 00143 Roma, Italy
Received 20 October 2008; accepted 14 September 2009
Available online 1 February 2010
Abstract
The Fortuna Basin is an example of a marginal Mediterranean basin with evaporitic sedimentation during the Late Tortonian and Messinian.This basin shows an early restriction event before the main Messinian Salinity Crisis (MSC) that allows the Tortonian Salinity Crisis (TSC) to beproposed as a tectonic uplift event isolating the eastern Betic basins. Four evaporitic events are present in the central part of the Fortuna Basin, frombottom to top: Los Baños Marls Formation (composed by Fenazar Conglomerate Bed, Lower Gypsum Member [Mb] and Sanel Mb), Tale GypsumFormation (Fm), Chicamo Diatomites and Gypsum Cycles Fm, and Rambla Salada Gypsum Fm. The present work documents the firstbiostratigraphic dating based on calcareous nannoplankton of these events. The lowest occurrence (LO) of Amaurolithus primus is registered at theupper part of the Sanel Mb, below the Tale Gypsum Fm. The LOs of Amaurolithus delicatus and Reticulofenestra rotaria, which mark the base ofthe Messinian, occur in the lower part of the Chicamo Cycles Fm, above the Tale Gypsum Fm, the Triquetrorhabdulus rugosus-Nicklithusamplificus integrate form and the LO of Nicklithus cf. amplificus in the upper part of the Chicamo Cycles Fm. Taking into account these results, anew calibration of the available magnetostratigraphic data is presented: the Chicamo Cycles Fm were formed during the reverse chron C3Ar andthe Tortonian-Messinian boundary should be found within the Tale Gypsum Fm or near the top of the Sanel Mb. The onset of the TSC, the firstrestriction phase of the Fortuna Basin, is represented by the Fenazar Conglomerate Bed, bottom of the Los Baños Fm, and not by the Tale GypsumFm, as previously considered.# 2010 Elsevier Masson SAS. All rights reserved.
Le bassin de Fortuna est un exemple de bassin méditerranéen marginal avec sédimentation évaporitique pendant le Tortonien supérieur et leMessinien. Il présente l’enregistrement d’événements de restriction avant la Crise de Salinité Messinienne (CSM), ce qui a permis de mettre enévidence une Crise de Salinité Tortonienne (CST) induite par un soulèvement tectonique qui a isolé le bassin. Dans la partie centrale du bassin, onpeut reconnaître quatre ensembles évaporitiques, avec de bas en haut, la Formation Los Baños (Couche Conglomératique de Fenazar, Membre[Mb] du Gypse Inférieur, Mb des Marnes de Sanel), la Formation (Fm) Gypse de Tale, la Fm Cycles de Diatomites et Gypse de Chicamo et la FmGypse de Rambla Salada. Ici, pour la première fois, nous datons ces ensembles sur la base d’une analyse biostratigraphique du nannoplanctoncalcaire. La première occurrence d’Amaurolithus primus est enregistrée dans la partie supérieure du Mb Sanel, au-dessous de la Fm Gypse de Tale.Les premières occurrences d’Amaurolithus delicatus et de Reticulofenestra rotaria, qui marquent la base du Messinien, sont localisées dans lapartie inférieure de la Fm Cycles de Chicamo, au-dessus de la Fm Gypse de Tale. Finalement, la transition Triquetrorhabdulus rugosus-Nicklithusamplificus et la première occurrence de Nicklithus cf. amplificus sont enregistrées dans la partie supérieure de la Fm Cycles de Chicamo. Sur la base
E-mail address: [email protected] (J.-E. Tent-Manclús).1 Professor Jesús Caracuel passed away in the course of this work. We dedicate it to his memory.
0016-6995/$ – see front matter # 2010 Elsevier Masson SAS. All rights reserved.doi:10.1016/j.geobios.2009.09.007
de ces résultats, nous proposons un nouveau calibrage des données magnétostratigraphiques disponibles. La Fm Cycles de Chicamo s’est déposéedurant le Chrone C3Ar. La limite tortonienne/messinienne est localisée près du toit du Mb Sanel ou dans la partie inférieure de la Fm Gypse de Tale.L’initiation de la CST, la première phase de restriction du bassin de Fortuna, est représentée par le Conglomérat de Fenazar, au bas de la Fm LosBaños et non pas par la Fm Gypse de Tale comme préalablement suggéré dans la littérature.# 2010 Elsevier Masson SAS. Tous droits réservés.
Mots clés : Cordillère Bétique ; Bassin de Fortuna ; Crise de Salinité tortonienne ; Messinien ; Nannofossiles calcaires
C. Lancis et al. / Geobios 43 (2010) 201–217202
1. Introduction
During the Late Miocene, the sedimentary basins of theEastern Betic Cordillera have registered two of the mostsingular events in the recent evolution of the Mediterranean.The most striking was the Messinian Salinity Crisis (MSC),manifested by the evaporitic sedimentation in the Mediterra-nean basins. In the Betics, the MSC is well-documented in theNijar Basin (Fortuin and Krijgsman, 2003; Aguirre andSánchez-Almazo, 2004), Sorbas Basin (Martín and Braga,1994; Riding et al., 1998, 1999, 2000; Fortuin et al., 2000;Krijgsman et al., 2001; Sierro et al., 2001; Flores et al., 2005),Vera Basin (Fortuin et al., 1995), Bajo Segura Basin (Feldmannand McKenzie, 1997; Lancis, 1998; Soria et al., 2003, 2008;Caracuel et al., 2004), and Fortuna Basin (Garcés et al., 1998;Tent-Manclús et al., 2007, 2008).
The other singular event was the recently documentedTortonian Salinity Crisis (TSC) named after Krijgsman et al.(2000). The TSC is prior to the MSC and has been defined byevaporite precipitation in the centre of the Lorca and Fortunabasins. The evaporites are related to a restriction phase due totectonics (Garcés et al., 1998; Dinarès-Turell et al., 1999;Garcés et al., 2001; Krijgsman et al., 2006; Tent-Manclús et al.,2008). The TSC was originally proposed as an important eventin the evolution of the Betic basins, because it marks the closureof the foreland North betic strait (Viseras et al., 2004). Recently,Tent-Manclús et al. (2008) related the TSC with the onset of theactivity of the Trans-Alboran Shear Zone (De Larouzière et al.,1988), thus affecting only the eastern portion of the cordillera;consequently this activity would not have an expression in thewestern basins. Likewise, during the Late Miocene, many Beticbasins as Granada, Guadix-Baza, Lorca, and Fortuna undergoemergence, indicating a regional tectonic uplift of the wholecordillera (Viseras et al., 2004).
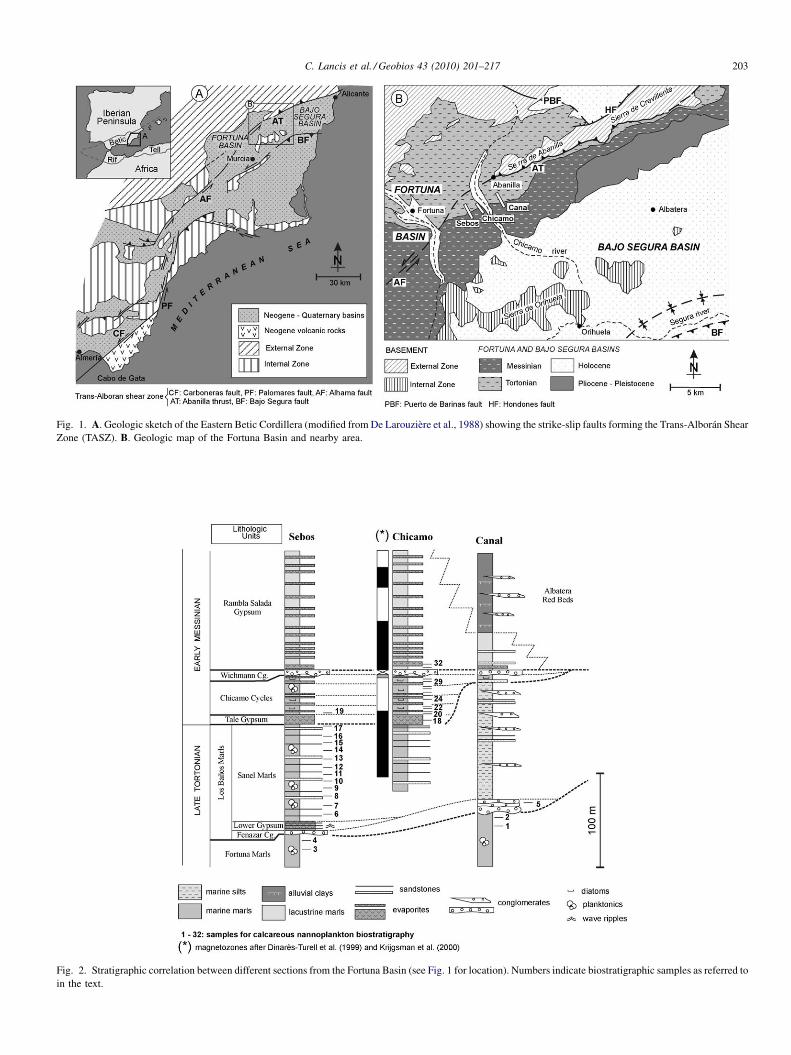
The Fortuna Basin is an intramontane basin developed in theeastern Betic Cordillera (southern Spain, Fig. 1). It is locatedalong the suture zone between the South-Iberian Palaeomargin(External Zone) and the Alboran Block (Internal Zone). Itssedimentary history started in the Early Tortonian as a pull-apart basin associated to the Puerto de Barinas strike-slip Fault(Tent-Manclús, 2003). Two infill periods can be distinguishedin the Fortuna Basin: the first one, during the Tortonian, wasmainly a marine basin where at least 600 m thick Fortuna Marlsaccumulated; and the second one, during the latest Tortonian-Pliocene, continental and restricted marine sedimentation ofshale, marl, gypsum, and conglomerate took place. These twoperiods are associated with different tectonic settings; the firstone was dominated by the transtensive activity of the right-
lateral Puerto de Barinas Fault, while the second one startedwhen the movement of this fault stopped and a new tectonicstress field, related to the Trans-Alborán Shear Zone, started(Tent-Manclús et al., 2008).
