Page 1
1
The Musical Brain: a Study
into Musical Perception
and Memory
By Martha Nye
Canterbury Christ Church University
Department of Music and Performing Arts
Bachelor of Music
Word Count: 11, 552
Date of Submission: Friday 3rd
May 2013
Page 2
2
Abstract This paper explores the neurological basis for the perception and memory for pitch, pitch
relationships and melodies. The phenomenon that musicians with similar training and
expertise excel in different areas of music leads to a question: if the training is similar, then
how is trainee or his methods different? This research explores the difference between, and
reasons for aptitude in, aural recall and sight-reading. The study shows the brain areas and
processes specific to music and the flexibility of musicians’ methods. The findings lead to a
personal understanding of proficiency or ineptitude in these skills.
Page 3
3
Table of Contents
Introduction ...................................................................................................................................... 4
Musical Neurophysiology ..................................................................................................................... 5
Chapter 1 – Pitch and Melody ..................................................................................................... 8
Recognition of Pitch ............................................................................................................................. 8
Memory Systems for Pitch ................................................................................................................. 11
Absolute Pitch ..................................................................................................................................... 14
Melody and Pitch Recognition ........................................................................................................... 16
Immediate Recall of Melodies ............................................................................................................ 18
Chapter Summary ............................................................................................................................... 21
Chapter 2 – Melody and Harmony ........................................................................................... 23
Melodic Contour ................................................................................................................................. 23
Scale Structure and Tonality ............................................................................................................... 24
Musical Notation and Structure .......................................................................................................... 27
Chapter Summary ............................................................................................................................... 34
Chapter 3 – Music in Action ....................................................................................................... 36
Sight-Reading and Working Memory ................................................................................................. 36
Imagery and Mental Rehearsal ........................................................................................................... 37
Brain Function and Structure .............................................................................................................. 40
Chapter Summary ............................................................................................................................... 40
Conclusion ...................................................................................................................................... 42
References ....................................................................................................................................... 44
Page 4
4
Introduction From the age of 18, my interest in brain organisation, function and mechanisms has grown,
especially in the field of music. Music has an emotional effect, an automatic effect and often
an unnoticed effect on our thoughts and feelings. The fact that music seems a phenomenon,
separate and far less understood from other brain studies, has appealed to me and led me to
search into the anatomical relationships of musical activity in the brain. My first interest was
that of ability – why could I easily pick up instruments, the reading of musical notation and
the act of singing when my peers, exposed to a similar level of musical activity, could not?
Why can I, after a similar amount and intensity of training to my sister, sight-read to near
perfection when she cannot? Yet she excels in aural tests, such as immediate recall of
melody and rhythm, and I perform poorly. What is it about our brains and their structure and
organisation that leads us to excel in different aspects of music? Which parts of my brain
have developed further or lesser to my sibling’s?
The study of memory has long interested me, both inside and outside of music, and
memory is intrinsically linked to the method and efficacy of perception and coding. A
particular issue of mine is that I cannot, and never have been able to, sing or play back a
melody I have just heard. As a music student I daily engage in many musical activities,
often spending a large proportion of my day exercising those musical structures in my brain.
I wanted to know which parts of my brain are stopping me either properly encoding or
remembering pitches and pitch sequences. The study of perception of melody may explain
to me where the deficiencies are. Perhaps the part of my brain which is lacking is also
implicated in other non-musical functions which are poor in performance, and this study
will lead me to understand where I excel and where I do not, and why.
Page 5
5
Musical Neurophysiology Music neurophysiology is an excellent tool for exploring brain structure, function and
organisation. Despite the fact that many become involved in musical training, only a small
percentage of the population develop a proficiency in music, which suggests that there is an
innate predisposition to musical skill or that something in a musician's brain plasticity is
different in nature to a non-musician’s (Peretz and Zatorre 2005). Similarly, the study of
brain organisation is a useful aid to the study of how the brain functions in music. Studies of
brain anomalies show the use of neural networks for music that are separated from non-
musical auditory and vocal functions. Using these two together we can discover some
neurological roots to a very human trait (Peretz and Zatorre 2005).
Many different parts of the brain are found to have been active during activities of
music processing. Just from the first sound entering the ear, structures like the cochlea,
brain stem, mid brain nuclei and mid brain cortex are activated and in a very short space of
time lead to perception. Different parts of the brain are involved in stages of the musical
processing sequence, and there has been evidence of segregation between these mechanisms
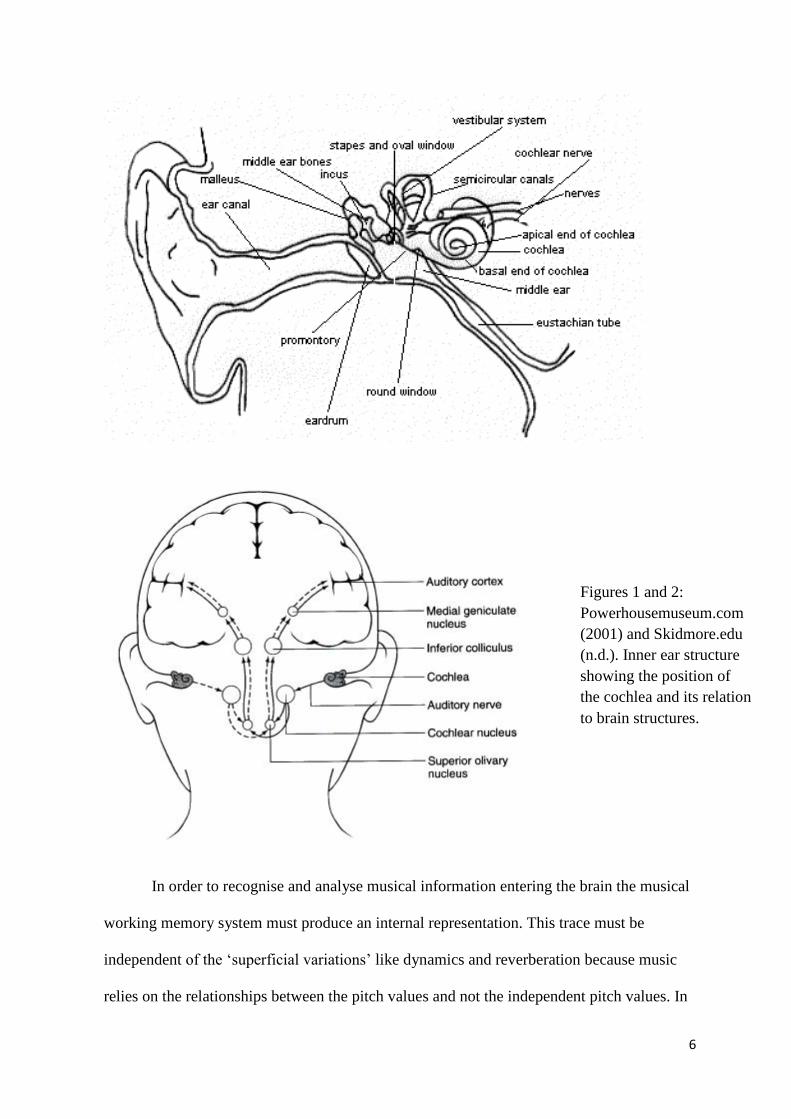
(Peretz and Zatorre 2005). The organisation of the systems within the cochlea (See Figures
1 and 2) appears to correspond to frequencies. The basal parts respond to high frequencies
while the apical parts respond to lower frequencies (Weinberger 1999). Music is processed
like any other auditory information, through the ascending auditory pathway to the auditory
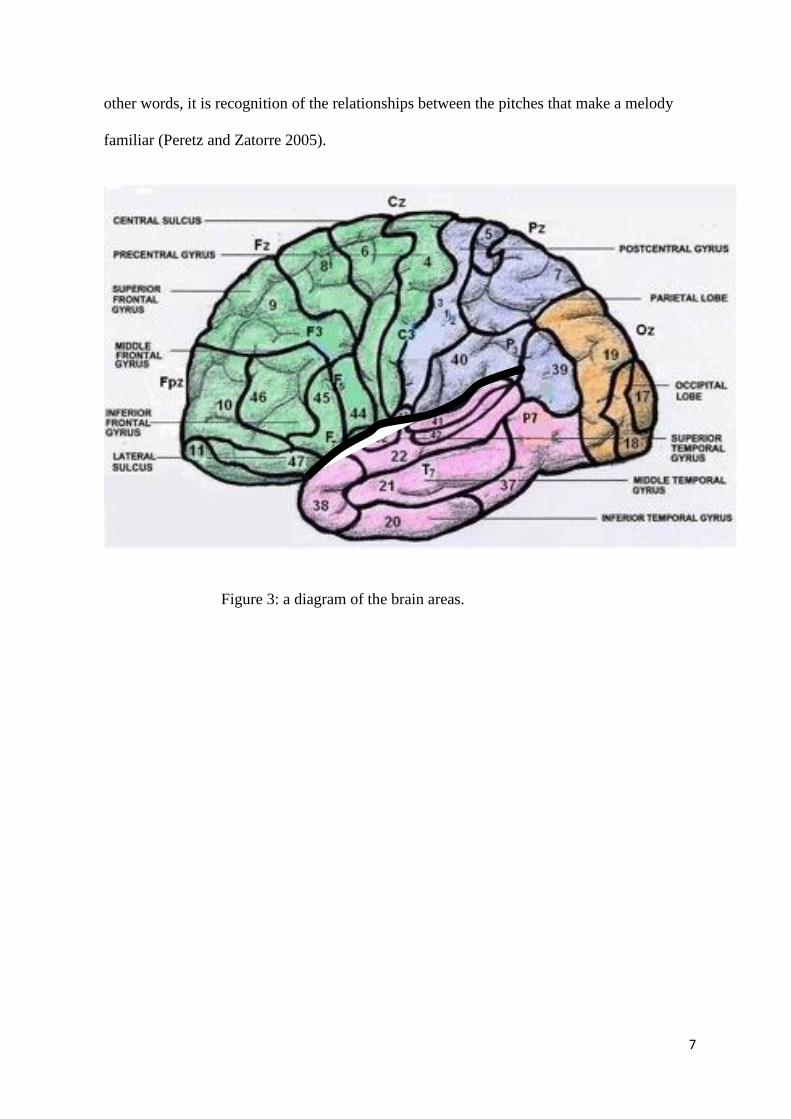
cortex. The cortex is on the superior temporal plane within the Sylvian fissure, demonstrated
by the thick line on Figure 3. The primary auditory cortex is in the superior temporal gyrus,
numbered 41 and 42 on the diagram (Figure 3). Laterally positioned to the primary auditory
cortex are the secondary auditory cortical areas (Stewart et al 2006).
