PALAIOS, 2014, v. 29, 401–413 Research Article DOI: http://dx.doi.org/10.2110/palo.2014.013 PALEOECOLOGY AND TAPHONOMY OF TRACE FOSSILS IN THE EOLIAN UPPER TRIASSIC/LOWER JURASSIC NUGGET SANDSTONE, NORTHEASTERN UTAH THOMAS R. GOOD AND A.A. EKDALE University of Utah, Department of Geology and Geophysics, Salt Lake City, Utah, 84112, USA e-mail: [email protected]ABSTRACT: The Upper Triassic/Lower Jurassic Nugget Sandstone represents a portion of a vast inland erg of eolian sand dunes that were populated by diverse vertebrates and invertebrates, represented primarily by their trace fossils. Body fossils are rare, making the trace fossil record essential for deciphering the paleoecology of the ancient dune system. Trace fossils in the Nugget Sandstone near Vernal, Utah, include invertebrate burrows (Entradichnus meniscus, Entradichnus isp., Planolites beverleyensis, Taenidium isp. ‘‘A’’, Taenidium isp. ‘‘B’’, Skolithos, and Planolites isp., burrow clusters, large oblique burrows, flared burrows) and trackways (Paleohelcura and Octopodichnus). Arthropods (insects and arachnids) probably were the tracemakers of most, if not all, the trace fossils. Sediment moisture must have played a key role in the production and preservation of the trace fossils, indicating that moisture was important for supporting such complex ecosystems. Extended wet climatic intervals must have persisted intermittently between arid intervals. New fossil evidence for plants (sphenophytes, cycads, and algal buildups), ichnologic evidence of herbivorous insects and carnivorous arachnids, as well as indirect evidence for environmental moisture content during deposition of the Nugget Sandstone, provide a picture of the paleoecology of this ancient sea of sand. INTRODUCTION Eolian deposits typically are devoid of abundant body fossils and trace fossils, so clues for interpreting details of paleoenvironmental conditions are few and far between. In such settings, trace fossils are preserved only under certain conditions, and they may provide important information about not only the inhabitants but also about the presence of moisture in the environment. The Nugget Sandstone is interpreted as a Upper Triassic/Lower Jurassic correlative of the Glen Canyon Group of the Colorado Plateau of western North America (Wingate, Kayenta, and Navajo Sandstones; Sprinkel et al. 2011). Together they represent one of the largest eolian environments to have ever existed. The Nugget Sandstone is one of the best exposed formations in the Dinosaur National Monument area of northeastern Utah (Fig. 1), where it is a light-colored, often bleached, prominent ridge-former varying in thickness from about 180–200 m (Gregson and Chure 2000). The Nugget Sandstone is dominated by large-scale, high-angle, trough cross-bedded eolian sandstone. Flat, finer-grained, thin, wavy, or irregular beds are not uncommon and are interpreted as interdune deposits (Doelger 1987). The Nugget paleoenvironment certainly was a desert, but it must have been, at least intermittently, a surprisingly tolerable environment. Most reports of trace fossils in the Nugget are cursory, mainly mentioning surficial trackways and trails (Table 1). Body fossils of invertebrates are extremely rare, but body fossils of vertebrates are locally abundant. The Saints and Sinners Quarry in northeastern Utah has yielded thousands of bones and bone fragments of ceolophysoids, drepanosaurs, and theropods, and even possible cycadeoid impressions, from an interdune lake deposit in the Nugget Sandstone of northeastern Utah. (Britt et al. 2010, 2011; Chambers et al. 2011; Engelmann et al. 2011, 2012). Sphenophyte remains have also been identified within Dinosaur National Monument (Daniel Chure, written communication 2012). Large tetrapod trackways in the Nugget and Navajo Sandstones have long been a subject of interest to both scientists and the general public, and smaller trace fossils afford clues to the community structure and ecosystem that supported these carnivorous vertebrates. Suggestions regarding climatic conditions during deposition of the Nugget Sandstone and its eolian equivalents have been offered by numerous workers (High and Picard 1975; Picard 1977b; Stokes 1991; Chandler et al. 1992; Chan and Archer 1999, 2000; Loope et al. 2001; Eisenberg 2003; Loope and Rowe 2003; Loope et al. 2004b; Wilkens 2008; Bryant 2011). TRACE FOSSILS Previously described traces from northeastern Utah include vertebrate trackways (Brasilichnium, Grallator, Eubrontes, Brachychirotherium, Pseudotetrasauropus, Tetrasauropus, and Otozoum tracks, Table 1). New trace fossil occurrences from this area include invertebrate burrows (Entradichnus meniscus, Entradichnus isp., Planolites beverleyensis, Taenidium isp. ‘‘A’’, Taenidium isp. ‘‘B’’, Skolithos, and Planolites isp., burrow clusters, large oblique burrows, flared burrows) and trackways (Paleohelcura, and Octopodichnus). Figure 2 shows the spatial and facies relationships of the ichnotaxa and body fossils of the Nugget Sandstone in this area. Table 2 describes the trace fossils of the Nugget Sandstone in northeastern Utah, and Table 3 outlines the interpreted dune facies, tracemakers, and sediment moisture content at time of production for each trace fossil. Entradichnus meniscus Entradichnus meniscus (Fig. 3A–B; Table 2) commonly is in close association with other burrows in local abundance, including Planolites Published Online: November 2014 Copyright E 2014, SEPM (Society for Sedimentary Geology) 0883-1351/14/029-401/$03.00
Transcript
PALAIOS, 2014, v. 29, 401–413
Research Article
DOI: http://dx.doi.org/10.2110/palo.2014.013
PALEOECOLOGY AND TAPHONOMY OF TRACE FOSSILS IN THE EOLIAN UPPER TRIASSIC/LOWERJURASSIC NUGGET SANDSTONE, NORTHEASTERN UTAH
THOMAS R. GOOD AND A.A. EKDALEUniversity of Utah, Department of Geology and Geophysics, Salt Lake City, Utah, 84112, USA
ABSTRACT: The Upper Triassic/Lower Jurassic Nugget Sandstone represents a portion of a vast inland erg of eolian sand dunesthat were populated by diverse vertebrates and invertebrates, represented primarily by their trace fossils. Body fossils are rare,making the trace fossil record essential for deciphering the paleoecology of the ancient dune system. Trace fossils in the NuggetSandstone near Vernal, Utah, include invertebrate burrows (Entradichnus meniscus, Entradichnus isp., Planolites beverleyensis,Taenidium isp. ‘‘A’’, Taenidium isp. ‘‘B’’, Skolithos, and Planolites isp., burrow clusters, large oblique burrows, flared burrows)and trackways (Paleohelcura and Octopodichnus). Arthropods (insects and arachnids) probably were the tracemakers of most, ifnot all, the trace fossils. Sediment moisture must have played a key role in the production and preservation of the trace fossils,indicating that moisture was important for supporting such complex ecosystems. Extended wet climatic intervals must havepersisted intermittently between arid intervals. New fossil evidence for plants (sphenophytes, cycads, and algal buildups),ichnologic evidence of herbivorous insects and carnivorous arachnids, as well as indirect evidence for environmental moisturecontent during deposition of the Nugget Sandstone, provide a picture of the paleoecology of this ancient sea of sand.
INTRODUCTION
Eolian deposits typically are devoid of abundant body fossils and tracefossils, so clues for interpreting details of paleoenvironmental conditionsare few and far between. In such settings, trace fossils are preserved onlyunder certain conditions, and they may provide important informationabout not only the inhabitants but also about the presence of moisture inthe environment.
The Nugget Sandstone is interpreted as a Upper Triassic/LowerJurassic correlative of the Glen Canyon Group of the Colorado Plateauof western North America (Wingate, Kayenta, and Navajo Sandstones;Sprinkel et al. 2011). Together they represent one of the largest eolianenvironments to have ever existed. The Nugget Sandstone is one of thebest exposed formations in the Dinosaur National Monument area ofnortheastern Utah (Fig. 1), where it is a light-colored, often bleached,prominent ridge-former varying in thickness from about 180–200 m(Gregson and Chure 2000). The Nugget Sandstone is dominated bylarge-scale, high-angle, trough cross-bedded eolian sandstone. Flat,finer-grained, thin, wavy, or irregular beds are not uncommon and areinterpreted as interdune deposits (Doelger 1987).
The Nugget paleoenvironment certainly was a desert, but it must havebeen, at least intermittently, a surprisingly tolerable environment. Mostreports of trace fossils in the Nugget are cursory, mainly mentioningsurficial trackways and trails (Table 1). Body fossils of invertebrates areextremely rare, but body fossils of vertebrates are locally abundant. TheSaints and Sinners Quarry in northeastern Utah has yielded thousands ofbones and bone fragments of ceolophysoids, drepanosaurs, andtheropods, and even possible cycadeoid impressions, from an interdunelake deposit in the Nugget Sandstone of northeastern Utah. (Britt et al.2010, 2011; Chambers et al. 2011; Engelmann et al. 2011, 2012).Sphenophyte remains have also been identified within Dinosaur NationalMonument (Daniel Chure, written communication 2012). Large tetrapod
trackways in the Nugget and Navajo Sandstones have long been a subjectof interest to both scientists and the general public, and smaller tracefossils afford clues to the community structure and ecosystem thatsupported these carnivorous vertebrates.
Suggestions regarding climatic conditions during deposition of theNugget Sandstone and its eolian equivalents have been offered bynumerous workers (High and Picard 1975; Picard 1977b; Stokes 1991;Chandler et al. 1992; Chan and Archer 1999, 2000; Loope et al. 2001;Eisenberg 2003; Loope and Rowe 2003; Loope et al. 2004b; Wilkens 2008;Bryant 2011).
TRACE FOSSILS
Previously described traces from northeastern Utah include vertebrate
Taenidium isp. ‘‘A’’, Taenidium isp. ‘‘B’’, Skolithos, and Planolites isp.,
burrow clusters, large oblique burrows, flared burrows) and trackways
(Paleohelcura, and Octopodichnus). Figure 2 shows the spatial and facies
relationships of the ichnotaxa and body fossils of the Nugget Sandstone
in this area. Table 2 describes the trace fossils of the Nugget Sandstone in
northeastern Utah, and Table 3 outlines the interpreted dune facies,
tracemakers, and sediment moisture content at time of production for
each trace fossil.
Entradichnus meniscus
Entradichnus meniscus (Fig. 3A–B; Table 2) commonly is in closeassociation with other burrows in local abundance, including Planolites
Published Online: November 2014Copyright E 2014, SEPM (Society for Sedimentary Geology) 0883-1351/14/029-401/$03.00
beverleyensis and Entradichnus isp. Created by burrowers moving instraight or gently curved lines on the lower dune slipface (Ekdale andPicard 1985), E. meniscus burrows generally are parallel to the foresetlaminae of the cross-bedded strata. In the Nugget Sandstone, E. meniscus
has no discernable preferential orientation, but a few examples showclusters of Entradichnus radiating from a central area (Fig. 3B). Thispattern can be observed in the Entradichnus meniscus of the NavajoSandstone of southern Utah (Ekdale et al. 2007). Unlike reports ofEntradichnus elsewhere, E. meniscus in the Nugget are preserved both innegative and positive hyporelief and epirelief, and sometimes they arepreserved without discernable relief, seen by a contrasting color to the
surrounding sediment. A majority of the horizontal, unwalled, un-branched traces in this area, which include Entradichnus isp. andPlanolites beverleyensis, are preserved in hyporelief. Grouping of menisciis observed in the best-preserved specimens of E. meniscus, a featurereferred to as megamenisci by Frey et al. (1984).