The Fortuna Basin has received much attention due to itsevaporitic sediments (Montenat, 1977; Santistebán Bové, 1981;Santistebán and Taberner, 1983; Lukowski, 1987; Müller andHsü, 1987; Lukowski and Poisson, 1990; Montenat et al., 1990;Garcés et al., 1998; Dinarès-Turell et al., 1999; Krijgsman et al.,2000; Playà et al., 2000; Garcés et al., 2001; Tent-Manclús,2003; Soria et al., 2005; Kuiper et al., 2006; Tent-Manclúset al., 2008), thought to be related to the MSC. Initially, theevaporites were considered to be Messinian (Santistebán Bové,1981; Santistebán and Taberner, 1983; Lukowski, 1987; Müllerand Hsü, 1987; Lukowski and Poisson, 1990). However, aspointed out early by Montenat (1977), palaeomagnetic studiesconcluded that the evaporites were older (Garcés et al., 1998,2001; Dinarès-Turell et al., 1999; Krijgsman et al., 2000). Threetemporal calibration options have been proposed for thepalaeomagnetic data obtained in the Rio Chicamo section, twodue to Dinarès-Turell et al. (1999) and one to Krijgsman et al.(2000). None of these three alternative calibrations weredirectly supported by the biostratigraphic data obtained fromthe section.
Due to the relevance of the TSC in the palaeogeographicevolution of the Betic Cordillera, it has been consideredimportant to establish the precise stratigraphy and age of thisevent in the Fortuna Basin, with well-exposed outcrops of theLate Tortonian-Early Messinian sediments. Our technique ofcentrifugation before production of smear slides offers the firstcalcareous nannoplankton biostratigraphic ages for the FortunaBasin evaporitic events, and the calibration of the previouslypublished magnetostratigraphy for the Rio Chicamo section(Dinarès-Turell et al., 1999; Krijgsman et al., 2000).
2. Methods
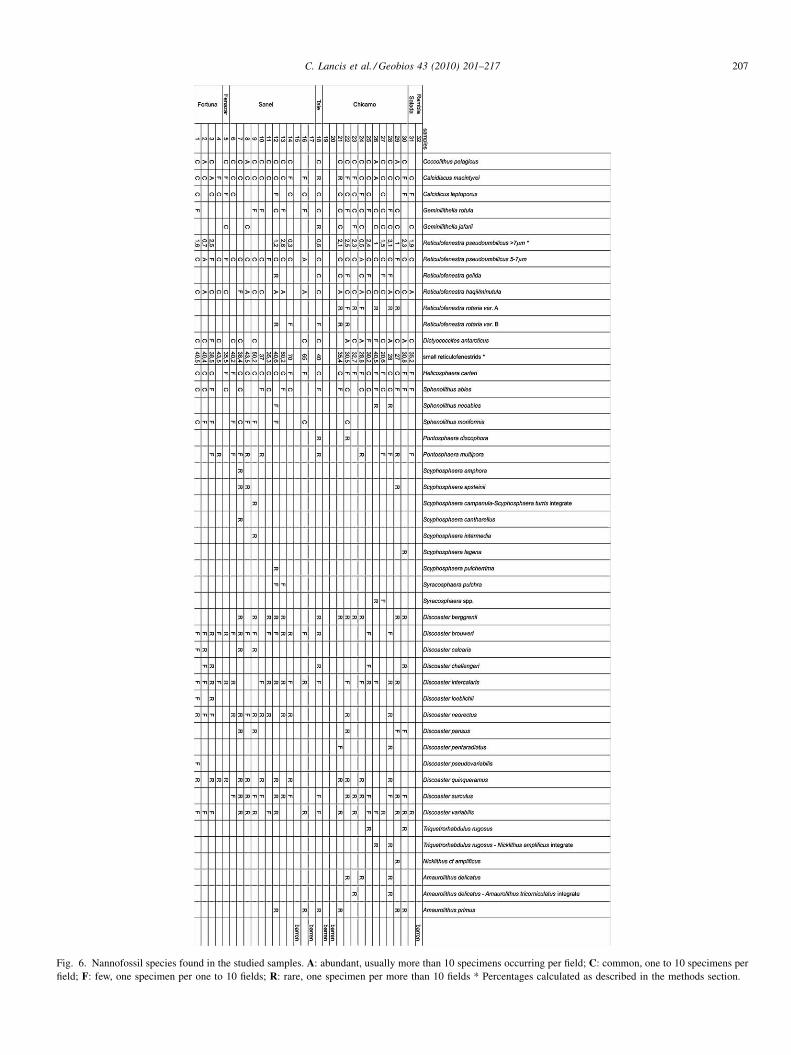
The central basin sediments of the Fortuna Basin are well-exposed in three sections distributed along the Tale ENE-WSWhill (Fig. 1(B)). The Rio Chicamo section is located 4 km southof the Abanilla village in the centre of the Fortuna Basin. Thelower part outcrops at the Chicamo River right margin and theupper part can be followed along the Abanilla-Santomera road(MU-414). The Canal section is located 3 km to the East of theprevious one, following an access road from the Abanilla-Albatera road to a waterway. The Sebos section runs along asecondary road to an industrial suet factory, 2 km west of the
Fig. 2. Stratigraphic correlation between different sections from the Fortuna Basin (see Fig. 1 for location). Numbers indicate biostratigraphic samples as referred toin the text.
Fig. 1. A. Geologic sketch of the Eastern Betic Cordillera (modified from De Larouzière et al., 1988) showing the strike-slip faults forming the Trans-Alborán ShearZone (TASZ). B. Geologic map of the Fortuna Basin and nearby area.
C. Lancis et al. / Geobios 43 (2010) 201–217 203
C. Lancis et al. / Geobios 43 (2010) 201–217204
first section. In all three sections, bedding is tilted 20–258towards the southeast.
Thirty-two samples were collected (interval around 4–5 m)from the Rio Chicamo, Sebos, and Canal sections to studynannofossil assemblages (Fig. 2). For each collected sample,four different smear slides were prepared, using a methodincreasing the nannofossil-to-silt ratio (Lancis, 1998). For thefirst two smear slides, 0.1 g of sediment was suspended in 10 mlof distilled water (pH 7); in one case, the direct suspension(without dilution) and spread on a 300 mm2 surface while, inthe other case, a 1/3 dilution of the suspension with distilledwater was used. The quality of the smear slides was improvedby a second procedure, as follows. A 10 ml suspension of 0.1 gof sediment in distilled water (pH 7) was centrifuged at1800 rpm (450 g) for 2 min at room temperature. After thesupernatant was discarded, distilled water was added to thepellet to make 10 ml of suspension, and the new mixture wassonicated for 8 sec. This centrifugation-sonication procedurewas repeated five times. Finally, 0.1 ml of the suspension wasdirectly spread over a 300 mm2 surface on the slide, withoutdilution for the third smear slide and after 1/3 dilution withdistilled water (pH 7) for the fourth one.
The four prepared smear slides of each collected sample weresemi-quantitatively analysed under � 1000 magnification, scan-ning the whole slide to find any rare biostratigraphic markers. Forcounting the ‘‘small reticulofenestrids’’ (< 3 mm), mean valueswere calculated using the percentage of those coccoliths found in
Fig. 3. The Rio Chicamo and Sebos sections with the rock-unit equivalents used in eage assignment in the different works (Tortonian deposits are in darker grey than
10 visual fields, counting around 3000 nannoliths. Finally,whether the percentage of Reticulofenestra pseudoumbilicus> 7 mm in the samples was higher than 2% was determined aftercounting 500 nannoliths larger than 3 mm.
3. Stratigraphy of the Fortuna Basin centre
3.1. Lithostratigraphic units
Montenat (1977) presented the first lithostratigraphic sketchof the Fortuna Basin, establishing three units: m5, the FortunaMarls, a marly unit including some gypsiferous intercalationsin its upper part; m5y, a gypsiferous unit forming ageomorphologic cuesta (Tale Gypsum, in Azéma andMontenat, 1975); and, finally, m6, the upper unit assigned tothe Messinian and formed mainly by continental facies.Subsequently, Santistebán Bové (1981) and Santistebán andTaberner (1983), Lukowski (1987), Müller and Hsü (1987),Dinarès-Turell et al. (1999), Krijgsman et al. (2000), andGarcés et al. (2001), used many alternative lithologic units, forwhich the relations are shown in Fig. 3.
The discontinuity on top of the Fortuna Marls Fm marks theonset of the second basin infill period. Four evaporitic eventscan be distinguished (Fig. 2) that change towards the east tocontinental red shales with conglomeratic levels (Albatera RedBeds Fm) that also overlie the evaporitic events in the basincentre. The first three can be interpreted as marine environ-
arlier works and in this work. The darker and light-grey backgrounds refer to theMessinian ones). For lithological symbols, see Fig. 2.
Fig. 4. Photography of the Rio Chicamo section, showing the top of the Sanel Marls Member, the Tale Gypsum Formation, the Chicamo Diatomites and GypsumCycles Fm.
C. Lancis et al. / Geobios 43 (2010) 201–217 205
ments but not the fourth, which is mainly a continental one(Figs. 2–4):
� First evaporitic event: Los Baños Marls Fm, composed by:� The Fenazar Conglomerate Bed, a marine unit formed by
Lithofaga-bored metamorphic clasts of Internal Zoneprovenance that erode the Fortuna Fm (Fig. 2);� The Lower Gypsum Mb, composed by gypsum beds and
some sandstone levels having oscillation ripples andhummocky cross-stratification to the eastern margin. Inthe Sebos section (basin centre), the terrigenous sedimentsdisappear and thus the evaporitic levels coalesce to form amarine gypsum bed. This is the first record of evaporiticsedimentation in the Fortuna Basin;� The Sanel Marls Mb, a marly marine unit containing
planktonic microfossils that intercalate some centimetre-thick gypsum beds:
Fig. 5. Mosaic of aerial photographs of the Cabezos Negros-El Tale area. The outcrovolcano), the Lower Gypsum Member, Tale Gypsum Formation, and Wichmann C
� Second evaporitic event: The Tale Gypsum Fm is a 30-m-thick marine gypsum level deposited over a discontinuity thatdeveloped on top of the Sanel Mb (Fig. 4). Krijgsman et al.(2000) consider this unit to be the result of the TSC;� Third evaporitic event: The Chicamo Diatomites and
Gypsum Cycles Fm is formed by five cycles of diatomiticmarls and gypsum beds with intercalated stromatolites. TheWichmann Conglomerate Bed is a thin conglomeratic unitlimited by two erosive surfaces, containing metamorphicclasts of Internal Zone provenance and coral-reef clastsoriginating from the basin margins. The diagenetic study bySantistebán and Taberner (1983) indicates a postdepositionalsubaerial exposure for this bed.;� Fourth evaporitic event: The Rambla Salada Gypsum Fm is
formed by shales and gypsum beds. According to Playà et al.(2000), these sediments were deposited in a mainly lacustrineevaporitic environment.
ps of the volcanic Fortuna lamproites (Tale, Derramadores, and Cabezos Negrosonglomerate Bed are shown.