Page 6
6
In order to recognise and analyse musical information entering the brain the musical
working memory system must produce an internal representation. This trace must be
independent of the ‘superficial variations’ like dynamics and reverberation because music
relies on the relationships between the pitch values and not the independent pitch values. In
Figures 1 and 2:
Powerhousemuseum.com
(2001) and Skidmore.edu
(n.d.). Inner ear structure
showing the position of
the cochlea and its relation
to brain structures.
Page 7
7
other words, it is recognition of the relationships between the pitches that make a melody
familiar (Peretz and Zatorre 2005).
Figure 3: a diagram of the brain areas.
Page 8
8
Chapter 1 – Pitch and Melody
Recognition of Pitch Pitch is one the fundamental elements of music, along with rhythm, and yet their processing
functions appear to be independent, whilst still relying on each other to make a melody.
Jones and Boltz found that in fully functioning brains, perception and memory of pitch is
quite rhythmical - perhaps aided by the rhythm – but that brain damage can cause a
deficiency in pitch comprehension and perception while the metric and rhythmic skills
remain intact (Peretz and Zatorre 2005, 91). This has been shown in a case study by
Fasanaro et al (1990). The subject was a 72-year-old professional musician playing several
instruments and was experienced in sight-reading. He suffered a loss of strength to his right
side and space-time disorientation. Neurological examinations showed mild right
hemiparesis, a weakness on one side of the body, including reduced sense of touch, sight
and a difficulty reading and naming. A brain scan showed decreased density in the
temporoparieto-occipital region (the junction between temporal, parietal and occipital
areas), with damage in the left medial region and splenium (in the very centre of the brain).
The following table (Table 1) shows how he performed in the musical ability tests and how
his pitch abilities are far decreased in comparison to his rhythmical abilities.
Page 9
9
A logical suggestion to make following this evidence is that pitch and rhythm
dimensions use separable subsystems within the musical mechanisms. Griffiths, Johnsrude,
Dean and Green (1999, in Peretz and Zatorre 2005) discovered similar activation patterns in
the brain in tests of pitch and time discrimination. Recall is never note perfect while it can
be harmonically and metrically consistent (Sloboda and Parker 2005). Evidence suggesting
that the operations are separable suggests that the brain areas that they do share are capable
of several simultaneous functions.
In studying how we compute pitch we can see that the right temporal neo-cortex,
located in the transverse temporal gyri, is heavily implicated. It is easy to see the function of
brain areas when comparing brain-damaged subjects to undamaged subjects. In a study by
Milner in 1962, those with focal brain excisions in the right temporal area were found to
have worse pitch relationship skills than those with excisions in the left. The impairment
was more than just a deficit in pitch discrimination, suggesting that it is implicated in more
than just this function (Peretz and Zatorre 2005, 91).
Table 1: Fasanaro
et al (1990, 268)
case study. A
table presenting
the results of the
subject’s musical
ability tests.
Page 10
10
In Heschl's gyri, (the area containing the temporal gyri in Figure 3), damage in the
right anterolateral section can cause further damage to pitch processing. Perception of notes
missing their fundamental is difficult, but subjects with this damage seem to be better at
determining the direction of pitch (Peretz and Zatorre, 2005). For recognition of a
frequency, the fundamental tone is not crucial as it can be identified without. For lower
notes, the high frequencies are more important and for high notes, the lower frequencies are
more dominant (Pierce 1999).
The posterior secondary auditory cortex may be involved in the analysis of pitch, as
it was shown to be active when comparing complex tones (a tone with harmonics), which
include some spectral change. Manipulation of this pitch leads to more response in the right
auditory regions, with manipulation of pitch height activating the posterior section and
analysis of pitch chroma (its qualities) taking place in the anterior section (Peretz and
Zatorre 2005, 92). All of this evidence points to the right secondary auditory cortex’s role in
processing of pitch relationships. This occurs even when attention is directed elsewhere as
Tervaniemi (2003) showed with electroencephalography (EEG) and magneto-
encephalography (MEG) scans, where activity in the brain was still occurring, although to a
lesser extent. In Tervaniemi’s studies, she found that ‘temporally and spectrally complex
sounds as well as their relations are automatically represented in the human auditory cortex’
(Tervaniemi 2003, 294) and that musical sounds are distinct from phonetic sounds in their
neural representations. The brain has an ability to encode and analyse complex sounds even
when the subject is focused on another task, and musical sounds continue to be differently
analysed (Tervaniemi 2003).
Pitch relationships include the intervals between them, and there are several means
of pitch and interval identification. ‘Magnitude Estimation’ occurs when the listeners group
close frequencies into the same magnitude (pitch) and then jump to another magnitude when
Page 11
11
the pitch gets too far out of range. ‘Absolute Identification’ is a skill held by those with
absolute pitch. These people can identify approximately 75 items, compared with those who
do not possess absolute pitch who can identify approximately 5 items. In interval
recognition for those with absolute pitch, ascending and descending intervals may be
perceived as the same, as it is merely the pitch categories that are needed for recognition.
The musicians tested could recognise the intervals (even those only with relative pitch) from
unison to a major tenth with 100% accuracy. ‘Category scaling’ meant the participants
shifted slightly narrower or wider intervals into equal temperament (Burns 1999, 221).
Memory Systems for Pitch Memory for pitch is a crucial part to our perception, comprehension and analysis of pitch
and melody. Whilst the distinct scale steps are not coded in the memory trace, they must be
perceived in order to make the representative trace, built on a framework of tonality and
contour (Dowling 1978). Music unfolds and develops over a time period, and therefore our
auditory mechanisms must maintain information correctly in order to relate certain elements
to those previous and subsequent. Deutsch (1970) found in her studies that the systems for
maintenance of pitch are dissociable from other memory parts, similarly to its perception as
we have seen above. A melody is analysed in terms of its pitch, dynamics, duration,
harmonic and melodic intervals, durational relationships, timbres, metre and other musical
elements by subdivisions, and each subdivision contains information on pitch, duration or
other elements. These are stored in parallel and the outputs combine at the time of retrieval
(Deutsch 1970). When one is determining whether two pitches are the same, the task
remains easy after a short interval. Although decay of the memory trace may have started, it
is not significant. When other tones are interpolated within a five-second delay, the task
becomes very difficult. The more tones that are interpolated, the more difficult the task
becomes. Pitch memory here is subject not only to time decay, but to interference. If
Page 12
12
difficulty arose from distraction of attention, then interpolated material of all kinds would
result in memory loss, but this did not happen. If it were held in a store with limited
capacity, then other non-musical materials would also be forgotten in the overload, but this
did not happen. A specialised pitch system is becoming overloaded with pitch information
only (Deutsch 1970).
This theory was tested in four groups. They were presented with a test pitch, a five-
second interval, and then a comparison pitch, which they then had to compare to the test
pitch. In group 1) six tones were interpolated in the interval before pitch comparison; in
group 2) six numbers were spoken in the interval before pitch comparison; in group 3) they
had the same as number 2, but were requested to both compare the pitches and recall the
numbers spoken in the interval; and in group 4) they were required to recall the numbers and
not compare the pitches. Table 2 shows the results.
Since group 3 had to memorise both numbers and pitches and had comparable pitch
memory impairment to group 2, and recalled numbers equally well as group 4, this shows
the different information was likely coded in separate systems. Group 1 had pitch memory
impairment from interpolated pitch, but no group had pitch memory impairment from
interpolated speech. Impairment could not be due to the lack of rehearsal as when speech
was interpolated error was far smaller. This suggests that pitch was processed in a
Table 2: Percentage error in
groups 1 to 4 (Deutsch 1970).
Group 1 shows substantial
impairment on pitch memory,
groups 2 and 3 show minimal
pitch memory impairment and
groups 3 and 4 show comparable
number recall error.
Page 13
13
specialised system, which was subject to interference from other pitch material, but not non-
musical material such as the spoken numbers (Deutsch 1970).