Entradichnus differs from Taenidium, which is a branching tunnel,cylindrical in cross section, and which incorporates ingested (or at leastdistinctly different) backfill. Taenidium typically does not strictly followbedding planes, and it usually is shorter and less straight thanEntradichnus. Some Taenidium ichnospecies have been described withexceptions to these generally accepted characteristics (D’Alessandro andBromley 1987). However, Entradichnus is differentiated from Taenidium
on the basis of the combined set of characteristics that differ fromtraditional and general diagnostic features of Taenidium.
FIG. 1.—Area map showing investigated outcrops of the Nugget Sandstone innortheastern Utah.
TABLE 1.— Trace fossils in the Nugget Sandstone.
Fossil Area or formation Reference
Trackways (Later named Paleohelcura(?) and Brasilichnium by Chure et al. 2014) N Utah Buss, 1921Trackways and burrows (Lacertoid tracks, Brasilichnium, Paleohelcura,
Octopodichnus, Undescribed trails)NW Colorado Faul and Roberts, 1951
Bilobed trail (Later named Gyrochorte by Picard 1975; Knapp 1976) C Wyoming Wroble, 1953Small vertebrate trackways NE Utah Stokes, 1957, 1959 (from Picard 1977a)Possible vertebrate track C Wyoming Kayser, 1964 (from Doelger 1987)‘‘Organic trails’’ C Wyoming Jordan, 1965 (from Picard 1977a)‘‘Burrow structures’’ E Utah Stanley et al., 1971Trackways and burrows (Later named Brasilichnium, Lacertoid tracks, Paleohelcura,
Octopodichnus, Entradichnus, by Chure et al. 2014)NE Utah Albers, 1975
Invertebrate trails NW Colorado Knapp, 1976Small vertical and horizontal burrows W Wyoming, N Utah Pacht, 1976Dinosaur and reptilian trackways, invertebrate burrows and trails Wyoming, Utah, Colorado, Idaho Stokes, 1978Small vertical and horizontal burrows Wyoming, Utah Dott, 1979Meniscate burrows C Wyoming Doelger, 1981Eubrontes, Grallator NE Utah Hamblin and Bilbey, 1999Trackways and burrows (Brasilichnium, Octopodichnus, Taenidium,
Entradichnus, Beaconites)N Utah Sanders and Picard, 1999
NE Utah Lockley et al., 1991; Lockley et al., 1992a-c;Lockley et al., 2001; Lucas et al., 2006
Brasilichnium NE Utah Engelmann et al., 2010Cheirothere tracks (Brachychirotherium) NE Utah Anderson, 2011Large burrows NE Utah Engelmann et al., 2014Entradichnus, Planolites, Taenidium, Skolithos, ‘burrow clusters’, and ‘flared
burrows’NE Utah This study
FIG. 2.—Idealized sketch of trace fossil and body fossil associations in theNugget Sandstone of northeastern Utah. Bc 5 burrow clusters; Br 5Brasilichnium; Ent 5 Entradichnus; Fb 5 flared burrows; G/E 5 Grallator andEubrontes; Gp 5 gastropods; Lb 5 large oblique burrows; Oct 5 Octopodichnus;Ph 5 Paleohelcura; Pl 5 Planolites beverleyensis; S/P 5 Skolithos and Planolitesisp.; Tn 5 Taenidium; Bb 5 bone bed.
402 T.R. GOOD AND A.A. EKDALE P A L A I O S
Entradichnus isp.
Entradichnus isp. (Fig. 3B–D; Table 2) exhibits similar characteristicsto, and is always associated with, Entradichnus meniscus and Planolitesbeverleyensis, but is distinguished by its distinctive feather stitch internalstructure. In the Nugget Sandstone, Entradichnus isp. occurs in negativeand positive hyporelief (Fig. 3C) and also with no relief as a contrastingcolor to the surrounding sediment (Fig. 3D). Entradichnus isp. is rarerthan both Entradichnus meniscus and Planolites beverleyensis, butmultiple specimens occur at two localities. One occurrence shows thefeather stitch backfill transition into the meniscate backfill of Entradich-nus meniscus within the same burrow, indicating that the same tracemakerwas capable of producing two types of backfill.
This feather stitch backfill may be the result of an organism pushing thesediment around its body with alternating appendage motion, ortransporting sediment behind its side-alternating posterior. E. isp.contrasts with E. meniscus, which is produced as sediment is pushed
around the body with simultaneous appendage motion, or somehowtransported behind the animal evenly.
Planolites beverleyensis
Planolites beverleyensis (Fig. 3E–F; Table 2) constitutes the majority ofthe horizontal traces in this area, where occurrences range from dense(hundreds of burrows per m2) to highly sparse (only one or two observedin a single locality). In dense occurrences they not only are grouped onsingle surfaces, but also in lateral depositional dip sections reaching asmuch as 30 m.
Planolites beverleyensis is the most commonly observed trace fossil inthe study area, and always occurs on lower slipface surfaces of ancientdunes. Its close association with Entradichnus suggests that P. bever-leyensis was produced by the same tracemaker. In this area thepreservation of internal structure in horizontal traces ranges from notpreserved to very well preserved.
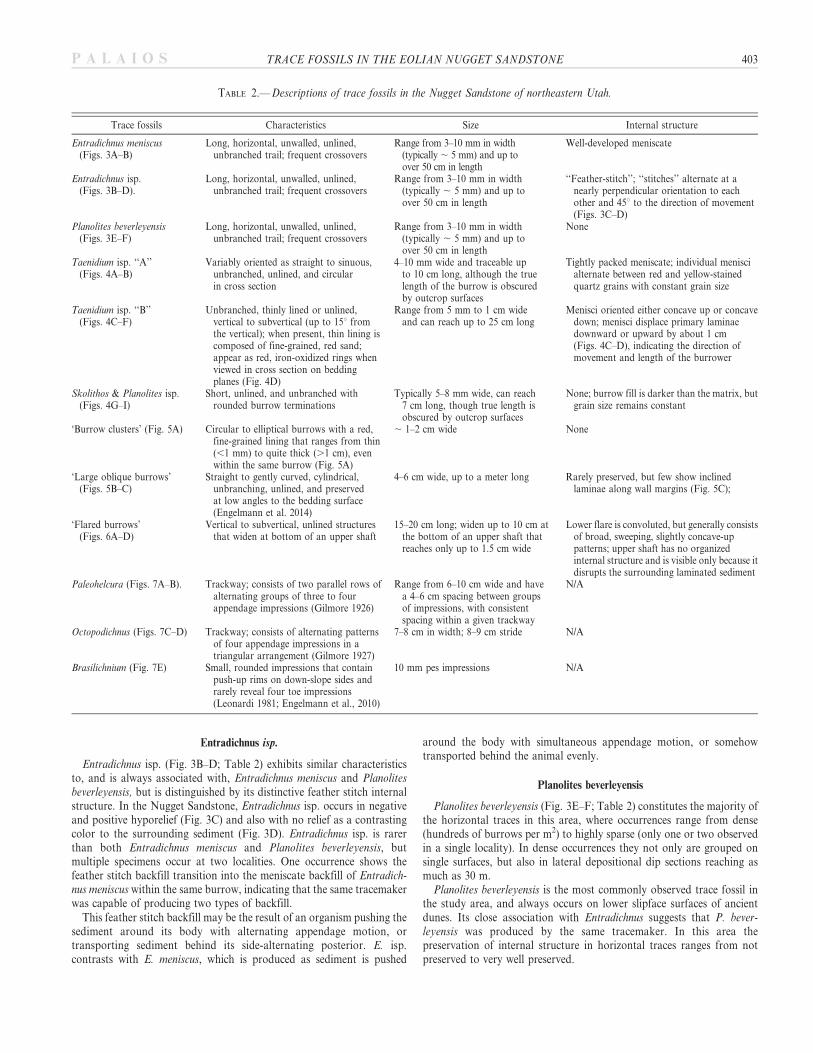
TABLE 2.—Descriptions of trace fossils in the Nugget Sandstone of northeastern Utah.
Range from 3–10 mm in width(typically , 5 mm) and up toover 50 cm in length
None
Taenidium isp. ‘‘A’’(Figs. 4A–B)
Variably oriented as straight to sinuous,unbranched, unlined, and circularin cross section
4–10 mm wide and traceable upto 10 cm long, although the truelength of the burrow is obscuredby outcrop surfaces
Tightly packed meniscate; individual meniscialternate between red and yellow-stainedquartz grains with constant grain size
Taenidium isp. ‘‘B’’(Figs. 4C–F)
Unbranched, thinly lined or unlined,vertical to subvertical (up to 15u fromthe vertical); when present, thin lining iscomposed of fine-grained, red sand;appear as red, iron-oxidized rings whenviewed in cross section on beddingplanes (Fig. 4D)
Range from 5 mm to 1 cm wideand can reach up to 25 cm long
Menisci oriented either concave up or concavedown; menisci displace primary laminaedownward or upward by about 1 cm(Figs. 4C–D), indicating the direction ofmovement and length of the burrower
Skolithos & Planolites isp.(Figs. 4G–I)
Short, unlined, and unbranched withrounded burrow terminations
Typically 5–8 mm wide, can reach7 cm long, though true length isobscured by outcrop surfaces
None; burrow fill is darker than the matrix, butgrain size remains constant
‘Burrow clusters’ (Fig. 5A) Circular to elliptical burrows with a red,fine-grained lining that ranges from thin(,1 mm) to quite thick (.1 cm), evenwithin the same burrow (Fig. 5A)
, 1–2 cm wide None
‘Large oblique burrows’(Figs. 5B–C)
Straight to gently curved, cylindrical,unbranching, unlined, and preservedat low angles to the bedding surface(Engelmann et al. 2014)
4–6 cm wide, up to a meter long Rarely preserved, but few show inclinedlaminae along wall margins (Fig. 5C);
‘Flared burrows’(Figs. 6A–D)
Vertical to subvertical, unlined structuresthat widen at bottom of an upper shaft
15–20 cm long; widen up to 10 cm atthe bottom of an upper shaft thatreaches only up to 1.5 cm wide
Lower flare is convoluted, but generally consistsof broad, sweeping, slightly concave-uppatterns; upper shaft has no organizedinternal structure and is visible only because itdisrupts the surrounding laminated sediment
Paleohelcura (Figs. 7A–B). Trackway; consists of two parallel rows ofalternating groups of three to fourappendage impressions (Gilmore 1926)
Range from 6–10 cm wide and havea 4–6 cm spacing between groupsof impressions, with consistentspacing within a given trackway
N/A
Octopodichnus (Figs. 7C–D) Trackway; consists of alternating patternsof four appendage impressions in atriangular arrangement (Gilmore 1927)
7–8 cm in width; 8–9 cm stride N/A
Brasilichnium (Fig. 7E) Small, rounded impressions that containpush-up rims on down-slope sides andrarely reveal four toe impressions(Leonardi 1981; Engelmann et al., 2010)
10 mm pes impressions N/A
TRACE FOSSILS IN THE EOLIAN NUGGET SANDSTONE 403P A L A I O S
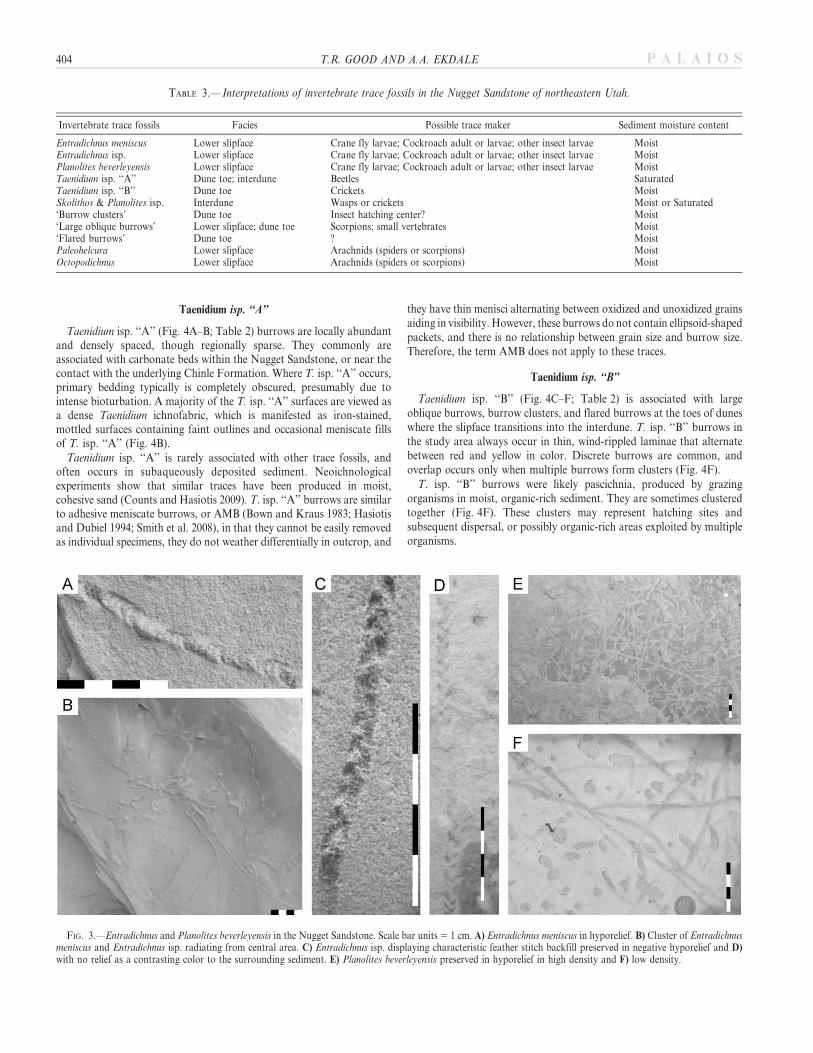
Taenidium isp. ‘‘A’’
Taenidium isp. ‘‘A’’ (Fig. 4A–B; Table 2) burrows are locally abundantand densely spaced, though regionally sparse. They commonly areassociated with carbonate beds within the Nugget Sandstone, or near thecontact with the underlying Chinle Formation. Where T. isp. ‘‘A’’ occurs,primary bedding typically is completely obscured, presumably due tointense bioturbation. A majority of the T. isp. ‘‘A’’ surfaces are viewed asa dense Taenidium ichnofabric, which is manifested as iron-stained,mottled surfaces containing faint outlines and occasional meniscate fillsof T. isp. ‘‘A’’ (Fig. 4B).