C. Lancis et al. / Geobios 43 (2010) 201–217206
3.2. Fortuna volcanic rocks
In the central part of the Fortuna Basin, there are threeoutcrops of volcanic lamproitic rocks: Cabezos Negros, El Tale,and Derramadores (Fig. 5). The El Tale and Derramadoresoutcrops have been considered to be subvertical dikes cuttingthe upper part of the Fortuna Fm (Fig. 5; Playà and Gimeno,2006). The Cabezos Negros outcrop has been interpreted as theneck of an individual domic volcano with minor lava-flowexpansions in its north-eastern sector (Fúster et al., 1967). Thedetailed study by Playà and Gimeno (2006) shows thecomplexity of the Cabezos Negros outcrop where severalvolcanic lithofacies can be differentiated, evidencing a widevariety of eruptive mechanisms.
As previously evidenced (Dinarès-Turell et al., 1999; Playàet al., 2000; Kuiper et al., 2006; Playà and Gimeno, 2006), theCabezos Negros volcano is hosted in the marly and carbonaticsediments of the lower part of the Rambla Salada Fm (Fig. 5).Other indirect criteria to establish the relative age of thevolcanic rocks are the lack of lamproite clasts in the WichmannConglomerate Bed, and the existence of a volcano-sedimentarylevel within the lowermost Rambla Salada Fm described byLukowski (1987), probably related to the Cabezos Negrosvolcano.
Bellon et al. (1983), found an absolute 40K/40Ar age of6.16 � 0.30 Ma for the Cabezos Negros outcrop. However,Kuiper et al. (2006) have recently dated samples from theCabezos Negros by the 40Ar/39Ar incremental heating method,reporting an absolute age of 7.71 � 0.11 Ma. These authorsattribute the discrepancy between the 40K/40Ar and 40Ar/39Arages to the intrinsic flaws of the 40K/40Ar method. Playà andGimeno (2006) explain the divergent absolute ages, ‘‘on the,supposedly, same outcrop’’, in terms of the genetic differencesin the eruptive mechanisms of the Cabezos Negros volcano.They propose two possible explanations:
� A successive arrival of two batches of slightly differentmagma during the same eruption that would have provided asame age of volcanism;� The presence of two different and successive eruptions of
lamproites detached some 1 Ma at Cabezos Negros.
In any case, Playà and Gimeno (2006) prefer, after thegeochemical evidence and the associated specific geodynamicrequirements, a restricted lapse of time involved in thelamproite genesis.
4. Nannofossil biostratigraphic results and discussion
The nannofossil assemblages found in the samples from thesuccessive lithostratigraphic units are shown in Fig. 6.
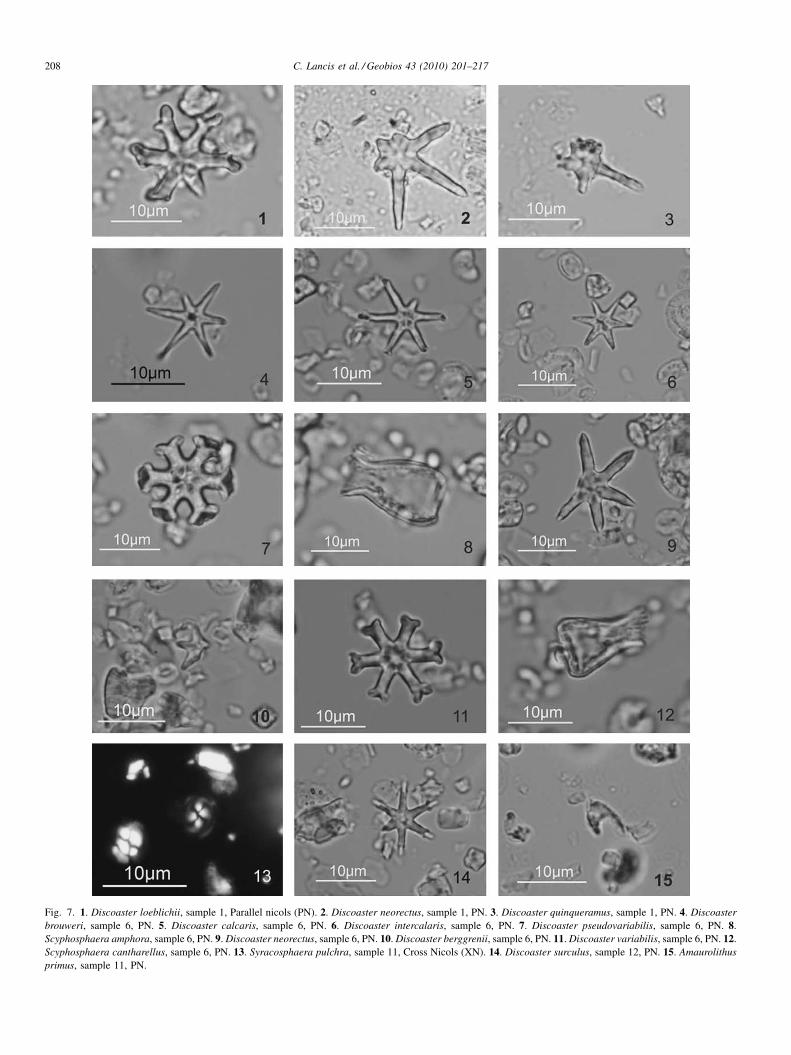
The bottom of the section, represented by the Fortuna Fmyields a well-diversified assemblage, containing Discoastercalcaris, D. neorectus (Fig. 7(2, 9)), D. brouweri (Fig. 7(4)),D. loeblichii (Fig. 7(1)), D. pseudovariabilis, D. quinqueramus(Fig. 7(3)), Coccolithus pelagicus, Sphenolithus moriformis,Sph. abies, Calcidiscus macintyrei, C. leptoporus, Helico-
sphaera carteri, Geminilithella rotula, Dictyococcites antarc-ticus, Pontosphaera multipora, R. pseudoumbilicus >7 mm,R. pseudoumbilicus 5–7 mm, R. haqii, R. minutula and veryabundant ‘‘small reticulofenestrids’’ (R. minuta and Dictyo-coccites productus). The asteroliths are relatively common,with great size and good preservation. The occurrence ofbroken specimens and overgrown nannofossils indicate somereworking in the samples.
The first Discoaster surculus (Fig. 7(14)) and D. berggrenii(Fig. 7(10)) can be observed in the bottom Sanel Mb samples,and the forms D. loeblichii and D. pseudovariabilis are absent.The presence of Scyphosphaera amphora (Fig. 7(8)), Scy.cantharellus (Fig. 7(12)), and Syracosphaera pulchra, and alsothe first rounded reticulofenestrids, as R. rotaria var. B (Lancis,1998) should be noted. The first Amaurolithus primus(Figs. 7(15) and 8(2)) occurs 80 m above the bottom of theSanel Mb.
R. rotaria var. A (Fig. 8(6, 7)) appears 10 m above the TaleGypsum Fm. Amaurolithus delicatus has been found in sample22 above the lowest occurence (LO) of R. rotaria var. A (sample21). The Triquetrorhabdulus rugosus-Nicklithus amplificusintegrate forms (Fig. 8(12, 13, 14)) and Nicklithus cf. amplificus(Fig. 8(15)) can be observed in the uppermost part of theChicamo Cycles Fm (sample 26 and upwards).
The ‘‘small reticulofenestrids’’ (R. minuta and Dictyococ-cites productus) represent about 40% of the nannoliths in theassemblages of the Fortuna Fm. Then the percentage increasesto 70% in some samples of the Sanel Fm and, finally, reduces to30% in the Chicamo Cycles.
The Wichmann Bed and Rambla Salada Fm, as transitionalor continental sediments, lack preserved nannofossils exceptthe first sample (31) above the Wichmann Bed, but its scarcenannofossils do not allow any biostratigraphic interpretations.
4.1. Asteroliths
The discoasterids are scarce in the Mediterranean during theMessinian, unlike in open ocean facies (Müller, 1978, 1990;Perch-Nielsen, 1985; Flores and Sierro, 1989; Flores et al.,1992; Raffi et al., 2003). Discoaster quinqueramus have beenidentified in the first sample of the Fortuna Fm, and despite thescarcity of this species, its First Appearance Datum (FAD) isclearly detected in the Neogene Betic basins (Martín-Pérez,1997; Lancis, 1998). This species is also associated withD. loeblichii, D. pseudovariabilis, D. calcaris, andD. neorectus. Discoaster quinqueramus is absent in thesediments of the Eastern Mediterranean (Theodoridis, 1984;Mazzei, 1985; Raffi et al., 2003), but can be recognized in theWestern Mediterranean (Rio et al., 1976; Müller, 1978; Müller,1990; Flores et al., 1992; Martín-Pérez, 1997; Lancis, 1998). Asmentioned by Müller (1978) concerning the MediterraneanD. quinqueramus, the ones of the Fortuna Fm are smaller andthe central knob is not as distinctly developed as in those oftropical areas. Discoaster quinqueramus and D. berggreniiappear together in our samples. Some workers observe the FADof D. berggrenii before the FAD of D. quinqueramus (Bukry,1971; Rio et al., 1990; Raffi and Flores, 1995) but in some
Fig. 6. Nannofossil species found in the studied samples. A: abundant, usually more than 10 specimens occurring per field; C: common, one to 10 specimens perfield; F: few, one specimen per one to 10 fields; R: rare, one specimen per more than 10 fields * Percentages calculated as described in the methods section.
biozonation, they have been considered as the same taxonomicunit (Müller, 1974; Theodoridis, 1984), the integrate formsbeing common (Rio et al., 1990; Raffi and Flores, 1995).
4.2. Acme of small reticulofenestrids
An increase in the small-sized reticulofenestrids (with thelong axis smaller than 3 mm) including R. minuta andDictyococcites productus specimens have been observed bymany authors (Backman, 1978; Flores, 1985; Flores and Sierro,1987, 1989; Flores et al., 1992; Lancis, 1998; Negri et al., 1999;Negri and Villa, 2000; Raffi et al., 2003; Wade and Bown,2006). The Acme Beginning can be found between the FADs ofD. berggrenii and A. primus (Lancis, 1998; Negri et al., 1999;Negri and Villa, 2000; Raffi et al., 2003). This increment isprobably due to ecological factors, since the R. minuta andD. productus are r-strategy species, considered able to flourishunder nutrient-rich conditions and to tolerate high environ-mental stress (Aubry, 1992; Flores et al., 1995, 2005; Wade andBown, 2006). Therefore, they tend to dominate in thenannoflora assemblages of the continental margins (Haq,1980). Flores et al. (1992) demonstrated that, instead of anecological factor, this is a synchronous event in someMediterranean and North Atlantic sections.