Deutsch (1973) also tested the error rate in pitch comparison when interpolated tones
were displaced by an octave. The tones of the same octave caused the greatest interference,
but the tones of the octave above were also highly disruptive. The least interference came
from the octave below. This could be explained by a suggestion that higher pitches attract
more attention, or that there is an independent process for interval analysis that is separate to
that of octave generalisation (Deutsch 1972) (and as we know this happens automatically
(Tervaniemi 2003)), meaning it becomes difficult to manage both tasks. The highest error
came from the test by Deutsch in 1974 when interpolated tones were played from all three
octaves, as the interval size was the largest (Deutsch 1999, 400).
Zatorre et al (1994) found that several regions are activated when working memory
load is high and that the frontal cortical region and posterior temporal regions are activated
in working memory for tones. Through measurements of Cerebral Blood Flow (CBF)
Zatorre and his team concluded that specialised neural systems that dealt with perceptual
analysis of melody were placed in the right superior temporal gyrus and pitch comparison is
computed in a specialised system in the right prefrontal cortex while retention of pitch
involves the right temporal and frontal cortices.
The auditory cortical areas are involved in high-information-load rehearsal, such as
in sight-reading, where one needs to retain pitch information while new information is
presented. In a study by Gaab et al (2003), subjects listened to six tones and were required
to judge whether the first and last tones were the same or different. Analysis of the
comparison caused activation patterns in the superior temporal gyrus and left inferior frontal
Page 14
14
gyrus, among other regions, measured through functional Magnetic Resonance Imaging
(fMRI) scans. These studies point to the existence of a working memory pitch subsystem.
Absolute Pitch A working memory pitch subsystem would involve the analysis of single pitches and related
pitches using the skills Relative Pitch (RP) and Absolute Pitch (AP). RP is the ability to
calculate pitches from their relation with a known pitch. The ability to recognise a pitch
without reference to another is known as perfect or absolute pitch (Bermudez and Zatorre
2009). Those who possess this trait report that it was not a learnt ability and that it was
acquired incidentally, which suggests they have a predilection affecting their perception of
information. It is likely that these ‘cognitive inclinations’ (Bermudez and Zatorre 2009) are
a part of the developmental, tuitional and genetic influences that lead to the development of
AP. Predispositions are very likely to be involved. Baharloo et al (2000) found sibling
recurrence of AP to be quite high. This suggests an innate neural substrate interacts with the
environment at a certain time in development. All possessors have been engaged in early
musical training (from under the age of six). This is when the central nervous system is most
adaptable and there seems to be more effect on the structures associated with the musical
activity at this age (Altenmueller and Schneider 2009). In their study of 113 siblings of AP
possessors, 50% also had the youthful training, and 50% of those also reported possessing
AP.
Miyazaki (1992) reported that AP impeded the development of RP, which is a more
important skill in musically meaningful situations. AP possessors were slower to recognise
or calculate intervals when the starting note was ‘out of tune’, suggesting that they were
perceived as two separate tones rather than a tonal relation. Several of the participants
counted up from one recognised note to the next in order to calculate the interval. However,
Page 15
15
many musicians have both AP and RP. AP was seen to be more prevalent in those deemed
to be extremely talented musicians and performers.
The brain activity of those possessing AP can be contrasted with those who do not,
as there is reduced electrical brain activity used in the working memory tasks of those
possessing AP. There also seems to be distinctly less response to pitch change. The right
frontal cortex, involved in monitoring pitch information in the working memory, is also less
active in those with AP. This suggests an element of efficiency or a lack of sensitivity, as
there is less analysis of pitch to be done (Peretz and Zatorre 2005). This is likely linked with
the categorical representation of pitch associated with AP as opposed to the continuous
trace, which allows for RP recognition. The right superior temporal gyrus and right inferior
frontal regions are activated when music is presented, and these are implicated in the
analysis of tonality. A link to tonal processing suggests that the initial perception of pitch
and auditory information is no different for a possessor of AP. The difference came when
single pitches were presented and activity in the left posterior dorsolateral frontal cortex was
noted in AP groups but not RP groups. This portion of the brain is associated with
conditional associative learning - learnt associations between a task and the appropriate
response (Sacchietti et al 2008; Zatorre et al 1998) - which may lead to a definition of AP as
the link of a stimulus (the pitch) to a response (the label or pitch name). This area was also
active in both groups when judging major or minor chords, which leads the authors to a
hypothesis that this area is implicated in verbal-tonal associations (Zatorre et al 1998).
The evidence of functional differences in the brain suggests that there may also be
some structural differences. A lateral asymmetry of the auditory cortical areas, which has
been evident in many studies, seems to be present in those possessing AP. There appears to
be either an enhancement of the left hand side or conversely a reduction on the right hand
side, which causes the difference (Keenan et al 2001). It is speculated that the early musical
Page 16
16
training is an influence of structural development. The size of the right-hand side was far
more consistent in the AP musicians tested than the left compared to the non musicians and
the RP musicians, suggesting that it is a ‘pruning’ of the right hand side which is the
influence on the development of AP (Keenan et al 2001, 1407).
Melody and Pitch Recognition In order to recognise melodies, a long-term trace of the pitch/time relations must be made
and maintained. Music is not based on a semantic system like language and instead the
representation must be one of form and structure. Melodies can be recognised at any pitch
and so are remembered as schema, not the absolute pitches, involving the intervals, contour
and scale degree context (Snyder 2009). We cannot use meaning to remember the melody –
meaning is conveyed through other means, such as our associations with memories and
emotions. As melodies are recognisable in different transpositions and with tempo and
instrumental change, the traces are likely to be abstract (Peretz and Zatorre 2005).
Processing of the semantics of music takes place in the middle temporal gyrus and the left
anterior temporal area (Koelsch and Siebel 2005).
However, Halpern discovered that we do often preserve some of the surface features
of music such as dynamics, tempo and some retention of the absolute pitch of tunes despite
insufficient verbal or visual cues for the pitch (Halpern 1989). Our memory’s duality in this
respect parallels our perception of music. We will never process every detail but we must be
aware of and consider the surface details of each unique performance of a piece. Pitch and
precise tempo can be retained in some form, but the memory of the entire piece is
remembered as more of a ‘gist’. The trace follows the structure of organisation (Peretz and
Zatorre 2005, 97) and uses the surface elements in order to relate these sections (Lamont
and Dibben 2001), which are used depending on the piece’s most memorable elements.
Lamont and Dibben (2001) also found that listeners are less likely to use more complex
Page 17
17
musical points, such as motivic development, in order to create mental representations of the
work. However, in order to appreciate and understand a unique performance, the surface
details like tempo and absolute pitch are required to be analysed. Both the structural and
surface details can be retained.
Further support for a musical memory subsystem comes from lesion studies. The
perception can remain intact whilst recognition of familiar melodies becomes impossible.
Sometimes this is even limited to music. Peretz (1996, 484) studied one lady who had a
bilateral auditory cortex lesion and was considered amusical (an inability to comprehend or
respond to music). She had a normal memory for lyrics when spoken, but was unable to
recognise the corresponding melodies. She had no other non-musical auditory deficits or
visual deficits, suggesting the damage was limited to this musical function. Zatorre and
Samson in 1992 found that after a right-sided temporal lobe lesion, retention of musical
memory quickly deteriorates. However, this right side is not as critical when the melody is
highly familiar. When recognition of a novel melody becomes difficult after damage to the
right temporal lobe, the left temporal lobe is used (Peretz and Zatorre 2005).
Bower in 1991 believed that mood might also have an effect on our memory for
music. Participants were placed in a positive and negative mood and then listened to a
melody judged as being happy or sad. In recognition there was a mood congruency effect
whereby the sad melodies were remembered better by those who were placed in a negative
mood, and vice versa (Houston and Haddock 2007). Houston and Haddock also found that
mood congruency effects are present in verbal and visual memory. This relates to the
activity on the right and left side of the amygdala that affects our moods, and this is held
deep inside the brain and connected to the auditory cortex (Scott et al 2000).
Page 18
18
Immediate Recall of Melodies Aural tests in instrumental exams require candidates to sing back a phrase or two after two
presentations. In the vast majority of my musical exams, I either just passed or failed on
these sections, so free recall is not a skill I have mastered. In music, free recall is an
interesting point of study when looking at the memory trace and the influences on and
products of the recall.
A study by Sloboda and Parker (2005) took eight young subjects, four who were
classified as musicians (trained performers) and four who were considered non-musicians
(they had not received any training but enjoyed listening to music). They listened to
excerpts from folk melodies. This study is interesting as they used real melodies as opposed
to pitch sequences designed specifically for the task, and is therefore more likely to show
how we react to and recall existing music. The excerpt was played six times and then the
subject was required to recall the sequence through singing six times. The authors found that
recalling more than one melody in a single session caused interference between the
materials, and so only the first melody recall was used in analysis. There was an intriguingly
high level of ‘intersong contamination’ (Sloboda and Parker 2005, 76), which was
unexpected.
The sequence was the first three phrases of the song ‘Sailor’, a Russian folk song
which was unfamiliar to the subjects. There were 21 notes, arranged in two-bar phrases, in
the form A1A2B. Section B shares much of the rhythmic patterns with A and so to an ear
attuned to Western tonality and structure, the melody would have been easily understood
and the structural groupings were clear. Transcriptions of the recalls were generated. Where
some subjects transposed their recall, it was ignored as unintentional and showed that
coding was in the context of pitch relationships, not actual pitch (Sloboda and Parker 2005,
77).