Taenidium isp. ‘‘A’’ is rarely associated with other trace fossils, andoften occurs in subaqueously deposited sediment. Neoichnologicalexperiments show that similar traces have been produced in moist,cohesive sand (Counts and Hasiotis 2009). T. isp. ‘‘A’’ burrows are similarto adhesive meniscate burrows, or AMB (Bown and Kraus 1983; Hasiotisand Dubiel 1994; Smith et al. 2008), in that they cannot be easily removedas individual specimens, they do not weather differentially in outcrop, and
they have thin menisci alternating between oxidized and unoxidized grainsaiding in visibility. However, these burrows do not contain ellipsoid-shapedpackets, and there is no relationship between grain size and burrow size.Therefore, the term AMB does not apply to these traces.
Taenidium isp. ‘‘B’’
Taenidium isp. ‘‘B’’ (Fig. 4C–F; Table 2) is associated with largeoblique burrows, burrow clusters, and flared burrows at the toes of duneswhere the slipface transitions into the interdune. T. isp. ‘‘B’’ burrows inthe study area always occur in thin, wind-rippled laminae that alternatebetween red and yellow in color. Discrete burrows are common, andoverlap occurs only when multiple burrows form clusters (Fig. 4F).
T. isp. ‘‘B’’ burrows were likely pascichnia, produced by grazingorganisms in moist, organic-rich sediment. They are sometimes clusteredtogether (Fig. 4F). These clusters may represent hatching sites andsubsequent dispersal, or possibly organic-rich areas exploited by multipleorganisms.
TABLE 3.— Interpretations of invertebrate trace fossils in the Nugget Sandstone of northeastern Utah.
Invertebrate trace fossils Facies Possible trace maker Sediment moisture content
Entradichnus meniscus Lower slipface Crane fly larvae; Cockroach adult or larvae; other insect larvae MoistEntradichnus isp. Lower slipface Crane fly larvae; Cockroach adult or larvae; other insect larvae MoistPlanolites beverleyensis Lower slipface Crane fly larvae; Cockroach adult or larvae; other insect larvae MoistTaenidium isp. ‘‘A’’ Dune toe; interdune Beetles SaturatedTaenidium isp. ‘‘B’’ Dune toe Crickets MoistSkolithos & Planolites isp. Interdune Wasps or crickets Moist or Saturated‘Burrow clusters’ Dune toe Insect hatching center? Moist‘Large oblique burrows’ Lower slipface; dune toe Scorpions; small vertebrates Moist‘Flared burrows’ Dune toe ? MoistPaleohelcura Lower slipface Arachnids (spiders or scorpions) MoistOctopodichnus Lower slipface Arachnids (spiders or scorpions) Moist
FIG. 3.—Entradichnus and Planolites beverleyensis in the Nugget Sandstone. Scale bar units 5 1 cm. A) Entradichnus meniscus in hyporelief. B) Cluster of Entradichnusmeniscus and Entradichnus isp. radiating from central area. C) Entradichnus isp. displaying characteristic feather stitch backfill preserved in negative hyporelief and D)with no relief as a contrasting color to the surrounding sediment. E) Planolites beverleyensis preserved in hyporelief in high density and F) low density.
404 T.R. GOOD AND A.A. EKDALE P A L A I O S
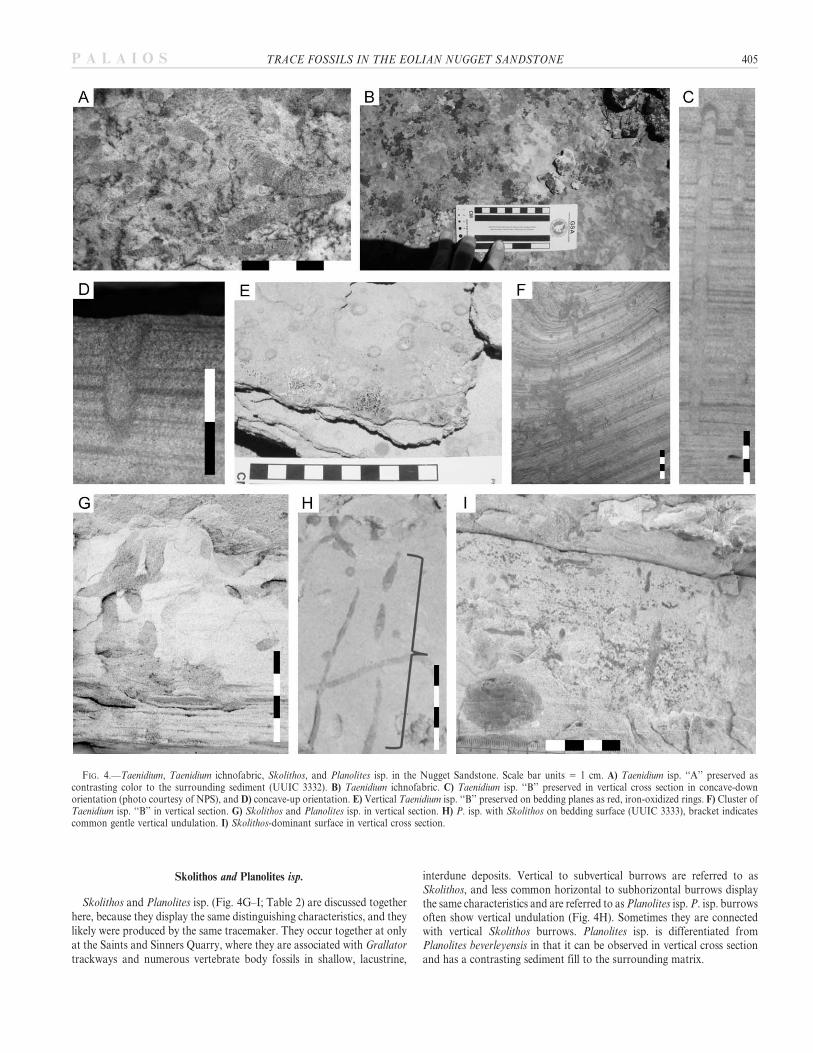
Skolithos and Planolites isp.
Skolithos and Planolites isp. (Fig. 4G–I; Table 2) are discussed togetherhere, because they display the same distinguishing characteristics, and theylikely were produced by the same tracemaker. They occur together at onlyat the Saints and Sinners Quarry, where they are associated with Grallator
trackways and numerous vertebrate body fossils in shallow, lacustrine,
interdune deposits. Vertical to subvertical burrows are referred to asSkolithos, and less common horizontal to subhorizontal burrows displaythe same characteristics and are referred to as Planolites isp. P. isp. burrowsoften show vertical undulation (Fig. 4H). Sometimes they are connectedwith vertical Skolithos burrows. Planolites isp. is differentiated fromPlanolites beverleyensis in that it can be observed in vertical cross sectionand has a contrasting sediment fill to the surrounding matrix.
FIG. 4.—Taenidium, Taenidium ichnofabric, Skolithos, and Planolites isp. in the Nugget Sandstone. Scale bar units 5 1 cm. A) Taenidium isp. ‘‘A’’ preserved ascontrasting color to the surrounding sediment (UUIC 3332). B) Taenidium ichnofabric. C) Taenidium isp. ‘‘B’’ preserved in vertical cross section in concave-downorientation (photo courtesy of NPS), and D) concave-up orientation. E) Vertical Taenidium isp. ‘‘B’’ preserved on bedding planes as red, iron-oxidized rings. F) Cluster ofTaenidium isp. ‘‘B’’ in vertical section. G) Skolithos and Planolites isp. in vertical section. H) P. isp. with Skolithos on bedding surface (UUIC 3333), bracket indicatescommon gentle vertical undulation. I) Skolithos-dominant surface in vertical cross section.
TRACE FOSSILS IN THE EOLIAN NUGGET SANDSTONE 405P A L A I O S
Burrow Clusters
Burrow clusters (Fig. 5A; Table 2) typically consist of hundreds ofburrows that occur on vertical and oblique surfaces in wind-rippledsediment consisting of thin red and white laminae. They are associatedwith Taenidium isp. ‘‘B’’, large oblique burrows, and flared burrows. Theobserved features are not plant root structures based on their clusteringhabit, invariability in size, and tendency to overlap. They also are notconsidered open chambers due to common overlapping.
Burrow clusters may represent hatching centers, much like theclustering of Taenidium isp. ‘‘B’’. These burrows cannot readily beassigned to an ichnogenus, because clustering cannot be a diagnosticfeature, and because the true morphology is indiscernible.