The samples of the Fortuna Fm and Sanel Mb (Fig. 6) showthat the ‘‘small reticulofenestrids’’ form about 40% of theassemblage, indicating that they are included in the period ofthe acme. The top of this interval cannot be precisely identified.
4.3. Paracme of Reticulofenestra pseudoumbilicus
R. pseudoumbilicus with maximum length between 7 and12 mm have been found from the Middle Miocene (from about13.6 Ma) to the Middle Pliocene (3.6 Ma) with an absenceinterval of about 2 Ma around the Late Miocene (Rio et al.,1990; Young, 1990; Gartner, 1992; Takayama, 1993; Raffi andFlores, 1995). Raffi et al. (2003) dated the Paracme Beginning(PB) at about 8.76 Ma and the Paracme End (PE) at about7.17 Ma. Krijgsman et al. (2000) used the paracme ofR. pseudoumbilicus > 7 mm in the Lorca Basin to calibrate themagnetostratigraphy of the Serrata section and the Chicamoand Chorrito sections of the Fortuna Basin.
The R. pseudoumbilicus >7 mm in a percentage higher than2% of the nannoflora assemblage (Fig. 6) occurs from thebottom of the studied section in the Fortuna Fm to the end of themarine sedimentation, possibly indicating the interval abovethe PE biohorizon.
4.4. First Appearance Datum, morphotypes, and range ofReticulofenestra rotaria
R. rotaria has been used for the Mediterranean Late Miocenenannofossil biostratigraphy since Theodoridis (1984) definedthe species (Young, 1990; Flores et al., 1992; Negri et al., 1999;Negri and Villa, 2000; Raffi et al., 2003; Morigi et al., 2007). Inthe Mediterranean Bajo Segura Basin Lancis (1998), observedtwo morphotypes of R. rotaria differing in the distal shield
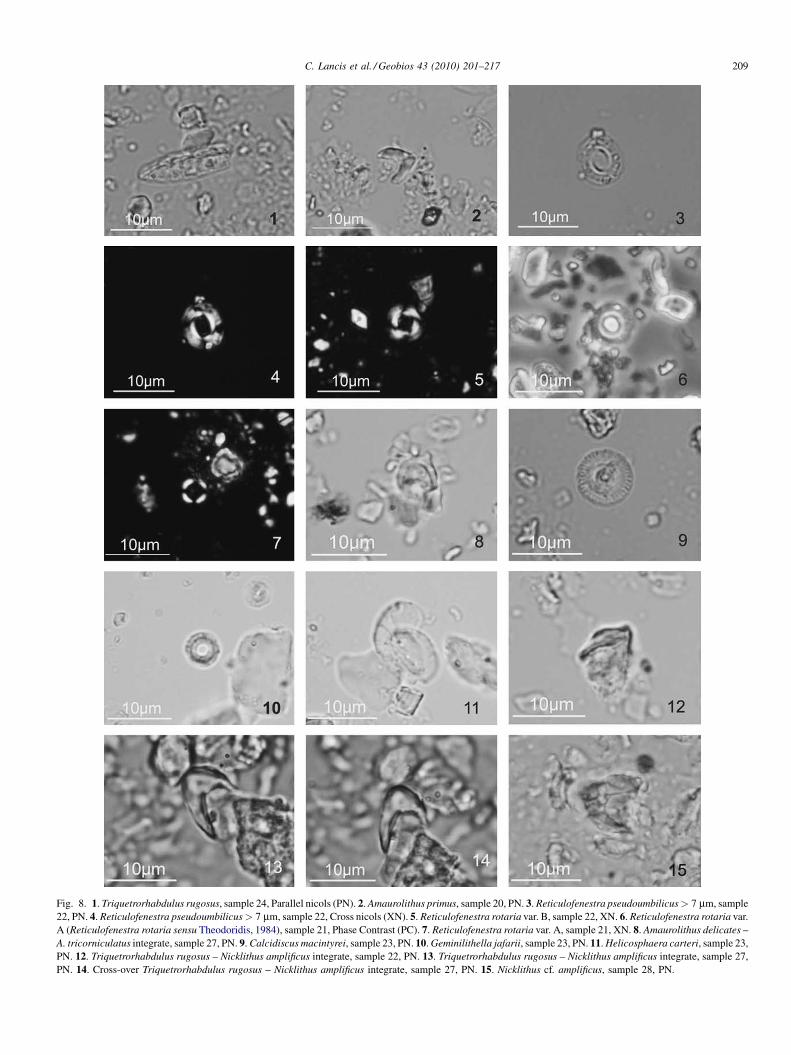
thickness in crossed nicols view (Fig. 8(5–7)). The older one,named R. rotaria var. B (Fig. 8(5)), has a wide central area withradially bent extinction lines and appears slightly above theA. primus FAD, while the R. rotaria var. A (Theodoridis, 1984;s.s.) (Fig. 8(6, 7)) has straight extinction lines and its FADcoincides with the FAD of A. delicatus. Some authors considerthe first event to be the FAD and the second the First CommonOccurrence (FCO) of R. rotaria (Negri et al., 1999; Hilgenet al., 2000; Negri and Villa, 2000; Raffi et al., 2003; Morigiet al., 2007) but they do not separate the two morphotypes. Thelast one is coincident with the FAD of A. delicatus used toestablish the Tortonian/Messinan boundary (Hilgen et al., 2000;Lourens et al., 2004). The Last Appearance Datum (LAD) ofR. rotaria is quite difficult to identify due to the scarcity of thespecies in these basins (Negri et al., 1999; Morigi et al., 2007).
The LO of R. rotaria var. B is found in sample 12 (SanelMb), coincident with the first A. primus; likewise, the LO ofR. rotaria var. A is found in sample 21 (Chicamo Cycles Fm),just before the LO of A. delicatus (sample 22).
4.5. Amaurolithus primus First Appearance Datum
The FAD of Amaurolithus spp. (A. primus) represents anevolutionary event which is very useful for biostratigraphicclassification worldwide (Raffi et al., 1998). This event, easilyidentified in the open ocean, has also been identified in theMediterranean Sea (Flores et al., 1992). The FAD of A. primushas been calibrated in Chron C3Br.2r (Negri et al., 1999; Negriand Villa, 2000; Raffi et al., 2003; Morigi et al., 2007), andLourens et al. (2004) established an age of 7.42 Ma. The FO ofA. primus occurs in the Sanel Mb (sample 12).
4.6. Amaurolithus delicatus First Appearance Datum
Theodoridis (1984) and also Flores et al. (1992) found theFAD of A. delicatus slightly above the FAD of A. primus andslightly below the FCO of R. rotaria. It coincides with the rapidinvasion of planoconvex forms included in the Globorotaliamiotumida-G. conoidea group (Sierro, 1985). The FAD ofA. delicatus can be calibrated in Chron C3Br.1r (Hilgen et al.,1995, 2000; Negri and Villa, 2000) and has generally beencorrelated with the Tortonian/Messinian boundary (Lourenset al., 2004). The LO of A. delicatus occurs in sample 22 in theChicamo Cycles Fm.
4.7. Nicklithus cf. amplificus First Appearance Datum
Sample 29 contains Nicklithus cf. amplificus (Fig. 8(15)).This species has a size of 9.5 mm, smaller than the originaldefinition between 12 and 20 mm (Bukry and Percival, 1971),and therefore we use the cf. term. For ‘‘Amaurolithusamplificus’’, Raffi et al. (1998) name proposed the new genus,Nicklithus because they found transition forms betweenTriquetrorhabdulus rugosus and Amaurolithus spp. at differentbiostratigraphic levels, indicating a polyphyletic origin for theceratoliths. In the Mediterranean Basin, Raffi and Rio (1979)found specimens of 9.3 mm, Martín-Pérez (1997) and Lancis
Fig. 9. Calcareous nannoplankton biozones of the Rio Chicamo/Sebos com-posite section.
C. Lancis et al. / Geobios 43 (2010) 201–217 211
(1998) found specimens between 9 and 11 mm, and Mansouriet al. (2008) reported specimens of 13 mm. The small size ofMediterranean specimens have been interpreted as the result ofrestricted marine conditions during the Messinian (Müller,1978, 1990). Gartner and Bukry (1975) located the FAD ofN. amplificus above the FADs of A. primus and A. delicatus. Rioet al. (1990), in the Indian Ocean, and Raffi and Flores (1995)and Raffi et al. (1995), in the Pacific, found the FAD ofN. amplificus after the FAD of A. delicatus.
Contrary to some authors (Benson and Rakic-El Bied, 1996;Negri et al., 1999; Hilgen et al., 2000), but in agreement withothers (Rio et al., 1990; Raffi and Flores, 1995), we found thatthe LO of N. cf. amplificus occurs after the LOs of R. rotaria varA and A. delicatus in the Fortuna Basin.
In the uppermost part of the Chicamo Cycles Fm (sample26 and upwards), Triquetrorhabdulus rugosus-Nicklithusamplificus integrate specimens (Fig. 8(12–14)) can beobserved. Perch-Nielsen (1977) suggested a phylogeneticrelationship between the Triquetrorhabdulus and Amaurolithus(Gartner and Bukry, 1975; Rio et al., 1990; Raffi and Flores,1995; Raffi et al., 1998).
Intermediate forms T. extensus-N. amplificus have beenreported by different workers in the open ocean (Rio et al.,1990; Raffi and Flores, 1995; Backman and Raffi, 1997) with adistribution slightly below and coincident with N. amplificus.Raffi et al. (1998) described T. rugosus as a speciesconsiderably varying in size and morphology, and theyconsidered the species T. extensus and T. finifer, observedand described by Theodoridis (1984) in the uppermostMiocene, to be aberrant morphovariants of T. rugosus.Triquetrorhabdulus rugosus-N. amplificus integrate forms havebeen also found in the Mediterranean basin (Lancis, 1998; Raffiet al., 2003) around the interval between 7.0 and 6.8 Ma, justbelow the N. amplificus FAD in the upper part of Chron C3Ar(Raffi et al., 2006).