Page 19
19
Firstly melodic contour was analysed. A melodic contour template was drawn up for
the three phrases and the subjects’ recalls were compared. Each correct note within a pattern
received one mark. When presented as a percentage, they found wide-ranging scores from
0-88%. Interestingly an example from one subject shows a very low melodic accuracy
score, but the subject seems to have created a new melody on the harmonic structure of the
original – this is analogous to linguistic recall, where it is possible to recall a sentence that
means the exact same thing but may contain very few of the same words: this new melody
essentially means the same thing (Bower 1976, cited in Sloboda and Parker 2005, 74).
Another subject’s recall seems to be based on the third phrase, which is section B, the
section containing different material to the previous two phrases. The third phrase recalled is
sometimes based on phrase one but the phrases have been rearranged. If the individual
phrase recalls were marked on the phrase they most resemble in the original, the score
becomes 57% as opposed to the original 2% (Sloboda and Parker 2005, 80). The subject's
initial error of recalling the third phrase first is maintained throughout the recalls, showing
how persistent a recall schema can be. This is most evident in the recalls of the other
melodies tested (which were not used in analysis), as the recalls were so disturbed by the
first melody that the materials became mixed through of the stubbornness of the recall
schema.
Looking at how a subject may improve over trials, it was found that while the notes
matching the melodies rose, the number of notes produced rose, too, meaning accuracy
stayed around the 50% mark. There is a constant freedom on the recall, which shows that
our recall is not highly restricted by the original surface material that we encode, suggesting
that it is not an absolute trace that we create. As discussed previously, our memory is much
more abstract (Peretz and Zatorre 2005, 97), based on underlying structures.
Page 20
20
Is the A1A2B harmonic and melodic structure helpful in recalling the melodic
patterns? The similarity between the phrases must be measured. If the structure is preserved
in recall, then phrase three will be significantly less similar than phrase two to phrase one. In
the authors’ analysis of the original, it was indeed shown that phrase three was less similar
but in the results, it is seen that the subjects make phrase three more similar to phrase one
than in the original, but phrase two is still more similar to phrase one than phrase three. This
evidence points to the structure being reproduced, but that the recall is far less varied
structurally, which shows a tendency of memory towards simplification (Sloboda and Parker
2005 , 85).
A search between the two groups should show a difference between the musicians
and non-musicians. Musicians outperformed on all analyses except the consistency in
breathing places, yet interestingly the superiority was not always significant. In summary,
the recall of a melody was never note-perfect. Recalls were closely related to the original
melody in many ways but the best-retained element was the metrical structure. They all
retain a feeling of four in a bar, and this excellent performance on metre memory is
indicative of the primary use of metre as a structure for melodic comprehension and recall.
The phrase divisions (which is governed by harmony and metre) are also particularly well
adhered to, and the A1A2B structure is maintained even when the melody itself is incorrect.
However, the harmonic and melodic patterns are of varying degrees of accuracy throughout
the subject sample. It suggests that harmonic and melodic structures can be attained
independently of the rhythm, supporting Griffiths et al’s (1999, cited in Peretz and Zatorre,
2005; Fasanaro et al 1990) view that pitch and rhythm and pitch and metre may be separable
in brain function. And furthering this idea, it seems that harmonic structure can be separable
from melodic information (Peretz 1993). Several subjects’ recalls showed a very different
melody that fitted the harmonic structure, and it was common for rhythm, metre and
Page 21
21
harmony to all remain intact while there were large variations on the melody. This retention
of harmony was the only significant difference between musicians and non-musicians,
suggesting training in Western music increases one's understanding of and memory for
harmonic and tonal structures (Sloboda and Parker 2005, 89).
This evidence suggests that we form models of structure in our memory where
surface detail is not retained (Peretz and Zatorre 2005; Sloboda and Parker 1985). Recall
involves filling in structural sections adhering to the 'rules' in that section. Different amounts
of training allow us to be sensible to the levels of structure. Due to this, the musicians can
code those harmonic relationships that are too complex for the non-musicians to
comprehend. However, there are certain elements that can be processed by both groups,
such as the basic melody and rhythm and the ascents and descents that are responsible for
the melodic contour. The main points of perception and memory for music that have been
discussed previously are here shown in action.
Chapter Summary In this chapter we have seen that the brain areas most implicated in the processing of
pitch are the right temporal cortex, in the anterolateral section, and particularly the superior
temporal gyrus as this contains the auditory cortex (Peretz and Zatorre 2005). Also
implicated are the left medial area and the splenium, an area deep inside the brain
‘underneath’ the auditory cortex. The junction between temporal, occipital and parietal
regions is also important, as damage here is correlated with a deficit in pitch processing
(Fasanaro et al 1990). In the right secondary auditory cortex, the anterior portion deals with
pitch chroma whilst the posterior portion deals with pitch height (Peretz and Zatorre 2005).
Melody is recalled as a series of pitch relationships, and the underlying structures such as
rhythm metre and harmony are much better recalled than the actual melody, even though the
melody is the most apparent factor in the music. Recall schemas are based on structures, not
Page 22
22
the surface features and musical training increases your understanding and therefore
memory for these structures (Sloboda and Parker 2005).
Deutsch’s 1970 study shows that non-musical information doesn't interfere with
pitch, while musical analysis does, supporting the specialised system for the memory of
pitch and melody. Zatorre et al (1994) show us that working memory load activates auditory
areas in the superior temporal gyrus, meaning the auditory cortices are implicated in high-
load musical working memory, supporting the separate subsystem hypothesis also presented
by Gaab et al (2003). If there is indeed a musical memory subsystem, it is active in the
auditory cortex and the right temporal lobe, with longer-term musical memory leaning
toward action in the left temporal lobe. This side deals with language (Scott et al 2000) and
semantics (Koelsch and Siebel (2005) implicated the left anterior temporal lobe in their
study), and therefore musical semantics relating to our associations.
For AP possessors, there is reduced brain activity for working memory compared to
those not possessing AP. This is related to the categorical representation of pitch in AP as
opposed to a continuous trace for RP, which requires more maintenance of information.
When a single pitch is presented, AP possessors show action in the left posterior dorso-
lateral frontal cortex, which works with stimulus and response processes, in this case pitch
and pitch label. There is brain asymmetry in possessors suggesting that the smaller right-
sided auditory cortex may influence AP development.
Page 23
23
Chapter 2 – Melody and Harmony
Melodic Contour Another method in which our brain processes melody is through the perception and memory
of melodic contour. The melodic contour is such a strong element in memory that a
reproduction of a non-Western melody, by a Westerner, would be produced with Western
elements, and yet the contour of the melody would be intact (Dowling 1978). Dowling
(1978) also points out that if the rhythm is incorrect or the intervals are displaced by an
octave (octave displacement: Deutsch (1972)) the tune may still be recognised. Deutsch
(1972) discovered when playing 'Yankee Doodle' with the notes randomly chosen from any
of three octaves, recognition was inhibited but still possible. She concluded that interval size
and preservation of contour are the most distinctive features for memory in our memory
trace. Long (1977) conjectured that a melodic contour with more directional change was less
effective in aiding the memory of pitch because more attention is required to the interval
size, where as a smoother, perhaps ‘V-shaped’, contour would create a shape memory trace
rather than intervallic memory trace.
The right superior temporal gyrus is strongly implicated in contour perception and
memory. Interval relations that make up the contour are processed in both the right and left
temporal gyri as discussed by Vignolo (2003), who found that the difference in accuracy in
the interval testing between those with right hemispheric lesions and those with left was
insignificant. Patel and Balaban in 1998 showed the cooperation between left and right
hemispheres in the processing of simple melody sequences (Peretz and Zatorre 2005, 92).
This asymmetry in the functions is already apparent in the tests on infants conducted by
Balaban et al (1998). The study found that reaction to a change that alters the contour of a
melody was stronger when presented in the left ear, while an interval change that preserved
the melodic contour provoked a stronger reaction when presented to the right ear (Balaban
Page 24
24
et al 1998, 48). This is consistent with asymmetry in contour and intervallic change
discovered in adults but Anderson in 1994 (Balaban 1998, 48). This evidence suggests that
perception of contour may be stronger on the right-hand side of the brain (as the information
came from the left ear) and that memory for contour is stronger on the left-hand side. Once
again, encoding of both contour and intervals does not require the attention of the listener
(Tervaniemi 2003).
A study by Epinosa and Gernstein in 1988 used three-note series in all of their
permutations (ABC; ACB; BAC etc.) and found local networks of neurons processing these
series, and they discovered up to 24 neurons spiking simultaneously. The effect of one
neuron’s activity on another was specific to each sequence: neuron-spike patterns of one
tone changed depending on the preceding tone (ABC compared to BAC). Connections
between the tones depended on the pattern of the tones, which points towards contour being
coded at a neuronal network level (Weinberger 1999).
Scale Structure and Tonality The scale structure involved in a tonal melody is a useful tool for listeners in the perception
and memory in pitch sequences and it is often learnt implicitly and used automatically to
influence incoming pitch, melody and interval information (Tillmann and Bharucha 2000).