Large Oblique Burrows
Large oblique burrows (Fig. 5B–C; Table 2) are associated withTaenidium isp. ‘‘B,’’ flared burrows, and burrow clusters, all of whichoccur exclusively in dune toe deposits. When burrows intersect theoutcrop surface, circular cross sections can be observed. Fill usuallyconsists of sand similar in texture and color to the surrounding matrix,
but it does not include laminae. Most are not preserved in relief, and thereis no evidence of bioglyphs or scratches. Lack of internal structureindicates that these were once open burrows.
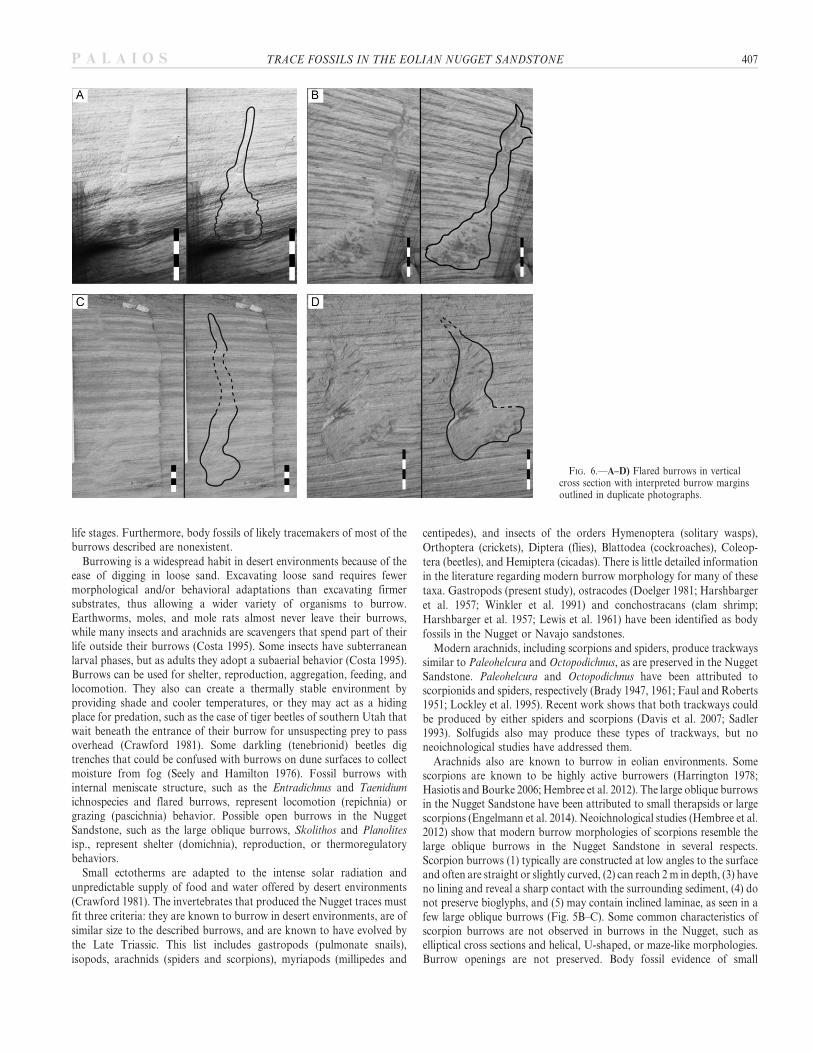
Flared Burrows
Flared burrows (Fig. 6A–D; Table 2) are associated with Taenidium isp.‘‘B’’, burrow clusters, and a large burrow, all of which occur in wind-rippled, dune toe deposits. Four flared burrows occur at two localitieswithin wind-rippled sediment consisting of thin red and white laminae.These burrows are perplexing, because it is unclear if the wide lowerportion was an open chamber, or just a broader disruption of the sedimentpossibly for feeding purposes. Close association with T. isp. ‘‘B’’ couldindicate production by the same tracemaker given that the shaft widths offlared burrows are similar to burrow widths of T. isp. ‘‘B’’. These featuresare distinguished from inorganic deformation features due to their sharpcontacts with the surrounding sediment, relative consistency in shape (thinupper shaft and flared bottom), and because no other evidence of soft-sediment deformation features in the area was observed.
Paleohelcura
Paleohelcura (Fig. 7A–B; Table 2) is associated with Octopodichnusand Brasilichnium. In the Nugget Sandstone, groups of impressionsgenerally contain three linearly arranged imprints, and no evidence ofbody or tail drags is present. Some trackways contain push-up rims on thedownslope side of impressions, suggesting that the tracemaker waswalking on a slope with more weight on its lower set of legs.
Octopodichnus
Octopodichnus (Fig. 7C–D; Table 2) is associated with Paleohelcuraand Brasilichnium. One side of some Octopodichnus trackways is poorlypreserved or absent, and many of the impressions show push-up rims onthe downslope side (Fig. 7C), suggesting that the organism was walkingon a slope with more weight on its lower set of legs. No evidence of bodyor tail drags is present.
Brasilichnium
Brasilichnium (Fig. 7E; Table 2) tracks in the Nugget Sandstone show apreference for traveling up the slipface (Engelmann et al. 2010). Theyoften include smaller manus impressions in addition to the larger pesimpressions. Brasilichnium are preserved on slipface surfaces in epireliefas impressions sometimes filled with coarser sand, and they typically areassociated with Paleohelcura and Octopodichnus. Brasilichnium is acommon trace fossil in Mesozoic eolian deposits, and it is attributed toquadrapedal synapsids (Lockley 2011).
IDENTITY OF TRACEMAKERS
Linking tracemakers with their trace fossils is facilitated by studies ofmodern burrowers in similar environments (Ahlbrandt et al. 1978;Tschinkel 2002; Davis et al. 2007; Smith et al. 2008; Smith and Hasiotis2008; Counts and Hasiotis 2009; Hembree 2009; Hembree et al. 2012).Current neoichnological studies offer information about only a fewexisting modern burrowers. Difficulties arise when comparing moderndune settings to the 200-million-year-old Nugget erg, primarily becausethere are no good modern analogues – environmental, ecologic, climatic,or sedimentologic. One reason for this is that the Nugget Sandstonepredates the evolution of angiosperms, social insects, and eutherianmammals, all of which dominate modern desert environments today.Interpretations also can be muddled by the fact that many organisms cancreate the same type of burrow, and that a single organism can produceseveral types of burrows. This is especially true for insects with multiple
FIG. 5.—Burrow clusters, large oblique burrows, and flared burrows in theNugget Sandstone. Scale bar units 5 1 cm. A) Burrow clusters on oblique section,inset shows close-up of burrow lining (similar to UUIC 3334). B) Large obliqueburrow in vertical section. C) Part of a large oblique burrow; arrow indicatesinclined laminae along margin.
406 T.R. GOOD AND A.A. EKDALE P A L A I O S
life stages. Furthermore, body fossils of likely tracemakers of most of theburrows described are nonexistent.
Burrowing is a widespread habit in desert environments because of theease of digging in loose sand. Excavating loose sand requires fewermorphological and/or behavioral adaptations than excavating firmersubstrates, thus allowing a wider variety of organisms to burrow.Earthworms, moles, and mole rats almost never leave their burrows,while many insects and arachnids are scavengers that spend part of theirlife outside their burrows (Costa 1995). Some insects have subterraneanlarval phases, but as adults they adopt a subaerial behavior (Costa 1995).Burrows can be used for shelter, reproduction, aggregation, feeding, andlocomotion. They also can create a thermally stable environment byproviding shade and cooler temperatures, or they may act as a hidingplace for predation, such as the case of tiger beetles of southern Utah thatwait beneath the entrance of their burrow for unsuspecting prey to passoverhead (Crawford 1981). Some darkling (tenebrionid) beetles digtrenches that could be confused with burrows on dune surfaces to collectmoisture from fog (Seely and Hamilton 1976). Fossil burrows withinternal meniscate structure, such as the Entradichnus and Taenidium
ichnospecies and flared burrows, represent locomotion (repichnia) orgrazing (pascichnia) behavior. Possible open burrows in the NuggetSandstone, such as the large oblique burrows, Skolithos and Planolitesisp., represent shelter (domichnia), reproduction, or thermoregulatorybehaviors.
Small ectotherms are adapted to the intense solar radiation and
unpredictable supply of food and water offered by desert environments
(Crawford 1981). The invertebrates that produced the Nugget traces must
fit three criteria: they are known to burrow in desert environments, are of
similar size to the described burrows, and are known to have evolved by
the Late Triassic. This list includes gastropods (pulmonate snails),
isopods, arachnids (spiders and scorpions), myriapods (millipedes and
centipedes), and insects of the orders Hymenoptera (solitary wasps),
et al. 1957; Winkler et al. 1991) and conchostracans (clam shrimp;
Harshbarger et al. 1957; Lewis et al. 1961) have been identified as body
fossils in the Nugget or Navajo sandstones.
Modern arachnids, including scorpions and spiders, produce trackwayssimilar to Paleohelcura and Octopodichnus, as are preserved in the NuggetSandstone. Paleohelcura and Octopodichnus have been attributed toscorpionids and spiders, respectively (Brady 1947, 1961; Faul and Roberts1951; Lockley et al. 1995). Recent work shows that both trackways couldbe produced by either spiders and scorpions (Davis et al. 2007; Sadler1993). Solfugids also may produce these types of trackways, but noneoichnological studies have addressed them.
Arachnids also are known to burrow in eolian environments. Somescorpions are known to be highly active burrowers (Harrington 1978;Hasiotis and Bourke 2006; Hembree et al. 2012). The large oblique burrowsin the Nugget Sandstone have been attributed to small therapsids or largescorpions (Engelmann et al. 2014). Neoichnological studies (Hembree et al.2012) show that modern burrow morphologies of scorpions resemble thelarge oblique burrows in the Nugget Sandstone in several respects.Scorpion burrows (1) typically are constructed at low angles to the surfaceand often are straight or slightly curved, (2) can reach 2 m in depth, (3) haveno lining and reveal a sharp contact with the surrounding sediment, (4) donot preserve bioglyphs, and (5) may contain inclined laminae, as seen in afew large oblique burrows (Fig. 5B–C). Some common characteristics ofscorpion burrows are not observed in burrows in the Nugget, such aselliptical cross sections and helical, U-shaped, or maze-like morphologies.Burrow openings are not preserved. Body fossil evidence of small
FIG. 6.—A–D) Flared burrows in verticalcross section with interpreted burrow marginsoutlined in duplicate photographs.
TRACE FOSSILS IN THE EOLIAN NUGGET SANDSTONE 407P A L A I O S
vertebrates is present in the Nugget Sandstone, so burrow production bysmall vertebrates cannot be ruled out.
Wasps can create open vertical, subvertical, or horizontal burrows,which may be test, sleeping, nesting, and false burrows (Ahlbrandt et al.1978). Modern wasps are particular about texture and cohesiveness of thesubstrate (Ahlbrandt et al. 1978), and they typically prefer to burrow inthe stoss sides of dunes (Loope and Rowe 2003). Skolithos and Planolitesisp. burrows could be attributed to wasps, although they are located in aninterdune interval that is thought to have been deposited in standingwater.
Crickets can create straight burrows, sometimes branching, orientedobliquely into damp, cohesive dune slopes on both stoss and lee sides(Ahlbrandt et al. 1978; Loope and Rowe 2003). Some sections of theburrows may be backfilled, while others remain open. Taenidium isp. ‘‘B’’,Skolithos, and Planolites isp. could be attributed to crickets.