5. Biostratigraphic calibration of themagnetostratigraphic data
5.1. Biostratigraphy
The uppermost portion of the Fortuna Fm studied in thiswork can be assigned to the lower portion of the CN9a subzone(Okada and Bukry, 1980), equivalent to the NN11a of Martini(1971), due to the co-occurrence of D. loeblichii, D. calcaris,D. pseudovariabilis, D. neorectus, and D. quinqueramus (ProtoDecima et al., 1978; Mazzei et al., 1979; Parker et al., 1985; Rioet al., 1990; Raffi and Flores, 1995). The Fenazar ConglomerateBed nannoflora is similar to the Fortuna Fm.
The lowest part of the Sanel Mb (sample 6) can be assignedto upper part of the CN9a/NN11a (Fig. 9) by the presence ofD. surculus. The boundary between the CN9a/CN9b (Okadaand Bukry, 1980), and NN11a/NN11b of Martini (1971) is
marked by the A. primus LO occurring in sample 12 (Lourenset al., 2004). The continuity of the acme of ‘‘smallreticulofenestrids’’ in Sanel Mb and the R. rotaria var. B LOin sample 12 justifies the Sanel Mb biozonation assignation tothe upper part of the CN9a and CN9b of the Okada and Bukry(1980) classification or the upper NN11a and NN11b of Martini(1971). The abundance of Scyphosphaera spp. and Ponto-sphaera multipora, related to shallow marine environments(Martini, 1965; Bukry, 1976; Siesser, 1977; Gartner et al., 1979;Flores, 1985; Lancis, 1998), and the upward decrease inDiscoaster spp., characteristic of oligotrophic environments ofdeeper waters (Hekel, 1973; Haq, 1980; Bukry, 1981; Lohmannand Carlson, 1981; Backman et al., 1986; Flores and Sierro,1987; Rio et al., 1990; Aubry, 1992; Young, 1998; Lancis,1998; Wade and Bown, 2006) indicate an upward increase inthe restriction of the depositional environment of the Sanel Mb.
The Tortonian/Messinian boundary is marked slightly belowthe FAD of A. delicatus (Lourens et al., 2004), the Sanel Mb areuppermost Tortonian and the Chicamo Cycles Fm areMessinian because they contain R. rotaria var A from thelowermost sample. Accordingly, the Tortonian/Messinianboundary may be located at the uppermost part of the SanelMb or within the Tale Gypsum Fm.
Raffi and Flores (1995) have used N. amplificus distributionpatterns to divide the Okada and Bukry (1980) subzone CN9bin three parts: CN9bA, from the FAD of A. primus to the FAD ofN. amplificus; CN9bB, from the FAD of N. amplificus to theLAD of N. amplificus; and CN9bC, from the LAD of
C. Lancis et al. / Geobios 43 (2010) 201–217212
N. amplificus to the LAD of D. quinqueramus. Nicklithusamplificus appears just below the Wichmann Bed, marking thebase of the CN9bB subzone (Okada and Bukry, 1980 emendedby Raffi and Flores, 1995).
Finally, the Rambla Salada samples yield no biostratigraphicelements.
5.2. Magnetostratigraphy
Two magnetostratigraphic studies of the Rio Chicamosection have been published (Dinarès-Turell et al., 1999;Krijgsman et al., 2000). Despite a general agreement of theretrieved polarity magnetozones, both interpretations differ intheir correlation to the standard Global Polarity Time Scale(GPTS, Fig. 10). However, two minor differences in the polaritysuccession exist between those two magnetostratigraphicstudies. The first one concerns the polarity around theWichmann Bed as in the Dinarès-Turell et al. (1999) study,in which a good-quality reverse sample is documented justbelow this Bed, suggesting that the reversal boundary betweenthe underlying reverse magnetozone that extents through theChicamo Cycles Fm and the normal magnetozone at the lowerpart of the Rambla Salada Gypsum Fm, occurs within theWichmann Bed or above (Fig. 10). Krijgsman et al. (2000),instead, interpreted the uppermost Chicamo Cycles Fm asnormal polarity (three samples) but they had no data for theWichmann Bed. In the former study by Dinarès-Turell et al.(1999), a clear reverse polarity is documented from samplesimmediately below the Wichmann Bed, although some
Fig. 10. Previous and proposed new correlation of the Rio Chicamo/Sebos combiostratigraphic and radiometric ages are shown. See text for discussion.
unreliable low-quality samples appear normal. A new samplingalong this interval confirms the poor quality of the magneticsignal from the marls just below the erosional surface at thebottom of the Wichmann Bed and therefore we prefer tomaintain the uncertainty of this portion (grey in Fig. 10). Thesecond difference between the two published studies is theupper extension, due to new roadcuts, of the magnetostrati-graphy by Krijgsman et al. (2000), which documents anadditional reverse chron which has been reproduced in ourrecent resampling (Fig. 10).
Dinarès-Turell et al. (1999) proposed two alternativecorrelations (Fig. 10):
� Tale Gypsum Fm within Chron C3Bn (6.935–7.091 Ma, orearliest Messinian);� Tale Gypsum Fm within Chron C3Br.1n (7.135–7.170 Ma, or
latest Tortonian).
On the other hand, Krijgsman et al. (2000) assigned the samenormal interval to Chron C4n.2n (7.650–8.072 Ma, LateTortonian). None of these correlations were substantiated inthe original works by direct biostratigraphic determinationsalong the Rio Chicamo section.
5.3. Calibration of the magnetostratigraphy withbiostratigraphic data
The nannofossil data presented here offers differentcalibration points (Fig. 10). The first one is the FAD of
posite section magnetozones with the Global Polarity Time Scale. Relevant
C. Lancis et al. / Geobios 43 (2010) 201–217 213
A. primus which occurred during Chron C3Br.2r based onanalyses of the stratotypic section (Hilgen et al., 2000; 7.341–
7.170 Ma). The second one is the FAD of A. delicatus assignedto Chron C3Br.1r (Hilgen et al., 2000; 7.135–7.091 Ma). ThenR. rotaria var. A, whom FAD (sample 21) is close to the FAD ofA. delicatus (Theodoridis, 1984; Flores et al., 1992; Lancis,1998; Lourens et al., 2004) and can be considered coincident.The FAD of N. amplificus in the equatorial Pacific is foundclose to the reversal between Chrons C3Ar and C3An.2n (Raffiand Flores, 1995; 6.567 Ma). The T. rugosus-N. amplificusintegrate forms appear slightly before N. amplificus in the upperpart of Chron C3Ar, and then disappear in the upper middle partof Chron C3An.2n (Berggren et al., 1995; Raffi and Flores,1995; Raffi et al., 1995, 2003, 2006; Backman and Raffi, 1997).Finally, the R. pseudoumbilicus > 7 mm PE occurs in thetransition between C3Bn and C3Ar (Raffi et al., 2006).
Three main discontinuities are found in the Rio Chicamosection. The first one appears at the base of the Tale GypsumFm and the next two at the bottom and top of the WichmannConglomerate Bed. The lower discontinuity separates samplesof normal polarity but the other two separate an unclear (grey)sample and a normal interval. None of the previous worksconsider specifically the possibility of any missing magneto-zone or hiatus associated to the discontinuities. However,taking into account the stratigraphic data, it seems likely thatthe discontinuities associated with the Wichmann Conglom-erate Bed could involve some hiatus. Consequently, in ourpreferred correlation of magnetozones, Chron C3r and/orChron C3An.2n could only be partly registered.
The LO of A. primus is located in the upper part of the SanelMb, not sampled in the Rio Chicamo section but sampled in theSebos section, which has no magnetostratigraphic data.Considering that the FAD of A. primus has been reported inChron C3Br.2r (Berggren et al., 1995; Raffi and Flores, 1995;Negri et al., 1999; Hilgen et al., 2000; Lourens et al., 2004) itcould be argued that the normal chron at the base of theChicamo section (Dinarès-Turell et al., 1999; Krijgsman et al.,2000) corresponds to Chron C3Bn or encompasses theC3Br.1n-C3Bn interval (Fig. 10).
Thus, the normal magnetozone of the upper Sanel Mb andthe Tale Gypsum Fm corresponds to the C3Bn plus eventuallythe C3Br.1n, and, in that case, the short reverse Chron C3Br.1rwas missing in the magnetostratigraphies by Dinarès-Turellet al. (1999) and Krijgsman et al. (2000). We note that someunreliable reverse samples exist a few meters below the base ofthe Tale Gypsum (Chicamo Gypsum) in the magnetostrati-graphic succession of Dinarès-Turell et al. (1999), which couldperhaps represent Chron C3Br.1r, although its position in theupper part of the thick normal magnetozone would in principleclash with the duration of the involved chrons in the standardGPTS (Fig. 10). However, the discontinuity at the base of theTale Gypsum could involve a hiatus and therefore part of anormal magnetozone (or even Chron C3Br.1r) could bemissing. Overall, it is not possible to precisely locate theTortonian/Messinian boundary with the available data and thus,we have tentatively located it at the base of the Tale GypsumFm in our figures.
Assuming the presence of A. delicatus from the lowermostpart of the Chicamo Cycles Fm, the normal/reverse inversionjust below the LO of A. delicatus could be correlated with theinversion C3Bn/C3Ar (second calibration line from the bottomin Fig. 10).
If the LO of N. amplificus is then considered, the ChicamoCycles Fm/Wichmann Bed upper reverse/normal inversioncorresponds to the C3Ar/C3An.2n reversal (third calibrationline in Fig. 10). Consequently, the reverse magnetozone of theChicamo Cycles Fm can be unambiguously calibrated to ChronC3Ar.
The upper magnetostratigraphic correlation of the RioChicamo section results by considering the Rambla Salada Fmas only Messinian (Garcés et al., 2001). In accordance, andfollowing a straightforward correlation to the GPTS, themagnetozones on top of the Wichmann Bed can be assigned asC3An.2n, C3Ar.1r, C3An.1n, and C3r, respectively (Fig. 10).
6. Discussion and conclusions
The Fenazar Conglomerate Bed and the Lower Gypsum Mbmark the beginning of the TSC, the first restriction phase of theFortuna Basin, and not the Tale Gypsum Fm as previouslyconsidered by Krijgsman et al. (2000).