Tonality can provide a ‘cognitive landmark’ (Snyder, 2009) and when compared with non-
musicians, although musicians' recall of music is superior, Cook found in 1987 that it is less
superior when they are presented with an atonal sequence (Lehmann et al 2007). Brain
damage can inhibit tonal knowledge while the interval and contour perception are spared
(Peretz 1993). In a case study by Peretz (1993) the subject who had a lesion in the left
temporal lobe and right frontal-opercular region (which includes the right temporal gyri)
could no longer judge the difference between tonality and atonality, and had a reduction in
pitch memory, but could still perceive the pitches and melody contour. There may be
Page 25
25
specialised neurons for processing of scale structure because it can be separable from other
functions. Janata, Birk, Van Horn, Leman, Tillmann and Bharucha (2002) suggested that the
medial prefrontal cortex was associated with knowledge of the key and tonality of a piece,
maintaining a tonality map. The authors discovered that the connections to the medial
prefrontal cortex were mainly coming from areas associated with auditory processing, and
that this auditory task-related activity was seen mainly in the right hemisphere of the
anterior superior temporal gyrus (Janata et al 2002).
Long (1977) conducted an experiment whereby he tested the effects of melody
length, contour, tonal structure and musical ability on memory for pitches within melodies,
which is different to other studies testing a sequence recall ability. Unsurprisingly, two of
the three groups that were comprised of mostly music students performed far better than the
one group consisting of mostly non-music students, supporting the view that musical
training would increase memory for pitch. Memory may be dependent on the learned
musical systems. He found that the length was not significant but that the tonality of
melodies, as opposed to atonality of others, increased the ease at which the pitch was
remembered. The learned system of tonality was a great aid to this memory (Long 1977).
A knowledge of tonal and harmonic principles lead to expectancies in harmonies.
Tramo, Bharucha and Musiek studied brain damaged subjects in 1990 and found that they
retained the ability to generate chord expectancies after a bilateral lesion in auditory cortex
but were deficient in pitch processing and had difficulty in judging intonation (Peretz and
Zatorre 2005, 93). Deviation from the chord expectancies causes robust potentials in the
brain, in the inferior frontal areas where the neurons generate a response. Koelsch, Gunter,
van Cramon, Zys-set, Lohmann and Friederici in 2002 showed that this happens bilaterally
(Peretz and Zatorre 2005, 93). The right hand side is typically implicated in yes/no decisions
and perhaps in inhibition responses to risk, which may be linked to the surprise of a
Page 26
26
harmonic deviation. The left hand-side is used in language functions and grammar (Scott et
al 2000), which corresponds to the ‘grammar’ of music, and the change in direction for the
music. Our grammar of music however, is limited to the musical culture in which we are
saturated. A Western-cultured person would reproduce a melody of entirely non-Western
elements inside the Western tonality schemes (Dowling 1978). Bartlett and Dowling in 1980
(Shuter-Dyson 1999) saw a recognition of key change, when the key was distant, in children
as young as 5 whilst children aged 9-10 showed a 90% accuracy in identifying a tonality
change.
Consonance and dissonance naturally play a part in our perception and recognition
of pitch relations. Intervals that are consonant have a simple frequency ratio (for example,
the fifth is 3:2) and dissonant relations have more complex frequency ratios (16:15 for the
minor second). The superior temporal gyri are again implicated here as dissonance appears
to be computed bilaterally in this area, possibly by specialised mechanisms. Fishman,
Volkov, Noh, Garell and Bakken in 2001 found dissonance caused activity in the temporal
gyri, but not consonance (Peretz and Zatorre 2005, 94), and a bilateral lesion of the auditory
cortex (inside this gyrus) has the effect of a loss in the subject's sense of dissonance (Peretz
et al 2001, in Peretz and Zatorre 2005, 94).
Some questions brought up here by Peretz and Zatorre (2005) are that of the point in
perception where computation of dissonance is critical, and whether our sensitivity is to
dissonant noises in general or to dissonant pitch relations. Is our definition of dissonance
down to our natural auditory-musical functions, or is it through learned associations? Lynch
and Eilers in 1991 tested western children's judgment of dissonance in western and Javanese
music, and found that while the western music was more accurate, they performed better
than the chance accuracy (predicted accuracy if purely guesswork) (Carterette and Kendall
Page 27
27
1999, 731), so there is some argument for a innate sense of dissonance, but with greater
sensitivity in the familiar and learned contexts.
Musical Notation and Structure Tonal structure's role in memory has already been discussed, which has its largest effect in
aural presentation. Does the visual representation have an effect on our memory similar to
that of the musical structure? Standard notation represents musical pitch and rhythm in a
visual manner. Ordered and 'good' musical structure is remembered far more easily than
'bad' or random structure (Halpern and Bower 1982).
Halpern and Bower used an interesting experiment on chess experts and chess
amateurs by Adriaan de Groot in 1965 showing results in how they perceive and remember
patterns. He found that to remember patterns and moves both during and after play, chess
Grand Masters use chunking, a memory method involving grouping information together to
create one 'chunk', rather than several smaller ones. The amateurs did not do this. It suggests
that other experts, such as musicians highly skilled in reading of notation, would also use
chunking in perception (in tasks like sight-reading) and memorisation while amateurs would
not. However, de Groot found that the experts’ memory is superior in meaningful patterns,
but no better than the amateurs’ when the patterns were random. Halpern and Bower (1982)
believed it would be the same for musicians when musical patterns and structures were
ordered and meaningful through a visual presentation.
In Halpern and Bower’s experiment they used sets of ten crotchets with no
accidentals or repeated notes and they were placed into pairs of 'good' structures and 'bad'
structures with very similar contours and intervals, meaning they are visually similar. These
melodies were judged to be 'good' or 'bad', both visually and aurally, by separate groups of
musical graduates and expert performers. The 'good' were judged significantly better than
the 'bad', which confirms their classification on both visual and aural presentation. They
Page 28
28
were also judged equally in visual complexity by non-musicians. 'Random' melodies were
composed in addition to the 'good' and 'bad' with larger intervals and more contour changes.
In the pilot study they discovered some possible encoding methods for the notation. Halpern
and Bower had a group of twelve musicians and twelve non-musicians. A slide of a 'good' or
a 'bad' melody was displayed for five seconds, and then recalled after a delay of fifteen
seconds. They found that musicians were overall better at recall than non-musicians and that
the 'good' melodies were overall remembered far more accurately than the' bad'. Whilst the
musicians remembered the 'good' melodies more than the 'bad', the non-musicians
performed equally poorly on both the categories, which supports the judgement that the
melodies were equally complex visually. The musicians’ memory for the 'bad' melodies was
better than the non-musicians’ performance on the 'bad'
In the second experiment, Halpern and Bower (1982) further investigated the effect
of the musical structure on the notation. Since the musicians recalled 'bad' melodies better
than the non-musicians, what kind would they recall equally? Random note placement in the
random melodies causes a far greater interval size average, which leads to an increase in
complexity of the visual presentation. This should affect both groups as while the non-
musicians only use visual regularity, musicians use both visual and musical regularity. The
random note placement inhibits the musical regularity as well as the visual. The musicians
showed that they performed well for 'good' melodies as they had both good visual and
musical regularity, moderately for the 'bad' melodies, which also has good visual regularity
but poor musical regularity, and badly on the random melodies that lacked all regularity.
The non-musicians performed moderately, and equally so, on the 'good' and 'bad' melodies
and badly on the random melodies. The musical differences had no effect on the non-
musicians, only the visual differences, and so they were worst performers on the random
melodies as there was little visual structure. Musicians were only mildly superior to the non-
Page 29
29
musicians on the random melodies, as there was no musical or visual regularity to follow,
which means melody had to be remembered in terms of pitch names. This was unfamiliar to
the non-musicians, probably explaining their poorer performance.
Absolute accuracy on recall was initially scored, but this did not allow for near
misses (like placing a note one line too far down on the stave) or for meaningful musical
mistakes (such as octave displacement), so the memory for the overall visual representation
of notation was difficult to judge. Instead it was marked on similarity – a melody placed
consistently one note too low on the stave in recognition would have been marked zero on
absolute, but actually it is very close to perfect so the memory was very good. This was
marked out of ten. The non-musicians scored 2-3/10 and the musicians’ score was far more
wide-ranging, but none were approaching perfect. The musicians’ scores were as predicted,
but their advantage over non-musicians decreased from 'good' to 'bad' to random. Their
similarity points were far higher than the non-musicians'. Musicians made some musical
errors. Often when memory was not intact, musically sensible substitutes were inserted. In
other examples, notes were encoded as an F major triad, for example, but recalled in the
wrong order. This is an example of the chunking of patterns within melodies. There were
fewer of these mistakes in the less structured 'bad' melodies as compared to the 'good', as
there was less musically structural information to encode.
Even when the recall was immediate, the musicians were better at recalling the
'good' melodies, which shows the immediate use of the learned structures as a framework
for memory. The effect of musical structure on working memory for pitch is shown in sight-
reading experiments by Sloboda (cited in Gabrielsson 1999, 512) where musicians
processed the music as a series of groups, both structural and musical. After a very brief
exposure to melody, the musicians were superior in recall of the simple patterns, and
reported that they seemed to be obvious in their patterns and groups.
Page 30
30
Two characteristics are likely to be helpful in memory for pitch sequences. Firstly,
the ease with which you can group the melody into sub-patterns and secondly the number of
patterns that are likely to be formed. Following the theory that musicians will use chunking
to remember patterns, the more easily sectioned melodies should be easier to remember, and
even easier if there are fewer chunks of information to maintain. It is expected that chunk
size is related to expertise (Lehmann et al 1007, 112). In a well-structured melody,
groupings would be easier and fewer, and thus more easily remembered than poorly
structured 'bad' melodies. Visual cues are unlikely to indicate the groupings, so non-
musicians, who are relying on the visual structures, are unlikely to use this method of
memory for recall.