Fly larvae burrow just beneath dune surfaces and produce meniscate-filled burrows (Ahlbrandt et al. 1978). Entradichnus meniscus has beenattributed to crane fly larvae (Ekdale et al. 2007; Seike et al. 2010). Onelocality in the Nugget Sandstone shows examples of Entradichnus
meniscus that transition into Entradichnus isp., interpreted as beingproduced by alternations in appendage movement, within the sameburrow. A similar style of backfill described by Bromley and Asgaard(1979) is interpreted as alternations in the direction of the animal’s anusas pulses of transported sediment are deposited, either by egestion orperistalsis. Entradichnus and Planolites beverleyensis, which may bepoorly preserved Entradichnus, can be attributed to fly larvae.
Both larval and adult cockroaches, like crane fly larvae, are known toburrow horizontally just beneath surfaces of sand dunes (Hawke andFarley 1973). Similarly Entradichnus meniscus, Entradichnus isp., andPlanolites beverleyensis could be attributed to cockroaches. Little isknown about the morphology of cockroach burrows, but if adultcockroaches produce horizontal meniscate traces, it is possible thatEntradichnus meniscus and Entradichnus isp. could have been produced bycockroaches.
Beetle larvae can produce straight, vertical to subvertical burrows withdensely packed meniscate fill in subaerial, moist, sandy soil (Counts andHasiotis 2009). Taenidium isp. ‘‘A’’ can be attributed to beetles.
Cicadas also can produce burrows similar to Taenidium isp. ‘‘A’’(Smith et al. 2008), though cicada burrows contain ellipsoid-shapedpackets not seen in T. isp. ‘‘A’’. Cicada burrows typically occur in alluvialdeposits and paleosols.
The flared burrows and burrow clusters remain enigmatic. They couldbe the work of taxa that no longer exist, behaviors that no longer exist, orbehaviors that have yet to be described in modern settings.
DISCUSSION
Role of Water in Preservation of Eolian Trace Fossils
Anyone who has visited a modern dune field might conclude that tracesin dry eolian sand have little potential for preservation. Trackwaysproduced during the night typically are blown away by midday, burrowsconstructed in loose sand immediately collapse unless reinforced biogeni-cally, and meniscate burrows are nearly impossible to produce in truly drysand due to lack of cohesion. Nevertheless, diverse trace fossil assemblagesmanage to appear in the fossil record of eolianites. Preservation, andprobably even much of the production of organism traces, can be explainedonly by the presence of water in the environment of deposition. In eolianenvironments, moisture in the sediment can result from precipitation, fog,dew, streams fed by adjacent highlands, or deep springs.
Most studies of trace fossil preservation in eolianites have consideredonly surficial trackways and impressions. Similar factors must affect thepreservation of shallow burrows, such as those described in this study.Two factors that enhance trace survivorship in eolian settings includecohesion of the substrate by the increased moisture content (McKee 1947;Ahlbrandt et al. 1978; Stokes and Madsen 1979; McKeever 1991; Sadler1993; Davis et al. 2007) and/or increased clay mineral content (Walkerand Harms 1972; Loope 1986; McKeever 1991; Davis et al. 2007). If atrace can retain its shape long enough for burial, which occurs frequentlyin the high sedimentation rates of eolian settings, it has a good chance ofpreservation.
Two studies suggesting that clay mineral content aids in tracepreservation are directly correlated to raindrop impressions (Walkerand Harms 1972; McKeever 1991), indicating that a moist substrate, inaddition to clay mineral content, also is likely an important factor inpreservation. Experiments conducted by Davis et al. (2007) show thattrackways in clay-enriched substrate are more resistant to wind, but arenot subjected to burial. Loope (1986) studied only large vertebratetrackways that disrupt multiple laminae, making the effect of claycoatings on small burrows and trackways unclear.
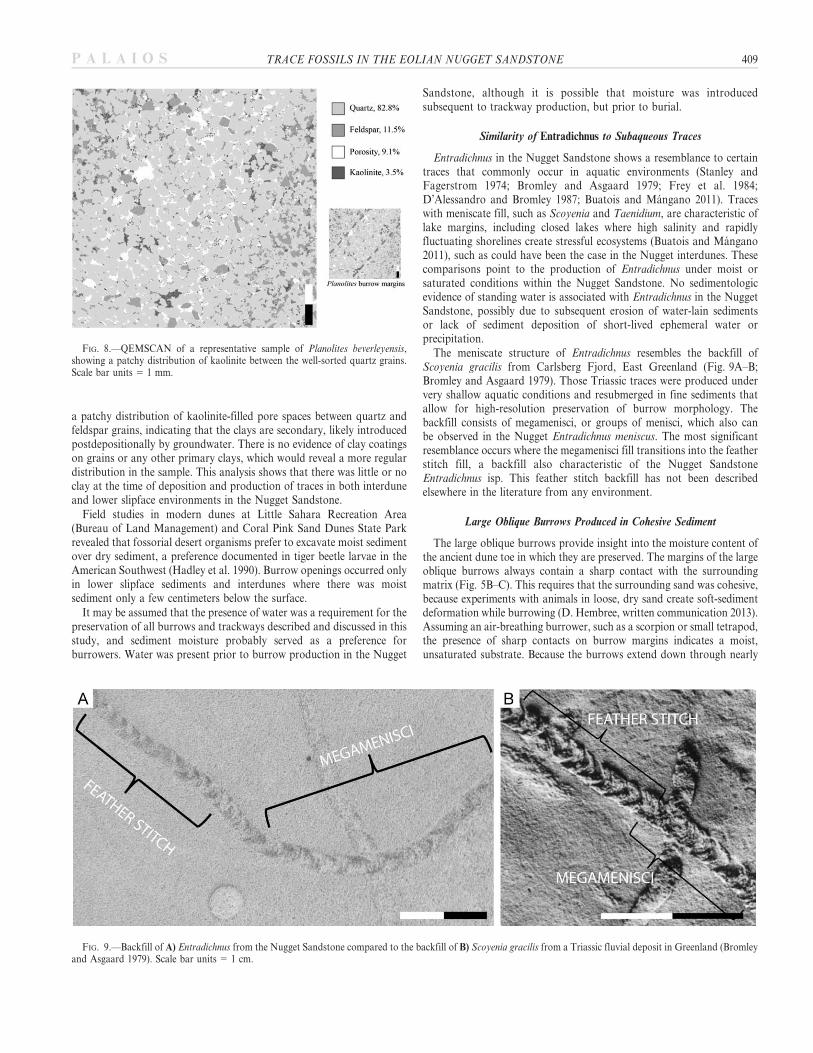
In order to investigate the role of clay in trace preservation,QEMSCANTM analysis was conducted on a representative sample ofPlanolites beverleyensis from the Nugget Sandstone (Fig. 8). Results show
FIG. 7.—Paleohelcura, Octopodichnus, and Brasilichnium trackways in theNugget Sandstone. Scale bar units 5 1 cm. A) Paleohelcura preserved in epireliefwith B) close-up image. Specimen UUIC 3337 is a cast of a portion of thistrackway. C) Octopodichnus (DNM 0486) showing a more complete preservationof the left side of the trackway with push-up rims indicating a downslope directionto the left of the photograph. D) Octopodichnus (cast UUIC 3336) showing a morecomplete preservation of the right side of the trackway with push-up rimsindicating a downslope direction to the right of the photograph. E) Brasilichniumwith individual toe impressions and push-up rims preserved in epirelief (photocourtesy of NPS).
408 T.R. GOOD AND A.A. EKDALE P A L A I O S
a patchy distribution of kaolinite-filled pore spaces between quartz andfeldspar grains, indicating that the clays are secondary, likely introducedpostdepositionally by groundwater. There is no evidence of clay coatingson grains or any other primary clays, which would reveal a more regulardistribution in the sample. This analysis shows that there was little or noclay at the time of deposition and production of traces in both interduneand lower slipface environments in the Nugget Sandstone.
Field studies in modern dunes at Little Sahara Recreation Area(Bureau of Land Management) and Coral Pink Sand Dunes State Parkrevealed that fossorial desert organisms prefer to excavate moist sedimentover dry sediment, a preference documented in tiger beetle larvae in theAmerican Southwest (Hadley et al. 1990). Burrow openings occurred onlyin lower slipface sediments and interdunes where there was moistsediment only a few centimeters below the surface.
It may be assumed that the presence of water was a requirement for thepreservation of all burrows and trackways described and discussed in thisstudy, and sediment moisture probably served as a preference forburrowers. Water was present prior to burrow production in the Nugget
Sandstone, although it is possible that moisture was introducedsubsequent to trackway production, but prior to burial.
Similarity of Entradichnus to Subaqueous Traces
Entradichnus in the Nugget Sandstone shows a resemblance to certaintraces that commonly occur in aquatic environments (Stanley andFagerstrom 1974; Bromley and Asgaard 1979; Frey et al. 1984;D’Alessandro and Bromley 1987; Buatois and Mangano 2011). Traceswith meniscate fill, such as Scoyenia and Taenidium, are characteristic oflake margins, including closed lakes where high salinity and rapidlyfluctuating shorelines create stressful ecosystems (Buatois and Mangano2011), such as could have been the case in the Nugget interdunes. Thesecomparisons point to the production of Entradichnus under moist orsaturated conditions within the Nugget Sandstone. No sedimentologicevidence of standing water is associated with Entradichnus in the NuggetSandstone, possibly due to subsequent erosion of water-lain sedimentsor lack of sediment deposition of short-lived ephemeral water orprecipitation.
The meniscate structure of Entradichnus resembles the backfill ofScoyenia gracilis from Carlsberg Fjord, East Greenland (Fig. 9A–B;Bromley and Asgaard 1979). Those Triassic traces were produced undervery shallow aquatic conditions and resubmerged in fine sediments thatallow for high-resolution preservation of burrow morphology. Thebackfill consists of megamenisci, or groups of menisci, which also canbe observed in the Nugget Entradichnus meniscus. The most significantresemblance occurs where the megamenisci fill transitions into the featherstitch fill, a backfill also characteristic of the Nugget SandstoneEntradichnus isp. This feather stitch backfill has not been describedelsewhere in the literature from any environment.
Large Oblique Burrows Produced in Cohesive Sediment
The large oblique burrows provide insight into the moisture content ofthe ancient dune toe in which they are preserved. The margins of the largeoblique burrows always contain a sharp contact with the surroundingmatrix (Fig. 5B–C). This requires that the surrounding sand was cohesive,because experiments with animals in loose, dry sand create soft-sedimentdeformation while burrowing (D. Hembree, written communication 2013).Assuming an air-breathing burrower, such as a scorpion or small tetrapod,the presence of sharp contacts on burrow margins indicates a moist,unsaturated substrate. Because the burrows extend down through nearly
FIG. 9.—Backfill of A) Entradichnus from the Nugget Sandstone compared to the backfill of B) Scoyenia gracilis from a Triassic fluvial deposit in Greenland (Bromleyand Asgaard 1979). Scale bar units 5 1 cm.
FIG. 8.—QEMSCAN of a representative sample of Planolites beverleyensis,showing a patchy distribution of kaolinite between the well-sorted quartz grains.Scale bar units 5 1 mm.
TRACE FOSSILS IN THE EOLIAN NUGGET SANDSTONE 409P A L A I O S
1 m or more of section, it may be concluded that at least 1 m of sedimentwas moist at the time of burrow production. This amount of moistureprobably would result only from recharge by heavy, possibly monsoonal,rainfall, or ephemeral rivers sourced in the adjacent highlands.