The age of the Fortuna Basin evaporitic deposits has been apoint of discussion for many years (Fig. 3). Dinarès-Turell et al.(1999) calibrated their magnetostratigraphic data using theCasa del Acero micromammal fauna, assigned to BiozoneMN12 (Agustí et al., 1985), for which the upper limit wasestablished as during Chron C3Ar (6.8 Ma, as considered byDinarès-Turell et al., 1999), corresponding to their optionB. For option A, they considered the numerical dating of thevolcanic rocks from the Cabezos Negros lamproites (Fig. 5) byBellon et al. (1983) at 6.16 � 0.3 Ma. Finally, Krijgsman et al.(2000) assigned the Tale Gypsum Fm to the Late Tortonian,considering a good magnetozone pattern to fit with the GPTSand the Casa del Acero site older than 6.8 Ma because in levelsof other sections of the same Fortuna Basin containingMN13 fauna a younger age was assigned by the palaeomagnet-ism (Garcés et al., 1998).
These important differences were due to the poor quality ofthe planktonic foraminifera and nannofossils registered in thesection, and to the recalibration of the mammal BiozoneMN12 of the Casa del Acero site with the GTPS (divergencebetween the Dinarès-Turell et al. (1999) and Krijgsman et al.(2000) interpretations).
Our nannoplankton biostratigraphical data permit a relativedating of the evaporitic events: the age of the first evaporiticevent is between the Late Tortonian (on top of the Fortuna Fm)and the Messinian/Tortonian boundary, placed probably nearthe bottom of the Tale Gypsum Fm (second evaporitic event),and finally the Chicamo Cycles Fm (third evaporitic event) isincluded in the Early Messinian.
These data also permit the recalibration of the previouslypublished magnetostratigraphic data (Dinarès-Turell et al.,1999; Krijgsman et al., 2000). The lowermost normalmagnetozone, including the top of the Sanel Mb, Tale Gypsum
C. Lancis et al. / Geobios 43 (2010) 201–217214
Fm, and the bottom part of the Chicamo Cycles Fm, correspondto Chrons C3Br.1n and C3Bn lacking the intermediate shortreverse Chron C3Br.1r. Chron C3Ar is identified in the rest ofthe Chicamo Cycles Fm. The Wichmann Bed, marking animportant desiccation event in the Fortuna Basin evolution,occurs during Chron C3Ar. The top of the Wichmann Bed, canbe calibrated as the reversal from Chron C3An.2n to C3r, allwithin the Messinian. This recalibration coincides mainly withthe option A of Dinarès-Turell et al. (1999).
This new calibration of the Rio Chicamo magnetostrati-graphy is compatible with that of Bellon et al. (1983), who gave6.16 � 0.3 Ma as the absolute dating of the Cabezos Negroslamproite, but this dating is incompatible with new absolutedating (7.71 � 0.11 Ma) by Kuiper et al. (2006). Thisincompatibility needs to be studied in detail. As stated byPlayà and Gimeno (2006), ‘‘without a more detailed radio-metric study of the volcanic rocks of Fortuna it is impossible toconfirm or to exclude a second volcanic event at CabezosNegros’’.
A recent magnetobiostratigraphical mammal scale calibra-tion for the Neogene by Agustí et al. (2001), correlating theMN12 unit with the topmost C4n.1n to the middle part of theC3Br.2r (7.44–7.22, Late Tortonian), also appears incompatiblewith our proposed calibration. It is clear that the mammal andmagnetostratigraphic intercalibration along this time intervalwould deserve a full in-depth consideration, which is beyondthe scope of the present paper. Nevertheless, the resolution ofthe nannoplankton biochronological scale appears higher thanthe mammal scale and permits a more reliable calibration.
Acknowledgements
This work have been supported by the projects CGL2006-08848, CGL2007-65832/BTE, CGL2006-10593, and GV06/093, and the research group GRUPOS03/085 (GeneralitatValenciana). The authors express their thanks to J.A. Flores,M. Cachao, I. Raffi, and the editor F. Quillévéré for theirvaluable suggestions and critical comments of this work. Weare indebted to David Nesbitt for correcting the English versionof the paper.
Appendix A
Taxonomic list of the calcareous nannoplankton of theFortuna Basin considered in this work (Young, 1998):
Dictyococcites antarcticus Haq, 1976D. productus (Kamptner, 1963). Backman, 1980Discoaster berggrenii Bukry, 1971D. brouweri Tan Sin Hok, 1927 emend. Bramlette and
Riedel, 1954D. calcaris Gartner, 1967D. intercalaris Bukry, 1971D. loeblichii Bukry, 1971D. neorectus Bukry, 1971D. pseudovariabilis Martini and Worsley, 1971D. quinqueramus Gartner, 1969D. surculus Martini and Bramlette, 1963D. variabilis Martini and Bramlette, 1963Geminilithella jafarii (Müller, 1974) Backman, 1980G. rotula (Kamptner, 1948) Backman, 1980Helicosphaera carteri (Wallich, 1877) Kamptner, 1954Nicklithus amplificus Raffi, Backman and Rio,
1998 = Amaurolithus amplificus (Bukry and Percival, 1971)Gartner and Bukry, 1975
Pontosphaera multipora (Kamptner, 1948) Roth, 1970Reticulofenestra gelida (Geitzenauer, 1972) Backman, 1978R. haqii Backman, 1978R. minuta Roth, 1970R. minutula (Gartner, 1967) Haq and Berggren, 1978R. pseudoumbilicus (Gartner, 1967) Gartner, 1969R. rotaria Theodoridis, 1984Scyphosphaera amphora Deflandre, 1942Scy. apsteinii Lohmann, 1902Scy. campanula Deflandre, 1942Scy. cantharellus Kamptner, 1955Scy. lagena Kamptner, 1955Scy. pulcherrima Deflandre, 1942Scy. turris Kamptner, 1955Sphenolithus abies Deflandre in Deflandre and Fert, 1954Sph. moriformis (Brönnimann and Stradner, 1960) Braml-
ette and Wilcoxon, 1967Sph. neoabies Bukry and Bramlette, 1969Syracosphaera pulchra Lohmann, 1902Triquetrorhabdulus rugosus Bramlette and Wilcoxon, 1967.Small reticulofenestrids (nannolith size smaller than 3 mm,
independently of central area properties): Dictyococcitesproductus (Kamptner, 1963) emend. Backman, 1980 plusR. minuta Roth, 1970)
References
Aguirre, J., Sánchez-Almazo, I.M., 2004. The Messinian post-evaporitic depos-its of the Gafares area (Almería-Níjar basin SE Spain). A new view of the‘‘Lago-Mare’’ facies. Sedimentary Geology 168, 71–95.
Agustí, J., Cabrera, L., Garcés, M., Krijgsman, W., Oms, O., Parés, J.M., 2001.A calibrated mammal scale for the Neogene of Western Europe State of theart. Earth-Science. Reviews 52, 247–260.
Agustí, J., Moyà-Solà, S., Gibert, J., Guillén, J., Labrador, M., 1985. Nuevosdatos sobre la bioestratigrafía del Neógeno continental de Murcia. Paleon-tología i Evolució. 18, 83–94.
Aubry, M.P., 1992. Late Paleogene calcareous nannoplankton evolution: a taleof climatic deterioration. In: Prothero, D.R., Berggren, W.A. (Eds.), Eo-cene-Oligocene Climatic and Biotic Evolution. Princeton University Press,Oxford, pp. 272–309.
C. Lancis et al. / Geobios 43 (2010) 201–217 215
Azéma, J., Montenat, C., 1975. Mapa y memoria explicativa de la hoja de 892(Fortuna) del Mapa geológico Nacional a escala 1:50.000. Instituto Geoló-gico y Minero de España, Madrid.
Backman, J., 1978. Late Miocene-Early Pliocene nannofossil biochronologyand biogeography in the Vera Basin SE Spain. Stockholm Contributions inGeology 32, 93–114.
Backman, J., Pestiaux, P., Zimmerman, H., Hermelin, O., 1986. Paleoclimaticand Paleogeographic development in the Pliocene North Atlantic: Discoas-ter accumulation and coarse fraction data. In: Summerhayes, C.P., Shack-leton, N.J., Hattori, T. (Eds.), North Atlantic Paleoceanography. GeologicalSociety Special Publication 21, London, pp. 231–242.
Backman, J., Raffi, I., 1997. Calibration of Miocene nannofossil eventsto orbitally-tuned cyclostratigraphies from Ceara Rise. In: Shackleton,N.J., Curry, W.B., Richter, C., Bralower, T.J. (Eds.), Proceedings of theOcean Drilling Program, Scientific Results 154, College Station, TX, pp.83–99.
Bellon, H., Bordet, P., Montenat, C., 1983. Chronologie du magmatismenéogène des Cordillères bétiques (Espagne méridionale). Bulletin de laSociété géologique de France 25, 205–217.
Benson, R.H., Rakic-El Bied, K., 1996. The Bou Regreg Section Morocco:Proposed Global Boundary Stratotype Section and Point of the Pliocene.Notes et Mémoires du Service géologique du Maroc 383, 51–150.
Berggren, W.A., Kent, D.V., Swisher III, C.C., Aubry, M.-P., 1995. A RevisedCenozoic Geochronology and Chronostratigraphy. In: Berggren, W.A.,Kent, D.V., Aubry, M.-P., Hardenbol, J. (Eds.), Geochronology, time scalesand global stratigraphic correlation. SEPM Society for Sedimentary Geol-ogy, Special Publication 54, Tulsa, pp. 129–212.
Bukry, D., 1971. Cenozoic calcareous nannofossils from the PacificOcean. Transactions of the San Diego Society Natural History 16, 303–
328.Bukry, D., 1976. Coccolith stratigraphy of Manihiki Plateau, Central Pacific,
Deep Sea Drilling Project, site 317. In: Schlanger, S.O., Jackson, E.D. et al.(Eds.),Initial Reports of the Deep Sea Drilling Project 33, U. S.GovernmentPrinting Office, Washington, pp. 493–499.
Bukry, D., 1981. Cenozoic Coccoliths from the Deep Sea Drilling Project.SEPM Society for Sedimentary Geology, Special Publication 32, Tulsa,335–353.
Bukry, D., Percival, S.E., 1971. New Tertiary calcareous nannofossils. TulaneStudies in Geology and Paleontology 8, 123–146.
Caracuel, J.E., Soria, J.M., Yébenes, A., 2004. Early Pliocene transgressivecoastal lags (Bajo Segura Basin Spain): a marker of the flooding after theMessinian salinity crisis. Sedimentary Geology 169, 121–128.
De Larouzière, F.D., Bolze, J., Bordet, P., Hernandez, J., Montenat, C., Ottd’Estevou, P., 1988. The Betic segment of the lithospheric Trans-Alboranshear zone during the Late Miocene. Tectonophysics 152, 41–52.