Using the same 'good' and 'bad' melodies from the previous tests, Halpern and Bower
(1982) presented these melodies on a sheet of paper in a random order to a group of subjects
(seven musicians and seven non-musicians) and instructed them to use vertical lines to
group the melodies as they wished. Consistency of absence or presence of a particular
division for both groups was marked along with the average number of groups for each
melody and the number of divisions each subject gave each melody (Table 3). Interestingly,
the difference between the two groups on consistency in the 'good' melodies was
insignificant, and in fact it was the non-musicians who were more consistent in their choices
for groupings. However, the non-musicians' choices seemed not to be musical decisions and
the consistency came from decisions like a visual division where the direction of the note-
stem changed. Another interesting finding was that the 'good' melodies were no more
consistently grouped than the 'bad' melodies, for either group, which shows that the musical
structure which makes it 'good' is not a visually obvious element, and therefore is not seen in
the 'good' melodies by the non-musicians, and that a musical division may be subjective and
differently judged by different musicians.
Page 31
31
To see whether the musicians were using the chunking strategy, recall scores from
immediate recall of melodies were compared with the grouping consistency marks for each
melody. As expected, the non-musicians showed no correlation between their grouping
consistency and accuracy in immediate recall whilst there was a significant correlation for
the musicians, pointing to a suggestion that chunks that are clear in the melody are used to
ease the memory process when a quick glance is the only presentation.
When there are fewer chunks, there is less information to maintain. Musicians
marked a significantly lower number of musical groups or chunks in the 'good' melodies
than the 'bad' melodies, but for the non-musicians it was the other way around. This was
explained after exploring the number of stem direction changes for each group of melodies
and the 'bad' melodies had fewer changes than the 'good' melodies. This shows the
importance of the musical structures for the musicians over the easier visual structures like
the grouping of the stem changes. The non-musicians were relying heavily on the visual
grouping strategies over musical groupings, as they showed a very high correlation between
the individual grouping numbers for each melody and the mean average throughout the
whole group. This suggests they used the same stimulus to mark segmentation in the
melody. The average number of groupings for each melody was then compared with the
recall marks and there was no correlation for the non-musicians. Clearly the stem direction
segmentation was not a method that was implemented by or that helped the non-musicians'
Table 3: Mean
consistency score
and number of
groupings for good
and bad melodies.
Halpern and Bower
(1982).
Page 32
32
recall performance. The musicians showed a significant negative correlation between
number of groupings and performance in both delayed and immediate recall (in which it was
even more significant), meaning they made fewer groups and therefore performed better.
The musicians grouped musically, not visually, and could take advantage of these groupings
in memory.
The inconsistency in the musicians’ groupings may have come from the lack of
musical features other than pitch. Segmentation, patterns and phrases are usually indicated
by larger scale contours and rhythms, neither of which are present in these melodies. The
fact that they ignored the visual cues of the stem directions (whilst the non-musicians took
advantage of this and consequently were more consistent) can explain the lack of
consistency through the group.
These experiments showed the negative correlation between the number of chunks
and recall accuracy in musicians' recall and therefore that chunking was a tool used by the
musicians, knowingly or otherwise, in order to recall melody. These relations are stronger
when the recall is immediate which, along with the findings that well-structured melodies
aid recall, show the very important and fast influence on memory trace that musical
structure can have, suggesting very early use of 'music-specific encoding' (Halpern and
Bower 1982, 46). It also shows that the non-musicians do not use these strategies, which
suggests either an innate brain function or an element of training is involved.
The 'grammar' of music (Halpern and Bower 1982, 47) has been learnt by the non-
musicians (to some extent) through listening to music and this is clear from the inter-
participant reliability of the non-musicians judging whether the melodies were 'good' or
'bad'. They were relatively consistent in deciding which were 'good' and 'bad', even though
the 'bad' were not 'blatantly discordant' (Halpern and Bower 1982, 47). Musicians have also
Page 33
33
gained this understanding and built upon it through their training, strengthening their
expectations and knowledge of structures and activities. This includes the names of relations
and structures, which can aid their memory for these features and elements of music. The
knowledge of the grammar involved in a 'good' melody and their associative knowledge of
how it is presented visually means recognition of groupings, patterns and structures occurs
immediately. Patterns which are well known (such as triads, or rising scales) can aid the
expectation of what is to come, and this would explain the existence of musically sound
mistakes in recall of melodies - the errors are not haphazard and come from interference
from similar musical patterns stored in the long term memory (Sloboda 1978). A melody is
presented as a ‘hierarchy of subsequences’ (Deutsch 1980, 2) and structure is utilised in
recall. Deutsch also found that the unstructured sequences caused a much heavier memory
load. The expectancies are quickly confirmed and the tonal hierarchy also helps us to
organise these chunks to aid the memory (Deutsch 1980). The chunks and musical structure
are often interlinked so when one strategy fails in retrieval, another can be used (Lehmann et
al 2007, 120).
Some studies suggest that the music is not stored in a simply visual or aural form but
in a more abstract manner recording contour (Dowling 1978) and tonality (as this is a well-
defined mental structure on which to place the melody) (Dowling 1978) along with other
musical elements. Sloboda’s 1978 example of 'chunks' is the sequence [C-D-E-F-G-D-B-C],
coded as:
a) C scale rising to dominant;
b) falling dominant chord;
c) return to tonic.
A novice musician unaware of tonal structure might encode:
a) C up to G;
b) G down to D;
Page 34
34
c) D down to B;
d) B up to C.
This comprises of more chunks and they have no connecting link between then. A
non-musician might then code the contour and approximate notation, as he has no access to
abstract frameworks. One builds an expectation on what you can see, and with confidence in
your training you may not need to consciously look at all the information (Lehmann et al
2007, 116; Tervaniemi 2003). Halpern and Bower (1982) suppose that the musicians use
these abstract methods and musical names rather than attempting to 'hear' the music in their
heads as this might cause interference (Sloboda and Parker 2005) as one would be
essentially sight-reading the presented melody, not concentrating on the memory trace.
Chapter Summary This chapter has shown the importance of preservation of contour for memory and for
recognition. The right superior temporal gyrus is activated in both perception and memory
for melodies and contour. The right brain seems to accommodate contour change while the
left preserves and recognises contour. The connections (interval change and direction)
between the pitches cause neurons to behave differently, which brings contour to a neuronal
level of perception, meaning it is a very detailed part of the function.
Tonal structure seems to be learnt automatically and acts as an aid in memory for
music. From brain damage studies we saw that a bilateral lesion in the temporal lobe led to a
loss of pitch and tonality recognition, but the tonal processing function remained, suggesting
that musical perception and memory belong to separable systems. A bilateral lesion also led
to a deficit in pitch processing but the ability to generate chord expectancies was retained,
showing that harmonic systems may be separable from the melodic systems. The right
anterior superior temporal gyrus is connected to knowledge of tonality (as well as working
memory) and so must be instrumental in sight-reading. The inferior frontal region is
activated in harmonic deviance bilaterally – on the right side because of surprise (linked to
Page 35
35
risks) and the left for the analysis of change in 'grammar'. Bilaterally, the superior temporal
gyrus is activated in dissonance – the right side for musical sound and the left for grammar
or semantics.
The tonal structure plays a big part in a musician's memory for music, and while the
visual structure can help (as 'bad' melodies were recalled better than 'random' melodies in
Halpern and Bower's 1982 experiment) it is secondary to music. The grouping of the
melodies showed that chunking, expectations and tonal knowledge are all methods
employed to remember a melodic sequence.
Page 36
36
Chapter 3 – Music in Action
Sight-Reading and Working Memory Sight-reading is an important musical activity, which involves very brief and complicated
processes in the brain. The sight-reading of music is functionally and neurally different to
our reading of words. For example, in some brain damage studies Cappelletti, Waley-
Cohen, Butterworth and Kopelman in 2000 found that symbol reading can remain intact
while musical notation reading is inhibited (Peretz and Zatorre 2005, 101). fMRI scans
revealed the involvement of the right occipital temporal region in deciphering musical
notation (Schon et al 2000, in Peretz and Zatorre 2005, 101). Sight-reading requires vast
amounts of information to be processed in a very short time for temporary use. One must
interpret both pitch and rhythm, sometimes on more than one stave, comprehend and
implement a key signature and metre and recognise the familiar patterns. Often these are
perceived peripherally, meaning errors that occur are 'musically meaningful' as they fit into
the perceived structure (Sloboda 1977, in Sloboda 2005, 40). The optimum time span for the
working memory to process music is 5-8 seconds and the surface detail is therefore partly
set-aside in order for the larger-scale structure to be comprehended (Snyder 2009). A
performance plan for motor translation must be created and the brain must anticipate correct
continuance of the piece in order to recognise mistakes automatically, using expectations
and imagery.
As has been discussed, some of these actions are distinguishable neurophysically.
After brain damage, pitch and rhythm perception can become separated (Fasanaro 1990).
There can be damage at different levels of perception and decoding, even as early as the
visual perception where pitch and rhythm are presented in different manners. It may happen
later on, when the brain attempts to create an imagined representation. Pitch is encoded
differently for different people. Some may give note names while others would use a motor
Page 37
37
function (like pressing the key on a piano) so it could be one of these functions that is
damaged (Peretz and Zatorre 2005, 102). However, since many can sight-read in different
media, through singing or several instruments, representation of pitch is likely to be quite
abstract and unrelated to the medium of performance. Koelsch and Siebel (2005) implicated
the frontal, parietal and premotor cortical areas in working memory, along with the
cerebellum.