Intervals of Increased Moisture
Fossil evidence in the Nugget Sandstone indicates short intervals ofincreased sediment moisture. Entradichnus meniscus and Planolitesbeverleyensis occur most often in dense populations on slipface surfacesthe Nugget Sandstone. Where outcrop availability allows, these surfacescan be traced to laterally and stratigraphically adjacent surfacescontaining identical fossil assemblages. These associations have beentraced stratigraphically to up to 10 m, while lateral depositional dipsections can reach up to 30 m, although outcrop availability generallyprohibits these extensive associations.
The necessary moisture to allow for production and preservation ofthese burrows could have resulted from rainfall associated with summermonsoon seasons or other unrelated climate intervals, or by theformation of dew during daily periods of increased atmospheric moisture.It has been suggested that monsoons developed during deposition of theNavajo Sandstone in southern Utah (Chandler et al. 1992; Chan andArcher 1999; Loope et al. 2001; Loope and Rowe 2003), but the idea ofdew production in the Nugget/Navajo erg has not been entertained in theliterature, perhaps because it is thought that dew alone would notproduce distinguishable sedimentologic features.
Burrows are preserved in thick lateral sections, so dune migration musthave been active during these moist intervals. Grainflows can originatewhen dry sand accumulates at the top of a dune in the cornice, whicheventually fails, producing an avalanche that buries the lower slipfacewhere the burrows would be preserved. Given that the stoss side of dunesis subjected to more wind and direct sunlight than the lee side,desiccation, transport and avalanche could occur before completedesiccation of lower slipface sediments during a hiatus in moisture input,allowing the shallow burrows to retain their morphology in moistsediment for burial.
It has been suggested that dune migration occurred during wet intervalsin the Navajo Sandstone (Loope et al. 2001; Bryant 2011). Modern dunestypically are stabilized to some degree by vegetation, such as grass andother angiosperms, preventing dune migration during wet periods. TheJurassic dunes were barren of this type of stabilizing vegetation, which didnot appear until the Cretaceous, and the dunes therefore could migratefreely if sufficient drying intervals allowed for desiccation of at least thestoss and crest sands of the dunes (Glennie and Evamy 1968; Loope 1988;Marzolf 1988).
The diversity of small vertebrates preserved at the Saints and SinnersQuarry, such as the drepanosaurs and sphenodontians, indicate the long-
term presence of the interdune lake they were deposited in (Daniel Chure,written communication 2012). This is because small vertebrates lack thevagility required to migrate opportunistically across extensive ergenvironments during short intervals to reach isolated and laterallyrestricted areas. They likely were inhabitants of a permanent, productiveecosystem afforded by a long-term shift in climate.
Paleoecology of the Nugget Sandstone
The wide diversity of trace fossils, including trackways and burrows, aswell as thousands of bones from the Saints and Sinners Quarry andpossible plant remains, provide a clear picture of the communitypaleoecology of the Nugget Sandstone. Although sufficient fossilevidence is available to reconstruct a functioning food web, likely gapsand biases in the fossil record inhibit a full reconstruction.
Stratigraphic correlations (Sprinkel et al. 2011) indicate that theNugget Sandstone was deposited over tens of millions of years, leavingfew traces of life within its various stratigraphic levels. Intervalscontaining fossil evidence are limited in lateral extent and are nearlyimpossible to correlate with other specific intervals, making it difficult todemonstrate that uncorrelated fossil occurrences contain taxa andichnotaxa that existed in different locales at exactly the same time. Highdiversities of fossils within the same interval provide evidence forinteractions between trace fossil producers. Fossil preservation in theNugget Sandstone indicates that most animal productivity occurredduring wet climatic intervals, and thus the organisms and theirbehaviors were preserved during similar climates. Because of this, it isassumed that most of the known taxa and ichnotaxa from variousintervals likely inhabited the dune and interdune environmentscontemporaneously.
Primary productivity is essential to support any ecosystem. Althoughthe preservational potential of plant remains in the Nugget Sandstone isextremely low, some evidence for the types of plants that persisted in theseancient ergs is available. Foliage impressions from the Saints and Sinnerslocality are interpreted as cycadeoid (Britt et al. 2010; Chambers et al.2011), and possible sphenophyte (fern) fossils have been found inDinosaur National Monument (Daniel Chure, personal communication2012). Possible algal buildups also were identified in this study (Fig. 10).Elsewhere, plant fossils in the Navajo Sandstone of Arizona, Idaho, andsouthern Utah include horsetails (Equisetum), conifers (araucarians), andcycads (Harshbarger et al. 1957; Oriel 1959; Gilland 1979; Loope 1979;Stokes 1991; Parrish and Falcon-Lang 2007; Wilkens 2008). Becauseconifer fossils occur in the Navajo Sandstone, the partially correlativesimilar environment represented by the Nugget Sandstone to the northmay have harbored conifers as well, in addition to the ferns and cycadsalready preserved there. Additionally, various types of algae, includingbrown algae, and stromatolites also have been described in the NavajoSandstone, adding to the list of potential primary producers in theNugget Sandstone (Stokes 1991; Eisenberg 2003; Loope et al. 2004a;Dorney and Parrish 2009).
With sufficient numbers of primary producers in the NuggetSandstone, primary consumers (herbivores) flourished and supported acomplex food chain. Primary consumers included insects and other smallherbivorous and detritivorous invertebrates that produced the smallertrace fossils described in this study. Larger herbivores, such as the basalsauropodomorph dinosaurs that produced Otozoum trackways in theupper Nugget Sandstone, and ornithischian dinosaurs that producedAnomoepus trackways in the partially correlative Navajo Sandstone, andtritylodonts that produced Brasilichnium, would have aided in energytransfer from primary producers to the top predators. Arachnids, such asthose that produced Octopodichnus and Paleohelcura, presumably wereinsectivorous predators. Drepanosaurs and sphenodontians may haveconsumed both insects and arachnids. Apex predators would have
FIG. 10.—Possible algal buildup (vertical view) preserved in carbonate layernear Taenidium isp. ‘‘A’’. Scale bar units 5 10 cm.
410 T.R. GOOD AND A.A. EKDALE P A L A I O S
included theropods, represented by the ichnotaxa Grallator andEubrontes, as well as theropod teeth and coelophysoid body fossils fromthe Saints and Sinners Quarry. These top predators may have fed on largearthropods or small vertebrates, such as sphenodontians and synapsids.
A complex trophic structure of the Nugget ecosystem would haverequired an adequate amount of water in the substrate as well as standingwater. Because the Nugget erg persisted in an arid climate, as evidencedby its large, cross-bedded dune deposits, populations of at least the largevertebrates likely were transient, inhabiting the erg only duringsufficiently wet and lengthy climatic intervals, and leaving the smallerherbivorous, insectivorous, and more desert-adapted animals to persistduring dryer intervals. In this way, organism diversity and food webcomplexity were increased during wetter climatic intervals, and theydecreased during more arid intervals.
SUMMARY AND CONCLUSIONS
The Upper Triassic/Lower Jurassic Nugget Sandstone represents aportion of a vast inland erg of eolian sand dunes that were populated by adiverse assemblage of taxa, including both vertebrates and invertebrates,represented primarily by their trace fossils. Body fossils from theseeolianites are extremely rare, making the trace fossil record essential fordeciphering the paleoecology of this ancient dune system.
This study describes 12 types of invertebrate trace fossils from 30 sitesin the Nugget Sandstone of northeastern Utah. Many of the invertebratetrace fossils are described in the Nugget Sandstone for the first time, andall are new to the studied field area. These trace fossils includeEntradichnus meniscus, Entradichnus isp., Planolites beverleyensis, Taeni-dium isp. ‘‘A’’, Taenidium isp. ‘‘B’’, Skolithos, and Planolites isp., burrowclusters, large oblique burrows, flared burrows, Paleohelcura andOctopodichnus. Possible tracemakers include arachnids and insects.Production of the traces was controlled by the dune facies (Fig. 2) andsediment moisture content (Table 3).
Small invertebrate burrows require at least some sediment cohesion forboth burrower preference and preservation. Multiple lines of evidencesupport the hypothesis that moisture provided sediment cohesion in theNugget Sandstone: (1) no evidence of clay grain coatings or other primaryclays was indicated in QEMSCAN analysis of Nugget burrows, indicatingthat electrostatic charges between clay particles were not the agent forsubstrate cohesion; (2) modern burrowers in Utah eolian environmentsshow a preference for moist sediment; (3) meniscate burrows in theNugget Sandstone resemble fluvial or shallow lacustrine burrows, such asScoyenia and Taenidium, indicating production in similarly moistsediment.
Extended periods of increased moisture are interpreted from trace andbody fossils in the Nugget Sandstone. Preservation of Planolitesbeverleyensis in lateral depositional dip sections reaching as much as30 m indicates a steady moisture input for hundreds to thousands ofyears, with active dune migration during this time. Additionally, the bodyfossils of small vertebrates from the Saints and Sinners Quarry representorganisms not adapted to extreme arid environments, suggesting that atleast one (and probably more) long-term wet climatic interval occurredduring deposition of the Nugget Sandstone.
With findings from this study, evidence for all parts of a functionalfood web is now available in the Nugget Sandstone. This includes primaryproduction by plants and possible algal mounds, primary consumption byinsects and other large herbivores, secondary consumption by arachnidsand small vertebrates, and tertiary consumption by theropods, as theapex predators.
Trace fossils play a critical role in interpreting the paleoecology of theancient dune environment represented by the Nugget Sandstone. Futurestudies in neoichnology will help refine interpretations of possibletracemakers in ancient ergs, and comparative studies of other early
Mesozoic eolianites will provide a more complete understanding of thespatial and temporal extent of their life habits.
ACKNOWLEDGMENTS
This project would not have happened without the work and support ofDaniel Chure and the National Park Service (NPS). This study greatlybenefited from insightful comments and edits by Randall Irmis and MarjorieChan. Kelly Good, Joshua Lively, Sherie Harding, and Warren Andersonprovided field and petrographic analysis assistance. Some field sites andfossils were discovered thanks to an NPS survey of the Nugget Sandstoneduring from 2009–2012 conducted in large part by Daniel Chure and GeorgeEngelmann. Other sites were located in 2011 in an additional survey of theNugget Sandstone conducted by Joanna Panosky and Keegan Melstrom,both of whom were part of the NPS’s Geoscientists-in-the-Parks (GIP)summer program. Funding was provided by an ExxonMobil GraduateStudent Research Grant, The Colorado Scientific Society, and the NationalScience Foundation’s GK-12 program, grant #DGE0841233.
REFERENCES
AHLBRANDT, T.S., ANDREWS, S., AND GWYNNE, D.T., 1978, Bioturbation in eoliandeposits: Journal of Sedimentary Petrology, v. 48, no. 3, p. 839–848.
ALBERS, S.H., 1975, Paleoenvironment of the Upper Triassic–Lower Jurassic (?) Nugget(?) Sandstone near Heber, Utah: Unpublished M.S. thesis, University of Utah, SaltLake City, 94 p.
ANDERSON, J.L., 2011, Terrestrial vertebrate trackways of the Early Jurassic NuggetFormation at Dinosaur National Monument, Utah: Geological Society of AmericaAbstracts with Programs, v. 43, no. 5, p. 85.
BOWN, T.M., AND KRAUS, M.J., 1983, Ichnofossils of the alluvial Willwood Formation(lower Eocene), Bighorn Basin, northwest Wyoming, U.S.A.: Palaeogeography,Palaeoclimatology, Palaeoecology, v. 43, no. 1–2, p. 95–128.
BRADY, L.F., 1947, Invertebrate tracks from the Coconino sandstone of northernArizona: Journal of Paleontology, v. 21, p. 150.