Dinarès-Turell, J., Ortí, F., Playà, E., Rossell, L., 1999. Palaeomagneticchronology of the evaporitic sedimentation in the Neogene Fortuna Basin(SE Spain): early restriction preceding the ‘Messinian Salinity Crisis’.Palaeogeography, Palaeoclimatology. Palaeoecology 154, 161–178.
Feldmann, M., McKenzie, J., 1997. Messinian stromatolite-thrombolite asso-ciations, Santa Pola SE Spain: an analogue for the Palaeozoic? Sedimen-tology 44, 893–914.
Flores, J.A., 1985. Nanoplancton calcáreo en el Neógeno del borde norocci-dental de la Cuenca del Guadalquivir (S.O. de España). Ph.D. Thesis,Universidad Salamanca (unpublished).
Flores, J.A., Sierro, F.J., 1987. Calcareous plankton in the Tortonian-Messiniantransition of the Northwestern edge of the Guadalquivir Basin (SW Spain).Abhandlungen der Geologischen Bundesanstalt 39, 64–87.
Flores, J.A., Sierro, F.J., 1989. Calcareous nannoflora and planktonicforaminifera in the Tortonian/Messinian boundary interval of East AtlanticDSDP sites and their relation to Spanish and Moroccan sections. In: VanHeck, S., Crux, J. (Eds.), Nannofossils and their applications. Ellis Hor-wood, London, pp. 249–266.
Flores, J.A., Sierro, F.J., Gabriel, M., Filippelli, G.M., Bárcena, M.A., Pérez-Folgado, M., Vázquez, A., Utrilla, R., 2005. Surface water dynamics andphytoplankton communities during deposition of cyclic late Messiniansapropel sequences in the western Mediterranean. Marine Micropaleontolo-gy 56, 50–79.
Flores, J.A., Sierro, F.J., Glaçon, G., 1992. Calcareous plankton analysis in thepre-evaporitic sediments of the ODP site 654 (Tyrrhenian Sea WesternMediterranean). Micropaleontology 38, 279–288.
Flores, J.A., Sierro, F.J., Raffi, I., 1995. Evolution of the calcareous nannofossilassemblage as a response to the paleoceanographic changes in the easternequatorial Pacific Ocean from 4 to 2 Ma (Leg 138 Sites 849 and 852). In:Pisias, N.G., Mayer, L.A., Janecek, T.R., Palmer-Julson, A., van Andel,T.H. (Eds.), Proceeding of the Ocean Drilling Program, Scientific Results138, College Station, TX, pp. 163–176.
Fortuin, A.R., Kelling, J.M.D., Roep, T.B., 1995. The enigmatic Messinian-Pliocene section of Cuevas del Almanzora (Vera Basin SE Spain) revisited-erosional features and strontium isotope ages. Sedimentary Geology 97,177–201.
Fortuin, A.R., Krijgsman, W., 2003. The Messinian of the Nijar Basin (SESpain): sedimentation, depositional environments and paleogeographicevolution. Sedimentary Geology 160, 213–242.
Fortuin, A.R., Krijgsman, W., Hilgen, F.J., Sierro, F.J., 2000. Late MioceneMediterranean desiccation: topography and significance of the ‘SalinityCrisis’ erosion surface on-land in southeast Spain: Comment. SedimentaryGeology 133, 167–174.
Fúster, J.M., Gastesi, P., Sagredo, J., Fermosi, M.L., 1967. Las rocas lamproí-ticas del SE de España. Estudios Geológicos 23, 35–69.
Garcés, M., Krijgsman, W., Agustí, J., 1998. Chronology of the late Turoliandeposits of the Fortuna Basin (SE Spain): implications for the Messinianevolution of the eastern Betics. Earth Planetary Science Letters 163, 69–81.
Garcés, M., Krijgsman, W., Agustí, J., 2001. Chronostratigraphic frameworkand evolution of the Fortuna basin (Eastern Betics) since the Late Miocene.Basin Research 13, 199–216.
Gartner, S., 1992. Miocene nannofossil chronology in the North Atlantic DSDPSite 608. Marine Micropaleontology 18, 307–331.
Gartner, S., Bukry, D., 1975. Morphology and phylogeny of the Coccolitho-phycean family Ceratolithaceae. United States Geological Survey. JournalResearch 3, 451–465.
Gartner, S., Chen, M.P., Stanton, R.J., 1979. Late Neogene history in theAmerican Mediterranean. VII International Congress Mediterranean Neo-gene Athens. Annales de Géologie des Pays Helléniques 1, 425–437.
Haq, B.U., 1980. Biogeographic history of Miocene calcareous nannoplanktonand paleoceanography of the Atlantic Ocean. Micropaleontology 26, 414–
443.Hekel, H., 1973. Nannofossil biostratigraphy, Leg 20, Deep Sea Drilling
Project. In: Heezen, B.C., MacGregor, I.D., Foreman, H.P., Forristal, G.,Hekel, H., Hesse, R., Hoskins, R.H., Jones, E.J.W., Kaneps, A., Krashe-ninnikov, V.A., Okada, H., Ruef, M.H. (Eds.), Initial Reports of the DeepSea Drilling Project 20, U.S. Government Printing Office, Washington,pp. 221–247.
Hilgen, F.J., Bissoli, L., Iaccarino, S., Krijgsman, W., Meijer, R., Negri, A.,Villa, G., 2000. Integrated stratigraphy and astrochronology of the Messi-nian GSSP at Oued Akrech (Atlantic Morocco). Earth and PlanetaryScience Letters 182, 237–251.
Hilgen, F.J., Krijgsman, W., Langereis, C.G., Lourens, L.J., Santerelli, A.,Zachariasse, W.J., 1995. Extending the astronomical (polarity) time scaleinto the Miocene. Earth and Planetary Science Letters 136, 495–510.
Krijgsman, W., Garcés, M., Agustí, J., Raffi, I., Taberner, C., Zachariasse, W.J.,2000. The ‘Tortonian salinity crisis’ of the eastern Betics (Spain). Earth andPlanetary Science Letters 181, 497–511.
Krijgsman, W., Hilgen, F.J., Fortuin, A., Sierro, F.J., 2001. Astrochronology forthe Messinian Sorbas Basin (SE Spain) and orbital (precessional) forcing forevaporite cyclicity. Sedimentary Geology 140, 43–60.
Krijgsman, W., Leewis, M.E., Garcés, M., Kouwenhoven, T.J., Kuiper, K.F.,Sierro, F.J., 2006. Tectonic control for evaporite formation in the EasternBetics (Tortonian; Spain). Sedimentary Geology 188/189, 155–170.
Kuiper, K.F., Krijgsman, W., Garcés, M., Wijbrans, J.R., 2006. Revised isotopic(40Ar/39Ar) age for the lamproite volcano of Cabezos Negros, Fortuna Basin(Eastern Betics, SE Spain). Palaeogeography Palaeoclimatology Palaeoe-cology 238, 53–62.
Lancis, C., 1998. El nanoplancton calcáreo de las cuencas neógenas orientalesde la Cordillera Bética. Ph.D. Thesis, Universidad Alicante, http://hdl.han-dle.net/10045/7944.
Lukowski, P., Poisson, A., 1990. Le bassin de Fortuna. In: Montenat, C. (Ed.),Les Bassins Néogènes du Domaine Bétique Oriental (Espagne). Docu-ments et Travaux, Institut géologique Albert de Lapparent 12/13, pp. 303–
311.Mansouri, M.E.H., Bessedik, M., Aubry, M.-P., Belkebir, L., Mansour, B.,
Beaufort, L., 2008. Contributions biostratigraphiques et paléoenvironne-mentales de l’étude des nannofossiles calcaires des dépôts tortono-messi-niens du bassin du Chélif (Algérie). Geodiversitas 30, 59–77.
Martín, J.M., Braga, J.C., 1994. Messinian events in the Sorbas basin ofSoutheastern Spain and their implications on the recent history the Medi-terranean. Sedimentary Geology 90, 257–268.
Martín-Pérez, J.A., 1997. Nanoplancton calcáreo del Mioceno de la CordilleraBética (Sector Oriental). Ph.D. Thesis, Universidad Granada (unpublished).
Martini, E., 1965. Mid-Tertiary calcareous nannoplankton from Pacific deep-seacores. Submarine Geology and Geophysics. In: Whittard, W.F., Bradshaw,R.B. (Eds.), Proceedings of the 17th Symposium Colston Research Society,London, pp. 393–411.
Martini, E., 1971. Standard Tertiary and Quaternary calcareous nannoplanktonzonation. In: Farinacci, A. (Ed.), Proceedings of second Planktonic confer-ence 2, Tecnoscienza, Roma, pp. 736–785.
Mazzei, R., 1985. The Miocene sequence of the Maltese islands: biostrati-graphic and chronostratigraphic reference based on nannofossils. Atti dellaSocietà Toscana di Scienze Naturali 92, 165–197.
Mazzei, R., Raffi, I., Rio, D., Hamilton, N., Cita, M.B., 1979. Calibration of lateNeogene calcareous plankton datum planes with the paleomagnetic recordof Site 397 and correlation with Moroccan and Mediterranean sections. In:von Rad, U., Ryan, W.B.F. et al., (Eds.), Initials Reports of the Deep SeaDrilling Project 47, U.S. Government Printing Office, Washington, pp. 375–
389.Montenat, C., 1977. Les bassins néogènes du levant d’Alicante et de Murcia
(Cordillères bétiques orientales Espagne). Stratigraphie, paléogéographie etévolution dynamique. Documents du Laboratoire de Géologie de la Facultédes Sciences de Lyon 69, 1–345.
Montenat, C., Ott d’Estevou, Ph., Coppier, G., 1990. Les basins néogènes entreAlicante et Cartagena. In: Montenat, C. (Ed.), Les Bassins Néogènes duDomaine Bétique Oriental (Espagne). Documents et Travaux, InstitutGéologique Albert de Lapparent 12/13, pp. 313–368.
Morigi, C., Negri, A., Giunta, S., Kouwenhoven, T., Krijgsman, W., Blanc-Valleron, M.-M., Orszag-Sperber, F., Rouchy, J.-M., 2007. Integratedquantitative biostratigraphy of the latest Tortonian-early Messinian Pissourisection (Cyprus): An evaluation of calcareous plankton bioevents. Geobios40, 267–279.
Müller, C., 1974. Calcareous nannoplankton, Leg 25 (Western Indian Ocean).In: Simpson, E.S.W., Schlich, R. et al. (Eds.), Initials Reports of the DeepSea Drilling Project 25, U.S. Government Printing Office, Washington,pp. 579–633.