According to McPherson in 1993, sight-reading is one the five skills all musicians
should possess (Kopiez et al 2006). Activities and skills relevant to sight-reading are aural
imagery, sight reading experience, your style of thinking and possession of an external locus
of control (Kornicke 1995, in Kopiez et al 2006). Sloboda (1974) noted that one’s ability to
read ahead is a condition and a sufficient short-term memory buffer must be present in order
to read ahead whilst playing the previous music. He also suggested that being left-handed
may be an advantage as left-handed people generally have less of a handicap on their right-
hand side than right-handed people do on their left side. Kilshaw and Annett in 1983 also
found that musical imagery appears to be highly related to sight-reading, as efficiency in
this department may help to hear the music and understand how it should sound before it is
executed, all within the small space of time (Kopiez et al 2006).
Imagery and Mental Rehearsal Musical working memory is essential for musical imagery, described as imagining music or
its attributes without hearing it (Peretz and Zatorre 2005, 97). Perceptual defects and
imagery defects are often found in common when a lesion in the auditory cortex is present,
which supports the view that imagery uses the perceptual mechanisms. In some tests with
brain imaging, it was discovered that the secondary auditory cortex is activated in both
imagery and rehearsal of melodies (Halpern and Zatorre 1999). They confirmed their
predictions that the right associative auditory cortex (which provides a realistic experience
Page 38
38
for the brain) accompanies musical imagery, and that the imagery cues retrieval from the
‘musical semantic memory’ (Halpern and Zatorre 1999, 702) and this increases CBF in the
right inferior frontal region and bilaterally the middle frontal region. When there is no need
for ‘semantic retrieval’ then the left frontal area is recruited. However, it is the associative
auditory cortex, not the primary, which is active in the imagery. They used melodies both
with and without lyrics. Those with lyrics activated the temporal cortex bilaterally whereas
those without activated the right temporal cortex. The right auditory cortex activation
supports the hypothesis that the right hemisphere mechanisms process the tonal information
(Halpern and Zatorre 1999, 702).
Highben and Palmer in 2004 found when they removed the auditory feedback of
rehearsal and requested the participants to imagine the feedback, that performance was
poorer than after rehearsal with the auditory feedback. This shows that the auditory
feedback is important for a successful performance. The pianists with excellent aural
imagery were least affected by the removal, supporting the view that aural imagery involves
the same brain sequences as actual aural information (Palmer 2005).
Janata (2001) found that the imagination of the continuance of a melody (where they
omitted an unexpected note) showed similar electrical functions in the brain to when
perception of a real tone occurs, which suggests that the auditory mechanisms which are
responsible for perception are also involved in imagery (Halpern and Zatorre 1999).
The neural networks involved in musical ability can be studied by teaching non-
musicians to play music (Peretz and Zatorre 2005). Musically untrained subjects were
allocated two hours of piano for five days, which, besides resulting in vast improvement in
performance, also resulted in increased activity in the motor areas associated with hand
muscles. Primary motor cortex neurons also showed increased activity, and repeated
Page 39
39
information input to the motor cortex resulted in long-term potentiation of the motor
neurons (Pascual-Leone 2001). Interestingly mental practice (‘the imagined rehearsal of a
motor act with the specific intent of learning or improving it’ (Pascual-Leone 2001, 321))
has similar effects:
‘Studies of regional Cerebral Blood Flow (rCBF) suggest that the prefrontal and
supplementary motor areas, basal ganglia, and cerebellum are part of the network
involved in the mental simulation of motor acts. Therefore, mental simulation of
movements activates some of the same central neural structures required for the
performance of the actual movements. In so doing, mental practice alone seems
to be sufficient to promote the modulation of neural circuits involved in the early
stages of motor skill learning. This modulation not only results in marked
improvement in performance, but also seems to place the subjects at an
advantage for further skill learning with minimal physical practice. The
combination of mental and physical practice leads to greater performance
improvement than does physical practice alone, a phenomenon for which our
findings provide a physiological explanation" (Pascual-Leone 2001, 321).
This shows the quick effects of training on brain plasticity. There are several
differences therefore in the way that musicians and non-musicians use the musical
mechanisms in the brain and listen to music. It seems that musicians recruit more neural
tissue for the activities and are more analytical in their listening. However, this is not
exclusive to musicians as non-musicians have shown that they too consider interval size
both consciously and automatically (Tervaniemi 2003). Even so, Peretz and Babai in 1992
found that musicians are more flexible in their listening, using their learned tonal and
structural knowledge to decide which method would be most appropriate for coding,
analysis and memory-trace generation (Peretz and Zatorre 2005, 105).
Page 40
40
Brain Function and Structure It is clear there is right-sided asymmetry for pitch-based processing but music uses a vast
network of regions on both right and left sides of the brain. Music and the activities
involved are both complex and numerous and it is not clear which part of the brain are
music-specific. Many of them share neurons and systems with other functions. For example,
Koelsch (2002) found that harmonic deviation activates Broca's area inside the frontal lobe,
which is a region associated with language, suggesting an element of syntactical procedure.
However, the area is vast, and could easily accommodate several networks, meaning the
musical function and the language function could be entirely unrelated in this area (Peretz
and Zatorre, 2005). The brain's structural adaptions to each function happen at a gross
structural level and appear during early-aged training (Altenmueller and Schneider, 2009).
Chapter Summary The studies in this section have shown that the auditory cortex is implicated in the imagery
of music and in working memory with the frontal, parietal and premotor cortical areas and
the cerebellum. Imagery needs the perceptual mechanisms in order to occur, like mental
rehearsal, which involves the primary motor cortex and the right associative auditory cortex
is involved in musical imagery. For musical semantic retrieval the right inferior frontal
region and bilateral mid-frontal region are used. When lyrics are used in perception, the left
and right auditory cortices are both involved as the left deals with language while right deals
with sound (in this case tonal information), and when without lyrics, only the right side is
used. In comparison with actual aural feedback, imagery causes the same activation in the
brain, and similarly when imagining a continuing melody, the same activation appears as
when actual perception is occurring.
Many separable processes are involved in sight-reading, but for them to work
simultaneously, one must have an ability to read ahead, an adequate short-term memory
Page 41
41
buffer and knowledge of the musical tonal and structural and frameworks to aid imagery and
generate accurate expectations.
Page 42
42
Conclusion
During this study, many activities and processes have been addressed in order to understand
the human brain’s means of dealing with musical information. It has led me to some
conclusions about my own patterns of perception, analysis and memory.
I may have been more able than my peers to acquire musical skills because my entire
family is musical. It is not necessarily genetic, but environmental. Studies have shown that
much musical information can be processed automatically, and therefore knowledge can be
unknowingly assimilated. Being surrounded by music in my very early years and onwards
must have had an effect on the development of musical structures and functions in my brain.
My ability to sight-read is likely due to a combination of things, but I believe the
main one may be an efficient short-term working memory buffer. This allows me to process,
perceive and reproduce the many facets of music in a very short space of time, and its
efficiency allows me to do this accurately. I have found in other areas of life, I am very able
to ‘juggle’ tasks: I can participate in several conversations, talk and read simultaneously and
even in more academic situations like mathematics, several activities were easily possible.
In comparison to my cognitive style, my sister cannot concentrate on more than one task,
but can concentrate for much longer. During aural recall, she can keep her mind on the
presented material without thinking about other things, and there are fewer facets to
perceive in this fashion of presentation. She is able to hold the information and recall it with
better accuracy. However, she cannot comprehend all the information involved in sight-
reading fast enough to reproduce the music accurately. A secondary reason could be an
aptitude in musical imagery. As I am very able to “hear” the music as I read it, it will aid my
understanding of the piece, and help to build those expectancies built on tonal frameworks.
When required to sing back a presented melody, I am aware that whilst the contour
of the recalled melody is usually correct (as was the case in Sloboda and Parker’s 2005
experiment), the tonal structure often is not. As I don’t have absolute pitch, my analysis of
Page 43
43
pitch relations and intervals to generate a trace, but these are based inside a tonal structure,
and it appears that the frameworks that I have understood and maintained are not being used
to aid my memory, unlike perception during sight-reading.
I think my abilities come down to my preference for the visual presentation of
music, as opposed to aural presentation. Essentially, aural recall and sight-reading amount to
the same processes – perception and reproduction. However, aural recall requires fast
perception, memory and recall of aural information, whereas sight-reading requires fast
perception of a larger amount of information through written music followed by
reproduction. Learning a piece of music by ear is impossible for me, and I find it incredibly
difficult to perform the most basic analysis without reading the score. In other tasks, such as
learning languages, I could pronounce and remember the word if I saw it. In an informal test
on myself, I could remember musical phrases such as in Halpern and Bower’s experiment
(1982) through seeing the musical patterns and chunks – a combination of the musical and
the visual methods. I think perhaps my difference in skill is not due to musical memory or
analysis, but a failing in aural perception, and a balancing superiority in visual perception.
Many brain areas, methods, processes and contextual effects (such as mood at the
time of testing) have an effect on performance and skill in music, so it is impossible to
pinpoint the reason for the difficulties I and other musicians have in certain activities, but
through knowledge of my own processes and through this study of perception and memory,
I think I have uncovered a path towards the answers.