BRADY, L.F., 1961, A new species of Palaeohelcura Gilmore from the Permian ofnorthern Arizona: Journal of Paleontology, v. 35, no. 1, p. 201–202.
BRITT, B.B., CHURE, D.J., ENGELMANN, G.F., SCHEETZ, R., AND HANSEN, R., 2010, Multi-taxic theropod bonebeds in an interdunal setting of the Early Jurassic eolian NuggetSandstone, Utah: Society of Vertebrate Paleontology, 70th Annual Meeting, Programsand Abstracts, Supplement to the online Journal of Vertebrate Paleontology, p. 65A.
BRITT, B.B., CHAMBERS, M., ENGELMANN, G.F., CHURE, D.J., AND SCHEETZ, R., 2011,Taphonomy of ceolophysoid theropod bonebeds preserved along the shoreline of anEarly Jurassic Lake in the Nugget Sandstone of NE Utah: Society of VertebratePaleontology, 71st Annual Meeting, Programs and Abstracts, Supplement to theonline Journal of Vertebrate Paleontology, p. 78.
BROMLEY, R.G., AND ASGAARD, U., 1979, Triassic freshwater ichnocoenoses fromCarlsberg Fjord, East Greenland: Palaeogeography, Palaeoclimatology, Palaeoecol-ogy, v. 28, p. 39–80.
BRYANT, G.C., 2011, Outcrop studies of soft-sediment deformation features in theNavajo Sandstone: Unpublished Ph.D. thesis, University of Toronto, 184 p.
BUATOIS, L.A., AND MANGANO, G.M., 2011, Ichnology: organism-substrate interactionsin space and time, New York, Cambridge University Press, p. 207–208.
BUSS, F., 1921, Footprints on the sands of time: Improvement Era, v. 25, no. 2, p. 159–161.CHAMBERS, M., KIMBERLY, H., BRITT, B.B., CHURE, D.J., ENGELMANN, G.F., AND
SCHEETZ, R., 2011, Preliminary taphonomic analysis of a Ceolophysoid theropoddinosaur bonebed in the Early Jurassic Nugget Sandstone of Utah: Geological Societyof America Abstracts with Programs, v. 42, no. 4, p. 16.
CHAN, M.A., AND ARCHER, A.W., 1999, Spectral analysis of eolian foreset periodicities:implications for Jurassic decadal-scale paleoclimatic oscillators: Paleoclimates, v. 3,no. 4, p. 239–255.
CHAN, M.A., AND ARCHER, A.W., 2000, Cyclic eolian stratification on the JurassicNavajo Sandstone, Zion National Park: Periodicities and implications for paleocli-mate, in Sprinkel, D.A., Chidsey, T.C., Jr., and Anderson, P.B., eds., Geology ofUtah’s Parks and Monuments, Vol. 28: Utah Geological Association Publication,p. 607–617.
CHANDLER, M.A., RIND, D., AND RUEDY, R., 1992, Pangaean climate during the EarlyJurassic: GCM simulations and the sedimentary record of paleoclimate: GeologicalSociety of America Bulletin, v. 104, p. 543–559.
CHURE, D.J., GOOD, T.R., AND ENGELMANN, G.F., 2014, A forgotten collection ofvertebrate and invertebrate ichnofossils from the Nugget Sandstone (?Late Triassic–?Early Jurassic), near Heber, Wasatch County, Utah, in Lockley, M.G., and Lucas,S.G., eds., Fossil Footprints of Western North America: New Mexico Museum ofNatural History and Science Bulletin no. 62, p. 181–196.
COSTA, G., 1995, Behavioural Adaptations of Desert Animals. Adaptations of DesertOrganisms: Berlin, Springer-Verlag, 198 p.
COUNTS, J.W., AND HASIOTIS, S.T., 2009, Neoichnological experiments with maskedchafer beetles (Coleoptera: Scarabaeidae): implications for backfilled continental tracefossils: PALAIOS, v. 24, no. 2, p. 74–91.
TRACE FOSSILS IN THE EOLIAN NUGGET SANDSTONE 411P A L A I O S
CRAWFORD, C.S., 1981, Biology of Desert Invertebrates: Berlin, Heidelberg, New York,Springer-Verlag, 314 p.
D’ALESSANDRO, A., AND BROMLEY, R.G., 1987, Meniscate trace fossils and theMuensteria-Taenidium problem: Journal of Paleontology, v. 30, no. 4, p. 743–763.
DAVIS, R.B., MINTER, N.J., AND BRADDY, S.J., 2007, The neoichnology of terrestrialarthropods: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 255, no. 3–4,p. 284–307.
DOELGER, N.M., 1981, Depositional environments of the Nugget Sandstone, RedCanyon Rim, Fremont County, Wyoming: Unpublished M.S. thesis, University ofWyoming, Laramie, 188 p.
DOELGER, N.M., 1987, The stratigraphy of the Nugget Sandstone: Wyoming GeologicalAssociation 38th Annual Field Conference Guidebook, p. 163–178.
DORNEY, L.J., AND PARRISH, J.T., 2009, Carbonate mound structures in the JurassicNavajo Sandstone of southeastern Utah: Geological Society of America Abstractswith Programs, v. 41, no. 6, p. 40.
DOTT, R.H., JR., 1979, Nugget-Navajo Sandstone environmental war: Can trace fossilshelp?: The American Association of Petroleum Geologists Bulletin, v. 63, no. 8,p. 1424–1425.
EISENBERG, L., 2003, Giant stromatolites and a supersurface in the Navajo Sandstone,Capitol Reef National Park, Utah: Geology, v. 31, no. 2, p. 111–114.
EKDALE, A.A., AND PICARD, M.D., 1985, Trace fossils in a Jurassic eolianite, EntradaSandstone, Utah, USA, in Curran, H.A., ed., The Use of Biogenic Structures inInterpreting Depositional Environments, Vol. 5: Society of Economic Paleontologistsand Mineralogists (SEPM) Special Publication, p. 3–12.
EKDALE, A.A., BROMLEY, R.G., AND LOOPE, D.B., 2007, Ichnofacies of an ancient erg: aclimatically influenced trace fossil association in the Jurassic Navajo Sandstone,southern Utah, USA, in Miller, W., ed., Trace Fossils, Concepts, Problems, Prospects:Amsterdam, Elsevier, p. 562–574.
ENGELMANN, G.F., CHURE, D.J., AND LOOPE, D.B., 2010, An occurrence of remarkablyabundant Brasilichnium tracks (Nugget Sandstone, Early Jurassic, Dinosaur NationalMonument) and their environmental context: Geological Society of AmericaAbstracts with Programs, v. 42, no. 5, p. 642.
ENGELMANN, G.F., CHURE, D.J., AND LOOPE, D.B., 2011, Interdune facies containing adinosaur bone bed in the Lower Jurassic Nugget Sandstone in northeastern Utah:Geological Society of America Abstracts with Programs, v. 43, no. 5, p. 263.
ENGELMANN, G.F., CHURE, D.J., BRITT, B.B., AND ANDRUS, A., 2012, The biostrati-graphic and paleoecological significance of a new drepanosaur from the Triassic–?Jurassic Nugget Sandstone of northeastern Utah: Geological Society of AmericaAbstracts with Programs, v. 44, no. 7, p. 604.
ENGELMANN, G.F., CHURE, D.J., AND GOOD, T.R., 2014, Large burrows in the dunes ofthe Nugget Sandstone, Early Jurassic?, of NE Utah, in Lockley, M.G., and Lucas,S.G., eds., Fossil Footprints of Western North America: New Mexico Museum ofNatural History and Science Bulletin no. 62.
FAUL, H., AND ROBERTS, W.A., 1951, New fossil footprints from the Navajo(?)Sandstone of Colorado: Journal of Paleontology, v. 25, p. 266–274.
FREY, R.W., PEMBERTON, G.S., AND FAGERSTROM, J.A., 1984, Morphological,ethological, and environmental significance of the ichnogenera Scoyenia andAncorichnus: Journal of Paleontology, v. 58, no. 2, p. 511–528.
GILLAND, J.K., 1979, Paleoenvironment of a carbonate lens in the lower NavajoSandstone near Moab, Utah: Utah Geology, v. 6, p. 29–38.
GILMORE, C.W., 1926, Fossil footprints from the Grand Canyon: SmithsonianMiscellaneous Collections, v. 77, p. 1–41.
GILMORE, C.W., 1927, Fossil footprints from the Grand Canyon: second contribution:Smithsonian Miscellaneous Collections, v. 80, p. 1–78.
GLENNIE, K.W., AND EVAMY, B.D., 1968, Plants and plant-root structures associatedwith aeolian sand: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 4, p. 77–87.
GREGSON, J.D., AND CHURE, D.J., 2000, Geology and paleontology of DinosaurNational Monument, Utah-Colorado, in Sprinkel, D.A., Chidsey, T.C., Jr., andAnderson, P.B., eds., Geology of Utah’s Parks and Monuments, Vol. 28: UtahGeological Association Publication, p. 155–188.
HADLEY, N.F., KNISLEY, C.B., SCHULTZ, T.D., AND PEARSON, D.L., 1990, Water relationsof tiger beele larvae (Cicindela marutha): correlations with habitat microclimate andburrowing activity: Journal of Arid Environments, v. 19, p. 189–197.
HAMBLIN, A.H., AND BILBEY, S.A., 1999, A dinosaur track site in the Navajo-NuggetSandstone, Red Fleet Reservoir, Uintah County, Utah, in Gillette, D.D., ed.,Vertebrate Paleontology in Utah: Utah Geological Survey Miscellaneous Publication,v. 99-1, p. 51–57.
HARRINGTON, A., 1978, Burrowing biology of the scorpion Cheloctonus jonesii(Arachnida: Scorpionida: Scorpionidae): Journal of Arachnology, v. 5, p. 243–249.
HARSHBARGER, J.W., REPENNING, C.A., AND IRWIN, J.H., 1957, Stratigraphy of theuppermost Triassic and the Jurassic rocks of the Navajo country: United StatesGeological Survey, Professional Paper 291, p. 74.
HASIOTIS, S.T., AND BOURKE, M.C., 2006, Continental trace fossils and museum exhibits:displaying burrows as organism behaviour frozen in time: The Geological Curator,v. 8, no. 211–226.
HASIOTIS, S.T., AND DUBIEL, R.F., 1994, Ichnofossil tiering in Triassic alluvial paleosols:implications for Pangean continental rocks and paleoclimate, in Beauchamp, B.,Embry, A.F., and Glass, D., eds., Pangea: Global Environments and Resources.Canadian Society of Petroleum Geologists Memoir, 17, p. 311–317.
HAWKE, S.D., AND FARLEY, R.D., 1973, Ecology and behavior of the desert burrowingcockroach, Arenivaga sp. (Dictyoptera, Polyphagidae): Oecologia, v. 11, no. 3, p. 263–279.
HEMBREE, D.I., 2009, Neoichnology of burrowing millipedes: linking modern burrowmorphology, organism behavior, and sediment properties to interpret continentalichnofossils: PALAIOS, v. 24, no. 7, p. 425–439.
HEMBREE, D.I., JOHNSON, L.M., AND TENWALDE, R.W., 2012, Neoichnology of the desertscorpion Hadrurus arizonensis: burrows to biogenic cross lamination: PalaeontologiaElectronica, v. 15, no. 1;10A, p. 34.
HIGH, L.R.J., AND PICARD, M.D., 1975, Sedimentary cycles in the Nugget Sandstone,northeastern Utah: Utah Geology, v. 2, no. 2, p. 117–124.