Müller, C., 1978. Neogene calcareous nannofossils from the Mediterranean –
Leg 42A of the Deep Sea Drilling Project. In: Hsü, K.J., Montadert, L. et al.(Eds.). Initials Reports of the Deep Sea Drilling Project 42, U.S. Govern-ment Printing Office, Washington, pp. 727–751.
Müller, C., 1990. Nannoplankton biostratigraphy and paleoenvironmentalinterpretations from the Tyrrhenian Sea, ODP Leg 107 (Western Mediter-ranean). In: Kastens, K.A., Mascle, J. et al., (Eds.), Proceedings of theOcean Drilling Program, Scientific Results 107, College Station, TX,p. 495–511.
Müller, D.W., Hsü, K.J., 1987. Event stratigraphy and paleoceanography in theFortuna basin (southeast Spain): a scenario for the Messinian salinity crisis.Paleoceanography 2, 679–696.
Negri, A., Giunta, S., Hilgen, F., Krijgsman, W., Vai, G.B., 1999. Calcareousnannofossil biostratigraphy of the M. del Casino section (northern Apen-nines Italy) and paleoceanographic conditions at times of Late Miocenesapropel formation. Marine Micropaleontology 36, 13–30.
Negri, A., Villa, G., 2000. Calcareous nannofossil biostratigraphy biochronol-ogy and paleoecology at the Tortonian/Messinan boundary of the Faner-omeni section (Crete). Palaeogeography, Palaeoclimatology, Palaeoecology156, 195–209.
Okada, H., Bukry, D., 1980. Supplementary modification and introduction ofcode numbers to the low-latitude coccolith biostratigraphic zonation(Bukry, 1973, 1975). Marine Micropaleontology 5, 321–325.
Parker, M.E., Clark, M., Wise, S.W. Jr., 1985. Calcareous nannofossils of DeepSea Drilling Project Sites 558 and 563, North Atlantic Ocean: biostratigra-phy and the distribution of Oligocene braarudosphaerids. In: Bougault, H.,Cande, S.C. et al., (Eds.), Initial Reports of the Deep Sea Drilling Project 82,U.S. Government Printing Office, Washington, pp. 559–589.
Perch-Nielsen, K., 1977. Albian to Pleistocene calcareous nannofossils from thewestern South Atlantic, DSDP Leg 39. In: Supko, R.R., Perch-Nielsen, K.et al. (Eds.), Initial Reports of the Deep Sea Drilling Project 39, U.S.Government Printing Office, Washington, pp. 699–823.
Perch-Nielsen, K., 1985. Cenozoic calcareous nannofossils. In: Bolli, H.M.,Saunders, J.B., Perch-Nielsen, K. (Eds.), Plankton Stratigraphy. CambridgeUniversity Press, Cambridge, pp. 427–554.
Playà, E., Gimeno, D., 2006. Evaporite deposition and coeval volcanism in theFortuna Basin. Sedimentary Geology 188/189, 205–218.
Playà, E., Ortí, F., Rosell, L., 2000. Marine to non-marine sedimentation in theupper Miocene evaporites of the Eastern Betics SE Spa sedimentologicalgeochemical evidence. Sedimentary Geology 133, 135–166.
Proto Decima, F., Medizza, F., Todesco, L., 1978. Southeastern Atlantic Leg40 calcareous nannofossils. In: Bolli, H.M., Ryan, W.B.F. et al., (Eds.),Initial Reports of the Deep Sea Drilling Project 40, U.S. GovernmentPrinting Office, Washington, pp. 571–634.
Raffi, I., Backman, J., Fornaciari, E., Pälike, H., Rio, D., Lourens, L., Hilgen, F.,2006. A review of calcareous nannofossil astrobiochronology encompass-ing the past 25 million years. Quaternary Science Reviews 25, 3113–3137.
Raffi, I., Backman, J., Rio, D., 1998. Evolutionary trends of calcareousnannofossils in the Late Neogene. Marine Micropaleontology 35, 17–41.
Raffi, I., Flores, J.A., 1995. Pleistocene through Miocene Calcareous nanno-fossils from eastern equatorial Pacific Ocean (Leg 138). In: Proceedings ofthe Ocean Drilling Program, Scientific Results 138, College Station, TX,pp. 233–285.
Raffi, I., Mozzato, C., Fornaciari, E., Hilgen, F.J., Rio, D., 2003. Late Miocenecalcareous nannofossil biostratigraphy and astrobiochronology for theMediterranean region. Micropaleontology 49, 1–26.
Raffi, I., Rio, D., 1979. Calcareous Nannofossils biostratigraphy of DSDP site132 Leg 13 (Tyrrhenian sea), western Mediterranean. Rivista Italiana diPaleontologia e Stratigrafia 85, 127–172.
Raffi, I., Rio, D., d’Afri, A., Fornaciari, E., Rochetti, S., 1995. Quantitativedistribution patterns and biomagnetostratigraphy of middle and late Mio-cene calcareous nannofossils from equatorial Indian and Pacific Ocean (legs115, 130 and 138). In: Pisias, N.G., Mayer, L.A., Janacek, T.R. et al. (Eds.),Proceedings of the Ocean Drilling Program, Scientific Results 138, CollegeStation, TX, pp. 479–502.
Riding, R., Braga, J.C., Martín, J.M., 1999. Late Miocene Mediterraneandesiccation: topography and significance of the ‘‘Salinity Crisis’’ erosionsurface on-land in southeast Spain. Sedimentary Geology 123, 1–7.
Riding, R., Braga, J.C., Martín, J.M., 2000. Late Miocene Mediterraneandesiccation: topography and significance of the ‘‘Salinity Crisis’’ erosionsurface on-land in southeast Spa Reply. Sedimentary Geology 133, 175–
184.Riding, R., Braga, J.C., Martín, J.M., Sánchez-Almazo, I.M., 1998. Mediterra-
nean Messinian Salinity Crisis: constraints from a coeval marginal basinSorbas, southeastern Spain. Marine Geology 146, 1–20.
Rio, D., Fornaciari, E., Raffi, I., 1990. Late Oligocene through early Pleistocenecalcareous nannofossils from western equatorial Indian Ocean (Leg 115).In: Duncan, R.A., Backman, J., Peterson, L.C. et al. (Eds.), Proceedings ofthe Ocean Drilling Program, Scientific Results 115, College Station, TX,pp. 175–235.
C. Lancis et al. / Geobios 43 (2010) 201–217 217
Rio, D., Mazzei, R., Palmieri, G., 1976. The stratigraphic position of theMediterranean Upper Miocene evaporites, based on nannofossils. Memoriedella Societá Geologica Italiana 16, 261–276.
Santistebán Bové, C., 1981. Petrología y sedimentología de los materiales delMioceno superior de la cuenca de Fortuna (Murcia), a la luz de la crisis desalinidad. Ph.D. Thesis, Universidad Barcelona (unpublished).
Santistebán, C., Taberner, C., 1983. Shallow marine and continental conglom-erates derived from coral reef complexes after desiccation of a deep marinebasin: the Tortonian-Messinian deposits of the Fortuna Basin SE Spain.Journal the Geological Society of London 140, 401–411.
Sierro, F.J., 1985. The replacement of the Globorotalia menardii group by theGloborotalia miotumida group: an aid to recognizing the Tortonian/Messi-nian boundary in the Mediterranean and adjacent Atlantic. Marine Micro-paleontology 9, 525–535.
Sierro, F.J., Hilgen, F.J., Krijgsman, W., Flores, J.A., 2001. The Abadcomposite (SE Spain): a Mediterranean and global reference section forthe Messinian. Palaeogeography, Palaeoclimatology, Palaeoecology 168,141–169.
Siesser, W.G., 1977. Calcareous nannofossils as age and paleoenvironmentalindicators Electron van. Sudelike Afrika. In: Proceedings V7, ElectronMicroscopy, Society South Africa, pp. 81–82.
Soria, J.M., Caracuel, J.E., Corbí, H., Dinarès-Turell, J., Lancis, C., Tent-Manclús, J.E., Viseras, C., Yébenes, A., 2008. The Messinian-early Plio-cene stratigraphic record in the southern Bajo Segura Basin (Betic Cordil-lera Spain). Implications for the Mediterranean salinity crisis. SedimentaryGeology 203, 267–288.
Soria, J.M., Tent-Manclús, J.E., Caracuel, J.E., Yébenes, A., Lancis, A.,Estévez, A., 2005. La crisis de salinidad Tortoniense: su registro en lazona de enlace entre las cuencas de Fortuna y del Bajo Segura. Geo-Temas8, 113–118.
Soria, J.M., Yébenes, A., Caracuel, J.E., 2003. Estratigrafía del Messiniense yPlioceno en el margen norte de la Cuenca del Bajo Segura (Cordillera Bética
oriental) Cambios paleogeográficos asociados a la crisis de salinidad delMediterráneo. Geo-Temas 5, 219–223.
Takayama, T., 1993. Notes on Neogene calcareous nannofossil biostratigra-phy of the Ontong Java Plateau and size variations of Reticulofenestracoccoliths. In: Berger, W.H., Kroenke, L.W., Mayer, L.A., Janecek, T.R.(Eds.), Proceedings of the Ocean Drilling Program, Scientific Results130, College Station, TX, pp. 179–229.
Tent-Manclús, J.E., 2003. Estructura y estratigrafía de las sierras de Crevillente,Abanilla y Algayat: su relación con la Falla de Crevillente. Ph.D. Thesis,Universidad Alicante , http.//hdl.handle.net/10045/10414.
Tent-Manclús, J.E., Lancis, C., Soria, J.M., Dinarès-Turell, J., Estévez, A.,Caracuel, J.E., Yébenes, A., 2007. Primeros datos bioestratigráficos de losgrupos evaporíticos de la Cuenca de Fortuna (Cordillera Bética). Geogaceta41, 231–234.
Tent-Manclús, J.E., Soria, J.M., Estévez, A., Lancis, C., Caracuel, J.E., Dinarès-Turell, J., Yébenes, A., 2008. Tortonian Salinity Crisis and tectonics: Aviewfrom the Fortuna Basin (SE Spain). Comptes Rendus Geoscience 340, 474–
481.Theodoridis, S.A., 1984. Calcareous nannofossils biozonation of the Miocene
and revision of the Helicoliths and Discoasters. Utrecht Micropaleontologi-cal Bulletin 31, 1–271.
Viseras, C., Soria, J.M., Fernández, J., 2004. Cuencas neógenas postorogénicasde la Cordillera Bética. In: Vera, J.E. (Ed.), Geología de España. SGE-IGME, Madrid, pp. 576–581.