Page 44
44
References Altenmueller, E. and S. Schneider (2009) ‘Planning and performance’ in Oxford Handbook
of Music Psychology. S. Hallam, I. Cross and M. Thaut (eds.) pp 332-343, Oxford:
Oxford University Press.
Baharloo, S., Susan K. Service, Neil Risch, Jane Gitschier and Nelson B. Freimer (2000)
Familial Aggregation of Absolute Pitch. The American Journal of Human Genetics, 67,
755–758.
Balaban, M. T., Linda M. Anderson and Amy B. Wisniewski (1998) ‘Lateral Asymmetries
in Infant Melody Perception’. Developmental Psychology, 34, 1:39-48.
Bermudez, P. and Robert J. Zatorre (2009) ‘The absolute pitch mind continues to reveal
itself’. Journal of Biology, 8:8, 75.
Burns, E.M. (1999) 'Intervals, Scales, and Tuning' in The Psychology of Music D. Deutsch
(ed.) pp. 251-264, London: Academic Press.
Carterette, Edward C. and Roger A. Kendall (1999) ‘Comparative Music Perception and
Cognition’ in Psychology of Music D. Deutsch (ed.). 725-791, Academic Press: London.
Deutsch, D. (1970) ‘Tones and Numbers: Specificity of Interference in Immediate
Memory.’ Science 168, 1604-1605.
Deutsch, D. (1972) ‘Octave generalization and tune recognition.’ Perception &
Psychophysics, 11, 411-412.
Deutsch, D. (1973) ‘Octave generalization of specific interference effects in memory for
tonal pitch.’ Perception and Psychophysics, 13:2, 271-275.
Deutsch, D. (1980) ‘The Processing of Structured and unstructured tonal sequences.’
Perception and Psychophysics, 28, 381-389.
Deutsch, D. (1999) ‘The Processing of Pitch Combinations’ in The Psychology of Music, D.
Deutsch (ed.) pp 349-412, Academic Press: London
Dowling, W. Jay (1978) ‘Scale and Contour: Two Components of a Theory of Memory for
Melodies.’ Psychological Review, 85:4, 341-354.
Fasanaro, A. M., D. L. A. Spitaleri, R. Valiani and D. Grossi (1990). ‘Dissociation in
Musical Reading: A Musician Affected by Alexia without Agraphia.’ Music Perception:
An Interdisciplinary Journal, 7:3, 259-272.
Gaab, N., Gaser, C., Zaehle, T., Jancke, L., Schlaug, G. (2003) ‘Functional anatomy of pitch
memory - an fMRI study with sparse temporal sampling.’ NeuroImage, 19, 1417–1426.
Gabrielsson, A (1999) ‘The Performance of Music’, in The Psychology of Music, D.
Deutsch (ed.), pp 501-602, Academic Press: London.
Page 45
45
Halpern, A.R. (1989) ‘Memory for the absolute pitch of familiar songs.’ Memory &
Cognition, 17:5, 572-581.
Halpern, A. and G. Bower (1982) ‘Musical Expertise and Melodic Structure in Memory for
Musical Notation.’ The American Journal of Psychology, 95:1, 31-50.
Halpern, A. and Robert J. Zatorre (1999) ‘When that tune runs through your head: a PET
investigation of auditory imagery for familiar melodies.’ Cerebral Cortex, 9, 697-704.
Houston, D. and Geoffrey Haddock (2007) ‘On auditing auditory information: the influence
of mood on memory for music.’ Psychology of Music, 35:2, 201-212.
Janata, P. (2001) ‘Brain Electrical Activity Evoked by Mental Formation of Auditory
Expectations and Images.’ Brain Topography, 13:3, 169-193.
Janata, P., Jeffrey L. Birk, John D. Van Horn, Marc Leman, Barbara Tillmann, Jamshed J.
Bharucha (2002) ‘The Cortical Topography of Tonal Structures Underlying Western
Music.’ Science, 298, 2167-2170.
Keenan, J.P., Ven Thangaraj, Andrea R. Halpern and Gottfried Schlaug (2001) ‘Absolute
Pitch and Planum Temporale.’ NeuroImage, 14, 1402–1408.
Koelsch, S. and Walter A. Siebel (2005) ‘Towards a neural basis of music perception.’
TRENDS in Cognitive Sciences, 9:12, 578-584.
Kopiez, R., C. Weihs, U. Ligges, and J.I. Lee (2006) ‘Classification of high and low
achievers in a music sight-reading task.’ Psychology of Music, 34:1, 5-26.
Lamont, A. and Nicola Dibben (2001) ‘Motivic Structure and the Perception of Similarity.’
Music Perception, 18:3, 245-274.
Lehmann, A., J. Sloboda and R. Woody (2007) Psychology for Musicians. Oxford: Oxford
University Press.
Long, A.P. (1977) ‘Relationships between Pitch Memory in Short Melodies and Selected
Factors.’ Journal of Research in Music Education, 25:4, 272-282.
Miyazaki, K. (1992) ‘Perception of Musical Intervals by Absolute Pitch Possessors.’ Music
Perception: an Interdisciplinary Journal, 9:4, 413-426.
Powerhousemuseum.com (2001) Cochlear implant. [online] Available at:
http://www.powerhousemuseum.com/hsc/cochlear/hearing.htm [Accessed: 27 Apr 2013]
Palmer, C. (2005) ‘Sequence Memory in Music Performance.’ Current Directions in
Psychological Science, 14:5, 247-250.
Pascual-Leone, A. (2001) ‘The brain that plays music and is changed by it.’ Available at:
http://tmslab.org/wp-content/files/PascualLeone_MUSICBRAIN_NYAcadSci.pdf
[Accessed: 26 Apr 2013].
Peretz, I. (1993) ‘Auditory Atonalia for Melodies.’ Cognitive Neuropsychology, 10:1, 21-56.
Page 46
46
Peretz, I. (1996) ‘Can We Lose Memory for Music? A Case of Music Agnosia in a
Nonmusician.’ Journal of Cognitive Neuroscience, 8:6, 481-496.
Peretz, I. & Robert J. Zatorre (2005) ‘Brain organization for music processing.’ Annual
Review of Psychology, 56, 89-114.
Pierce, J. R. (1999) ‘The Nature of Musical Sound’ in The Psychology of Music Deutsch, D.
(ed.) pp. 1-24 London: Academic Press
Sacchetti, E., Alessandro Galluzzo, Adelaide Panariello, Giovanni Parrinello and Stefano
Francesco Cappa (2008). ‘Self-ordered pointing and visual conditional associative
learning tasks in drug-free schizophrenia spectrum disorder patients.’ BMC Psychiatry,
8:6.
Scott, S.K., C.C. Blank, S. Rosen and R.J.S. Wise (2000) ‘Identification of a pathway for
intelligible speech in the left temporal lobe’. Brain, 123, 2400-2406.
Shuter-Dyson, R. (1999) 'Musical Ability' in Psychology for Musicians D. Deutsch (ed.) pp
627-651, London: Academic Press.
Skidmore.edu (n.d.) Auditory Stimulus & Anatomy. [online] Available at:
http://www.skidmore.edu/~hfoley/PercLabs/Auditory1.htm [Accessed: 26 Apr 2013].
Sloboda, J. (1974) ‘The Eye-Hand Span – An Approach to the Study of Sight-Reading.’
Psychology of Music, 2:2, 4-10.
Sloboda, J (1978) 'The Psychology of Music' in Exploring the Musical Mind J. Sloboda
(ed.), 3-26, New York: Oxford University Press.
Sloboda, J (2005) Exploring the Musical Mind. New York: Oxford University Press.
Sloboda, J and D.H.H. Parker (2005) 'Immediate Recall of Melodies' in Exploring the
Musical Mind J. Sloboda (ed.), 71-96, New York: Oxford University Press.
Snyder, B (2009) 'Memory for Music' in Oxford Handbook of Music Psychology. S. Hallam,
I. Cross and M. Thaut (eds.), pp. 107-118, Oxford: Oxford University Press.
Stewart, L., K. von Kriegstein, J.D. Warren, and T.D. Griffiths (2006) ‘Music and the brain:
disorders of musical listening.’ Brain, 129, 2533-2553.
Tervaniemi, M. (2003) ‘Musical Sound Processing: EEG and MEG evidence’ in The
Cognitive Neuroscience of Music Peretz and Zatorre (ed.), pp. 294-309, Oxford: Oxford
University Press.
Tillmann, B. and Jamshed J. Bharucha (2000) ‘Implicit Learning of Tonality: A self-
organizing approach.’ Psychological Review, 107:4, 885-913.
Vignolo, Luigi A. (2003) ‘Music Agnosia and Auditory Agnosia: Dissociations in Stroke
Patients.’ Annals of the New York Academy of Sciences, 999, 50-57.
Page 47
47
Weinberger, Norman M. (1999) ‘Music and the Auditory System’ in The Psychology of
Music Deutsch, D. (ed.), pp. 47-89, London: Academic Press.
Zatorre, R.J., Alan C Evans and Ernst Meyer (1994) ‘Neural Mechanisms Underlying
Melodic Perception and Memory for Pitch.’ The Journal of Neuroscience, 14:4, 1908-
1919.
Zatorre, R. J., David W. Perry, Christine A. Beckett, Christopher F. Westbury and Alan C.
Evans (1998) ‘Functional anatomy of musical processing in listeners with absolute pitch
and relative pitch.’ Neurobiology, 95, 3172-3177.