JORDAN, W.M., 1965, Regional environmental study of the early Mesozoic Nugget andNavajo Sandstones: Unpublished Ph.D. thesis, University of Wisconsin, Madison,206 p.
KAYSER, R.B., 1964, Sedimentary petrology of the Nugget Sandstone (Jurassic),northern Utah, western Wyoming and eastern Idaho: Unpublished M.S. thesis,University of Utah, Salt Lake City, 65 p.
KNAPP, R.R., 1976, Depositional environments and diagenesis of the Nugget Sandstone,south-central Wyoming, northeast Utah and northwest Colorado: Unpublished M.S.thesis, University of Wyoming, Laramie, 67 p.
LEONARDI, G., 1981, Brasilichnium elusivum gen. n., sp. n.: pistas de tetrapode mesozoicoguardadas nas coleccoes do Museu Nacional do Rio de Janeiro: Anais da AcademiaBrasileira de Ciencias, v. 53, no. 4, p. 793–805.
LEWIS, G.E., IRWIN, J.H., AND WILSON, R.F., 1961, Age of the Glen Canyon Group(Triassic and Jurassic) on the Colorado Plateau: Geological Society of AmericaBulletin, v. 72, no. 1437–1440.
LOCKLEY, M.G., 2011, The ichnotaxonomic status of Brasilichnium with specialreference to occurrences in the Navajo Sandstone (Lower Jurassic) in the westernUSA: New Mexico Museum of Natural History and Science, Bulletin 53, p. 306–315.
LOCKLEY, M.G., CONRAD, K., AND PAQUETTE, M., 1991, Distribution and significance ofMesozoic vertebrate trace fossils in Dinosaur National Monument, in Plumb, G., ed.,University of Wyoming National Park Service Research Center, 15th Annual Report,p. 85–90.
LOCKLEY, M.G., CONRAD, K., AND PAQUETTE, M., 1992a, New discoveries of fossilfootprints at Dinosaur National Monument: Park Science, v. 12, no. 3, p. 4–5.
LOCKLEY, M.G., CONRAD, K., PAQUETTE, M., AND FARLOW, J.O., 1992b, Distribution andsignificance of Mesozoic vertebrate trace fossils in Dinosaur National Monument, inPlumb, G., and Harlow, H.J., eds., 16th Annual Report of the National Park ServiceResearch Center, University of Wyoming, p. 74–85.
LOCKLEY, M.G., CONRAD, K., PAQUETTE, M., AND HAMBLIN, A.H., 1992c, Late Triassicvertebrate tracks in the Dinosaur National Monument area: Utah Geological Survey,Miscellaneous Publication, v. 92-3, p. 383–391.
LOCKLEY, M.G., HUNT, A.P., HAUBOLD, H., AND LUCAS, S.G., 1995, Fossil footprints inthe DeChelly Sandstone of Arizona: with paleoecological observations on theichnology of dune facies, in Lucas, S.G., and Heckert, A.B., eds., Early PermianFootprints and Facies: New Mexico Museum of Natural History and Science BulletinNo. 6, p. 225–233.
LOCKLEY, M.G., WRIGHT, J.L., HUNT, A.P., AND LUCAS, S.G., 2001, The Late Triassicsauropod track record comes into focus: old legacies and new paradigms: Geology ofthe Llano Estacado: New Mexico Geological Society Guidebook, v. 52, p. 181–190.
LOOPE, D.B., 1979, Fossil wood and probably root casts in the Navajo Sandstone:Geological Society of America Abstracts with Programs, v. 11, p. 278.
LOOPE, D.B., 1986, Recognizing and utilizing vertebrate tracks in cross section:Cenozoic hoofprints from Nebraska: PALAIOS, v. 1, p. 141–151.
LOOPE, D.B., 1988, Rhizoliths in ancient eolianites, in Kocurek, G., ed., Late Paleozoicand Mesozoic Eolian Deposits of the Western Interior of the United States, Vol. 56:Sedimentary Geology, p. 301–314.
LOOPE, D.B., AND ROWE, C.M., 2003, Long-lived pluvial episodes during deposition ofthe Navajo Sandstone: The Journal of Geology, v. 111, p. 223–232.
LOOPE, D.B., ROWE, C.M., AND JOECKEL, R.M., 2001, Annual monsoon rains recordedby Jurassic dunes: Nature, v. 412, p. 64–66.
LOOPE, D.B., EISENBERG, L., AND WAISS, E., 2004a, Navajo sand sea of near-equatorialPangea: Tropical westerlies, slumps, and giant stromatolites, in Nelson, E.P., andErslev, E.A., eds., Field Trips in the Southern Rocky Mountains, USA: GeologicalSociety of America Field Guide 5, p. 1–13.
LOOPE, D.B., STEINER, M.B., ROWE, C.M., AND LANCASTER, N., 2004b, Tropicalwesterlies over Pangaean sand seas: Sedimentology, v. 51, no. 2, p. 315–322.
LUCAS, S.G., LOCKLEY, M.G., HUNT, A.P., AND TANNER, L.H., 2006, Biostratigraphicsignificance of tetrapod footprints from the Triassic–Jurassic Wingate Sandstone onthe Colorado Plateau: New Mexico Museum of Natural History and Science, Bulletin37, p. 194–201.
MARZOLF, J.E., 1988, Controls on late Paleozoic and early Mesozoic eolian deposition ofthe western United States: Sedimentary Geology, v. 56, p. 167–191.
MCKEE, E.D., 1947, Experiments on the development of tracks in fine cross-beddedsand: Journal of Sedimentary Petrology, v. 17, p. 23–28.
MCKEEVER, P.J., 1991, Trackway preservation in eolian sandstones from the Permian ofScotland: Geology, v. 19, p. 726–729.
ORIEL, S.S., 1959, Problems of the Nugget Sandstone, in McKee et al., eds., PaleotectonicMaps of the Triassic System: U.S. Geological Survey Miscellaneous GeologicInvestigations, Map I-300, p. 23–24.
PACHT, J.A., 1976, Depositional environments and diagenesis of the Nugget Sandstone:western Wyoming and north central Utah: Unpublished M.S. thesis, University ofWyoming, Laramie, 99 p.
PARRISH, J.T., AND FALCON-LANG, H.J., 2007, Coniferous trees associated with interdunedeposits in the Jurassic Navajo Sandstone Formation, Utah, USA: Palaeontology,v. 50, no. 4, p. 829–843.
412 T.R. GOOD AND A.A. EKDALE P A L A I O S
PICARD, M.D., 1975, Facies, Petrography and Petroleum Potential of Nugget Sandstone(Jurassic), Southwestern Wyoming and Northeastern Utah: Rocky MountainAssociation of Geologists Symposium, p. 109–127.
PICARD, M.D., 1977a, Petrology of the Jurassic Nugget Sandstone, northeast Utah andsouthwest Wyoming: Wyoming Geological Association 29th Annual Field ConferenceGuidebook, p. 239–258.
PICARD, M.D., 1977b, Stratigraphic analysis of the Navajo Sandstone: a discussion:Journal of Sedimentary Petrology, v. 47, no. 1, p. 475–483.
SADLER, C.J., 1993, Arthropod trace fossils from the Permian DeChelly Sandstone,Northeastern Arizona: Journal of Paleontology, v. 67, p. 240–249.
SANDERS, C.R., AND PICARD, M.D., 1999, Sedimentology and ichnology of a bioturbatedinterval in the Nugget Sandstone, a Jurassic eolianite, near Peoa, northeastern Utah:Utah Geological Association Publication, v. 27, p. 123–140.
SEELY, M.K., AND HAMILTON, W.J., 1976, Fog catchment sand trenches constructed bytenebrionid beetles, lepidochora, from the Namib Desert: Science, v. 193, p. 484–486.
SEIKE, K., HASEGAWA, H., AND ICHINNOROV, N., 2010, Preferred orientation of the tracefossil Entradichnus meniscus in eolian dune strata (Djadokhta Formation) atTugrikiin Shiree, southern Mongolia and its paleoecological implications: IslandArc, v. 19, no. 4, p. 622–627.
SMITH, J.J., AND HASIOTIS, S.T., 2008, Traces and burrowing behaviors of the cicadanymph Cicadetta callipe: neoichnology and paleoecological significance of extant soil-dwelling insects: PALAIOS, v. 23, p. 503–513.
SMITH, J.J., HASIOTIS, S.T., KRAUS, M.J., AND WOODY, D.T., 2008, Naktodemasis bowni:new ichnogenus and ichnospecies for adhesive meniscate burrows (AMB), andpaleoenvironmental implications, Paleogene Willwood Formation, Bighorn Basin,Wyoming: Journal of Paleontology, v. 82, no. 2, p. 267–278.
SPRINKEL, D.A., KOWALLIS, B.J., AND JENSEN, P.H., 2011, Correlation and age of theNugget Sandstone and Glen Canyon Group, Utah: Utah Geological AssociationPublication, v. 40, p. 131–149.
STANLEY, K.O., AND FAGERSTROM, J.A., 1974, Miocene invertebrate trace fossils from abraided river environment, western Nebraska, U.S.A.: Palaeogeography, Palaeocli-matology, Palaeoecology, v. 15, p. 63–82.
STANLEY, K.O., JORDAN, W.M., AND DOTT, R.H., JR., 1971, New hypothesis of EarlyJurassic paleogeography and sediment dispersal for Western United States: TheAmerican Association of Petroleum Geologists Bulletin, v. 55, no. 1, p. 10–19.
STOKES, W.L., 1957, Jurassic System of the southern flank of the Uinta Mountains, inGuidebook to the Geology of the Uinta Basin: Intermountain Association ofPetroleum Geologists, p. 92–96.
STOKES, W.L., 1959, Jurassic rocks of the Wasatch Range and vicinity: 10th FieldConference Guidebook: Intermountain Association of Petroleum Geologists, p. 109–114.
STOKES, W.L., 1978, Animal tracks in the Navajo-Nugget Sandstone: Contributions toGeology: University of Wyoming, v. 16, no. 2, p. 103–107.
STOKES, W.L., 1991, Petrified mini-forests of the Navajo Sandstone, east-central Utah:Survey Notes: Utah Geological and Mineral Survey, v. 25, no. 1, p. 14–19.
STOKES, W.L., AND MADSEN, J., JR., 1979, Environmental significance of pterosaurtracks in the Navajo Sandstone, Grand County, Utah: Brigham Young UniversityGeological Studies, v. 26, p. 21–26.
TSCHINKEL, W.R., 2002, Subterranean ant nests: trace fossil past and future?:Palaeogeography, Palaeoclimatology, Palaeoecology, v. 192, p. 321–333.
WALKER, T.R., AND HARMS, J.C., 1972, Eolian origin of flagstone beds, LyonsSandstone (Permian), type area, Boulder County, Colorado: The MountainGeologist, v. 9, p. 279–288.
WILKENS, N.D., 2008, Paleoecology of Early Jurassic Navajo Sandstone interdunedeposits: Unpublished Ph.D. thesis, Arizona State University, Phoenix, 416 p.
WINKLER, D.A., JACOBS, L.L., CONGLETON, J.D., AND DOWNS, W.R., 1991, Life in a sandsea: biota from Jurassic interdunes: Geology, v. 19, p. 889–892.
WROBLE, J.L., 1953, Stratigraphy and sedimentation of the Popo Agie, Nugget, andSundance Formations in central Wyoming: Unpublished M.A. thesis, University ofWyoming, Laramie, 151 p.
Received 30 January 2014; accepted 24 July 2014.
TRACE FOSSILS IN THE EOLIAN NUGGET SANDSTONE 413P A L A I O S